Evidencia electrofisiológica para la
Suppression of Highly Salient Distractors
Brad T. Stilwell1
, Howard Egeth2, and Nicholas Gaspelin1
Abstracto
■ There has been a longstanding debate as to whether salient
stimuli have the power to involuntarily capture attention. Como un
potential resolution to this debate, the signal suppression hypoth-
esis proposes that salient items generate a bottom–up signal that
automatically attracts attention, but that salient items can be
suppressed by top–down mechanisms to prevent attentional cap-
tura. Despite much support, the signal suppression hypothesis
has been challenged on the grounds that many prior studies
may have used color singletons with relatively low salience that
are too weak to capture attention. The current study addressed
this by using previous methods to study suppression but increased
the set size to improve the relative salience of the color singletons.
To assess whether salient distractors captured attention, electro-
physiological markers of attentional allocation (the N2pc compo-
próximo) and suppression (the PD component) were measured. El
results provided no evidence of attentional capture, but instead
indicated suppression of the highly salient singleton distractors,
as indexed by the PD component. This suppression occurred even
though a computational model of saliency confirmed that the
color singleton was highly salient. Altogether, this supports the
signal suppression hypothesis and is inconsistent with stimulus-
driven models of attentional capture. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
oh
C
norte
_
a
_
0
1
8
2
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
A vigorously debated question about visual attention has
been whether physically salient stimuli can automatically
capture attention (Luck, Gaspelin, Folk, Remington, &
Theeuwes, 2021). Por un lado, from a phenomeno-
logical standpoint, salient stimuli often feel as if they have
an inherent power to attract attention. Por esta razón,
salient stimuli such as brightly colored objects are often
used as visual warning signals in applied settings, como
a red stop sign or a neon orange construction cone. Sobre el
other hand, salient stimuli must be ignored to accomplish
everyday visually guided behavior. If attention were cap-
tured by every salient stimulus, simple tasks like operating
a motor vehicle or shopping for items in the grocery store
would be nearly impossible. This perplexing puzzle has
led to longstanding debate about the nature of attentional
control in vision.
As a potential resolution to the attentional capture
debate, the signal suppression hypothesis has proposed
that salient stimuli automatically compete for attention,
but can be proactively suppressed before they capture
atención (Gaspelin & Luck, 2018b, 2019). Studies sup-
porting this account have been recently challenged on
the grounds that the salient stimuli may have not been
highly salient and therefore were easy to suppress ( Wang
& Theeuwes, 2020; see also the work of Theeuwes, 2004).
The current study will therefore evaluate whether highly
1State University of New York at Binghamton, 2Johns Hopkins
Universidad
© 2022 Instituto de Tecnología de Massachusetts
salient stimuli elicit electrophysiological indices of atten-
tional capture or suppression.
The Attentional Capture Debate
Initial studies of attentional capture supported stimulus-
driven accounts, which suggest that certain classes of
physically salient stimuli have an automatic power to
capture visual attention, even when completely task irrel-
evant (Theeuwes, 1991, 1992; Yantis & En Jonia, 1984). Para
ejemplo, Theeuwes (1992) used an additional singleton
paradigma, in which participants searched for a unique
shape target (p.ej., a diamond) among a homogenous set
of distractor shapes (p.ej., circles) and reported the orien-
tation of a tilted line inside the target. En tono rimbombante, en un
subset of trials, one of the nontarget shapes was rendered
in a unique color. This color singleton was never the target
stimulus and therefore should have been ignored. Cómo-
alguna vez, manual response times (RTs) were slower when
the color singleton was present than when it was absent.
This singleton-presence cost was interpreted to indicate
that the color singleton automatically captured attention,
slowing detection of the target when it was present
(Theeuwes, 1991, 1992, 2004).
Subsequent studies, sin embargo, provided opposing evi-
dence that salient stimuli do not have the ability to auto-
matically capture attention (Bacon & Egeth, 1994; Folk,
Remington, & Johnston, 1992). According to goal-driven
accounts, physically salient stimuli will only capture atten-
tion when they match the attentional control settings of
Revista de neurociencia cognitiva 34:5, páginas. 787–805
https://doi.org/10.1162/jocn_a_01827
the observer. Por ejemplo, when participants establish an
attentional set for the color red, any red item—including
salient red color singletons—could capture attention;
mientras, any colored item falling outside of the attentional
colocar, such as a salient blue color singleton, would not (Folk
& Remington, 2010; Folk et al., 1992). To explain the afore-
mentioned studies using the additional singleton para-
digm (Theeuwes, 1992), goal-driven accounts proposed
that participants may have inadvertently been encouraged
to establish an attentional set for salience more generally.
Eso es, participants searched for the uniquely shaped
objetivo (a shape singleton), and this may have caused
observers to search for any unique stimulus. This singleton
detection mode (Pashler, 1988) may have caused the color
singleton to capture attention. Later studies modified the
additional singleton paradigm to discourage singleton
detection mode by making the nontarget shapes hetero-
geneous, thereby encouraging a more stringent atten-
tional set for the specific target feature (Bacon & Egeth,
1994). This feature-search mode completely eliminated
the singleton-presence cost, suggesting that the singleton
distractor no longer captured attention. This study and
others led to the conclusion that physically salient distrac-
tors do not automatically capture attention unless made
task relevant (Moher, Abrams, Egeth, Yantis, & Stuphorn,
2011; Egeth, leonardo, & Leber, 2010; Lien, Ruthruff, &
Johnston, 2010; Lien, Ruthruff, Goodin, & Remington,
2008; Leber & Egeth, 2006; lamy & Egeth, 2003; Folk,
Leber, & Egeth, 2002).
Por décadas, the debate between stimulus-driven and
goal-driven accounts of the attentional capture remained
largely unresolved. This was problematic because
stimulus-driven accounts and goal-driven accounts make
directly competing predictions about whether salient
stimuli can automatically attract attention.
The Signal Suppression Hypothesis
As a potential resolution, a hybrid model of attentional
capture was proposed: the signal suppression hypothesis
(Luck et al., 2021; Gaspelin & Luck, 2018b, 2019). Accord-
ing to this account, physically salient stimuli automatically
generate a bottom–up signal that will capture attention.
Sin embargo, salient stimuli can be suppressed via top–down
mechanisms to prevent attentional capture. De este modo, if a
physically salient stimulus is not suppressed, it will auto-
matically capture attention, which is consistent with
stimulus-driven accounts. Además, if the physically
salient stimulus is successfully suppressed, then there will
be no resulting attentional capture, which is consistent
with goal-driven accounts.
Support for this account has accumulated through con-
verging evidence from studies of psychophysics (Adán,
patel, Rangan, & serencias, 2021; Chang & Egeth, 2019,
2021; Won & Geng, 2020; Gaspelin, leonardo, & Luck,
2015), eye movements (Gaspelin, Gaspar, & Luck, 2019;
Gaspelin & Luck, 2018a; Gaspelin, leonardo, & Luck,
2017), ERPs (van Moorselaar, Daneshtalab, & Slagter,
2021; Feldmann-Wüstefeld, Busch, & Schubö, 2020;
Gaspelin & Luck, 2018C; Weaver, van Zoest, & Hickey,
2017; Gaspar, Christie, Prime, Jolicœur, & McDonald,
2016; Gaspar & McDonald, 2014; Sawaki & Luck, 2010),
and single-unit recordings in monkeys (Cosman, Lowe,
Woodman, & Schall, 2018). Además, an emerging
body of literature suggests that observers can learn to
suppress salient items based on their locations (camioneta
Moorselaar & Slagter, 2019; Ruthruff & Gaspelin, 2018;
Wang & Theeuwes, 2018a, 2018b), simple features (Adán
& serencias, 2021; Gaspelin et al., 2019; Stilwell, Bahle, &
Vecera, 2019; Gaspelin & Luck, 2018a; Cunningham &
Egeth, 2016; Vatterott & Vecera, 2012) and possibly by
learning to anticipate their mere presence more generally
(van Moorselaar, Lampers, Cordesius, & Slagter, 2020;
Won, Forloines, zhou, & Geng, 2020; Won, Kosoyan, &
Geng, 2019; Vatterott, Mozer, & Vecera, 2018; Moher
et al., 2011).
One line of support for the signal suppression hypoth-
esis has come from the capture-probe paradigm (p.ej.,
Stilwell & Gaspelin, 2021; Gaspelin et al., 2015). This task
involves intermixing frequent search trials with infrequent
probe trials. On search trials, participants search for a tar-
get stimulus and attempt to ignore a salient item such as a
color singleton. On probe trials, probe letters are briefly
superimposed on top of the search items and participants
report as many letters as possible in an untimed task. El
assumption is that participants should be more likely to
report letters at attended locations and less likely to report
letters at suppressed locations. The typical finding is that
probe report accuracy is lower for letters appearing at the
singleton location than for letters appearing at the average
nonsingleton location: a probe suppression effect (Stilwell
& Gaspelin, 2021; Gaspelin et al., 2015). This suggests that
singletons are suppressed below baseline levels to prevent
attentional capture.
Further evidence for the signal suppression hypothesis
has come from ERP studies of the N2 posterior contralat-
eral (N2pc) and distractor positivity (PD) componentes.
The N2pc component is an index of covert attentional
allocation (Eimer, 2014; Luck, 2012; Hickey, Di Lollo, &
McDonald, 2009; Eimer & Kiss, 2008; Woodman & Luck,
2003; Hillyard, Vogel, & Luck, 1998; Luck & Hillyard,
1994). It occurs as a negative-going deflection at elec-
trode sites that are contralateral to the attended visual
hemifield around 200–300 msec following stimulus pre-
sentation. En cambio, the PD component is a putative
index of attentional suppression that appears as a
positive-going deflection contralateral to the ignored
hemifield (Hickey, McDonald, & Theeuwes, 2006). El
scalp distribution of the PD component is similar to the
N2pc component (Sawaki & Luck, 2010; Hickey et al.,
2009). Además, magnetoencephalography and fMRI
studies in humans seem to indicate that source activity
underlying the N2pc and PD components propagates
from higher levels of visual cortex such as IT and V4
788
Revista de neurociencia cognitiva
Volumen 34, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
oh
C
norte
_
a
_
0
1
8
2
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(Hopf et al., 2006; Luck, Chelazzi, Hillyard, & Desimone,
1997) to lower levels of visual cortex such as V1
(Donohue, Schoenfeld, & Hopf, 2020; see also the work
of Adam & serencias, 2021).
Several studies have demonstrated that task-irrelevant
salient stimuli elicit a PD component, indicating that they
are suppressed (Drisdelle & Eimer, 2021; van Moorselaar
et al., 2021; Berggren & Eimer, 2020; Feldmann-Wüstefeld
et al., 2020; Henare, Kadel, & Schubö, 2020; Gaspelin &
Luck, 2018C; Barras & Kerzel, 2016; Gaspar et al., 2016;
Gaspar & McDonald, 2014; Jannati, Gaspar, & McDonald,
2013; Sawaki & Luck, 2010; Hickey et al., 2009). Para examen-
por ejemplo, Gaspar and McDonald (2014) found that task-
irrelevant color singletons elicited a PD component, pero
no corresponding N2pc component, which was taken to
suggest that the salient item was proactively inhibited to
prevent attentional capture (see also the work of Jannati
et al., 2013; Sawaki & Luck, 2010). Además, Gaspelin
and Luck (2018C) demonstrated that the magnitude of
the PD component was positively correlated with the
magnitude of probe suppression effects across partici-
pants in the capture-probe paradigm (see also the work
of Feldmann-Wüstefeld et al., 2020). Además, Weaver
and colleagues (2017) have shown that successful eye
movements away from salient distractors are preceded by
a PD component, and this overt suppression depended
on the moment the saccade was elicited: When saccades
were triggered before the PD component, there was no evi-
dence for overt suppression (es decir., saccade deviation). De este modo,
the PD component seems to reflect covert suppression
independent of overt suppression. Colectivamente, hay
an abundance of evidence that salient items can be sup-
pressed, at least under certain circumstances, and that the
PD component indexes this covert suppressive process.
Can Highly Salient Stimuli Be Suppressed?
The signal suppression hypothesis, sin embargo, ha sido
recently challenged on the grounds that the singleton dis-
tractor may have been only weakly salient in past studies.
According to Theeuwes and associates, the search displays
used by Gaspelin, Luck, and colleagues were small, típicamente
containing only four to six elements. This small set size may
have reduced the salience of the color singleton. To sup-
port their argument, Wang and Theeuwes (2020) usado
the capture-probe paradigm to compare probe suppression
effects at low display set sizes of four and six, cual tiene
been used previously, and a higher set size of 10, representación
the singleton distractor more salient relative to the nonsin-
gleton distractors. They found probe suppression effects at
the lower set size of four, but probe capture effects (es decir.,
higher probe recall for the singleton distractor than non-
singleton distractors) at the higher set size of 10. Ellos
concluded that highly salient items cannot be suppressed.
There are a few reasons to be skeptical of the above
afirmar. Primero, Gaspelin et al. ran control experiments that
showed that the color singletons in four-item displays
were salient enough to capture attention under singleton
detection mode (Gaspelin et al., 2015, 2017). Más-
más, Gaspelin and Luck (2018a, Experimento 4) presentado
their usual suppression effect with displays of size six; después
acerca de 20 ensayos, eye movements to the singletons were
eliminated. The important point is that during the first
20 ensayos, these same singletons were sufficiently salient
to capture overt attention. All of these studies suggest that
singletons were sufficiently salient to attract attention but
that participants could learn to suppress them under con-
ditions that encourage top–down control. Además,
Chang, Niebur, and Egeth (2021) have demonstrated that
the color singletons used in these studies seem to be
salient as assessed by computational models of saliency.
Además, Stilwell and Gaspelin (2021) provided evi-
dence that singletons can even be suppressed at high set
sizes and that the results of Wang and Theeuwes (2020)
may have been due, at least in part, to a design flaw
whereby increasing the number of probed items caused
a type of floor effect. In Stilwell and Gaspelin (2021), par-
ticipants performed a capture-probe paradigm with large
set sizes of 10 elementos, which should have rendered the sin-
gleton highly salient. En tono rimbombante, the number of probe
letters was independently manipulated to be either 10 o
four letters. The four-letter probe displays allowed probe
accuracy to remain relatively high, preventing any kind of
floor effect that would conceal underlying suppression of
the color singleton. En efecto, probe suppression effects
were observed in the four-letter displays but were greatly
reduced in 10-letter displays, largely because of low
overall probe accuracy. This pattern of results is consistent
with a floor effect and demonstrates that even highly
salient singletons seem to be suppressible, at least under
certain conditions.
En resumen, some recent studies have suggested that
highly salient color singletons cannot be suppressed
( Wang & Theeuwes, 2020; see also the work of Theeuwes,
2004). Although there has been some evidence challeng-
ing this claim (Chang et al., 2021; Stilwell & Gaspelin,
2021), further evidence is needed to definitively demon-
strate that covert attention is not automatically allocated
to highly salient color singletons. The current study will
use ERPs to assess covert attentional allocation to highly
salient color singletons. A benefit of ERP measures is that
they provide a continuous measure of visual processing,
which allows one to assess whether the salient item was
ever covertly attended during an experimental trial. Él
also allows one to assess whether putative indices of
suppression—such as the Pd component—are elicited
by highly salient color singletons.
The Current Study
Participants performed a version of the additional single-
ton task that has previously been shown to produce atten-
tional suppression via the PD component (Gaspelin &
Luck, 2018C), but the task was modified to improve the
Stilwell, Egeth, and Gaspelin
789
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
oh
C
norte
_
a
_
0
1
8
2
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
bottom–up salience of the color singleton. Namely, el
search displays were modified to include additional
target-colored distractors, which should increase relative
bottom-up contrast with the color singleton distractor
(Chang et al., 2021; Stilwell & Gaspelin, 2021; Wang &
Theeuwes, 2020). We independently verified that this
manipulation increased the bottom-up salience of the
distractor using a computational model of salience
(Kotseruba, Wloka, Rasouli, & Tsotsos, 2020; Hou, Harel,
& Koch, 2012). Two separate experiments then assessed
whether this highly salient item elicited a PD component
(indicating suppression) or instead an N2pc component
(indicating attentional capture). If highly salient items
cannot be suppressed as claimed by stimulus-driven
accounts, they should elicit an N2pc component.
EXPERIMENT 1
Experimento 1 used an additional singleton paradigm sim-
ilar to Gaspelin and Luck (2018C) but modified to improve
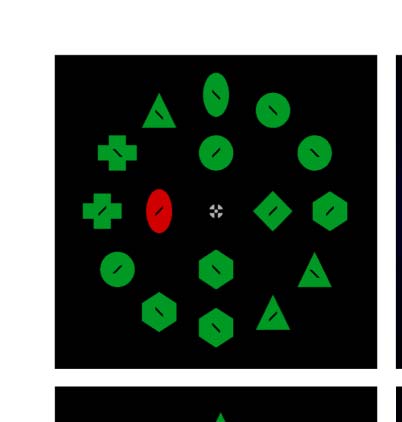
the salience of the color singleton (Cifra 1). Participantes
searched for a specific target shape (p.ej., green diamond)
among a set of heterogeneous distractor shapes (verde
triangles, hexagons, ovals, etc.) and made a speeded
button press regarding the tilt of an inscribed line (izquierda
vs. bien). On some trials, a color singleton distractor
appeared at a nontarget location. The primary search
display consisted of four items, similar to the work of
Gaspelin and Luck (2018C). To improve the salience of
the color singleton, an additional 12 target-colored
dummy shapes were presented around this search display
(see also the work of Stilwell & Gaspelin, 2021). Neither
the target nor the singleton ever appeared in this outer
ring of dummy shapes, which was only used to improve
the salience of the color singleton.
Lateralized ERPs were used to assess attentional alloca-
ción. The singleton and target could appear either on the
vertical midline or at lateralized location. When an item
appears on the vertical midline, the EEG activity elicited
Cifra 1. Stimuli and procedure from Experiment 1. Participantes
searched an array of 16 shapes for the target (p.ej., green diamond) y
made a speeded button press indicating the orientation of a black line
segment inside (as in the work of Gaspelin & Luck, 2018C). En tono rimbombante,
the target and singleton could only appear in the inner ring. The outer
ring was included to boost the salience of the singleton distractor.
by a stimulus should be equal in both visual hemifields,
effectively cancelling out any lateralized difference in
the ERP waveform. When a stimulus is lateralized, cómo-
alguna vez, an N2pc component should be elicited if it is
attended, whereas a PD component should be elicited if
it is suppressed.
Stimulus-driven accounts and the signal suppression
hypothesis make competing predictions about ERPs
on trials where the singleton distractor is lateralized,
and the target appears on the midline (singleton
lateral/target midline trials). Stimulus-driven accounts pre-
dict that the singleton should automatically capture atten-
ción, resulting in an N2pc component (Hickey et al., 2006).
En cambio, the signal suppression hypothesis predicts
that the singleton should be proactively suppressed and
should therefore result in a PD component on singleton
lateral/target midline trials (Gaspelin & Luck, 2018C;
Gaspar & McDonald, 2014; Sawaki & Luck, 2010).
Métodos
Participantes
A sample of 20 participants from State University of New
York at Binghamton participated for monetary compensa-
ción. In the final sample, 14 were women and six were men
(edad media = 18.8 años). Our sample size was determined
a priori based on a power analysis of the pooled effect size
(Cohen’s dz = 1.51) from four recent studies of the PD
component (Sawaki & Luck, 2010, dz = 1.44; Gaspelin &
Luck, 2018C, dz = 1.83; Drisdelle & Eimer, 2021, dz = 1.42;
Kerzel & Burra, 2020, dz = 1.39). With this effect size, 11
participants would be needed to achieve 99% fuerza. Nosotros
chose to err on the side of caution and collect a larger
sample size.
All participants had normal or corrected-to-normal
visual acuity and had normal color vision as indicated by
an Ishihara test. All experimental protocols were approved
by a university ethics board.
Stimuli and Procedure
Stimuli were presented using PsychToolbox for MATLAB
(Brainard, 1997) on an Asus VG245H LCD monitor with
a black background at a viewing distance of 100 cm. El
timing delay of the stimulus presentation system was mea-
sured using a photosensor, and event codes were shifted
off-line to compensate for this delay (12 mseg).
As depicted in Figure 1, each search display contained
16 shapes that were arranged in two concentric rings
(Theeuwes, 2004). The inner ring consisted of four shapes
arranged equidistantly around a notional circle with a
radius of 2.0°, and the outer ring consisted of twelve
shapes arranged in a notional circle with a radius of 4.0°.
The individual shapes were triangles (1.2° by 1.2°), hexa-
gons (1.2° by 1.2°), ovals (1.5° by 0.9°), crosses (1.2° by
1.2°), diamonds (1.2° by 1.2°), and circles (1.2° diameter).
790
Revista de neurociencia cognitiva
Volumen 34, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
oh
C
norte
_
a
_
0
1
8
2
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
The shapes were either red (30.0 cd/m2, x = .627, y =
.330) or green (30.0 cd/m2, x = .292, y = .631), cual
were photometrically isoluminant. Each shape contained
a black line segment (0.4° × 0.1°) tilted either 45° to the
left or right, seleccionado al azar. A gray fixation cross
(0.5° diameter; 30.0 cd/m2, x = .306, y = .320) was con-
tinuously visible throughout the trial.
The target shape (circle or diamond) and target color
(green or red) were held constant for the entire experi-
mental session, and all four potential combinations were
counterbalanced across participants. The inner ring con-
tained a target shape and three distractors. On each trial,
the three distractors were randomly selected from a pool
of the five nontarget shapes without replacement (p.ej.,
triangle, hexagon, oval, cross, or circle in the diamond
target condition). The outer ring contained dummy
shapes that were meant to boost the relative salience
of the singleton distractor (Stilwell & Gaspelin, 2021).
These shapes were also randomly selected from the non-
target shapes. To ensure high distractor heterogeneity,
no shape could be selected more than 3 times in the
outer ring.
On singleton-present trials (75% of trials), a distractor in
the inner ring was rendered in a unique color, creating a
singleton distractor. On singleton-absent trials (25% de
ensayos), todo 16 shapes (in both rings) were the target color.
This was meant to ensure a high number of trials in which
the singleton was present for the primary analysis in this
estudiar (singleton lateral/target midline trials). The location
of the target shape and location of singleton distractor
were selected at random with the exception that that the
singleton distractor could not appear at the target location.
These two items could appear only in the inner ring of
four shapes. This ensured that the singleton distractor
and target shape appeared equally distant from fixation
to avoid introducing a spatial bias (p.ej., see the work of
Woodman & Luck, 1999).
To encourage feature-search mode, the distractor
shapes were carefully selected to reduce search asymme-
intentos. Específicamente, to eliminate any possible use of single-
ton detection mode, we selected distractor shapes that
were both linear and curvilinear to contrast the diamond
and circle target shapes, respectivamente. If only linear shapes
were used as distractors, the circle target might “pop out”
at high set sizes because it would be a curvilinear single-
tonelada, and this could potentially induce singleton detection
mode (Bacon & Egeth, 1994).
Trials began with a fixation cross for 1000 mseg. Próximo,
the search array appeared, and participants attempted to
locate the target shape as quickly as possible. Participantes
responded to the orientation of the line segment inside
the target (left vs. bien) via a speeded manual response
on a gamepad using the left- and right-shoulder buttons,
respectivamente. If participants were too slow to respond (RT
was greater than 2000 mseg), a “Too Slow” screen was
presented for 500 mseg, along with a 200-Hz tone lasting
300 mseg. Incorrect responses were followed by the same
tone for the same duration. The interval between trials
randomly varied between 0 y 500 mseg.
At the beginning of each session, participants were
instructed that the singleton distractor would never be
the target and should be ignored. Además, Participantes
were instructed that the target would only appear in the
inner ring of shapes. Eye movements were discouraged
using the on-line horizontal EOG waveforms; la experiencia-
menter reminded participants to maintain fixation through-
out the trial whenever eye movements were detected.
Each session began with one practice block of 192 ensayos.
This was followed by nine regular blocks of 192 ensayos
(1728 trials in total). This yielded 1296 singleton-present
y 432 singleton-absent trials per participant. The key
hypotheses in the current experiment relate to trials
where the singleton was presented at a lateral location
and the target was on the vertical midline. For each par-
ticipant, there were approximately 324 of these singleton
lateral/target midline trials, which should yield high statis-
tical power to detect any significant effect. Each block was
divided into four miniature blocks of 48 trials that were
separated by 15-sec breaks to allow participants to blink
and/or adjust seating position. At the end of each full
block, a 5-min break was provided. Participants received
block-by-block feedback on mean RT and accuracy.
Electrophysiological Recording and Analysis
The EEG was recorded using active Ag/AgCl electrodes
(Brain Products actiCHamp system) de 27 scalp sites
(FP1, FP2, F7, F3, Fz, F4, F8, C3, Cz, C4, P9, P7, P5, P3,
Pz, P4, P6, P8, P10, PO7, PO3, POz, PO4, PO8, O1, Oz,
and O2, according to the modified 10–20 system; Ameri-
can Electroencephalographic Society, 1994) referenced to
each mastoid. To detect eye movements and blinks, el
EOG was recorded from electrodes placed at the outer
canthi of each eye and below the right eye. All signals were
recorded in single-ended mode using a customized
version of the PyCorder recording software and then
referenced off-line. The EEG was filtered on-line with a
cascaded integrator–comb antialiasing filter with a half-
power cutoff at 130 Hz and then digitized with a 500-Hz
sampling rate.
After data acquisition, all analyses were conducted using
ERPLAB Toolbox (Lopez-Calderon & Luck, 2014) y
EEGLAB Toolbox (Delorme & Makeig, 2004). EEG signals
were referenced to the average of the left and right mas-
toids, and the four EOG signals were averaged for each
direction (horizontal and vertical) for each corresponding
pair of electrodes. These signals were then filtered off-
line using a noncausal Butterworth high-pass filter (half-
amplitude cutoff: 0.1 Hz, slope: 12 dB/octave). Usando un
600-msec epoch, beginning 200 msec before stimulus
onset, averaged ERP waveforms were computed. Para
plotting purposes only and to maximize temporal preci-
sión, a low-pass filter (half-amplitude cutoff: 30 Hz, slope:
12 dB/octave) was applied to the averaged ERPs.
Stilwell, Egeth, and Gaspelin
791
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
oh
C
norte
_
a
_
0
1
8
2
7
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Trials were excluded if an incorrect behavioral result
was produced or if manual RT was faster than 200 mseg
or longer than 1500 mseg. Además, trials were excluded
if a blink or eye movement was detected during the trial in
either the horizontal or vertical EOG channels as indexed
by step-like voltages in the EEG signal. Blinks were identi-
fied as a voltage step exceeding 80 μV between 200 y
400 msec in channels FP2 and VEOG. Saccades were iden-
tified as voltage steps exceeding 16 μV for saccades
entre 100 y 400 msec in the bipolar HEOG channel.
To ensure that all eye movements were eliminated, nosotros
used an approach similar to Woodman and Luck (2003).
Grand averaged bipolar HEOG waveforms were computed
for left- and right-target trials. Participants were replaced if
bipolar HEOG deviated more than 3.2 μV between 100 y
400 msec poststimulus. Two participants were replaced
for this reason. This approach ensures that eye move-
ments did not exceed ±0.1° in the direction of the target
(Lins, Picton, Iceberg, & Scherg, 1993) during the critical time
windows used to assess the N2pc and PD components (ver
also the work of Gaspelin & Luck, 2018C). We planned to
replace any participant with more than 25% of trials
excluded because of any combination of exclusion criteria
aforementioned. Sin embargo, in this experiment, no partici-
pants had to be replaced for this reason. In the final set of
Participantes, an average of 6.8% of trials were excluded
because of artifacts (range = 0.4–19.5%).
All subsequent EEG analyses were performed using the
PO7 and PO8 electrode sites, which were chosen a priori
based on previous studies of the PD component (Gaspelin
& Luck, 2018C; Gaspar & McDonald, 2014; Sawaki & Luck,
2010). The N2pc and PD components were calculated by
subtracting the ipsilateral waveform from contralateral wave-
form to create difference waveforms over time windows
chosen a priori based on the work of Sawaki and Luck
(2010). Eso es, the N2pc component was measured as the
mean amplitude between 225 y 300 mseg, and the PD
component amplitude was quantified as the mean ampli-
tude of the difference waveform between 115 y 225 mseg.
This strategy of using an earlier time window for the PD ERP
component than the N2pc component is consistent with
several previous studies (Drisdelle & Eimer, 2021; Gaspelin
& Luck, 2018C; Weaver et al., 2017; Sawaki & Luck, 2010).
To avoid issues of sphericity, all reported p values in this
article were Greenhouse–Geisser corrected for ANOVAs
with more than two levels of a given factor. When appro-
priate, we adjusted the partial eta squared in our reports of
effect size for positive bias (Mordkoff, 2019). For within-
subject t tests, Cohen’s dz was used to measure effect size
(Lakens, 2013).
Resultados
Behavioral Results
Figure 2A depicts mean RTs for singleton-present and
singleton-absent trials. As can be seen, RTs were
Cifra 2. Manual RT results for Experiment 1. Mean manual RT as a
function of singleton presence for Experiment 1. Error bars represent
within-subject confidence intervals.
significantly faster on singleton-present trials (679 mseg)
than singleton-absent trials (701 mseg), t(19) = 6.95,
pag < .001, dz = 1.40. This 22-msec singleton-presence
benefit replicates previous studies using displays that
encourage feature-search mode (Chang & Egeth, 2019,
2021; Stilwell & Gaspelin, 2021; Gaspelin & Luck, 2018c;
Gaspelin et al., 2015; Vatterott & Vecera, 2012). This pat-
tern suggests that the color singleton was proactively sup-
pressed, as if reducing the effective set size of the search
display by one search item when it was present. Error rates
were not significantly different between singleton-present
(3.1%) and singleton-absent (3.3%) trials, t(19) = 0.62,
p = .54, dz = 0.14. Altogether, the behavioral results dem-
onstrate no evidence of attentional capture by the color
singleton and instead suggest that it was suppressed.
Electrophysiological Results
Figure 3 depicts grand-averaged ERP waveforms for the lat-
eral occipital sites (PO7 and PO8) for targets and singleton
distractors. Separate waveforms are shown for electrodes
contralateral and ipsilateral relative to the search item of
interest. For example, the ipsilateral waveform for the tar-
get was the average of the right hemisphere electrode
when the target appeared in the right visual field and the
left hemisphere electrode when the target appeared in the
left visual field. The contralateral waveform was the aver-
age of the right hemisphere electrode when the target
appeared in the left visual field and the left hemisphere
electrode when the target appeared in the right visual
field. Trials in which the singleton distractor and target
were both lateralized were excluded from the analyses
because the lateralized N2pc component to the target
and lateralized PD component to the singleton distractor
would summate (e.g., Hickey et al., 2009), preventing a
dissociation of target enhancement versus singleton
suppression.
792
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Electrophysiological results for Experiment 1. (A–C) depict ipsilateral and contralateral waveforms for each potential search display
configuration. In the schematics of search displays, the target was the green diamond and the singleton distractor was the uniquely colored item.
(D) depicts contra-minus-ipsi difference waveforms for each display configuration. All waveforms in this article were low-pass filtered to improve
the visibility of the effects (Butterworth noncausal filter, half-amplitude cutoff = 30 Hz, slope = 12 dB/octave) but were analyzed using unfiltered
waveforms.
When the target was presented at a lateralized location
and the singleton was absent (Figure 3A), there was a
negative-going deflection in the contralateral waveform
compared to the ipsilateral waveform beginning at approx-
imately 200 msec poststimulus: an N2pc component. This
N2pc component appeared with approximately the same
amplitude and time course when the target was lateralized
and the singleton was present (Figure 3B). Conversely,
when the singleton distractor was lateralized, and the tar-
get appeared on the vertical midline (Figure 3C) during
the interval beginning at approximately 130 msec post-
stimulus, there was an increased positive voltage in the
contralateral waveform compared to the ipsilateral wave-
form: a PD component. When the target is on the vertical
midline, the lateralized activity across hemifields cancels
out, isolating the ERP waveform to the singleton distractor.
Figure 3D depicts contralateral-minus-ipsilateral differ-
ence waveforms for direct comparison of lateralization
across conditions. Similar to previous studies (Drisdelle
& Eimer, 2021; Gaspelin & Luck, 2018c; Sawaki & Luck,
2010), the positivity elicited by the singleton distractor
peaks before the negativity elicited by the target, lending
further support for the notion that the time window of
the PD component should be earlier than the N2pc
component.
Mean Amplitude during the N2pc Time Window
The N2pc components were measured as the mean
amplitudes from 225 to 300 msec poststimulus in the
contralateral-minus-ipsilateral difference waveforms.
These mean amplitudes were then analyzed using a one-
way within-subject ANOVA comparing the three trial types
(target lateral/singleton absent, target lateral/singleton
midline, and singleton lateral/target midline). There was
a significant main effect of Trial Type, F(2, 38) = 25.50,
p < .001, ηp
2 = .551. Preplanned t tests compared the
difference waveforms between trial types. The N2pc
component was not significantly different on target
lateral/singleton absent trials (−1.1 μV ) and target
lateral/singleton midline trials (−1.2 μV), t(19) = 0.76,
p = .46, dz = 0.17. This provides preliminary evidence that
the singleton distractor did not capture attention. If the
singleton had captured attention, target detection should
have been impaired on trials where the singleton was pres-
ent, reducing or perhaps delaying the N2pc component
evoked by the lateralized target on singleton present trials
compared to singleton absent trials. However, no such
pattern was observed. On singleton lateral/target midline
trials, the amplitude of the singleton-evoked activity was
both positive in polarity (+0.1 μV) and was significantly
larger than the amplitude of the target-evoked negativity
both on target lateral/singleton midline trials, t(19) =
5.71, p < .001, dz = 1.28, and target lateral/singleton
absent trials, t(19) = 4.94, p < .001, dz = 1.10.
Planned one-sample t tests compared ipsilateral-
minus-contralateral difference waveforms against zero
for each trial type (target lateral/singleton absent, target
lateral/singleton midline, and singleton lateral/target
midline). There was a significant N2pc component
on both target lateral/singleton absent trials (−1.1 μV),
t ( 1 9 ) = 5 . 3 5 , p < . 0 0 1 , d z = 1 . 2 0 , a n d t a r g e t
lateral/singleton midline trials (−1.2 μV), t(19) = 5.50,
p < .001, dz = 1.22. Critically, there was not a significant
N2pc component on singleton lateral/target midline
trials (+0.1 μV), t(19) = 0.63, p = .54, dz = 0.14.
Stilwell, Egeth, and Gaspelin
793
Mean Amplitude during the PD Time Window
The same analyses from the previous section were con-
ducted, but mean amplitudes were calculated between
115 and 225 msec in the contralateral-minus-ipsilateral
difference waveforms to assess the PD component. These
mean amplitudes were analyzed using a one-way within-
subject ANOVA comparing the three trial types (target
lateral/singleton absent, target lateral/singleton midline,
and singleton lateral/target midline). There was a main
effect of Trial Type, F(2, 38) = 14.07, p < .001, ηp
2 = .395.
Preplanned t tests compared the contralateral-minus-
ipsilateral difference waveforms between trial types. The
magnitude of the PD component to the target stimulus
did not differ on target lateral/singleton absent (−0.2 μV)
and target lateral/singleton midline trials (−0.3 μV ),
t(19) = 1.38, p = .19, dz = 0.31. The PD component was
larger on singleton lateral/target midline trials (+0.4 μV)
than target lateral/singleton midline trials, t(19) = 4.69,
p < .001, dz = 1.05, and target lateral/singleton absent trials,
t(19) = 3.41, p = .003, dz = 0.76.
Planned one-sample t tests compared contralateral-
minus-ipsilateral difference waveforms against zero for
each trial type (singleton lateral/target midline, target
lateral/singleton absent, and target lateral/singleton mid-
line). Critically, there was a significant PD component on
singleton lateral/target midline trials (+0.4 μV), t(19) =
4.36, p < .001, dz = 0.98. No such PD component was
observed on trials where the target was lateralized. If
anything, there was a lateralized negativity. On target
lateral/singleton midline trials, there was significant
negative-going deflection evoked (−0.3 μV), t(19) = 3.33,
p = .004, dz = 0.74. On target lateral/singleton absent trials,
there was no statistically reliable positive- or negative-going
deflection (−0.2 μV), t(19) = 1.58, p = .13, dz = 0.35.
Exploratory Analysis: Post-PD Negativity
As shown in Figure 3D, on singleton lateral/target mid-
line trials, there is a negative-going deflection that
occurred after the PD component from approximately
275–375 msec. We did not have any specific hypotheses
about this post-PD negativity. It has occurred in some previ-
ous studies (Drisdelle & Eimer, 2021; Feldmann-Wüstefeld
et al., 2020; Gaspelin & Luck, 2018c, Experiment 3; Kerzel &
Burra, 2020) but not others (Gaspelin & Luck, 2018c;
Gaspar & McDonald, 2014, Experiment 1; Sawaki & Luck,
2010). The exact cognitive process underlying this post-PD
negativity is currently unknown. Some have proposed that
it could reflect attentional capture by the singleton distrac-
tor on a small portion of trials or participants, producing a
combination of an N2pc and a PD component in the grand
average waveform even though they never occurred on
the same trial (Gaspelin & Luck, 2018b). Others have sug-
gested that this negativity may result from a search strategy
at low set sizes, whereby the two locations on the horizon-
tal midline are prioritized during visual search. The second
negativity could represent either a secondary shift of atten-
tion to the singleton (Kerzel & Burra, 2020) or delayed
suppression of the nonsingleton distractor opposite of
the singleton distractor (Drisdelle & Eimer, 2021).
We conducted an exploratory analysis to assess the sig-
nificance of the negativity from 275–375 msec using a one-
sample t test on the difference waveform on singleton
lateral/target midline trials. This indicated the negativity
(−0.7 μV) was statistically significant, t(19) = 5.36, p <
.001, dz = 1.20. The post-PD component did not signifi-
cantly correlate with the magnitude of the singleton-
presence benefit, (r = .29, p = .22). Ultimately, it is
currently unknown what cognitive process the post-PD
negativity reflects, but it seems to be unrelated to distrac-
tor suppression.
Saliency Maps
Saliency maps were used to independently verify the
salience of the color singleton (see the work of Chang
et al., 2021; Stilwell & Gaspelin, 2021). A set of 1000 search
displays were generated (250 for each counterbalancing
condition) using the stimulus code. The resulting images
were then processed using the Image Signature Toolbox
in MATLAB (Hou et al., 2012) to generate saliency maps.
This toolbox was chosen because it performs similarly to
human observers in its ability to detect singletons in artifi-
cial images, unlike many canonical saliency models
(Kotseruba et al., 2020). The default settings were used,
with the exception of the mapWidth() parameter, which
was adjusted to accommodate the image resolution
(1920 × 1080). The output was a series of saliency maps
that resemble heatmaps depicting the relative distribution
of salience across the image (Figure 4). For each saliency
map, a circular region of interest (1.8° in diameter) was
Figure 4. Saliency map results for both experiments. Stimulus images
(n = 1,000 for each experiment) were analyzed using the Image
Signature Toolbox (Hou et al., 2012) for each experiment to determine
if the singleton was truly salient. In this example, the saliency map
indicated a high salience score at the location of the color singleton.
794
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
defined encompassing each of the 16 search items. The
mean salience score at each location was calculated by
averaging the pixels in the interest area.
Mean salience scores were used to calculate two metrics
of salience: the global saliency index (GSI) and the single-
ton win rate. First, the GSI was computed as the mean
salience score at the singleton distractor minus the mean
salience score at the average of all nonsingleton locations
(Kotseruba et al., 2020). The difference score was normal-
ized by dividing by the sum of the salience scores of all
search items. The GSI is a continuous metric ranging from
−1 to 1. A positive value indicates that the singleton is
more salient than the average nonsingleton item, whereas
negative values indicate that the singleton is less salient
than the average nonsingleton item. A value of zero indi-
cates that the singleton is as salient as the average of all of
the other items. The average GSI across all images was
0.81 (SE = 0.003), t(999) = 320.67, p < .001, dz = 10.14,
indicating the singleton had an average salience score that
was much higher than the other search items. The second
metric to assess salience was the singleton win rate, which
is the percentage of images in which the singleton had
the highest salience score of all items in the saliency
map. The singleton win rate was 100%. This means that
the singleton distractor was selected as the most salient
item in the display in every singleton simulation image.
We also compared singleton salience in 16-item displays
of the current study with four-item displays (resembling
the work of Gaspelin & Luck, 2018c). The basic goal was
to ascertain whether increasing the set size actually
increased the salience of the color singleton compared
to previous studies. Four-item displays were created by
removing the outer ring of dummy shapes. Four-item dis-
plays produced an average GSI of 0.47 (SE = 0.005), which
was significantly lower than the GSI of 0.81 (SE = 0.003)
produced in the current study, t(999) = 59.72, p < .001,
dz = 1.89. However, the singleton win rate was still 100%
of trials, even in the four-item displays. These results sug-
gest that the singletons in the current study were more
salient than the four-item displays of Gaspelin and Luck
(2018c). However, it is important to highlight that even
the displays of Gaspelin and Luck (2018c) seem to suggest
that the singletons used were highly salient: The GSI of
0.47 was significantly above 0, t(999) = 93.99, p < .001,
dz = 2.97, and the singleton win rate was 100%.
Discussion
Experiment 1 tested whether highly salient color single-
tons can be suppressed or instead capture attention. To
improve the salience of the color singleton, we increased
the set size compared to previous studies from four items
to 16 items (Stilwell & Gaspelin, 2021; Wang & Theeuwes,
2020). Indeed, computational models of salience sug-
gested that increasing the display set size improved the
salience of the color singleton. Despite increasing the
salience of the singleton distractor, the singleton did not
capture attention. First, manual RTs were significantly
faster when the singleton was present than absent, sug-
gesting suppression of the singleton. Second, there was
a significant PD component on singleton lateral/target
midline trials, reflecting suppression of the salient single-
ton. Altogether, these results suggest that the salient sin-
gleton distractor was proactively suppressed, supporting
the signal suppression hypothesis and refuting stimulus-
driven accounts.
EXPERIMENT 2
In Experiment 1, the color singleton elicited a PD compo-
nent, which, along with the behavioral results, would seem
to indicate that the color singleton was suppressed. It is
possible, however, that adding an outer ring of dummy
shapes may have encouraged participants to restrict visual
processing to only the inner ring of shapes. If the saliency
computations between the singleton and nonsingleton
distractors were somehow restricted to the inner ring of
shapes, the singleton would be no more salient than pre-
vious demonstrations using four-item search arrays (such
as in the work of Gaspelin & Luck, 2018c).
Experiment 2 was therefore designed to increase the
salience of the singleton distractor by increasing the num-
ber of items at task-relevant locations. The search displays
were similar to Experiment 1 but had an inner ring of eight
items rather than four items (see Figure 5). This manipu-
lation guarantees that the singleton would be more
salient than in Experiment 1. The theoretical predictions
are identical to Experiment 1. According to the signal sup-
pression hypothesis, the singleton distractor should be
suppressed, resulting in a PD component when it is later-
alized. According to the stimulus-driven account, how-
ever, the singleton distractor should capture attention,
resulting in an N2pc component when it is lateralized. In
addition, the increased set size of the inner ring of shapes
also allows us to test a recent alternative account of the PD
component (see the “Idiosyncratic Search Hypothesis: PD
Horizontal Midline Analysis” in the Results section; Kerzel
& Burra, 2020).
Figure 5. Example search array from Experiment 2. The search displays
in Experiment 2 were identical to Experiment 1 except for the addition
of four distractors in the inner ring, raising the set size of the inner ring
from four to eight items. The overall display contained four more
distractors, rendering the color singleton distractor even more salient
than Experiment 1.
Stilwell, Egeth, and Gaspelin
795
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
addition, lateralized color singletons elicited a PD compo-
nent. Importantly, this PD component occurred at all
lateralized positions supporting the signal suppression
hypothesis.
Behavioral Results
Mean RTs for Experiment 2 are depicted in Figure 6. Over-
all RTs were slower in Experiment 2 (840 msec) than
Experiment 1 (690 msec), t(38) = 5.90, p < .001, dz =
1.86, suggesting that search was more difficult in Experi-
ment 2 because of the higher set size (e.g., Wolfe, 2021;
Wolfe & Horowitz, 2017) and/or increased visual crowding
( Whitney & Levi, 2011).
In Experiment 2, mean RTs were numerically faster on
singleton-present trials (837 msec) than singleton-absent
trials (843 msec) but this difference was nonsignificant,
t(19) = 1.64, p = .12, dz = 0.37. This lack of a singleton
presence cost suggests that the color singleton did not
capture attention. It is interesting to note that the
singleton-presence benefit was smaller in Experiment 2
(−6 msec) than Experiment 1 (−22 msec), t(38) = 3.43,
p = .001, dz = 1.08. This is consistent with the notion that
that suppressing the singleton is akin to reducing the set
size by an item. The net benefit of this suppression would
thereby be weaker in Experiment 2 (reducing the relevant
set size from eight to seven items) than Experiment 1
(reducing the relevant set size from four to three items).
Error rates were not significantly different between
singleton-present (2.9%) and singleton-absent (2.6%) tri-
als, t(19) = 1.60, p = .13, dz = 0.36.
Electrophysiological Results
The same exclusion criteria (i.e., artifact rejection and
participant replacement) from Experiment 1 were used
in Experiment 2. In the final set of participants (six
Methods
Participants
A new sample of 20 participants from State University of
New York at Binghamton (16 women and 4 men; mean
age = 21.5 years) participated for monetary compensa-
tion. The sample size was determined a priori to match
Experiment 1. A power analysis of the effect size observed
for the PD component in Experiment 1 (dz = 0.98) sug-
gested this sample size would yield 99% power to detect
a significant effect.
Stimuli and Procedure
The experimental task was identical to Experiment 1,
except that four nonsingleton distractor shapes were
added to the inner ring (see Figure 5). These shapes
appeared at the four diagonal positions between the four
cardinal positions used in Experiment 1. All search items in
the inner ring were equidistant from central fixation. On
each trial, the target shape was equally likely to appear at
each inner ring location. The remaining five distractor
shapes were randomly selected to appear in the remaining
inner ring and outer ring positions with the constraint that
a given distractor shape could only be used at most two
times in the inner ring or three times in the outer ring. This
was meant to maximize heterogeneity of distractors and
encourage feature-search mode.
In ERP studies, it is important to ensure that there are
enough trials per condition to prevent statistical noise that
could lead to a Type I error (Luck & Gaspelin, 2017). By
adding four items to inner ring of the search display, this
reduced the number of trials in which a search items
appeared on the vertical or horizontal midline. To improve
statistical power, the three lateralized locations on either
side of the vertical midline were collapsed to form a single
lateralized condition for each hemifield. For example, sin-
gleton lateral/target midline trials included any trial in
which the singleton appeared at one of the three latera-
lized locations. Similar approaches have been used in
previous studies that use high set sizes (e.g., Feldmann-
Wüstefeld, Weinberger, & Awh, 2021; Gaspar et al., 2016;
Gaspar & McDonald, 2014). This approach did not signifi-
cantly alter the results: The results of these full-display anal-
yses were similar to the results of Experiment 1 and the
partial-display analyses (i.e., restricting analyses to items
appearing on the vertical and horizontal midlines) of
Experiment 2, albeit noisier. In addition, for the key ERP
component (singleton lateral/target midline), we analyzed
separate waveforms for when the singleton was and was
not on the horizontal midline, which allows for a direct
comparison with Experiment 1 (see “Idiosyncratic Search
Hypothesis: PD Horizontal Midline Analysis” and Figure 8).
Results
To summarize, the basic results are similar to Experiment 1.
There was no singleton-presence cost on mean RT. In
Figure 6. Manual RT results for Experiment 2. Mean manual RT as a
function of singleton presence for Experiment 2. Error bars represent
within-subject confidence intervals.
796
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Electrophysiological results for Experiment 2. (A–C) depict ipsilateral and contralateral waveforms for each potential search display
configuration. In the schematics of search displays, the target was the green diamond and the singleton distractor was the uniquely colored item. (D)
depicts contra-minus-ipsi difference waveforms for each display configuration.
participants were replaced because of eye movements),
an average of 5.8% of trials were excluded because of arti-
facts (range = 0.0–20.8%).
Figure 7 depicts grand-averaged ERP waveforms for the
lateral occipital sites (PO7 and PO8) for targets and single-
ton distractors. When the target was presented at a later-
alized location, there was an N2pc component starting at
approximately 200 msec poststimulus, which was similar
in magnitude when the singleton was present or absent
(Figure 7A and 7B). Conversely, when the singleton
distractor was presented at a lateralized location and
the target appeared on the vertical midline, there was a
PD component beginning at 130 msec with no corre-
sponding N2pc component (Figure 7C).
Mean Amplitude during the N2pc Time Window
Mean amplitudes from 225 to 300 msec were analyzed
using a one-way within-subject ANOVA comparing the
three trial types (target lateral/singleton absent, target
lateral/singleton midline, and singleton lateral/target
midline). There was a significant main effect of Trial Type,
F(2, 38) = 6.23, p = .005, ηp
2 = .207. Preplanned t tests
compared the contralateral-minus-ipsilateral difference
waveforms between trial types. Importantly, the N2pc
component evoked by the lateralized target did not differ
significantly based upon whether a singleton appeared
on the midline (i.e., target lateral/singleton absent and
target lateral/singleton midline conditions, −0.4 μV for
both), t(19) = 0.31, p = .76, dz = 0.07. On singleton
lateral/target midline trials, there was a negative-going
deflection (−0.1 μV) that was significantly smaller than
the N2pc component on both target lateral/singleton mid-
line trials (−0.4 μV), t(19) = 3.48, p = .003, dz = 0.78, and
target lateral/singleton absent trials (−0.4 μV), t(19) =
2.46, p = .024, dz = 0.55.
Planned one-sample t tests compared ipsilateral-
minus-contralateral difference waveforms against zero
for each trial type (target lateral/singleton absent, target
lateral/singleton midline, and singleton lateral/target
midline). There was a significant N2pc component on both
target lateral/singleton absent trials (−0.4 μV), t(19) = 3.79,
p = .001, dz = 0.85, and target lateral/singleton midline
trials (−0.4 μV, t(19) = 4.64, p < .001, dz = 1.04. Criti-
cally, there was not a significant N2pc component on
singleton lateral/target midline trials (−0.1 μV), t(19) =
0.73, p = .47, dz = 0.16.
Mean Amplitude during the PD Time Window
Mean amplitudes were calculated between 115 and
225 msec in the contralateral-minus-ipsilateral difference
waveforms to assess the PD component. These mean
amplitudes were analyzed using a one-way within-subject
ANOVA comparing the three trial types (target lateral/
singleton absent, target lateral/singleton midline, and sin-
gleton lateral/target midline). There was a main effect of
Trial Type, F(2, 38) = 12.57, p < .001, ηp
2 = .367. Pre-
planned t tests compared the contralateral-minus-
ipsilateral difference waveforms between trial types. The
magnitude of the PD component to the target stimulus
did not differ significantly between the two singleton
conditions (i.e., target lateral/singleton absent, −0.1 μV,
and target lateral/singleton midline conditions, 0.0 μV),
t(19) = 0.65, p = .52, dz = 0.15. The PD component was larger
on singleton lateral/target midline trials (+0.4 μV) than tar-
get lateral/singleton midline trials (−0.1 μV), t(19) = 4.20,
p < .001, dz = 0.94, and target lateral/singleton absent
trials (0.0 μV), t(19) = 4.99, p < .001, dz = 1.12.
Stilwell, Egeth, and Gaspelin
797
Planned one-sample t tests compared contralateral-
minus-ipsilateral difference waveforms against zero for
each trial type (singleton lateral/target midline, target
lateral/singleton absent, and target lateral/singleton mid-
line). Critically, there was a significant PD component on
singleton lateral/target midline trials (+0.4 μV), t(19) =
5.37, p < .001, dz = 1.20. As in Experiment 1, no such
PD component was observed on trials where the target
was lateralized. If anything, there was a lateralized
negativity. On target lateral/singleton absent trials,
there was a nonsignificant negative-going deflection
evoked (−0.1 μV), t(19) = 1.48, p = .16, dz = 0.33. On
target lateral/singleton midline trials, there was no statisti-
cally significant positive- or negative-going deflection
(0.0 μV), t(19) = 0.31, p = .76, dz = 0.07.
Experiment 1 versus Experiment 2
As can be seen in Figures 3D and 7D, there was a decrease
in the magnitude of the target-elicited N2pc components
from Experiment 1 to Experiment 2. This suggests that the
target was more difficult to locate in Experiment 2,
because of the higher set size (e.g., Wolfe, 2021; Wolfe
& Horowitz, 2017) and/or increased visual crowding of
search items ( Whitney & Levi, 2011). This difficulty in
locating the target would cause nontarget items to be ini-
tially attended more frequently than in Experiment 1,
decreasing the magnitude of the N2pc component. This
interpretation is also consistent with the increased mean
RT from Experiment 1 to Experiment 2.
To formally assess this, the mean amplitudes of the
contra-minus-ipsi difference waveforms were compared
during the N2pc time window for trials with lateralized tar-
gets (i.e., target lateral/singleton absent and target
lateral/singleton midline) between each experiment using
independent-samples t tests. On target lateral/singleton
absent trials, the mean amplitude of the N2pc component
was reduced from Experiment 1 (−1.1 μV) to Experi-
ment 2 (−0.4 μV), t(38) = 3.10, p = .004, dz = 0.98.
Similarly, on target lateral/singleton midline trials, the
mean amplitude of the N2pc component was reduced from
Experiment 1 (−1.2 μV) to Experiment 2 (−0.4 μV), t(38) =
3.25, p = .002, dz = 1.03.
Interestingly, the magnitude of the singleton-elicited PD
component was unaffected by the increase in set size
between experiments. The mean amplitude of the PD
components on singleton lateral/target midline trials
were similar in Experiment 1 (+0.4 μV) and Experi-
ment 2 (+0.4 μV), t(38) = 0.37, p = .716, dz = 0.12.
This is consistent with the notion that the singleton was
suppressed in both experiments: It was quickly detected
on a preattentive feature map and suppressed before
the first shift of covert attention to prevent any atten-
tional orienting to that item.
Idiosyncratic Search Hypothesis: PD Horizontal
Midline Analysis
The increased set size of items in the inner display also
allowed us to test an alternative account of the PD compo-
nent (see Figure 8A). Recently, Kerzel and Burra (2020)
proposed the idiosyncratic search hypothesis in which
low set sizes encourage an unusual search strategy of pri-
oritizing search items on the horizontal midline. This bias
toward the two items on the horizontal midline causes an
issue on trials where the singleton distractor appears on
the horizontal midline. The nonsingleton distractor at
the opposite location will automatically attract attention
because it matches the target color, yielding an N2pc com-
ponent. This N2pc component (a negative-going deflec-
tion) could easily be confused with a PD component
(i.e., a positive-going deflection) to the singleton distrac-
tor on the opposite side. Thus, by this account, the “PD”
component to the singleton distractor in Gaspelin and
Luck (2018c) is actually an N2pc component to the nonsin-
gleton distractor opposite to the singleton distractor on
the horizontal midline.
The idiosyncratic search hypothesis makes an impor-
tant prediction in Experiment 2: This problem should be
eliminated on trials where the singleton is not on the hor-
izontal midline. On these trials, the two locations on the
horizontal midline are occupied by nonsingleton distrac-
tors, and they should have, on average, an equal atten-
tional priority. As a result, there should be no bias of an
N2pc component to one lateralized search item on the
horizontal midline. Thus, according to this hypothesis,
Figure 8. PD horizontal midline analysis for Experiment 2. (A) The singleton distractor could appear either on a location that is on the horizontal
midline or that is not on the horizontal midline. The white dashed line shows the horizontal midline for illustrative purposes and was not included in
the actual experiment. (B) The contra-minus-ipsi difference waveforms on singleton lateral/target midline trials as a function of whether or not the
singleton appeared on the horizontal midline.
798
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
the PD component—which is purportedly an N2pc com-
ponent to a nonsingleton distractor on the horizontal
midline—should be eliminated on trials where the single-
ton distractor is not on the horizontal midline.
As can be observed in Figure 8B, a PD component was
evoked by the singleton distractor regardless of whether
or not it appeared on the horizontal midline. One-sample
t tests comparing the mean amplitude (115–225 msec) of
each PD component to zero were performed. There was a
significant PD component evoked by the singleton distrac-
tor both when the singleton appeared on the horizontal
midline (+0.3 μV ), t(19) = 2.48, p = .023, dz = 0.56,
and critically, when it did not (+0.4 μV), t(19) = 5.44,
p < .001, dz = 1.22. Interestingly, the mean amplitude
of the PD component was numerically larger when the
singleton was not on the midline than when it was;
however, there was no significant difference between
the magnitude of these two PD components, t(19) =
1.31, p = .21, dz = 0.29. These results are inconsistent with
the idiosyncratic search hypothesis, and instead support
the signal suppression hypothesis.
Exploratory Analysis: Post-PD Negativity
In Experiment 1, there was a negative-going deflection
after the initial PD component on singleton lateral/target
midline trials, which has been observed in some previous
studies. This post-PD negativity was not apparent in Exper-
iment 2 (Figure 7D). An exploratory analysis was con-
ducted to assess the significance of the negativity from
275–375 msec using a one-sample t test on the difference
waveforms. This analysis indicated the late negativity
(−0.1 μV ) was nonsignificant, t(19) = 0.69, p = .50,
dz = 0.15. This late negativity was significantly smaller in
mean amplitude than Experiment 1, t(38) = 4.04, p < .001,
dz = 1.28. Its magnitude did not correlate with the magni-
tude of the singleton-presence benefit (r = −.21, p = .37).
Altogether, these results indicate that the post-PD nega-
tivity did not appear consistently in our two experiments
(see also the work of Gaspelin & Luck, 2018c) and seems
to be unrelated to distractor suppression.
Saliency Maps
As depicted in Figure 4, saliency maps were again used to
independently assess the salience of the color singleton
(see the work of Chang et al., 2021; Stilwell & Gaspelin,
2021). We generated 1000 search arrays, and the resulting
images were processed using the Image Signature Tool-
box in MATLAB (Hou et al., 2012). We calculated two
saliency metrics: the GSI and the singleton win rate. The
average GSI across all images from Experiment 2 was 0.82
(SE = 0.002), indicating the singleton had an average
salience score that was much higher than the other search
items. This GSI was significantly higher than a four-item
display, t(1998) = 62.64, p < .001, dz = 2.80. It was also
slightly larger than Experiment 1 (GSI = 0.81, SE =
0.003), t(1998) = 3.68, p < .001, dz = 0.16. The singleton
win rate in Experiment 2 was 100%, which is equivalent
to Experiment 1 and the four-item displays matching
Gaspelin and Luck (2018c). Together, these saliency
map results suggest that the singleton was highly salient,
and if anything, the singleton was more salient in Experi-
ment 2 than in Experiment 1 and previous studies using
four-item displays (Gaspelin & Luck, 2018c).
Discussion
To boost the salience of the singleton distractor, Experi-
ment 2 increased the number of items in the inner (task-
relevant) ring to test between two competing models: the
stimulus-driven account and the signal suppression
hypothesis. Despite the increased salience of the single-
ton distractor, there was no evidence of a singleton pres-
ence cost in the manual RTs. There was also no evidence of
attentional capture in the ERPs. The N2pc component to
lateralized target was unaffected by the presence of a sin-
gleton on the midline. In addition, the singleton distractor
evoked a PD component, which has been previously estab-
lished to indicate suppression. All of these results are
inconsistent with a stimulus-driven account, which would
directly predict that the singleton distractor capture atten-
tion and interfere with search.
It is worth noting that the increased set size in Experi-
ment 2 seemed to impact target processing independent
of suppression of the salient singleton. Manual RTs were
slower in Experiment 2 than Experiment 1, and the N2pc
components elicited by the lateralized target were
reduced in mean amplitude from Experiment 1 to 2. How-
ever, the mean amplitude of the PD component elicited by
the salient singleton distractor was unaffected by the
increased set size between experiments. Together, these
results suggest that increasing the number of task relevant
items in Experiment 2 rendered the target more difficult to
localize than in Experiment 1, but this set size manipula-
tion did not impact suppression of the singleton distrac-
tor. This pattern is consistent with the signal suppression
hypothesis that predicts that the singleton distractor
was proactively suppressed. Furthermore, this pattern of
results adds to a growing literature suggesting that
distractor processing is independent of target processing
(Adam & Serences, 2021; Stilwell & Vecera, 2020; Chang
& Egeth, 2019).
GENERAL DISCUSSION
According to the signal suppression hypothesis, salient
stimuli generate a bottom–up salience signal that automat-
ically competes for attentional allocation, but this salience
signal can be proactively suppressed to prevent attentional
capture. Previous evidence in support of the signal
suppression hypothesis (e.g., Gaspelin & Luck, 2018c;
Gaspelin et al., 2015, 2017) has been challenged on the
Stilwell, Egeth, and Gaspelin
799
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
grounds that the small set sizes used in these studies may
have yielded color singletons that were insufficiently
salient to capture attention (Theeuwes in Luck et al.,
2021; Wang & Theeuwes, 2020). Although there has been
some evidence refuting such an interpretation (Chang
et al., 2021; Stilwell & Gaspelin, 2021), further evidence
is needed to definitively determine whether highly salient
distractors can be suppressed. The current study therefore
assessed whether highly salient color singletons elicit
electrophysiological indices of attentional capture (the
N2pc component) or attentional suppression (the PD
component).
Experiment 1 used an additional singleton paradigm
similar to Gaspelin and Luck (2018c), which previously
produced electrophysiological evidence of distractor sup-
pression, but modified the task to make the singleton dis-
tractor more salient by increasing the set size. This higher
set size should increase the relative contrast of the single-
ton distractor with other search items. The results indi-
cated that the singleton distractor was suppressed and
did not capture attention. Manual RTs were, if anything,
faster when the singleton distractor was present than
when it was absent: a singleton-presence benefit. The
ERP data also indicated that singleton distractors were
suppressed. First, lateralized singleton distractors elicited
a PD component, indicative of proactive distractor sup-
pression (Drisdelle & Eimer, 2021; van Moorselaar et al.,
2021; Feldmann-Wüstefeld et al., 2020; Gaspelin & Luck,
2018c; Gaspar & McDonald, 2014; McDonald, Green,
Jannati, & Di Lollo, 2013; Sawaki, Geng, & Luck, 2012;
Hickey et al., 2009). Second, lateralized targets elicited
N2pc components that did not differ as a function of
singleton distractor presence. If the singleton distractor
mandatorily captured attention, this should have dis-
rupted target processing and delayed and/or reduced
the magnitude of the N2pc component. Altogether, the
results suggest that the singleton distractor was sup-
pressed and did not interfere with search for the target,
consistent with the signal suppression hypothesis.
Experiment 2 provided a more stringent test of whether
increasing salience of the singleton distractor would
lead to attentional capture. In Experiment 1, the set size
was increased—compared to prior studies that used
four-item displays (e.g., Gaspelin & Luck, 2018c)—by
surrounding the central search array with a ring of dummy
shapes to boost the low-level feature contrast between
the singleton and nonsingleton items (i.e., distractor–
distractor dissimilarity; see the work of Duncan &
Humphreys, 1989). However, because of the outer ring’s
task-irrelevance, visual processing may have been
restricted to the inner ring of shapes. Experiment 2 there-
fore also increased the number of task-relevant items in
the inner ring (from four to eight items). The results
largely replicated those of Experiment 1. Lateralized
salient distractors elicited a PD component, indicating sup-
pression. Lateralized targets elicited N2pc components
that were unaffected by the presence of a singleton
distractor. These results suggest that the singleton was
suppressed to prevent attentional capture, which is incon-
sistent with a stimulus-driven account but consistent with
a signal suppression account.
The current results fit nicely with prior studies demon-
strating suppression of salient singleton distractors.
Singleton distractors did not evoke an N2pc component
but instead evoked PD components, consistent with the
previous literature on distractor suppression (Drisdelle
& Eimer, 2021; van Moorselaar et al., 2021; Feldmann-
Wüstefeld et al., 2020; van Moorselaar & Slagter, 2019;
Gaspelin & Luck, 2018c; Gaspar & McDonald, 2014;
McDonald et al., 2013; Sawaki et al., 2012; Hickey et al.,
2009). Furthermore, both the current ERP results and
the results of studies using the capture-probe paradigm
(Chang & Egeth, 2019, 2021; Stilwell & Gaspelin, 2021;
Gaspelin et al., 2015) converge to suggest that salient
singleton distractors are proactively suppressed. More-
over, the current study adds to a growing body of literature
aimed at uncovering the nature of saliency and its role in
models of attentional capture (Chang et al., 2021; Luck
et al., 2021; Egeth et al., 2010).
It is interesting to note that the mean amplitude of the
singleton-elicited PD component was unchanged by the
set size manipulations in the current study. The mean
amplitude of the PD component was equivalent at the
set size 16 of Experiment 1 (+0.4 μV) and the set size 20
of Experiment 2 (+0.4 μV). This may be because both
experiments used relatively large set sizes that maximized
the salience of the color singleton. We therefore also com-
pared the magnitude of the PD component to a previous
study (e.g., Gaspelin & Luck, 2018c; Experiment 1), which
was nearly identical to the current study but used a much
smaller set size of four items. The mean amplitude of
the singleton-elicited PD component was quite similar
(+0.5 μV) to the current experiments. At face value, this
suggests that increasing the salience of the singleton does
not necessarily impact suppression. However, future
studies may be needed to more directly manipulate the
salience of the singleton distractor systematically and test
whether suppression is modulated by degrees of salience.
In both experiments, we provide independent support
for the notion that the singleton distractor was indeed
salient using a computational model of salience (Chang
et al., 2021; Kotseruba et al., 2020; Hou et al., 2012). The
GSI (GSI = 0.81 in Experiment 1 and GSI = 0.82 in
Experiment 2) and the singleton win rate (100% in both
Experiments 1 and 2) indicated that the singleton had a
high salience score in all of the images. We additionally
compared the salience scores to displays of set size 4, sim-
ilar to Gaspelin and Luck (2018c). The results indicated
that the singleton was actually more salient in the current
experiments than in past experiments using a set size of
four (GSI = 0.47). Together, these analyses lend further
support to the claim that these singleton distractors were
highly salient in the current study and were more salient
than previous studies of distractor suppression (e.g.,
800
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Gaspelin & luck, 2018c). A shortcoming of this approach
is that it depends on the validity of the computational
model of salience (see the work of Jeck, Qin, Egeth, &
Niebur, 2019). Therefore, future research may be needed
to establish new metrics to measure bottom–up saliency
based upon human performance and/or electrophysi-
ological measures.
The increased the number of task-relevant items in
Experiment 2 allowed us to test an alternative account of
the PD component: the idiosyncratic search hypothesis
(Kerzel & Burra, 2020). According to this account, the
low set sizes used in previous studies cause the locations
on the horizontal midline to be searched first and the PD
component does not reflect suppression of the salient
item, but rather an N2pc enhancement of the nonsingle-
ton distractor on the horizontal midline. However, this
account seems unlikely for a few reasons. First, Experi-
ment 2 found that the singleton distractor elicited a reli-
able PD component even when it was not on the horizontal
midline. Second, perhaps more directly, Drisdelle and
Eimer (2021) used a similar design to the current study
but compared conditions where the target could appear
at horizontal locations to conditions where the target
could only appear at vertical locations, which should dis-
courage a horizontal search bias. They still found that the
PD components elicited by the singleton distractors were
the same across conditions. Last, the idiosyncratic search
hypothesis fails to account for the data from higher set
size displays where the incentive to attend to items on
the horizontal midline should be reduced (e.g., Gaspar
& McDonald, 2014). Altogether, there might be an atten-
tional bias to search items on the horizontal midline at low
set sizes, but the evidence that this could unambiguously
account for the PD component seems relatively weak.
It has been recently claimed that displays of heteroge-
neous shapes—as used in the current study—may render
the target stimulus nonsalient. As a result, participants may
stop using the attentional priority map to locate the target
and instead adopt an approach of randomly searching
items (clump scanning; Liesefeld, Liesefeld, & Müller,
2021a, 2021b). This, in turn, causes items with high atten-
tional priority because of bottom–up salience to fail to cap-
ture attention. Although interesting, there are some major
shortcomings of this model. First, the target is enhanced
above baseline levels in heterogeneous displays as indi-
cated by measures of probe report (Stilwell & Gaspelin,
2021; Gaspelin et al., 2015) and eye movements (Gaspelin
et al., 2017, 2019). If participants were not using the prior-
ity map, these target enhancement effects should not
occur: Search should be effectively “unguided” and there-
fore random. Second, this account cannot explain how
attentional allocation is guided away from color singletons.
If participants were not using a priority map, then atten-
tion should behave randomly and would not be biased
toward or away from any kind of stimulus feature. Ulti-
mately, the clump scanning hypothesis needs to be tested
more directly, but without a direct measure of the
attentional priority map and whether participants are
using it, this account will be difficult to falsify.
An interesting finding of the current study and prior
investigations is the late negativity appearing approxi-
mately 275–375 msec post stimulus on trials containing a
lateralized singleton distractor. This post-PD negativity
seems inconsistent: Some studies have found it (Drisdelle
& Eimer, 2021; Feldmann-Wüstefeld et al., 2020; Gaspelin
& Luck, 2018c, Experiment 3; Kerzel & Burra, 2020)
whereas others have not (Gaspelin & Luck, 2018c, Exper-
iment 1; Gaspar & McDonald, 2014; Sawaki & Luck,
2010). In this study, it occurred in Experiment 1 but not
in Experiment 2. It is currently unknown exactly what this
lateralized negativity reflects. Some have suggested that
the lateralized singleton distractor is suppressed on the
majority of trials, but on the trials when the singleton
captures attention, the waveforms blend together produc-
ing a later negativity (Gaspelin & Luck, 2018c; Gaspar &
McDonald, 2014). Another possibility is that the late neg-
ativity reflects a second PD component to the nonsingleton
distractor opposite the singleton distractor (Drisdelle &
Eimer, 2021). The current study does not offer definitive
evidence for either explanation; therefore, future studies
will likely be needed to test more directly what this ERP
component reflects.
The signal suppression hypothesis claims the singleton
distractor is proactively suppressed to prevent attentional
capture. It is important to note that the terminology “pro-
active suppression” could indicate one of two mecha-
nisms. Proactive suppression could mean that suppression
is implemented before any stimulus is presented, effec-
tively suppressing a specific location or feature before
the presentation of the search array. For example, Eimer
and Grubert (2014) demonstrated that feature-based
attentional templates are activated in anticipation of the
search array. Alternatively, proactive suppression could
mean that salient stimuli are suppressed after the stimuli
onset, but before the initial shift of covert attention. This
study was not designed to distinguish between these two
forms of proactive suppression, which would require
some measurement of the attentional template before
the search array appears. Therefore, further studies will
be needed to distinguish between these models.
It is important to highlight that the current study does
contrast with forms of reactive inhibition (e.g., Moher &
Egeth, 2012) whereby distractors must first be attended
before they can be ignored (e.g., the “search-and-destroy
model) or rapidly disengaged from following attentional
capture (e.g., Theeuwes, 2010). If the singleton was
attended before it was inhibited, this should have resulted
in some indication of attentional allocation, such as an
N2pc component, before suppression was observed. As
demonstrated by Weaver and colleagues (2017), covert
suppression can function before and independent of overt
behavioral measures such as overt eye movements. The
current study isolated covert shifts of attention by prevent-
ing overt attentional shifts (i.e., requiring participants to
Stilwell, Egeth, and Gaspelin
801
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
maintain central fixation) and found no evidence that the
salient distractors were covertly attended. These results,
and others, suggest that reactive inhibition is not the sole
mechanism for suppression of salient distractors. Impor-
tantly, it should be highlighted that reactive and proactive
inhibition are not mutually exclusive mechanisms (for a
review see the work of Geng, 2014).
In summary, the current study clearly demonstrates that
highly salient color singleton distractors are proactively
suppressed, as indexed by the PD component. Further-
more, the current study provides converging evidence in
support of the signal suppression hypothesis, helping
steer the field toward a potential resolution to the
decades-long debate on attentional capture, and high-
lights the importance of addressing physical salience in
models of attentional control.
Acknowledgments
All data and stimulus presentation programs in this article are
available on the Open Science Framework at https://osf.io
/9xnsz/.
Reprint requests should be sent to Brad T. Stilwell, Department
of Psychology, State University of New York at Binghamton, PO
Box 6000, Binghamton, NY 13902–6000, or via e-mail:
stilwell@binghamton.edu.
Funding Information
This project was supported by the National Science Foun-
dation (https://dx.doi.org/10.13039/100000001), grant
number: BCS-2045624 to N.G.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender identi-
fication of first author/last author) publishing in the Journal
of Cognitive Neuroscience ( JoCN) during this period were
M(an)/M = .407, W(oman)/M = .32, M/W = .115, and
W/W = .159, the comparable proportions for the articles
that these authorship teams cited were M/M = .549,
W/M = .257, M/W = .109, and W/W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance.
REFERENCES
Adam, K. C. S., Patel, T., Rangan, N., & Serences, J. T. (2021).
Classic visual search effects in an additional singleton task: An
open dataset. Journal of Cognition, 4, 34. https://doi.org/10
.5334/joc.182, PubMed: 34396037
Adam, K. C. S., & Serences, J. T. (2021). History modulates early
sensory processing of salient distractors. Journal of
Neuroscience, 41, 8007–8022. https://doi.org/10.1523
/JNEUROSCI.3099-20.2021, PubMed: 34330776
American Electroencephalographic Society. (1994). American
Electroencephalographic Society guidelines in
electroencephalography, evoked potentials, and
polysomnography. Journal of Clinical Neurophysiology,
11, 1–147.
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven
attentional capture. Perception & Psychophysics, 55,
485–496. https://doi.org/10.3758/BF03205306, PubMed:
8008550
Barras, C., & Kerzel, D. (2016). Active suppression of
salient-but-irrelevant stimuli does not underlie resistance to
visual interference. Biological Psychology, 121, 74–83.
https://doi.org/10.1016/j.biopsycho.2016.10.004, PubMed:
27756581
Berggren, N., & Eimer, M. (2020). Spatial filtering restricts the
attentional window during both singleton and feature-based
visual search. Attention, Perception & Psychophysics, 82,
2360–2378. https://doi.org/10.3758/s13414-020-01977-5,
PubMed: 31993978
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163
/156856897X00357, PubMed: 9176952
Chang, S., & Egeth, H. E. (2019). Enhancement and suppression
flexibly guide attention. Psychological Science, 30,
1724–1732. https://doi.org/10.1177/0956797619878813,
PubMed: 31693453
Chang, S., & Egeth, H. E. (2021). Can salient stimuli really be
suppressed? Attention, Perception, & Psychophysics, 83,
260–269. https://doi.org/10.3758/s13414-020-02207-8,
PubMed: 33241528
Chang, S., Niebur, E., & Egeth, H. (2021). Standing out in a
small crowd: The role of display size in attracting attention.
Visual Cognition, 29, 587–591. https://doi.org/10.1080
/13506285.2021.1918810, PubMed: 34707459
Cosman, J. D., Lowe, K. A., Woodman, G. F., & Schall, J. D.
(2018). Prefrontal control of visual distraction. Current
Biology, 28, 414–420. https://doi.org/10.1016/j.cub.2017.12
.023, PubMed: 29358071
Cunningham, C. A., & Egeth, H. E. (2016). Taming the white
bear: Initial costs and eventual benefits of distractor
inhibition. Psychological Science, 27, 476–485. https://doi
.org/10.1177/0956797615626564, PubMed: 26893292
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
Donohue, S. E., Schoenfeld, M. A., & Hopf, J.-M. (2020). Parallel
fast and slow recurrent cortical processing mediates target
and distractor selection in visual search. Communications
Biology, 3, 689. https://doi.org/10.1038/s42003-020-01423-0,
PubMed: 33214640
Drisdelle, B. L., & Eimer, M. (2021). PD components and
distractor inhibition in visual search: New evidence for the
signal suppression hypothesis. Psychophysiology, 58, e13878.
https://doi.org/10.1111/psyp.13878, PubMed: 34110022
Duncan, J., & Humphreys, G. W. (1989). Visual search and
stimulus similarity. Psychological Review, 96, 433–458.
https://doi.org/10.1037/0033-295X.96.3.433, PubMed:
2756067
Egeth, H. E., Leonard, C. J., & Leber, A. B. (2010). Why salience
is not enough: Reflections on top–down selection in vision.
Acta Psychologica, 135, 130–132. https://doi.org/10.1016/j
.actpsy.2010.05.012, PubMed: 20580341
Eimer, M. (2014). The neural basis of attentional
control in visual search. Trends in Cognitive Sciences,
802
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
18, 526–535. https://doi.org/10.1016/j.tics.2014.05.005,
PubMed: 24930047
Eimer, M., & Grubert, A. (2014). The gradual emergence of
spatially selective target processing in visual search: From
feature-specific to object-based attentional control. Journal
of Experimental Psychology: Human Perception and
Performance, 40, 1819–1831. https://doi.org/10.1037
/a0037387, PubMed: 24999612
Eimer, M., & Kiss, M. (2008). Involuntary attentional capture is
determined by task set: Evidence from event-related brain
potentials. Journal of Cognitive Neuroscience, 20,
1423–1433. https://doi.org/10.1162/jocn.2008.20099,
PubMed: 18303979
Feldmann-Wüstefeld, T., Busch, N. A., & Schubö, A. (2020).
Failed suppression of salient stimuli precedes behavioral
errors. Journal of Cognitive Neuroscience, 32, 367–377.
https://doi.org/10.1162/jocn_a_01502, PubMed: 31702429
Feldmann-Wüstefeld, T., Weinberger, M., & Awh, E. (2021).
Spatially guided distractor suppression during visual search.
Journal of Neuroscience, 41, 3180–3191. https://doi.org/10
.1523/JNEUROSCI.2418-20.2021, PubMed: 33653697
Folk, C. L., Leber, A. B., & Egeth, H. E. (2002). Made you blink!
Contingent attentional capture produces a spatial blink.
Perception & Psychophysics, 64, 741–753. https://doi.org/10
.3758/BF03194741, PubMed: 12201333
Folk, C. L., & Remington, R. (2010). A critical evaluation of the
disengagement hypothesis. Acta Psychologica, 135, 103–105.
https://doi.org/10.1016/j.actpsy.2010.04.012, PubMed:
20510849
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992).
Involuntary covert orienting is contingent on attentional
control settings. Journal of Experimental Psychology:
Human Perception and Performance, 18, 1030–1044. https://
doi.org/10.1037/0096-1523.18.4.1030, PubMed: 1431742
Gaspar, J. M., Christie, G. J., Prime, D. J., Jolicœur, P., &
McDonald, J. J. (2016). Inability to suppress salient distractors
predicts low visual working memory capacity. Proceedings of
the National Academy of Sciences, U.S.A., 113, 3693–3698.
https://doi.org/10.1073/pnas.1523471113, PubMed: 26903654
Gaspar, J. M., & McDonald, J. J. (2014). Suppression of salient
objects prevents distraction in visual search. Journal of
Neuroscience, 34, 5658–5666. https://doi.org/10.1523
/JNEUROSCI.4161-13.2014, PubMed: 24741056
Gaspelin, N., Gaspar, J. M., & Luck, S. J. (2019). Oculomotor
inhibition of salient distractors: Voluntary inhibition cannot
override selection history. Visual Cognition, 27, 227–246.
https://doi.org/10.1080/13506285.2019.1600090, PubMed:
31745389
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2015). Direct
evidence for active suppression of salient-but-irrelevant
sensory inputs. Psychological Science, 26, 1740–1750. https://
doi.org/10.1177/0956797615597913, PubMed: 26420441
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2017). Suppression
of overt attentional capture by salient-but-irrelevant color
singletons. Attention, Perception, & Psychophysics, 79,
45–62. https://doi.org/10.3758/s13414-016-1209-1, PubMed:
27804032
distractors. Journal of Cognitive Neuroscience, 30, 1265–1280.
https://doi.org/10.1162/jocn_a_01279, PubMed: 29762104
Gaspelin, N., & Luck, S. J. (2019). Inhibition as a potential
resolution to the attentional capture debate. Current
Opinion in Psychology, 29, 12–18. https://doi.org/10.1016/j
.copsyc.2018.10.013, PubMed: 30415087
Geng, J. J. (2014). Attentional mechanisms of distractor
suppression. Current Directions in Psychological Science,
23, 147–153. https://doi.org/10.1177/0963721414525780
Henare, D. T., Kadel, H., & Schubö, A. (2020). Voluntary control
of task selection does not eliminate the impact of selection
history on attention. Journal of Cognitive Neuroscience, 32,
2159–2177. https://doi.org/10.1162/jocn_a_01609, PubMed:
32662724
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009).
Electrophysiological indices of target and distractor
processing in visual search. Journal of Cognitive
Neuroscience, 21, 760–775. https://doi.org/10.1162/jocn.2009
.21039, PubMed: 18564048
Hickey, C., McDonald, J. J., & Theeuwes, J. (2006).
Electrophysiological evidence of the capture of visual
attention. Journal of Cognitive Neuroscience, 18,
604–613. https://doi.org/10.1162/jocn.2006.18.4.604,
PubMed: 16768363
Hillyard, S. A., Vogel, E. K., & Luck, S. J. (1998). Sensory gain
control (amplification) as a mechanism of selective attention:
Electrophysiological and neuroimaging evidence.
Philosophical Transactions of the Royal Society of London,
Series B: Biological Sciences, 353, 1257–1270. https://doi.org
/10.1098/rstb.1998.0281, PubMed: 9770220
Hopf, J. M., Luck, S. J., Boelmans, K., Schoenfeld, M. A.,
Boehler, C. N., Rieger, J., et al. (2006). The neural site of
attention matches the spatial scale of perception. Journal
of Neuroscience, 26, 3532–3540. https://doi.org/10.1523
/JNEUROSCI.4510-05.2006, PubMed: 16571761
Hou, X., Harel, J., & Koch, C. (2012). Image signature:
Highlighting sparse salient regions. IEEE Transactions on
Pattern Analysis and Machine Intelligence, 34, 194–201.
https://doi.org/10.1109/TPAMI.2011.146, PubMed: 21788665
Jannati, A., Gaspar, J. M., & McDonald, J. J. (2013). Tracking
target and distractor processing in fixed-feature visual search:
Evidence from human electrophysiology. Journal of
Experimental Psychology: Human Perception and
Performance, 39, 1713–1730. https://doi.org/10.1037
/a0032251, PubMed: 23527999
Jeck, D. M., Qin, M., Egeth, H., & Niebur, E. (2019). Unique
objects attract attention even when faint. Vision Research,
160, 60–71. https://doi.org/10.1016/j.visres.2019.04.004,
PubMed: 31047908
Kerzel, D., & Burra, N. (2020). Capture by context elements, not
attentional suppression of distractors, explains the PD with
small search displays. Journal of Cognitive Neuroscience, 32,
1170–1183. https://doi.org/10.1162/jocn_a_01535, PubMed:
31967520
Kotseruba, I., Wloka, C., Rasouli, A., & Tsotsos, J. K. (2020). Do
saliency models detect odd-one-out targets? New datasets
and evaluations. ArXiv Preprint ArXiv:2005.06583.
Gaspelin, N., & Luck, S. J. (2018a). Distinguishing among
Lakens, D. (2013). Calculating and reporting effect sizes to
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
potential mechanisms of singleton suppression. Journal
of Experimental Psychology: Human Perception and
Performance, 44, 626–644. https://doi.org/10.1037
/xhp0000484, PubMed: 29035072
Gaspelin, N., & Luck, S. J. (2018b). The role of inhibition in
avoiding distraction by salient stimuli. Trends in Cognitive
Sciences, 22, 79–92. https://doi.org/10.1016/j.tics.2017.11.001,
PubMed: 29191511
Gaspelin, N., & Luck, S. J. (2018c). Combined electrophysiological
and behavioral evidence for the suppression of salient
facilitate cumulative science: A practical primer for t-tests and
ANOVAs. Frontiers in Psychology, 4, 863. https://doi.org/10
.3389/fpsyg.2013.00863, PubMed: 24324449
Lamy, D., & Egeth, H. E. (2003). Attentional capture in
singleton-detection and feature-search modes. Journal of
Experimental Psychology: Human Perception and
Performance, 29, 1003–1020. https://doi.org/10.1037/0096
-1523.29.5.1003, PubMed: 14585019
Leber, A. B., & Egeth, H. E. (2006). It’s under control:
Top–down search strategies can override attentional capture.
Stilwell, Egeth, and Gaspelin
803
Psychonomic Bulletin & Review, 13, 132–138. https://doi.org
/10.3758/BF03193824, PubMed: 16724780
Lien, M.-C., Ruthruff, E., Goodin, Z., & Remington, R. W. (2008).
Contingent attentional capture by top–down control settings:
Converging evidence from event-related potentials. Journal
of Experimental Psychology: Human Perception and
Performance, 34, 509–530. https://doi.org/10.1037/0096-1523
.34.3.509, PubMed: 18505320
Lien, M.-C., Ruthruff, E., & Johnston, J. C. (2010). Attentional
capture with rapidly changing attentional control settings.
Journal of Experimental Psychology: Human Perception
and Performance, 36, 1–16. https://doi.org/10.1037
/a0015875, PubMed: 20121291
Liesefeld, H. R., Liesefeld, A. M., & Müller, H. J. (2021a).
Preparatory control against distraction is not feature-based.
Cerebral Cortex, bhab341. https://doi.org/10.1093/cercor
/bhab341, PubMed: 34585718
Liesefeld, H. R., Liesefeld, A. M., & Müller, H. J. (2021b).
Attentional capture: An ameliorable side-effect of searching
for salient targets. Visual Cognition, 29, 600–603. https://doi
.org/10.1080/13506285.2021.1925798
Lins, O. G., Picton, T. W., Berg, P., & Scherg, M. (1993). Ocular
artifacts in EEG and event-related potentials. I: Scalp
topography. Brain Topography, 6, 51–63. https://doi.org/10
.1007/BF01234127, PubMed: 8260327
Lopez-Calderon, J., & Luck, S. J. (2014). ERPLAB: An
open-source toolbox for the analysis of event-related
potentials. Frontiers in Human Neuroscience, 8, 213.
https://doi.org/10.3389/fnhum.2014.00213, PubMed:
24782741
Luck, S. J. (2012). Electrophysiological correlates of the
focusing of attention within complex visual scenes: N2pc and
related ERP components. In S. J. Luck & E. S. Kappenman
(Eds.), The Oxford handbook of event-related potential
components (pp. 329–360). New York: Oxford University
Press.
Luck, S. J., Chelazzi, L., Hillyard, S. A., & Desimone, R. (1997).
Neural mechanisms of spatial selective attention in areas V1,
V2, and V4 of macaque visual cortex. Journal of
Neurophysiology, 77, 24–42. https://doi.org/10.1152/jn.1997
.77.1.24, PubMed: 9120566
Luck, S. J., & Gaspelin, N. (2017). How to get statistically
significant effects in any ERP experiment (and why you
shouldn’t). Psychophysiology, 54, 146–157. https://doi.org
/10.1111/psyp.12639, PubMed: 28000253
Luck, S. J., Gaspelin, N., Folk, C. L., Remington, R. W., &
Theeuwes, J. (2021). Progress toward resolving the attentional
capture debate. Visual Cognition, 29, 1–21. https://doi.org/10
.1080/13506285.2020.1848949, PubMed: 33574729
Luck, S. J., & Hillyard, S. A. (1994). Electrophysiological
correlates of feature analysis during visual search.
Psychophysiology, 31, 291–308. https://doi.org/10.1111/j.1469
-8986.1994.tb02218.x, PubMed: 8008793
McDonald, J. J., Green, J. J., Jannati, A., & Di Lollo, V. (2013). On
the electrophysiological evidence for the capture of visual
attention. Journal of Experimental Psychology: Human
Perception and Performance, 39, 849–860. https://doi.org/10
.1037/a0030510, PubMed: 23163789
Moher, J., Abrams, J., Egeth, H. E., Yantis, S., & Stuphorn, V.
(2011). Trial-by-trial adjustments of top–down set modulate
oculomotor capture. Psychonomic Bulletin & Review, 18,
897–903. https://doi.org/10.3758/s13423-011-0118-5, PubMed:
21691926
Moher, J., & Egeth, H. E. (2012). The ignoring paradox: Cueing
distractor features leads first to selection, then to inhibition
of to-be-ignored items. Attention, Perception, &
Psychophysics, 74, 1590–1605. https://doi.org/10.3758/s13414
-012-0358-0, PubMed: 22893004
Mordkoff, J. T. (2019). A simple method for removing bias from
a popular measure of standardized effect size: Adjusted
partial eta squared. Advances in Methods and Practices in
Psychological Science, 2, 228–232. https://doi.org/10.1177
/2515245919855053
Pashler, H. (1988). Cross-dimensional interaction and
texture segregation. Perception & Psychophysics, 43,
307–318. https://doi.org/10.3758/BF03208800, PubMed:
3362658
Ruthruff, E., & Gaspelin, N. (2018). Immunity to attentional
capture at ignored locations. Attention, Perception, &
Psychophysics, 80, 325–336. https://doi.org/10.3758/s13414
-017-1440-4, PubMed: 29116615
Sawaki, R., Geng, J. J., & Luck, S. J. (2012). A common neural
mechanism for preventing and terminating the allocation
of attention. Journal of Neuroscience, 32, 10725–10736.
https://doi.org/10.1523/JNEUROSCI.1864-12.2012, PubMed:
22855820
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression of
attention by salient singletons: Electrophysiological evidence
for an automatic attend-to-me signal. Attention, Perception,
& Psychophysics, 72, 1455–1470. https://doi.org/10.3758/APP
.72.6.1455, PubMed: 20675793
Stilwell, B. T., Bahle, B., & Vecera, S. P. (2019). Feature-based
statistical regularities of distractors modulate attentional
capture. Journal of Experimental Psychology: Human
Perception and Performance, 45, 419–433. https://doi.org/10
.1037/xhp0000613, PubMed: 30802131
Stilwell, B. T., & Gaspelin, N. (2021). Attentional suppression
of highly salient color singletons. Journal of Experimental
Psychology: Human Perception and Performance, 47,
1313–1328. https://doi.org/10.1037/xhp0000948, PubMed:
34766817
Stilwell, B. T., & Vecera, S. P. (2020). Learned distractor
rejection in the face of strong target guidance. Journal of
Experimental Psychology: Human Perception and
Performance, 46, 926–941. https://doi.org/10.1037
/xhp0000757, PubMed: 32391708
Theeuwes, J. (1991). Exogenous and endogenous control of
attention: The effect of visual onsets and offsets. Perception
& Psychophysics, 49, 83–90. https://doi.org/10.3758
/BF03211619, PubMed: 2011456
Theeuwes, J. (1992). Perceptual selectivity for color and form.
Perception & Psychophysics, 51, 599–606. https://doi.org/10
.3758/BF03211656, PubMed: 1620571
Theeuwes, J. (2004). Top–down search strategies cannot
override attentional capture. Psychonomic Bulletin & Review,
11, 65–70. https://doi.org/10.3758/BF03206462, PubMed:
15116988
Theeuwes, J. (2010). Top–down and bottom–up control of
visual selection. Acta Psychologica, 135, 77–99. https://doi
.org/10.1016/j.actpsy.2010.02.006, PubMed: 20507828
van Moorselaar, D., Daneshtalab, N., & Slagter, H. A. (2021).
Neural mechanisms underlying distractor inhibition on the
basis of feature and/or spatial expectations. Cortex, 137,
232–250. https://doi.org/10.1016/j.cortex.2021.01.010,
PubMed: 33640854
van Moorselaar, D., Lampers, E., Cordesius, E., & Slagter, H. A.
(2020). Neural mechanisms underlying expectation-
dependent inhibition of distracting information. eLife, 9,
e61048. https://doi.org/10.7554/eLife.61048, PubMed:
33320084
van Moorselaar, D., & Slagter, H. A. (2019). Learning what is
irrelevant or relevant: Expectations facilitate distractor
inhibition and target facilitation through distinct neural
mechanisms. Journal of Neuroscience, 39, 6953–6967.
https://doi.org/10.1523/JNEUROSCI.0593-19.2019, PubMed:
31270162
804
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Vatterott, D. B., Mozer, M. C., & Vecera, S. P. (2018). Rejecting
salient distractors: Generalization from experience. Attention,
Perception, & Psychophysics, 80, 485–499. https://doi.org/10
.3758/s13414-017-1465-8, PubMed: 29230673
Vatterott, D. B., & Vecera, S. P. (2012). Experience-dependent
attentional tuning of distractor rejection. Psychonomic
Bulletin & Review, 19, 871–878. https://doi.org/10.3758
/s13423-012-0280-4, PubMed: 22696250
Wang, B., & Theeuwes, J. (2018a). Statistical regularities
modulate attentional capture. Journal of Experimental
Psychology: Human Perception and Performance, 44,
13–17. https://doi.org/10.1037/xhp0000472, PubMed:
29309194
Wang, B., & Theeuwes, J. (2018b). How to inhibit a distractor
location? Statistical learning versus active, top–down
suppression. Attention, Perception, & Psychophysics, 80,
860–870. https://doi.org/10.3758/s13414-018-1493-z, PubMed:
29476331
1060–1092. https://doi.org/10.3758/s13423-020-01859-9,
PubMed: 33547630
Wolfe, J. M., & Horowitz, T. S. (2017). Five factors that guide
attention in visual search. Nature Human Behaviour, 1, 1–8.
https://doi.org/10.1038/s41562-017-0058
Won, B.-Y., Forloines, M., Zhou, Z., & Geng, J. J. (2020). Changes
in visual cortical processing attenuate singleton distraction
during visual search. Cortex, 132, 309–321. https://doi.org/10
.1016/j.cortex.2020.08.025, PubMed: 33010740
Won, B.-Y., & Geng, J. J. (2020). Passive exposure attenuates
distraction during visual search. Journal of Experimental
Psychology: General, 149, 1987–1995. https://doi.org/10.1037
/xge0000760, PubMed: 32250138
Won, B.-Y., Kosoyan, M., & Geng, J. J. (2019). Evidence for
second-order singleton suppression based on probabilistic
expectations. Journal of Experimental Psychology: Human
Perception and Performance, 45, 125–138. https://doi.org/10
.1037/xhp0000594, PubMed: 30596437
Wang, B., & Theeuwes, J. (2020). Salience determines attentional
Woodman, G. F., & Luck, S. J. (1999). Electrophysiological
orienting in visual selection. Journal of Experimental Psychology:
Human Perception and Performance, 46, 1051–1057. https://doi
.org/10.1037/xhp0000796, PubMed: 32757594
Weaver, M. D., van Zoest, W., & Hickey, C. (2017). A temporal
dependency account of attentional inhibition in oculomotor
control. Neuroimage, 147, 880–894. https://doi.org/10.1016/j
.neuroimage.2016.11.004, PubMed: 27836709
Whitney, D., & Levi, D. M. (2011). Visual crowding: A
fundamental limit on conscious perception and object
recognition. Trends in Cognitive Sciences, 15, 160–168.
https://doi.org/10.1016/j.tics.2011.02.005, PubMed: 21420894
Wolfe, J. M. (2021). Guided Search 6.0: An updated model
of visual search. Psychonomic Bulletin & Review, 28,
measurement of rapid shifts of attention during visual search.
Nature, 400, 867–869. https://doi.org/10.1038/23698,
PubMed: 10476964
Woodman, G. F., & Luck, S. J. (2003). Serial deployment of
attention during visual search. Journal of Experimental
Psychology: Human Perception and Performance, 29,
121–138. https://doi.org/10.1037/0096-1523.29.1.121,
PubMed: 12669752
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and
selective attention: Evidence from visual search. Journal of
Experimental Psychology: Human Perception and
Performance, 10, 601–621. https://doi.org/10.1037/0096-1523
.10.5.601, PubMed: 6238122
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
7
8
7
2
0
0
4
6
3
9
/
j
o
c
n
_
a
_
0
1
8
2
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Stilwell, Egeth, and Gaspelin
805