A Predictive Processing Account of Card Sorting: Fast
Proactive and Reactive Frontoparietal Cortical Dynamics
during Inference and Learning of Perceptual Categories
Francisco Barceló
Abstract
■ For decades, a common assumption in cognitive neuroscience
has been that prefrontal executive control is mainly engaged dur-
ing target detection [Posner, M. I., & Petersen, S. E. The attention
system of the human brain. Annual Review of Neuroscience, 13,
25–42, 1990]. More recently, predictive processing theories of
frontal function under the Bayesian brain hypothesis emphasize
a key role of proactive control for anticipatory action selection
(i.e., planning as active inference). Here, we review evidence of
fast and widespread EEG and magnetoencephalographic fronto-
temporo-parietal cortical activations elicited by feedback cues
and target cards in the Wisconsin Card Sorting Test. This evidence
is best interpreted when considering negative and positive feed-
back as predictive cues (i.e., sensory outcomes) for proactively
updating beliefs about unknown perceptual categories. Such pre-
dictive cues inform posterior beliefs about high-level hidden
categories governing subsequent response selection at target
onset. Quite remarkably, these new views concur with Don
Stuss’ early findings concerning two broad classes of P300 cortical
responses evoked by feedback cues and target cards in a comput-
erized Wisconsin Card Sorting Test analogue. Stuss’ discussion of
those P300 responses—in terms of the resolution of uncertainty
about response (policy) selection as well as the participants’
expectancies for future perceptual or motor activities and their
timing—was prescient of current predictive processing and active
(Bayesian) inference theories. From these new premises, a
domain-general frontoparietal cortical network is rapidly engaged
during two temporarily distinct stages of inference and learning of
perceptual categories that underwrite goal-directed card sorting
behavior, and they each engage prefrontal executive functions in
fundamentally distinct ways. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
“Perhaps the most widely accepted measure to show
executive functional deficits is the sorting task.” (Stuss
& Benson, 1984, p. 18)
INTRODUCTION
In recent years, there has been a paradigm shift in the cog-
nitive neurosciences motivated by views of the brain as a
prediction machine, whose working principle is to make
active (Bayesian) inferences about the causes of its sensory
inputs (Friston, 2005, 2010). In this paper, the implica-
tions of this new theory of cortical responses are examined
for a deeper understanding of the Wisconsin Card Sorting
Test ( WCST), one of the most distinctive tests of frontal
lobe function (Milner, 1963). In doing so, we hope to
showcase the potential of these new views to solve some
paradoxes of the frontal lobe riddle—and to resolve long-
lived controversies in the literature (Teuber, 1964/2009;
Stuss & Alexander, 2007; Donchin & Coles, 1988). As with
other shifts in paradigm, predictive processing has led
many to update our beliefs and think differently about
old findings (Barceló, Periáñez, & Knight, 2002), recasting
them in the light of the new ideas. In this spirit, we review
University of the Balearic Islands
EEG and magnetoencephalographic evidence about the
fast neural dynamics underlying two processing stages
during WCST performance, as indexed by two broad
classes of scalp-recorded P300 cortical responses to infor-
mative feedback and target stimuli. These two classes of
P3-like responses show distinct scalp topographies along
a frontoparietal axis, with discrete contributions from
frontal and posterior multimodal association cortices
(Knight, 1997). Crucially, these P3-like responses can be
explained under a common overarching principle of
surprise minimization at either higher (frontal) or lower
(nonfrontal) levels in cortical hierarchies (Friston, 2005).
Hence, given the relevance of surprise minimization un-
der active inference, the thread of this paper will revolve
around the P300 family of cortical responses, one of the
most widely used EEG indexes of cognition, which has
been linked to surprise and uncertainty resolution ever
since its discovery (Donchin, 1981; Sutton, Braren, Zubin,
& John, 1965).
The new theory of cortical responses allows us to recast
card sorting behavior simply in terms of two temporarily
distinct information processing stages of inference and
learning of perceptual categories ruling goal-directed action
selection (Friston, 2005). Crucially, on this view, perception
and action are closely intertwined into perception–action
cycles (Fuster, 2013), thus reinstating old ideas about
© 2020 by the Massachusetts Institute of Technology. Published under
a Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 33:9, pp. 1636–1656
https://doi.org/10.1162/jocn_a_01662
reafference and corollary discharge in the neuropsychology
of the frontal lobes (Teuber, 1964/2009; Luria, 1966).
Furthermore, the information processing demands placed
on the participant being assessed do not rigidly depend on
the relevant (“attend”) or irrelevant (“ignore”) task condi-
tions as instructed by the examiner. Instead, demands are
flexibly linked to the participant’s internal model of the
statistical structure of the task and its sensorimotor contin-
gencies (Parr, Rikhye, Halassa, & Friston, 2019; O’Regan &
Noe, 2001). These represent major departures from tradi-
tional serial processing schemes that consider behavioral
responses as the final output of stimulus-feature competi-
tion and intermediate cognitive operations (Norman &
Shallice, 1986), with little influence on earlier information
processes (Barceló & Cooper, 2018b). As a result, predic-
tions from the new theory differ from those of conven-
tional schemes and offer solid grounds to explain many
paradoxical results—for example, that frontal damage does
not always impair detection of relevant targets (Knight,
1997; cf. Posner & Petersen, 1990) or that “irrelevant”
distracters and ancillary feedback cues can both tax
working memory capacity and engage frontal resources
more than “relevant” target stimuli (Barceló & Knight,
2007a; Barceló, Escera, Corral, & Periánez, 2006).
In the following sections, we describe how the new the-
ory of cortical responses allows us to recast card sorting in
terms of predictive action selection (i.e., planning as active
inference; Friston, FitzGerald, Rigoli, Schwartenbeck, &
Pezzulo, 2017; Botvinick & Toussaint, 2012). In this context,
Don Stuss’ extensive work with frontal lobe patients has
been a source of inspiration for our own studies on the fast
neural dynamics subserving WCST performance. There is,
however, one early and less well-known study where he em-
ployed ERPs to measure the cortical responses in a card
sorting analogue of the WCST (Stuss & Picton, 1978). In ret-
rospect, it is remarkable that Don and Terence’s discussion
of their P3-like responses—to informative feedback cues—
in terms of the resolution of uncertainty about “correct
response selection,” and as linked to the “contextual updat-
ing” in the participants’ expectancies for “future perceptual
or motor activity,” preempted current predictive processing
and active inference theories of perceptual categorization.
These new views can elegantly subsume diverse frontal
lobe functions such as energization, task setting, and
monitoring (Stuss & Alexander, 2007), and also executive
attention, inhibitory control, working memory, and decision-
making (Fuster, 2019; Stuss, Shallice, Alexander, & Picton,
1995; Stuss & Benson, 1984), all of which are deployed dur-
ing WCST performance as well as in many other complex
forms of goal-directed behavior that are disrupted by
frontal lobe lesions (Luria, 1966).
ACTIVE INFERENCE AND PREFRONTAL
EXECUTIVE FUNCTIONS
Clark, 2013), together with the revival of enactivism in cog-
nitive science (O’Regan & Noe, 2001), underwrite action-
oriented cognition with concepts such as sensorimotor
contingencies and perception–action cycles for a full un-
derstanding of higher cognitive functions in humans
(Fuster, 2013). According to these views, a common ruling
principle underlies perception (i.e., sensory state estima-
tion), action (i.e., response policy selection), and learning
(i.e., perceptual and reinforcement learning), which is to
minimize the same information theory quantity in our
brains called the “free energy.” The free energy principle
posits that brains must minimize surprise when sampling
sensory data given some internal generative model
(Friston, 2010). A generative model is a probabilistic and
formal rendition of traditional schemas (Stuss et al., 1995;
Norman & Shallice, 1986) and neuronal models (Sokolov,
1963) that have been long used to explain brain and behav-
ioral responses in the exchanges with our surroundings.
The basic idea is appealing in its simplicity: The brain sets
up a number of competing hypotheses or predictions
about the causes of its sensory inputs and then actively up-
dates these predictions through action on the basis of
bottom–up prediction errors. These errors result from a
mismatch between what is predicted and what is actually
observed and can thus be thought of as “surprise signals”
being transmitted through ascending connections up the
neural hierarchies (Friston, 2019). This recursive exchange
of descending predictions and ascending prediction errors
results in information transmission (or “neuronal message
passing”) between high and low levels across the neural
hierarchies. This information exchange evolves dynamically
over time and terminates when the generative model is up-
dated (referred to as “belief updating”) and now encodes
the belief that more accurately predicts the hidden cause
of those sensations, thus minimizing surprise, as mandated
by the free energy principle (Friston, 2005, 2010).
A complementary account of the imperative to minimize
surprise (as scored by free energy) is apparent when one
appreciates that free energy is also (a mathematical bound
on) the log likelihood of sensory input, under our internal
or generative model. In statistics, this (marginal) likelihood
is known as model evidence, whereas in machine learning,
it is known as evidence bound (Winn & Bishop, 2005). In
short, the imperative is to minimize surprise, which is ex-
actly the same as maximizing the evidence for our models
of the world—sometimes referred to as self-evidencing
(Hohwy, 2016). When considering the consequences of
any action on the world, the imperative to minimize sur-
prise becomes nuanced: In other words, the imperative be-
comes the minimization of uncertainty1 (i.e., surprise
expected after an action). This lends free energy minimiza-
tion an epistemic aspect, in which most things that we do
are in the service of resolving uncertainty about how our
sensations are generated (K. Friston, personal communica-
tion, 2020).
The active inference and predictive processing frameworks
(Friston, 2005, 2019; Hohwy, 2019; Friston et al., 2017;
The simplicity of this basic scheme is also appealing to
many of us who have used the WCST (Figure 1) as a
Barceló
1637
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
an integral constituent of recursive perception–action
cycles (Friston et al., 2017; Fuster, 2013). Once the partic-
ipant infers the correct sorting category, the examiner
keeps on reinforcing the same category for the next
10 cards or so, thus favoring a context where there is little
uncertainty about the correct course of action. As the
participant becomes increasingly confident about the
reward contingencies, his or her precision2 in selecting
among competing perceptual categories and response
policies increases, leading to greater behavioral efficiency.
At this point, the epistemic (i.e., exploratory) affordance of
any policy gives way to the pragmatic (i.e., exploitative)
affordance of securing rewards. This second stage reflects
“context learning,” when the agent’s actions primarily
serve a pragmatic or exploitative function (Friston et al.,
2016). Here, we contend that these two temporarily dis-
tinct stages of inference and learning can be readily iden-
tified during WCST performance, and they each engage
prefrontal executive functions in fundamentally different
ways. Note the correspondence with accounts of execu-
tive functions in terms of nonroutine and routine activi-
ties, respectively (Stuss et al., 1995).
In this paper, existing evidence will be reappraised in the
light of this new theory of cortical responses, in the hope of
illustrating its potential to explain apparently contradictory
findings and to resolve long-lasting dialectics in the litera-
ture. An introduction to probability theory and Bayesian
inference is beyond the scope of this paper (see Doya &
Ishii, 2007). However, it will be useful to clarify some key
concepts from the outset. First, in active inference, the
causes of sensory inputs are called “hidden” states or vari-
ables because they cannot be directly observed and need to
be inferred from the sensory exchanges between an agent
and its environment, “which means computing the poste-
rior probability of (unknown or hidden) causes, given
observed outcomes” (Friston et al., 2017, p. 7). An exam-
ple of a hidden variable is the various ways to sort a card
depicting two blue circles (i.e., whether the “correct”
response depends on the color, the form, or the num-
ber of elements in the card).
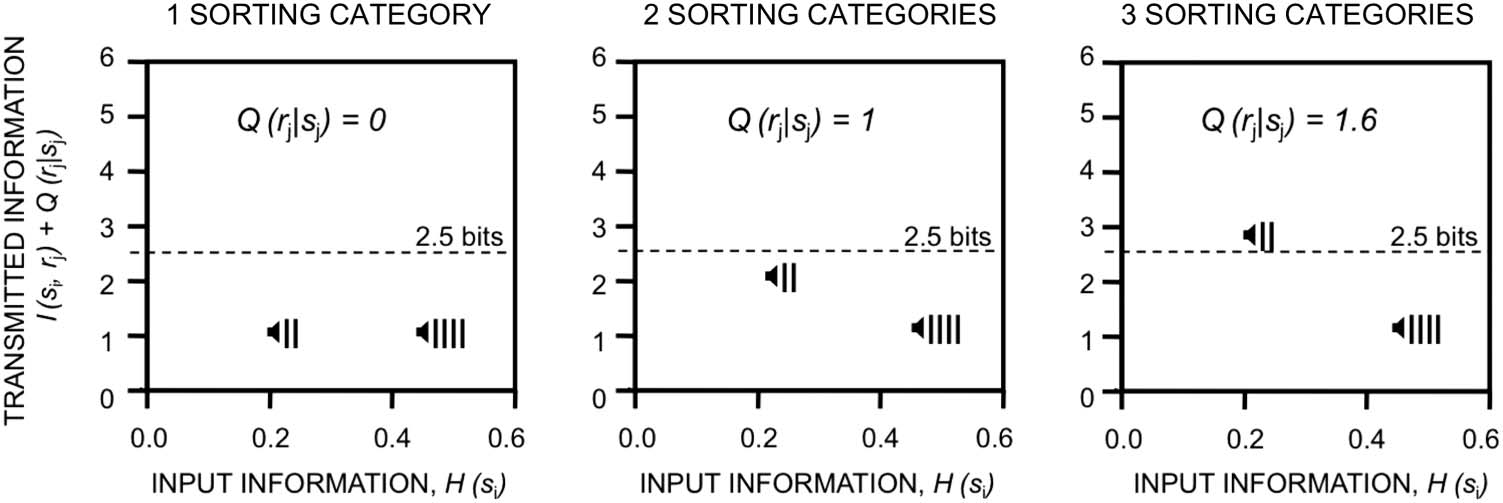
Second, there is a straightforward mathematical corre-
spondence between Bayes’ theorem and the joint and
conditional probabilities used to compute the mutual in-
formation between any two variables (Doya & Ishii,
2007). Hence, estimates of average Bayesian surprise be-
ing transmitted across frontal and posterior multimodal
association cortices during WCST performance may also
be expressed as the mutual information between hidden
states and sensory outcomes (Friston et al., 2017; cf.
Figure 2). Similar formalisms have been used to estimate
mental capacity limits in humans (Koechlin & Summerfield,
2007) and were one of the founding cornerstones of infor-
mation processing views in cognitive neuroscience (Miller,
1956).
Third, although active inference is a powerful scheme
to examine belief updating and surprise minimization at
any level in the neural hierarchies, here we will focus on
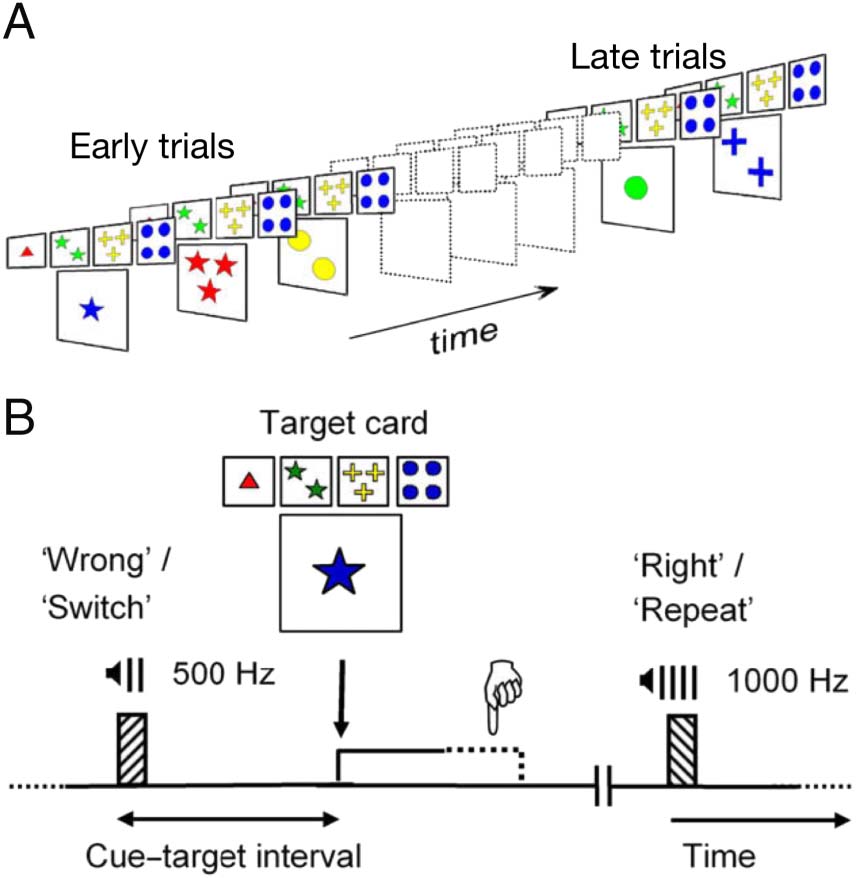
Figure 1. Computerized WCST version adapted for recording cortical
responses. (A) Schematic of one card sorting series where early and late
trials broadly map onto the stages of inference and learning of
perceptual categories. (B) Schematic of one card sorting trial where
simple tonal sounds can be instructed either as negative and positive
feedback or as “switch” and “repeat” cues informing about probabilistic
updates in the policy for responding to the ensuing target card.
Adapted from Nyhus and Barceló (2009), with permission.
neuropsychological tool for examining prefrontal execu-
tive functions in research and clinical contexts. As will be
further explained below, this is an open-ended test where
the participant (called “agent” in active inference) is re-
quested to sort a pack of cards without clear instructions
about what is the correct course of action. In these set-
tings, agents need to engage in active inference and set
up hypotheses to predict which of several perceptual
categories—that are “hidden” in the sensory cues from
the cards—will be most likely rewarded by the examiner.
Then, the participant actively tests each of his or her pre-
dictions one after another to resolve his or her uncertainty
and disambiguate among plausible alternatives. This is
an active and recursive process, where actions generate
informative prediction errors, until the examiner provides
confirmatory “correct” feedback about the rewarded per-
ceptual category. In this first stage of perceptual inference,
which normally takes several trials, participants use actions
in an epistemic or exploratory fashion to disambiguate
the rewarded category hidden in the cards (Friston et al.,
2017).
Crucially, in active inference, the feedback provided by
the examiner has “epistemic affordance.” In other words,
choosing one card or another can resolve uncertainty about
the contingencies currently in play, above and beyond
the pragmatic value of choosing the correct card. In turn,
reducing uncertainty about the card sorting contingencies
enables some more confident inference about “what
should I do next?” In short, response selection becomes
1638
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
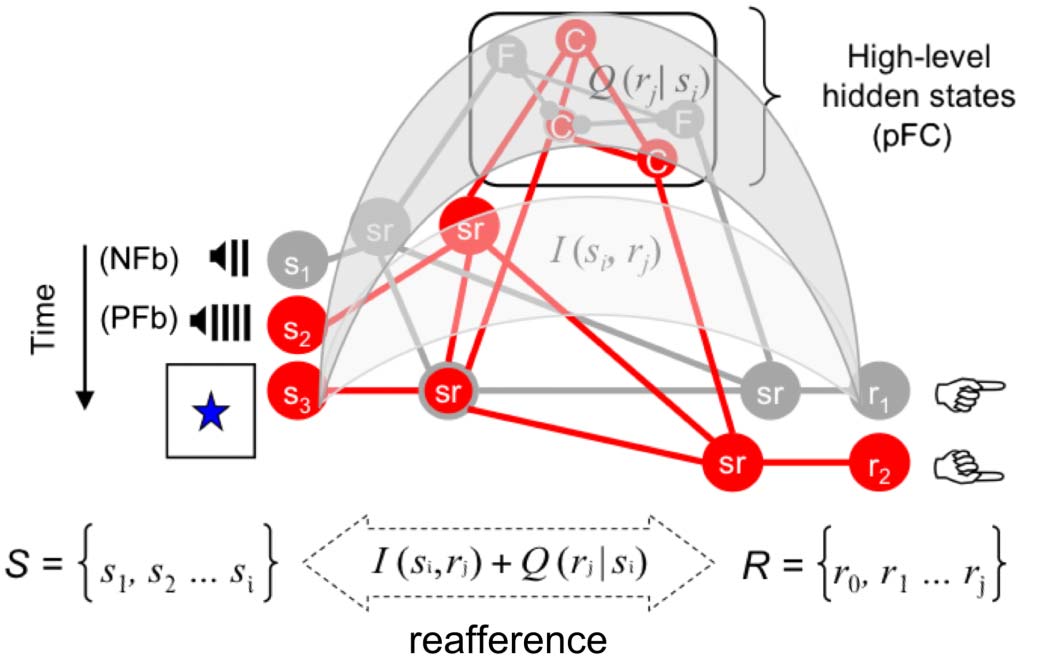
Figure 2. Hierarchical model of
prefrontal function (adopted
from Miller & Cohen, 2001).
High- and low-level belief
updates at frontal and posterior
multimodal association cortices
can be modeled as the mutual
information between hidden
states and sensory outcomes
transmitted through higher [Q(rj
| si)] and lower [I(si, rj)] levels in
the neural hierarchies,
respectively (Barceló & Cooper,
2018b). The quantity Q(rj | si)
corresponds to the psychological
notion of cognitive control
(Koechlin & Summerfield, 2007).
For simplicity, only two
perceptual categories, color (C) and form (F), are illustrated, together with only three stimuli and two motor responses from the pool of all stimuli (S ) and
responses (R) in our WCST analogue. Red indicates active units or pathways. Small circles represent conflict between two antagonistic high-level units.
Negative feedback (NFb) cues are very informative because they inform about updates in the hidden perceptual category and corresponding response
policy selection. In turn, positive feedback (PFb) cues and target cards are comparatively less informative events for response selection. The bidirectional
arrow captures the concept of “reafference,” or feedback–feedforward connectivity (Stuss & Benson, 1984), through mutually informed exteroceptive,
proprioceptive, and interoceptive sensory outcomes (Friston, 2010). Adapted from Nyhus and Barceló (2009), with permission.
two broad classes of scalp-recorded P3-like cortical re-
sponses with distinct topographies overlying frontal and
posterior multimodal association cortices, respectively
(Knight, 1997). We will refer to these two classes of cortical
responses as “anterior” and “posterior” P3-like responses,
respectively, or P3a and P3b for short (Polich, 2007;
Squires, Squires, & Hillyard, 1975). This anatomical distinc-
tion should suffice to match the research aims of many
brain lesion studies (Milner, 1963). This also agrees with
hierarchical models of prefrontal function that explicitly
distinguish between high-level hidden states represented
at pFC from low-level states at posterior association corti-
ces (see Figure 2; cf. Stuss, Picton, & Alexander, 2001).
Finally, the family of P3-like responses has long been
regarded as a proxy for surprise minimization, that is,
reduction in the uncertainty generated by surprising events
(Sutton et al., 1965), but also of learning and memory con-
solidation (Donchin, 1981). Hence, P3-like responses offer
sufficient topographical and temporal resolution to assess
high- and low-level belief updating at frontal and posterior
multimodal association cortices, during the inference and
learning of perceptual categories in card sorting.
A GENERATIVE MODEL OF CARD SORTING
In this section, card sorting will be framed in terms of active
inference—that is, the updating of the participant’s beliefs
about how his or her sensations are caused during WCST
performance. Card sorting can be seen as a paradigmatic
example of goal-directed uncertainty resolution, whereby
perception and action are jointly deployed to reduce
contextual uncertainty. This resolution of uncertainty rests
upon inferring the perceptual categories “hidden” in the
cards (in the parietal hierarchies) and then using this infer-
ence to select a response on the basis of inferred outcomes
under each plausible choice (in prefrontal hierarchies).
This entails solving the dual problem of (1) inferring some
hitherto unknown sorting categories and (2) learning the
statistical structure of the task, which underwrites policy
selection (Friston et al., 2017). For this, active inference
rests on a generative model of observed sensory outcomes,
which is just a context-sensitive, time-dynamic, and revis-
able hypothesis about how observed sensory outcomes
are generated, while evaluating competing hypotheses
about their hidden causes. Crucially, unlike other schemes,
these generative models have a hierarchical structure and
incorporate actions, responses, or decisions. This means
that sensory observations depend on actions (e.g., which
card you select), which requires the generative model to
entertain expectations about sensory outcomes under
different action plans or response “policies” (Friston et al.,
2016, 2017).
A great deal of clinical and brain imaging studies on the
WCST have been inspired by Milner’s (1963) seminal work,
and many of them have adopted similar testing materials
and procedures. Milner used four keycards differing in col-
or, form, and number: one red triangle, two green stars,
three yellow crosses, and four blue circles. These were
placed in front of the patient, who was also given a pack
of 128 choice cards that varied along these same dimen-
sions (Figure 1). The only instruction given to the patient
was to match each choice card with one of the four key-
cards, following some hitherto undeclared sorting princi-
ple. Then, the patient was to rely on the information
provided by the examiner’s positive (“right”) or negative
(“wrong”) feedback—to discover the hidden sorting rule
and try to get as many correct card sorts as possible.
Patients received no further information about how to
proceed, and many other important task variables were ini-
tially unknown to the patient. Crucially, one such hidden
Barceló
1639
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
variable was the examiner’s policy for reinforcing each of
the perceptual categories in the cards, which are the color,
number, and type of forms in the card. In addition, Milner
arbitrarily used the following (hidden) sequence: color –
form – number – color – form – number. Likewise, and un-
known to the patient, Milner changed the reinforced sorting
category after 10 consecutive correct sorts—and did so with-
out giving any warning to the patient (Milner, 1963).
Test administration terminated when the patient suc-
cessfully completed the six sorting categories or when all
128 choice cards had been placed on top of the keycards.
The main test scores were the total number of categories
achieved as well as the number of perseverative and non-
perseverative (i.e., “set-loss”) errors. A perseverative error
was scored when a card was sorted using the previously
reinforced—but now irrelevant—sorting category or when
there was a continued tendency to respond using one
and the same category. All other errors were considered
as nonperseverative (Milner, 1963). Note that successful
performance in this test requires that patients efficiently in-
fer and learn some critical hidden variables ruling their
(sensorimotor) information exchanges with their testing
environment.
Over the years, this original version of the WCST has
been adapted in many different ways to best examine the
componential structure of the information processes
underlying brain and behavioral responses in healthy and
clinical populations. For instance, Nelson (1976) removed
the choice cards that shared more than one attribute with
the keycards, thus eliminating response ambiguity and
simplifying the scoring of errors. This version consists of
two packs of 24 cards each, categories are scored with only
six consecutive correct sorts, and patients are told when to
change the category, but the actual criterion is not declared.
In other versions, patients are informed beforehand of the
three sorting categories (Stuss et al., 2000). Clearly, these
modifications facilitate inference about sorting categories.
In fact, there are nearly as many subtle adaptations of the
original test as published studies, and it would be difficult
to review them all here. A more cost-effective approach is
to examine the basic elements of a generative model of per-
ceptual categorization during card sorting (Rigoli, Pezzulo,
Dolan, & Friston, 2017) and see how these elements help us
to interpret the frontal and posterior P3-like responses
evoked during the two temporarily distinct stages of infer-
ence and learning. Put simply, a generative model defines
sensory inputs as a function of the underlying hidden
causes (i.e., states or variables) in the environment
(Friston, 2005, 2010):
ð
e ¼ g v; θ
Þ;
(1)
where e represents the sensory evidence (e.g., exterocep-
tive, proprioceptive, and interoceptive sensory inputs) and
g(v, θ) is a nonlinear probabilistic function that generates
sensory inputs from their hidden causes. These causes
are represented by a vector v, which is only a list of the
unknown or hidden variables in a typical testing situation
(i.e., v = variable type and number of sorting categories,
variable length of trial sequences, variable ISIs, variable ori-
entation and color of visual features in the cards, variable
pitch of “right” and “wrong” feedback sounds, variable
room luminance, and so on). The generative model is fur-
ther elaborated to incorporate a hierarchical structure and
temporal dynamics, meaning that some of those hidden
variables will be disambiguated early by belief updating at
low levels in the cortical hierarchy (e.g., primary sensory
cortices), whereas other variables will entail a longer-lasting
exchange of predictions and prediction errors at higher
levels in the hierarchy (e.g., prefrontal and posterior associ-
ation cortices; Figure 2), until belief updating is complete
and surprise is minimized. The parameters (θ) in the model
encode the contingencies or relationships among those var-
iables and need to be learned through practice (e.g., that
the sorting category changes after 10 correct sorts).
From the extant literature, one might predict that pFC
will be mostly engaged when resolving the uncertainty
about the abstract sorting categories and also in setting
up the corresponding response policy (i.e., “task setting”—
the sensorimotor mapping between attributes in the card
and response selection as made explicit by the keycards;
cf. Stuss & Alexander, 2007; Stuss et al., 1995). Furthermore,
from conventional views on the anatomy of the executive
attention system, one might assume maximal prefrontal
engagement when the target card is on display for its
appraisal (cf. Posner & Petersen, 1990). In turn, one might
be less inclined to expect frontal involvement in response
to changes in the sensory attributes of the cards and the
feedback and might dismiss other variables (i.e., room
luminance) as largely irrelevant for evaluating frontal lobe
functions.
These are some aspects where hierarchical generative
models depart from traditional schemes because predic-
tions and prediction errors—and how high up in the
neural hierarchy these surprise signals are transmitted—
depend on nonlinear interactions between those hidden
variables (Friston et al., 2017). Very roughly, active infer-
ence relies on Bayes’ rule [p(h|e) = p(e|h) p(h)/p(e)] to
formalize belief updating from a prior belief about a hy-
pothesis, p(h), to a posterior belief, p(h|e), based on the
likelihood that the available evidence was generated by
that hypothesis, p(e|h). The terms p(h) and p(h|e) are
known as “prior” and “posterior” probabilities, respec-
tively and, together with the actual evidence, p(e), allow
us to quantify Bayesian surprise (e.g., a prediction error3)
as the divergence between the prior and posterior prob-
abilities (Hohwy, 2019). Below, we show how this simple
rule may be recursively applied on a trial-by-trial basis to nar-
row down the participants’ hypotheses about the hidden
sorting category based on evidence from the feedback—
and, indeed, beliefs about what to do next and the requisite
task parameters that have to be learned.
To determine which aspects of the testing situation will
generate Bayesian surprise signals large enough to engage
1640
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
pFC rests on computing the Kullback–Leibler divergence
between prior and posterior distributions (Itti & Baldi,
2009). This can be approximated by computing the mutual
information between hidden variables and observed sen-
sory outcomes, that is, the information gained about un-
observable hidden states from observable outcomes
(Barceló & Cooper, 2018a; Friston et al., 2017; Figure 2).
These estimations of the informativeness of task events
are far more fine-tuned than traditional stimulus taxon-
omies in terms of targets, distracters, feedback cues, and
so on. However, to date, very few studies have attempted
to quantify the information gain of task events during card
sorting (cf. Kopp & Lange, 2013; Nyhus & Barceló, 2009).
As an alternative, below we present some intuitive exam-
ples to illustrate under which testing conditions Bayesian
surprise signals (i.e., precision-weighted prediction errors)
can be large enough to engage pFC as opposed to posteri-
or multimodal association cortices during the inference
and learning of the sorting categories. For an accurate
recording of behavioral and cortical responses, we will
assume the examiner employs a computerized version of
the WCST for testing the participant (see Figure 1).
Inference About the Sorting Category
To infer the sorting category, the participant needs to find
out about the response policy (course of action) being re-
warded by the examiner. For this, the participant needs to
resolve his or her uncertainty about the current context
(i.e., the “hidden” correct sorting category) to know what
to do next, and his or her actions should therefore fulfill
exploratory or epistemic imperatives (Friston et al., 2017).
The P3a and P3b responses associated with this type of
belief updating index rapid evidence accumulation over a
scale of milliseconds (Friston, 2005) and engage prefrontal
and posterior multimodal association cortices (Barceló
et al., 2002; Stuss & Picton, 1978), as part of a more widely
distributed neural network (Parr et al., 2019; Friston et al.,
2017). Now, let us examine an example of how this in-
ferential process entails the planning of a sequence of
perception–action cycles over several card sorting trials.
To start with, the participant needs to rely on his or her
prior knowledge on how to sort things, including cards,
together with the scarce instructions from the examiner
that there is some “correct” way of sorting the cards.
Then, he or she makes a first educated guess (based on prior
beliefs about the nature of these kinds of tasks) for
“number” and matches the first choice card with the first
keycard by pressing Button 1 on the keypad. For this, the
participant has visually scanned the scene, sampling the
evidence, and has correctly categorized three perceptual
dimensions in the card. This also allows him or her to
estimate a prior probability p(h) = 1/3 for number. She also
knows that the probability of a positive feedback is p(e) =
1/2, although the likelihood of this first hypothesis fitting
the evidence from feedback, p(e|h), is rather low. In any
case, he or she must wait to hear the informative first
negative feedback to update his or her belief and discard
(inhibit) “number” as a hypothesis. In doing so, his or
her brain computes a posterior probability, p(h|e), in re-
sponse to ascending prediction errors sufficiently precise
to engage pFC (Barceló et al., 2002; Stuss & Picton, 1978;
see Figure 3, left). Now, the posterior of this first card sort
becomes the prior for the next trial, and the perception–
action cycle starts again.
In fact, the participant decides to bet for “form” just after
hearing the first negative feedback and before the second
card is on display. Hence, he or she quickly presses Button
3 for form when she sees the second card. This action is like
a hypothesis-testing experiment (Parr, Rees, & Friston,
2018; Friston, Adams, Perrinet, & Breakspear, 2012), but
it does not disambiguate the correct sorting category yet.
Instead, it generates another very informative outcome:
a second negative feedback, which also sends off ascend-
ing prediction errors to pFC to update posterior beliefs
(Figure 4). By now, an attentive and efficient participant,
who has kept track of all previous sensorimotor contin-
gencies, will be fairly certain that there is only one remain-
ing category to choose from. Thus, when the third card is
displayed, the participant sorts it by color on the fourth
pile. Again, this new action does not resolve his or her
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
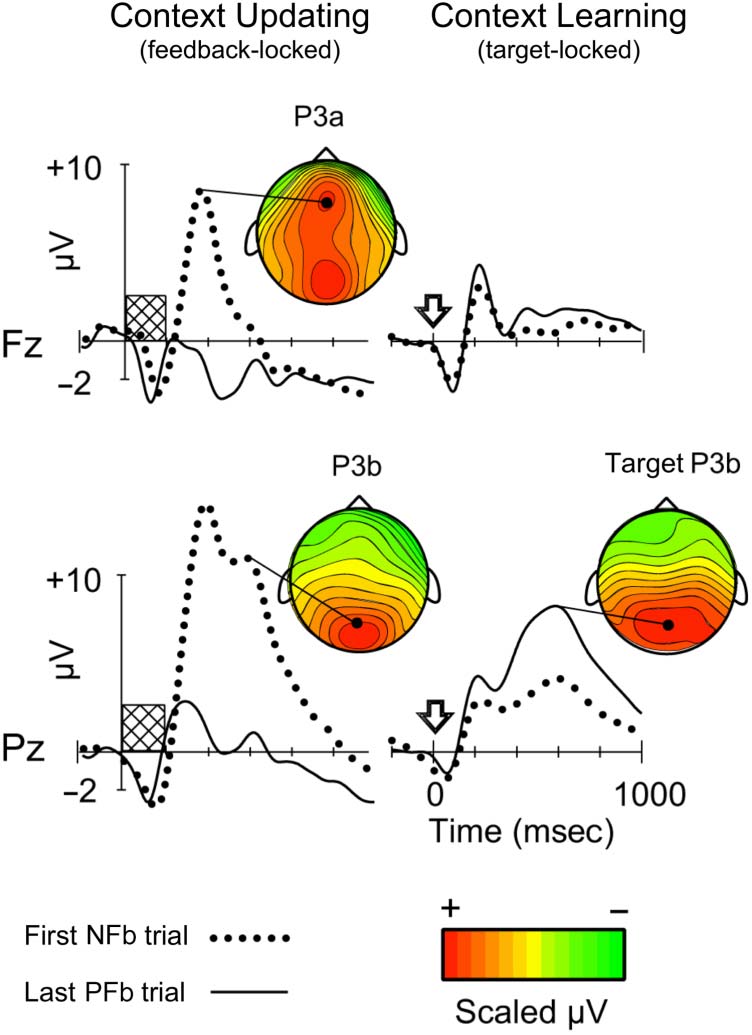
Figure 3. Cortical responses to feedback cues and WCST target cards.
Grand-averaged ERPs time-locked to feedback cues (shaded rectangle)
and target cards (wide arrow) are displayed for first negative feedback
(NFb) trials and last positive feedback (PFb) trials in a card sorting series,
at midfrontal (Fz) and midparietal (Pz) regions. Voltages are in microvolts
(μV). Scalp potential maps are shown for mean P3a and P3b responses to
first NFb cues and for mean target P3b responses to the last correct target
card in the series. The color scale is in normalized units. Early NFb trials
foster perceptual inference, whereas late PFb trials foster context learning.
Adapted with permission from Barceló et al. (2002).
Barceló
1641
correspondence between this stage of inference and fron-
tal lobe processes such as “energization” and “task setting”
(Shallice, Stuss, Picton, Alexander, & Gillingham, 2008;
Stuss & Alexander, 2007).
For subsequent cycles of perceptual inference, the first
negative feedback will normally evoke larger surprise sig-
nals than the second negative and subsequent feedbacks
(Barceló et al., 2002). This gradual reduction depends
not only on progressive belief updating, and hence a lesser
magnitude of prediction errors for these conditions, but
also on interactions with other hidden variables. For
instance, whereas the first negative feedback is unpredict-
able (i.e., the participant does not know the length of trial
series yet), the temporal onset of the next feedback can be
easily predicted after the first card sort. Furthermore, a first
positive feedback can trigger prediction errors of similar or
even larger magnitude than a second negative feedback.
This happens when the participant tests the correct cate-
gory after the first negative feedback and because of an
interaction with a sensory change in the feedback cue
from negative to positive (Lange, Seer, & Kopp, 2017; Kopp
& Lange, 2013; Periáñez & Barceló, 2009; Barceló et al., 2002).
Crucially, the engagement of pFC does not depend on the
type of feedback as delivered by the examiner but on the
magnitude (precision) of prediction errors during percep-
tual inference. In other words, it is not the sensory evi-
dence per se that determines belief updating—it is the
degree to which that evidence calls for a revision of pos-
terior beliefs (that are quintessentially time and context
dependent).
This scheme suggests that feedback stimuli are very in-
formative predictive cues that resolve the uncertainty
about ongoing predictions—and guide the planning of
subsequent actions toward the inference of the hidden
sorting category (Botvinick & Toussaint, 2012). In contrast,
the target cards are merely the workbench, where those
hypotheses are tested through action (Friston et al.,
2017), and they are comparatively less salient and informa-
tive than feedback cues for inferring the sorting category
(Barceló & Cooper, 2018a, 2018b). Note that, in this
scheme, proprioceptive inputs from actions need to be in-
tegrated with exteroceptive (i.e., visual, auditory) inputs
from the cards and feedback cues, as they all inform the
inference of the same hidden variable. Frontoparietal
multimodal association cortices are candidate structures
for this multisensory integration (Andersen & Cui, 2009),
as they generate top–down predictions in the form of
corollary discharges to suppress exteroceptive and propri-
oceptive prediction errors (Fuster, 2013; Friston, Shiner,
et al., 2012; Teuber, 1964/2009). In summary, there is a di-
rect correspondence between this recursive belief updat-
ing process—to infer the sorting category—and planning
as active inference, whereby an agent plans a desired future
state of affairs as the joint probability over the available
perceptual categories, her goal-directed actions, and the
looked-after reward from the examiner (Rigoli et al.,
2017; Botvinick & Toussaint, 2012).
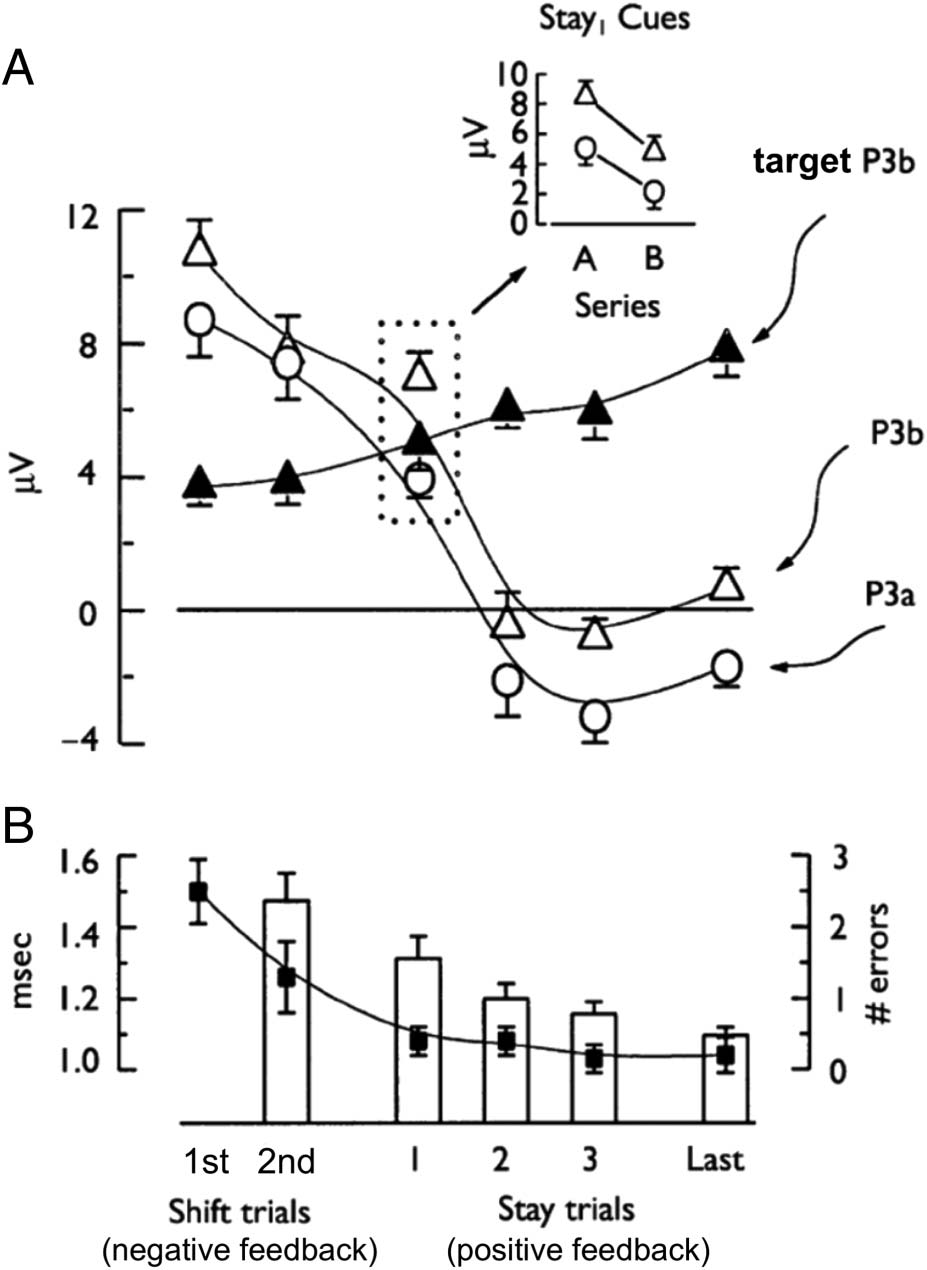
Figure 4. Cortical and behavioral responses to negative and positive
feedback trials in a card sorting series. (A) Group-averaged mean ± SEM
amplitudes of feedback-locked P3a and P3b as well as target-locked
P3b responses plotted across negative feedback (shift) and positive
feedback (stay) trials. Mean P3a and P3b amplitudes were measured
from midfrontal (Fz) and midparietal (Pz) scalp regions, respectively.
(B) Mean ± SEM RTs from efficiently completed WCST series without
errors (solid squares) and mean ± SEM number of set-loss errors from
failed series (bars) are shown during the inference and learning of the
sorting category. Adapted with permission from Barceló et al. (2002).
residual uncertainty. Only the ensuing first positive feed-
back confirms the participant’s prediction, although this
is still likely to generate ascending surprise signals large en-
ough to reach pFC (Li, Wang, Du, & Cao, 2018; Kopp &
Lange, 2013; Barceló et al., 2002). It is normally the second
positive feedback that fully matches the participant’s pre-
dictions about the hidden sorting category, thus terminat-
ing perceptual inference. This moment is marked by the
phenomenon of repetition suppression4 of frontal P3a re-
sponses to the second and subsequent positive feedbacks
in the card sorting series (Figure 4). This is reminiscent of
the extinction of cortical orienting responses (Sokolov,
1963), and it ensues from the suppression of prediction
errors about the newly disclosed perceptual category
by top–down predictions from higher cortical regions
(Friston, 2005). Note that this “planning as active infer-
ence” fits well with the role of pFC in the manipulation
and input gating (i.e., updating) of information in working
memory (Badre & Nee, 2018; Fuster, 2013). Note also the
1642
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Learning of the Sorting Category
Context learning (i.e., the learning of the sorting category)
proceeds after inference is completed by high-level belief
updating and once the participant is confident about the
course of action being rewarded by the examiner. Now,
the participant still needs to resolve the uncertainty about
the parameters in the generative model. These are the
values of hidden quantities that do not change over time,
such as the stimulus–response mappings, the length of
trial sequences, the ISIs, and so on. This type of uncer-
tainty reduction resolves the participant’s ignorance
about the probabilistic structure of the task and enables
his or her actions to fulfill an exploitative or pragmatic func-
tion (Friston et al., 2017). In one sense, resolving ignorance
about quantities that change slowly calls on the same prin-
ciples as resolving uncertainty about quantities that change
quickly. In what follows, we will explore the notion that
resolving uncertainty about model parameters necessarily
renders inferences about hidden perceptual categories
progressively more precise—and therefore more evident
in belief updating—and electrophysiologically (K. Friston,
personal communication, 2020).
The parietal P3b responses associated with this type of
long-term belief updating reflect evidence accumulation
evolving over slower time scales of seconds or longer
(Friston, 2005) and engage posterior multimodal associa-
tion cortices (Barceló et al., 2002; Figure 3, right), as part
of a widely distributed neural network (Parr et al., 2019;
Friston et al., 2017). Critically, the same imperative of
minimizing surprise also applies to this second stage of
learning. Let us examine how this type of uncertainty reso-
lution entails the accumulation of evidence through recur-
sive perception–action cycles over several trials under the
same sorting category.
Once the participant has inferred the first sorting cate-
gory, this variable is parameterized with a fixed value (e.g.,
the stimulus–response mapping for “color”), thus providing
a stable context for fulfilling the goal rewarded by the exam-
iner. With each new card sort under the correct sorting
category, the visual scanning of the scene becomes more
and more efficient as the participant learns the spatial
arrangement of the keycards. This learning speeds up but-
ton presses and improves behavioral efficiency as the par-
ticipant practices the corresponding stimulus–response
mappings (Barceló et al., 2002; Barceló, Muñoz-Céspedes,
Pozo, & Rubia, 2000; Figure 4A and B) and also gradually
learns the parameters of his or her generative model
(e.g., length of trial sequences, ISIs, visual attributes in
the cards). Note that residual uncertainty about all these
task parameters does not prevent the participant from
securing the rewarded behavioral goal by repeating the
same response policy. Importantly, now the participant’s
actions simply confirm predictions about the ongoing
sorting category, which results in repetition suppression
of P3a and P3b responses to the positive feedback and in
repetition enhancement of P3b responses to the target
cards (Figure 4A and B). Whereas the former phenomenon
reflects suppression of prediction errors to the positive
feedback, the enhancement of target P3b responses reflects
a gradual increase in the precision of predictions at higher
levels in the cortical hierarchy, as more and more evidence
about task parameters is accumulated within a card sorting
series and in successive series (Auksztulewicz & Friston,
2016).
Thus, an efficient participant will become gradually
more confident about the task parameters as he or she
successfully infers and learns more sorting categories.
Task parameters are encoded at different levels in the
neural hierarchy in the form of probability distributions
represented by their mean and precision (the inverse
of variance). The higher the precision, the narrower the
distribution is around the mean (Hohwy, 2019; Friston
et al., 2017). This may eventually lead the participant to
become quite confident that, for example, there are only
three sorting categories to choose from, that the category
will change every 10 correct sorts, that there is a constant
delay between the feedback and the card, and so on. In
other words, the precision of posterior expectations
about many hidden variables will gradually increase over
the course of testing (Auksztulewicz & Friston, 2016).
Despite this, the participant cannot predict which of
the many target cards will follow the feedback, nor which
button press will have to be selected to close the
perception–action cycle under the current sorting cate-
gory. Hence, top–down predictions cannot suppress pre-
diction errors to the onset of target cards, which is why
these evoke target P3b responses over posterior associa-
tion cortices. Crucially, this explains why these target P3b
responses never “habituate” (Donchin, 1981), and in-
stead show repetition enhancement, indexing a gradual
increase in the precision of predictions about many task
parameters being learned along the card sorting series
(Auksztulewicz & Friston, 2016; Figures 4 and 5).
Note that this stage of context learning may proceed
with less belief updating in frontal cortices during the main-
tenance of information in working memory. This is consis-
tent with evidence that target P3b responses are largely
preserved after prefrontal damage (Barceló & Knight,
2007a; Knight, 1997), given that output gating of informa-
tion from working memory relies more on posterior asso-
ciation cortices and related subcortical structures (Badre
& Nee, 2018; Stuss & Alexander, 2000). Note also the
correspondence of this second stage of learning with pro-
cesses such as “monitoring” (Shallice et al., 2008; Stuss &
Alexander, 2007; Stuss et al., 1995).
Dynamic Balance between Inference and Learning
From the foregoing description, one might be tempted
to associate the first stage of inference to the negative feed-
back early in the card sorting series and the second stage of
learning to the target cards later on in the series (Figures 3
and 4). However, this is another aspect where active
Barceló
1643
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
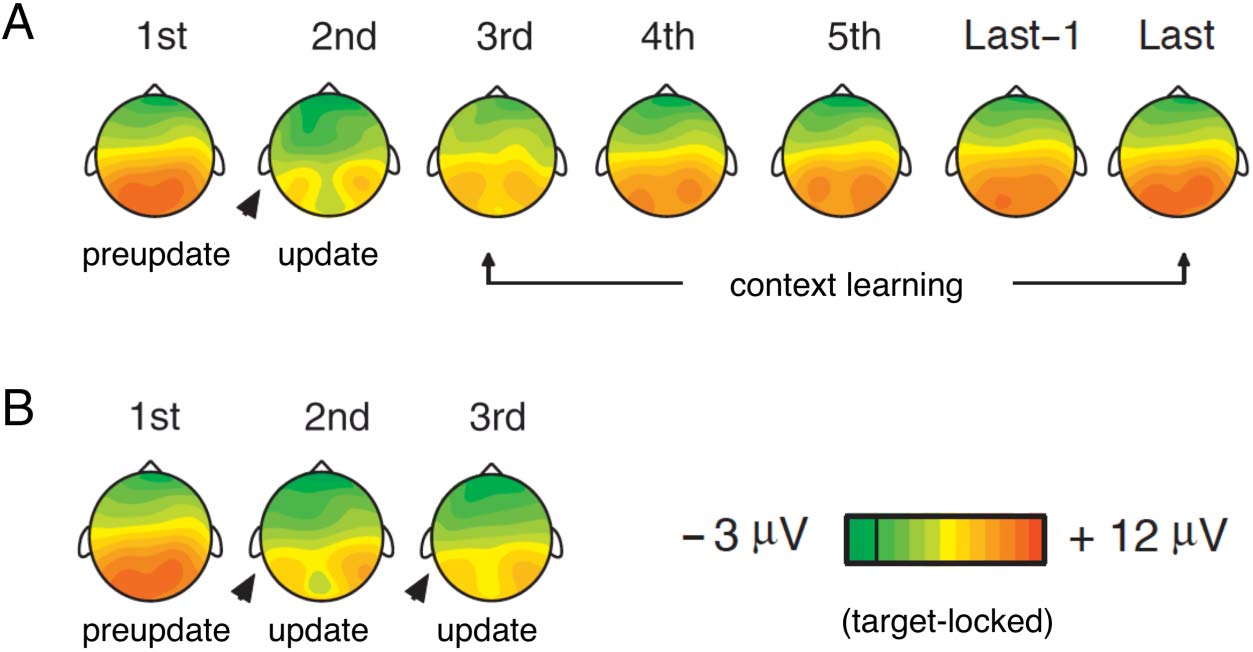
Figure 5. Voltage maps of mean target P3b amplitudes during the inference and learning of perceptual categories. Arrowheads mark target trials
preceded by a negative feedback (i.e., update trials). (A) Series with only one negative feedback trial. (B) Series with two negative feedback trials
(postupdate trials not shown). Card sorting series (A) and (B) evoked similar repetition enhancement of target P3b responses after the first
positive feedback. Note the large belief update after the first positive feedback in the third trial of series (A) indexing the correct inference of the
sorting category. Adapted with permission from Barceló et al. (2000).
inference departs from conventional theories of frontal
lobe function. Crucially, in active inference, the dynamic
balance between inference (information exploration) and
learning (information exploitation) does not depend solely
on the type of stimulus or task condition as defined by the
examiner (cf. Posner & Petersen, 1990). Instead, this
balance depends on the magnitude of the ascending sur-
prise signals that will ultimately result in either high- or
low-level belief updating—engaging either pFC or posterior
multimodal association cortices, respectively (Maisto,
Friston, & Pezzulo, 2019).
In other words, whether or not an agent’s generative
model will undergo high-level belief updating at pFC does
not depend solely on the type of stimulus that generated
the sensory inputs, such as a target card, a distracter stim-
ulus, a positive or negative feedback, and so on. As will be
shown below, any of these stimuli can potentially generate
both anterior P3a and posterior P3b responses, indexing
the inference and learning of perceptual categories,
respectively. Hence, the critical question is what is the
threshold magnitude of surprise signals above which they
will engage pFC? In active inference, the magnitude of
those prediction errors, and how high up in the neural
hierarchy they penetrate, hinges upon dynamic trial-by-trial
updates in the participant’s generative model (Figures 4
and 5).
These belief updates do not depend solely on the sensory
evidence, like the mean stimulus probability (Duncan-
Johnson & Donchin, 1977), but they are a function of the
likelihood of sensory outcomes given their expected
hidden causes and the prior probability of those causes
(Friston et al., 2017). Furthermore, the dynamic balance
between inference and learning also depends on the inter-
actions between concurrent hidden variables during WCST
performance, such as the number of sorting categories, the
length of the card sorting series, ISIs, and so on. All of these
factors determine the precision of posterior beliefs. For
example, a precise belief that the sorting rule changes on
every 10th trial is very different from an imprecise belief
that the rule might change at some point in the future.
How these factors determine the precision of ascending
surprise signals and whether there may be an information
threshold above which these signals may potentially en-
gage pFC will be addressed next.
COGNITIVE FLEXIBILITY IN
ACTIVE INFERENCE
The WCST has long been considered as a gold standard for
the assessment of cognitive flexibility (Diamond, 2013;
Miller & Cohen, 2001), which may be compromised mostly
in two ways: one, when repeating a sorting category after
negative feedback, which is scored as a perseverative error,
and second, when switching the sorting category after a
positive feedback, which is scored as a set-loss error. In
her seminal study, Milner (1963) concluded that patients
with frontal lesions were more prone to perseverative
errors than patients with nonfrontal lesions. Although the
“specificity” of WCST error scores as markers of frontal
lobe function has been questioned on several grounds
(Nyhus & Barceló, 2009; Barceló & Knight, 2002), their
“sensitivity” to frontal damage has also been confirmed
many times over the years (Glascher, Adolphs, & Tranel,
2019; Demakis, 2003; Stuss et al., 2000), thus lending
support to Milner’s seminal findings. Many metabolic brain
imaging studies have examined the neural correlates of
cognitive flexibility using WCST analogues ( Wang, Cao,
Cai, Gao, & Li, 2015; Buchsbaum, Greer, Chang, & Berman,
2005; Konishi et al., 1998). Notably, lesion and brain
imaging studies do not inform about the fast neural
dynamics of inference and learning of perceptual catego-
ries, nor about their subtle trade-off during WCST perfor-
mance. Here, we contend that knowledge about the fast
dynamics of inference and learning of the sorting
1644
Journal of Cognitive Neuroscience
Volume 33, Number 9
categories can improve our understanding of specific dis-
ruptions in information processes underlying the com-
mission of WCST errors (Lange, Seer, & Kopp, 2017;
Lange, Seer, Muller-Vahl, & Kopp, 2017; Kopp & Lange,
2013; Barceló, 1999).
In the literature on executive functions, the stages of
inference and learning of perceptual categories can be
broadly assimilated within the cognitive processes of task-
set switching and task-set maintenance, respectively
(Glascher et al., 2019; Gajewski, Ferdinand, Kray, &
Falkenstein, 2018; Diamond, 2013; Shallice et al., 2008;
Barceló, 2003). From traditional views, the information pro-
cessing demands in a cognitive task are typically linked to
nominal stimulus taxonomies as defined by the testing con-
ditions and experimental procedures (i.e., a target card, a
distracter, a negative feedback, switch and repeat cues).
For instance, a larger frontal engagement for processing
target stimuli is generally assumed (Posner & Petersen,
1990). In turn, the participant’s motor responses and the
ensuing feedback signals are often considered as the end
product of earlier stimulus-feature competition and inter-
mediate cognitive operations (Barceló & Cooper, 2018b).
In sharp contrast, in active inference, proprioceptive inputs
from actions are an integral part in the recursive exchanges
of information to disambiguate the hidden causes of sensa-
tions (see Figure 2). Hence, in active inference, cognitive
flexibility depends on a dynamic balance between inference
and learning (Maisto et al., 2019). Let us now consider some
intuitive examples to illustrate how the brain can rapidly
alternate between context learning and perceptual infer-
ence, even midtrial, in the relatively well-structured condi-
tions of WCST performance.
Whereas perceptual inference can be mainly linked to
the negative feedback, a positive feedback can also trigger
large prediction errors that elicit frontal P3a responses,
especially when there is residual uncertainty about the hid-
den category (Kopp & Lange, 2013; Cunillera et al., 2012).
Likewise, in a volatile context with frequently changing
sorting categories, target cards may also elicit an anterior
P3a indexing perceptual inference (Kopp, Lange, Howe,
& Wessel, 2014; Adrover-Roig & Barceló, 2010; Barceló,
Sanz, Molina, & Rubia, 1997). This can be best seen in
the first target after a predictive cue (Brydges & Barceló,
2018; Barceló et al., 2006), or with WCST analogues that
employ more than three sorting categories and short trial
sequences, which forces the participant to adopt an episte-
mic response policy that prevails over context learning
(Kopp et al., 2014; Stuss & Picton, 1978). Such an epistemic
policy can be fostered with WCST versions that use ambig-
uous choice cards that share more than one stimulus di-
mension with the keycards (Stuss et al., 2000; Milner,
1963). This is because ambiguous choice cards reduce
the efficacy of the participant’s actions to infer the correct
sorting category (Figure 1A).
Moreover, nonlinear interactions between hidden vari-
ables, such as the number of sorting categories and correct
trial length, may explain some adjustments in the timing of
perceptual inference. For instance, if midway through the
testing session the participant learns and becomes confi-
dent (i.e., forms precise posterior beliefs) about the con-
stant length of trial sequences, then eventually he or she
may correctly anticipate an upcoming change in category.
In this case, high-level belief updating will be endogenously
generated after the last correct card sort (Barceló et al.,
2000). An extreme example is when a mischievous partic-
ipant decides to sort the cards in the pile that matches
none of the perceptual categories (Barceló et al., 2000).
Here, one would not expect to see a frontal P3a to the con-
tinual negative feedbacks, because they are perfectly pre-
dicted by the participant. In turn, target cards will continue
to elicit parietal P3b responses, as their contents cannot be
predicted by the participant. In fact, always sorting on the
nonmatching pile provides a stable context for learning
under the same response policy, although the examiner
might find the participant’s strategy highly surprising.
This speaks to the importance of the participant’s genera-
tive model to estimate trial-by-trial changes in the “subjec-
tive surprise” associated with a sensory outcome given its
hidden causes (cf. Friston et al., 2017; Itti & Baldi, 2009;
Duncan-Johnson & Donchin, 1977). These examples illus-
trate that the dynamic balance between inference and
learning depends on the precision of prediction errors.
This can vary from trial to trial as a function of the partic-
ipant’s prior beliefs, the sensorimotor contingencies
between task events, and the interactions between con-
current hidden variables.
In healthy participants, cognitive flexibility hinges on an
adaptive and context-sensitive balance between perceptual
inference and context learning or between the exploration
of novel information versus the exploitation of familiar in-
formation (Ebitz, Sleezer, Jedema, Bradberry, & Hayden,
2019). Next, we use some more examples to show how a
well-practiced participant, who has been efficiently sorting
cards for a while, will quickly revert his or her pragmatic
policy into an epistemic policy in the face of novel, unex-
pected, or surprising information. For instance, if unknown
to the participant the examiner slips a gray card among the
regular color cards, or suddenly rings a bell, these events
will trigger perceptual inference and the accompanying
frontal P3a responses—indexing large ascending surprise
signals, very much like those elicited by the negative feed-
back (Wessel & Aron, 2017; Barceló et al., 2006).
The participant might even stop sorting and turn toward
the examiner asking for help on how to sort gray cards
( Wessel et al., 2016). Similar P3a responses are elicited if
the keyboard button gets stuck and does not lead to the
expected outcome and the closure of the perception–
action cycle. Furthermore, a very gradual change in room
luminance may go unnoticed, but if the room lights are
suddenly switched off, again the participant will likely stop
sorting and turn her head around looking to disambiguate
the hidden cause of sudden darkness. All these are com-
mon examples of epistemic affordances that elicit orienting
responses to salient sensory cues (Sokolov, 1963). In other
Barceló
1645
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
words, the very small likelihood of any such unexpected
sensory outcomes, under the current hypothesis, gener-
ates precise prediction errors that engage pFC (Barceló
& Knight, 2007a; Friedman, Cycowicz, & Gaeta, 2001).
Ignoring such surprising changes in the environment,
and to perseverate sorting the cards, could be considered
as a clear sign of cognitive inflexibility.5 In other words, per-
ceptual inference of unexpected and highly informative
distracters introduces a change in context that interrupts
ongoing behavior and leads to a reevaluation of current
goals and goal-directed action selection (Maisto et al.,
2019; Rigoli et al., 2017).
Finally, active inference could potentially offer a parsi-
monious explanation for both perseverative and set-loss
errors as lapses in control because of transient information
overload, either during inference of the sorting category or
during context learning, respectively (Dehais et al., 2019).
A perseverative error could thus result from a highly pre-
cise prior belief that has accumulated over several trials un-
der the previous sorting category (Friston et al., 2016).
Conversely, a set-loss error could be because of the low
precision of (or confidence in) posterior beliefs for the
newly acquired sorting category in the face of distracting
sensory inputs. From these premises, one would predict
perseverative card sorts to evoke large posterior P3b re-
sponses typical of context learning and, conversely, card
sorts with set-loss errors to evoke frontal P3a responses
typical of perceptual inference. Indeed, this was the main
pattern of results found when comparing P3-like responses
to target WCST cards resulting in either type of error
(Barceló, 1999). More recently, the amplitude of both
P3a and P3b responses to negative feedback cues has been
shown to be inversely correlated with the number of per-
severative errors in patients with Parkinson disease (Lange,
Seer, & Kopp, 2017; Lange et al., 2016). These findings are
consistent with the implication of the domain-general
frontoparietal control network in the regulation of both
perseverative and set-loss errors (Dehais et al., 2019). In
summary, perseverative and set-loss errors could be seen
as transient information overloads in the participant’s
exchanges with his or her testing environment either
during perceptual inference or during context learning,
respectively (Ebitz et al., 2019; Friston et al., 2017).
WHEN NEUROPSYCHOLOGY
MEETS ELECTROPHYSIOLOGY
Neuropsychologists have traditionally relied on structural
and functional brain imaging techniques to explore the
neural substrates of cognition (Stuss & Alexander, 2007).
In turn, electrophysiological techniques offer split-second
temporal resolution into the fast neural dynamics of the
inference and learning stages of perceptual categorization
as well as sufficient topographical specificity to discern
anterior P3a from posterior P3b responses (Knight, 1997).
In a series of ERP studies, we used a computerized version
of the WCST to examine the neural dynamics underlying
the inference and learning of perceptual categories. One
common theme in these studies was that negative and
positive feedback stimuli were regarded as predictive cues
that proactively prompted for a switch, or repetition, in the
ongoing sorting category in anticipation of the next card
sort (Garcia-Garcia, Barceló, Clemente, & Escera, 2011;
Periáñez & Barceló, 2009; Barceló et al., 2002, 2006;
Barceló, 2003). In one early study, Barceló et al. (2002) used
the 24 choice cards that can be unambiguously matched
with the four keycards based on only one perceptual
dimension (cf. Nelson, 1976). These 24 choice cards were
semirandomly arranged into 18 series, each ruled by a dif-
ferent sorting category. Each series contained between six
and eight target cards so as to provide a stable context for
learning and also to prevent the anticipation of the next se-
ries. A fixed cue-target interval of 1600 msec allowed suffi-
cient preparation time—and minimized the effects of
temporal uncertainty upon target detection. Target cards
were displayed for 1500 msec, or until a response was
issued, and intertrial intervals between button presses
and the next feedback varied randomly between 1500
and 2000 msec (Barceló, 2003; Figure 1B).
With these task parameters, healthy participants can
comfortably sort the cards with high accuracy rates.
The correct sorting category was initially unknown to
the participants and changed randomly from one series
to the next, so participants were forced into a perceptual
inference stage, and they had to rely on the negative and
positive feedback to disambiguate the hidden sorting cat-
egory. Our findings revealed two broad classes of P3-like
responses (Figures 3 and 4): A frontal–central P3a and a
posterior P3b that were time-locked to negative feedback
stimuli early in the card sorting series, during perceptual
inference, and showed rapid repetition suppression
(Figure 3, left). Then, a posterior P3b time-locked to
the target cards that showed repetition enhancement
during context learning later on in the card sorting series,
whenever the context remained stable for a sufficient
number of trials (Figure 3, right). Despite major differences
in testing materials and procedures, similar findings have
been consistently reported by many authors using WCST
analogues (Li et al., 2018; Lange, Seer, & Kopp, 2017;
Lange, Seer, Muller-Vahl, et al., 2017; Lange et al., 2016;
Vila-Ballo et al., 2015; Kopp et al., 2014; Kopp & Lange,
2013; Cunillera et al., 2012; Adrover-Roig & Barceló,
2010; Kopp, Tabeling, Moschner, & Wessel, 2006).
For some time, we thought that our treatment was an
original and intuitive, if unconventional, way to interpret
these two classes of P3-like responses to the feedback cues
and target cards. In those days, the prevailing ideas about
the type of P3-like responses elicited by task-relevant
(target) and task-irrelevant (distracter) stimuli in cognitive
tasks—together with mainstream ideas about the anatom-
ical substrates of the anterior and posterior attention
systems—clashed with the interpretation of our findings.
In short, we were reporting a gradually enhanced parietal
P3b upon detection of “relevant” target WCST cards, whose
1646
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
processing was then assumed to be mediated by the ante-
rior executive system (cf. Posner & Petersen, 1990).
Moreover, our frontal–central P3a to ancillary negative
feedback cues resembled novelty P3 potentials to task-
irrelevant distracters in simple oddball tasks (Polich,
2007; Friedman et al., 2001). As the reader can imagine,
it was not an easy enterprise to convince reviewers about
the soundness of our evidence, in the face of prevailing hy-
potheses about frontal lobe function and the putative roles
of these two classes of P3-like responses (Barceló et al.,
2002; Donchin & Coles, 1988).
It was in this historical context when we were fortunate
enough to meet Don Stuss for the first time: He had kindly
accepted our invitation as a keynote speaker in a confer-
ence we organized in Mallorca in 2004. In his talk, entitled
“Reflections on the Value of Frontal Lobe Tests: Suggestions
for the Future,” he offered an overview of his work on
patients with brain lesions assessed with several traditional
tests of frontal function (such as the WCST, Trail Making
Test, Stroop, and verbal fluency tasks). In their well-known
patient study on the WCST, Stuss and collaborators (2000)
largely replicated and extended Milner’s (1963) seminal
findings: Apart from confirming the sensitivity of the
WCST to left and right dorsolateral prefrontal lesions, they
also reported that a group with lesions in superior medial
frontal cortex was the most impaired on nearly every mea-
sure. In the ensuing discussion, although admitting that
such a multifactorial test as the WCST is unlikely to be sen-
sitive only to frontal lobe lesions, he advised the analysis of
the component processes involved to understand why and
how lesions in different brain regions could lead to differ-
ent types of errors in this test. In his opinion, the WCST
could still provide a reasonable index of frontal lobe func-
tioning, if there was control of some of the nonfrontal pro-
cesses involved in WCST performance (Stuss et al., 2000).
When do these nonfrontal processes intervene and how
they can be isolated from frontal processes remain pressing
questions nowadays, and thus the contents of his talk are
still very much up-to-date. In the poster session after the
conference, Don showed a special interest in one of our
ERP studies on the WCST. Initially, we thought the reason
for his interest was that the study was coauthored by his
friend and colleague Bob Knight (Stuss & Knight, 2013).
Then, pointing to the figure with the P3-like responses elic-
ited by the negative feedback (Figure 3, left), he asked this
rather cryptic question with his warm and friendly smile:
“So why do you label those two peaks as P3a and P3b?”
This question revealed in Don a much deeper knowl-
edge in cognitive electrophysiology than the typical neuro-
psychologist has, and that was how we found out about his
earlier ERP study using an analogue of the WCST as part of
his PhD thesis (Stuss & Picton, 1978). In this study, Don
and Terence reported P3-like responses to negative feed-
back cues and target cards showing a similar timing,
morphology, and scalp topography to those reported in our
2002 study. Although they labeled their feedback-locked
peaks as P3 and P4, respectively, we both agreed that these
P3-like responses were related to the resolution of uncer-
tainty about the choice of upcoming motor responses in
the next card sort and that they indexed functionally differ-
ent neural operations each (Barceló et al., 2002; Stuss &
Picton, 1978). Likewise, we both agreed that the growing
parietal target P3b to the cards indexed rehearsal and over-
learning of the same sorting rule over several correct card
sorts, although in their study, they did not find a significant
enhancement in their target P3b amplitudes probably
because of their short trial series (cf. Stuss & Picton,
1978). At the time, neither of us could call on active infer-
ence to help us interpret those two classes of P3-like
responses. In addition, none of us had quantified the
high- and low-level Bayesian surprise associated with our
feedback cues and target cards (Barceló & Cooper,
2018b; cf. Figure 2), nor how this surprise changes dynam-
ically on a trial-by-trial basis as a function of interactions
between hidden variables such as the length of trial series
or the transition probabilities between the sorting catego-
ries (Maheu, Dehaene, & Meyniel, 2019).
Looked at from the perspective of active inference, these
two ERP studies on the WCST offer some valuable insights
into prefrontal executive functions. First, there is instant
neural activation across a widely distributed fronto-
temporo-parietal cortical network in response to the feed-
back cues, with a lesser frontal involvement in response to
the ensuing target cards. Second, the feedback-locked P3a
and P3b responses seem to index two distinct predictive
processing operations in preparation for the next card sort,
given their distinct timing, scalp topographies, and re-
sponse to task variables. The candidate operations are (a)
the inference of (or resolution of uncertainty about) the
hidden sorting category and ( b) the updating of
stimulus–response mappings in preparation for the next
card sort, respectively (cf. Barceló et al., 2002; Stuss &
Picton, 1978). In particular, the frontal P3a could be seen
as an index of high-level surprise minimization during per-
ceptual categorization (cf. Rigoli et al., 2017). This agrees
with views of P3a as a cortical orienting response to novel
percepts (Polich, 2007; Sokolov, 1963). Third, the parietal
P3b to the target cards seems to index low-level surprise
minimization over posterior association cortices (as partic-
ipants cannot predict the contents of the card), and it
shows repetition enhancement as the participant learns
the task context over several trials (Figure 5). Finally, these
P3-like responses could be regarded as domain-general
signatures of perceptual inference and learning in many
cognitive tasks (Friston, 2005).
In the following sections, we will answer Don’s question
in the light of evidence supporting a functional and
anatomical dissociation between the stages of perceptual
inference and context learning during card sorting. Most
studies used analogues of the WCST, or related task-
switching paradigms (Kopp & Lange, 2013; Kopp et al.,
2006), and they normally measured P3-like responses to
both feedback cues and target cards. As shown below,
Barceló
1647
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
the active inference framework offers a radically different,
although much more flexible, dynamic and richer account
of P3a and P3b responses than conventional views in terms
of task-irrelevant (“ignore”) and task-relevant (“attend”)
stimulus conditions (Donchin & Coles, 1988).
FAST NEURAL DYNAMICS OF INFERENCE AND
LEARNING OF THE SORTING CATEGORIES
From conventional views on the anatomy of executive
attention (Posner & Petersen, 1990), we initially searched
for an ERP correlate of frontal function in response to the
target cards (Barceló et al., 1997, 2000; Barceló, 1999;
Barceló & Rubia, 1998). These studies followed prior work
by Mattes, Cohen, Berg, Canavan, and Hopmann (1991),
who measured several slow cortical potentials, including
P300, to WCST choice cards, keycards, and the feedback
cues. They compared early and late correct trials in each
card sorting series as well as early incorrect trials that lead
to the inference of the category. Quite unexpectedly at the
time, target P3b responses were neither sensitive to task
conditions nor discriminated between healthy controls
and schizophrenic patients. In contrast, negative and
positive feedback cues early in the WCST series elicited
large frontal P3a and parietal P3b responses that were
much reduced in the patients. These effects were inter-
preted in terms of impaired context updating and “inability
to use feedback information to modify subsequent behav-
ior” (Mattes et al., 1991, p. 203). Thus, although these
authors considered the feedback “at the end of the trial,”
they also assumed it could modify subsequent behavior.
Importantly, their frontal–central P3a to the feedback was
regarded as an index of context updating, despite prevail-
ing views linking context updating to the “task-relevant”
and parietally distributed target P3b (Donchin & Coles,
1988).
To further explore earlier findings of conspicuous parie-
tal target P3b responses to the WCST cards (Barceló &
Rubia, 1998; Barceló et al., 1997), the study of Barceló
et al. (2000) compared early and late card sorts within
the WCST series with two control conditions. They found
that the gradual buildup in target P3b amplitude along the
WCST series was related to the learning of the sorting
category after its endogenous inference after negative
feedback (see Figure 5). In turn, when the category was
explicitly disclosed with the first card, or when participants
always sorted cards on the nonmatching pile (thus gener-
ating continual but fully predictable negative feedbacks),
both early and late target cards elicited a full-blown parietal
P3b. These findings agree with the hypothesis that the rep-
etition enhancement of target P3b reflects a gradual
increase in the precision of endogenous predictions as
the participant becomes gradually more confident about
the ongoing sorting category and more evidence about
task parameters is accumulated along the card sorting
series (Auksztulewicz & Friston, 2016). Importantly, the
repetition enhancement of target P3b may go easily
unnoticed when all target trials are averaged together
(Mattes et al., 1991) or when the sorting category changes
frequently, as a volatile context hampers learning (Lange,
Seer, Muller-Vahl, et al., 2017; Kopp et al., 2014; Periáñez &
Barceló, 2009; Stuss & Picton, 1978). Hence, the repetition
enhancement of target P3b is best observed when ERP
waveforms are obtained from long card sorting series
without any errors (cf. Barceló et al., 2000; Figure 5).
The insensitivity of target P3b to cognitive variables trig-
gering high-level belief updating, and even to prefrontal
damage (Barceló & Knight, 2007a; Knight, 1997), eventually
led most researchers to focus on the feedback-locked
P3a and P3b responses (Li et al., 2018; Lange, Seer, &
Kopp, 2017; Lange, Seer, Muller-Vahl, et al., 2017; Lange
et al., 2016; Vila-Ballo et al., 2015; Kopp et al., 2006, 2014;
Kopp & Lange, 2013; Cunillera et al., 2012; Garcia-Garcia
et al., 2011; Periáñez & Barceló, 2009; Barceló, 2003;
Stuss & Picton, 1978). In general, these studies are consis-
tent with the hypothesis that P3a indexes the inference of
the sorting category (i.e., high-level belief updating),
whereas P3b indexes the updating of stimulus–response
mappings in preparation for the next card sort. Note that
there is a consistent delay between the peak latency of P3a
(approximately 250–350 msec) and P3b (approximately
400–800 msec).6 This speaks of two temporally distinct
stages of perceptual inference and context learning, with
inference preceding and being a prerequisite for learning
(Barceló & Cooper, 2018b; Friston, 2005). Accordingly,
frontal P3a responses would index surprise minimization
over hidden perceptual categories (e.g., response poli-
cies), whereas parietal P3b responses would index sur-
prise minimization over task parameters (e.g., stimulus–
response mappings; Friston et al., 2017).
Source localization and magnetoencephalographic
studies suggest that the early P3a aspect recruits activity
from a distributed fronto-temporo-parietal network with
key frontal nodes at the inferior frontal gyrus, middle fron-
tal gyrus, anterior insula, and ACCs (Diaz-Blancat, Garcia-
Prieto, Maestú, & Barceló, 2018; Bayless, Gaetz, Cheyne, &
Taylor, 2006; Periáñez et al., 2004), as well as the hippo-
campus for retrieval of the new sorting category (Knight,
1996) and the BG for inhibition of the old response policy
and the selection of a new one (Lange et al., 2016; Wessel
et al., 2016). In turn, the later P3b aspect recruits activity
from the TPJ, with key nodes at the supramarginal gyrus,
superior temporal gyrus, inferior parietal lobe, and precu-
neus (Diaz-Blancat et al., 2018; Bayless et al., 2006;
Periáñez et al., 2004). These structures map well onto
the functional anatomy of active inference (Friston et al.,
2017) and are broadly consistent with metabolic brain
imaging studies of the WCST (Buchsbaum et al., 2005;
Monchi, Petrides, Petre, Worsley, & Dagher, 2001;
Konishi et al., 1998). In summary, the evidence supports
the implication of a domain-general frontoparietal
network in the elicitation of feedback-locked P3a and
P3b responses during perceptual inference, compatible
with that engaged by surprising distracters in simpler
1648
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
categorization tasks, such as oddball tasks (Barceló et al.,
2006; Bledowski et al., 2004).
Methodological, procedural, and technical differences
between ERP studies can explain even gross differences
in the timing, morphology, and scalp topography of
feedback-locked P3-like responses. This is because the
magnitude of ascending surprise signals, and hence, the
relative engagement of frontal and posterior multimodal
association cortices—during actively inferring the sorting
category—critically depends on the inference and learning
of other states and task parameters, such as the number
and length of sorting categories, ISIs, and so on. Despite
this, there are remarkable consistencies across studies.
For instance, when switch and repeat precues are inter-
spersed with feedback postcues, similar P3a and P3b
responses are elicited by the switch cues, the negative
feedback, and the first positive feedback because all these
stimuli help resolve the participant’s uncertainty about the
same hidden sorting category (Vila-Ballo et al., 2015; Kopp
& Lange, 2013; Cunillera et al., 2012).
In general, the larger the number of sorting categories to
be disambiguated, the larger and more frontally distributed
the P3a responses (Kopp & Lange, 2013; Barceló et al.,
2002, 2006). In turn, larger P3b responses are elicited with
more complex tasks and stimulus–response mappings
(Barceló & Cooper, 2018a; Kopp & Lange, 2013).
However, frontal P3a and parietal P3b may also be elicited
by precues and postcues that merely prompt for a change
in the hand used for sorting the card (Kopp et al., 2006).
This evidence concurs with active inference, in that simple
sensory cues may result in high-level belief updating
whenever these resolve the participant’s uncertainty about
binary response policies conveying maximal sensorimotor
ambiguity (Kopp & Lange, 2013; Friston, 2010). This
speaks of the importance of sensorimotor contingencies
and reafference to fully account for feedback-locked P3a
and P3b responses (cf. Figure 2), over and above the sur-
prise conveyed by exteroceptive stimuli alone. Remarkably,
Stuss and Picton also interpreted their frontal P3 to negative
feedback cues in terms of context updating and the resolu-
tion of the participant’s uncertainty about the correctness
of her motor responses. Furthermore, regarding their pari-
etal P4, they pointed out that “a feedforward or ‘corollary’
discharge […] output may be generated to modify the ex-
pectancies for future perceptual or motor activity” (Stuss
& Picton, 1978, p. 157). Such an interpretation anticipated
modern views of planning as active inference (Friston et al.,
2017; Botvinick & Toussaint, 2012).
SURPRISE MINIMIZATION AND THE P300 IN
CARD SORTING
In response to Don’s question, we labeled our feedback-
locked P3a and P3b after early work on the P300 to infre-
quent sounds in “attend” conditions (Squires et al., 1975).
In fact, there has been much variability in the labeling of the
early and late aspects of feedback-locked P3-like responses
in studies with WCST analogues: P3 and P4 (Stuss & Picton,
1978), P3a and P3b (Kopp & Lange, 2013; Barceló et al.,
2002), early and late P3 (Cunillera et al., 2012; Periáñez &
Barceló, 2009), early and late novelty P3 (Garcia-Garcia
et al., 2011; Barceló et al., 2006), P3a and sustained poten-
tial (Kopp et al., 2006), P3a and sustained parietal positivity
(Lange et al., 2016; Kopp et al., 2014), and P3a and poste-
rior switch positivity (Lange, Seer, & Kopp, 2017). In some
studies, the early and late P3-like aspects are mingled, and
they are referred to as P3 and P300 despite their large
intensities over frontal scalp regions ( Vila-Ballo et al.,
2015; Mattes et al., 1991). To add up to this chaotic taxon-
omy, gross differences in the timing, morphology, and
scalp distribution of P3-like responses across studies could
raise doubts about their comparability under a strict defini-
tion of ERP component (Luck & Kappenman, 2012).
Despite these nominal discrepancies, most authors con-
sistently interpreted these frontal P3a and posterior P3b to
feedback and switch cues in WCST analogues as indexing an
updating in the contents of working memory, in reference
to the “context updating” hypothesis of P300 (Donchin &
Coles, 1988). Paradoxically, though, such an account chal-
lenges long-held views of context updating as linked to “rel-
evant” target stimuli that elicit P3b potentials with parietal
maxima (Donchin, 1981). Here, a crucial question is: When
is the task context updated? In response to the feedback
cues, during perceptual inference? Or in response to the
target stimuli, during context learning? In line with other
authors, we also agree with Stuss and Picton (1978) in that
the context updating P300 does engage frontal cortices in
response to negative feedback during perceptual categori-
zation. Crucially, this is because predictive feedback cues
are more informative and salient than target cards for ruling
complex goal-directed card sorting behavior (Barceló &
Cooper, 2018a, 2018b; Figures 2 and 6).
Hence, active inference, with its computational defini-
tion of surprise minimization, offers a solid integrative
framework to dilute lexical disparities and resolve long-
lasting controversies regarding the role of P3a and P3b
responses in context updating in terms of high- and low-
level belief updating at prefrontal and posterior multimodal
association cortices, respectively. Furthermore, these
new views can also accommodate recent accounts of sim-
ilar P3-like positivities in terms of evidence accumulation
(O’Connell, Dockree, & Kelly, 2012). Such an overarching
integrative framework is much needed for a theory-guided
interpretation of frontal and nonfrontal P3-like responses,
when examining higher cognitive functions with different
testing procedures and stimulus materials (Parr et al., 2018;
Friston, 2005; Stuss et al., 2000; Luria, 1966).
The integrative potential of active inference owes to the
same principle of surprise minimization being applied to
both inference and learning. Under these new views, inter-
oceptive and proprioceptive inputs generated by card sort-
ing actions need to be integrated with exteroceptive inputs
from the visual cards and feedback sounds as they all
inform the inference of the same perceptual category.
Barceló
1649
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Estimations of
information gain for predictive
feedback cues as a function of
the number of perceptual
categories held in working
memory. Estimates assume
information transmission
through higher [Q(rj | si)] and
lower [I(si, rj)] levels in the
neural hierarchies (cf. Figure 2).
The more perceptual categories
associated with negative
feedback cues (▪◂||), the larger
the amount of information being
conveyed by these cues for
anticipatory response (policy)
selection. Adapted from Nyhus
and Barceló (2009), with
permission.
Frontoparietal association cortices are candidate structures
for this multisensory integration (Andersen & Cui, 2009),
as they generate top–down predictions in the form of cor-
ollary discharges that suppress exteroceptive and proprio-
ceptive prediction errors to minimize surprise at both
prefrontal and posterior association cortices (Teuber,
1964/2009; cf. Figure 2). From here, feedback-locked P3a
and P3b responses can be seen as belief updates at pre-
frontal and posterior association cortices, respectively.
The former resolves the uncertainty about the new sorting
category, and the latter resolves the uncertainty about the
corresponding stimulus–response mappings in prepara-
tion for the next card sort. In both cases, these high- and
low-level belief updates are informed by beliefs about a fu-
ture desired outcome: the rewarded response policy
(Friston et al., 2017). This concurs with Bayesian accounts
of perceptual categorization (Rigoli et al., 2017) as well as
with views of planning as active inference (Botvinick &
Toussaint, 2012).
The same general principle of surprise minimization
applies to context learning once the participant is confi-
dent about the correct sorting category. In this situation,
the target P3b indexes low-level surprise minimization at
posterior association cortices because the participant
cannot predict the contents of the next target card, nor
the corresponding button press to close the perception–
action cycle. Crucially, this explains why these target P3b
responses never “habituate” (Donchin, 1981), and instead,
they show repetition enhancement, indexing a gradual
increase in the precision of predictions about many task
parameters being learned along the card sorting series
(Auksztulewicz & Friston, 2016). Furthermore, the key dif-
ference between feedback-locked P3b and target P3b is
that the former contributes to the inference of the sorting
category and is informed by beliefs about the future (pre-
diction), whereas the latter indexes context learning and is
informed by beliefs about the past (postdiction; Friston
et al., 2017). This idea concurs with dual-mode models that
define cognitive control in terms of two temporarily dis-
tinct stages of proactive and reactive control (Barceló &
Cooper, 2018a; Braver, 2012). Note that the existence of
two functionally distinct types of P3b solves a long-lasting
dialectic about two competing hypotheses of P3b elicita-
tion in terms of context updating and context closure
(Donchin & Coles, 1988; Verleger, 1988). Furthermore,
inference and learning each entail accumulation of evi-
dence over different time scales (Friston, 2005). Hence,
active inference can also accommodate evidence about a
centroparietal P3-like positivity that has been interpreted
in terms of evidence accumulation during perceptual
decision-making. The use of task designs that foster a
stable context may explain why this centroparietal posi-
tivity preferentially shows a centroparietal rather than a
frontal–central scalp distribution (O’Connell et al., 2012).
In fact, the scalp-recorded target P3b has long been re-
ferred to as a “late positive complex” consisting of manifold
component operations (Polich, 2007; Sutton & Ruchkin,
1984). Recent single-trial EEG decomposition has shown
that target P3-like responses can be divided into stimulus-
locked, response-locked, and latency variable P3-like sub-
components (Verleger, Grauhan, & Smigasiewicz, 2016),
each putatively indexing fast cycles of belief updating in
the participant’s generative model in response to within-
trial changes in exteroceptive, proprioceptive, and intero-
ceptive sensory outcomes (Brydges & Barceló, 2018).
Some of these target P3-like subcomponents show a more
frontal distribution on first target trials after predictive cues
prompting for a switch in context (Barceló & Cooper,
2018a), consistent with the larger magnitudes of prediction
errors during perceptual inference in volatile contexts. In
these cases, the frontal target P3a aspect peaks 50–150 msec
earlier than the ensuing parietal target P3b aspect
(Brydges & Barceló, 2018; cf. Donchin, 1981, Figures 15
and 16; Verleger et al., 2016), suggesting that the stage of
perceptual inference always precedes the stage of context
learning (Friston, 2005). These studies also suggest that
both high- and low-level belief updating may be endoge-
nously generated, being time-locked to neither a stimulus
nor a response (cf. Johnson & Donchin, 1985). Hence, a
finer grained decomposition of target P3-like responses
1650
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
to various sources of sensory evidence during the stages of
inference and learning of perceptual categories will foster
our understanding of the complex workings of frontopar-
ietal cortical networks subserving cognitive control (Brydges
& Barceló, 2018; Friston, 2005).
Traditional views on the P300 portray a sharp dichotomy
between anterior P3a and posterior P3b responses that is
rigidly constrained by the “ignore” versus “attend” task con-
ditions instructed by the examiner (Polich, 2007). In con-
trast, active inference defines perceptual categorization in
a much flexible, time-dynamic, and context-sensitive way
toward the efficient control of complex forms of goal-
directed behavior (Rigoli et al., 2017; Luria, 1966). One
may assume that, when certain information threshold is
surpassed, surprise signals will result in high-level belief up-
dates at pFC. This threshold seems to be a function of the
likelihood of sensory outcomes given their expected hid-
den causes, the prior probability of those causes, and their
relative precision (Friston et al., 2017; Figure 2). This new
formal scheme allows for a graded engagement of pFC dur-
ing perceptual inference and speaks to the importance of
quantifying sensorimotor information for modeling
task-averaged behavioral and brain responses and their
trial-by-trial dynamics (Barceló & Cooper, 2018a, 2018b;
cf. Figures 2 and 6).
To date, few studies on the WCST have attempted to
quantify the sensorimotor information conveyed by task
events about the hidden perceptual categories (Kopp &
Lange, 2013; Nyhus & Barceló, 2009). This shall be neces-
sary to model cortical responses in terms of information
transmission across prefrontal and posterior association
cortices during the inference and learning of the sorting
categories (Figure 2). For instance, using a task-switching
version of the WCST, Kopp and Lange (2013) reported that
frontal P3a amplitudes varied with the entropy of switch
cues, whereas posterior P3b amplitudes correlated with
the surprise of switch cues. However, such modeling
approach was based on a simple neuronal model of the
stimulus (Sokolov, 1963) and did not consider the partici-
pant’s “expectancies for future perceptual or motor ac-
tivity” (Stuss & Picton, 1978, p. 157).
Paradoxically, motor components and the reactions of
skeletal muscles were originally seen as essential character-
istics of the orienting reflex, defined as a “mechanism that
facilitates the collection and transmission of information”
(Sokolov, 1966, p. 351). Although Sokolov (1966) also
relied on Bayes’ theorem in his attempt to identify an infor-
mation threshold for elicitation of the orienting reflex, he
overlooked the importance of reafference between extero-
ceptive and proprioceptive sensory inputs for perceptual
categorization (cf. Luria, 1966). In turn, when sensorimotor
information is considered, then this threshold might well
correspond with our capacity limits for processing informa-
tion (Miller, 1956; Figure 6). This idea was examined in a
study that modeled the frontoparietal distribution of
P3-like responses to predictive cues and target stimuli in
a simplified task-switching WCST analogue (Barceló &
Cooper, 2018a). This study reported the largest frontal
P3a responses to predictive cues in the most complex task
condition conveying maximal sensorimotor information
for anticipatory response (policy) selection during percep-
tual inference. In contrast, the largest reactive target P3b
responses were obtained in the simplest task condition,
consistent with maximal precision of predictions during
context learning (Barceló & Cooper, 2018b).
In summary, the evidence reviewed here supports the
notion of two broad classes of P3-like responses that index
high- and low-level belief updating at prefrontal and poste-
rior association cortices during the inference and learning
of perceptual categories. Further modeling work will be
needed to examine the existence of some information
threshold in a participant’s generative model for the elici-
tation of frontally distributed P3-like responses during
perceptual categorization (Friston, 2005; Sokolov, 1966;
see Figure 6).
CONCLUSIONS
The WCST has been one of the most distinctive tools for
the assessment of frontal lobe functions in clinical and
research contexts. Its extensive use has generated a very
rich database of behavioral and brain imaging results, some
of which seemed paradoxical or contradictory under tradi-
tional theories of executive attention and frontal lobe func-
tion. In this paper, we aimed to solve some of those
paradoxes adopting a new theory of cortical responses
inspired on the Bayesian brain hypothesis and the free
energy principle (Friston, 2005, 2010). The new theory
can explain changes in the frontoparietal scalp distribution
of two broad classes of P300 cortical responses that had
long been associated with uncertainty resolution (Sutton
et al., 1965) and memory consolidation (Donchin, 1981)
in many cognitive domains. The reviewed evidence sug-
gests that, when sorting cards, there is instant activation
across a distributed frontoparietal network (Barceló et al.,
2002; Stuss & Picton, 1978), which jointly with other
cortical and subcortical structures ( Wessel et al., 2016;
Buchsbaum et al., 2005; Knight, 1996), is differentially
engaged during two temporally distinct stages: (1) during
the inference of the hidden perceptual category and (2)
during learning of the task context or task parameters
(Friston, 2005).
In retrospect, it is striking that Don and Terence’s inter-
pretation of their frontal–central P300 responses to the
feedback cues in terms of context updating was prescient
of modern views of planning as active inference. Much
confusion in the literature owes to rigid attempts to de-
fine a unitary “task-relevant” P300 component based on
its maximal parietal intensity (Donchin & Coles, 1988). In
contrast, active inference offers a more flexible and richer
picture of P3-like cortical responses as proxies of neural
activity in a frontoparietal network that is differentially
engaged during both inference and learning of perceptual
categories. However, to date, few studies have attempted
Barceló
1651
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
to dissociate these two stages, which may have been inad-
vertently combined in the task-averaged ERP waveforms.
This confound would foreseeably produce posterior P3b
responses with parietal maxima indexing context learning,
together with earlier frontal–central P3a responses with
lesser intensities indexing perceptual inference in a pro-
portion of the averaged trials (cf. Barceló et al., 2006,
Figure 2; Barceló et al., 1997, Figure 1; Donchin, 1981,
Figures 4, 15, and 16; Duncan-Johnson & Donchin, 1977,
Figure 1B and C; Johnson & Donchin, 1985, Figures 1–3).
Future research could aim to undo potential confounds
in the recording of P3-like responses during the stages of
perceptual inference and learning in cognitive tasks. For
this, it will be important to model Bayesian surprise and
to identify an information threshold above which there will
be a gradually larger frontal involvement in the elicitation of
anterior P300 responses during perceptual inference
(Barceló & Cooper, 2018a; Sokolov, 1966). In this respect,
the Bayesian brain hypothesis reinstates pioneering infor-
mation processing views in cognitive neuroscience (Miller,
1956; Figure 6). A quantitative estimation of such a thresh-
old would allow researchers to examine how this quantity
depends on individual variables such as age, memory span,
fluid intelligence, or brain damage. Furthermore, an accu-
rate report of P3-like responses requires their dissociation
from other overlapping cortical responses, like negative ex-
pectancy waves (Gajewski et al., 2018; Luck & Kappenman,
2012; Stuss & Picton, 1978).
From these new premises, frontal and nonfrontal P3-like
responses offer promising biomarkers of epistemic (explor-
atory) and pragmatic (exploitative) behavior during the
inference and learning of perceptual categories in card sort-
ing and, more generally, as basic tools in the neuropsycho-
logical assessment of higher cortical functions in humans
(Parr et al., 2018; Stuss & Benson, 1984; Luria, 1966).
Acknowledgments
The author wishes to express his gratitude to Karl Friston for his
many edits, clarifying comments, and valuable suggestions on an
early version of this paper. Thanks also to the reviewers and editor
of this Special Issue for their thoughtful feedback.
Reprint requests should be sent to Francisco Barceló, Laboratory
of Neuropsychology, Cra. Valldemossa, km 7, 5 – University of the
Balearic Islands, Mallorca, Spain, or via e-mail: f.barcelo@uib.es.
Funding Information
This work was funded by the Spanish Ministry of Science
and Innovation (PID2019-106045GB-I00/AEI/10.13039/
501100011033).
Notes
1. Technically, in information theory, surprise is known as
self-information and expected surprise corresponds to entropy.
Entropy is a measure of uncertainty. This means that minimiz-
ing expected surprise corresponds to resolving uncertainty (cf.
Sutton et al., 1965).
2. Precision is an important quantity that can be read as the
complement of uncertainty (i.e., the precision of a Gaussian
probability density is the inverse of its variance or dispersion).
3. Technically, the Bayesian surprise can be associated with a
precision-weighted prediction error. In other words, the degree
of belief updating in predictive processing schemes is deter-
mined by the magnitude of prediction errors weighted by their
precision. This means a precise prediction error will have more
influence on belief updating as it ascends the cortical hierarchy.
In turn, this is usually interpreted in terms of a larger evoked
response as measured electrophysiologically (K. Friston, per-
sonal communication, 2020).
4. The conventional term “habituation” is not fully appropri-
ate because it normally refers to a type of nonassociative learn-
ing, whereas P3-like responses are best described in terms of
classical Hebbian associative learning in the cortical hierarchies
(Friston, 2010).
In active inference, there is an intimate relationship be-
5.
tween cognitive flexibility and the deployment of executive at-
tention. This follows because attention is thought to be
mediated by setting the precision of various beliefs at different
levels in the cortical hierarchies. Please see Parr and Friston
(2017) and Parr et al. (2019) for details (K. Friston, personal
communication, 2020).
6. The earlier latency of P3a has been attributed to bidirec-
tional hyperdirect prefrontotectal pathways ( Wessel et al.,
2016; Barceló & Knight, 2007a, 2007b). These hyperdirect path-
ways convey fast prior information about the spatio-temporal
context of incoming sensory signals ascending through regular
geniculocortical pathways. In other words, pFC holds prior be-
liefs about whether the incoming sensory evidence belongs to
the immediate perceptual context—and is thus part of active
working memory—or else, whether it is something completely
unexpected that calls for new perceptual inference and the elic-
itation of an orienting response (Sokolov, 1963). For a review of
frontal connectivity with tectal, thalamic, and other brainstem
structures, see Stuss and Benson (1984).
REFERENCES
Adrover-Roig, D., & Barceló, F. (2010). Individual differences in
aging and cognitive control modulate the neural indexes of
context updating and maintenance during task switching.
Cortex, 46, 434–450. DOI: https://doi.org/10.1016/j.cortex
.2009.09.012, PMID: 19889406
Andersen, R. A., & Cui, H. (2009). Intention, action planning,
and decision making in parietal–frontal circuits. Neuron, 63,
568–583. DOI: https://doi.org/10.1016/j.neuron.2009.08.028,
PMID: 19755101
Auksztulewicz, R., & Friston, K. (2016). Repetition suppression
and its contextual determinants in predictive coding. Cortex,
80, 125–140. DOI: https://doi.org/10.1016/j.cortex.2015
.11.024, PMID: 26861557, PMCID: PMC5405056
Badre, D., & Nee, D. E. (2018). Frontal cortex and the hierarchical
control of behavior. Trends in Cognitive Sciences, 22, 170–188.
DOI: https://doi.org/10.1016/j.tics.2017.11.005, PMID:
29229206, PMCID: PMC5841250
Barceló, F. (1999). Electrophysiological evidence of two different
types of error in the Wisconsin card sorting test. NeuroReport,
10, 1299–1303. DOI: https://doi.org/10.1097/00001756
-199904260-00027, PMID: 10363943
Barceló, F. (2003). The Madrid card sorting test (MCST): a task
switching paradigm to study executive attention with event-
related potentials. Brain Research Protocols, 11, 27–37. DOI:
https://doi.org/10.1016/S1385-299X(03)00013-8
Barceló, F., & Cooper, P. S. (2018a). An information theory
account of late frontoparietal ERP positivities in cognitive
1652
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
control. Psychophysiology, 55, e12814. DOI: https://doi.org
/10.1111/psyp.12814, PMID: 28295342
Barceló, F., & Cooper, P. S. (2018b). Quantifying contextual
information for cognitive control. Frontiers in Psychology, 9,
1693. DOI: https://doi.org/10.3389/fpsyg.2018.01693, PMID:
30250445, PMCID: PMC6139323
Barceló, F., Escera, C., Corral, M. J., & Periánez, J. A. (2006).
Task switching and novelty processing activate a common
neural network for cognitive control. Journal of Cognitive
Neuroscience, 18, 1734–1748. DOI: https://doi.org/10.1162
/jocn.2006.18.10.1734, PMID: 17014377
Barceló, F., & Knight, R. T. (2002). Both random and
perseverative errors underlie WCST deficits in prefrontal
patients. Neuropsychologia, 40, 349–356. DOI: https://
doi.org/10.1016/S0028-3932(01)00110-5
Barceló, F., & Knight, R. T. (2007a). An information–theoretical
approach to contextual processing in the human brain:
evidence from prefrontal lesions. Cerebral Cortex,
17(Suppl. 1), i51–i60. DOI: https://doi.org/10.1093/cercor
/bhm111, PMID: 17726004
Barceló, F., & Knight, R. T. (2007b). Theoretical sequelae of a
chronic neglect and unawareness of prefrontotectal pathways
in the human brain. Behavioral and Brain Sciences, 30,
83–85. DOI: https://doi.org/10.1017/S0140525X07000921
Barceló, F., Muñoz-Céspedes, J. M., Pozo, M. A., & Rubia, F. J.
(2000). Attentional set shifting modulates the target P3b
response in the Wisconsin card sorting test. Neuropsychologia,
38, 1342–1355. DOI: https://doi.org/10.1016/S0028-3932(00)
00046-4
Barceló, F., Periáñez, J. A., & Knight, R. T. (2002). Think
differently: A brain orienting response to task novelty.
NeuroReport, 13, 1887–1892. DOI: https://doi.org/10.1097
/00001756-200210280-00011, PMID: 12395085
Barceló, F., & Rubia, F. J. (1998). Non-frontal P3b-like activity
evoked by the Wisconsin Card Sorting Test. NeuroReport, 9,
747–751. DOI: https://doi.org/10.1097/00001756-199803090
-00034, PMID: 9559950
Barceló, F., Sanz, M., Molina, V., & Rubia, F. J. (1997). The
Wisconsin card sorting test and the assessment of frontal
function: A validation study with event-related potentials.
Neuropsychologia, 35, 399–408. DOI: https://doi.org/10
.1016/S0028-3932(96)00096-6
Bayless, S. J., Gaetz, W. C., Cheyne, D. O., & Taylor, M. J. (2006).
Spatiotemporal analysis of feedback processing during a card
sorting task using spatially filtered MEG. Neuroscience
Letters, 410, 31–36. DOI: https://doi.org/10.1016/j.neulet.2006
.09.073, PMID: 17055155
Bledowski, C., Prvulovic, D., Hoechstetter, K., Scherg, M.,
Wibral, M., Goebel, R., et al. (2004). Localizing P300
generators in visual target and distractor processing: A
combined event-related potential and functional magnetic
resonance imaging study. Journal of Neuroscience, 24,
9353–9360. DOI: https://doi.org/10.1523/JNEUROSCI.1897
-04.2004, PMID: 15496671, PMCID: PMC6730097
Botvinick, M., & Toussaint, M. (2012). Planning as inference.
Trends in Cognitive Sciences, 16, 485–488. DOI: https://
doi.org/10.1016/j.tics.2012.08.006, PMID: 22940577
Braver, T. S. (2012). The variable nature of cognitive control: A
dual mechanisms framework. Trends in Cognitive Sciences,
16, 106–113. DOI: https://doi.org/10.1016/j.tics.2011.12.010,
PMID: 22245618, PMCID: PMC3289517
Brydges, C. R., & Barceló, F. (2018). Functional dissociation
of latency-variable, stimulus- and response-locked target P3
sub-components in task-switching. Frontiers in Human
Neuroscience, 12, 60. DOI: https://doi.org/10.3389/fnhum
.2018.00060, PMID: 29515383, PMCID: PMC5826240
Buchsbaum, B. R., Greer, S., Chang, W. L., & Berman, K. F. (2005).
Meta-analysis of neuroimaging studies of the Wisconsin
card-sorting task and component processes. Human Brain
Mapping, 25, 35–45. DOI: https://doi.org/10.1002/hbm.20128,
PMID: 15846821, PMCID: PMC6871753
Clark, A. (2013). Whatever next? Predictive brains, situated
agents, and the future of cognitive science. Behavioral and
Brain Sciences, 36, 181–204. DOI: https://doi.org/10.1017
/S0140525X12000477, PMID: 23663408
Cunillera, T., Fuentemilla, L., Perianez, J., Marco-Pallares, J.,
Kramer, U. M., Camara, E., et al. (2012). Brain oscillatory
activity associated with task switching and feedback
processing. Cognitive, Affective & Behavioral Neuroscience,
12, 16–33. DOI: https://doi.org/10.3758/s13415-011-0075-5,
PMID: 22160843
Dehais, F., Hodgetts, H. M., Causse, M., Behrend, J., Durantin,
G., & Tremblay, S. (2019). Momentary lapse of control:
A cognitive continuum approach to understanding and
mitigating perseveration in human error. Neuroscience &
Biobehavioral Reviews, 100, 252–262. DOI: https://doi.org
/10.1016/j.neubiorev.2019.03.006, PMID: 30878500
Demakis, G. J. (2003). A meta-analytic review of the sensitivity
of the Wisconsin card sorting test to frontal and lateralized
frontal brain damage. Neuropsychology, 17, 255–264.
DOI: https://doi.org/10.1037/0894-4105.17.2.255, PMID:
12803431
Diamond, A. (2013). Executive functions. Annual Review of
Psychology, 64, 135–168. DOI: https://doi.org/10.1146
/annurev-psych-113011-143750, PMID: 23020641, PMCID:
PMC4084861
Diaz-Blancat, G., Garcia-Prieto, J., Maestú, F., & Barceló, F.
(2018). Fast neural dynamics of proactive cognitive control in
a task-switching analogue of the Wisconsin card sorting test.
Brain Topography, 31, 407–418. DOI: https://doi.org/10
.1007/s10548-017-0607-6, PMID: 29124546
Donchin, E. (1981). Surprise! Surprise? Psychophysiology, 18,
493–513. DOI: https://doi.org/10.1111/j.1469-8986.1981
.tb01815.x, PMID: 7280146
Donchin, E., & Coles, M. G. H. (1988). Is the P300 component a
manifestation of context updating? Behavioral and Brain
Sciences, 11, 343–356. DOI: https://doi.org/10.1017
/S0140525X00058027
Doya, K., & Ishii, K. (2007). A probability primer. In K. Doya, S.
Ishii, A. Pouget, & R. P. N. Rao (Eds.), Bayesian brain:
Probabilistic approaches to neural coding (pp. 3–13).
Cambridge, MA: MIT Press. DOI: https://doi.org/10.7551
/mitpress/9780262042383.003.0001
Duncan-Johnson, C. C., & Donchin, E. (1977). On quantifying
surprise: The variation of event-related potentials with
subjective probability. Psychophysiology, 14, 456–467. DOI:
https://doi.org/10.1111/j.1469-8986.1977.tb01312.x, PMID:
905483
Ebitz, R. B., Sleezer, B. J., Jedema, H. P., Bradberry, C. W., &
Hayden, B. Y. (2019). Tonic exploration governs both
flexibility and lapses. PLoS Computational Biology, 15,
e1007475. DOI: https://doi.org/10.1371/journal.pcbi.1007475,
PMID: 31703063, PMCID: PMC6867658
Friedman, D., Cycowicz, Y. M., & Gaeta, H. (2001). The novelty
P3: An event-related brain potential (ERP) sign of the brain’s
evaluation of novelty. Neuroscience & Biobehavioral
Reviews, 25, 355–373. DOI: https://doi.org/10.1016/S0149
-7634(01)00019-7
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 360, 815–836. DOI: https://doi.org
/10.1098/rstb.2005.1622, PMID: 15937014, PMCID:
PMC1569488
Friston, K. (2010). The free-energy principle: A unified brain
theory? Nature Reviews in Neuroscience, 11, 127–138. DOI:
https://doi.org/10.1038/nrn2787, PMID: 20068583
Barceló
1653
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Friston, K. (2019). Waves of prediction. PLoS Biology, 17,
e3000426. DOI: https://doi.org/10.1371/journal.pbio.3000426,
PMID: 31581195
Friston, K., Adams, R. A., Perrinet, L., & Breakspear, M. (2012).
Perceptions as hypotheses: Saccades as experiments. Frontiers
in Psychology, 3, 151. DOI: https://doi.org/10.3389/fpsyg.2012
.00151, PMID: 22654776, PMCID: PMC3361132
Friston, K., FitzGerald, T., Rigoli, F., Schwartenbeck, P.,
O’Doherty, J., & Pezzulo, G. (2016). Active inference and
learning. Neuroscience & Biobehavioral Reviews, 68,
862–879. DOI: https://doi.org/10.1016/j.neubiorev.2016
.06.022, PMID: 27375276, PMCID: PMC5167251
Friston, K., FitzGerald, T., Rigoli, F., Schwartenbeck, P., &
Pezzulo, G. (2017). Active inference: A process theory.
Neural Computation, 29, 1–49. DOI: https://doi.org/10
.1162/NECO_a_00912, PMID: 27870614
Friston, K., Shiner, T., FitzGerald, T., Galea, J. M., Adams, R.,
Brown, H., et al. (2012). Dopamine, affordance and active
inference. PLoS Computational Biology, 8, e1002327.
DOI: https://doi.org/10.1371/journal.pcbi.1002327, PMID:
22241972, PMCID: PMC3252266
Fuster, J. M. (2013). Cognitive functions of the prefrontal
cortex. In D. T. Stuss & R. T. Knight (Eds.), Principles of
frontal lobe function (2nd ed., pp. 11–22). New York:
Oxford University Press. DOI: https://doi.org/10.1093/med
/9780199837755.003.0002
Fuster, J. M. (2019). The prefrontal cortex in the neurology
clinic. In M. D’Esposito & J. H. Grafman (Eds.), Handbook
of clinical neurology: The frontal lobes (3rd ed., Vol. 163,
pp. 3–15). Amsterdam: Elsevier. DOI: https://doi.org
/10.1016/B978-0-12-804281-6.00001-X, PMID: 31590737
Gajewski, P. D., Ferdinand, N. K., Kray, J., & Falkenstein, M.
(2018). Understanding sources of adult age differences in
task switching: Evidence from behavioral and ERP studies.
Neuroscience & Biobehavioral Reviews, 92, 255–275. DOI:
https://doi.org/10.1016/j.neubiorev.2018.05.029, PMID:
29885425
Garcia-Garcia, M., Barceló, F., Clemente, I. C., & Escera, C.
(2011). COMT and ANKK1 gene–gene interaction modulates
contextual updating of mental representations. Neuroimage,
56, 1641–1647. DOI: https://doi.org/10.1016/j.neuroimage
.2011.02.053, PMID: 21352928
Glascher, J., Adolphs, R., & Tranel, D. (2019). Model-based
lesion mapping of cognitive control using the Wisconsin Card
Sorting Test. Nature Communications, 10, 20. DOI: https://
doi.org/10.1038/s41467-018-07912-5, PMID: 30604744,
PMCID: PMC6318292
Hohwy, J. (2016). The self-evidencing brain. Noûs, 50, 259–285.
DOI: https://doi.org/10.1111/nous.12062
Hohwy, J. (2019). Prediction error minimization in the brain. In
M. Sprevak & M. Colombo (Eds.), The Routledge handbook
of the computational mind (pp. 159–172). Abingdon Oxon,
UK: Routledge.
Itti, L., & Baldi, P. (2009). Bayesian surprise attracts human
attention. Vision Research, 49, 1295–1306. DOI: https://doi
.org/10.1016/j.visres.2008.09.007, PMID: 18834898
Johnson, R., Jr., & Donchin, E. (1985). Second thoughts: Multiple
P300s elicited by a single stimulus. Psychophysiology, 22,
182–194. DOI: https://doi.org/10.1111/j.1469-8986.1985
.tb01584.x, PMID: 3991845
Knight, R. T. (1996). Contribution of human hippocampal
region to novelty detection. Nature, 383, 256–259. DOI:
https://doi.org/10.1038/383256a0, PMID: 8805701
Knight, R. T. (1997). A distributed cortical network for visual
attention. Journal of Cognitive Neuroscience, 9, 75–91. DOI:
https://doi.org/10.1162/jocn.1997.9.1.75, PMID: 23968181
Koechlin, E., & Summerfield, C. (2007). An information
theoretical approach to prefrontal executive function. Trends
in Cognitive Sciences, 11, 229–235. DOI: https://doi.org/10
.1016/j.tics.2007.04.005, PMID: 17475536
Konishi, S., Nakajima, K., Uchida, I., Kameyama, M., Nakahara,
K., Sekihara, K., et al. (1998). Transient activation of inferior
prefrontal cortex during cognitive set shifting. Nature
Neuroscience, 1, 80–84. DOI: https://doi.org/10.1038/283,
PMID: 10195114
Kopp, B., & Lange, F. (2013). Electrophysiological indicators of
surprise and entropy in dynamic task-switching environments.
Frontiers in Human Neuroscience, 7, 300. DOI: https://doi
.org/10.3389/fnhum.2013.00300, PMID: 23840183, PMCID:
PMC3699721
Kopp, B., Lange, F., Howe, J., & Wessel, K. (2014). Age-related
changes in neural recruitment for cognitive control. Brain
and Cognition, 85, 209–219. DOI: https://doi.org/10.1016
/j.bandc.2013.12.008, PMID: 24434022
Kopp, B., Tabeling, S., Moschner, C., & Wessel, K. (2006).
Fractionating the neural mechanisms of cognitive control.
Journal of Cognitive Neuroscience, 18, 949–965. DOI:
https://doi.org/10.1162/jocn.2006.18.6.949, PMID: 16839302
Lange, F., Seer, C., & Kopp, B. (2017). Cognitive flexibility in
neurological disorders: Cognitive components and event-
related potentials. Neuroscience & Biobehavioral Reviews,
83, 496–507. DOI: https://doi.org/10.1016/j.neubiorev.2017
.09.011, PMID: 28903059
Lange, F., Seer, C., Loens, S., Wegner, F., Schrader, C., Dressler,
D., et al. (2016). Neural mechanisms underlying cognitive
inflexibility in Parkinson’s disease. Neuropsychologia, 93,
142–150. DOI: https://doi.org/10.1016/j.neuropsychologia
.2016.09.021, PMID: 27693667
Lange, F., Seer, C., Muller-Vahl, K., & Kopp, B. (2017). Cognitive
flexibility and its electrophysiological correlates in Gilles de la
Tourette syndrome. Developmental Cognitive Neuroscience,
27, 78–90. DOI: https://doi.org/10.1016/j.dcn.2017.08.008,
PMID: 28863370, PMCID: PMC6987949
Li, F., Wang, J., Du, B., & Cao, B. (2018). Electrophysiological
response to the informative value of feedback revealed in a
segmented Wisconsin card sorting test. Frontiers in Psychology,
9, 57. DOI: https://doi.org/10.3389/fpsyg.2018.00057, PMID:
29459841, PMCID: PMC5807434
Luck, S. J., & Kappenman, E. S. (2012). Handbook of event-
related potential components. Oxford: Oxford University
Press. DOI: https://doi.org/10.1093/oxfordhb/9780195374148
.001.0001
Luria, A. R. (1966). Higher cortical functions in man. London:
Tavistock Publications.
Maheu, M., Dehaene, S., & Meyniel, F. (2019). Brain signatures
of a multiscale process of sequence learning in humans. eLife,
8, e41541. DOI: https://doi.org/10.7554/eLife.41541, PMID:
30714904, PMCID: PMC6361584
Maisto, D., Friston, K., & Pezzulo, G. (2019). Caching mechanisms
for habit formation in active inference. Neurocomputing, 359,
298–314. DOI: https://doi.org/10.1016/j.neucom.2019.05.083,
PMID: 32055104, PMCID: PMC7001981
Mattes, R., Cohen, R., Berg, P., Canavan, A. G. M., & Hopmann,
G. (1991). Slow cortical potentials (SCPS) in schizophrenic
patients during performance of the Wisconsin Card-Sorting
Test ( WCST). Neuropsychologia, 29, 195–205. DOI: https://
doi.org/10.1016/0028-3932(91)90021-Y
Miller, G. A. (1956). The magical number seven plus or
minus two: Some limits on our capacity for processing
information. Psychological Review, 63, 81–97. DOI:
https://doi.org/10.1146/annurev.neuro.24.1.167, PMID:
11283309
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202. DOI: https://doi.org/10.1037/h0043158, PMID:
13310704
1654
Journal of Cognitive Neuroscience
Volume 33, Number 9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Milner, B. (1963). Effects of different brain lesions on card
sorting. Archives of Neurology, 9, 100–110. DOI: https://
doi.org/10.1001/archneur.1963.00460070100010
Monchi, O., Petrides, M., Petre, V., Worsley, K., & Dagher, A.
(2001). Wisconsin Card Sorting revisited: Distinct neural
circuits participating in different stages of the task identified
by event-related functional magnetic resonance imaging.
Journal of Neuroscience, 21, 7733–7741. DOI: https://doi
.org/10.1523/JNEUROSCI.21-19-07733.2001, PMID: 11567063,
PMCID: PMC6762921
Nelson, H. E. (1976). A modified card sorting test sensitive to
frontal lobe defects. Cortex, 12, 313–324. DOI: https://doi
.org/10.1016/S0010-9452(76)80035-4
Norman, D. A., & Shallice, T. (1986). Attention to action: Willed
and automatic control of behavior. In R. J. Davidson, G. E.
Schwartz, & D. Shapiro (Eds.), Consciousness and self-
regulation (Vol. 4). New York: Plenum. DOI: https://doi.org
/10.1007/978-1-4757-0629-1_1
Nyhus, E., & Barceló, F. (2009). The Wisconsin Card Sorting
Test and the cognitive assessment of prefrontal executive
functions: A critical update. Brain and Cognition, 71,
437–451. DOI: https://doi.org/10.1016/j.bandc.2009.03.005,
PMID: 19375839
O’Connell, R. G., Dockree, P. M., & Kelly, S. P. (2012). A
supramodal accumulation-to-bound signal that determines
perceptual decisions in humans. Nature Neuroscience, 15,
1729–1735. DOI: https://doi.org/10.1038/nn.3248, PMID:
23103963
O’Regan, J. K., & Noe, A. (2001). A sensorimotor account of
vision and visual consciousness. Behavioral and Brain
Sciences, 24, 939–973. DOI: https://doi.org/10.1017
/S0140525X01000115, PMID: 12239892
Parr, T., & Friston, K. J. (2017). Working memory, attention, and
salience in active inference. Scientic Reports, 7, 14678.
DOI: https://doi.org/10.1038/s41598-017-15249-0, PMID:
29116142, PMCID: PMC5676961
Parr, T., Rees, G., & Friston, K. J. (2018). Computational
neuropsychology and Bayesian inference. Frontiers in
Human Neuroscience, 12, 61. DOI: https://doi.org/10
.3389/fnhum.2018.00061, PMID: 29527157, PMCID:
PMC5829460
Parr, T., Rikhye, R. V., Halassa, M. M., & Friston, K. J. (2019).
Prefrontal computation as active inference. Cerebral Cortex,
30, 682–695. DOI: https://doi.org/10.1093/cercor/bhz118,
PMID: 31298270, PMCID: PMC7444741
Periáñez, J. A., & Barceló, F. (2009). Updating sensory versus
task representations during task-switching: Insights from
cognitive brain potentials in humans. Neuropsychologia, 47,
1160–1172. DOI: https://doi.org/10.1016/j.neuropsychologia
.2009.01.014, PMID: 19350711
Periáñez, J. A., Maestu, F., Barcelo, F., Fernandez, A., Amo, C., &
Ortiz Alonso, T. (2004). Spatiotemporal brain dynamics
during preparatory set shifting: MEG evidence. Neuroimage,
21, 687–695. DOI: https://doi.org/10.1016/j.neuroimage
.2003.10.008, PMID: 14980570
Polich, J. (2007). Updating P300: An integrative theory of P3a
and P3b. Clinical Neurophysiology, 118, 2128–2148. DOI:
https://doi.org/10.1016/j.clinph.2007.04.019, PMID: 17573239,
PMCID: PMC2715154
Posner, M. I., & Petersen, S. E. (1990). The attention system of the
human brain. Annual Review of Neuroscience, 13, 25–42.
DOI: https://doi.org/10.1146/annurev.ne.13.030190.000325,
PMID: 2183676
Rigoli, F., Pezzulo, G., Dolan, R., & Friston, K. (2017). A
goal-directed Bayesian framework for categorization.
Frontiers in Psychology, 8, 408. DOI: https://doi.org
/10.3389/fpsyg.2017.00408, PMID: 28382008, PMCID:
PMC5360703
Shallice, T., Stuss, D. T., Picton, T. W., Alexander, M. P., &
Gillingham, S. (2008). Mapping task switching in frontal
cortex through neuropsychological group studies. Frontiers
in Neuroscience, 2, 79–85. DOI: https://doi.org/10.3389
/neuro.01.013.2008, PMID: 18982110, PMCID: PMC2570079
Sokolov, E. N. (1963). Perception and the conditioned reflex.
Oxford: Pergamon Press.
Sokolov, E. N. (1966). Orienting reflex as information regulator.
In A. N. Leontiev, A. R. Luria, & A. A. Smirnov (Eds.),
Psychological research in the U.S.S.R. (pp. 334–360).
Moscow: Progress Publishers.
Squires, N. K., Squires, K. C., & Hillyard, S. A. (1975). Two varieties
of long-latency positive waves evoked by unpredictable
auditory stimuli in man. Electroencephalography and Clinical
Neurophysiology, 38, 387–401. DOI: https://doi.org/10.1016
/0013-4694(75)90263-1
Stuss, D. T., & Alexander, M. P. (2000). Executive functions and
the frontal lobes: A conceptual view. Psychological Research,
63, 289–298. DOI: https://doi.org/10.1007/s004269900007,
PMID: 11004882
Stuss, D. T., & Alexander, M. P. (2007). Is there a dysexecutive
syndrome? Philosophical Transactions of the Royal Society
of London, Series B: Biological Sciences, 362, 901–915. DOI:
https://doi.org/10.1098/rstb.2007.2096, PMID: 17412679,
PMCID: PMC2430005
Stuss, D. T., & Benson, D. F. (1984). Neuropsychological
studies of the frontal lobes. Psychological Bulletin, 95, 3–28.
DOI: https://doi.org/10.1037/0033-2909.95.1.3, PMID:
6544432
Stuss, D. T., & Knight, R. T. (Eds.). (2013). Principles of
frontal lobe function (2nd ed.). New York: Oxford
University Press. DOI: https://doi.org/10.1093/med
/9780199837755.001.0001
Stuss, D. T., Levine, B., Alexander, M. P., Hong, J., Palumbo, C.,
Hamer, L., et al. (2000). Wisconsin card sorting test
performance in patients with focal frontal posterior brain
damage: Effects of lesion location and test structure on
separable cognitive. Neuropsychologia, 38, 388–402. DOI:
https://doi.org/10.1016/S0028-3932(99)00093-7
Stuss, D. T., & Picton, T. W. (1978). Neurophysiological correlates
of human concept formation. Behavioral Biology, 23, 135–162.
DOI: https://doi.org/10.1016/S0091-6773(78)91734-0
Stuss, D. T., Picton, T. W., & Alexander, D. C. (2001).
Consciousness, self-awareness, and the frontal lobes. In S. P.
Salloway, P. F. Malloy, & J. D. Duffy (Eds.), The frontal lobes
and neuropsychiatric illness (pp. 101–109). Washington,
DC: American Psychiatric Publishing.
Stuss, D. T., Shallice, T., Alexander, M. P., & Picton, T. W.
(1995). A multidisciplinary approach to anterior attentional
functions. Annals of the New York Academy of Sciences, 769,
191–211. DOI: https://doi.org/10.1111/j.1749-6632.1995.
tb38140.x, PMID: 8595026
Sutton, S., Braren, M., Zubin, J., & John, E. R. (1965). Evoked-
potential correlates of stimulus uncertainty. Science, 150,
1187–1188. DOI: https://doi.org/10.1126/science.150.3700
.1187, PMID: 5852977
Sutton, S., & Ruchkin, D. S. (1984). The late positive complex.
Advances and new problems. Annals of the New York
Academy of Science, 425, 1–23. DOI: https://doi.org/10.1111
/j.1749-6632.1984.tb23520.x, PMID: 6588819
Teuber, H. L. (1964/2009). The riddle of frontal lobe function in
man. Neuropsychology Review, 19, 25–46. DOI: https://doi
.org/10.1007/s11065-009-9088-z, PMID: 19459235
Verleger, R. (1988). Event-related potentials and cognition:
A critique of the context updating hypothesis and an
alternative interpretation of P3. Behavioral and Brain
Sciences, 11, 343–356. DOI: https://doi.org/10.1017
/S0140525X00058015
Barceló
1655
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Verleger, R., Grauhan, N., & Smigasiewicz, K. (2016). Is P3 a
strategic or a tactical component? Relationships of P3 sub-
components to response times in oddball tasks with go, no-go
and choice responses. Neuroimage, 143, 223–234. DOI: https://
doi.org/10.1016/j.neuroimage.2016.08.049, PMID: 27570107
Vila-Ballo, A., Cunillera, T., Rostan, C., Hdez-Lafuente, P.,
Fuentemilla, L., & Rodriguez-Fornells, A. (2015).
Neurophysiological correlates of cognitive flexibility and
feedback processing in violent juvenile offenders. Brain
Research, 1610, 98–109. DOI: https://doi.org/10.1016
/j.brainres.2015.03.040, PMID: 25839762
Wang, J., Cao, B., Cai, X., Gao, H., & Li, F. (2015). Brain
activation of negative feedback in rule acquisition revealed
in a segmented Wisconsin card sorting test. PLoS One,
10, e0140731. DOI: https://doi.org/10.1371/journal.pone
.0140731, PMID: 26469519, PMCID: PMC4607368
Wessel, J. R., & Aron, A. R. (2017). On the globality of motor
suppression: Unexpected events and their influence on
behavior and Cognition. Neuron, 93, 259–280. DOI: https://
doi.org/10.1016/j.neuron.2016.12.013, PMID: 28103476,
PMCID: PMC5260803
Wessel, J. R., Jenkinson, N., Brittain, J. S., Voets, S. H., Aziz, T. Z., &
Aron, A. R. (2016). Surprise disrupts cognition via a fronto-basal
ganglia suppressive mechanism. Nature Communications, 7,
11195. DOI: https://doi.org/10.1038/ncomms11195, PMID:
27088156, PMCID: PMC4837448
Winn, J., & Bishop, C. M. (2005). Variational message passing.
Journal of Machine Learning Research, 6, 661–694.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
9
1
6
3
6
1
9
5
6
1
4
0
/
/
j
o
c
n
_
a
_
0
1
6
6
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
1656
Journal of Cognitive Neuroscience
Volume 33, Number 9