Effects of Multisession Prefrontal Transcranial Direct
Current Stimulation on Long-term Memory and
Working Memory in Older Adults
Jacky Au1, Rachel N. Smith-Peirce1, Elena Carbone2, Austin Moon3
Michelle Evans4, John Jonides4, and Susanne M. Jaeggi1
,
Abstracto
■ Transcranial direct current stimulation (tDCS) is a noninvasive
form of electrical brain stimulation popularly used to augment the
effects of working memory (W.M.) training. Although success has
been mixed, some studies report enhancements in WM perfor-
mance persisting days, semanas, or even months that are actually
more reminiscent of consolidation effects typically observed in
the long-term memory (LTM) domain, rather than WM improve-
ments per se. Although tDCS has been often reported to enhance
both WM and LTM, these effects have never been directly com-
pared within the same study. Sin embargo, given their considerable
neural and behavioral overlap, this is a timely comparison to make.
This study reports results from a multisession intervention in older
adults comparing active and sham tDCS over the left dorsolateral
pFC during training on both an n-back WM task and a word learn-
ing LTM task. We found strong and robust effects on LTM, pero
mixed effects on WM that only emerged for those with lower base-
line ability. En tono rimbombante, mediation analyses showed an indirect
effect of tDCS on WM that was mediated by improvements in con-
solidation. We conclude that tDCS over the left dorsolateral pFC
can be used as an effective intervention to foster long-term learning
and memory consolidation in aging, which can manifest in perfor-
mance improvements across multiple memory domains. ■
INTRODUCCIÓN
Transcranial direct current stimulation (tDCS) is a nonin-
vasive method of electrical brain stimulation that can
influence cognitive functioning via modulation of cortical
excitability. tDCS targeted over the dorsolateral pFC
(DLPFC) has been shown to affect a variety of memory
funciones (Huo et al., 2021; Buch et al., 2017; Mancuso,
Ilieva, hamilton, & Farah, 2016; Brasil-Neto, 2012), cual
can be a critical boon for older adults looking for methods
to mitigate age-related cognitive decline. Por ejemplo, nuestro
previous work, although in younger adults, demonstrated
sustained performance enhancements when administer-
ing DLPFC tDCS during an n-back working memory
( W.M.) task over the course of a week-long intervention
(Au et al., 2016), an effect that was replicated by an inde-
pendent group in a similar experiment (Ruf, Fallgatter, &
Plewnia, 2017). Other WM training studies have been
similarly successful as well, both in younger (Ke et al.,
2019; Richmond, Wolk, Chein, & Olson, 2014) así como
adultos mayores (Esteban & Berryhill, 2016; jones, Esteban,
Alam, Bikson, & Berryhill, 2015).
Despite these initial successes, a follow-up study by our
group failed to replicate our original training effects in
younger adults (Au et al., 2021), and a large randomized
controlled trial of 123 individuals also did not observe
1Universidad de California, Irvine, 2University of Padova, 3universidad-
sity of California, Riverside, 4University of Michigan
© 2022 Instituto de Tecnología de Massachusetts
significant improvements in older adults (Nilsson,
Lebedev, Rydström, & Lövdén, 2017). Además, meta-
analyses suggest that effects of tDCS on WM are small at
best (Colina, Fitzgerald, & Hoy, 2016; Mancuso et al., 2016)
or unreliable at worst (Horvath, Forte, & Carretero, 2015).
This is not to suggest that tDCS-enhanced WM is not a
worthwhile pursuit, but rather to highlight the challenges
in the field and the difficulty in understanding the specific
conditions that will most likely elicit an effect. A diferencia de,
tDCS effects on long-term memory (LTM) seem to be
more promising, with stronger and more robust meta-
analytic effects, especially among older adults (Huo
et al., 2021; Galli, Vadillo, Sirota, Feurra, & Medvedeva,
2019; Summers, Kang, & Cauraugh, 2016). Además,
there is accumulating evidence that tDCS can increase
long-term potentiation (LTP; Kronberg, Bridi, Abel, Bikson,
& Parra, 2017; Podda et al., 2016; Rohan, Carhuatanta,
McInturf, Miklasevich, & Jankord, 2015; Ranieri et al.,
2012) and LTP-like plasticity in the cortex (Frase et al.,
2021; Agboada, Mosayebi-Samani, kuo, & Nitsche, 2020;
Monte-Silva et al., 2013), which may facilitate the consoli-
dation and long-term retention of material learned during
stimulation. De hecho, even the WM training effects we orig-
inally reported (Au et al., 2016) displayed properties that
were actually more reminiscent of LTM consolidation than
WM per se, such as a spaced learning pattern in which gains
were greater over a weekend compared with consecutive
weekdays, as well as maintenance effects that persisted
Revista de neurociencia cognitiva 34:6, páginas. 1015–1037
https://doi.org/10.1162/jocn_a_01839
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
up to a year later even in the absence of further stimulation
(Katz et al., 2017). The main purpose of the current study,
por lo tanto, is to compare the relative effects of tDCS on both
WM and LTM during a combined memory intervention in
older adults who may be more susceptible to tDCS modu-
lation and to investigate the extent to which improved LTM
actually mediates any enhancements in WM performance.
In considering the effects of tDCS across memory
dominios, one important issue to recognize is that the pro-
cesses that underlie WM and LTM are not completely dis-
tinct and have a bidirectional relationship (Bergmann,
Rijpkema, Fernández, & Kessels, 2012; Cabeza, Dolcos,
graham, & Nyberg, 2002). WM processing relies on
retrieval from LTM stores to contextualize incoming infor-
formación, which in turn, with enough repetition or salience,
can be re-encoded back into LTM (Miyake & Shah, 1999;
van der Linden, 1998). Además, activity in the pFC dur-
ing a WM task has been shown to predict LTM formation
(Blumenfeld, 2006; Ranganath, cohen, & Brozinsky,
2005), even of memoranda separate from those used in
the WM task (Melrose et al., 2020). En cambio, actividad
in medial-temporal lobe structures such as the hippocam-
pus typically thought to subserve LTM (Squire, Rígido, &
clark, 2004) have also been implicated in the maintenance
of WM (Bergmann et al., 2012; Nichols, Kao, Verfaellie, &
Gabrieli, 2006; Cabeza et al., 2002). So, the two memory
domains work together cooperatively, which is also
reflected by their shared neural substrates (Nee & En Jonia,
2008, 2013; Hannula & Ranganath, 2008). This is further
underscored by findings of improvements in certain
aspects of LTM measures after WM training (Rudebeck,
Bor, Ormond, O’Reilly, & Sotavento, 2012; Richmond, Morrison,
Chein, & Olson, 2011; Buschkuehl et al., 2008), así como
recent studies that show how WM performance can exhibit
sleep-dependent enhancements reminiscent of LTM con-
solidation. Específicamente, several studies have documented
that training-related improvements on the n-back WM task
were only observed if the interval between sessions
included sleep or a nap, but not wake (Zinke, Noack, &
Born, 2018; Lau, Wong, Lau, Hui, & Tseng, 2015). Más-
más, these performance improvements were associated
with increased slow wave activity during sleep (Ferrarelli
et al., 2019; Sattari, Whitehurst, Ahmadi, & Mednick,
2019; Pugin et al., 2015), which is a critical factor in the
consolidation of perceptual and motor procedural skills
(Määttä et al., 2010; Crupi et al., 2009; Huber, Felice
Ghilardi, Massimini, & Tononi, 2004). De este modo, we hypothe-
size that the long-term retention of performance benefits
observed from the use of tDCS on WM tasks may arise
from similar mechanisms as those observed on LTM tasks.
These processes may relate to the consolidation of new
cognitive routines (Gathercole, Dunning, holmes, &
Norris, 2019), which may include declarative components
of WM training such as strategy learning or procedural
components such as developing stimulus–response map-
pings or the proficiency of updating between shifting rules
and goals (p.ej., Sali & Adecuado, 2020; Oberauer, 2009).
Given the overlapping neural architecture between the
two memory domains, it is perhaps not surprising that
tDCS delivered over the same left DLPFC area can enhance
both WM (reviewed in Mancuso et al., 2016) as well as LTM
(Huo et al., 2021; Galli et al., 2019) actuación. Sin embargo,
a nuestro conocimiento, a direct comparison between the two
memory domains has never been made within a single
estudiar. De este modo, the goal of the current study was to carry
out this comparison in the context of a five-session inten-
sive memory training intervention. Although we expected
improvements in both memory domains based on pre-
vious literature (p.ej., Perceval, Martín, Copland, Laine, &
Meinzer, 2020; Au et al., 2016; Sandrini et al., 2016;
jones, Esteban, et al., 2015; Javadi & cheng, 2013), nosotros
hypothesized stronger and more robust effects would
emerge within the LTM domain because of the facilitatory
effects of tDCS on memory consolidation (Au, Karsten,
Buschkuehl, & Jaeggi, 2017; Podda et al., 2016). Además,
if the long-term improvements in both memory domains
arise from similar consolidation mechanisms, si
directly or indirectly, then we hypothesized that the
strength of this consolidation, as measured by LTM
retention, might mediate at least part of the relationship
between tDCS and long-term WM performance.
Finalmente, we sought to build upon our previous work
that has shown baseline WM performance and spaced
training to be influential moderators of tDCS efficacy (katz
et al., 2017; Au et al., 2016). With respect to the former, nosotros
previously found that individuals with lower baseline
ability benefitted more from tDCS, a finding generally
corroborated by the literature (p.ej., Perceval et al., 2020;
McConathey et al., 2017; Looi et al., 2016; Tseng et al.,
2012), although evidence for a high-baseline advantage
also exists ( jones, Gözenman, & Berryhill, 2015; jones &
Berryhill, 2012). With respect to the spacing issue, we pre-
viously reported that spacing training sessions apart by a
few days was associated with greater training gains com-
pared with daily training, but only in the presence of tDCS
(Au et al., 2016). It is known that varying the intersession
interval between repeated bouts of tDCS can influence the
size and direction of effects, although most studies to date
have manipulated this interval on the order of minutes
rather than days (Goldsworthy, Pitcher, & Ridding, 2015;
Monte-Silva et al., 2013). With the higher-order learning
involved in cognitive training protocols, we previously
postulated that spacing tDCS sessions apart by days could
facilitate consolidation processes that occur during that
same time frame (Au et al., 2016, 2017). Sin embargo, our pre-
vious study found this spacing effect with WM training
gains, and it is important to replicate this phenomenon
with memoranda encoded into LTM before endorsing an
effect of spaced tDCS on consolidation.
De este modo, the overarching goals of the current study are
tres. Primero, we seek to establish the efficacy of tDCS on
improving WM and LTM in a sample of healthy older
adultos, both on trained and untrained measures. Segundo,
we seek to compare the effects of tDCS between LTM and
1016
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
WM and determine the degree to which LTM consolida-
tion mediates long-term improvements on n-back train-
En g. Finalmente, we explore the possible moderating influences
that baseline ability and intersession spacing interval have
on the efficacy of tDCS. In tackling these goals, nosotros
included pretest and posttest measures of WM and LTM
before and after the tDCS intervention, respectivamente, y
we also randomly assigned older adult participants
between the ages of 65 y 85 years to come into the
lab either daily or every other day. Although there is no
definitive evidence that tDCS is more or less effective in
older compared with younger populations, there is some
suggestion that lower-performing populations such as
clinical and older adult populations may have a higher
response rate to tDCS (Huo et al., 2021; Galli et al.,
2019; Dedoncker, Brunoni, Baeken, & Vanderhasselt,
2016; Hill et al., 2016). Por lo tanto, not only does testing
older adults potentially increase our chances of finding a
study effect, but it also affords us the opportunity to foster
memory skills in a population that is susceptible to cogni-
tive decline and is in great need of such interventions.
MÉTODOS
Participantes
In a joint collaboration between the University of California,
Irvine, and the University of Michigan, 55 neurologically and
psychologically healthy right-handed older adults (entre
ages of 65 y 85 años; edad media = 71.32 años, 73%
women) were recruited from the local communities before
the start of the COVID-19 global pandemic, which halted
our recruitment. De estos 55, one scored above threshold
on the Geriatric Depression Screener (Scogin, Rohen, &
Bailey, 2000), and two others dropped out during the
course of the study. Their data were excluded from all
analiza. In the end, our analytic sample comprised 24
individuals randomized to receive active tDCS and 28 ran-
domized to receive sham tDCS. Of the active tDCS group,
12 were randomized to train daily, y 12 were to train
every other day. Of the sham group, 13 were randomized
to train daily, y 15 were to train every other day. Rarely, a
participant who could not make a certain training session
was allowed to reschedule within 1 día, once during their
training period. This only affected three of our participants
(two sham/one active). All research procedures were
approved by the institutional review boards at both univer-
ciudades, and each participant provided informed consent.
General Procedure
In our between-subject design, participants were ran-
domly assigned to one of two stimulation conditions
(active or sham tDCS) and one of two spacing conditions
(daily or every-other-day training). In accordance with our
previous procedure (Au et al., 2016), participants were
not informed of the possibility of a sham condition. Todo
participants were told they were receiving active stimula-
tion and advised that the sensations associated with
stimulation were generally subtle with considerable
interindividual variability. Upon conclusion of study
procedures, participants were debriefed about the sham
procedure and asked to guess if they received active or
sham stimulation.
The intervention itself consisted of five sessions of WM
and LTM training, which occurred either within a calendar
week for the daily training group or within two calendar
weeks for the every-other-day group. Each training session
lasted approximately an hour, including setup and
cleanup, and started with word list learning, seguido por
n-back training. Stimulation was applied at the beginning
of word learning and spilled over into part of n-back train-
En g, lasting for a fixed duration of 25 min regardless of how
far along participants had progressed on the training. Este
was done to ensure a comparable stimulation duration
across all participants and also to replicate our previous
trabajar, which showed 25 min of stimulation to be an effec-
tive dosage (Au et al., 2016). Además, we aimed to
avoid overstimulation, which has been observed to
reverse the direction of tDCS effects (Monte-Silva et al.,
2013). After n-back training, participants received a brief
5-min break during which the tDCS electrodes were
removed. At the end of the session, participants were once
again asked to recall as many words as they could remem-
ber from the beginning of that day’s session. At the begin-
ning of the next session, before learning the new word list
for that day, participants were again asked to recall as many
words as they could remember, but this time cumulatively
from all previous sessions.
Pretests and posttests consisting of trained and
untrained outcome measures took place within a few days
before the first or after the last training sessions. A final
follow-up appointment was scheduled 3 months after
the last training session to repeat the outcome measures.
Because of the unexpected COVID-19 global pandemic
and the subsequent lockdown, the follow-up sessions
were not completed by all participants and the data
were not analyzed. Sin embargo, descriptive data for the
participants who did finish are provided in Appendix A
(Table A1). A schematic of the study design is provided
En figura 1.
tDCS
Stimulation was administered via the Oasis Pro tDCS
device by MIND Alive, Cª, usando 5 × 7 cm sponge elec-
trodes placed horizontally on the head. The anode was
placed over the left DLPFC (corresponding to position
F3 in the international 10–20 EEG system), and the cath-
ode was placed over the contralateral supraorbital area
(site Fp2). Electrode positions were identified using a
BraiNet 10/20 placement cap (bio-medical.com), cual
was individually fitted to a participant’s head based on
their head circumference. Stimulation lasted 25 min with
Au et al.
1017
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. (A) Overall study design. General study procedures are depicted. Lightning bolts represents time period during which participants receive
either active or sham stimulation. (B) Structure of a training session. A snapshot of the training intervention is portrayed, in which participants
are randomized to train either daily or every other day under active or sham stimulation. The intervention itself consisted of both a word learning
and n-back task under stimulation, followed by a delayed recall approximately 30 min later, and again at the beginning of the next session (either
24 o 48 hr later depending on the spacing condition) before repeating the training session all over again with a new set of words for the day. El
24–48 hr delayed recall is cumulative and consists of all words learned up to that point in training. Both delayed recalls are performed in the absence
of stimulation.
a current intensity of 2 mA, which ramped up and down for
the first and last 15 segundo de estimulación. Sham tDCS was set
up in the same way, except the current was shut off
unknown to participants between the 15-sec ramping
periods. The active and sham conditions were prepro-
grammed into the device using codes that only the first
author knew. De este modo, both the experimenters and partici-
pants were blind to the participants’ conditions.
use as new, unstudied words for the recognition task,
counterbalanced between posttest and follow-up.
A similar procedure was used to generate three lists of
60 words each for the metamemory outcome measure. A
different list was used at pretest, posttest, and follow-up.
Training Tasks
LTM
Word Generation Procedure
We used the same procedure to generate word lists for the
LTM training task, metamemory outcome measure, y
recognition outcome measure. We selected words from
a variety of established databases (Brysbaert, Warriner, &
Kuperman, 2014; Warriner, Kuperman, & Brysbaert, 2013;
Reilly & Kean, 2007; Stadthagen-Gonzalez & davis, 2006;
Bird, franklin, & Howard, 2001) to create separate word
lists that were matched on key lexical features such as
word length, familiarity, arousal and valence, concreto-
ness, and imageability. For the training task, 150 palabras
were selected and divided into five matched lists of 30.
Eighteen of these 30 words were kept for use during each
training session, whereas the remainder was retained for
The word-list learning task comprised a total of 90 palabras,
which were separated into five lists of 18 words each. Cada
list was presented during one of the five training sessions.
With the exception of the first training day, each subse-
quent session started with cumulative recall of all words
learned during previous sessions. tDCS was not adminis-
tered during cumulative recall. Following that, the tDCS
device was set up in accordance with the participant’s
group assignment, and a new word list for the day was then
visually presented on a computer screen at a rate of one
cada 3 segundo. Participants verbally recalled as many words
as they could remember immediately after the list while
a researcher recorded their answers. The list was then
repeated twice more, for a total of three rounds of
1018
Revista de neurociencia cognitiva
Volumen 34, Número 6
immediate recall. After a delay of approximately 30 mín.,
during which participants trained on the n-back task
(see below), participants were once more asked to ver-
bally recall as many words as possible.
The dependent variables of interest were the number of
words recalled after three rounds of immediate recall, después
a 30-min delay, and at cumulative recall at the beginning of
the next session. Throughout the remainder of the article,
these time points will be respectively referred to as imme-
diate word learning (es decir., the third immediate recall),
delayed recall, and cumulative recall.
At posttest and 3-month follow-up, a self-paced recogni-
tion test was administered in which participants were
given 30 old words learned during training and 30 nuevo
palabras. Each word was displayed on a computer screen
until participants made a response indicating whether
the current word was old or new.
W.M.
Participants trained on a tablet-based version of an n-back
tarea, identical to the training task used in the study of
Jaeggi et al. (2020), which used pictures of everyday objects
as stimuli and required indicating whether a presented pic-
ture was the same as that presented in trials previously.
Stimuli were presented in a moving window that lasted
para 1000 msec with an ISI of 2500 mseg. The difficulty level
continuously adapted to individual performance across
sessions. Participants needed to achieve accuracy scores
de 90% or greater to advance to the next round; puntuaciones de
70% or lower demoted them to the previous round. Cada
n-level comprised three rounds such that participants had
to demonstrate 90% or greater accuracy three times before
incrementing to the next n-level. This was done to encour-
age mastery of an n-back level before advancing to the next.
The only exception was the 1-back level, which only con-
tained one round. Each training session consisted of 10
rounds, and each round consisted of five target trials and
aproximadamente 20 nontarget trials.
A brief practice consisting of four rounds was given to
participants at pretest to explain instructions and familiar-
ize them with the upcoming training. At posttest and at
3-month follow-up, the training task was readministered
but started over at 1-back instead of continuing adaptively
from the last training day’s n-level. This was done to ensure
comparability across participants at posttest and 3-month
hacer un seguimiento. The dependent variable of interest for this task
was the average n-level achieved during each session.
Because of a combination of technical problems and exper-
imenter error, posttest data from eight participants (four
active/four sham) were not collected or were lost.
Incidental Memory
Embedded within the n-back WM task was an incidental
memory task in which participants were unexpectedly
asked to recall as many of the n-back stimuli as they could
at the end of training. These stimuli consisted of everyday
objects such as shoes, clocks, fruits, vegetables, animals,
Etcétera. Each session comprised a total of 16 unique
objects, which rotated each training day for a total of 80
unique objects by Session 5. Participants were given a max-
imum of 2 min to recall these objects, and this was done at
two time points—after their last training session as well as
at the beginning of posttest and 3-month follow-up.
Because of experimenter error, six participants (five
active/one sham) were not administered the incidental
memory test after their last training session and were
therefore not included in the analyses. The dependent var-
iable of interest was the total number of objects recalled
per time point.
Untrained Outcome Measures
Number n-Back
The number n-back task was an untrained variant of the
trained n-back task that used numbers instead of everyday
objects as stimuli. Numbers were presented visually at the
center of the screen at a rate of one every 3 segundo. The task
comprised one warm-up round of 1-back, seguido por
three rounds of 2-back and 3-back. Each round consisted
de 20 + n trials, and the dependent variable was the accu-
racy rate, or the proportion of hits minus the proportion of
false alarms for the 2- and 3-back rounds (Snodgrass &
Corwin, 1988).
Sternberg Item Recognition
In the item recognition task (Sternberg, 1969), participar-
pants were shown 4–10 uppercase letters equidistant from
a fixation cross located in the center of the computer
pantalla. Set sizes varied between trials in a pseudorandom
order that was unpredictable for the participant. Presenta-
tion time varied between set sizes. For set size 4, estímulos
were displayed for 1300 mseg, and this presentation time
incremented up by 325 msec for each one-step increase in
set size. A single, lower-case probe letter was displayed
afterward, and participants had to indicate whether or
not the probe was contained within the initial set of letters.
There were three blocks of 20 trials each. Set sizes 4 y 5
were included primarily as a warm-up and were excluded
from analyses because of near-ceiling performance for all
Participantes. De este modo, the dependent variable was the median
RT for correct trials averaged across set sizes 6–10.
Metamemory
The metamemory task is a modified word list learning task
modeled after that used in McGillivray and Castel (2011)
and further described in Parlett-Pelleriti, lin, jones,
Linstead, and Jaeggi (2019). It consisted of learning five
12-word lists presented one word at a time on a computer
pantalla. Sin embargo, after encoding each word, Participantes
Au et al.
1019
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
were additionally asked to give a confidence rating
entre 0 y 9, indicating their degree of confidence
in their ability to subsequently recall that word. A flashing
red box appeared around the word after 3 sec if participants
had not yet responded. Participants were not penalized for
slow responses, nor did the task move on without their
respuesta. This was merely done to encourage quick
responses to ensure comparable encoding times among
Participantes. At the end of each 12-word list, Participantes
were asked to recall as many of the words from that list as
they could recall. Participants gave all confidence and recall
responses verbally while an experimenter typed in their
respuestas. This was done to control for typing speed and
ability between participants and allow participants to focus
solely on the recall task at hand. Parallel test versions were
administered at pretest, posttest, and follow-up (see Word
Generation Procedure above). The dependent variable was
the total number of correctly recalled words. Confidence
ratings and other potential variables of interest (Parlett-
Pelleriti et al., 2019) were not relevant to the present hypoth-
eses and were not analyzed in the current article.
Analytical Approach
Statistical analyses were conducted using STATA Version
13 (StataCorp, 2013). To interrogate training and transfer
efectos, we used linear mixed-effects models, cuales son
generalizations of ordinary least-squares linear regression,
but allow for the inclusion of random deviations other
than those associated with the overall error term. Specifi-
cally, our analyses accounted for participant-level random
intercepts that shift the regression line up or down accord-
ing to each participant’s starting ability. The general equa-
tion used was as follows:
Taskij ¼ β0 þ β1Sessionij þ β2Conditionij
þ β3Moderator þ β4−6Session XX Moderatorij
þ μ
þ εij
0i
Taskij represents the dependent measure for the ith par-
ticipant for the jth session. β0 represents the overall
regression intercept. β1–2 represent vectors of fixed effect
beta weight coefficients for the session and condition pre-
dictors. β3 represents fixed effect beta weight coefficients
for the baseline or spacing predictors, which are both used
in separate models to assess the extent to which these var-
iables moderate the results. β4–6 represents the fixed
effect beta weight coefficients for all the double and triple
interactions between the aforementioned predictors. El
fixed effect coefficients from β1–6 are equivalent to those
in a standard ordinary least squares regression. Finalmente, μ0i
represents the participant-level random intercept, and εij
represents a vector of error terms.
The predictor condition was coded as a dummy
variable representing the tDCS group referenced to sham,
whereas the predictor spacing was coded as a dummy
variable representing more spacing (es decir., training every
other day) referenced to less spacing (es decir., training daily).
Baseline was a continuous predictor based on pretest per-
formance on either the untrained number n-back task to
index baseline WM performance or the metamemory task
to index baseline LTM performance. Además, base-
line was mean-centered to zero so that interpretations of
regression coefficients are made with respect to average
baseline ability. A diferencia de, for the session predictor, zero
was referenced to the last session rather than to the mean
so that interpretations of regression coefficients can be
made with respect to the end of training and not the mid-
dle. All variables were left unstandardized in their natural
units, with the exception of baseline ability, as indicated in
all the table legends in Appendix A.
Three separate models were run for each analysis, a
main effects model and a separate interaction model for
each moderator of interest (baseline and spacing). El
main effects model was run because the interaction
models alone do not include estimates of main effects,
only partial effects and interactions. Separate interaction
models were run for each moderator because we were
not interested in interactions between baseline and
spacing. Además, using separate models reduces the
complexity of each model and avoids four-way interac-
ciones, which our analyses are underpowered to properly
detect or interpret.
Mediation Model
To interrogate the potential mediating role of LTM consol-
idation on long-term WM performance, we conducted
mediation analyses using ordinary least squares regression
via the SEM builder in STATA using full information maxi-
mum likelihood. In the absence of complete 3-month
follow-up data because of the COVID-19 lockdown, nosotros
used posttest performance on the trained n-back task as
our dependent measure of long-term WM performance.
The independent predictor variable was the dichotomous
variable Condition (active/sham) and the mediator was the
total number of n-back stimuli incidentally recalled (aver-
aged between both time points at the end of training and
at posttest). Baseline WM performance at pretest on the
untrained n-back task was also put in the model as an inde-
pendent variable to control for general relationships
between LTM and WM performance. With the exception
of the dummy variable, Condition, all variables in the
model were standardized as z-scores, and effects are
reported as standardized beta weights. Bias-corrected
95% confidence intervals were calculated for each direct
and indirect effect based on 5000 bootstrap samples
(Hayes, 2018).
As an additional sensitivity analysis, word learning re-
tention was used as an alternate mediator. This was done
to evaluate whether specific memory of n-back stimuli
was necessary to manifest a mediation effect of LTM on
WM performance or whether a more general measure of
individual consolidation strength would also show the
same effects. In this model, word learning retention was
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
1020
Revista de neurociencia cognitiva
Volumen 34, Número 6
across all five training sessions. We regressed this value
on the following predictors: condición, base, and spac-
En g. In two separate models, we also evaluated the interac-
tions between condition and baseline as well as condition
and spacing.
tDCS had little to no impact on intraday word learning as
our analyses found no main or interaction effects of condi-
ción ( ps > .19). See Table A2 for all regression coefficients.
Intraday Delayed Word Recall
To measure intraday delayed recall, we summed the total
number of words participants were able to remember after
a 30-min delay, across all five training sessions. Using this
value as the dependent variable, the same analytic proce-
dure was followed as described above for immediate word
aprendiendo. Despite null effects on immediate word recall,
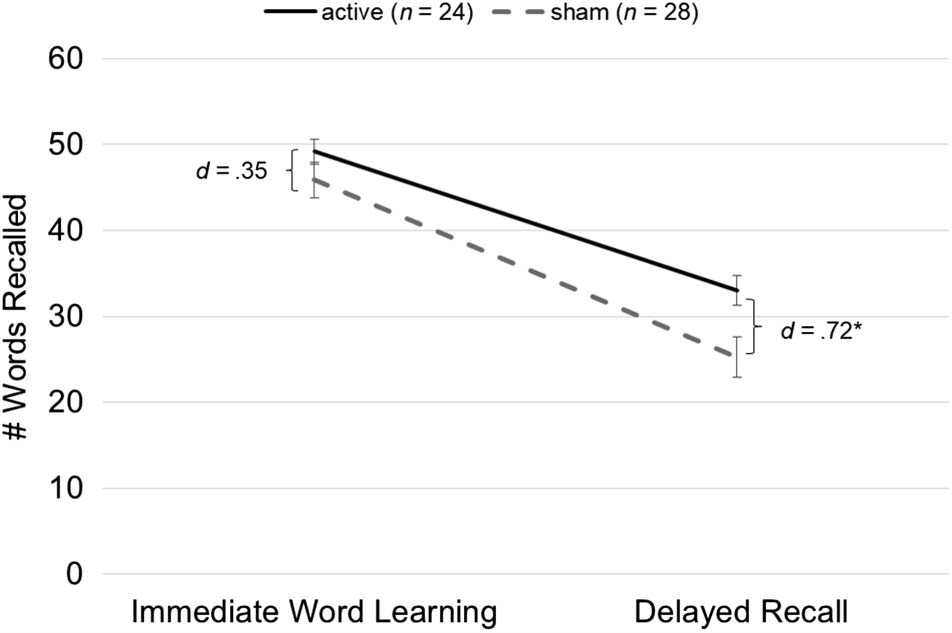
tDCS was effective in boosting delayed recall (Cifra 2).
Our main effects model revealed a significant effect of
Condition (b = 6.56, z = 2.57, pag = .01, re = 0.72) demostración
greater performance in the tDCS group. Neither the in-
teraction of Condition with Baseline nor Spacing was sig-
nificant ( ps > .095; Table A3).
Between-day Cumulative Word Recall
To test the effects of tDCS on cumulative word recall
between days, we regressed the total number of previously
learned words that participants were able to recall at the
beginning of each session on the following predictors:
Session, Condition, Base, and Spacing. Además de
testing the main effects, we also ran two separate models
testing the interactions between Session, Condition, y
Baseline as well as Session, Condition, and Spacing.
operationalized as a latent variable based on word reten-
tion at delayed recall, as well as at cumulative recall the
next day. Aquí, retention was defined specifically as the
total number of words recalled at each time point divided
by the total number of words initially learned after three
rounds of immediate recall, averaged across all five ses-
siones. At the cumulative recall time point measured the
next day, only words recalled from the previous session
were counted. De este modo, this is a measure of overnight consol-
idation, averaged over five sessions, rather than a cumula-
tive measure as used in the main results. Retention scores
were calculated in this way rather than simply using the
total number of words recalled to isolate time-dependent
consolidation processes from general cognitive ability. Para
ejemplo, a high-ability participant might initially learn 10
words and retain five, whereas a lower-ability participant
might learn only eight words but retain four. De este modo, control-
ling for general ability, both participants perform equally
and retain half their learning.
RESULTADOS
Descriptive data for pretest, posttest, and follow-up are
included in Table A1. Significant main and interaction
effects involving tDCS are reported below. All other
efectos, significant or not, are not reported in the main text
but are included in Tables A1–A11. Unless otherwise noted
in the figures or table captions, the analytic sample size
comprised 24 individuals who received active tDCS and
28 who received sham. Data at follow-up were unfortu-
nately not completed because of research disruption by
the COVID-19 pandemic. Although descriptive data on
the partial sample collected are reported in Table A1, sta-
tistical analyses were not carried out and are not reported.
Debriefing
Forty participants (20 active, 20 sham) were debriefed
about the existence of a sham group at the 3-month
hacer un seguimiento, 13 of whom were contacted by phone because
of the lockdown restrictions imposed by the COVID-19
pandemic. Del 20 active tDCS participants, solo 10
successfully guessed their true condition. Del 20 sham
Participantes, only nine successfully guessed their true con-
condición. There was no difference between groups in their
guesses (χ2
1,norte = 40 = 0.10, pag = .75). Además, allá
was also no difference between groups in their confidence
ratings concerning their guesses (mean active: 6.75/10;
mean sham: 6.2/10), t(38) = 0.69, pag = .25.
Training Effects
Intraday Immediate Word Learning
To measure intraday immediate word learning, nosotros
summed the total number of words participants were able
to learn after three rounds of immediate word recall,
Cifra 2. Intraday word recall. The total number of words recalled,
summed across all five training sessions, is shown on the y-axis.
Immediate word learning refers to the number of words recalled on
the third (and final) round of immediate recall. No significant effects
were observed, but the tDCS advantage approximately doubled and
became significant 30 min later at delayed recall. Error bars represent
SEM. * denotes significant Group effect ( pag < .05).
Au et al.
1021
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
with no main or interaction effects involving Condition
( ps > .55).
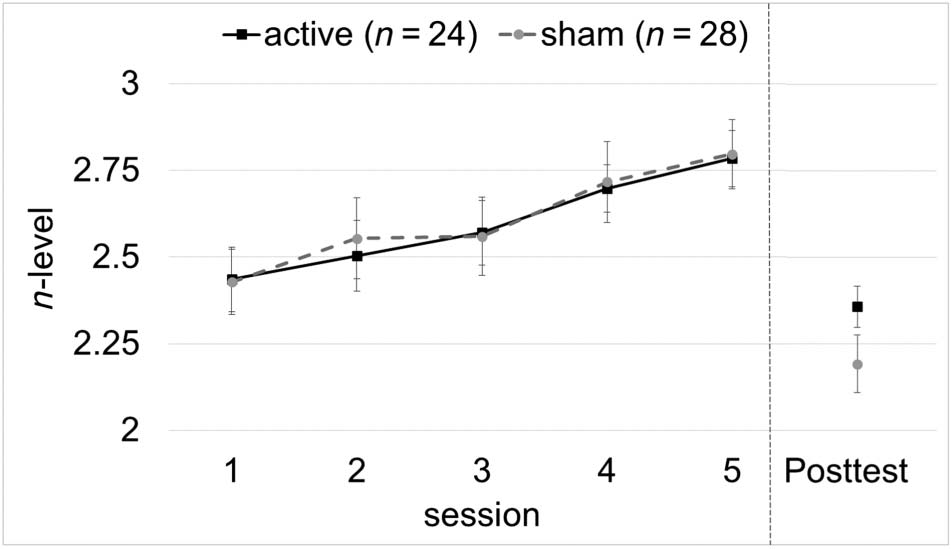
n-Back WM Training
To evaluate the effects of tDCS during n-back training, nuestro
linear mixed-effects model regressed the average n-back
level achieved each training day on the following predic-
tores: Session, Condition, Base, and Spacing. De nuevo, después
running the main effects model, we then ran two separate
interaction models testing first the interactions between
Session, Condition, and Baseline and then the interactions
between Session, Condition, and Spacing. Over the train-
ing period, tDCS had no observable effect on n-back per-
rendimiento (Cifra 4), with no main or interaction effects
( ps > .54). See Table A6 for all regression coefficients.
Posttest n-Back
We modeled n-back performance on the trained task sep-
arately at posttest because tDCS was not administered at
this time point and also the n-back level was reset back to
the beginning rather than continuing adaptively from the
last training session; de este modo, it did not continue the same lin-
ear trend from training. The analytic model at this time
point was the same as described above, except with the
omission of the Session predictor. Además, the ana-
lytic sample was smaller (norte = 44) because data from eight
Participantes (four active/four sham) were lost or not col-
lected because of a combination of technical glitches
and experimenter error.
Although there was no significant main effect of Condi-
ción ( pag = .17), we did observe a trending interaction with
Cifra 3. (A) Cumulative recall. The total number of words cumulatively
recalled at the beginning of each session, including posttest, is shown on
the y-axis. These words include all words learned since the beginning
of training up until the current session (de este modo, Session 1 is excluded from
this figure). The group receiving active tDCS consistently outperformed
the sham group in each session, with the largest difference manifesting
at posttest. (B) Every-other-day spacing condition. tDCS appeared
less effective when training sessions were separated by 2 días, con un
nonsignificant group difference by posttest (re = 0.40). (C) Daily spacing
condición. tDCS was more effective when training sessions were only
separated by 1 día, with a significant group difference by posttest (re = 1.33).
Error bars represent SEM. * denotes significant Group effect ( pag < .05).
In addition to promoting delayed recall intraday, tDCS
also improved cumulative recall throughout the training
period (Figure 3). There was a main effect of Condition,
showing a tDCS advantage across all sessions (b = 3.187,
z = 2.40, p = .016). There were also significant Session ×
Condition interactions in both the baseline and spacing
regression models ( ps < .001), showing an increasing
tDCS advantage over time, culminating in a large effect size
difference by posttest (d = 0.82). Finally, despite the lack of
a main effect of Spacing ( p = .15), we observed both a
Condition × Spacing interaction (b = −6.47, z = −2.12,
p = .034) as well as a Condition × Session × Spacing
interaction (b = −1.17, z = −1.98, p = .048), which
indicate that every-other-day spacing significantly reduced
the overall advantage of tDCS, as well as the marginal
advantage per session. See Table A4 for all regression
coefficients.
Word Recognition
In addition to free recall, we also tested the effects of tDCS
on recognition memory at posttest (Table A5). We
regressed recognition accuracy on the following predic-
tors: Condition, Baseline, and Spacing. After running the
main effects model, we tested for interactions between
Condition and each of the other two predictors in two
separate models. Our analyses found no evidence that
tDCS influenced performance on recognition memory,
Figure 4. Trained n-back task. Training performance on the n-back
task was very similar between groups across all five sessions and
also was not significantly different at posttest. Posttest n-back was
administered without stimulation and also without adaptively continuing
from the previous session (i.e., all participants started over at 1-back).
At pretest (not shown), participants were introduced to four rounds
of n-back training to habituate them to the task before training and
stimulation. The first training session continued adaptively from this
pretest exposure, which explains why Session 1 performance is greater
than at posttest.
1022
Journal of Cognitive Neuroscience
Volume 34, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Baseline (b = −0.17, z = −1.93, p = .054). Because of our
a priori hypothesis and the robust literature support for a
baseline-dependent effect of tDCS (Krebs, Peter, Wyss,
Brem, & Klöppel, 2021; Perceval et al., 2020; Arciniega,
Gözenman, Jones, Stephens, & Berryhill, 2018; Katz
et al., 2017; Gözenman & Berryhill, 2016; Heinen et al.,
2016; Looi et al., 2016; Minichino et al., 2015; Hsu, Tseng,
Liang, Cheng, & Juan, 2014; Tseng et al., 2012), we con-
ducted a post hoc analysis on this interaction, which
revealed that lower-baseline tDCS participants (lowest
50th percentile) outperformed lower-baseline sham
participants, t(22) = 2.50, p = .02, d = 1.06, whereas
the same comparison among high-baseline performers
yielded no difference, t(18) = 0.74, p = .47, d = 0.33.
The overall group difference, irrespective of baseline per-
formance, was d = 0.47. See Table A7 for all regression
coefficients.
n-Back Stimuli Incidental Memory Recall
To evaluate incidental memory during n-back training, we
regressed the total number of correctly recalled stimuli,
both on the last training day and at posttest, on the follow-
ing predictors: Session, Condition, Baseline (based on
pretest metamemory performance), and spacing. Once
again, interactions between Session, Condition, and Base-
line as well as Session, Condition, and Spacing were tested
in separate models after testing for main effects. Addition-
ally, the analytic sample was smaller (n = 46) because data
from six participants (five active/one sham) were not col-
lected at the end of training because of experimenter error
and were removed from the entire analysis.
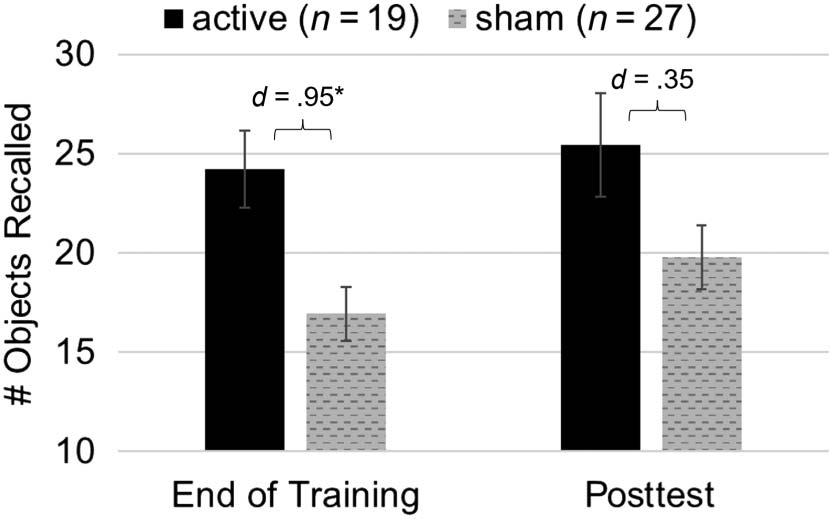
We found that tDCS improved incidental memory recall of
stimuli encountered throughout n-back training (Figure 5),
with a main effect of Condition (b = 4.89, z = 2.43, p =
.015) but no significant interactions ( ps > .21). Ver
Table A8 for all regression coefficients.
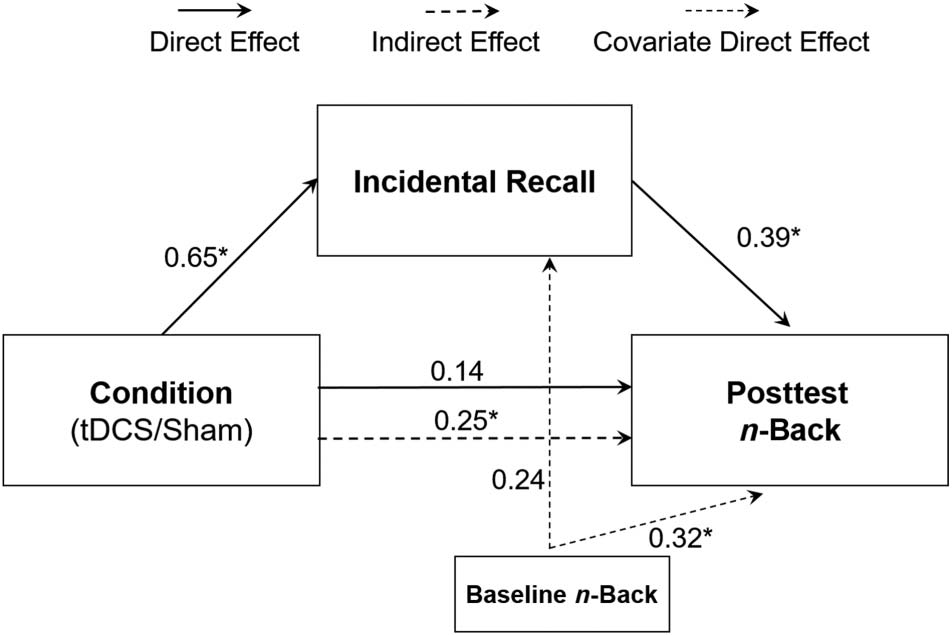
Cifra 6. Mediation model. We found significant direct effects from
condition to incidental recall and from incidental recall to posttest
n-back, resulting in a significant indirect effect from condition to
posttest n-back (β = 0.25). The bias-corrected confidence interval did
not include zero (95% CI [0.03, 0.67]) después 5000 bootstrap samples.
Baseline performance on the untrained n-back variant was included to
control for general correlations between LTM and WM performance
that may directly or indirectly influence posttest n-back scores.
* denotes significant path (bias-corrected confidence interval excludes
zero).
Mediators of Long-term n-Back Improvements
To understand the relationship between tDCS and long-
term WM improvements, we tested a mediation model
evaluating the role of LTM as a mediating variable between
condición (active/sham) and posttest n-back (Cifra 6). Todo
variables were standardized, and paths are reported as
standardized beta weights, with the exception of condi-
ción, whose paths are unstandardized. Incidental memory
of n-back stimuli was used as the mediator, and baseline
scores on the untrained n-back variant were included in
the model as a control. We found significant direct effects
of Condition on incidental recall (b = 0.65, 95% inclinación-
corrected CI [0.09, 1.21]) and incidental recall on posttest
n-back (β = 0.39, 95% bias-corrected CI [0.09, 0.71]),
resulting in a significant indirect effect of Condition on
posttest n-back (b = 0.25, 95% bias-corrected CI [0.03,
0.67]). We also ran a sensitivity analysis with an alternate
mediator, using a latent variable derived from word reten-
tion scores measured 30 min and 24 hr after initial word
aprendiendo. A similar pattern of results emerged, with a signif-
icant direct effect of Condition on word retention (b =
0.84, 95% bias-corrected CI [0.37, 1.32]), a marginal direct
effect of word retention on posttest n-back (β = 0.45, 95%
bias-corrected CI [−0,06, 0.88]), and a significant indirect
effect of Condition on posttest n-back (b = 0.38, 95%
bias-corrected CI [0.05, 0.98])
Cifra 5. Incidental recall. Participants receiving tDCS demonstrated
an advantage on incidental recall of n-back stimuli. Despite the lack of a
significant group difference at the posttest time point alone, there is no
interaction with Session, and this advantage is statistically significant
en general. Error bars represent SEM. * denotes significant Group effect
( pag < .05).
Transfer Effects
For each of the transfer tasks, the untrained n-back, meta-
memory, and the Sternberg, we regressed the dependent
Au et al.
1023
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
variable of interest on the following predictors: condition
and spacing. After running the main effects model, we
tested for interactions between Condition and each of
the other two predictors in two separate models. No
transfer effects were detected on any task; specifically,
there were no main effects of Condition nor any significant
interactions (all ps > .23; Tables A10–A12). Base
interaction models were not tested because baseline is
colinear with pretest scores.
Power Analysis
Since the COVID-19 pandemic paused research activities
and cut our recruitment short, we conducted a retrospec-
tive power analysis using the R package pwr to determine
the level of power we had in detecting our main training
efectos. For LTM effects, we considered our effect size esti-
mates derived from the average delayed recall over five
sessions (re = 0.72) and the cumulative recall at posttest
(re = 0.82), which were both within the confidence interval
of meta-analytic estimates (re = 0.625, 95% CI [0.250,
0.999]) of tDCS-induced episodic memory improvement
in older adults (Huo et al., 2021). Using sample sizes of
24 y 28 and a two-sided significance threshold of α =
.05, we found that we had power (1 − β) entre .60
y .82 to detect effect sizes ranging between meta-
analytic and our own observed effects. Power for WM train-
ing effects were calculated with the same parameters,
except we used the overall effect size at posttest of d =
0.47, which again was within the confidence interval of
meta-analytic estimates (re = 0.28, 95% CI [0.06, 0.52])
of tDCS-induced improvements on WM training (Mancuso
et al., 2016). Aquí, fuerza (1 − β) ranged between .18 y
.38, suggesting we may have been underpowered to
detect a true effect if it existed. Sample sizes of approxi-
mately 72–188 participants per group would have been
required to detect effects in this range with traditionally
accepted statistical power (1 − β = .80). Despite having
our recruitment cut short from the pandemic and being
underpowered in some of our analyses, we note that our
sample size exceeded many other tDCS studies of WM
training, which ranged from 10 a 21 per group (Mancuso
et al., 2016), and was comparable to many other studies
of LTM, which ranged from 10 a 48 per group (Huo
et al., 2021; Galli et al., 2019).
DISCUSIÓN
The current study set out with three initial aims: to quan-
tify the relative efficacy of tDCS on LTM and WM training in
older adults over the course of five training sessions, a
evaluate the extent to which LTM consolidation mediates
WM improvements, and to identify potential moderators
of the tDCS effect. With respect to the first aim, we found
tDCS to be effective in boosting LTM performance, con
effects about 1.5 times stronger (d = 0.72–0.82) than
those observed after WM training (re = 0.47), which were
not significant. The LTM improvements were observed in
free recall but not recognition, in line with previous
reports (Perceval et al., 2020; Galli et al., 2019; Leshikar
et al., 2017; Matzen, Trumbo, Leach, & Leshikar, 2015),
which collectively suggest that tDCS affects LTM by facili-
tating processes related to elaborate recollection rather
than surface-level familiarity (cf. Yonelinas, 2002). Más-
más, these effects were time dependent in that no signif-
icant differences were observed between groups at
immediate recall directly after word learning (re = 0.35;
Cifra 2), but a tDCS advantage started appearing at the
30-min delayed recall (re = 0.72; Cifra 2), which persisted
for the duration of the week-long intervention and was
most pronounced at posttest (re = 0.82; Cifra 3). Este
suggests that tDCS decreases the rate of forgetting or,
put another way, increases the strength of consolidation.
The lack of transfer to a novel word list at posttest rules out
general improvements in strategies or processing ability,
suggesting specificity to the words learned during stimula-
ción. En tono rimbombante, benefits were observed both explicitly
when participants were aware of the eventual word recall
tareas, as well as incidentally when participants passively
encoded a variety of stimuli during n-back training without
being told of an eventual recall task (Cifra 5). En general, nuestro
results add to accumulating evidence demonstrating that
prefrontal tDCS can enhance LTM consolidation in
humanos (Huo et al., 2021; Galli et al., 2019) and is consis-
tent with animal work that shows that tDCS increases hip-
pocampal LTP and levels of brain-derived neurotrophic
factor, an important protein for learning and consolidation
(Cocco, Podda, & Grassi, 2018; Podda et al., 2016). Cómo-
alguna vez, a critical difference between human and animal work
is that the human hippocampus is too deep to be directly
targeted by tDCS. De este modo, to the extent that mechanisms of
human tDCS are analogous to animal models, effects may
derive from increased LTP-like plasticity in superficial cor-
tical areas or may indirectly stem from hippocampal LTP
via hippocampal-prefrontal circuitry (Sigurdsson &
Duvarci, 2016; Blumenfeld & Ranganath, 2007).
Despite the robust effects of tDCS on LTM, Había
no discernable effects on WM at the group level (Cifra 4).
Performance during the training period was virtually iden-
tical between groups, although see Moderation section
further below for a discussion of possible baseline-
dependent effects. In considering the reasons for this
overall null effect, one advantage of the current study
design is that we are able to rule out explanations related
to an ineffective tDCS montage. Because of the black box
nature of delivering a current through the scalp, dónde
even small displacements in electrode positioning can dra-
matically alter current density over the desired cortical
región (Ramaraju, Roula, & McCarthy, 2018), it is often dif-
ficult to ascertain the extent to which the experimental
manipulation successfully targets the cortical ROI at an
individual level. This can be one major source of variability
between studies that can account for some of the
1024
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
unreliability in WM effects throughout the literature. Cómo-
alguna vez, because of the robust tDCS-related improvements in
LTM, we can largely rule out this concern and focus on
alternative explanations. Por ejemplo, one possibility we
cannot rule out is interference effects or neural competi-
tion between LTM and WM since participants perform the
two tasks back-to-back during training. Interference
between different memory domains, even with distinct
memoranda, has been documented before (Marrón &
Robertson, 2007) and may have eclipsed any potential
tDCS effects. This would be consistent with why we
observe the greatest numeric difference at posttest (ver
Cifra 4) where there is no LTM task administered before
n-back, despite performance that is almost completely
overlapping during training. Además, a recent article
demonstrates that LTM consolidation during sleep dimin-
ishes the benefit of sleep toward improving WM training
actuación (Chen, Niknazar, Alaynick, Whitehurst, &
Mednick, 2021). Sin embargo, the extent to which these fac-
tors truly influence our results is beyond the ability of our
study design to investigate, and a large randomized con-
trolled trial of tDCS in older adults has also previously
failed to observe benefits to WM training even in the
absence of LTM tasks (Nilsson et al., 2017). De este modo, we will
refrain from further speculation here.
Mediation Effects
Regardless of the specific reason why we did not observe a
significant effect of tDCS on WM training, our mediation
analyses suggest that any effect that did exist may actually
have been driven by LTM rather than any direct improve-
ments in WM per se. Específicamente, we observed that individ-
uals assigned to the active tDCS condition performed an
average of 0.65 SDs higher than their sham counterparts
on the incidental recall task embedded in the n-back train-
En g (ver figura 6). Sucesivamente, each standard deviation
increase in recall scores resulted in a 0.39-SD increase in
n-back performance at posttest. The indirect effect of
tDCS on posttest n-back scores was thus an average
mejora de 0.33 SDs and was statistically significant.
En otras palabras, even though we found no evidence that
tDCS directly improved the ability to manipulate items
held in WM during the n-back task, it did improve the
long-term retention of these items, which in turn pre-
dicted better WM performance at posttest. From this,
one might be tempted to speculate that increased familiar-
ity with the items in LTM facilitated WM performance
involving those same items, as has been demonstrated
antes (Oberauer, Awh, & Sutterer, 2017). Although this
may be true to some extent, especially given reciprocal
interactions between the hippocampus and pFC in both
LTM and WM ( Jin & Marén, 2015), our sensitivity analysis
revealed the same pattern of results even when using word
retention as the mediator rather than n-back stimuli recall.
Por lo tanto, what these mediators may actually be measur-
ing are individual differences in the ability to consolidate
information more generally. This is in line with a body of
research demonstrating that sleep-dependent consolida-
tion augments WM training benefits (Ferrarelli et al.,
2019; Sattari et al., 2019; Pugin et al., 2015). A pesar de
our study is unable to pinpoint what exactly is being con-
solidated that benefits WM training, there has been spec-
ulation in the literature that WM training benefits may arise
from the acquisition of new cognitive routines, akin to the
acquisition of new skill sets, rather than improvements in
existing processes (Gathercole et al., 2019). Our current
data fit within this framework.
En tono rimbombante, our mediation models controlled for a
second independent variable, baseline n-back perfor-
mance, which itself could have predictive value for both
posttest n-back as well as LTM recall scores because of
shared variance between WM and LTM task performance
(Unsworth, 2010; Ranganath & Blumenfeld, 2005). De este modo,
controlling for baseline rules out alternative explanations
that the mediation effect could simply be related to gen-
eral correlations between WM and LTM. De hecho, we did
observe a small indirect effect (0.12 SDs) of baseline
n-back performance on posttest n-back, suggesting that
this shared variance does play a role in the interrelation-
ships between LTM and WM performance (es decir., those
who perform well on one memory task are likely to per-
form well on another memory task). Sin embargo, el
indirect effect of tDCS on n-back performance remained
significant above and beyond any confounding influences
of these interrelationships.
Finalmente, a weakness to our mediation model is the lack of
a preexisting relationship between tDCS and posttest
n-back. This is not a statistical weakness, as such a relation-
ship is not a necessary prerequisite for probing indirect
effects in mediation analysis (Hayes, 2018; O’Rourke &
MacKinnon, 2018; Rucker, Preacher, Tormala, & Petty,
2011), and in fact, it is in cases where a total effect is absent
where mediation analyses can be especially informative by
proposing an alternate causal chain other than the original
independent-dependent variable relationship (Pieters,
2017). Sin embargo, it does hamper our efforts to explain
the long-term effects of tDCS on WM performance in
other studies that “did” observe an overall effect on
WM enhancement (p.ej., Ruf et al., 2017; Au et al., 2016;
Esteban & Berryhill, 2016; jones, Esteban, et al., 2015;
Parque, SEO, kim, & Ko, 2014; Martin et al., 2013), porque
the mechanisms of action in these studies may potentially
differ. Although our current results suggest that the effect
of tDCS on WM is fully mediated through its effect on con-
solidation, it would be important for future studies that do
find a strong overall effect on WM to parcel out the relative
contributions of the direct and indirect effects of tDCS.
This understanding could serve to inform future studies
and to increase the precision of training and transfer
efectos. Por ejemplo, if a large portion of what is actually
being modulated by tDCS actually pertains to processes
related to LTM rather than WM per se, then training can
be spaced appropriately to allow for consolidation to
Au et al.
1025
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
occur, and transfer can focus on the declarative and
procedural components of the trained task rather than
any WM-specific skills. The current study provides a prime
example of this in that we found significant effects on the
declarative recall of stimuli used during n-back training
despite a lack of an overall effect on the WM portion of
the task itself.
Moderation Effects
Although tDCS did not seem to enhance WM training at
the group level, we found tentative evidence ( pag = .054)
at posttest for a selective benefit in those who started with
lower baseline WM performance. This is consistent with
our previous work in young adults (Katz et al., 2017), como
well as in a body of other tDCS work that also shows a
low-baseline advantage across a variety of tasks (krebs
et al., 2021; Perceval et al., 2020; Arciniega et al., 2018;
Gözenman & Berryhill, 2016; Heinen et al., 2016; Looi
et al., 2016; Minichino et al., 2015; Hsu et al., 2014; Tseng
et al., 2012). De este modo, despite the marginal nature of the cur-
rent findings, the cumulative evidence for a moderating
influence of baseline ability is compelling and should be
an important consideration in future studies for identify-
ing potential responders and nonresponders to tDCS.
In addition to baseline, the spacing interval between
sessions was also found to moderate the effects of tDCS.
There was a selective tDCS advantage for the daily training
subgroup in the cumulative word recall task (Figure 3B–
C), which involved learning between days, but no effects
on delayed or immediate word recall, which only involved
intraday learning (and thus an effect of spacing would not
be expected). Correspondingly, the daily training sub-
group also outperformed the every-other-day group dur-
ing n-back training as well, but there were no differential
effects as a function of tDCS. Although these results
should be interpreted with caution because of the small
spacing subgroup sizes (entre 12 y 15 per group;
Figure 3B–3C), it is notable that the effect size of daily
spacing was over three times greater than every-other-
day spacing on the cumulative recall task by posttest (re =
1.33 vs. re = 0.40). It may be that training every other day
left too much time for forgetting to occur in our cohort of
adultos mayores, which may have eclipsed any effect tDCS could
have had on boosting between-session consolidation. A
related issue to consider is that longer spacing intervals
are generally optimized for longer retention intervals
(Cepeda, Vul, Rohrer, Wixted, & Pashler, 2008). De este modo, con
the short retention interval (∼1–2 weeks) of our current
estudiar, there may not have been enough passage of time
to show any advantage of every-other-day spacing above
and beyond the benefits of daily spacing. Although we did
have a longer retention interval planned as a 3-month
follow-up in our study, we were unable to complete data
collection because of COVID-19 and unfortunately are
unable to draw any meaningful conclusions. Sin embargo, ver
Table A1 for descriptive data on the follow-up measures.
We note that this advantage of daily spacing stands in
contrast with our previous study on young adults (Au
et al., 2016), in which almost all of the tDCS benefit on
n-back training appeared over a weekend (∼72-hr spac-
En g) rather than between consecutive weekdays. Cómo-
alguna vez, this contrast is not necessarily contradictory, como el
two studies assessed different populations and different
memory domains (LTM vs. W.M.). Desafortunadamente, we did
not find an overall tDCS effect or a tDCS × Spacing inter-
action within the WM domain in the current study, cual
would have afforded a more direct comparison to our
previous results. Once more, sin embargo, we reiterate that
these subgroup analyses, both in our current and previous
resultados, contain rather small sample sizes and should be
interpreted cautiously when informing future study
designs. So, although we do not advise overreliance on
the specific spacing intervals reported in our studies, especialmente-
cially given that the optimal spacing and retention intervals
will vary because of a variety of factors such as task selec-
ción, población, and study length, we do admonish that
spacing is an important and underexplored moderator in
the tDCS literature that has both theoretical and empirical
basis to influence responsiveness to tDCS (Goldsworthy
et al., 2015; Monte-Silva et al., 2013; Alonzo, Brassil, taylor,
Martín, & Loo, 2012; Monte-Silva, kuo, Liebetanz, Paulus,
& Nitsche, 2010) and should be considered when design-
ing longitudinal tDCS studies meant to act upon between-
session consolidation.
Conclusions
The current study found that tDCS is effective for improv-
ing memory performance among older adults. As we pre-
dicted, there were stronger and more robust effects on
LTM than WM, presumably because of the role tDCS plays
in enhancing consolidation processes. This is underscored
by the observation that, despite the lack of an overall effect
on n-back WM performance, we did observe a strong
effect on the incidental recall of the stimuli used in the
tarea n-back. En otras palabras, even though the short-term
relational characteristics of WM stimuli were not better
encoded (es decir., their n position), their long-term semantic
characteristics were. Además, our mediation model sug-
gested that any influence tDCS did have on WM training at
posttest was mediated through its effects on enhancing
consolidation.
Además, both of our hypothesized moderators,
baseline and spacing, were influential in different contexts.
Primero, we found modest evidence that tDCS was selectively
beneficial for individuals with low baseline WM abilities.
Segundo, we also found that tDCS was selectively beneficial
for LTM consolidation in older adults when applied daily
rather than every other day. We posit that there is likely
an optimal spacing interval for tDCS to act upon after
enough time has passed for memory consolidation to
reach a certain strength but before too much forgetting
ocurre. This optimal interval, sin embargo, can be variable
1026
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
for different tasks and different populations and is largely
an unknown factor in most studies.
En general, our results add to a line of existing literature
that has documented an association between prefrontal
activity and LTM (reviewed in Blumenfeld & Ranganath,
2007). Además, the use of tDCS in our study to modulate
this prefrontal activity goes beyond these previous neuro-
psychological and neuroimaging studies by implicating a
causal role of the pFC in LTM formation and consolidation.
Given the ubiquity of age-related memory decline (Pequeño,
2001) and its predictive value in everyday functioning in
old age (Borella et al., 2017), it is imperative that we
develop new treatments and interventions to mitigate this
decline. The use of tDCS to enhance learning and consol-
idation in older adults is one step in that direction and can
be used to facilitate the acquisition of new skills, knowl-
borde, and hobbies that may help older adults maintain
active and stimulating lives well into their golden years.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
oh
C
norte
_
a
_
0
1
8
3
9
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
APPENDIX A
Table A1. Descriptive Data
Active tDCS Group
Sham Group
Pre
(norte = 24)
–
Correo
(norte = 24)
Follow-up
(norte = 13)
2.36 (0.27)
2.41 (0.39)
Pre
(norte = 28)
–
Correo
(norte = 28)
Follow-up
(norte = 14)
2.19 (0.41)
2.50 (0.61)
24.21 (8.36)
25.42 (11.36)
20.18 (15.09)
16.93 (7.09)
19.78 (8.43)
11.92 (8.25)
–
–
16.39 (9.05)
8.15 (11.00)
0.67 (0.09)
0.62 (0.07)
–
–
9.63 (7.59)
2.07 (2.87)
0.69 (0.08)
0.57 (0.08)
Trained n-backa
(n-level)
n-Back stimuli recallb
(No. of objects)
Cumulative recall
(No. of words)
Word recognition
(% correcto)
Untrained n-back (Pr)
0.62 (0.17)
0.67 (0.09)
0.62 (0.07)
0.61 (0.20)
0.76 (0.15)
0.73 (0.10)
Sternberg item
recognition (segundo)
1190.67
(136.40)
0.75 (0.17)
0.75 (0.13)
1195.53
(210.96)
1171.22
(183.23)
1138.92
(193.26)
Metamemory
(% correcto)
0.29 (0.14)
1147.98
(126.27)
1116.20
(102.54)
0.29 (0.15)
0.33 (0.15)
0.12 (0.17)
Descriptive data are provided for pretest, posttest, and follow-up. Values in parentheses are standard deviations. Pr values for the untrained n-back
are measures of accuracy as described in the Methods.
a Because of experimenter error, the analytic sample for the trained n-back task at posttest comprised 20 active and 24 sham participants.
b For n-back stimuli recall, the pretest column refers to the first measurement after Training Session 5. Because of technical glitches and experi-
menter error, the analytic sample sizes for the active and sham groups were n = 19 and n = 27, respectivamente.
Au et al.
1027
Table A2. Intraday Immediate Word Learning
Modelo
Main effects
Base
Spacing
Variable
Condition
Baseline LTM
Spacing
Condition
Baseline LTM
Condition × Baseline
Condition
Spacing
Condition × Spacing
B
2.24
4.71
−1.85
2.30
4.82
−0.14
4.78
−0.99
−3.02
SE B
2.25
1.12
2.23
2.17
1.83
2.15
3.61
4.16
4.98
pag
.32
<.001* .41 .29 .008* .95 .19 .81 .55 n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham and daily spacing. Baseline is a continuous variable mean-centered to zero and standardized. * p < .05. l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / 3 4 6 1 0 1 5 2 0 1 9 2 3 2 / Table A3. Intraday Delayed Word Recall Model Main effects Baseline Spacing Variable Condition Baseline LTM Spacing Condition Baseline LTM Condition × Baseline Condition Spacing Condition × Spacing B 6.56 4.87 −3.25 6.66 5.05 −0.18 12.26 0.39 −9.06 SE B 2.57 1.28 2.55 2.51 2.06 2.50 3.93 4.63 5.42 / j o c n _ a _ 0 1 8 3 9 p d . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 p .011* <.001* .20 .008* .014* .94 .002* .93 .095 n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham and daily spacing. Baseline is a continuous variable mean-centered to zero and standardized. * p < .05. 1028 Journal of Cognitive Neuroscience Volume 34, Number 6 Table A4. Between-day Cumulative Recall Model Main effects Baseline Spacing Variable Session Condition Baseline LTM Spacing Session Condition Baseline LTM Session × Condition Session × Baseline Condition × Baseline Session × Condition × Baseline Session Condition Spacing Session × Condition Session × Spacing Condition × Spacing Session × Condition × Spacing B 2.00 3.18 2.07 −1.89 1.54 5.17 2.56 0.97 0.47 1.79 0.33 1.41 9.12 0.10 1.68 0.15 −6.47 −1.17 SE B 0.15 1.32 0.66 1.32 0.20 1.46 0.97 0.29 0.19 1.46 0.29 0.29 2.18 2.07 0.42 0.40 3.05 0.59 p <.001* .016* .002* .15 <.001* <.001* .008* <.001* .014* .22 .24 <.001* <.001* .96 <.001* .70 .034* .048* n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham and daily spacing. Baseline is a continuous variable mean-centered to zero and standardized. Session is an unstandardized, continuous variable, referenced to the last session rather than the mean for ease of interpretation. * p < .05. Table A5. Word Recognition Model Main effects Baseline Spacing Variable Condition Baseline LTM Spacing Condition Baseline LTM Condition × Baseline Condition Spacing Condition × Spacing B −0.03 0.05 −0.05 −0.03 0.03 0.03 −0.02 −0.05 −0.01 SE B 0.02 0.01 0.02 0.02 0.01 0.02 0.03 0.03 0.04 p .10 .001* .012* .14 .015* .10 .64 .14 .76 n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham and daily spacing. Baseline is a continuous variable mean-centered to zero and standardized. * p < .05. Au et al. 1029 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / 3 4 6 1 0 1 5 2 0 1 9 2 3 2 / / j o c n _ a _ 0 1 8 3 9 p d . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Table A6. n-Back WM Training Model Main effects Baseline Spacing Variable Session Condition Baseline WM Spacing Session Condition Baseline WM Session × Condition Session × Baseline Condition × Baseline Session × Condition × Baseline Session Condition Spacing Session × Condition Session × Spacing Condition × Spacing Session × Condition × Spacing B 0.09 −0.03 0.18 −0.07 0.09 −0.03 0.14 0.001 0.01 0.21 −0.05 0.13 −0.09 −0.29 −0.05 −0.08 0.11 0.08 SE B 0.01 0.11 0.06 0.12 0.02 0.15 0.09 0.02 0.02 0.15 0.03 0.02 0.19 0.18 0.03 0.03 0.26 0.05 p <.001* .78 .002* .57 <.001* .83 .14 .98 .47 .18 .10 <.001* .65 .10 .18 .013* .67 .10 n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham and daily spacing. Baseline is a continuous variable mean-centered to zero and standardized. Session is an unstandardized, continuous variable, referenced to the last session rather than the mean for ease of interpretation. * p < .05. Table A7. Posttest n-Back Model Main effects Baseline Spacing Variable Condition Baseline WM Spacing Condition Baseline WM Condition × Baseline Condition Spacing Condition × Spacing B 0.13 0.15 0.02 0.12 0.20 −0.17 0.07 −0.17 0.19 SE B 0.09 0.05 0.10 0.09 0.06 0.09 0.14 0.16 0.20 p .17 .003* .82 .17 .001* .054 .62 .28 .33 n = 44. Because of a combination of technical glitches and experimenter error, data were lost or not collected for eight participants. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham and daily spacing. Baseline is a continuous variable mean-centered to zero and standardized. * p < .05. 1030 Journal of Cognitive Neuroscience Volume 34, Number 6 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / 3 4 6 1 0 1 5 2 0 1 9 2 3 2 / / j o c n _ a _ 0 1 8 3 9 p d . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Table A8. n-Back Stimuli Incidental Memory Recall Model Main effects Baseline Spacing Variable Session Condition Baseline LTM Spacing Session Condition Baseline LTM Session × Condition Session × Baseline Condition × Baseline Session × Condition × Baseline Session Condition Spacing Session × Condition Session × Spacing Condition × Spacing Session × Condition × Spacing B 2.17 4.89 3.33 −3.78 3.13 4.01 4.27 −2.59 2.50 1.28 0.72 2.92 2.98 −5.35 −3.10 −0.12 4.62 3.42 SE B 1.10 2.02 0.94 1.97 1.31 2.32 1.43 2.06 1.27 2.20 1.95 2.11 3.45 3.20 3.05 2.83 5.00 4.42 p .05* .015* >.001*
.054
.017*
.084
.003
.21
.049*
.56
.71
.17
.39
.095
.31
.97
.36
.44
norte = 46. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectivamente, to sham
and daily spacing. Baseline is a continuous variable mean-centered to zero and standardized. Session is an unstandardized, continuous variable,
referenced to the last session rather than the mean for ease of interpretation.
* pag < .05.
Table A9. Mediation Effects
Independent Variable
Direct Effect
(a → b)
Direct Effect
(b → c)
Direct Effect
(a → c)
Indirect Effect
(a → c)
Condition
Baseline
Condition
Baseline
Mediator: Incidental Recall
0.65 [0.09, 1.21]*
0.39 [0.09, 0.71]*
0.14 [−0.42, 0.61]
0.25 [0.03, 0.67]*
0.24 [−0.07, 0.69]
0.39 [0.09, 0.71]*
0.32 [0.11, 0.60]*
0.12 [0.03, 0.29]*
Mediator: Word Retention
0.84 [0.37, 1.32]*
0.45 [−0.06, 0.88]
−0.09 [−0.89, 0.60]
0.38 [0.05, 0.98]*
0.27 [−0.01, 0.52]
0.45 [−0.06, 0.88]
0.28 [−0.02, 0.71]
0.12 [0.01, 0.33]*
n = 52. Regression coefficients for direct and indirect effects are shown for both mediation models (incidental recall and word retention). The
independent variable is represented by a, the mediator by b, and the dependent variable (posttest n-back) by c. All variables are standardized except
for Condition, which is a dummy variable referenced to sham. 95% bias-corrected confidence intervals are included in brackets.
* p < .05
Au et al.
1031
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
A1. Transfer Effects
Table A10. Untrained n-Back
Model
Main effects
Interaction
Variable
Session
Condition
Spacing
Session
Condition
Spacing
Session × Condition
Session × Spacing
Condition × Spacing
Session × Condition × Spacing
B
0.14
0.004
−0.01
0.13
0.04
−0.05
−0.04
0.01
−0.07
0.06
SE B
0.02
0.04
0.04
0.04
0.11
0.11
0.06
0.06
0.16
0.08
p
<.001*
.93
.76
.001*
.70
.63
.53
.82
.68
.44
n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham
and daily spacing. Session is an unstandardized, continuous variable, referenced to the last session rather than the mean for ease of interpretation.
* p < .05.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table A11. Sternberg Item Recognition
Model
Main effects
Interaction
Variable
Session
Condition
Spacing
Session
Condition
Spacing
Session × Condition
Session × Spacing
Condition × Spacing
Session × Condition × Spacing
B
−38.56
−9.61
46.45
−63.36
−16.04
−61.77
16.02
72.52
53.92
−63.23
SE B
16.70
42.96
42.79
63.98
146.64
138.58
93.24
88.16
205.43
130.27
p
.021*
.82
.28
.32
.91
.66
.86
.41
.79
.63
n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham
and daily spacing. Session is an unstandardized, continuous variable, referenced to the last session rather than the mean for ease of interpretation.
* p < .05.
1032
Journal of Cognitive Neuroscience
Volume 34, Number 6
Table A12. Metamemory
Model
Main effects
Interaction
Variable
Session
Condition
Spacing
Session
Condition
Spacing
Session × Condition
Session × Spacing
Condition × Spacing
Session × Condition × Spacing
B
0.05
0.03
0.02
−0.01
−0.01
−0.04
0.06
0.07
−0.10
−0.01
SE B
0.02
0.04
0.04
0.04
0.11
0.10
0.06
0.06
0.16
0.09
p
.033*
.43
.65
.82
.92
.67
.31
.23
.54
.89
n = 52. Regression coefficients are displayed for each predictor. Condition and Spacing are both dummy variables referenced, respectively, to sham
and daily spacing. Session is an unstandardized, continuous variable, referenced to the last session rather than the mean for ease of interpretation.
* p < .05.
Acknowledgments
The authors would like to thank Dr. Grace Lin (Massachusetts
Institute of Technology) for her help in putting together and
matching the various word lists used in this study. We would
also like to thank the many undergraduate and postbaccalaure-
ate research assistants in the Working Memory and Plasticity
Lab at the University of California, Irvine, as well as the Cogni-
tive Neuroimaging Lab at the University of Michigan.
Reprint requests should be sent to Jacky Au, University of
California, Irvine School of Education, 3200 Education Building,
Irvine CA 92697, or via e-mail: jwau@uci.edu.
Author Contributions
Jacky Au: Conceptualization; Formal analysis; Methodology;
Project administration; Writing—Original draft. Rachel N.
Smith-Peirce: Project administration; Writing—Review &
editing. Elena Carbone: Project administration; Writing—
Review & editing. Austin Moon: Project administration;
Writing—Review & editing. Michelle Evans: Project admin-
istration; Writing—Review & editing. John Jonides: Con-
ceptualization; Funding acquisition; Resources; Software;
Supervision; Writing—Review & editing. Susanne M. Jaeggi:
Conceptualization; Funding acquisition; Resources; Soft-
ware; Supervision; Writing—Review & editing.
to this work. No other authors declare any conflicts of inter-
ests or sources of funding.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .541, W/M = .213,
M/ W = .098, and W/ W = .148.
Funding Information
REFERENCES
This work was supported by The National Institutes of
Health, grant number: TL1TR001415 to J. A., The National
Institute on Aging (https://dx.doi.org/10.13039/100000049),
grant numbers: 1R01AG049006, 1K02AG054665 to S. M. J,
as well as the National Science Foundation (https://dx.doi
.org/10.13039/100000001), grant number: 1658268 to J. J. S.
M. J. has an indirect financial conflict of interest with
the MIND Research Institute, CA, whose interests are related
Agboada, D., Mosayebi-Samani, M., Kuo, M.-F., & Nitsche, M. A.
(2020). Induction of long-term potentiation-like plasticity in
the primary motor cortex with repeated anodal transcranial
direct current stimulation—Better effects with intensified
protocols? Brain Stimulation, 13, 987–997. https://doi.org/10
.1016/j.brs.2020.04.009, PubMed: 32325264
Alonzo, A., Brassil, J., Taylor, J. L., Martin, D., & Loo, C. K.
(2012). Daily transcranial direct current stimulation (tDCS)
leads to greater increases in cortical excitability than second
daily transcranial direct current stimulation. Brain
Au et al.
1033
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Stimulation, 5, 208–213. https://doi.org/10.1016/j.brs.2011.04
.006, PubMed: 22037139
Arciniega, H., Gözenman, F., Jones, K. T., Stephens, J. A., &
Berryhill, M. E. (2018). Frontoparietal tDCS benefits visual
working memory in older adults with low working memory
capacity. Frontiers in Aging Neuroscience, 10, 57. https://doi
.org/10.3389/fnagi.2018.00057, PubMed: 29593522
Au, J., Karsten, C., Buschkuehl, M., & Jaeggi, S. M. (2017).
Optimizing transcranial direct current stimulation protocols
to promote long-term learning. Journal of Cognitive
Enhancement, 1, 65–72. https://doi.org/10.1007/s41465-017
-0007-6
Au, J., Katz, B., Buschkuehl, M., Bunarjo, K., Senger, T., Zabel,
C., et al. (2016). Enhancing working memory training with
transcranial direct current stimulation. Journal of Cognitive
Neuroscience, 28, 1419–1432. https://doi.org/10.1162/jocn_a
_00979, PubMed: 27167403
Au, J., Katz, B., Moon, A., Talati, S., Abagis, T. R., Jonides, J.,
et al. (2021). Post-training stimulation of the right
dorsolateral prefrontal cortex impairs working memory
training performance. Journal of Neuroscience Research,
99, 2351–2363. https://doi.org/10.1002/jnr.24784, PubMed:
33438297
Bergmann, H. C., Rijpkema, M., Fernández, G., & Kessels, R. P. C.
(2012). Distinct neural correlates of associative working
memory and long-term memory encoding in the medial
temporal lobe. Neuroimage, 63, 989–997. https://doi.org/10
.1016/j.neuroimage.2012.03.047, PubMed: 22484305
Bird, H., Franklin, S., & Howard, D. (2001). Age of acquisition
and imageability ratings for a large set of words, including
verbs and function words. Behavior Research Methods,
Instruments, & Computers, 33, 73–79. https://doi.org/10.3758
/BF03195349, PubMed: 11296722
Blumenfeld, R. S. (2006). Dorsolateral prefrontal cortex promotes
long-term memory formation through its role in working
memory organization. Journal of Neuroscience, 26, 916–925.
https://doi.org/10.1523/JNEUROSCI.2353-05.2006, PubMed:
16421311
Blumenfeld, R. S., & Ranganath, C. (2007). Prefrontal cortex
and long-term memory encoding: An integrative review
of findings from neuropsychology and neuroimaging.
Neuroscientist, 13, 280–291. https://doi.org/10.1177
/1073858407299290, PubMed: 17519370
Borella, E., Cantarella, A., Joly, E., Ghisletta, P., Carbone, E.,
Coraluppi, D., et al. (2017). Performance-based everyday
functional competence measures across the adult lifespan:
The role of cognitive abilities. International Psychogeriatrics,
29, 2059–2069. https://doi.org/10.1017/S1041610217000680,
PubMed: 28595679
Brasil-Neto, J. P. (2012). Learning, memory, and transcranial
direct current stimulation. Frontiers in Psychiatry, 3. https://
doi.org/10.3389/fpsyt.2012.00080, PubMed: 22969734
Brown, R. M., & Robertson, E. M. (2007). Off-line processing:
Reciprocal interactions between declarative and procedural
memories. Journal of Neuroscience, 27, 10468–10475.
https://doi.org/10.1523/JNEUROSCI.2799-07.2007, PubMed:
17898218
Brysbaert, M., Warriner, A. B., & Kuperman, V. (2014).
Concreteness ratings for 40 thousand generally known English
word lemmas. Behavior Research Methods, 46, 904–911.
https://doi.org/10.3758/s13428-013-0403-5, PubMed: 24142837
Buch, E. R., Santarnecchi, E., Antal, A., Born, J., Celnik, P. A.,
Gerloff, C., et al. (2017). Effects of tDCS on motor learning
and memory formation: A consensus and critical position
paper. Clinical Neurophysiology, 128, 589–603. https://doi
.org/10.1016/j.clinph.2017.01.004, PubMed: 28231477
Buschkuehl, M., Jaeggi, S. M., Hutchison, S., Perrig-Chiello, P.,
Däpp, C., Müller, M., et al. (2008). Impact of working memory
training on memory performance in old-old adults. Psychology
and Aging, 23, 743–753. https://doi.org/10.1037/a0014342,
PubMed: 19140646
Cabeza, R., Dolcos, F., Graham, R., & Nyberg, L. (2002).
Similarities and differences in the neural correlates of
episodic memory retrieval and working memory. Neuroimage,
16, 317–330. https://doi.org/10.1006/nimg.2002.1063, PubMed:
12030819
Cepeda, N. J., Vul, E., Rohrer, D., Wixted, J. T., & Pashler, H.
(2008). Spacing effects in learning: A temporal ridgeline of
optimal retention. Psychological Science, 19, 1095–1102.
https://doi.org/10.1111/j.1467-9280.2008.02209.x, PubMed:
19076480
Chen, P.-C., Niknazar, H., Alaynick, W. A., Whitehurst, L. N., &
Mednick, S. C. (2021). Competitive dynamics underlie
cognitive improvements during sleep. Proceedings of the
National Academy of Sciences, U.S.A., 118. https://doi.org/10
.1073/pnas.2109339118, PubMed: 34903651
Cocco, S., Podda, M. V., & Grassi, C. (2018). Role of BDNF
signaling in memory enhancement induced by transcranial
direct current stimulation. Frontiers in Neuroscience, 12,
427. https://doi.org/10.3389/fnins.2018.00427, PubMed:
29997473
Crupi, D., Hulse, B. K., Peterson, M. J., Huber, R., Ansari, H.,
Coen, M., et al. (2009). Sleep-dependent improvement in
visuomotor learning: A causal role for slow waves. Sleep,
32, 1273–1284. https://doi.org/10.1093/sleep/32.10.1273,
PubMed: 19848357
Dedoncker, J., Brunoni, A. R., Baeken, C., & Vanderhasselt, M.-A.
(2016). The effect of the interval-between-sessions on
prefrontal transcranial direct current stimulation (tDCS)
on cognitive outcomes: A systematic review and
meta-analysis. Journal of Neural Transmission, 123,
1159–1172. https://doi.org/10.1007/s00702-016-1558-x,
PubMed: 27145765
Ferrarelli, F., Kaskie, R., Laxminarayan, S., Ramakrishnan, S.,
Reifman, J., & Germain, A. (2019). An increase in sleep slow
waves predicts better working memory performance in
healthy individuals. Neuroimage, 191, 1–9. https://doi.org/10
.1016/j.neuroimage.2019.02.020, PubMed: 30753924
Frase, L., Mertens, L., Krahl, A., Bhatia, K., Feige, B., Heinrich, S. P.,
et al. (2021). Transcranial direct current stimulation induces
long-term potentiation-like plasticity in the human visual
cortex. Translational Psychiatry, 11, 1–11. https://doi.org/10
.1038/s41398-020-01134-4, PubMed: 33414402
Galli, G., Vadillo, M. A., Sirota, M., Feurra, M., & Medvedeva, A.
(2019). A systematic review and meta-analysis of the effects of
transcranial direct current stimulation (tDCS) on episodic
memory. Brain Stimulation, 12, 231–241. https://doi.org/10
.1016/j.brs.2018.11.008, PubMed: 30503376
Gathercole, S. E., Dunning, D. L., Holmes, J., & Norris, D.
(2019). Working memory training involves learning new
skills. Journal of Memory and Language, 105, 19–42. https://
doi.org/10.1016/j.jml.2018.10.003, PubMed: 31235992
Goldsworthy, M. R., Pitcher, J. B., & Ridding, M. C. (2015).
Spaced noninvasive brain stimulation prospects for inducing
long-lasting human cortical plasticity. Neurorehabilitation
and Neural Repair, 29, 714–721. https://doi.org/10.1177
/1545968314562649, PubMed: 25505220
Gözenman, F., & Berryhill, M. E. (2016). Working memory
capacity differentially influences responses to tDCS and
HD-tDCS in a retro-cue task. Neuroscience Letters, 629,
105–109. https://doi.org/10.1016/j.neulet.2016.06.056,
PubMed: 27369325
Hannula, D. E., & Ranganath, C. (2008). Medial temporal lobe
activity predicts successful relational memory binding.
Journal of Neuroscience, 28, 116–124. https://doi.org/10
.1523/JNEUROSCI.3086-07.2008, PubMed: 18171929
1034
Journal of Cognitive Neuroscience
Volume 34, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hayes, A. F. (2018). Introduction to mediation, moderation,
and conditional process analysis: A regression-based
approach (2nd ed.). New York: Guilford Press.
Heinen, K., Sagliano, L., Candini, M., Husain, M., Cappelletti, M.,
& Zokaei, N. (2016). Cathodal transcranial direct current
stimulation over posterior parietal cortex enhances distinct
aspects of visual working memory. Neuropsychologia, 87,
35–42. https://doi.org/10.1016/j.neuropsychologia.2016.04
.028, PubMed: 27143222
Hill, A. T., Fitzgerald, P. B., & Hoy, K. E. (2016). Effects of anodal
transcranial direct current stimulation on working memory: A
systematic review and meta-analysis of findings from healthy
and neuropsychiatric populations. Brain Stimulation, 9,
197–208. https://doi.org/10.1016/j.brs.2015.10.006, PubMed:
26597929
Horvath, J. C., Forte, J. D., & Carter, O. (2015). Quantitative
review finds no evidence of cognitive effects in healthy
populations from single-session transcranial direct current
stimulation (tDCS). Brain Stimulation, 8, 535–550. https://
doi.org/10.1016/j.brs.2015.01.400, PubMed: 25701175
Hsu, T.-Y., Tseng, P., Liang, W.-K., Cheng, S.-K., & Juan, C.-H.
(2014). Transcranial direct current stimulation over right
posterior parietal cortex changes prestimulus alpha
oscillation in visual short-term memory task. Neuroimage,
98, 306–313. https://doi.org/10.1016/j.neuroimage.2014.04
.069, PubMed: 24807400
Huber, R., Felice Ghilardi, M., Massimini, M., & Tononi, G.
(2004). Local sleep and learning. Nature, 430, 78–81. https://
doi.org/10.1038/nature02663, PubMed: 15184907
Huo, L., Zhu, X., Zheng, Z., Ma, J., Ma, Z., Gui, W., et al. (2021).
Effects of transcranial direct current stimulation on episodic
memory in older adults: A meta-analysis. Journals of
Gerontology: Series B, 76, 692–702. https://doi.org/10.1093
/geronb/gbz130, PubMed: 31782505
Jaeggi, S. M., Buschkuehl, M., Parlett-Pelleriti, C. M., Moon, S. M.,
Evans, M., Kritzmacher, A., et al. (2020). Investigating the
effects of spacing on working memory training outcome:
A randomized, controlled, multisite trial in older adults.
Journals of Gerontology: Series B, 75, 1181–1192. https://doi
.org/10.1093/geronb/gbz090, PubMed: 31353413
Javadi, A. H., & Cheng, P. (2013). Transcranial direct current
stimulation (tDCS) enhances reconsolidation of long-term
memory. Brain Stimulation, 6, 668–674. https://doi.org/10
.1016/j.brs.2012.10.007, PubMed: 23137702
Jin, J., & Maren, S. (2015). Prefrontal-hippocampal interactions
in memory and emotion. Frontiers in Systems Neuroscience,
9, 170. https://doi.org/10.3389/fnsys.2015.00170, PubMed:
26696844
Jones, K. T., & Berryhill, M. E. (2012). Parietal contributions to
visual working memory depend on task difficulty. Frontiers
in Psychiatry, 3. https://doi.org/10.3389/fpsyt.2012.00081,
PubMed: 22973241
Jones, K. T., Gözenman, F., & Berryhill, M. E. (2015). The
strategy and motivational influences on the beneficial effect
of neurostimulation: A tDCS and fNIRS study. Neuroimage,
105, 238–247. https://doi.org/10.1016/j.neuroimage.2014.11
.012, PubMed: 25462798
Jones, K. T., Stephens, J. A., Alam, M., Bikson, M., & Berryhill,
M. E. (2015). Longitudinal neurostimulation in older adults
improves working memory. PLoS One, 10, e0121904. https://
doi.org/10.1371/journal.pone.0121904, PubMed: 25849358
Katz, B., Au, J., Buschkuehl, M., Abagis, T., Zabel, C., Jaeggi, S. M.,
et al. (2017). Individual differences and long-term consequences
of tDCS-augmented cognitive training. Journal of Cognitive
Neuroscience, 29, 1498–1508. https://doi.org/10.1162/jocn_a
_01115, PubMed: 28253083
Ke, Y., Wang, N., Du, J., Kong, L., Liu, S., Xu, M., et al. (2019).
The effects of transcranial direct current stimulation (tDCS)
on working memory training in healthy young adults.
Frontiers in Human Neuroscience, 13, 19. https://doi.org/10
.3389/fnhum.2019.00019, PubMed: 30774590
Krebs, C., Peter, J., Wyss, P., Brem, A.-K., & Klöppel, S. (2021).
Transcranial electrical stimulation improves cognitive training
effects in healthy elderly adults with low cognitive performance.
Clinical Neurophysiology, 132, 1254–1263. https://doi.org/10
.1016/j.clinph.2021.01.034, PubMed: 33875372
Kronberg, G., Bridi, M., Abel, T., Bikson, M., & Parra, L. C.
(2017). Direct current stimulation modulates LTP and LTD:
Activity dependence and dendritic effects. Brain Stimulation,
10, 51–58. https://doi.org/10.1016/j.brs.2016.10.001, PubMed:
28104085
Lau, E. Y. Y., Wong, M. L., Lau, K. N. T., Hui, F. W. Y., & Tseng,
C. (2015). Rapid-eye-movement-sleep (REM) associated
enhancement of working memory performance after a
daytime nap. PLoS One, 10. https://doi.org/10.1371/journal
.pone.0125752, PubMed: 25970511
Leshikar, E. D., Leach, R. C., McCurdy, M. P., Trumbo, M. C.,
Sklenar, A. M., Frankenstein, A. N., et al. (2017). Transcranial
direct current stimulation of dorsolateral prefrontal cortex
during encoding improves recall but not recognition
memory. Neuropsychologia, 106, 390–397. https://doi.org/10
.1016/j.neuropsychologia.2017.10.022, PubMed: 29056368
Looi, C. Y., Duta, M., Brem, A.-K., Huber, S., Nuerk, H.-C., &
Cohen Kadosh, R. (2016). Combining brain stimulation and
video game to promote long-term transfer of learning and
cognitive enhancement. Scientific Reports, 6, 22003. https://
doi.org/10.1038/srep22003, PubMed: 26902664
Määttä, S., Landsness, E., Sarasso, S., Ferrarelli, F., Ferreri, F.,
Ghilardi, M. F., et al. (2010). The effects of morning training
on night sleep: A behavioral and EEG study. Brain Research
Bulletin, 82, 118–123. https://doi.org/10.1016/j.brainresbull
.2010.01.006, PubMed: 20105456
Mancuso, L. E., Ilieva, I. P., Hamilton, R. H., & Farah, M. J.
(2016). Does transcranial direct current stimulation improve
healthy working memory? A meta-analytic review. Journal of
Cognitive Neuroscience, 28, 1063–1089. https://doi.org/10
.1162/jocn_a_00956, PubMed: 27054400
Martin, D. M., Liu, R., Alonzo, A., Green, M., Player, M. J.,
Sachdev, P., et al. (2013). Can transcranial direct current
stimulation enhance outcomes from cognitive training?
A randomized controlled trial in healthy participants.
International Journal of Neuropsychopharmacology, 16,
1927–1936. https://doi.org/10.1017/S1461145713000539,
PubMed: 23719048
Matzen, L. E., Trumbo, M. C., Leach, R. C., & Leshikar, E. D.
(2015). Effects of non-invasive brain stimulation on
associative memory. Brain Research, 1624, 286–296. https://
doi.org/10.1016/j.brainres.2015.07.036, PubMed: 26236022
McConathey, E. M., White, N. C., Gervits, F., Ash, S., Coslett, H. B.,
Grossman, M., et al. (2017). Baseline performance predicts
tDCS-mediated improvements in language symptoms
in primary progressive aphasia. Frontiers in Human
Neuroscience, 11, 347. https://doi.org/10.3389/fnhum.2017
.00347, PubMed: 28713256
McGillivray, S., & Castel, A. D. (2011). Betting on memory leads
to metacognitive improvement by younger and older adults.
Psychology and Aging, 26, 137–142. https://doi.org/10.1037
/a0022681, PubMed: 21417541
Melrose, R. J., Zahniser, E., Wilkins, S. S., Veliz, J., Hasratian, A. S.,
Sultzer, D. L., et al. (2020). Prefrontal working memory activity
predicts episodic memory performance: A neuroimaging
study. Behavioural Brain Research, 379, 112307. https://doi
.org/10.1016/j.bbr.2019.112307, PubMed: 31678217
Minichino, A., Bersani, F. S., Bernabei, L., Spagnoli, F., Vergnani,
L., Corrado, A., et al. (2015). Prefronto–cerebellar transcranial
direct current stimulation improves visuospatial memory,
Au et al.
1035
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
executive functions, and neurological soft signs in patients
with euthymic bipolar disorder. Neuropsychiatric Disease
and Treatment, 11, 2265–2270. https://doi.org/10.2147/NDT
.S79108, PubMed: 26356034
Pieters, R. (2017). Meaningful mediation analysis: Plausible
causal inference and informative communication. Journal of
Consumer Research, 44, 692–716. https://doi.org/10.1093/jcr
/ucx081
Miyake, A., & Shah, P. (Eds.). (1999). Models of working
Podda, M. V., Cocco, S., Mastrodonato, A., Fusco, S., Leone, L.,
memory: Mechanisms of active maintenance and executive
control. Cambridge University Press. https://doi.org/10.1017
/CBO9781139174909
Monte-Silva, K., Kuo, M.-F., Hessenthaler, S., Fresnoza, S.,
Liebetanz, D., Paulus, W., et al. (2013). Induction of late
LTP-like plasticity in the human motor cortex by repeated
non-invasive brain stimulation. Brain Stimulation, 6,
424–432. https://doi.org/10.1016/j.brs.2012.04.011, PubMed:
22695026
Monte-Silva, K., Kuo, M. F., Liebetanz, D., Paulus, W., & Nitsche,
M. A. (2010). Shaping the optimal repetition interval for
cathodal transcranial direct current stimulation (tDCS).
Journal of Neurophysiology, 103, 1735–1740. https://doi.org
/10.1152/jn.00924.2009, PubMed: 20107115
Nee, D. E., & Jonides, J. (2008). Neural correlates of access to
short-term memory. Proceedings of the National Academy
of Sciences, U.S.A., 105, 14228–14233. https://doi.org/10.1073
/pnas.0802081105, PubMed: 18757724
Nee, D. E., & Jonides, J. (2013). Trisecting representational
states in short-term memory. Frontiers in Human
Neuroscience, 7, 796. https://doi.org/10.3389/fnhum.2013
.00796, PubMed: 24324424
Nichols, E. A., Kao, Y.-C., Verfaellie, M., & Gabrieli, J. D. E.
(2006). Working memory and long-term memory for faces:
Evidence from fMRI and global amnesia for involvement of
the medial temporal lobes. Hippocampus, 16, 604–616.
https://doi.org/10.1002/hipo.20190, PubMed: 16770797
Nilsson, J., Lebedev, A. V., Rydström, A., & Lövdén, M. (2017).
Direct-current stimulation does little to improve the outcome
of working memory training in older adults. Psychological
Science, 28, 907–920. https://doi.org/10.1177
/0956797617698139, PubMed: 28509625
Oberauer, K. (2009). Design for a working memory. In B. H. Ross
(Ed.), Psychology of learning and motivation (Vol. 51,
pp. 45–100). Cambridge, MA: Academic Press. https://doi.org
/10.1016/S0079-7421(09)51002-X
Oberauer, K., Awh, E., & Sutterer, D. W. (2017). The role of
long-term memory in a test of visual working memory:
Proactive facilitation but no proactive interference. Journal
of Experimental Psychology: Learning, Memory, and
Cognition, 43, 1–22. https://doi.org/10.1037/xlm0000302,
PubMed: 27685018
O’Rourke, H. P., & MacKinnon, D. P. (2018). Reasons for testing
mediation in the absence of an intervention effect: A research
imperative in prevention and intervention research. Journal
of Studies on Alcohol and Drugs, 79, 171–181. https://doi.org
/10.15288/jsad.2018.79.171, PubMed: 29553343
Park, S.-H., Seo, J.-H., Kim, Y.-H., & Ko, M.-H. (2014). Long-term
effects of transcranial direct current stimulation combined
with computer-assisted cognitive training in healthy older
adults. NeuroReport, 25, 122–126. https://doi.org/10.1097
/ WNR.0000000000000080, PubMed: 24176927
Parlett-Pelleriti, C., Lin, G. C., Jones, M. R., Linstead, E., & Jaeggi,
S. M. (2019). Exploring age-related metamemory differences
using modified brier scores and hierarchical clustering. Open
Psychology, 1, 215–238. https://doi.org/10.1515/psych-2018
-0015, PubMed: 33693310
Perceval, G., Martin, A. K., Copland, D. A., Laine, M., & Meinzer,
M. (2020). Multisession transcranial direct current
stimulation facilitates verbal learning and memory
consolidation in young and older adults. Brain and
Language, 205, 104788. https://doi.org/10.1016/j.bandl.2020
.104788, PubMed: 32199339
Barbati, S. A., et al. (2016). Anodal transcranial direct
current stimulation boosts synaptic plasticity and memory
in mice via epigenetic regulation of Bdnf expression.
Scientific Reports, 6. https://doi.org/10.1038/srep22180,
PubMed: 26908001
Pugin, F., Metz, A. J., Wolf, M., Achermann, P., Jenni, O. G., &
Huber, R. (2015). Local increase of sleep slow wave activity
after three weeks of working memory training in children and
adolescents. Sleep, 38, 607–614. https://doi.org/10.5665/sleep
.4580, PubMed: 25669190
Ramaraju, S., Roula, M. A., & McCarthy, P. (2018). Modelling the
effect of electrode displacement on transcranial direct
current stimulation (tDCS). Journal of Neural Engineering,
15, 016019. https://doi.org/10.1088/1741-2552/aa8d8a,
PubMed: 28925375
Ranganath, C., & Blumenfeld, R. S. (2005). Doubts about
double dissociations between short- and long-term memory.
Trends in Cognitive Sciences, 9, 374–380. https://doi.org/10
.1016/j.tics.2005.06.009, PubMed: 16002324
Ranganath, C., Cohen, M. X., & Brozinsky, C. J. (2005). Working
memory maintenance contributes to long-term memory
formation: Neural and behavioral evidence. Journal of
Cognitive Neuroscience, 17, 994–1010. https://doi.org/10
.1162/0898929054475118, PubMed: 16102232
Ranieri, F., Podda, M. V., Riccardi, E., Frisullo, G., Dileone, M.,
Profice, P., et al. (2012). Modulation of LTP at rat
hippocampal CA3-CA1 synapses by direct current stimulation.
Journal of Neurophysiology, 107, 1868–1880. https://doi.org
/10.1152/jn.00319.2011, PubMed: 22236710
Reilly, J., & Kean, J. (2007). Formal distinctiveness of high- and
low-imageability nouns: Analyses and theoretical implications.
Cognitive Science, 31, 157–168. https://doi.org/10.1080
/03640210709336988, PubMed: 21635291
Richmond, L. L., Morrison, A. B., Chein, J. M., & Olson, I. R.
(2011). Working memory training and transfer in older
adults. Psychology and Aging, 26, 813–822. https://doi.org/10
.1037/a0023631, PubMed: 21707176
Richmond, L. L., Wolk, D., Chein, J., & Olson, I. R. (2014).
Transcranial direct current stimulation enhances verbal
working memory training performance over time and near
transfer outcomes. Journal of Cognitive Neuroscience, 26,
2443–2454. https://doi.org/10.1162/jocn_a_00657, PubMed:
24742190
Rohan, J. G., Carhuatanta, K. A., McInturf, S. M., Miklasevich, M. K.,
& Jankord, R. (2015). Modulating hippocampal plasticity
with in vivo brain stimulation. Journal of Neuroscience, 35,
12824–12832. https://doi.org/10.1523/JNEUROSCI.2376-15
.2015, PubMed: 26377469
Rucker, D. D., Preacher, K. J., Tormala, Z. L., & Petty, R. E.
(2011). Mediation analysis in social psychology: Current
practices and new recommendations: Mediation analysis
in social psychology. Social and Personality Psychology
Compass, 5, 359–371. https://doi.org/10.1111/j.1751-9004
.2011.00355.x
Rudebeck, S. R., Bor, D., Ormond, A., O’Reilly, J. X., & Lee, A. C. H.
(2012). A potential spatial working memory training task
to improve both episodic memory and fluid intelligence.
PLoS One, 7, e50431. https://doi.org/10.1371/journal.pone
.0050431, PubMed: 23209740
Ruf, S. P., Fallgatter, A. J., & Plewnia, C. (2017). Augmentation of
working memory training by transcranial direct current
stimulation (tDCS). Scientific Reports, 7, 876. https://doi.org
/10.1038/s41598-017-01055-1, PubMed: 28432349
1036
Journal of Cognitive Neuroscience
Volume 34, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Sali, A. W., & Egner, T. (2020). Declarative and procedural
Stephens, J. A., & Berryhill, M. E. (2016). Older adults improve
working memory updating processes are mutually facilitative.
Attention, Perception, & Psychophysics, 82, 1858–1871.
https://doi.org/10.3758/s13414-019-01887-1, PubMed:
31875313
Sandrini, M., Manenti, R., Brambilla, M., Cobelli, C., Cohen,
L. G., & Cotelli, M. (2016). Older adults get episodic memory
boosting from noninvasive stimulation of prefrontal cortex
during learning. Neurobiology of Aging, 39, 210–216. https://
doi.org/10.1016/j.neurobiolaging.2015.12.010, PubMed:
26923418
Sattari, N., Whitehurst, L. N., Ahmadi, M., & Mednick, S. C.
(2019). Does working memory improvement benefit from
sleep in older adults? Neurobiology of Sleep and Circadian
Rhythms, 6, 53–61. https://doi.org/10.1016/j.nbscr.2019.01
.001, PubMed: 31236520
Scogin, F., Rohen, N., & Bailey, E. (2000). Geriatric depression
scale. In Handbook of psychological assessment in primary
care settings (pp. 491–508). Erlbaum.
Sigurdsson, T., & Duvarci, S. (2016). Hippocampal-prefrontal
interactions in cognition, behavior and psychiatric disease.
Frontiers in Systems Neuroscience, 9, 190. https://doi.org/10
.3389/fnsys.2015.00190, PubMed: 26858612
Small, S. A. (2001). Age-related memory decline: Current
concepts and future directions. Archives of Neurology, 58,
360–364. https://doi.org/10.1001/archneur.58.3.360, PubMed:
11255438
Snodgrass, J. G., & Corwin, J. (1988). Pragmatics of
measuring recognition memory: Applications to dementia
and amnesia. Journal of Experimental Psychology: General,
117, 34–50. https://doi.org/10.1037/0096-3445.117.1.34,
PubMed: 2966230
Squire, L. R., Stark, C. E. L., & Clark, R. E. (2004). The medial
temporal lobe. Annual Review of Neuroscience, 27, 279–306.
https://doi.org/10.1146/annurev.neuro.27.070203.144130,
PubMed: 15217334
Stadthagen-Gonzalez, H., & Davis, C. J. (2006). The Bristol
norms for age of acquisition, imageability, and familiarity.
Behavior Research Methods, 38, 598–605. https://doi.org/10
.3758/BF03193891, PubMed: 17393830
on everyday tasks after working memory training and
neurostimulation. Brain Stimulation, 9, 553–559. https://doi
.org/10.1016/j.brs.2016.04.001, PubMed: 27178247
Sternberg, S. (1969). Memory-scanning: Mental processes
revealed by reaction-time experiments. American Scientist,
57, 421–457. PubMed: 5360276
Summers, J. J., Kang, N., & Cauraugh, J. H. (2016). Does
transcranial direct current stimulation enhance cognitive and
motor functions in the ageing brain? A systematic review and
meta- analysis. Ageing Research Reviews, 25, 42–54. https://
doi.org/10.1016/j.arr.2015.11.004, PubMed: 26607412
Tseng, P., Hsu, T.-Y., Chang, C.-F., Tzeng, O. J. L., Hung, D. L.,
Muggleton, N. G., et al. (2012). Unleashing potential:
Transcranial direct current stimulation over the right
posterior parietal cortex improves change detection in
low-performing individuals. Journal of Neuroscience, 32,
10554–10561. https://doi.org/10.1523/JNEUROSCI.0362-12
.2012, PubMed: 22855805
Unsworth, N. (2010). On the division of working memory and
long-term memory and their relation to intelligence: A latent
variable approach. Acta Psychologica, 134, 16–28. https://doi
.org/10.1016/j.actpsy.2009.11.010, PubMed: 20022311
van der Linden, M. (1998). The relationships between working
memory and long-term memory. Comptes Rendus de
l’Académie Des Sciences–Series III–Sciences de La Vie, 321,
175–177. https://doi.org/10.1016/S0764-4469(97)89818-6
Warriner, A. B., Kuperman, V., & Brysbaert, M. (2013). Norms of
valence, arousal, and dominance for 13,915 English lemmas.
Behavior Research Methods, 45, 1191–1207. https://doi.org
/10.3758/s13428-012-0314-x, PubMed: 23404613
Yonelinas, A. P. (2002). The nature of recollection and
familiarity: A review of 30 years of research. Journal of
Memory and Language, 46, 441–517. https://doi.org/10.1006
/jmla.2002.2864
Zinke, K., Noack, H., & Born, J. (2018). Sleep augments
training-induced improvement in working memory in
children and adults. Neurobiology of Learning and Memory,
147, 46–53. https://doi.org/10.1016/j.nlm.2017.11.009,
PubMed: 29175513
Au et al.
1037
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
6
1
0
1
5
2
0
1
9
2
3
2
/
/
j
o
c
n
_
a
_
0
1
8
3
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3