A Special Role of Syllables, But Not Vowels or Consonants,
for Nonadjacent Dependency Learning
Ivonne Weyers1,2
and Jutta L. Mueller1,2
Abstract
■ Successful language processing entails tracking (morpho)
syntactic relationships between distant units of speech, so-called
nonadjacent dependencies (NADs). Many cues to such depen-
dency relations have been identified, yet the linguistic elements
encoding them have received little attention. In the present
investigation, we tested whether and how these elements, here
syllables, consonants, and vowels, affect behavioral learning
success as well as learning-related changes in neural activity in
relation to item-specific NAD learning. In a set of two EEG studies
with adults, we compared learning under conditions where
either all segment types (Experiment 1) or only one segment
type (Experiment 2) was informative. The collected behavioral
and ERP data indicate that, when all three segment types are
available, participants mainly rely on the syllable for NAD
learning. With only one segment type available for learning,
adults also perform most successfully with syllable-based
dependencies. Although we find no evidence for successful
learning across vowels in Experiment 2, dependencies between
consonants seem to be identified at least passively at the
phonetic-feature level. Together, these results suggest that
successful item-specific NAD learning may depend on the
availability of syllabic information. Furthermore, they highlight
consonants’ distinctive power to support lexical processes.
Although syllables show a clear facilitatory function for NAD
learning, the underlying mechanisms of this advantage require
further research. ■
INTRODUCTION
Processing dependencies between temporally distant
units of speech (e.g., “he sings” or “The girl the boy kissed
ran away.”) is essential to human language. The hierar-
chical structure of language requires tracking the relation-
ships between units, such as words or phrases, beyond
the directly adjacent environment and across variable
numbers of intervening elements. By studying the cogni-
tive processes involved in the detection and learning of
these so-called nonadjacent dependencies (NADs), we
can learn about some of the most basic mechanisms
supporting language processing and acquisition. Although
it is known that NADs can be learned in principle, and
many external cues have been identified that guide learn-
ing (Wilson et al., 2018), comparatively little research has
explored the role of the speech sounds themselves that
carry the dependency. The specific acoustic features of
the input are particularly relevant during early acquisition
of NADs because, initially, phonetic surface-level forms
have to be identified as re-occurring patterns in the input.
The resulting early, item-specific representations form the
basis for the later development of more abstract, categor-
ical relations (Mueller, ten Cate, & Toro, 2020; Culbertson,
Koulaguina, Gonzalez-Gomez, Legendre, & Nazzi, 2016).
In other words, the ability to detect and recognize depen-
dencies between phonetic elements in linguistic input
1University of Vienna, 2University of Osnabrück
© 2022 Massachusetts Institute of Technology
can be understood as a precursor or an initial “bootstrap-
ping” process that paves the way for the acquisition of
higher-level syntactic rules. This study aims to compare
learning-related changes in neural activity during NAD
learning and processing and their dependence on the
segmental level at which they are encoded, namely, sylla-
bles, consonants, and vowels.
In this line of research, artificial grammar learning (AGL)
paradigms have proven a useful means to isolate surface-
level structural processing from semantic meaning and
syntactic function, while also controlling for effects of
previous language learning (e.g., Frost & Monaghan,
2016; Newport & Aslin, 2004; Marcus, Vijayan, Bandi
Rao, & Vishton, 1999; Reber, 1967). Gómez (2002), for
example, used sequences of nonword triplets ( pel kicey
rud, vot wadim jic) in which the first (A) and third (B)
word encoded a simple AXB NAD across a variable middle
word (X). After passive auditory exposure to these strings,
both adult and infant participants showed behavioral evi-
dence of learning by successfully discriminating between
consistent ( pel kicey rud) and inconsistent ( pel kicey jic)
exemplars. Since then, many studies have confirmed that
adults (Frost & Monaghan, 2016; Vuong, Meyer, &
Christiansen, 2016; Mueller, Friederici, & Männel, 2012;
van den Bos, Christiansen, & Misyak, 2012; Citron,
Oberecker, Friederici, & Mueller, 2011; Mueller, Oberecker,
& Friederici, 2009; Peña, Bonatti, Nespor, & Mehler, 2002),
infants (Marchetto & Bonatti, 2015; Mueller et al., 2012;
Gómez & Maye, 2005), and even nonhuman primates
Journal of Cognitive Neuroscience 34:8, pp. 1467–1487
https://doi.org/10.1162/jocn_a_01874
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
(Malassis, Rey, & Fagot, 2018; Milne et al., 2016) are able to
learn such arbitrary AXB NAD relations from speech (for a
review, see Mueller, Milne, & Männel, 2018).
A variety of NADs and their learnability have been inves-
tigated, differing, for example, in the type of relationship
between dependent units (e.g., repeated elements [AXA]
or item-specific dependencies [AXB]) and the complexity
of the structure that is encoded (e.g., simple AXB relation-
ships or crossed dependencies [A1A2A3B1B2B3]; for a
review, see Wilson et al., 2018). Many studies have further
focused on identifying circumstances or cues that facilitate
(or hinder) the learning of NADs. They are learned better,
for instance, when the variability of intervening X ele-
ments is high (Gómez & Maye, 2005; Onnis, Monaghan,
Christiansen, & Chater, 2004; Gómez, 2002), when the
dependent elements appear in edge positions (Endress,
Nespor, & Mehler, 2009), when they are highlighted by
pauses (Mueller, Bahlmann, & Friederici, 2008; Peña
et al., 2002) or other prosodic cues (Grama, Kerkhoff,
& Wijnen, 2016; Mueller, Bahlmann, & Friederici, 2010),
or when they are perceptually similar (Creel, Newport, &
Aslin, 2004; Newport & Aslin, 2004). Few studies, however,
have systematically investigated the role of the specific lin-
guistic elements encoding the NAD and their impact on
learning success and processing. In all of the studies cited
above, the encoding elements were artificial monosyllabic
or multisyllabic units. Yet, whether NADs are coded by
those units or rather by segments forming those units, that
is, consonants and vowels, is not known. Thus, in this
study, we ask whether syllables, consonants, and vowels
are equally suitable computational units for NAD learning.
Before we turn to this study aiming to answer these ques-
tions, we briefly review the relevant previous literature on
the role of linguistic segments in speech and particularly
in NAD learning. To this aim, we consider both behavioral
and neurophysiological experiments as both may provide
complementary information about the nature of the
involved cognitive processes.
The general notion that syllables are linguistic units rel-
evant for both language comprehension and production
is not new (e.g., Bertoncini & Mehler, 1981; Mehler,
Dommergues, Frauenfelder, & Segui, 1981; Hooper,
1972). Word production models largely agree that the syl-
lable plays a role in the speech production process and
merely disagree on when syllabic information is made
available (e.g., Schiller & Costa, 2006; Levelt, Roelofs, &
Meyer, 1999; Dell, 1986, 1988; Shattuck-Hufnagel, 1983).
In Levelt’s model of speech production (Levelt, 1989),
for instance, it is presumed that for frequently used sylla-
bles of a given language, independent representations
and motor programs are stored in the so-called mental
syllabary, the activation of which allows for effective pho-
netic encoding and fluent articulation (e.g., Cholin, Dell, &
Levelt, 2011; Cholin, 2008; Levelt et al., 1999; Levelt, 1992).
Such articulatory motor programs at the syllable level have
received support from both modeling (Guenther, 2016;
Guenther, Ghosh, & Tourville, 2006) and experimental
work (Ziegler, Aichert, & Staiger, 2010; Cholin, Levelt, &
Schiller, 2006; Carreiras & Perea, 2004). More recently,
studies using EEG have shown that cortical activity
recorded during continuous speech perception tracks lin-
guistic structure at different levels, including syllable and
word boundaries (Batterink, 2020; Choi, Batterink, Black,
Paller, & Werker, 2020; Poeppel & Assaneo, 2020; Ding,
Melloni, Zhang, Tian, & Poeppel, 2016; Ding & Simon,
2014); in fact, Giraud and Poeppel (2012) showed that
syllabic structure is tracked already in primary auditory
cortex, and a number of studies have related the precision
of syllable tracking to language skills, particularly reading
(Goswami, 2011; Abrams, Nicol, Zecker, & Kraus, 2009),
which further supports the relevance of the syllabic pro-
cessing level. Similarly, a recent computational approach
highlights a possible role of (acoustic) syllables even
in prelinguistic perception and speech sequencing
(Räsänen, Doyle, & Frank, 2018).
Because of its apparent role as a basic perceptual and
production unit, the syllable has been a natural target unit
for AGL studies concerned with NAD learning (e.g.,
Mueller et al., 2012; de Diego-Balaguer, Toro, Rodriguez-
Fornells, & Bachoud-Lévi, 2007; Endress & Bonatti, 2007;
Peña et al., 2002). Mueller et al. (2012), for instance, used a
classic oddball design and auditorily presented adult
listeners with a segmented stream of syllable sequences
encoding an item-specific AXB NAD ( fikato, lerobu),
which was interspersed with few deviant items in which
the final syllable violated the AXB dependency ( fiwebu,
lekoto). While participants performed a target detection
task, their EEG response was recorded. For those partici-
pants who showed behavioral evidence of learning, devi-
ant detection was indexed by an N2/P3 complex in the
ERPs. De Diego-Balaguer et al. (2007) found similar ERP
effects, across both learners and nonlearners, using com-
parable items (nulade vs. delanu) and design.
With regard to the smaller segmental level, there is evi-
dence that consonants and vowels are not mere superim-
posed linguistic categories we use to classify speech
sounds but actually constitute separable classes also at
the neural level (Caramazza, Chialant, Capasso, & Miceli,
2000; Boatman, Hall, Goldstein, Lesser, & Gordon,
1997), which are processed by distinct neural mechanisms
(Carreiras, Dunabeitia, & Molinaro, 2009; Carreiras, Gillon-
Dowens, Vergara, & Perea, 2009; Carreiras & Price, 2008;
Carreiras, Vergara, & Perea, 2007). Carreiras and Price
(2008) presented participants with written words in
which either consonants (e.g., PRIVAMERA) or vowels
(PRIMEVARA) were transposed. Participants had to either
read the words aloud or perform a lexical decision task
while MRI brain scans were acquired. Whereas vowel
changes induced increased relative activation in the STS
during reading out loud, consonant changes exhibited
increased activation in the right middle frontal cortex
in the lexical decision task. The authors concluded
that vowel changes placed additional demands on areas
relevant for prosodic processing—possibly because of
1468
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
self-monitoring processes engaged during production.
Consonant changes, on the other hand, additionally
taxed inhibitory control mechanisms during lexical deci-
sion, indicating more difficulties with lexico-semantic
processing.
Consonants and vowels have been compared exten-
sively concerning their functional role in word segmenta-
tion (Nazzi, Poltrock, & Von Holzen, 2016; Toro, Nespor,
Mehler, & Bonatti, 2008; Mehler, Peña, Nespor, &
Bonatti, 2006; Bonatti, Peña, Nespor, & Mehler, 2005;
Newport & Aslin, 2004; Peña et al., 2002) and word
identification/lexical selection (Delle Luche et al., 2014;
Havy, Serres, & Nazzi, 2014; Carreiras, Dunabeitia, et al.,
2009; Carreiras, Gillon-Dowens, et al., 2009; New, Araújo,
& Nazzi, 2008; Cutler, Sebastian-Galles, Soler-Vilageliu, &
Van Ooijen, 2000). When asked to reconstruct a word from
a nonword (kebra) by changing a single phoneme, for
instance, adult participants prefer to make a vowel change
(cobra) rather than a consonant change (zebra; Sharp,
Scott, Cutler, & Wise, 2005; Cutler et al., 2000; Van Ooijen,
1996). This observation holds even cross-linguistically,
both for languages like Spanish with a larger number of
consonants than vowels and for languages with a relatively
equal consonant–vowel ratio, such as Dutch (Cutler et al.,
2000). Similarly, adults can exploit co-occurrence statistics
(transitional probabilities) between consonants to seg-
ment a continuous speech stream into word-like units
(Nazzi et al., 2016; Toro, Nespor, et al., 2008; Mehler
et al., 2006; Bonatti et al., 2005). Equivalent transitional
probabilities between vowels can only be exploited for this
purpose under highly redundant conditions, and in a
direct comparison with equal distributional information
across both segments, adults preferentially extract words
based on consonant rather than vowel frames (Bonatti
et al., 2005; Newport & Aslin, 2004).
This asymmetry is addressed by the “consonant–vowel
(CV) hypothesis,” which proposes that consonants and
vowels assume at least partially distinct functions in lin-
guistic processing: Consonants primarily encode lexical
information, whereas vowels carry sentence prosody and
thereby supply information about syntactic constituency
and sentence structure (Nespor, Peña, & Mehler, 2003).
Although there is abundant evidence for the former
assumption (see above), evidence for the latter remains
scarce. A possible structural role of vowels has mainly been
tested with the help of AGLs encoding item-independent
repetition rules (e.g., fefufu, kufefe). These are deemed
good examples of structural learning for two reasons: They
require generalization of a regularity (ABB/ABA) beyond
specific items (e.g., lumifi vs. lumifa), and vowel repeti-
tions specifically can be conceptualized as an extreme case
of vowel harmony, a phonetic assimilation operation that
provides cues to morphosyntactic constituency in some
languages (e.g., Turkish, Hungarian, Finnish). There is
tentative evidence1 that adults learn such reduplication
rules better when they are encoded by vowels rather than
consonants (Monte-Ordoño & Toro, 2017a; Toro, Nespor,
et al., 2008; Toro, Shukla, Nespor, & Endress, 2008),
although participants in these studies were speakers of
Catalan–Spanish (Monte-Ordoño & Toro, 2017a) and
Italian (Toro, Nespor, et al., 2008; Toro, Shukla, et al.,
2008), languages that do not typically harmonize.
Item-specific NADs between vowels and consonants
have only scarcely been researched. Newport and Aslin
(2004) reported successful segmentation of a continuous
syllable stream into trisyllabic words based on transitional
probabilities between nonadjacent consonant ( p_g_t_)
and vowel (_a_u_e) frames. The authors concede, how-
ever, that the dependencies they employed may not
exactly qualify as “nonadjacent” at the segmental level,
as the entire consonant/vowel frame always remained
fixed and the middle segment did not vary (i.e., pxxxtx).
Specifically, if the assumed statistical learning mechanism
operated on separate representations of consonant and
vowel tiers, or if the segments were simply grouped
together because of their perceptual similarity, the given
dependencies would actually exist between adjacent
units (Newport & Aslin, 2004).
This Study
Our aim in this study was to compare syllables, conso-
nants, and vowels as carriers of item-specific NADs. We
focused on item-specific dependencies of the type AXB,
for example, as used by Gómez (2002). These lend them-
selves well to study NADs in natural language, because
particularly at the local, morphosyntactic level, such NADs
often exist between specific units (e.g., she is running)
whose phonetic surface-level form and arbitrary relation-
ship need to be learned.
The summarized previous studies have shown that audi-
torily presented NADs at the syllable level can be learned
by adults. From these results, it remains unclear, however,
whether the relevant learning mechanism operates on the
syllable level or whether NAD learning is possibly biased or
guided by the lower segmental level. We addressed this
question in Experiment 1, using an experimental design
with alternating learning and test phases. In the learning
phases, participants were exclusively exposed to a
syllable-based NAD. In the test phases, they were also
tested for the consonant- and vowel-based dependencies
inherent in this syllable-based NAD. If participants also
showed signs of discrimination for either of these, this
would suggest a special role for segments smaller than
the syllable in the learning of syllable-based NADs.
On the basis of the available evidence from studies
comparing consonants and vowels, it seems that redun-
dancies or repetition regularities are learned better across
vowels, whereas dependencies between nonrepetitive,
distinctive features are learned better across consonants.
These previous studies have so far only compared the role
of consonants and vowels in segmentation and/or repeti-
tion detection tasks. None of them have evaluated their
role in NAD learning specifically and asked whether, in
Weyers and Mueller
1469
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
principle, item-specific dependencies can be learned
across these segments. We focused on this second ques-
tion in Experiment 2, using an oddball paradigm to expose
three separate groups of adults to input from which only
one type of dependency (syllable/consonant/vowel-based)
could be learned.
During both experiments, we recorded participants’
EEG. The relatively low number of participants showing
behavioral evidence of learning across previous studies
(e.g., Mueller et al., 2012, 10/46 learners; de Diego-
Balaguer et al., 2007, 8/16 learners) underscores the
importance of an additional measure such as EEG. EEG
can provide important insights into online processing
even in the absence of offline learning success and into
possible qualitative differences in the neural processes
that support learning across these segments.
In line with the cited studies, we expected to find high
accuracy rates along with ERP evidence of learning for
syllable NADs in both experiments. With regard to the
two segmental conditions/groups, hypotheses are more
difficult to formulate. One could tentatively expect that
item-based dependencies, which rely on the identifica-
tion, association, and storage of specific segmental units,
are learned better if they are encoded by consonants, as
these appear to have larger distinctive power in the con-
text of word identification and lexical selection. An advan-
tage for vowels has mainly been postulated in the learning
of repetitions. Although we do not employ repetitions, we
purposefully chose phonetically similar segments for our
stimulus material. It is thus conceivable that perceptual
similarity in the vowel condition/group serves as a similar
(albeit weaker) cue to the dependency relationship. As
both of these options are conceivable, we did not have
any specific hypotheses with regard to performance or
neurophysiological responses in the two segmental
conditions/groups.
EXPERIMENT 1
Methods
Participants
The experiment was approved by the ethics committee of
the University of Osnabrück and adheres to the guidelines
of the Declaration of Helsinki (2013). Participants were
recruited from the University of Osnabrück, gave written
informed consent before participating in the experiment,
and received either course credit or payment as compen-
sation for their participation. All tested participants were
native speakers of German, were right-handed, and had
normal hearing, normal or corrected-to-normal vision,
and no history of neurological conditions. On the basis
of the average number of participants in similar previous
studies (e.g., Citron et al., 2011; Mueller et al., 2009;
Mueller, Bahlmann, et al., 2008), we aimed for a minimum
of 25 participants entering the final analysis. Five of the
34 tested participants had to be excluded from analysis
because of technical difficulties or high artifact rate in
the EEG. The remaining 29 participants (2 men, 27
women) were between 18 and 29 years old (mean =
21.62 years, SD = 2.6 years).
Stimuli and Procedure
The stimulus material consisted of trisyllabic sequences of
individually recorded CV syllables spoken by a trained
female speaker. Recordings of similar length and pitch
were selected from several recorded exemplars, digitized
(44.1 kHz/16-bit sampling rate, mono), normalized to
the same sound intensity, and cut to the same length
(380 msec). The two syllable frames bi X pe and go X ku
served as standards of an AXB-type NAD. Within items,
syllables were separated by 50-msec pauses, and items
were separated by 700-msec interstimulus pauses
(Mueller, Bahlmann, et al., 2008; Peña et al., 2002). In an
attempt to boost learning of this pairwise association,
several cues known to aid NAD learning were integrated:
The phonemes coding the dependency were selected for
their perceptual similarity, that is, the consonants differed
only in voicing (b–p, g–k) and the vowels were both either
rounded back (o–u) or unrounded front (i–e) vowels
(Creel et al., 2004; Newport & Aslin, 2004); the relevant
units A and B were placed in edge positions (Endress
et al., 2009); variability of the middle element X was high
with 24 different syllables (la, ma, na, ra, sa, ta, dä, nä,
rä, sä, tä, wä, dö, lö, mö, sö, tö, wö, dü, lü, mü, nü, rü,
wü; Gómez, 2002); and attention to stimuli was required
given the active design (Mueller et al., 2012).
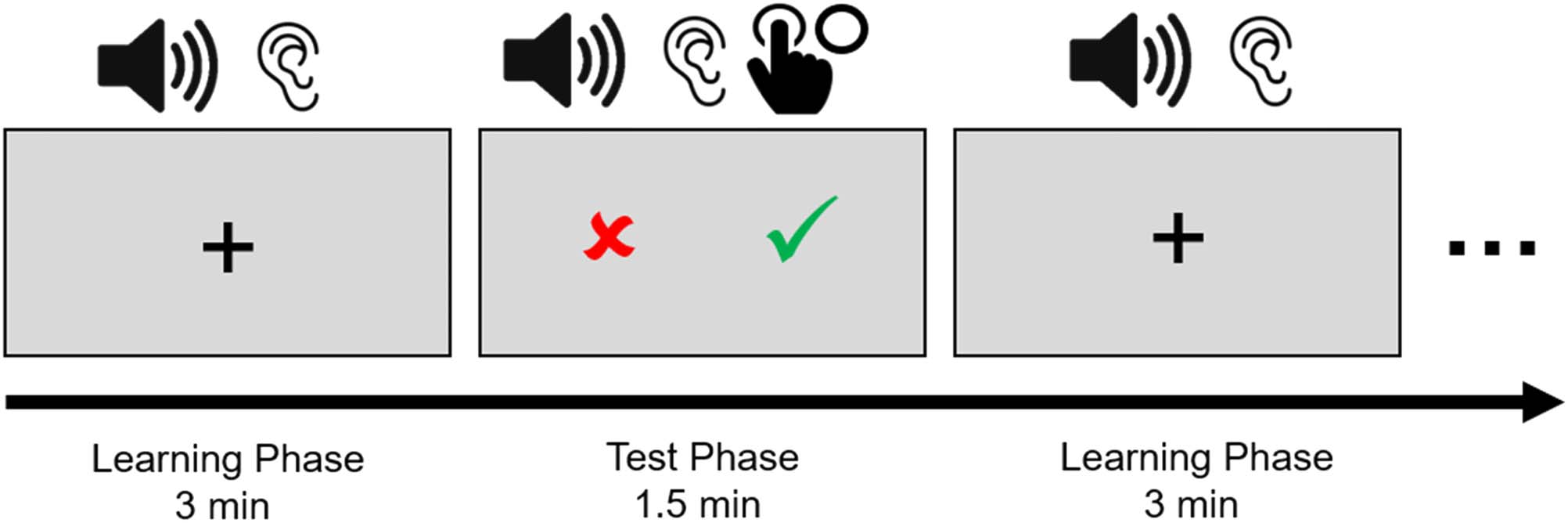
Eight learning phases alternated with eight test phases.
In each learning phase, the same 48 correct syllable items
(see Table 1 for examples) were repeated twice in pseudo-
random order while a fixation cross was shown on the
screen (see Figure 1). Participants were instructed to listen
attentively and detect a regularity inherent in the input,
based on which they would have to make a grammaticality
judgment in the test phases. During the test phases, items
were presented individually,2 and after a short delay of
900 msec, a response cue appeared on the screen (see
Figure 1). No feedback was provided. The test items com-
prised correct (e.g., bidape) and incorrect (e.g., bidaku)
exemplars of the syllable dependency as well as vowel-
based and consonant-based variants of the AXB depen-
dency. In correct exemplars of this segmental-level
dependency, either the vowel dependency (e.g., kowabu)
or the consonant dependency (e.g., gewako) remained
constant compared to the learning phase syllable NADs.
Incorrect exemplars of these lower-level NADs violated the
respective dependency on either the final consonant (e.g.,
gewapi) or the final vowel (e.g., kowage).3 In contrast to
Newport and Aslin’s (2004) technically adjacent segmental
dependencies ( p_g_t_/_a_u_e), our segmental-level
dependencies were truly nonadjacent. Eight separate
middle syllables (da, wa, lä, mä, nö, rö, sü, tü) were used
for the test phase items. Each test phase comprised 24
1470
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Correct and Incorrect Exemplars by Condition, With Violations Underlined
Learning Phases
bilape
bidäpe
bidöpe
bimüpe
gomaku
gowäku
gosöku
gorüku
Syllables
Test Phases
Consonants
Vowels
correct
bidape
biläpe
biröpe
gowaku
gomäku
gotüku
incorrect
bidaku
biläku
biröku
gowape
gomäpe
gotüpe
correct
budapi
buläpi
buröpi
gewako
gemäko
getüko
incorrect
budako
buläko
buröko
gewapi
gemäpi
getüpi
correct
pidage
piläge
piröge
kowabu
komäbu
kotübu
incorrect
pidabu
piläbu
piräbu
kowage
komäge
kotüge
test items, holding correct and incorrect exemplars of the
three conditions. We did not control for an exactly equal
distribution of items per condition in each test phase of
the four item lists created but restricted the number of
items per segmental condition in each learning phase to
a minimum of four and a maximum of 12.4
During the experiment, participants were seated in a
chair at a distance of 100 cm from a computer screen while
the stimuli were played via loudspeakers.
Data Acquisition and Preprocessing
The continuous EEG was recorded from a 64 Ag/AgCl elec-
trode cap (TMSI B.V.; International 10–20 system of elec-
trode placement), using a TMSi 72 Refa amplifier system
and the TMSi Polybench recording software. The data
were recorded with an implicit average online reference
of all electrodes. The ground electrode was placed on
the left collar bone; two additional single electrodes were
placed on both temples, as well as one placed above and
one below the left eye that recorded the horizontal and
vertical EOG. Impedances of all electrodes were kept
below 5 kΩ, and the data were sampled at a rate of
512 Hz with no hardware filters (except for antialiasing)
in place.
The EEG data were processed offline with MATLAB
( Version R2017a, The MathWorks Inc., 2010) and the
EEGLAB open source toolbox (Version 14.1.1b; Delorme
& Makeig, 2004). Before averaging, the continuous data
were rereferenced to average mastoids, detrended, and
filtered with two separate digital windowed-sinc finite
impulse response filters (window type: Kaiser), one
high-pass filter (−6 dB half-amplitude cutoff, 0.1-Hz cutoff
frequency, filter order: 9274), and a low-pass filter (−6 dB,
30 Hz, 188), to remove slow drifts and line noise. For ERP
averaging, epochs from −100 to 1000 msec after the onset
of the final syllable (or vowel in case of the vowel condi-
tion) were cut out. Independent component analysis
(ICA) was performed on the individual participant data
to remove eye movement artifacts.5 Trials containing any
remaining artifacts were selected using a semiautomatic
procedure coupled with visual inspection and excluded
from further analysis. A baseline correction (−100 to
0 msec) was applied to the cleaned data, which were then
averaged by participant for each experimental condition.
The average number of epochs per participant entering
the final analysis amounted to 28.86 (SD = 3.03) correct
Figure 1. Experimental procedure in Experiment 1.
Weyers and Mueller
1471
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
and 28.21 (SD = 3.65) incorrect syllable items, 28.55 (SD =
3.42) correct and 28.69 (SD = 3.17) incorrect consonant
items, and 28.76 (SD = 3.86) correct and 29.17 (SD =
2.88) incorrect vowel
items and did not significantly
differ within or between conditions.
Data Analysis
The behavioral data were analyzed using RStudio (R Core
Team, 2020) and the lme4 package (Bates, Mächler,
Bolker, & Walker, 2015). A generalized linear mixed-
effects model (GLMM) including a binomial link function
was fitted to investigate the effects of condition (syllable,
vowel, consonant), phase (1–8), and list (1–4), as well as
the interaction between condition and phase, on the
response accuracy data. The predictor phase was interval
scaled and centered to facilitate model fitting. The factor
list was included to confirm that the distribution of items
between lists did not affect learning. Participants and items
were included in the model as intercept-only random
effects. Standard treatment contrasts were implemented
with the consonant condition and list A as reference levels
of the respective predictors. In total, 5568 data points (29
participants, eight test phases with 24 items each) were
entered into the model. To test whether, across the whole
group, participants’ response accuracy rates in the three
conditions exceeded chance level, separate intercept
models were fitted to the accuracy data of each condition.
Each of these models comprised 1856 data points.
p Values for fixed effects were calculated via Wald tests
(standard for glmer in lme4).
Participants were then split into learners (response
accuracy ≥ 64%, indicated by a binomial test for chance
response, p = .033) and nonlearners based on their
behavioral performance in each condition. Because of
the fact that the vowel and consonant dependencies
were inherent in the syllable condition items, participants
who learn either of these two dependencies should also
be able to correctly evaluate the syllable condition items.
Participants were thus classified as “syllable” learners,
“vowel + syllable” learners, or “consonant + syllable”
learners.6 We further tested whether the latter two groups
already performed above chance level in both conditions
in the first test phase. If so, this might be indicative of
segment-based NAD learning; if not, it may indicate
sequential learning effects. To this end, we fit additional
intercept models to the accuracy data of the learner
groups, separately for the first and second test phases.
For the vowel + syllable learners, the respective models
included 216 data points, whereas 72 data points were
entered into the consonant + syllable learner models.
T h e F i e l d T ri p t o o l b o x f or E E G / M E G a n a l y s i s
(Oostenveld, Fries, Maris, & Schoffelen, 2011) was used
for statistical analyses of the EEG data. Separate nonpara-
metric cluster-based permutation tests using dependent
samples t tests were run for each segmental condition,
comparing ERP responses to correct and incorrect items.
All electrodes, except for the EOG and reference elec-
trodes, were included in the analysis. Because previous
literature does not provide specific expectations as to
the timing and location of possible effects, the ERP analysis
was exploratory. Only the latency range of 0–50 msec was
excluded from analysis to increase power (Groppe,
Urbach, & Kutas, 2011), because we were not interested
in any auditory brain stem or primary auditory cortex
responses in this very early time window (for a review,
see Pratt, 2012; Picton, 2011). The initial sample-specific
test statistic threshold was set to 0.05. The minimum num-
ber of neighboring channels to be included in a cluster was
set to two, and neighboring channels were identified with
a spatial neighborhood template by use of the triangula-
tion method. For the cluster statistic permutation test,
we employed the maximum sum approach and set the
alpha level of the permutation test to .05 (distributed over
both tails) and the number of draws from the permutation
distribution to 2000 (Monte Carlo sampling).
Results
Behavioral Results
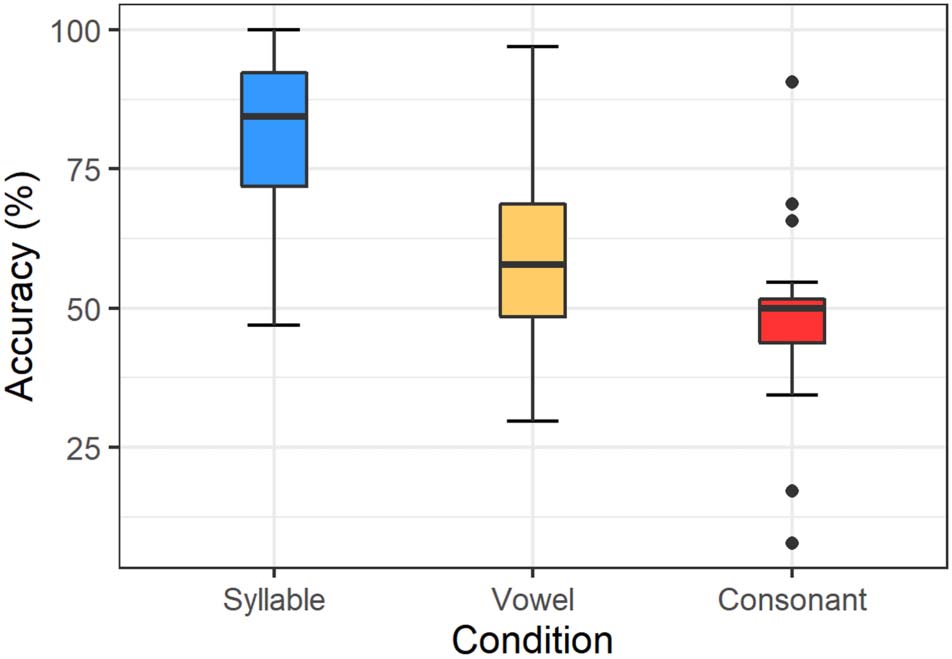
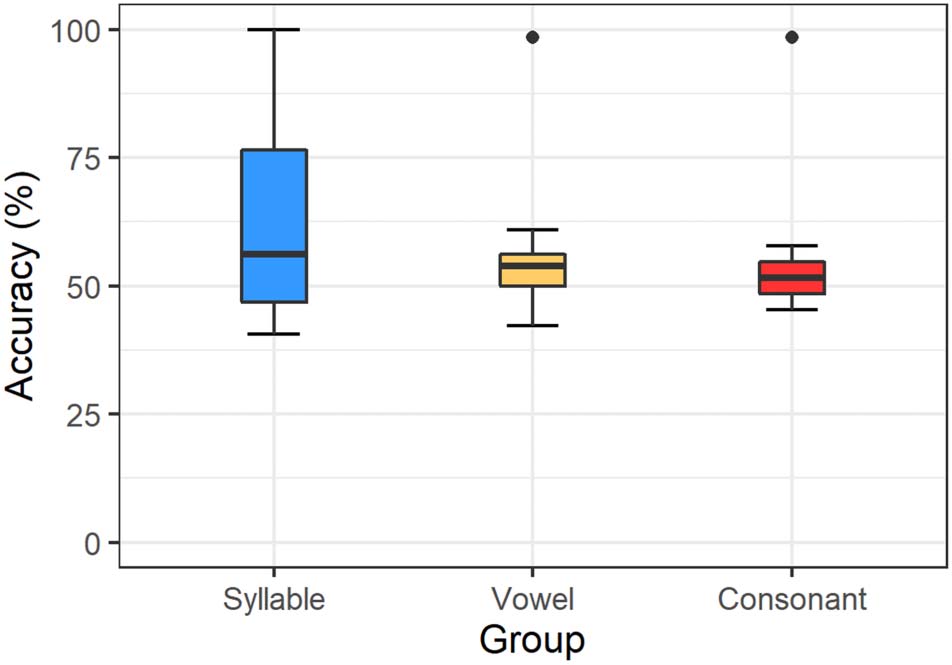
Accuracy rates were at 80.8% (SD = 14.4%) in the syllable
condition, 59.4% (SD = 16.1%) in the vowel condition, and
47.9% (SD = 14.4%) in the consonant condition. Figure 2
further illustrates the distribution of the participant aver-
ages. In the consonant condition, the median is at 50%
accuracy and the low dispersion and range of the data
(except for a few outliers) suggest little deviation from
chance level. The median response accuracy in the vowel
condition is higher with 57.8%, and both dispersion and
range are greater than for the consonants, but the two
boxes still overlap slightly and the lower quartile range
Figure 2. Average response accuracy separated by condition in
Experiment 1. The length of each box represents the interquartile range
(IQR), limited by the upper and lower quartiles; horizontal lines mark
the median; whiskers illustrate data outside the IQR (maximum: 1.5 ×
IQR); and outlier participants are indicated by dots (any data points
outside 1.5 × IQR).
1472
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
of the vowel plot includes chance level. The accuracy plot
of the syllable condition prominently differs from those of
the other two conditions. Although the interquartile
ranges are similar to the vowel condition, there is no
overlap with either of the other two conditions as median
accuracy lies at 84.4%.
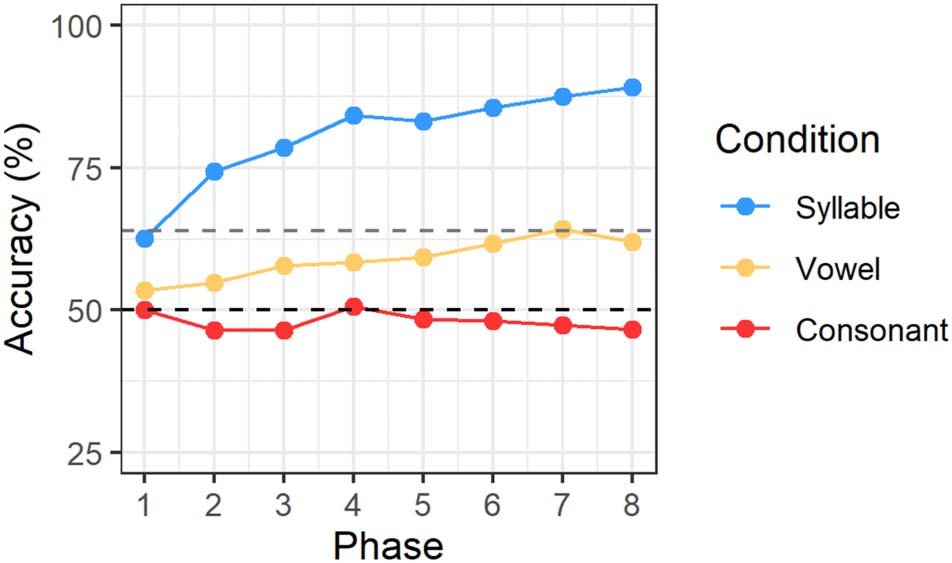
Figure 3 depicts the development of average response
accuracies by condition across test phases. It is clear that,
in the consonant condition, accuracy remained at chance
level (black dashed line) throughout the entire experi-
ment. Although accuracy seems to increase slightly across
phases in the vowel condition, it remained below the
significantly above-chance threshold (64%) up to the pen-
ultimate phase. For the syllable condition, average perfor-
mance already exceeded chance in the first test phase and
improved almost continuously from thereon, suggesting a
clear learning effect. The previously specified model
showed a significant interaction between phase and con-
dition for both the syllable and vowel conditions, as well as
a significant syllable effect, when compared against the
consonant condition (see Table 2). The factor list was
nonsignificant. To further investigate the encountered
interactions post hoc, separate GLMMs were fitted for
each condition with phase as a predictor (not centered
this time). The parameter list was excluded, but partici-
pants and items were again entered as random effects
on the intercept. After correcting for multiple compari-
sons via the Bonferroni method ( p < .008), both the
intercept (β0 = 2.02, SE = 0.31, z = 6.44, p < .0001)
and the fixed effect phase (β1 = 0.61, SE = 0.07, z =
8.40, p < .0001) were significant for the syllable condition.
Phase was also significant for the vowel condition (β1 =
0.19, SE = 0.06, z = 3.34, p < .001; but not the intercept
[β0 = 0.62, SE = 0.38, z = 1.64, p = .10]), suggesting a
phase effect on response accuracy for both the syllable
and vowel conditions. No significant effects were found
for the consonant condition (β0 = −0.12, SE = 0.40,
z = −0.31, p = .76; β1 = −0.03, SE = 0.06, z = −0.59,
p = .55). The additionally fitted intercept models for each
segmental condition revealed that the estimated intercept
Figure 3. Average response accuracies per test phase (1–8) separated
by condition in Experiment 1. The black dashed line indicates chance
level; the gray dashed line marks significantly above-chance level (64%).
Table 2. Results of the GLMM Examining the Effects of
Condition, Phase, and List on Behavioral Response Accuracy
FE
SE
z Value
p
Syllable
1.817
.390
4.655
<.001
Vowel
Phase
List B
List C
List D
Syllable × Phase
Vowel × Phase
0.635
−0.027
0.161
0.378
0.236
0.548
0.195
.388
.053
.225
.226
.232
.086
.075
1.637
−0.517
0.715
1.677
1.015
.102
.605
.475
.094
.310
6.373
<.001
2.593
<.01
Bold print indicates significant effects ( p < .05). FE = fixed effect
estimates; SE = standard error.
was significantly different from zero only for the syllable
condition (β0 = 1.89, SE = 0.29, z = 6.43, p < .0001), sug-
gesting above-chance performance in this condition
(vowels: β0 = 0.61, SE = 0.38, z = 1.64, p = .10; conso-
nants: β0 = −0.12, SE = 0.40, z = −0.30, p = .76).
Through the categorization of participants into learners
and nonlearners, we identified 25 people who successfully
learned the syllable dependency, nine of whom also
performed well in the vowel condition and three who
qualified as learners in both the consonant and syllable
conditions. Four participants were nonlearners in all of
the conditions. The models fitted to the vowel + syllable,
consonant + syllable, and syllable learner data to investi-
gate above-chance performance in the first and second
test phases could not be fitted as initially described
because of issues with singularity. We therefore reduced
the models’ complexity and fitted simple generalized lin-
ear models instead, omitting the random effects terms for
participants and items (Bates, Kliegl, Vasishth, & Baayen,
2015). Bonferroni-corrected ( p < .008) results for the esti-
mated intercepts showed that the response accuracies in
the syllable condition were already above chance in the
first phase for both syllable (n = 25; β0 = 0.50, SE =
0.16, z = 3.09, p < .002) and vowel + syllable (n = 9;
β0 = 0.78, SE = 0.26, z = 2.98, p < .003) learners. The
latter group’s response accuracy for vowels, however, only
surpassed chance in the second test phase (β0 = 0.74,
SE = 0.26, z = 2.85, p < .004). For the sake of complete-
ness, intercept models for the consonant + syllable
learners were fitted as well, although the group size was
admittedly very small (n = 3). The intercept models
revealed chance-level responses in the syllable condition
in the first test phase (β0 = −5.23e-17, SE = 4.71e-01,
z = 0, p > .008), which rose significantly above chance
only in the second test phase (β0 = 1.39, SE = 0.50, z =
2.77, p < .006). Performance for the consonant items
remained at chance in both phases (Phase 1: β0 = −1.04,
SE = 0.48, z = −2.19, p > .008; Phase 2: β0 = 1.20, SE =
0.47, z = 2.59, p > .008).
Weyers and Mueller
1473
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ERP Results
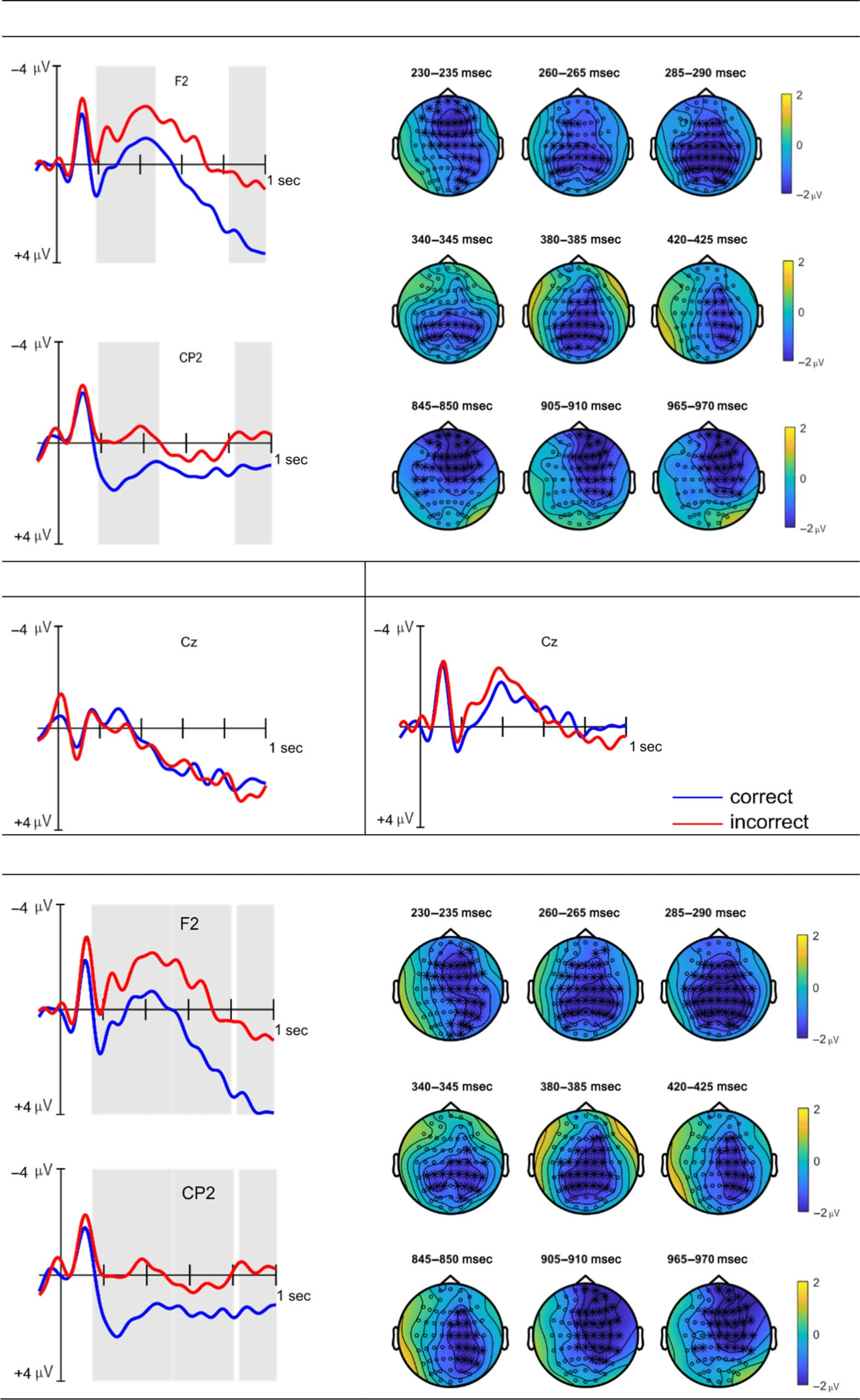
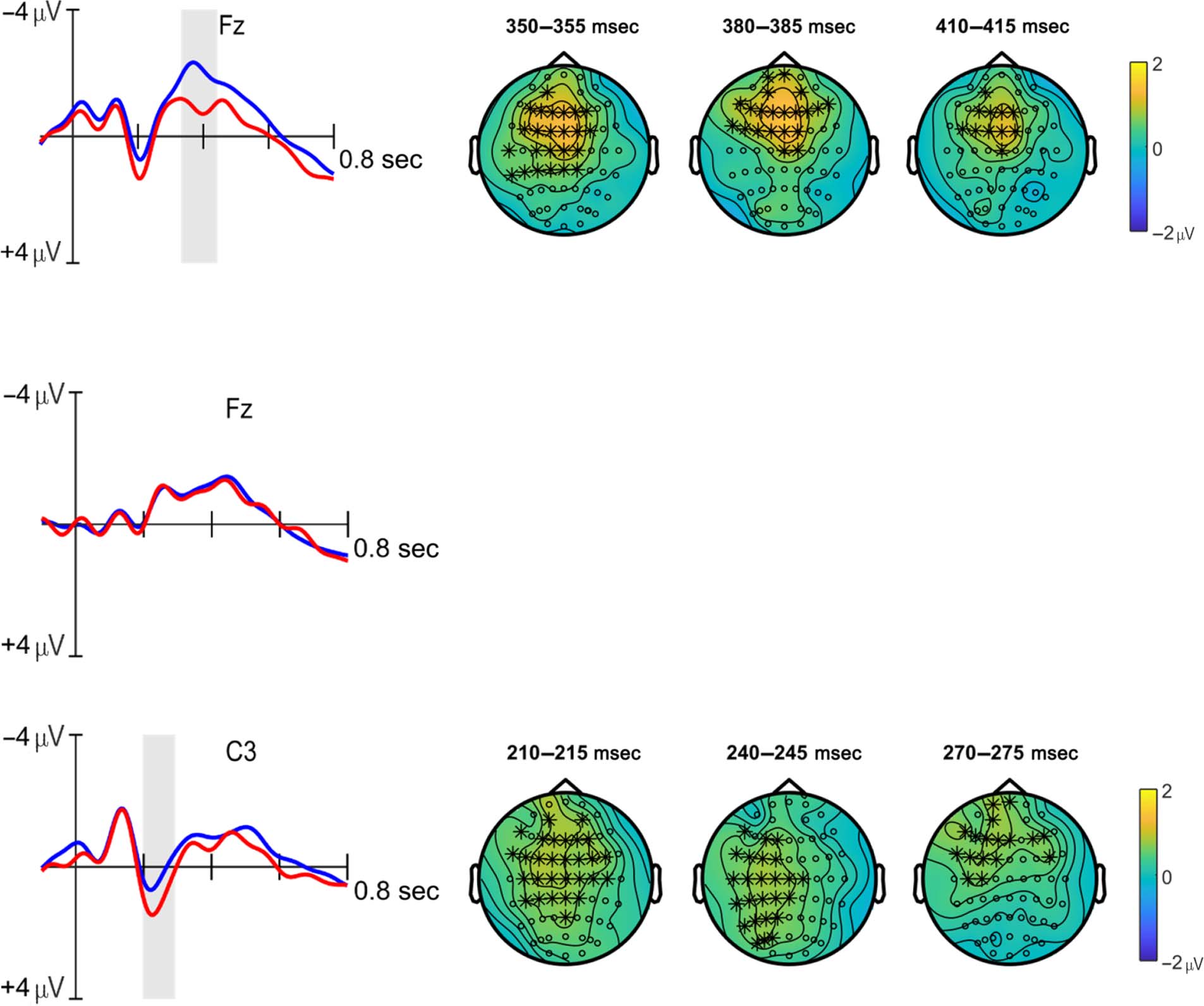
The cluster-based permutation test comparing the aver-
aged ERP responses to correct and incorrect syllable con-
dition items revealed a significant grammaticality effect
( p < .05), corresponding to two clusters. The first cluster
spanned a time window of approximately7 190–470 msec
and was observed as a negativity in the incorrect condition.
Inspection of both the ERP and topographical plots
revealed, however, that the test seemed to have grouped
two separate effects into one based on the parameters
specified above (specifically the minimum number of
neighboring channels being set to two) and a spatial
overlap between the two clusters. In particular, two
separate peaks were clearly visible at frontal electrodes
(Figure 4A, i).8 The first effect began frontally and devel-
oped into a broadly distributed effect including centro-
parietal electrodes (Figure 4A, iii), peaking around
230–240 msec. The second effect began with a centro-
parietal focus (hence the overlap), peaking between
370 and 380 msec (Figure 4A, iv), but developed into
a right-lateralized effect with a broad frontal-to-parietal
distribution. The second cluster began at around 830 msec,
lasted until the end of the epoch, and was observed as a
negativity with a fronto-central, right-lateralized distribu-
tion (Figure 4A, v).
The equivalent comparisons in the consonant and
vowel conditions did not yield any significant differ-
ences. The syllable learners (n = 25) showed a similar
significant grammaticality effect ( p < .05) as reported
for the whole group, except that here, three clusters
were identified. The first cluster began around 150 msec
and was observed to last until approximately 520 msec
with a broad distribution. The second cluster was
observed between 530 msec and roughly 800 msec as
a negativity with a fronto-central, right-lateralized distri-
bution. The third cluster, also visible as a negativity,
occurred between approximately 830 and 1000 msec,
also with a right-lateralized fronto-central focus (see
Figure 4). Cluster-based permutation tests performed
on the averaged data of the successful vowel (n = 9)
and consonant (n = 3) learners’ averaged data were
nonsignificant.
Discussion
In Experiment 1, we investigated adults’ learning of audi-
torily presented NADs from segmented streams of trisyl-
labic AXB items. Whereas the syllable condition tested
for successful learning of the syllable dependency as pre-
sented in the learning phase, the vowel and consonant
conditions aimed at assessing whether the basis of this
learned association lay with either of these smaller seg-
mental units. In other words, we tested whether, given
the availability of all three segments, participants would
memorize entire syllable frames or build representations
based on vowels or consonants, respectively.
The behavioral data showed that participants were by
far most successful at distinguishing correct and incorrect
syllable items at test. We thereby replicated previous find-
ings (Mueller et al., 2012; de Diego-Balaguer et al., 2007;
Endress & Bonatti, 2007; Peña et al., 2002). Neither of
the other two smaller segments seem to have been partic-
ularly accessible for NAD learning. If at all, correct and
incorrect exemplars of the vowel-based NAD were suc-
cessfully differentiated offline by a larger number of
learners (n = 9) than consonant-based NADs (n = 3).9
However, the comparison of these learners’ accuracy rates
for syllables and vowels in the first two test phases showed
a sequential learning pattern: Although their accuracy
rates for syllable items already exceeded chance in the first
phase, they only performed above chance level starting
from the second test phase in the vowel condition. The
specific design employed here possibly invited strategic
evaluation of (early) test phase exemplars, resulting in
the (later) application of a learned regularity that was
not built exclusively on learning phase input. The data
available from the few consonant + syllable learners
(n = 3) were less clear but tentatively suggested simul-
taneous onset of above-chance performance in both
conditions.
The EEG data supported the conclusion that partici-
pants likely built a syllable-based and not a consonant- or
vowel-based representation, because the only significant
ERP effects were found in the syllable condition. In the
latter, we encountered a broadly distributed negativity
followed by another late negativity with a fronto-central
focus in response to incorrect items. We interpreted the
first negative shift as two separate effects, namely, an
N200 with a broad distribution followed by an N400-like
effect with a typical centro-parietal topography. This com-
bination and distribution of effects has typically been
found in auditory speech processing studies investigating
semantic violations within the sentence context (Van Den
Brink, Brown, & Hagoort, 2001; Hagoort & Brown, 2000;
Connolly, Phillips, & Forbes, 1995; Connolly & Phillips,
1994; Connolly, Phillips, Stewart, & Brake, 1992; Connolly,
Stewart, & Phillips, 1990; McCallum, Farmer, & Pocock,
1984). Van Den Brink et al. (2001) specifically investigated
the differentiation of the two effects by comparing partic-
ipants’ ERP responses to sentences with semantically
incongruous (target: penseel/ brush) but phonetically
congruous final words (De schilder kleurde de details in
met een klein pensioen/The painter colored the details
with a small pension) to those elicited by semantically
and phonetically incongruous sentence-final words (De
schilder kleurde de details in met een klein doolhof/
The painter colored the details with a small labyrinth).
Whereas the N400 appeared in response to words at odds
with the semantic sentence context, the additional N200
was elicited whenever the target word also constituted a
mismatch with the expected word on the phonological
level. As a result, the N200 was interpreted as an index of
phonological processing that interacted with semantic
1474
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. ERP waveforms10 of data averaged across all participants for the syllable (A.i–A.ii), vowel (B.i), and consonant (C.i) conditions at
representative electrodes, and waveforms of data averaged across syllable learners (n = 25) only (D.i–D.ii). Significant differences between correct
and incorrect conditions are shaded in gray. Topographical plots of the syllable condition (A.iii–A.v/D.iii–D.v) show 5-msec averaged time windows
that best illustrate distribution and time course of the respective effects; significant electrodes contributing to the effects are marked with asterisks.
Weyers and Mueller
1475
context effects in the lexical selection process ( Van Den
Brink et al., 2001).
More recently and in the context of AGL tasks, the N200
has been established more generally as an attention-
dependent marker of novelty detection or sequence
matching (for a review, see Folstein & Van Petten, 2008).
Mueller et al. (2012), for instance, found an N200 effect in
response to incorrect nonadjacent syllable combinations
in their previously described oddball experiment, which
employed stimulus material very similar to the present
input. Thus, it is likely that, in our experiment, the N200
reflects two aspects: First, because the effect is attention
dependent, it shows participants’ attentional focus on
the stimulus material and specifically the relevant final unit
in the syllable condition; and second, it suggests auditory
discrimination processes that identify the final syllable of
the incorrect AXB sequences as a mismatch with the pre-
viously acquired phonemic template of the NAD. No such
effect was visible for the vowel and consonant conditions,
which is likely because here both correct and incorrect test
items included an unexpected phonemic mismatch with
the learning phase items (see Table 1).
The second negative effect, which we identified as an
N400-like component, is also in line with previous
research. Although the N400 component is typically asso-
ciated with lexical and semantic processing (e.g., Lau,
Phillips, & Poeppel, 2008), similar N400-like effects have
also been reported in a number of AGL studies (Citron
et al., 2011; Mueller et al., 2009; Mueller, Girgsdies, &
Friederici, 2008; de Diego-Balaguer et al., 2007;
Cunillera, Toro, Sebastián-Gallés, & Rodríguez-Fornells,
2006; Sanders, Newport, & Neville, 2002). These studies
have shown that the effect does not depend on the avail-
ability of semantic meaning but is also sensitive to prese-
mantic levels of processing, specifically lexical access
(i.e., the identification of familiar word forms). In word
segmentation tasks, for example, nonwords elicited an
N400-like effect, which is explained by lexical search pro-
cesses failing to match them with previously established
lexical items (de Diego-Balaguer et al., 2007; Cunillera
et al., 2006; Sanders et al., 2002). A set of NAD learning
studies (Citron et al., 2011; Mueller et al., 2009) in which
German native speakers successfully learned a morpho-
syntactic dependency from mere exposure to Italian
sentences also reported such a lexically interpreted
N400-like component in response to violations of this
NAD. On the basis of this evidence, we assume that
our participants built a representation of the AXB sylla-
ble dependency. When the final syllable failed to match
it, difficulties with lexical access resulted in the N400-
like response. This interpretation also aligns with recent
accounts that more generally assume surprisal (e.g.,
Kuperberg, 2016; Frank, Otten, Galli, & Vigliocco, 2015)
or prediction error (e.g., Bornkessel-Schlesewsky &
Schlesewsky, 2019; Rabovsky, Hansen, & McClelland,
2018) as the basis for the N400. Under this view, partici-
pants in our experiment experienced surprisal, that is, a
low match between their probabilistic prediction and the
bottom–up input, upon encountering the final syllable of
an incorrect syllable item, inciting them to update their
internal model.
Interestingly, however, we did not find a late positivity,
contrary to what has been reported in combination with
the N400-like response in some of the cited studies
(Citron et al., 2011; Mueller et al., 2009; Mueller,
Bahlmann, et al., 2008; de Diego-Balaguer et al., 2007).
Mueller, Bahlmann et al. (2008) found such a negativity–
positivity complex in response to deviant items (e.g., tile
puwo moku) after exposure to a segmented, rule-based
stream of bisyllabic nonwords encoding an item-specific
AXB dependency (e.g., tile puwo semi). The authors inter-
preted the positivity as indicating controlled structural
processes, akin to a P600 effect, which has been estab-
lished as a marker of sentence-level syntactic (and seman-
tic) integration difficulty (Friederici, 2011). Recently, it
has become subject of considerable debate whether the
P600 is actually specific to language or constitutes a more
general marker of incongruency detection for complex
structured sequences (e.g., Christiansen, Conway, & Onnis,
2012; Coulson, King, & Kutas, 1998) similar or even identi-
cal to the P300 (e.g., Sassenhagen, Schlesewsky, &
Bornkessel-Schlesewsky, 2014; Bornkessel-Schlesewsky
et al., 2011). Although this discussion is outside the scope
of this article, an aspect that is relevant to the present inves-
tigation is the notion that the process underlying the P600 is
one of (predictive) structured sequence processing, be it
domain general or domain specific (cf. Christiansen et al.,
2012, for a similar argument). If one assumed that item-
specific NAD learning in general was a somewhat structural
processing task, one might have expected a P600-like
positivity also in the context of this study. There are sev-
eral reasons this might not have been the case: First, our
stimulus material might simply not have induced such
structural processing operations. Because Mueller
et al.’s (2008; and Citron et al.’s [2011]) stimulus mate-
rial consisted of larger units encoding the dependency, it
is conceivable that their mere size was decisive in trig-
gering rather structural (syntactic-like) processing strate-
gies, whereas our smaller units more closely resembled
words and warranted lexically based operations.
Second, and apart from functional differences in pro-
cessing dependent on the type of input, the experimental
design might have played a role. Citron et al. (2011)
reported the positivity only for a design with a single
prolonged learning phase, but not for a design with
alternating learning and test phases (here, they only report
the N400-like effect), as we used in Experiment 1. De
Diego-Balaguer et al. (2007) further found the negativity–
positivity complex only when deviant items were inserted
into an oddball-like stream, but not when presented in
isolation (here, only an N400 modulation was reported).
These differences in results between research designs
might be related to the finding that both the P600
(e.g., Hahne & Friederici, 1999) and the P300 (e.g.,
1476
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Duncan-Johnson & Donchin, 1982; Duncan-Johnson &
Donchin, 1977) are sensitive to the conditional probability
of occurrence of a target (or violation). As such, these
positivities might appear primarily in paradigms that spe-
cifically manipulate the frequency of occurrence of target
items and render them highly unexpected.
Furthermore, second-language learners initially show
N400 effects in response to violations of morphosyntactic
rules at early stages of learning, which may later develop
into a more native-like N400/ P600 complex (Morgan-
Short, Sanz, Steinhauer, & Ullman, 2010; Mueller et al.,
2009; Osterhout, McLaughlin, Pitkänen, Frenck-Mestre,
& Molinaro, 2006). From the available evidence, it is
impossible to determine, however, whether this differ-
ence in effects actually suggests functional differences in
how item-specific NADs of different sizes are processed,
whether they are attributable mainly to the specific exper-
imental designs used, or whether they reflect learners’
relative “proficiency.”
The late negative effect in the syllable condition is less
tangible than the two previously classified effects, at least
from what is typically seen in AGL designs. Because of the
effect’s late appearance, relatively long duration, and
fronto-central topography, two possible candidates come
to mind. First, the reorienting negativity, which reflects
the process of reorienting one’s attention from an unex-
pected or unpredicted distractor back toward task-
relevant information, including its retrieval from working
memory (Bendixen, Schröger, Ritter, & Winkler, 2012;
Escera & Corral, 2007; Munka & Berti, 2006; Berti, Roeber,
& Schröger, 2004; Escera, Alho, Schröger, & Winkler,
2000; Schröger & Wolff, 1998). Typically, however, the
reorienting negativity has only been reported when the
deviant is employed as a behavioral distractor that is to
be ignored (e.g., tonal changes in a visual task) and not
when the distractor is part of the target stimulus set. In
the present experimental design, however, it is conceiv-
able that the incorrect syllable items acted as distractors.
After extended exposure to correct AXB sequences
(∼3 min in each learning phase), an incorrect syllable
item, although task relevant, was highly unexpected and
required participants to reallocate attention to the mainte-
nance of the correct representation.
An alternative but related explanation comes from the
working memory literature, where not only the retention
of tones (Lefebvre et al., 2013) or simple acoustic features
such as pitch (Guimond et al., 2011) or timbre (Nolden
et al., 2013) but also the retention of verbal information
(Ruchkin et al., 1997; Lang, Starr, Lang, Lindinger, &
Deecke, 1992) in auditory working memory have elicited
sustained anterior negative waves during the retention
period. These fronto-central slow waves have typically
been interpreted as reflecting active maintenance of
stimulus information in working memory, a process that
includes both the sustained (re)activation of stimulus
representations and their rehearsal (for a review, see
Kaiser, 2015). It is possible that, in the present experiment,
encountering an unexpected stimulus in the test phase
initiated such maintenance and rehearsal processes to
avoid “contamination” of the established correct target
representation.
The fact that we did not find any ERP effects across the
whole group to suggest differentiation of correct and
incorrect vowel or consonant items, respectively, indi-
cates that the given input did not induce a learning pro-
cess that is intuitively or preferentially based on either of
the two segments. The small number of participants who
successfully distinguished items in these conditions did
not provide an appropriate signal-to-noise ratio to obtain
any significant ERP effects, rendering such a comparison
uninformative. These results suggest that, with all three
segments available, the syllable was the most relevant unit
for NAD learning. The sequential learning pattern for
syllables and vowels and the absence of ERP effects in
the vowel condition further suggest that participants
likely built a syllable-based representation, which only a
subgroup of learners was able to flexibly access and apply
also to the segment-based test items.
EXPERIMENT 2
In Experiment 2, we employed a between-participant
design to test whether adults are capable of learning
NADs under conditions where only one type of segmental
information, namely, syllables, vowels, or consonants,
respectively, is available as a learning cue. To prevent
the previously explained possibly confounding effects of
strategic learning from test items, we decided to abolish
the learning phase/test phase design in favor of an oddball
paradigm. The oddball task was followed by a forced-
choice grammaticality judgment task (GJT) akin to a test
phase in the previously used design. We additionally
administered a small debriefing questionnaire after the
experiment, asking participants if they had identified a
regularity in the input and, if so, if they could spell it
out. Most importantly, they were asked to indicate when
during the experiment, approximately, they thought they
had identified the regularity, with “test phase” being one
of the possible answers. Such retrospective evaluations of
the learning process have been shown to yield valuable
information not only on what was learned but also to
which degree the learned representation was consciously
accessible (Rebuschat, 2013).
Methods
Participants
The same circumstances and requirements as in Experi-
ment 1 were applied. On the basis of similar previous
studies (e.g., Monte-Ordoño & Toro, 2017a; de Diego-
Balaguer et al., 2007), we aimed for a minimum of 20
participants entering the final analysis per group. Note
that the smaller number of participants compared with
Weyers and Mueller
1477
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Experiment 1 was justified given that, in the between-
participant oddball design, participants were exposed
to an overall larger number of deviant items. Seventy-
seven adults participated in Experiment 2, 10 of whom
had to be excluded because of technical difficulties or
high artifact rate in the EEG recording. The remaining
67 participants (15 men, 52 women) were between 18
and 33 years old (M = 21.8 years, SD = 3.10 years). Par-
ticipants were randomly assigned to one of three exper-
imental groups (SYL: n = 23, CON: n = 22, VOW: n =
22), which did not significantly differ with regard to age
or sex.
Stimuli
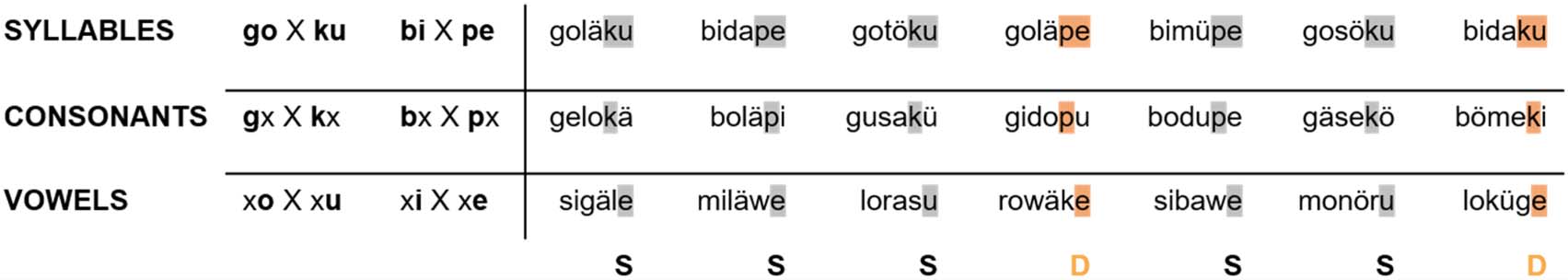
For the second experiment, a number of additional sylla-
bles were used, which had already been recorded for
Experiment 1. Four hundred forty-eight stimuli were used
for each experimental group, with a distribution of 384
standard trisyllabic sequences interspersed with 64 devi-
ants (∼14%). In the syllable group, standard and deviant
items were identical to the correct and incorrect syllable
condition items from Experiment 1, except for a different
set of 32 middle syllables (/d, l, m, n, r, s, t, w/ each
combined with /a, ä, ö, ü/). In the vowel group, standard
sequences were defined as the fixed vowel combinations
xi X xe and xo X xu, whereas all other slots in the CVCVCV
structure were filled equally often with the consonants /b,
g, k, l, m, r, s, w/ (and the vowels /a, ä, ö, ü/ in the middle
syllable) in a nonrepetitive manner. The consonant
group items were built correspondingly, filling the open
positions with /d, l, m, s/ and /a, e, i, o, u, ä, ö, ü/ (see
Figure 5).
Procedure
Four different item lists were created per group, with
the sequence of items pseudoranzomized according to
the number of constraints. The first 16 items of each list
consisted of standards for familiarization. Each deviant
item was preceded by a minimum of four and a maxi-
mum of eight standards (see Figure 5). To focus partic-
ipants’ attention on the input, they were instructed to
attentively listen to the input, determine a regularity
inherent in it, and perform a target detection task
(Mueller et al., 2012). Throughout the continuous stim-
ulus presentation, they would press a button whenever
they detected an item deviating from the regularity. The
oddball task was followed by a forced-choice GJT com-
prising 64 items (half standards and half deviants). Test
items were presented individually and, after a 900-msec
delay, had to be evaluated as adhering to or deviating
from the regularity previously identified. Stimulus pre-
sentation and all other external conditions were the
same as in Experiment 1. After the experiment, partici-
pants received the previously mentioned debriefing
questionnaire, asking whether they thought they had
learned a regularity (“Yes,” “No,” “Not sure”) and, if
“Yes” or “Not sure,” when they had learned it (“Begin-
ning,” “Middle,” “End,” “Test Phase”) and if they could
spell it out.
Data Acquisition and Preprocessing
The same software, parameters, and preprocessing steps
as in Experiment 1 were used, except that, for ERP averag-
ing, epochs from −100 to 800 msec after onset of the final
syllable (or vowel in case of the vowel condition) were cut
out from the oddball task. Data epochs were cut shorter
than in Experiment 1 to avoid contamination of the signal
by the button press during exposure. Standard items
immediately after a deviant were excluded from analysis
to avoid effects of refamiliarization. The average number
of epochs per participant entering the final analysis in each
group was as follows: 311.22 (SD = 16.14) standards and
62.04 (SD = 3.57) deviants in the syllable group, 310.95
(SD = 7.56) standards and 61.77 (SD = 1.85) deviants in
the consonant group, and 308.09 (SD = 9.40) standards
and 60.59 (SD = 2.65) deviants in the vowel group. Differ-
ences in the number of standard/deviant items between
groups were not significant.
Data Analysis
Similar to Experiment 1, accuracy scores were calculated
for each participant for the GJT items. For the responses
0 was additionally calculated
during the oddball phase, d
as an index of sensitivity. To identify learners and non-
0 > 1)
learners, responses in the target detection task (d
and/or accuracy in the GJT (response accuracy ≥ 64%,
Figure 5. Exemplary series of standard (S) and deviant (D) stimuli by group.
1478
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
indicated by a binomial test for chance response, p =
.033) were taken into account. Because we were mainly
interested in whether participants had learned the
respective dependency they had been exposed to, visi-
ble in above-chance performance in the GJT, we fitted
generalized linear mixed-effects intercept models includ-
ing a binomial link function to the accuracy data of
each group, with participants and items as random
effects. Furthermore, 1472 (syllable condition) and
1408 (consonant and vowel condition) data points were
entered into these models, respectively. p Values for
fixed effects were calculated via Wald tests (standard
for glmer in lme4). With regard to the EEG data, sepa-
rate cluster-based permutation tests with the same
parameters stated above were run for each group com-
paring responses to standard and deviant items in the
oddball task.
Table 3. Results of the GLMMs Separately Investigating
Divergence from Chance-Level Response in Each Group
FE
SE
z Value
p
Syllable
Intercept
1.037
.368
2.814
<.005
Intercept
0.254
.148
1.715
.086
Vowel
Consonant
Intercept
0.167
.117
1.432
.152
Bold print indicates significant effects ( p < .05). FE = fixed effect
estimates; SE = standard error.
Results
Behavioral Results
Average response accuracy in the GJT was at 63.6% (SD =
21.3%) in the syllable group, at 54.9% (SD = 10.8%) in the
vowel group, and at 53.6% (SD = 10.7%) in the consonant
group. The boxplots in Figure 6 provide further descrip-
tive evidence for largely chance-level responses in both
the vowel and consonant groups: Both medians are close
to 50% accuracy, and range and dispersion of the data
are low. Only one outlier per condition is visible, each
with an accuracy rate close to ceiling. Whereas the lower
quartile range of the syllable condition overlaps with the
other two boxes and the median is similar (56.3%), range
and dispersion of the accuracy rates are greater in this
Figure 6. Average response accuracy separated by groups in
Experiment 2. The length of each box represents the interquartile range
(IQR), limited by the upper and lower quartiles; horizontal lines mark
the median; whiskers illustrate data outside the IQR (maximum: 1.5 ×
IQR); and outlier participants are indicated by dots (any data points
outside 1.5 × IQR).
group and the data are visibly skewed toward the upper
percentages.
The fitted intercept models revealed that the estimated
intercept was significantly different from zero only for the
syllable group, whereas the other two models did not pro-
vide evidence for above-chance performance in the vowel
and consonant groups (see Table 3). We identified seven
learners in the syllable group (detection rate: 48.2%, SD =
32.9%; grammaticality judgment: 92.4%, SD = 12.6%), one
in the vowel group (detection rate: 85.9%; grammaticality
judgment: 98.4%), and one in the consonant group (detec-
tion rate: 21.9%; grammaticality judgment: 98.4%). The
debriefing questionnaire revealed that five of the seven
syllable group learners were able to correctly spell out
the syllable dependency, as were the individual learners
in the vowel and consonant groups, respectively. The syl-
lable learners and the vowel learner further believed to
have learned the regularity “at the beginning” or “in the
middle” of the experiment, whereas the consonant learner
perceived detection to have happened only “at the end” of
the experiment.
ERP Results
The comparison between the evoked responses of
standard and deviant items in the syllable group indicated
a significant difference ( p < .05). The difference was
manifested in a fronto-centrally distributed positivity,
based on a cluster between approximately 335 and
440 msec (Figure 7A, i–ii).10 The cluster-based test yielded
no significant differences in the vowel group (Figure 7B, i)
but exposed a significant difference between conditions
in the consonant group ( p < .05), corresponding to a
fronto-central to centro-parietal, slightly left-lateralized
positive cluster between around 200 and 290 msec
(Figure 7C, i–ii).
Weyers and Mueller
1479
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. ERP waveforms11 of data averaged across all participants of the syllable (A.i), vowel (B.i), and consonant (C.i) groups at the representative
electrodes Fz and C3. Significant differences between standard and deviant condition are shaded in gray. Topographical plots of the syllable (A.ii) and
consonant (C.ii) groups show 5-msec averaged time windows that best illustrate distribution and time course of the respective effects; significant
electrodes contributing to the effects are marked with asterisks.
DISCUSSION
Experiment 2 aimed at assessing adult listeners’ ability to
detect and evaluate NADs between specific syllables,
vowels, or consonants online. In both behavioral and neu-
rophysiological results, the syllable again clearly emerged
as the unit from which adults discerned the dependency
most successfully.11 We thereby closely replicated Mueller
et al.’s (2012) findings and extended them by showing
that explicit instructions specifying which units are rele-
vant to the regularity (the authors hinted at the order of
the syllables) are not necessary for successful learning.
The corresponding EEG data further highlighted the role
of the syllable unit. We interpreted the encountered pos-
itivity in response to deviant syllable items as a P3 effect,
which is typical in oddball designs and indicates
discrimination of the infrequently occurring deviant
(target) from the frequently occurring standard (for a
review, see Polich, 2007). It mirrored the effect Mueller
et al. (2012) found and, similarly, the effect reported by
Monte-Ordoño and Toro (2017b) in an oddball paradigm
in response to stimuli that violated a syllable-based ABB
repetition rule.
The P3 is often classified as P3a or P3b depending on its
distribution. Whereas the former shows a frontal distribu-
tion and is suggestive of a stimulus-driven (unconscious)
attention switch to previously unattended material or
stimulus features (Escera & Corral, 2007; Debener,
Kranczioch, Herrmann, & Engel, 2002; Escera, Alho,
Winkler, & Näätänen, 1998), the latter is centro-parietally
distributed and indexes task-relevant conscious attention
to stimuli and subsequent working memory updating
1480
Journal of Cognitive Neuroscience
Volume 34, Number 8
processes (Ferdinand, Mecklinger, & Kray, 2008; Sergent,
Baillet, & Dehaene, 2005; Wetter, Polich, & Murphy, 2004).
The fact that we found a fronto-central distribution of
the P3 suggests a mixture of P3a and P3b probably attrib-
utable to averaging across the entire participant group, of
which only seven people consciously discriminated the
items, resulting only in a slight shift of the effect toward
central electrode sites. Nevertheless, the effect was
significant across the entire syllable group, suggesting that
even those who did not demonstrate behavioral evidence
of learning possibly passively recognized the depen-
dency violation.
When it comes to the smaller segmental level, we found
no evidence, either behavioral or neurophysiological, that
vowels are accessible for item-based NAD learning. Only a
single participant performed well at test, and there were
no significant ERP effects across the whole group that
would suggest any kind of differentiation between
standards and deviants. Because the experimental design
prevented strategic learning from test items, these results
support our suspicion that the behavioral learning effects
seen in the vowel condition in Experiment 1 were largely
provoked by the experimental design.
In the consonant group, we also only found a single par-
ticipant who exhibited behavioral evidence of learning.
Nonetheless, the ERPs computed for the entire group
showed a significant P2 effect in response to deviants com-
pared to standards—a component that has been found to
signal selective attention to acoustic stimulus change and
feature detection in a variety of auditory tasks (see e.g.,
Paulmann, Bleichner, & Kotz, 2013; Cunillera et al., 2006;
Yingling & Nethercut, 1983). Cunillera et al. (2006), for
example, encountered a P2 effect during segmentation
of stressed words from a continuous stream when com-
pared to the same unstressed word stream. Although this
is evidence for an emergent dissociation, the lack of a con-
current behavioral response or P3(b) effect similar to the
syllable group suggests that whichever consonant-based
representation participants might have built, it was not
accessible to be translated into an offline response. One
might assume a less sophisticated NAD representation
than in the syllable group, for instance, one that rested
merely on phonological memory of the perceptual simi-
larity of the paired consonants.
In any case, the finding confirmed the tentatively formu-
lated expectation that the use of item-based dependencies
between specific units might result in a slight advantage
for consonants over vowels in NAD learning because of
their previously shown prevalence in word identification
and lexical selection (Delle Luche et al., 2014; Havy
et al., 2014; Carreiras, Dunabeitia, et al., 2009; Carreiras,
Gillon-Dowens, et al., 2009; New et al., 2008; Cutler
et al., 2000), where item specificity is of importance. In
other words, the fact that the dependency relationship
between consonants was more salient to adults than that
between vowels is suggestive of lexical processes having
been at play during learning.
GENERAL DISCUSSION
The goal of this study was to compare syllables, conso-
nants, and vowels as carriers of item-specific NADs
and to identify possible differences between the three
segments with regard to online processing and offline
learning success for NADs. We used an artificial grammar
consisting of trisyllabic sequences from which a depen-
dency relationship between specific units had to be
learned. In Experiment 1, we reported evidence that
adults preferentially learn the syllable-based dependency
when all three segments are available for NAD learning
and likely build a syllable-based representation of the
input. We found no evidence that would suggest that
item-specific NAD learning was biased or guided by the
smaller segmental level. From Experiment 1 alone, it
remained unclear whether it is the syllable per se that
receives a special role in NAD learning or whether adults
simply focused on the largest informative unit. We further
investigated this in Experiment 2, where three separate
groups of participants were exposed to material contain-
ing either a syllable-, consonant-, or vowel-based depen-
dency that could be learned. Again, the syllable clearly
emerged as the unit from which the dependency was
learned most successfully. When it was not available and
the only units informative to the nonadjacency relation-
ship were consonants or vowels, respectively, there was
little evidence for successful learning. We therefore con-
clude that it was not relative informativeness that triggered
syllable-based NAD learning in Experiment 1 but rather a
more general (attentional) preference for the syllable unit
over the two smaller segments.
A caveat to be considered in this regard is that, in both of
our experiments, the syllables within the trisyllabic AXB
units were separated by 50-msec pauses, possibly resulting
in a perceptual bias toward the syllable unit. Recent
oscillation-based approaches to natural speech processing
have shown, however, that neuronal oscillations auto-
matically track the dynamics of continuous speech input
at the syllable rate even without such artificially inserted
segmentation markers (Poeppel & Assaneo, 2020;
Giraud & Poeppel, 2012). Although these oscillator
models cannot explain how isolated subword units such
as the ones used here are decoded, they do highlight a
general possible perceptual advantage for syllables in
competent language users. Whether this advantage is
based on a signal-driven, bottom–up mechanism or
whether it constitutes a learned, top–down process is
subject of considerable debate and is beyond the scope
of this article (for a discussion, see, e.g., Räsänen et al.,
2018; Giraud & Poeppel, 2012).
With regard to vowels as carriers of NADs in Experi-
ment 2, we found no evidence for a “perceptual similarity
effect” as hypothesized initially: It seems that nonre-
peated but phonetically close vowels do not suffice to
induce the learning advantage vowels have tentatively
demonstrated over consonants in the context of studies
Weyers and Mueller
1481
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
using repetition rules (Monte-Ordoño & Toro, 2017a;
Toro, Nespor, et al., 2008; Toro, Shukla, et al., 2008).
As such, vowels do not seem to be particularly accessible
for the identification and memorization of item-specific
dependency relations when they are the only informative
segment. What is intriguing, however, is that at least a
small group of participants in Experiment 1 was able to
correctly evaluate the vowel test items by identifying
them as subcomponents of the syllable dependency. That
is, once a syllable-based representation of the NAD has
been built, it seems that a matching vowel-based depen-
dency is recognized more easily than a matching
consonant-based dependency. The question remains
whether this is because of mere perceptual differences
in the two segments (i.e., relative saliency of vowels) or
because of a top–down mechanism that identifies such
“word”-internal regularities and preferentially operates
across vowels. One might speculate that if working mem-
ory and specifically articulatory rehearsal processes play a
role for such explicit, strategic consideration of the stim-
ulus material, it might be easier to rehearse and connect
individual vowels than consonants. Single vowels can be
full syllables in some cases (e.g., in a word like “a” or an
exclamation like “uh”) and thus serve as independent
production units, whereas consonants typically occur in
conjunction with vowels (e.g., even during the produc-
tion of individual consonants in spelling out loud).
When it comes to consonants, the ERP evidence for
deviant processing (at least at an acoustic level) found in
Experiment 2 further substantiates the superior role of
consonants as “identifiers,” as formulated, for instance,
in the CV hypothesis (Nespor et al., 2003). Together with
the lexically related ERP effects found for syllables in
Experiment 1, these results additionally show that the pro-
cessing of item-specific NADs seems to prompt lexical
rather than (morphosyntactic-like) structure-related pro-
cesses. The question remains whether the given ERP pat-
terns were because of the “low proficiency” attained by the
learners throughout the experiment, that is, similar to
what Morgan-Short et al. (2010) and Osterhout et al.
(2006) find for second-language learners at early stages
of morphosyntactic learning, or whether they are attribut-
able to the nature of the stimulus material. A potential
methodological issue to be mentioned in this context is
the fact that consonants always preceded vowels in our
stimulus material. In combination with the mentioned
pauses between units, this might have provided conso-
nants with a positional advantage and increased their
relative salience. To exclude this possibility, future investi-
gations could directly compare CVXCV and VCXVC
structures.
Future research could further investigate which features
of the syllable specifically facilitate the extraction of depen-
dencies in speech. Are specific acoustic properties of the
syllable responsible, for example, its duration (providing
sufficient time for working memory processes) or
spectro-temporal complexity (providing a unique and rich
representation for memory), or is it its association with
articulatory gestures (e.g., in the mental syllabary) that
supports inner rehearsal processes during NAD learning?
Overall, it seems that, at the smaller segmental level,
vowels are susceptible to conscious, strategic manipula-
tions in working memory, whereas the processing of rela-
tionships between consonants instead induces implicit
learning processes. Thus, we can conclude that, on the
basis of the present evidence, syllables, consonants, and
vowels clearly do not constitute equally suitable computa-
tional units for NAD learning. Indeed, we were able to
show that syllables are by far the most accessible unit
for the learning of such relatively local, item-specific
dependencies.
Reprint requests should be sent to Ivonne Weyers, Department of
Linguistics, University of Vienna, Sensengasse 3A, Vienna 1090,
Austria, or via e-mail: ivonne.weyers@univie.ac.at.
Funding Information
Deutsche Forschungsgemeinschaft (https://dx.doi.org/10
.13039/501100001659), grant number: MU 3112/3-1.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender
identification of first author/ last author) publishing in
the Journal of Cognitive Neuroscience ( JoCN ) during this
period were M(an)/M = .407, W(oman)/M = .32, M/ W =
.115, and W/ W = .159, the comparable proportions for the
articles that these authorship teams cited were M/M =
.549, W/M = .257, M/ W = .109, and W/ W = .085 (Postle
and Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN
encourages all authors to consider gender balance explic-
itly when selecting which articles to cite and gives them
the opportunity to report their article’s gender citation
balance.
Notes
1.
In all of the cited studies, the authors used only eight
(four: Toro, Shukla, et al. [2008]) pairs of test items in the gen-
eralization test, which makes their average correct response
rates tentative indicators of learning at best. The authors
reported average response accuracies of 66.6% (SD = 11.2;
Toro, Nespor, et al., 2008), 61.6% (SD = 12.9; Toro, Shukla,
et al., 2008), and 61.18% (SD = 22.78; Monte-Ordoño & Toro,
2017a) for NAD generalizations across vowels, that is, as low as
five hits out of eight trials. A simple binomial test reveals that
the cumulative probability of P(X ≥ 5) = .363, suggesting there
is a 36% probability that the average result was obtained by
chance.
2. To test for learning, previous behavioral studies have typi-
cally used a two-alternative forced-choice task (e.g., Peña et al.,
2002; Saffran, Newport, & Aslin, 1996). This task, in which two
1482
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
items are presented to the participant sequentially, is problem-
atic when simultaneously using EEG, however, because it addi-
tionally taxes working memory. Therefore, we chose a standard
ERP violation paradigm, in which items are presented and eval-
uated individually.
3. Note that other similar EEG studies have sometimes used
novel phonemes in their violation items (e.g., Monte-Ordoño &
Toro, 2017a, 2017b) and subsequently compared ERP
responses to phonemically very different sets of correct (e.g.,
fefuku) and incorrect (e.g., mimomi) exemplars. Because this
can lead to confounding phoneme change effects, we ensured
our stimulus design would allow comparing responses to the
same phonemes in the relevant positions (e.g., pi and ko in
the consonant condition; see Table 1).
4. Even an exactly equal distribution of items per segmental
condition would likely result in a perceived imbalance of cor-
rect and incorrect items by the participant, for example, assum-
ing they learn the vowel dependency and correctly evaluate the
vowel and syllable items but reject all consonant condition
items.
ICA was run on 1-Hz high-pass filtered (−6 dB, 1 Hz, 930)
5.
data to improve decomposition performance ( Winkler,
Debener, Muller, & Tangermann, 2015). The resulting ICA
weights were then applied to the 0.1-Hz filtered data sets.
6. Theoretically even “vowel + consonant + syllable”
learners, although this could arguably be evidence for a
syllable-based representation and subsequent access to its seg-
mental subcomponents rather than segment-specific function-
alities.
7. Note that it is in the nature of the cluster-based permuta-
tion method that the reported cluster T-statistic does not allow
any inferences with regard to the specific temporal or spatial
distribution of the identified cluster(s), since there is no error
rate control for the inclusion of individual sample statistics in a
specific cluster. The spatiotemporal characteristics of the
effects established by the test are, however, likely to be highly
correlated with those of the true effect (Sassenhagen &
Draschkow, 2019; Groppe et al., 2011; Maris & Oostenveld,
2007).
8. An additional 10-Hz low-pass filter (−6 dB, 10, 620) was
applied to the averaged data exclusively for plotting to improve
visibility.
In this context, it should be mentioned that a few partici-
9.
pants actually performed below chance level in one of the two
conditions (vowel or consonant; see Figure 1) but show high
response accuracy in the respective other condition. Appar-
ently, these participants (n = 3 for vowels, n = 2 for conso-
nants) established (perceptual) classes of elements ( Wilson
et al., 2018), evaluating any combination of o/u and i/e (or b/
p, g/k) as correct, regardless of the vowels’ (or consonants’)
specific positions in the items. Therefore, they evaluated the
incorrect consonant items bu X ko/ge X pi (vowel items pi X
bu/ko X ge) as correct because the vowel pairings were intact
but rejected their correct counterparts bu X pi/ge X ko (pi X
ge/ko X bu for vowels), because these violated the pairings
they had learned.
10. An additional 10-Hz low-pass filter (−6 dB, 10, 620) was
applied to the averaged data exclusively for plotting of the ERP
graphs to improve readability.
11. On the basis of the information provided in the debriefing
questionnaire after Experiment 2, it seems that at least some
participants (SYL: n = 5, CON: n = 6, VOW: n = 2) were dis-
tracted by the use of umlaut in the middle positions irrelevant
for the NAD and pressed a button whenever no umlaut
appeared in a stimulus. This is in line with the finding that, ini-
tially, adults primarily identify the distributional properties of an
input and only extract the conditional regularities potentially at
odds with them after extended exposure (Endress & Bonatti,
2016). Even so, the average detection rates of the remaining
participants (consciously) undisturbed by the umlaut did not
exceed chance in either the vowel or consonant group.
REFERENCES
Abrams, D. A., Nicol, T., Zecker, S., & Kraus, N. (2009).
Abnormal cortical processing of the syllable rate of speech
in poor readers. Journal of Neuroscience, 29, 7686–7693.
https://doi.org/10.1523/JNEUROSCI.5242-08.2009, PubMed:
19535580
Bates, D. M., Kliegl, R., Vasishth, S., & Baayen, H. (2015a).
Parsimonious mixed models. ArXiv. https://doi.org/10.48550
/arXiv.1506.04967
Bates, D. M., Mächler, M., Bolker, B. M., & Walker, S. C. (2015b).
Fitting linear mixed-effects models using lme4. Journal of
Statistical Software, 67, 1–48. https://doi.org/10.18637/jss
.v067.i01
Batterink, L. (2020). Syllables in sync form a link: Neural
phase-locking reflects word knowledge during
language learning. Journal of Cognitive Neuroscience,
32, 1735–1748. https://doi.org/10.1162/jocn_a_01581,
PubMed: 32427066
Bendixen, A., Schröger, E., Ritter, W., & Winkler, I. (2012).
Regularity extraction from non-adjacent sounds. Frontiers in
Psychology, 3, 143. https://doi.org/10.3389/fpsyg.2012.00143,
PubMed: 22661959
Berti, S., Roeber, U., & Schröger, E. (2004). Bottom–up influences
on working memory: Behavioral and electrophysiological
distraction varies with distractor strength. Experimental
Psychology, 51, 249–257. https://doi.org/10.1027/1618-3169.51
.4.249
Bertoncini, J., & Mehler, J. (1981). Syllables as units in infant
speech perception. Infant Behavior and Development, 4,
247–260. https://doi.org/10.1016/S0163-6383(81)80027-6
Boatman, D., Hall, C., Goldstein, M. H., Lesser, R., & Gordon, B.
(1997). Neuroperceptual differences in consonant and vowel
discrimination: As revealed by direct cortical electrical
interference. Cortex, 33, 83–98. https://doi.org/10.1016/S0010
-9452(97)80006-8
Bonatti, L. L., Peña, M., Nespor, M., & Mehler, J. (2005). Linguistic
constraints on statistical computations. Psychological Science,
16, 451–459. https://doi.org/10.1111/j.0956-7976.2005.01556.x,
PubMed: 15943671
Bornkessel-Schlesewsky, I., Kretzschmar, F., Tune, S., Wang, L.,
Genç, S., Philipp, M., et al. (2011). Think globally: Cross-
linguistic variation in electrophysiological activity during
sentence comprehension. Brain and Language, 117,
133–152. https://doi.org/10.1016/j.bandl.2010.09.010,
PubMed: 20970843
Bornkessel-Schlesewsky, I., & Schlesewsky, M. (2019). Toward
a neurobiologically plausible model of language-related,
negative event-related potentials. Frontiers in Psychology,
10, 298. https://doi.org/10.3389/fpsyg.2019.00298, PubMed:
30846950
Caramazza, A., Chialant, D., Capasso, R., & Miceli, G. (2000).
Separable processing of consonants and vowels. Nature,
403, 428–430. https://doi.org/10.1038/35000206, PubMed:
10667794
Carreiras, M., Dunabeitia, J. A., & Molinaro, N. (2009a).
Consonants and vowels contribute differently to visual word
recognition: ERPs of relative position priming. Cerebral
Cortex, 19, 2659–2670. https://doi.org/10.1093/cercor
/bhp019, PubMed: 19273459
Carreiras, M., Gillon-Dowens, M., Vergara, M., & Perea, M.
(2009b). Are vowels and consonants processed differently?
Event-related potential evidence with a delayed letter
Weyers and Mueller
1483
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
paradigm. Journal of Cognitive Neuroscience, 21, 275–288.
https://doi.org/10.1162/jocn.2008.21023, PubMed: 18510451
Carreiras, M., & Perea, M. (2004). Naming pseudowords in
Spanish: Effects of syllable frequency. Brain and Language,
90, 393–400. https://doi.org/10.1016/j.bandl.2003.12.003,
PubMed: 15172555
Carreiras, M., & Price, C. J. (2008). Brain activation for
consonants and vowels. Cerebral Cortex, 18, 1727–1735.
https://doi.org/10.1093/cercor/bhm202, PubMed: 18234690
Carreiras, M., Vergara, M., & Perea, M. (2007). ERP correlates of
transposed-letter similarity effects: Are consonants processed
differently from vowels? Neuroscience Letters, 419, 219–224.
https://doi.org/10.1016/j.neulet.2007.04.053, PubMed: 17507160
Choi, D., Batterink, L. J., Black, A. K., Paller, K. A., & Werker, J. F.
(2020). Preverbal infants discover statistical word patterns at
similar rates as adults: Evidence from neural entrainment.
Psychological Science, 31, 1161–1173. https://doi.org/10.1177
/0956797620933237, PubMed: 32865487
Cholin, J. (2008). The mental syllabary in speech production: An
integration of different approaches and domains. Aphasiology,
22, 1127–1141. https://doi.org/10.1080/02687030701820352
Cholin, J., Dell, G. S., & Levelt, W. J. M. (2011). Planning and
articulation in incremental word production: Syllable-
frequency effects in English. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 37, 109–122.
https://doi.org/10.1037/a0021322, PubMed: 21244111
Cholin, J., Levelt, W. J. M., & Schiller, N. O. (2006). Effects of
syllable frequency in speech production. Cognition, 99,
205–235. https://doi.org/10.1016/j.cognition.2005.01.009,
PubMed: 15939415
Christiansen, M. H., Conway, C. M., & Onnis, L. (2012). Similar
neural correlates for language and sequential learning:
Evidence from event-related brain potentials. Language and
Cognitive Processes, 27, 231–256. https://doi.org/10.1080
/01690965.2011.606666, PubMed: 23678205
Citron, F. M. M., Oberecker, R., Friederici, A. D., & Mueller, J. L.
(2011). Mass counts: ERP correlates of non-adjacent
dependency learning under different exposure conditions.
Neuroscience Letters, 487, 282–286. https://doi.org/10.1016/j
.neulet.2010.10.038, PubMed: 20971159
Connolly, J. F., & Phillips, N. A. (1994). Event-related potential
components reflect phonological and semantic processing of
the terminal word of spoken sentences. Journal of Cognitive
Neuroscience, 6, 256–266. https://doi.org/10.1162/jocn.1994
.6.3.256, PubMed: 23964975
Connolly, J. F., Phillips, N. A., & Forbes, K. A. K. (1995). The
effects of phonological and semantic features of sentence-
ending words on visual event-related brain potentials.
Electroencephalography and Clinical Neurophysiology, 94,
276–287. https://doi.org/10.1016/0013-4694(95)98479-R,
PubMed: 7537200
Connolly, J. F., Phillips, N. A., Stewart, S. H., & Brake, W. G.
(1992). Event-related potential sensitivity to acoustic and
semantic properties of terminal words in sentences. Brain
and Language, 43, 1–18. https://doi.org/10.1016/0093-934X,
PubMed: 1643505
Connolly, J. F., Stewart, S. H., & Phillips, N. A. (1990). The
effects of processing requirements on neurophysiological
responses to spoken sentences. Brain and Language, 39,
302–318. https://doi.org/10.1016/0093-934X(90)90016-A,
PubMed: 2224497
Coulson, S., King, J. W., & Kutas, M. (1998). Expect the
unexpected: Event-related brain response to morphosyntactic
violations. Language and Cognitive Processes, 13, 21–58.
https://doi.org/10.1080/016909698386582
Creel, S. C., Newport, E. L., & Aslin, R. N. (2004). Distant
melodies: Statistical learning of nonadjacent dependencies
in tone sequences. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 30, 1119–1130. https://
doi.org/10.1037/0278-7393.30.5.1119, PubMed: 15355140
Culbertson, J., Koulaguina, E., Gonzalez-Gomez, N., Legendre,
G., & Nazzi, T. (2016). Developing knowledge of nonadjacent
dependencies. Developmental Psychology, 52, 2174–2183.
https://doi.org/10.1037/dev0000246, PubMed: 27893252
Cunillera, T., Toro, J. M., Sebastián-Gallés, N., & Rodríguez-
Fornells, A. (2006). The effects of stress and statistical cues on
continuous speech segmentation: An event-related brain
potential study. Brain Research, 1123, 168–178. https://doi
.org/10.1016/j.brainres.2006.09.046, PubMed: 17064672
Cutler, A., Sebastian-Galles, N., Soler-Vilageliu, O., & Van
Ooijen, B. (2000). Constraints of vowels and consonants on
lexical selection: Cross-linguistic comparisons. Memory and
Cognition, 28, 746–755. https://doi.org/10.3758/BF03198409,
PubMed: 10983448
Debener, S., Kranczioch, C., Herrmann, C. S., & Engel, A. K.
(2002). Auditory novelty oddball allows reliable distinction
of top–down and bottom–up processes of attention.
International Journal of Psychophysiology, 46, 77–84.
https://doi.org/10.1016/S0167-8760(02)00072-7, PubMed:
12374648
Dell, G. S. (1986). A spreading-activation theory of retrieval in
sentence production. Psychological Review, 93, 283–321.
https://doi.org/10.1037/0033-295X.93.3.283, PubMed: 3749399
Dell, G. S. (1988). The retrieval of phonological forms in
production: Tests of predictions from a connectionist model.
Journal of Memory and Language, 27, 124–142. https://doi
.org/10.1016/0749-596X(88)90070-8, PubMed: 3749399
Delle Luche, C., Poltrock, S., Goslin, J., New, B., Floccia, C., &
Nazzi, T. (2014). Differential processing of consonants and
vowels in the auditory modality: A cross-linguistic study.
Journal of Memory and Language, 72, 1–15. https://doi.org
/10.1016/j.jml.2013.12.001
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
de Diego-Balaguer, R., Toro, J. M., Rodriguez-Fornells, A., &
Bachoud-Lévi, A. C. (2007). Different neurophysiological
mechanisms underlying word and rule extraction from
speech. PLoS One, 2, e1175. https://doi.org/10.1371/journal
.pone.0001175, PubMed: 18000546
Ding, N., Melloni, L., Zhang, H., Tian, X., & Poeppel, D. (2016).
Cortical tracking of hierarchical linguistic structures in
connected speech. Nature Neuroscience, 19, 158–164.
https://doi.org/10.1038/nn.4186, PubMed: 26642090
Ding, N., & Simon, J. Z. (2014). Cortical entrainment to
continuous speech: Functional roles and interpretations.
Frontiers in Human Neuroscience, 8, 311. https://doi.org/10
.3389/fnhum.2014.00311, PubMed: 24904354
Duncan-Johnson, C. C., & Donchin, E. (1977). On quantifying
surprise: The variation of event-related potentials with
subjective probability. Psychophysiology, 14, 456–467. https://
doi.org/10.1111/j.1469-8986.1977.tb01312.x, PubMed: 905483
Duncan-Johnson, C. C., & Donchin, E. (1982). The P300
component of the event-related brain potential as an index of
information processing. Biological Psychology, 14, 1–52.
https://doi.org/10.1016/0301-0511(82)90016-3
Endress, A. D., & Bonatti, L. L. (2007). Rapid learning of syllable
classes from a perceptually continuous speech stream.
Cognition, 105, 247–299. https://doi.org/10.1016/j.cognition
.2006.09.010, PubMed: 17083927
Endress, A. D., & Bonatti, L. L. (2016). Words, rules, and
mechanisms of language acquisition. Wiley Interdisciplinary
Reviews: Cognitive Science, 7, 19–35. https://doi.org/10.1002
/wcs.1376, PubMed: 26683248
1484
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Endress, A. D., Nespor, M., & Mehler, J. (2009). Perceptual and
memory constraints on language acquisition. Trends in
Cognitive Sciences, 13, 348–353. https://doi.org/10.1016/j.tics
.2009.05.005, PubMed: 19647474
Escera, C., Alho, K., Schröger, E., & Winkler, I. (2000).
Involuntary attention and distractibility as evaluated with
event-related brain potentials. Audiology and Neuro-Otology,
5, 151–166. https://doi.org/10.1159/000013877, PubMed:
10859410
Escera, C., Alho, K., Winkler, I., & Näätänen, R. (1998). Neural
mechanisms of involuntary attention to acoustic novelty and
change. Journal of Cognitive Neuroscience, 10, 590–604.
https://doi.org/10.1162/089892998562997, PubMed: 9802992
Escera, C., & Corral, M. J. (2007). Role of mismatch negativity
and novelty-P3 in involuntary auditory attention. Journal of
Psychophysiology, 21, 251–264. https://doi.org/10.1027/0269
-8803.21.34.251, PubMed: 10859410
Ferdinand, N. K., Mecklinger, A., & Kray, J. (2008). Error and
deviance processing in implicit and explicit sequence
learning. Journal of Cognitive Neuroscience, 20, 629–642.
https://doi.org/10.1162/jocn.2008.20046, PubMed: 18052785
Folstein, J. R., & Van Petten, C. (2008). Influence of cognitive
control and mismatch on the N2 component of the ERP: A
review. Psychophysiology, 45, 152–170. https://doi.org/10
.1111/j.1469-8986.2007.00602.x, PubMed: 17850238
Frank, S. L., Otten, L. J., Galli, G., & Vigliocco, G. (2015). The
ERP response to the amount of information conveyed by
words in sentences. Brain and Language, 140, 1–11. https://
doi.org/10.1016/j.bandl.2014.10.006, PubMed: 25461915
Friederici, A. D. (2011). The brain basis of language processing:
From structure to function. Physiological Reviews, 91,
1357–1392. https://doi.org/10.1152/physrev.00006.2011,
PubMed: 22013214
Frost, R. L. A., & Monaghan, P. (2016). Simultaneous segmentation
and generalisation of non-adjacent dependencies from
continuous speech. Cognition, 147, 70–74. https://doi.org/10
.1016/j.cognition.2015.11.010, PubMed: 26638049
Giraud, A. L., & Poeppel, D. (2012). Cortical oscillations and
speech processing: Emerging computational principles and
operations. Nature Neuroscience, 15, 511–517. https://doi
.org/10.1038/nn.3063, PubMed: 22426255
Gómez, R. L. (2002). Variability and detection of invariant
structure. Psychological Science, 13, 431–436. https://doi.org
/10.1111/1467-9280.00476, PubMed: 12219809
Gómez, R. L., & Maye, J. (2005). The developmental trajectory
of nonadjacent dependency learning. Infancy, 7, 183–206.
https://doi.org/10.1207/s15327078in0702_4, PubMed:
33430549
Goswami, U. (2011). A temporal sampling framework for
developmental dyslexia. Trends in Cognitive Sciences, 15,
3–10. https://doi.org/10.1016/j.tics.2010.10.001, PubMed:
21093350
Grama, I. C., Kerkhoff, A., & Wijnen, F. (2016). Gleaning
structure from sound: The role of prosodic contrast
in learning non-adjacent dependencies. Journal of
Psycholinguistic Research, 45, 1427–1449. https://doi.org/10
.1007/s10936-016-9412-8, PubMed: 26861215
Groppe, D. M., Urbach, T. P., & Kutas, M. (2011). Mass
univariate analysis of event-related brain potentials/fields I: A
critical tutorial review. Psychophysiology, 48, 1711–1725.
https://doi.org/10.1111/j.1469-8986.2011.01273.x, PubMed:
21895683
Guenther, F. H. (2016). Neural control of speech. In F. H.
Guenther (Ed.), Neural control of speech. Cambridge, MA:
MIT Press. https://doi.org/10.7551/mitpress/10471.001.0001
Guenther, F. H., Ghosh, S. S., & Tourville, J. A. (2006). Neural
modeling and imaging of the cortical interactions underlying
syllable production. Brain and Language, 96, 280–301.
https://doi.org/10.1016/j.bandl.2005.06.001, PubMed:
16040108
Guimond, S., Vachon, F., Nolden, S., Lefebvre, C., Grimault, S.,
& Jolicoeur, P. (2011). Electrophysiological correlates of
the maintenance of the representation of pitch objects
in acoustic short-term memory. Psychophysiology, 48,
1500–1509. https://doi.org/10.1111/j.1469-8986.2011.01234.x,
PubMed: 21824153
Hagoort, P., & Brown, C. M. (2000). ERP effects of listening
to speech: Semantic ERP effects. Neuropsychologia, 38,
1518–1530. https://doi.org/10.1016/S0028-3932(00)00052-X
Hahne, A., & Friederici, A. D. (1999). Electrophysiological
evidence for two steps in syntactic analysis. Early automatic
and late controlled processes. Journal of Cognitive
Neuroscience, 11, 194–205. https://doi.org/10.1162
/089892999563328, PubMed: 10198134
Havy, M., Serres, J., & Nazzi, T. (2014). A consonant/vowel
asymmetry in word-form processing: Evidence in childhood
and in adulthood. Language and Speech, 57, 254–281. https://
doi.org/10.1177/0023830913507693, PubMed: 25102609
Hooper, J. B. (1972). The syllable in phonological theory,
Language, 48, 525. https://doi.org/10.2307/412031
Kaiser, J. (2015). Dynamics of auditory working memory.
Frontiers in Psychology, 6, 613. https://doi.org/10.3389/fpsyg
.2015.00613, PubMed: 26029146
Kuperberg, G. R. (2016). Separate streams or probabilistic
inference? What the N400 can tell us about the comprehension
of events. Language, Cognition and Neuroscience, 31,
602–616. https://doi.org/10.1080/23273798.2015.1130233,
PubMed: 27570786
Lang, W., Starr, A., Lang, V., Lindinger, G., & Deecke, L. (1992).
Cortical DC potential shifts accompanying auditory and visual
short-term memory. Electroencephalography and Clinical
Neurophysiology, 82, 285–295. https://doi.org/10.1016/0013
-4694(92)90108-T
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network
for semantics: (De)constructing the N400. Nature Reviews
Neuroscience, 9, 920–933. https://doi.org/10.1038/nrn2532,
PubMed: 19020511
Lefebvre, C., Vachon, F., Grimault, S., Thibault, J., Guimond, S.,
Peretz, I., et al. (2013). Distinct electrophysiological indices
of maintenance in auditory and visual short-term memory.
Neuropsychologia, 51, 2939–2952. https://doi.org/10.1016/j
.neuropsychologia.2013.08.003, PubMed: 23938319
Levelt, W. J. M. (1989). Speaking: From intention to
articulation. Cambridge, MA: MIT Press. https://doi.org/10
.2307/1423219
Levelt, W. J. M. (1992). Accessing words in speech production:
Stages, processes and representations. Cognition, 42, 1–22.
https://doi.org/10.1016/0010-0277(92)90038-J, PubMed:
1582153
Levelt, W. J. M., Roelofs, A., & Meyer, A. S. (1999). A theory
of lexical access in speech production. Behavioral
and Brain Sciences, 22, 1–75. https://doi.org/10.1017
/S0140525X99001776, PubMed: 11301520
Malassis, R., Rey, A., & Fagot, J. (2018). Non-adjacent
dependencies processing in human and non-human
primates. Cognitive Science, 42, 1677–1699. https://doi.org
/10.1111/cogs.12617, PubMed: 29781135
Marchetto, E., & Bonatti, L. L. (2015). Finding words and word
structure in artificial speech: The development of infants’
sensitivity to morphosyntactic regularities. Journal of Child
Language, 42, 873–902. https://doi.org/10.1017
/S0305000914000452, PubMed: 25300736
Marcus, G. F., Vijayan, S., Bandi Rao, S., & Vishton, P. M. (1999).
Rule learning by seven-month-old infants. Science, 283,
77–80. https://doi.org/10.1126/science.283.5398.77, PubMed:
9872745
Weyers and Mueller
1485
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190. https://doi.org/10.1016/j.jneumeth
.2007.03.024, PubMed: 17517438
McCallum, W. C., Farmer, S. F., & Pocock, P. V. (1984). The
effects of physical and semantic incongruites on auditory
event-related potentials. Electroencephalography and
Clinical Neurophysiology/Evoked Potentials, 59, 477–488.
https://doi.org/10.1016/0168-5597(84)90006-6
Mehler, J., Dommergues, J. Y., Frauenfelder, U., & Segui, J.
(1981). The syllable’s role in speech segmentation. Journal
of Verbal Learning and Verbal Behavior, 20, 298–305.
https://doi.org/10.1016/S0022-5371(81)90450-3
Mehler, J., Peña, M., Nespor, M., & Bonatti, L. (2006). The “soul”
of language does not use statistics: Reflections on vowels and
consonants. Cortex, 42, 846–854. https://doi.org/10.1016
/S0010-9452(08)70427-1
Milne, A. E., Mueller, J. L., Männel, C., Attaheri, A., Friederici,
A. D., & Petkov, C. I. (2016). Evolutionary origins of
non-adjacent sequence processing in primate brain
potentials. Scientific Reports, 6, 1–10. https://doi.org/10.1038
/srep36259, PubMed: 27827366
Monte-Ordoño, J., & Toro, J. M. (2017a). Different ERP profiles
for learning rules over consonants and vowels.
Neuropsychologia, 97, 104–111. https://doi.org/10.1016/j
.neuropsychologia.2017.02.014, PubMed: 28232218
Monte-Ordoño, J., & Toro, J. M. (2017b). Early positivity signals
changes in an abstract linguistic pattern. PLoS One, 12,
e0180727. https://doi.org/10.1371/journal.pone.0180727,
PubMed: 28678863
Morgan-Short, K., Sanz, C., Steinhauer, K., & Ullman, M. T.
(2010). Second language acquisition of gender agreement in
explicit and implicit training conditions: An event-related
potential study. Language Learning, 60, 154–193. https://
doi.org/10.1111/j.1467-9922.2009.00554.x, PubMed:
21359123
Mueller, J. L., Bahlmann, J., & Friederici, A. D. (2008a). The role
of pause cues in language learning: The emergence of
event-related potentials related to sequence processing.
Journal of Cognitive Neuroscience, 20, 892–905. https://doi
.org/10.1162/jocn.2008.20511, PubMed: 18201121
Mueller, J. L., Bahlmann, J., & Friederici, A. D. (2010).
Learnability of embedded syntactic structures depends on
prosodic cues. Cognitive Science, 34, 338–349. https://doi.org
/10.1111/j.1551-6709.2009.01093.x, PubMed: 21564216
Mueller, J. L., Friederici, A. D., & Männel, C. (2012). Auditory
perception at the root of language learning. Proceedings of
the National Academy of Sciences, U.S.A., 109, 15953–15958.
https://doi.org/10.1073/pnas.1204319109, PubMed: 23019379
Mueller, J. L., Girgsdies, S., & Friederici, A. D. (2008). The
impact of semantic-free second-language training on ERPs
during case processing. Neuroscience Letters, 443, 77–81.
https://doi.org/10.1016/j.neulet.2008.07.054, PubMed:
18201121
Mueller, J. L., Milne, A. E., & Männel, C. (2018). Non-adjacent
auditory sequence learning across development and primate
species. Current Opinion in Behavioral Sciences, 21,
112–119. https://doi.org/10.1016/j.cobeha.2018.04.002
Mueller, J. L., Oberecker, R., & Friederici, A. D. (2009). Syntactic
learning by mere exposure an ERP study in adult learners.
BMC Neuroscience, 10, 1–9. https://doi.org/10.1186/1471
-2202-10-89, PubMed: 19640301
Mueller, J. L., ten Cate, C., & Toro, J. M. (2020). A comparative
perspective on the role of acoustic cues in detecting language
structure. Topics in Cognitive Science, 12, 859–874. https://
doi.org/10.1111/tops.12373, PubMed: 30033636
Munka, L., & Berti, S. (2006). Examining task-dependencies
of different attentional processes as reflected in the P3a
and reorienting negativity components of the human
event-related brain potential. Neuroscience Letters, 396,
177–181. https://doi.org/10.1016/j.neulet.2005.11.035,
PubMed: 16356637
Nazzi, T., Poltrock, S., & Von Holzen, K. (2016). The
developmental origins of the consonant bias in lexical
processing. Current Directions in Psychological Science, 25,
291–296. https://doi.org/10.1177/0963721416655786
Nespor, M., Peña, M., & Mehler, J. (2003). On the different roles
of vowels and consonants in speech processing and language
acquisition. Lingue e Linguaggio, 2, 201–227. https://doi.org
/10.1418/10879
New, B., Araújo, V., & Nazzi, T. (2008). Differential processing of
consonants and vowels in lexical access through reading.
Psychological Science, 19, 1223–1227. https://doi.org/10.1111
/j.1467-9280.2008.02228.x, PubMed: 19121127
Newport, E. L., & Aslin, R. N. (2004). Learning at a distance I.
Statistical learning of non-adjacent dependencies. Cognitive
Psychology, 48, 127–162. https://doi.org/10.1016/S0010-0285
(03)00128-2, PubMed: 14732409
Nolden, S., Bermudez, P., Alunni-Menichini, K., Lefebvre, C.,
Grimault, S., & Jolicoeur, P. (2013). Electrophysiological
correlates of the retention of tones differing in timbre in
auditory short-term memory. Neuropsychologia, 51,
2740–2746. https://doi.org/10.1016/j.neuropsychologia.2013
.09.010, PubMed: 24036359
Onnis, L., Monaghan, P., Christiansen, M. H., & Chater, N.
(2004). Variability is the spice of learning, and a crucial
ingredient for detecting and generalizing in nonadjacent
dependencies. Proceedings of the Annual Meeting of the
Cognitive Science Society, 26, 1047–1052.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis
of MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869. https://doi.org/10.1155/2011/156869, PubMed:
21253357
Osterhout, L., McLaughlin, J., Pitkänen, I., Frenck-Mestre, C., &
Molinaro, N. (2006). Novice learners, longitudinal designs,
and event-related potentials: A means for exploring the
neurocognition of second language processing. Language
Learning, 56, 199–230. https://doi.org/10.1111/j.1467-9922
.2006.00361.x
Paulmann, S., Bleichner, M., & Kotz, S. A. (2013). Valence,
arousal, and task effects in emotional prosody processing.
Frontiers in Psychology, 4, 345. https://doi.org/10.3389/fpsyg
.2013.00345, PubMed: 23801973
Peña, M., Bonatti, L. L., Nespor, M., & Mehler, J. (2002). Signal-
driven computations in speech processing. Science, 298,
604–607. https://doi.org/10.1126/science.1072901, PubMed:
12202684
Picton, T. W. (2011). Human auditory evoked potentials. Plural
Publishing.
Poeppel, D., & Assaneo, M. F. (2020). Speech rhythms and their
neural foundations. Nature Reviews Neuroscience, 21,
322–334. https://doi.org/10.1038/s41583-020-0304-4, PubMed:
32376899
Polich, J. (2007). Updating P300: An integrative theory of P3a
and P3b. Clinical Neurophysiology, 118, 2128–2148. https://
doi.org/10.1016/j.clinph.2007.04.019, PubMed: 17573239
Pratt, H. (2012). Sensory ERP components. In S. J. Luck &
E. S. Kappenman (Eds.), The Oxford handbook of ERP
components (pp. 89–114). Oxford University Press.
Rabovsky, M., Hansen, S. S., & McClelland, J. L. (2018).
Modelling the N400 brain potential as change in a
probabilistic representation of meaning. Nature Human
Behaviour, 2, 693–705. https://doi.org/10.1038/s41562-018
-0406-4, PubMed: 31346278
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
1486
Journal of Cognitive Neuroscience
Volume 34, Number 8
Räsänen, O., Doyle, G., & Frank, M. C. (2018). Pre-linguistic
segmentation of speech into syllable-like units. Cognition,
171, 130–150. https://doi.org/10.1016/j.cognition.2017.11
.003, PubMed: 29156241
R Core Team. (2020). A language and environment for
statistical computing (Vol. 2). R Foundation for Statistical
Computing https://www.r-project.org.
Reber, A. S. (1967). Implicit learning of artificial grammars.
Journal of Verbal Learning and Verbal Behavior, 6,
855–863. https://doi.org/10.1016/S0022-5371(67)80149-X
Rebuschat, P. (2013). Measuring implicit and explicit knowledge
in second language research. Language Learning, 63,
595–626. https://doi.org/10.1111/lang.12010
The Production of Speech, 109–136. https://doi.org/10.1007
/978-1-4613-8202-7_6
Toro, J. M., Nespor, M., Mehler, J., & Bonatti, L. L. (2008).
Finding words and rules in a speech stream: Functional
differences between vowels and consonants. Psychological
Science, 19, 137–144. https://doi.org/10.1111/j.1467-9280
.2008.02059.x, PubMed: 18271861
Toro, J. M., Shukla, M., Nespor, M., & Endress, A. D. (2008). The
quest for generalizations over consonants: Asymmetries
between consonants and vowels are not the by-product
of acoustic differences. Perception & Psychophysics, 70,
1515–1525. https://doi.org/10.3758/PP.70.8.1515, PubMed:
19064494
Ruchkin, D. S., Berndt, R. S., Johnson, R., Ritter, W., Grafman, J.,
van den Bos, E., Christiansen, M. H., & Misyak, J. B.
& Canoune, H. L. (1997). Modality-specific processing
streams in verbal working memory: Evidence from
spatio-temporal patterns of brain activity. Cognitive Brain
Research, 6, 95–113. https://doi.org/10.1016/S0926-6410(97)
00021-9, PubMed: 9450603
Saffran, J. R., Newport, E. L., & Aslin, R. N. (1996). Word
segmentation: The role of distributional cues. Journal of
Memory and Language, 35, 606–621. https://doi.org/10.1006
/jmla.1996.0032
Sanders, L. D., Newport, E. L., & Neville, H. J. (2002).
Segmenting nonsense: An event-related potential index
of perceived onsets in continuous speech. Nature
Neuroscience, 5, 700–703. https://doi.org/10.1038/nn873,
PubMed: 12068301
Sassenhagen, J., & Draschkow, D. (2019). Cluster-based
permutation tests of MEG/EEG data do not establish
significance of effect latency or location. Psychophysiology,
56, e13335. https://doi.org/10.1111/psyp.13335, PubMed:
25151545
Sassenhagen, J., Schlesewsky, M., & Bornkessel-Schlesewsky, I.
(2014). The P600-as-P3 hypothesis revisited: Single-trial
analyses reveal that the late EEG positivity following
linguistically deviant material is reaction time aligned. Brain
and Language, 137, 29–39. https://doi.org/10.1016/j.bandl
.2014.07.010, PubMed: 25151545
Schiller, N. O., & Costa, A. (2006). Different selection principles
of freestanding and bound morphemes in language
production. Journal of Experimental Psychology: Learning,
Memory, and Cognition, 32, 1201–1207. https://doi.org/10
.1037/0278-7393.32.5.1201, PubMed: 16938057
Schröger, E., & Wolff, C. (1998). Attentional orienting and
reorienting is indicated by human event-related brain
potentials. Neuroreport, 9, 3355–3358. https://doi.org/10
.1097/00001756-199810260-00003, PubMed: 9855279
Sergent, C., Baillet, S., & Dehaene, S. (2005). Timing of the
brain events underlying access to consciousness during the
attentional blink. Nature Neuroscience, 8, 1391–1400.
https://doi.org/10.1038/nn1549, PubMed: 16158062
Sharp, D. J., Scott, S. K., Cutler, A., & Wise, R. J. S. (2005).
Lexical retrieval constrained by sound structure: The role
of the left inferior frontal gyrus. Brain and Language, 92,
309–319. https://doi.org/10.1016/j.bandl.2004.07.002,
PubMed: 15721963
Shattuck-Hufnagel, S. (1983). Sublexical units and
suprasegmental structure in speech production planning.
(2012). Statistical learning of probabilistic nonadjacent
dependencies by multiple-cue integration. Journal of Memory
and Language, 67, 507–520. https://doi.org/10.1016/j.jml.2012
.07.008
Van Den Brink, D., Brown, C. M., & Hagoort, P. (2001).
Electrophysiological evidence for early contextual
influences during spoken-word recognition: N200 versus
N400 effects. Journal of Cognitive Neuroscience, 13,
967–985. https://doi.org/10.1162/089892901753165872,
PubMed: 11595099
Van Ooijen, B. (1996). Vowel mutability and lexical selection in
English: Evidence from a word reconstruction task. Memory
and Cognition, 24, 573–583. https://doi.org/10.3758
/BF03201084, PubMed: 8870528
Vuong, L. C., Meyer, A. S., & Christiansen, M. H. (2016).
Concurrent statistical learning of adjacent and nonadjacent
dependencies. Language Learning, 66, 8–30. https://doi.org
/10.1111/lang.12137
Wetter, S., Polich, J., & Murphy, C. (2004). Olfactory, auditory,
and visual ERPs from single trials: No evidence for
habituation. International Journal of Psychophysiology, 54,
263–272. https://doi.org/10.1016/j.ijpsycho.2004.04.008,
PubMed: 15331217
Wilson, B., Spierings, M., Ravignani, A., Mueller, J. L., Mintz,
T. H., Wijnen, F., et al. (2020). Non-adjacent dependency
learning in humans and other animals. Topics in Cognitive
Science, 12, 843–858. https://doi.org/ttps://doi.org/10.1111
/tops.12381, PubMed: 32729673
Winkler, I., Debener, S., Muller, K.-R., & Tangermann, M.
(2015). On the influence of high-pass filtering on ICA-based
artifact reduction in EEG-ERP. In 2015 37th Annual
International Conference of the IEEE Engineering in
Medicine and Biology Society (EMBC), pp. 4101–4105.
https://doi.org/10.1109/EMBC.2015.7319296, PubMed:
26737196
Yingling, C. D., & Nethercut, G. E. (1983). Evoked responses
to frequency shifted tones: Tonotopic and contextual
determinants. International Journal of Neuroscience, 22,
107–118. https://doi.org/10.3109/00207459308987389,
PubMed: 6668128
Ziegler, W., Aichert, I., & Staiger, A. (2010). Syllable- and
rhythm-based approaches in the treatment of apraxia of
speech. Perspectives on Neurophysiology and Neurogenic
Speech and Language Disorders, 20, 59–66. https://doi.org
/10.1044/nnsld20.3.59
Weyers and Mueller
1487
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
6
7
2
0
3
3
1
7
0
/
/
j
o
c
n
_
a
_
0
1
8
7
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3