A Causal Role of the Right Superior Temporal
Sulcus in Emotion Recognition
From Biological Motion
Rochelle A. Basil
1
2
, Margaret L. Westwater
1
, Martin Wiener
, and James C. Thompson
1
1George Mason University
2Department of Psychiatry, Addenbrooke’s Hospital, University of Cambridge
a n o p e n a c c e s s
j o u r n a l
Keywords: biological motion, superior temporal sulcus, emotion recognition, transcranial magnetic
stimulation
ABSTRACT
Understanding the emotions of others through nonverbal cues is critical for successful social
interactions. The right posterior superior temporal sulcus (pSTS) is one brain region thought
to be key in the recognition of the mental states of others based on body language and facial
expression. In the present study, we temporarily disrupted functional activity of the right
pSTS by using continuous, theta-burst transcranial magnetic stimulation (cTBS) to test
the hypothesis that the right pSTS plays a causal role in emotion recognition from body
movements. Participants (N = 23) received cTBS to the right pSTS, which was individually
localized using fMRI, and a vertex control site. Before and after cTBS, we tested participants’
ability to identify emotions from point-light displays (PLDs) of biological motion stimuli and a
nonbiological global motion identification task. Results revealed that accurate identification
of emotional states from biological motion was reduced following cTBS to the right pSTS,
but accuracy was not impaired following vertex stimulation. Accuracy on the global motion
task was unaffected by cTBS to either site. These results support the causal role of the right
pSTS in decoding information about others’ emotional state from their body movements
and gestures.
INTRODUCTION
The dynamics and kinematics of body movements play a vital role in the expression and per-
ception of emotion and other social cues. Indeed, the ability to detect, identify, and respond
appropriately to these dynamic nonverbal cues is central to social competence (Mehrabian &
Ferris, 1967; Rosenthal, Hall, DiMatteo, Rogers, & Archer, 1979). Neuroimaging evidence
has long implicated the right posterior superior temporal sulcus (pSTS) as a cortical region
central to the perception of social cues from body movements (Bonda, Ostry, & Evans, 1996;
Grossman & Blake, 2002; Puce, Allison, Bentin, Gore, & McCarthy, 1998). This region
shows a preference for dynamic bodies and faces over static, and it is sensitive to the configu-
ration of a moving body (Pitcher, Dilks, Saxe, Triantafyllou, & Kanwisher, 2011; Thompson,
Clarke, Stewart, & Puce, 2005). While evidence indicates that both the left pSTS (Saygin et al.,
2007; van Kemenade, Muggleton, Walsh, & Saygin, 2012) and right pSTS are involved in the
perception of body movement, there is evidence for the response to body motion to be more
consistent in the right hemisphere (Bonda et al., 1996; Engell & McCarthy, 2013; Grossman
& Blake 2002; Pavlova, Lutzenberger, Sokolov, & Birbaumer, 2004; Pelphrey, Morris, &
Citation: Basil, R. A., Westwater, M. L.,
Wiener, M., & Thompson, J. C. (2017).
A Causal Role of the Right Superior
Temporal Sulcus in Emotion
Recognition From Biological Motion.
Open Mind: Discoveries in Cognitive
Science, 2(1), 26–36. https://doi.org/
10.1162/opmi_a_00015
DOI:
https://doi.org/10.1162/opmi_a_00015
Received: 11 November 2016
Accepted: 6 September 2017
Competing Interests: The authors
declare no competing financial
interests.
Corresponding Author:
James C. Thompson
jthompsz@gmu.edu
Copyright: © 2017
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
STS Role in Emotion Recognition Basil et al.
McCarthy 2004; Pitcher, Duchaine, & Walsh, 2014; Puce et al., 1998). Greater functional
magnetic resonance imaging (fMRI) response selectivity to point-light displays (PLDs) of bio-
logical motion in the right pSTS is also associated with larger, more complex social networks,
implying that the coding of the movements of others in this region is important for social
abilities (Dziura & Thompson, 2014). However, common neuroimaging techniques, such as
cross-sectional fMRI, are correlational and therefore limited in their ability to establish the
causal role of a brain region in a perceptual or cognitive function. Here, we used fMRI-guided
continuous, theta-burst transcranial magnetic stimulation (cTBS), a specific repetitive tran-
scranial magnetic stimulation (rTMS) protocol, to examine whether the right pSTS is causally
involved in the perception of emotional states conveyed by body movements.
Understanding the causal role of the right pSTS in the perception of dynamic social stim-
uli is important, as impaired processing of face and body stimuli in this region has been linked
to social deficits in autism spectrum disorder (ASD; Alaerts et al., 2014; Koldewyn, Whitney,
& Rivera, 2011; Pavlova, 2012) and schizophrenia (Kim, Doop, Blake, & Park, 2005; Kim,
Norton, McBain, Ongur, & Chen, 2013). More recently, decreased fMRI activity to point-light
biological motion stimuli in the right pSTS has been proposed as a “neurobiomarker” for ASD
(Björnsdotter, Wang, Pelphrey, & Kaiser, 2016). Studies of brain structure also point to altered
cortical thickness of the pSTS in neurodevelopmental disorders (Zilbovicius et al., 2006), and
protracted maturation of this region has been associated with altered functional network dif-
ferentiation in children and adolescents with ASD (Shih et al., 2011). Shih et al.
(2011)
postulate that atypical development of the pSTS could indicate impaired functional specificity
of this region in individuals with ASD, and this might contribute to the inability of some with
neurodevelopmental disorders to discriminate dynamic social cues, such as subtle shifts in
body language, that convey the intentions or emotions of another. As such, increased under-
standing of the role of the right pSTS in the perception and discrimination of social information
from biological motion is important for future work with these clinical populations.
Several studies have investigated biological motion perception using offline repetitive
transcranial magnetic stimulation (rTMS) to the pSTS, where “offline” indicates that stimula-
tion does not occur simultaneously with the task. Accordingly, “online” stimulation protocols
administer stimulation concurrently with the task itself, such that stimulation occurs at a prede-
termined point in time during the task protocol, typically with either a single pulse of TMS, or
a brief, high-frequency burst of stimulation. Online stimulation protocols are preferred when
the causal role of a brain region has been well established and the precise timing of a region is
under investigation. Offline stimulation protocols are useful when the goal is to first establish
the causal involvement of a brain region in a specific cognitive function. Grossman, Battelli,
and Pascual-Leone (2005) previously reported that 1 Hz repetitive TMS (rTMS) to the right pSTS
impaired discrimination of upright point-light biological motion when presented in noise. Sim-
ilarly, Vangeneugden et al.
(2014) used 1 Hz rTMS to show that the right pSTS was involved
in discriminating walking direction from biological motion. One study (van Kemenade et al.,
2012) used cTBS to examine the role of left pSTS in biological motion discrimination from
noise, finding a nonsignificant trend for reduced detection of biological motion from noise.
Potential advantages of cTBS include similar effect sizes to online stimulation, signifi-
cantly shorter stimulation periods (<1 min), and longer duration of effect (60–90 min)
compared to 1 Hz rTMS (Huang, Edwards, Rounis, Bhatia, & Rothwell, 2005; Thut Pascual-
Leone, 2010). Given prior work, we sought determine if pSTS function extends further to
the coding identification actions conveying emotional information. We examined the
effects disruption by continuous theta-burst (cTBS) on a point-light emotion discrim-
ination task (Atkinson, Dittrich, Gemmell, Young, 2004). chose this because the
OPEN MIND: Discoveries in Cognitive Science
27
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
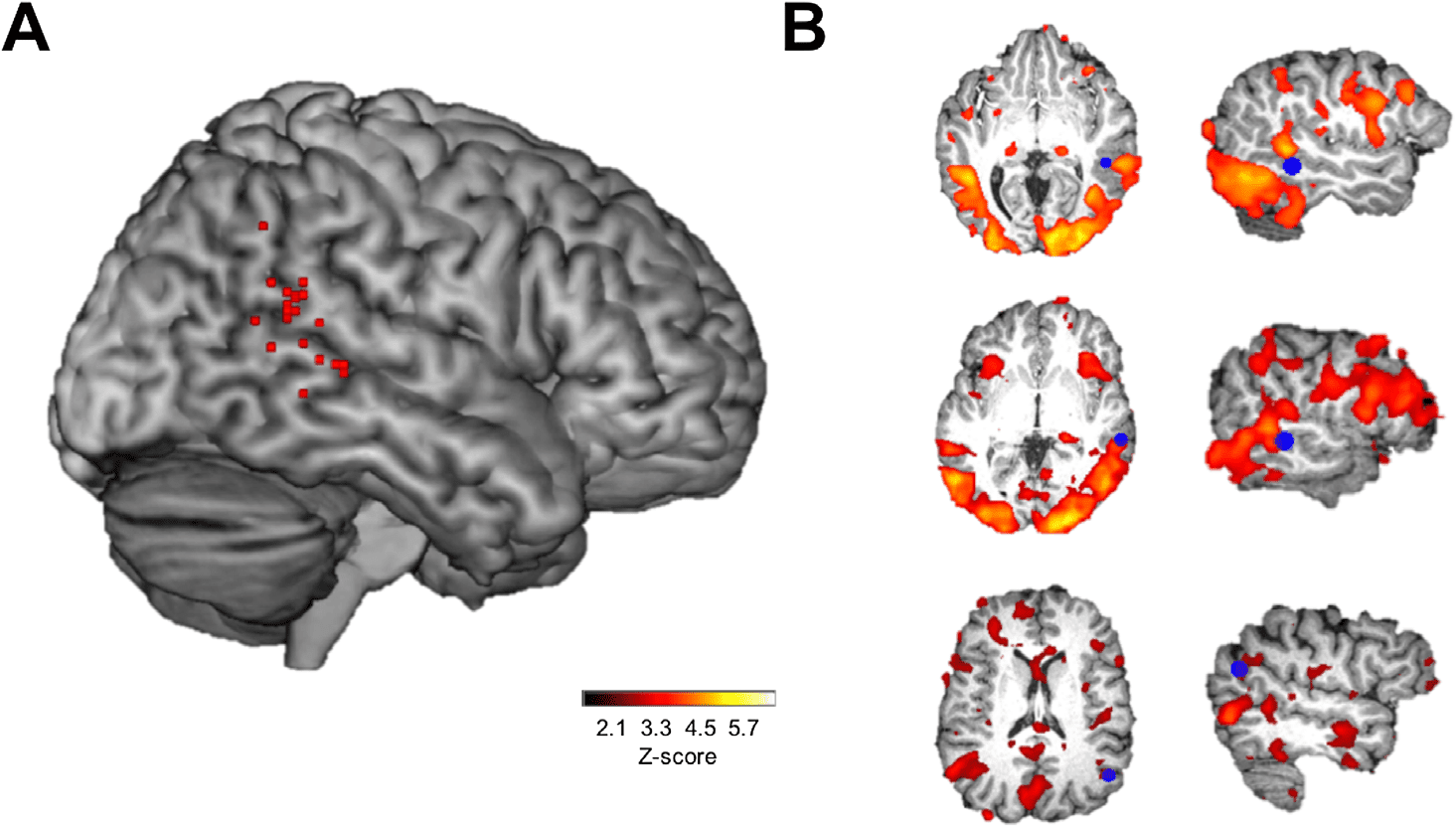
> scrambled con-
trast. (A) Subjects (N = 23) cTBS stimulation sites overlaid on the MNI-152 template brain.
(B) Uncorrected Z-statistic maps of intact > scrambled contrast for three representative subjects.
Activation maps were thresholded for visualization, and regions of interest (ROI) were overlaid.
OPEN MIND: Discoveries in Cognitive Science
29
STS Role in Emotion Recognition Basil et al.
previous studies (Borckardt et al., 2008). Participants were provided hearing protection for
both stimulation sessions.
Procedure
In a randomized and counterbalanced design, participants were presented with dynamic stim-
uli of PLDs, which comprised the biological stimuli, as well as global nonbiological motion
stimuli in different directions in separate alternating blocks (see Figure 2C). Stimuli were pre-
sented on a Dell E1911 19” Monitor with 1440 × 900 resolution and a refresh rate of 60 Hz,
using Neurobehavioral Systems Presentation Software (Version 17.2; 2015). Each stimulus
was presented for 1,200 ms, and each block contained 48 trials (16 trials per condition). Each
block was 3 min in duration.
On each trial, participants completed a three-alternative forced choice task, where they
were instructed to choose the emotion (in the case of biological motion) or the type of motion
(in the nonbiological motion trials) as quickly and accurately as possible. For the biological
motion task, PLDs performed actions to convey one of the three emotions (Figure 2A). There
were 10 actions for each of the three emotions, and stimuli came from a set of full-body 13-dot
PLDs created by and validated by Atkinson and colleagues (2004). The stimulus set included,
among others, shaking of the fist or arms (angry), a vertical jump or skipping (happy), and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
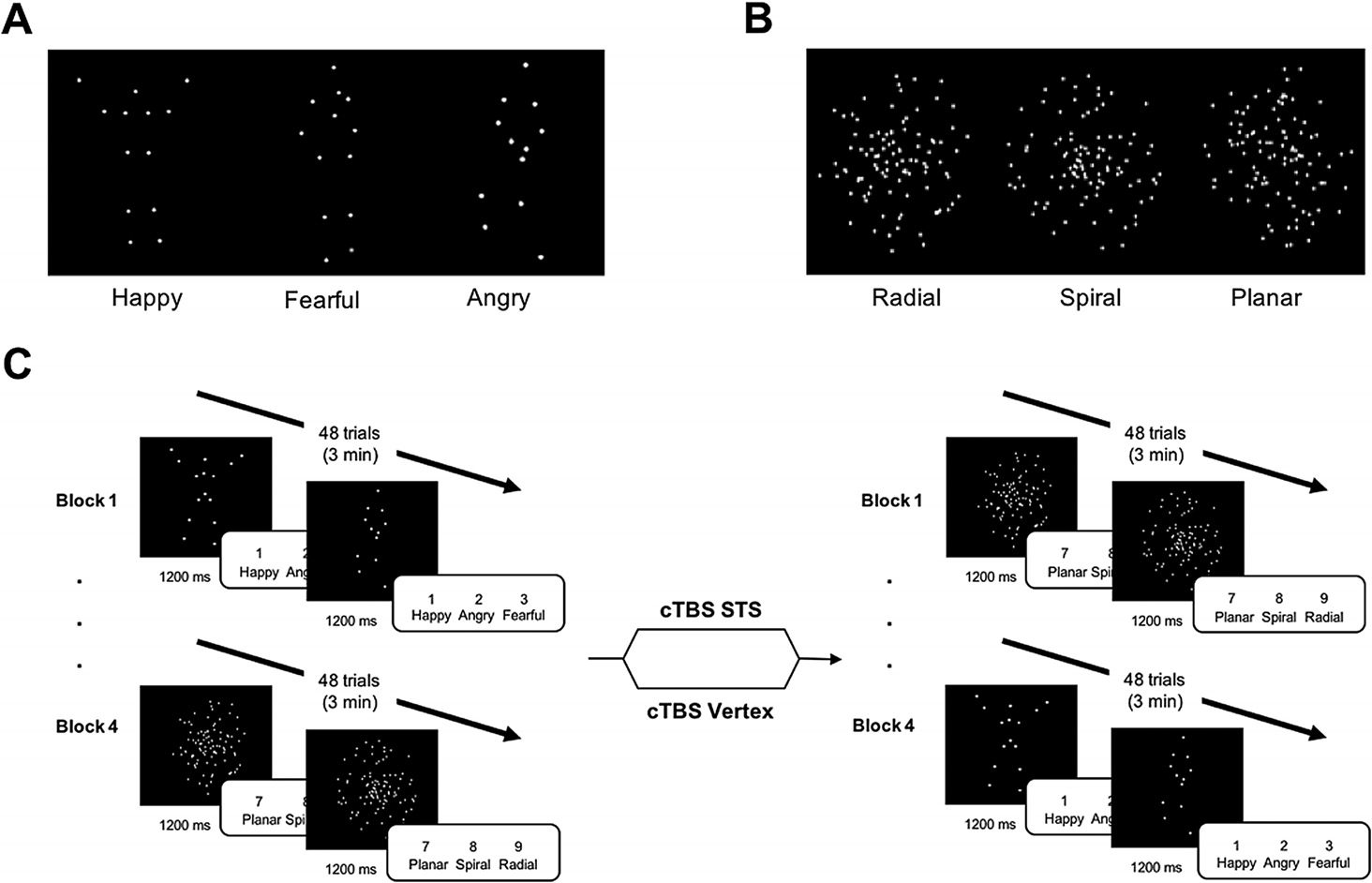
Figure 2. Biological and nonbiological motion stimuli and task paradigm. (A) Biological stimuli
were PLDs (point-light displays) that conveyed three emotions: happy, fearful, and angry. (B) Non-
biological motion stimuli consisted of dots moving with the same pattern in three conditions: radial,
spiral, and planar. Radial motion was defined as dots moving with the same rho but separate theta
values. Dots for spiral motion had the same theta but separate rho values. Dots had the same rho
and theta for planar motion. Each condition had 10 actions. (C) Participants completed four ran-
domized task blocks (two biological motion; two nonbiological motion) prior to cTBS (continuous,
theta-burst transcranial magnetic stimulation) to either the right pSTS (posterior superior temporal
sulcus) or vertex. Four more randomized task blocks (two biological motion; two nonbiological mo-
tion) were completed after cTBS. Each block was 3 min. Each participant completed the paradigm
twice for cTBS of the pSTS and vertex. Stimulation site was counterbalanced across participants.
OPEN MIND: Discoveries in Cognitive Science
30
STS Role in Emotion Recognition Basil et al.
◦
◦
◦
/sec, with a lifetime of 7 frames, within a circular aperture of 12
cowering or turning from camera (fearful). The global motion task required subjects to view a
series of dots and choose whether the motion of the dots appeared to be unidirectional (i.e.,
planar), radiating from a center point (radial), or circulating around a center point (spiral; see
Figure 2B). Stimuli for the global motion control task were created in MATLAB (v.R2012a).
Stimuli consisted of 100 white dots (0.2
) on a black background. Dots moved at a speed
of 2
. Noise dots (60%)
moved in a random direction for the duration of their lifetime. Coherent dots (radial, rotational,
or planar) moved with the same pattern, with the pattern first determined in polar coordi-
nates and converted to Cartesian coordinates for drawing each frame. Coherent radial motion
(expansion/contraction) was determined by making all dots move with the same rho
(Spearman’s rank-ordered correlation coefficient) but with each dot having a separate theta.
Coherent spiral motion (clockwise/counterclockwise) was created by making all dots move
with the same theta and separate rho. Planar motion (left or right) was created by making dots
move with the same theta and rho. As in the biological motion task, there were ten actions per
condition. Participants were seated 75 cm in front of a desktop screen when completing the
biological and nonbiological motion tasks. Each participant remained seated for cTBS stimu-
◦
lation. The visual angle of the stimuli was 9.4
, and it remained the same for all conditions.
Subjects were asked to respond using designated response keys on a keyboard. Numeric keys
1, 2, and 3 corresponded with biological motion stimuli, and keys 6, 7, and 8 were used for
nonbiological motion trials.
Before the experiment began, participants completed a training session to familiarize
In total, subjects completed four
themselves with both the stimuli and the button coding.
blocks of biological motion (two pre-cTBS, two post-cTBS) and four blocks of nonbiological
motion (two pre-cTBS, two post-cTBS) in a counterbalanced fashion. Continuous TBS was
performed upon completion of the first four blocks (see Figure 2C). Subjects remained seated
while the experimenter and an assistant located the stimulation site (i.e., vertex, pSTS), posi-
tioned the coil, and applied stimulation. Following cTBS, subjects were repositioned, facing
the computer screen, and instructed to complete the remaining task blocks. Each session lasted
approximately 40 min in total.
RESULTS
The cTBS manipulation aimed to temporarily disrupt participants’ ability to identify emotional
states from biological motion stimuli following cTBS over the right pSTS. As an active con-
trol, the vertex was stimulated. We did not anticipate a decrease in subjects’ ability to label
emotional states after cTBS to the vertex. A paired samples t test indicated nonsignificant differ-
ences between accurate identification of biological (M = 0.86, SD = 0.07) and nonbiological
(M = 0.88, SD = 0.16) stimuli before cTBS, t(22) = 0.928, p = .329. As such, any observed
differences in accuracy across the task conditions were hypothesized to reflect cTBS stimula-
tion, not task difficulty. Statistical analyses of task session data were completed in IBM SPSS
Statistics (v.19). We conducted two, two-by-two repeated measures ANOVAs to examine the
effect of cTBS site (right pSTS and vertex) and stimulus type (biological or nonbiological) on
accuracy and reaction time (RT). Mean accuracy and RT values for each condition are shown
in Table 1.
The two-by-two repeated measures ANOVA of accuracy showed a significant main
effect of cTBS site, F(1, 22) = 10.87, p = .003. A main effect of stimulus type also reached
statistical significance, F(1, 22) = 8.50, p = .008. Analyses further indicated a significant
OPEN MIND: Discoveries in Cognitive Science
31
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
STS Role in Emotion Recognition Basil et al.
Table 1. Accuracy and reaction time values for biological and nonbiological motion conditions
pre- and post-cTBS.
Accuracy
Biological
Nonbiological
Reaction time
Biological
Nonbiological
pSTS
Vertex
Pre-cTBS
88 ± 6%
90 ± 10%
Post-cTBS
84 ± 9%
93 ± 9%
Pre-cTBS
84 ± 7%
86 ± 20%
Post-cTBS
87 ± 8%
90 ± 15%
1,262 ± 280 ms
1,146 ± 286 ms
1,137 ± 226 ms
980 ± 222 ms
1,308 ± 259 ms
1,158 ± 297 ms
1,203 ± 202 ms
1,014 ± 184 ms
Notes: Accuracy (% correct) and reaction time (ms) values are presented as M ± SD. cTBS = con-
tinuous, theta-burst transcranial magnetic stimulation; pSTS = posterior superior temporal sulcus.
two-way interaction of cTBS site and stimulus type, F(1, 22) = 4.93, p = .037. These effects are
shown in Figure 3. Post-hoc paired samples t tests revealed that the change in accuracy when
cTBS was targeted over the right pSTS significantly differed between the two stimulus types,
t(22) = −4.60, p < .001. Post-hoc comparisons further demonstrated that participants’ ability
to accurately identify emotions from biological motion was significantly impaired following
cTBS to the right pSTS but not to the vertex, t(22) = −5.77, p < .001. Additional post-hoc
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Mean differences in percentage accuracy change by stimulus condition (biological
motion and nonbiological motion) cTBS site. Continuous transcranial magnetic stimulation (TBS)
over right posterior superior temporal sulcus (pSTS) impaired emotional recognition through biolog-
ical motion, but not nonbiological motion. The y axis denotes mean change in accuracy (% correct)
from pre- to post-cTBS (continuous, theta-burst transcranial magnetic stimulation), where a value
of 0 would indicate no change. An asterisk denotes a significant (p < .001) difference. Error bars
indicate SEM.
OPEN MIND: Discoveries in Cognitive Science
32
STS Role in Emotion Recognition Basil et al.
Figure 4. Mean differences in change in reaction time by condition (biological and non-
biological motion) by cTBS site. Reaction time (RT; ms) to either stimulus type was not significantly
changed following continuous, theta-burst transcranial magnetic stimulation (cTBS) to the right
posterior superior temporal sulcus (pSTS) and vertex. The y axis denotes a change in RT from pre- to
post-cTBS, where more negative values indicate greater decreases in RT (i.e., faster responding) from
pre- to post-cTBS. Error bars indicate SEM.
comparisons indicated that accurate discrimination of nonbiological motion stimuli did not
differ significantly by cTBS site, t(22) = −0.45, p = .66.
A two-by-two repeated measures ANOVA of RT did not demonstrate a main effect of
cTBS site, F(1, 22) = 0.37, p = .547, or stimulus type, F(1, 22) = 2.89, p = .103. The two-way
interaction of cTBS site and stimulus type was also nonsignificant, F(1, 22) = 0.00, p = .984.
These findings are illustrated in Figure 4.
DISCUSSION
The present study found that disruption of the right pSTS using cTBS leads to selective impair-
ment in the recognition of emotions conveyed by human movements. We used fMRI to target
a right pSTS region in each participant, as this region has been previously implicated in the vi-
sual processing of biological motion and in “social networks” of the brain. We found that right
pSTS-targeted cTBS reduced the accuracy of the identification of different emotional point-
light stimuli. The detrimental effects of cTBS to this region did not extend to the recognition of
nonbiological motion, as we found no significant impairment in the identification of different
global motion stimuli. The effect of cTBS to the right pSTS was limited to recognition accuracy;
RT was not significantly changed by cTBS. This absence of an RT effect is not unprecedented,
as similar findings were reported by both Grossman et al.
(2005) and Vangeneugden et al.
(2014). The findings of this study indicate that the right pSTS subserves the coding of dynamic
social information, such as emotion, conveyed by the body movements of another person.
OPEN MIND: Discoveries in Cognitive Science
33
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
STS Role in Emotion Recognition Basil et al.
Previous work has shown that 1 Hz rTMS to the pSTS region can impair discrimination
of biological motion from noise (Grossman et al., 2005). Additional work has indicated that
rTMS to the pSTS can decrease walking direction discrimination from biological motion while
leaving facing orientation unaffected (Vangeneugden et al., 2014). Taken together, these stud-
ies underscore the importance of a subregion of the pSTS to the detailed visual processing of
biological motion; however, it remains unclear whether the causal role of this pSTS region ex-
tends to the coding of social information, such as emotion. It is possible that the pSTS serves as
a high-level visual processor that is not central to the processing of social meaning from such
stimuli. The selectivity of the fMRI response of the pSTS region to the biological motion region
does appear to be linked to social abilities (Pelphrey, Morris, & McCarthy, 2004; Saxe, Xiao,
Kovacs, Perrett, & Kanwisher, 2004), and even the size and complexity of social networks
(Dziura & Thompson, 2014), suggesting a more diffuse role in social cognition. Assessment
of right STS grey matter volume further supports the region as a predictor of social cognition,
where increased volume has been associated with biological motion discrimination (Gilaie-
Dotan, Kanai, Bahrami, Rees, & Saygin, 2013) and social network size (Kanai, Bahrami,
Roylance, & Rees, 2012). More recently, an fMRI study showed that the pSTS region is in-
volved in the processing of emotion conveyed by body movements (Goldberg, Christensen,
Flash, Giese, & Malach, 2015). Together with the findings of the present study, which demon-
strate a causal role in emotion recognition from biological motion, it seems that the visual
coding of human movements represents one facet of a broader social cognitive role played by
the pSTS region (Allison, Puce, & McCarthy, 2000).
Consistent with a broader social cognitive role of the right pSTS region, this region
has also been considered by many as part of a face-processing network (Haxby, Hoffman,
& Gobbini, 2000, 2002; Hoffman & Haxby, 2000). The original model by Haxby and
colleagues (2000) proposed that the right pSTS represents changeable aspects of the face, such
as emotional expression, eye-gaze, and mouth movements, while ventral temporal fusiform
cortex represents invariant aspects of faces, such as identity. This model was modified by
O’Toole and colleagues (2002), who suggested that the right pSTS might also encode facial
identity based on dynamic motion signatures. The pSTS face area (pSTS-FA) overlaps consid-
erably with the pSTS region that represents biological motion (Deen, Koldewyn, Kanwisher, &
Saxe, 2015; Engell & McCarthy, 2013; Grosbras, Beaton, & Eickhoff, 2012), and a causal
role for the pSTS-FA in recognizing facial emotional expressions has been demonstrated using
rTMS by Pitcher and colleagues (2014). Recent work by Deen et al. (2015) reported functional
overlap between STS modules that respond selectively to biological motion and face stimuli;
however, reliable differences in the spatial distribution of these modules were also observed.
In future work, it will be important to determine the degree of overlap between pSTS modules
that exhibit response selectivity for biological movement, emotional expression derived from
said motion, and facial expression.
Dysfunction of the processing of biological motion in the right pSTS region has been
suggested to be a potential diagnostic “biomarker” for social communication deficits in ASD
(Björnsdotter et al., 2016), and it has been implicated in social deficits in schizophrenia (Kim
et al., 2013). Our finding of a causal role of this region in emotion recognition from biological
motion in healthy adults strengthens the rationale for examining right pSTS functionality when
determining the basis of social and emotional deficits in clinical populations.
It should be
noted that, while we did not examine the potential role of the left pSTS in emotion recognition
from kinetic movement, functional connectivity analyses suggest that this region coactivates
with the right pSTS in response to social stimuli (Lahnakoski et al., 2012). Further examination
of the causal role of the left pSTS in emotion perception is warranted. In pursuing this work,
OPEN MIND: Discoveries in Cognitive Science
34
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
STS Role in Emotion Recognition Basil et al.
findings will provide a neurobiological basis to guide future treatment interventions for social
and affective processing.
ACKNOWLEDGMENTS
This work was supported by Office of Naval Research Award N00014-10-1-0198.
AUTHOR CONTRIBUTIONS
RAB, MW, and JCT designed the study. RAB and MLW carried out the experimental procedures
and completed data analysis. RAB and MLW drafted the manuscript. JCT and MW provided
critical feedback on the manuscript, and all authors contributed to the final draft of the
manuscript.
REFERENCES
Alaerts, K., Woolley, D. G., Steyaert, J., Di Martino, A., Swinnen,
S. P., & Wenderoth, N. (2014). Underconnectivity of the superior
temporal sulcus predicts emotion recognition deficits in autism.
Social Cognitive and Affective Neuroscience, 9, 1589–1600. doi:
10.1093/scan/nst156
Allison, T., Puce, A., & McCarthy, G. (2000). Social perception from
visual cues: role of the STS region. Trends in Cognitive Sciences,
4, 267–278. doi:10.1016/S1364-6613(00)01501-1
Atkinson, A. P., Dittrich, W. H., Gemmell, A. J., & Young, A. W.
(2004). Emotion perception from dynamic and static body ex-
pressions in point-light and full-light displays. Perception, 33,
717–746. doi:10.1068/p5096
Björnsdotter, M., Wang, N., Pelphrey, K., & Kaiser, M. D.
(2016).
Evaluation of quantified social perception circuit activity as
JAMA
a neurobiological marker of autism spectrum disorder.
Psychiatry, 73, 614–621.
doi:10.1001/jamapsychiatry.2016.
0219
Borckardt,
Bonda, E., Ostry, D., & Evans, A.
Journal of Neuroscience, 16, 3737–3744.
(1996). Specific involvement of
human parietal systems in the perception of biological motion
and the amygdala.
J.
J., Reeves, S. T., Weinstein, M., Smith, A. R.,
Shelley, N., Kozel, F. A., . . . George, M. S. (2008). Significant
analgesic effects of one session of postoperative left prefrontal
cortex repetitive transcranial magnetic stimulation: A replication
study. Brain Stimulation, 1, 122–127. doi:10.1016/j.brs.2008.
04.002
Brainsight Image Analysis and Neuronavigation Software (Version
2.3). (2016). Montreal, Canada: Rogue Research. Available from
www.rogue-research.com
Deen, B., Koldewyn, K., Kanwisher, N., & Saxe, R.
(2015). Func-
tional organization of social perception and cognition in the
superior temporal sulcus. Cerebral Cortex, 25, 4596–4609. doi:
10.1093/cercor/bhv111
Dziura, S. L., & Thompson, J. C. (2014). Social-network complexity
in humans is associated with the neural response to social infor-
mation. Psychological Science, 25, 2095–2101. doi:10.1177/
0956797614549209
Engell, A. D., & McCarthy, G.
(2013). Probabilistic atlases for
face and biological motion perception: An analysis of their re-
liability and overlap. NeuroImage, 74, 140–151. doi:10.1016/j.
neuroimage.2013.02.025
Gilaie-Dotan, S., Kanai, R., Bahrami, B., Rees, G., & Saygin,
A. P.
(2013). Neuroanatomical correlates of biological mo-
tion detection. Neuropsychologia, 51, 457–463. doi:10.1016/j.
neuropsychologia.2012.11.027
Goldberg, H., Christensen, A., Flash, T., Giese, M. A., & Malach,
R.
(2015). Brain activity correlates with emotional perception
induced by dynamic avatars. NeuroImage, 122, 306–317. doi:
10.1016/j.neuroimage.2015.07.056
Grosbras, M. H., Beaton, S., & Eickhoff, S. B. (2012). Brain regions
involved in human movement perception: A quantitative voxel-
based meta-analysis. Human Brain Mapping, 33, 431–454. doi:
10.1002/hbm.21222
Grossman, E. D., Battelli, L., & Pascual-Leone, A.
(2005). Repet-
itive TMS over posterior STS disrupts perception of biological
motion. Vision Research, 45, 2847–2853. doi:10.1016/j.visres.
2005.05.027
Grossman, E. D., & Blake, R. (2002). Brain areas active during visual
perception of biological motion. Neuron, 35, 1167–1175. doi:
10.1016/S0896-6273(02)00897-8
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The distributed
human neural system for face perception. Trends in Cognitive
Sciences, 4, 223–233. doi:10.1016/S1364-6613(00)01482-0
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I.
(2002). Human
neural systems for face recognition and social communica-
tion. Biological Psychiatry, 51, 59–67. doi:10.1016/S0006-
3223(01)01330-0
Hoffman, E. A., & Haxby, J. V.
(2000). Distinct representations of
eye gaze and identity in the distributed human neural system for
face perception. Nature Neuroscience, 3, 80–84. doi:10.1038/
71152
OPEN MIND: Discoveries in Cognitive Science
35
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
STS Role in Emotion Recognition Basil et al.
(2005).
Huang, Y., Edwards, M., Rounis, E., Bhatia, K., & Rothwell,
J. C.
Theta burst stimulation of the human motor
cortex. Neuron, 45, 201–206. Retrieved from http://papers2://
publication/uuid/2B96EA86-FE20-4A0C-9DD6-58267BF0F8F9
(2012). Online
social network size is reflected in human brain structure. Pro-
ceedings of the Royal Society B—Biological Sciences, 279, 1327–
1334. doi:10.1098/rspb.2011.1959
Kanai, R., Bahrami, B., Roylance, R., & Rees, G.
Kim, J., Doop, M. L., Blake, R., & Park, S.
Impaired
visual
recognition of biological motion in schizophrenia.
Schizophrenia Research, 77, 299–307. doi:10.1016/j.schres.
2005.04.006
(2005).
Kim, J., Norton, D., McBain, R., Ongur, D., & Chen, Y.
(2013).
Deficient biological motion perception in schizophrenia: Re-
sults from a motion noise paradigm. Frontiers in Psychology, 4,
153–164. doi:10.3389/fpsyg.2013.00391
Koldewyn, K., Whitney, D., & Rivera, S. M.
(2011). Neural cor-
relates of coherent and biological motion perception in autism.
Developmental Science, 14, 1075–1088. doi:10.1111/j.1467-
7687.2011.01058.x
Lahnakoski, J. M., Glerean, E., Salmi, J., Jääskeläinen, I. P., Sams, M.,
Hari, R., & Nummenmaa, L. (2012). Naturalistic fmri mapping
reveals superior temporal sulcus as the hub for the distributed
brain network for social perception. Frontiers in Human Neuro-
science, 6, 233. doi:10.3389/fnhum.2012.00233
MATLAB (Version R2012a). (2012). Natick, MA: Mathworks. Avail-
able from www.mathworks.com
Mehrabian, A., & Ferris, S. R.
Inference of attitudes from
(1967).
nonverbal communication in two channels. Journal of Consulting
Psychology, 31, 248–252. doi:10.1037/h0024648
Neurobehavioral Systems Presentation Software (Version 17.2).
(2015). Berkeley, CA: Neurobehavioral Systems, Inc. Available
from www.neurobs.com
O’Toole, A. J., Roark, D. A., & Abdi, H. (2002). Recognizing moving
faces: A psychological and neural synthesis. Trends in Cognitive
Sciences, 6, 261–266. doi:10.1016/S1364-6613(02)01908-3
Pavlova, M. A. (2012). Biological motion processing as a hallmark
of social cognition. Cerebral Cortex, 22, 981–995. doi:10.1093/
cercor/bhr156
Pavlova, M., Lutzenberger, W., Sokolov, A., & Birbaumer, N. (2004).
recognizable and non-
Dissociable cortical processing of
recognizable biological movement: Analysing gamma MEG
activity. Cerebral Cortex, 14, 181–188. doi:10.1093/cercor/
bhg117
Pelphrey, K. A., Morris, J. P., & McCarthy, G.
(2004). Grasping
the intentions of others: The perceived intentionality of an action
influences activity in the superior temporal sulcus during social
perception. Journal of Cognitive Neuroscience, 16, 1706–1716.
doi:10.1162/0898929042947900
Pitcher, D., Dilks, D. D., Saxe, R. R., Triantafyllou, C., & Kanwisher,
N.
(2011). Differential selectivity for dynamic versus static in-
formation in face-selective cortical regions. NeuroImage, 56,
2356–2363. doi:10.1016/j.neuroimage.2011.03.067
Pitcher, D., Duchaine, B., & Walsh, V. (2014). Combined TMS and
fMRI reveal dissociable cortical pathways for dynamic and static
face perception. Current Biology, 24, 2066–2070. doi:10.1016/
j.cub.2014.07.060
Pitcher, D., Garrido, L., Walsh, V., & Duchaine, B. C.
(2008).
Transcranial magnetic stimulation disrupts the perception and
embodiment of facial expressions. Journal of Neuroscience, 28,
8929–8933. doi:10.1523/JNEUROSCI.1450-08.2008
Puce, A., Allison, T., Bentin, S., Gore, J. C., & McCarthy, G. (1998).
Temporal cortex activation in humans viewing eye and mouth
movements. Journal of Neuroscience, 18, 2188–2199. Retrieved
from http://www.ncbi.nlm.nih.gov/pubmed/9482803
Rosenthal, R., Hall, J. A., DiMatteo, M. R., Rogers, P. L., & Archer, D.
(1979). Sensitivity to nonverbal cues: The PONS test. Baltimore,
MD: Johns Hopkins University Press.
Saxe, R., Xiao, D. K., Kovacs, G., Perrett, D., & Kanwisher, N. (2004).
A region of right posterior superior temporal sulcus responds
to observed intentional actions. Neuropsychologia, 42, 1435–
1446. doi:10.1016/j.neuropsychologia.2004.04.015
Saygin, A. P., Adolphs, R., Damasio, H., Tranel, D., Cooper, G.,
Damasio, A., . . . Wechsler, D. (2007). Superior temporal and
premotor brain areas necessary for biological motion perception.
Brain: A Journal of Neurology, 130(Pt. 9), 2452–2461. doi:10.
1093/brain/awm162
Shih, P., Keehn, B., Oram, J. K., Leyden, K. M., Keown, C. L., &
Müller, R.-A.
Functional differentiation of posterior
superior temporal sulcus in autism: A functional connectivity
magnetic resonance imaging study. Biological Psychiatry, 70,
270–277. doi:10.1016/j.biopsych.2011.03.040
(2011).
Thompson, J. C., Clarke, M., Stewart, T., & Puce, A. (2005). Config-
ural processing of biological motion in human superior temporal
sulcus. Journal of Neuroscience, 25, 9059–9066. doi:10.1523/
JNEUROSCI.2129-05.2005
Thut, G., & Pascual-Leone, A.
(2010). A review of combined
TMS-EEG studies to characterize lasting effects of repetitive TMS
and assess their usefulness in cognitive and clinical neuro-
science. Brain Topography, 22, 219–232. doi:10.1007/s10548-
009-0115-4
Vangeneugden, J., Peelen, M. V., Tadin, D., & Battelli, L.
(2014).
Distinct neural mechanisms for body form and body motion
Journal of Neuroscience, 34, 574–585. doi:
discriminations.
10.1523/JNEUROSCI.4032-13.2014
van Kemenade, B. M., Muggleton, N., Walsh, V., & Saygin, A. P.
(2012). Effects of TMS over premotor and superior temporal
Journal of Cognitive
cortices on biological motion perception.
Neuroscience, 24, 896–904. doi:10.1162/jocn_a_00194
Zilbovicius, M., Meresse, I., Chabane, N., Brunelle, F., Samson, Y.,
& Boddaert, N. (2006). Autism, the superior temporal sulcus and
social perception. Trends in Neurosciences, 29, 359–366. doi:
10.1016/j.tins.2006.06.004
OPEN MIND: Discoveries in Cognitive Science
36
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
2
6
1
8
6
8
3
1
5
o
p
m
_
a
_
0
0
0
1
5
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3