Early Top–Down Control of Visual Processing Predicts
Working Memory Performance
Aaron M. Rutman, Wesley C. Clapp, James Z. Chadick,
and Adam Gazzaley
Abstracto
■ Selective attention confers a behavioral benefit on both per-
ceptual and working memory ( W.M.) actuación, often attrib-
uted to top–down modulation of sensory neural processing.
Sin embargo, the direct relationship between early activity modu-
lation in sensory cortices during selective encoding and subse-
quent WM performance has not been established. To explore
the influence of selective attention on WM recognition, we used
electroencephalography to study the temporal dynamics of
top–down modulation in a selective, delayed-recognition para-
digm. Participants were presented with overlapped, “double-
exposed” images of faces and natural scenes, and were in-
structed to either remember the face or the scene while simulta-
neously ignoring the other stimulus. Aquí, we present evidence
that the degree to which participants modulate the early P100
(97–129 msec) event-related potential during selective stimulus
encoding significantly correlates with their subsequent WM rec-
ognition. These results contribute to our evolving understanding
of the mechanistic overlap between attention and memory. ■
INTRODUCCIÓN
Goal-directed selective attention influences the magni-
tude and speed of neural processing in cortical regions
where sensory information is actively represented, a través de
a process known as top–down modulation (Gazzaley,
Cooney, McEvoy, Caballero, & DʼEsposito, 2005; Kastner &
Ungerleider, 2000; Luck, Chelazzi, Hillyard, & Desimone,
1997; Desimone & Duncan, 1995). Many studies have
capitalized on the high temporal resolution of electro-
encephalography (EEG) to reveal early influences of top–
down control on visual processing in humans (Hillyard &
Anllo-Vento, 1998), and more recently to establish a direct
relationship between neural measures of modulation and
indicators of behavioral performance, such as the speed
of stimulus detection (Talsma, Mulckhuyse, Slagter, &
Theeuwes, 2007; Thut, Nietzel, Brandt, & Pascual Leone,
2006). Además, evidence has emerged that demon-
strates a mechanistic overlap between the processes of
selective attention and working memory ( W.M.). Several
studies have revealed a major role of WM in the control of
visual selective attention (Awh & En Jonia, 2001; de fockert,
rees, Frith, & La vida, 2001; Desimone, 1996), mientras
others have shown that selective attention is a key compo-
nent of WM (Awh & En Jonia, 2001). Recent studies utilizing
EEG have investigated the time course of attentional involve-
ment in WM, presenting a model in which attention is uti-
lized throughout the WM maintenance period (Sreenivasan,
Universidad de California, San Francisco
katz, & Jha, 2007; Jha, 2002), likely by biasing cortical
processing of relevant sensory representations and ac-
tivity modulation of distractors (Sreenivasan & Jha, 2007).
Although data have revealed that WM maintenance may de-
pend on temporally early attentional factors (Sreenivasan
et al., 2007), notably for distracting information (Zanto
& Gazzaley, 2009), a direct correlation between early neu-
ral measures of selective activity modulation during en-
coding and subsequent WM performance has not yet been
descrito.
Selective attention results in activity modulation at
very early stages of visual processing (Schoenfeld, Hopf,
Martinez, & Mai, 2007; Martinez et al., 2006; Khoe, mitchell,
Reynolds, & Hillyard, 2005; López, Rodríguez, & Valdés-
Sosa, 2004; Pinilla, Cobo, Torres, & Valdes-Sosa, 2001;
Valdes-Sosa, Bobes, Rodriguez, & Pinilla, 1998), incluido
amplitude modulations of the P100 (∼100 msec) y
N170 (∼170 msec) event-related potential (ERP) compo-
nents (see Hillyard & Anllo-Vento, 1998), which have been
localized to visual cortical areas in lateral extrastriate cor-
tex (Di Russo, Martínez, Sereno, Pitzalis, & Hillyard, 2002;
Gomez Gonzalez, clark, Admirador, Luck, & Hillyard, 1994). Nosotros
hypothesize that such early top–down modulation of corti-
cal activity reflects the fidelity of sensory representations of
relevant information in such a manner that it confers a be-
havioral benefit on maintaining that information in mind.
Here we explore how early markers of visual processing
that are modulated when attention is selectively directed
a complejo, real-world visual objects (es decir., human faces
or natural scenes) relate to subsequent WM recognition
© 2009 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 22:6, páginas. 1224–1234
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
6
mi
–
1
pag
2
d
2
F
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
oh
2
C
4
norte
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
5
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
t
F
.
.
/
.
oh
norte
1
8
METRO
a
y
2
0
2
1
actuación. Our study utilized a delayed recognition task
in which participants were instructed to remember two
estímulos (800 msec each) over the course of a 4-sec delay
período (Cifra 1). We used overlapping transparent
images of faces and scenes, with either the face or the
scene relevant (and the other irrelevant) for the WM task,
in a design similar to previous studies of object-based
atención (Furey et al., 2006; Hacer & Chun, 2005; serencias,
Schwarzbach, Courtney, Golay, & Yantis, 2004; OʼCraven,
Downing, & Kanwisher, 1999). Recording posterior EEG
measures while participants viewed the overlapped stimuli
during the encoding period (equivalent bottom–up input
with variations only in instructions) enabled us to evalu-
ate the timing of top–down modulation and correlate
these measures with recognition accuracy recorded after
the delay period.
MÉTODOS
Participantes
Nineteen healthy, right-handed individuals (edad media =
22.9 años; range = 18–34 years; 10 hombres) with normal
or corrected-to-normal vision volunteered, gave consent,
and were monetarily compensated to participate in the
estudiar. Participants were prescreened, and none used any
medication known to affect cognitive state.
Estímulos
The stimuli consisted of grayscale images of faces and
natural scenes. All face and scene stimuli were novel
across all tasks, across all runs, and across all trials of
the experiment. Images were 225 pixels wide and 300
pixels tall (14 × 18 cm), and were presented foveally,
subtending a visual angle of 3° from a small cross at the
center of the image. The face stimuli consisted of a vari-
ety of neutral-expression male and female faces across a
large age range. Hair and ears were removed digitally,
and a blur was applied along the contours of the face
as to remove any potential non-face-specific cues. El
sex of the face stimuli was held constant within each trial.
Images of scenes were not digitally modified beyond re-
sizing and gray-scaling. For the tasks consisting of over-
lapped faces and scenes, one face and one scene were
randomly paired, made transparent, and digitally over-
lapped using Adobe Photoshop CS2 such that both the
Cifra 1. Experimental
paradigma. Five different
tasks were presented in a
delayed-recognition task
diseño. All trials involved
viewing two images (Stim-1,
Stim-2) (encode), seguido
by a 4-sec period (delay),
and concluded with a third
imagen (probe). Encoding
stimuli in FM and SM
consisted of isolated pictures
of faces and natural scenes,
whereas encoding stimuli
in the three overlap tasks
(FM-O, PV-O, and SM-O)
consisted of an overlapped
scene and a face picture.
For the four memory tasks,
participants were instructed
to remember the relevant
encoding stimuli, maintain
the images in mind over the
delay period, and respond
with a button press whether
or not the probe image
matched one of the two
encoding period images. Para
the passive view task (PV-O),
participants were instructed
to relax and view the
double-exposed images
without trying to remember
a ellos, after which they
responded to the direction of
an arrow with a button press.
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
6
mi
–
1
pag
2
d
2
F
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
oh
2
C
4
norte
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
5
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
.
t
F
.
/
oh
norte
1
8
METRO
a
y
2
0
2
1
Rutman et al.
1225
face and the scene were equally visible. Overlapped and
isolated images were randomly assigned to the different
tareas.
Experimental Procedures
The experimental paradigm was comprised of five different
tasks in a delayed-recognition WM task design (Cifra 1).
Each task consisted of the same temporal sequence with
only the instructions differing across tasks. All tasks in-
volved viewing two images (Stim-1, Stim-2), each being
displayed for 800 mseg (with a 200-msec ISI). These im-
ages were followed by a 4-sec period (delay) en el cual
the images were to be held in mind (mentally rehearsed).
After the delay, a third image appeared (probe). The par-
ticipant was instructed to respond with a button press (como
quickly as possible without sacrificing accuracy) si
or not the probe image matched one of the previous two
images (Stim-1, Stim-2). This was followed by an intertrial
interval (ITI) lasting 4 segundo.
For three of the five tasks, the Stim-1 and Stim-2 images
were composed of both a scene and a face superimposed
upon each other. For these double-exposed images, el
participants were instructed to focus their attention on
and hold in mind either the face or the scene, while ignor-
ing the other. In the face memory-overlap task (FM-O), el
faces were held in mind while the scenes were ignored,
and vice versa in the scene memory-overlap task (SM-O).
When the probe image appeared, it was composed of an
isolated face in the FM-O task, or an isolated scene in the
SM-O task. For the passive view (PV-O) tarea, Participantes
were instructed to relax and view the double-exposed
images without trying to hold them in mind, después de lo cual
they responded to an arrow direction with a button press.
For the other two tasks, the Stim-1 and Stim-2 images were
each composed of a single stimulus without any distract-
ing information: a face in the face memory task (FM) y
a scene in the scene memory task (SM). The task was
presented in three separate runs, each run consisting
of each of the five task sets presented in blocks and
counterbalanced in random order across all participants.
Each task set consisted of a block of 20 trials of that task
(60 total trials per task condition for all 3 carreras, 120 total
encode period images). Each blocked task set was pre-
ceded by an instruction screen cueing the subject to
the specific memory goal of the task (es decir., “remember
the faces”).
Following the main experiment, participants performed
a surprise postexperiment recognition test in which they
viewed 320 nonoverlapped images, incluido 160 faces
y 160 escenas. Eighty of the faces and 80 of the scenes
were novel stimuli that were not included in the main ex-
perimento. Había 20 faces each from the FM, FM-O,
SM-O, and PV-O tasks, y 20 scenes each from the SM,
SM-O, FM-O, and PV-O tasks. No encoded stimulus was in-
cluded that was also a match during a trial of the main
experimento, so that no stimuli in the postexperiment test
were seen more than once before. All included face and
scene stimuli (both novel images and images from the ex-
perimento) were randomly ordered, and participants were
asked to rate their confidence of recognition of each im-
age as follows: 1 = definitely did not see the image during
the course of the experiment; 2 = think that the image
was not seen during the experiment; 3 = think that the
image was seen during the course of the experiment;
y 4 = definitely saw the image during the experiment.
An incidental long-term memory recognition index for
each stimulus was calculated by subtracting the rating of
novel stimuli for each participant.
Eye-movement Control Experiment
Eye tracking was performed on five participants (re-
cruited with the same exclusionary criteria) while they
performed the main experiment with identical instruc-
ciones. Data were collected on an ASL EYE-TRAC6 (Applied
Science Laboratories, Bedford, MAMÁ) sampled at 60 Hz. Eye
blinks were removed and data were high-pass filtered at
0.5 Hz using a fifth-order Butterworth filter to remove drift
using MATLAB (MathWorks, Natick, MAMÁ). Across-condition
time-series analysis was performed using paired t tests
with an uncorrected alpha value of .05. Analyses of var-
iance (ANOVAs) were calculated using a two-way repeated
measures ANOVA, and post hoc t tests were performed
for eye-position differences between conditions, using an
alpha value of .05 with Tukey–Kramer correction.
Electrophysiological Recordings
Neural data were recorded at 1024 Hz through a 24-bit
BioSemi ActiveTwo 64-channel Ag–AgCl active elec-
trode EEG acquisition system in conjunction with BioSemi
ActiView software (CortechSolutions, LLC, Wilmington,
CAROLINA DEL NORTE). Electrode offsets were maintained between ±20 mV.
Precise markers of stimulus presentation were acquired
using a photodiode. Trials with excessive peak-to-peak de-
flections, amplifier clipping, or excessive high-frequency
(EMG) activity were excluded prior to analysis.
Electrophysiological Data Analysis
Preprocessing was conducted through the EEGLAB toolbox
(Swartz Center for Computational Neuroscience, UCSD, La
Jolla, California) for MATLAB. Off-line, the raw EEG data were high-
pass filtered (0.5 Hz), referenced to an average reference,
and segmented into epochs beginning 200 msec before
stimulus onset and ending 800 msec after stimulus onset.
Single epochs were baseline-corrected using an average
from −200 to 0 msec before stimulus appearance. Eye move-
ments and artifacts were removed through an independent
1226
Revista de neurociencia cognitiva
Volumen 22, Número 6
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
6
mi
–
1
pag
2
d
2
F
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
oh
2
C
4
norte
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
5
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
/
F
t
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
component analysis by excluding components consistent
with topographies for blinks and eye movements and elec-
trooculogram time series. Artifact-free data epochs were
then split by task, filtered (1–30 Hz), and averaged, to cre-
ate stimulus-locked ERPs.
ERP amplitudes were then calculated as the area ±4 msec
from the peak latency. Across-participant ERP ANOVA and
t test statistics were calculated using amplitudes and la-
tencies from each participantʼs EOI.
Localizer Task
An independent functional localizer task was used to de-
fine electrodes of interest (EOIs) for each participant
(Liu, harris, & Kanwisher, 2002). The localizer task con-
sisted of a 1-back design in which participants attended
to seven blocks of 20 faces and seven blocks of 20 escenas.
Participants were instructed to attend to the stimuli and
to indicate when each 1-back match occurred by pressing
a button with both forefingers. Face and scene blocks
were randomly intermixed. Face and scene trials were
then segmented separately and averaged. Epochs to re-
peated stimuli were not included in the average in order
to prevent motor contamination in the ERP. The P100
component was identified at lateral posterior electrodes
as the first positive deflection appearing between 50 y
150 msec after stimulus onset. The N170 component
was identified at posterior sites as the maximal negative
pico entre 120 y 220 msec after stimulus onset.
As revealed in previous studies, we found a significant
preference for faces at both 100 mseg (Herrmann, Ehlis,
Ellgring, & Fallgatter, 2005; Liu et al., 2002), y 170 mseg
after stimulus onset (Herrmann et al., 2005; Liu et al.,
2002; Bentín, alison, Chip, Pérez, & McCarthy, 1996) en
components for all posterior-lateral electrodes, such that
they revealed significantly larger amplitudes for faces
versus scenes (electrodes P10, PO8, P8, O2, P9, PO7, P7,
O1; all p values < .02). The lateral posterior electrode that
showed the largest P100 and N170 amplitude difference
between faces and scenes was defined as that participantʼs
P100 EOI and N170 EOI, respectively. EOIs included the
following electrodes: P8, P10, PO4, PO8, O2, P7, P9,
PO7, and O1.
Event-related Potentials
Epochs from each task of the main experimental task were
separately segmented, baselined at −200 to 0 msec rela-
tive to stimulus onset, and then averaged. Only encoding-
period segments (Stim-1, Stim-2) from correct trials were
included. ERPs from each of the tasks included a mean
of 116 averaged epochs per participant per task (range
80–120). The peak of the P100 ERP component for each
posterolateral electrode was defined as the maximal posi-
tive voltage of the first positive deflection appearing be-
tween 50 and 150 msec after stimulus onset, whereas the
peak of the N170 component was defined as the maximal
negative voltage between 120 and 220 msec after stimulus
onset. After the peak was identified for each individual,
Statistical Analysis
Behavioral and ERP data were each subjected to a repeated
measures 2 × 2 ANOVA (with stimuli type and overlap as
factors) and checked against a normal distribution using
a Lilliefors test. Post hoc two-tailed t tests were corrected
for multiple comparisons using Tukeyʼs honestly signifi-
cant difference criterion and an alpha of .05. Time windows
for significant divergence of face and scene localizer data
were calculated using paired t tests for each time point.
These were not corrected for multiple comparisons under
the assumption that time-dependent measures are not
independent comparisons.
RESULTS
Behavioral Results
WM accuracy and response time (RT) data were subjected
to separate, repeated measures 2 × 2 ANOVA with the type
of stimulus attended (face vs. scene) and overlap status
(overlapped vs. nonoverlapped) as factors. WM accuracy
revealed a main effect of overlap [F(1, 18) = 55.05, p <
.0001], such that accuracy was significantly reduced in tasks
with overlapped stimuli relative to tasks with face and
scene stimuli presented in isolation (FM-O: 82.7% vs. FM:
89.5%, p < .01; SM-O: 83.9% vs. SM: 92.9%, p < .01; Fig-
ure 2A). This WM performance reduction for the over-
lapping stimuli was also evident as an increased RT for
overlap tasks [F(1, 18) = 15.09, p < .001] (FM-O: 1096 msec
vs. FM: 1055 msec, p = .09; SM-O: 1103 msec vs. SM:
1029 msec, p < .01; Figure 2B).
There was a main effect of stimulus for WM accuracy
[F(1, 18) = 4.8, p < .05], but no interaction between stim-
ulus and overlap [F(1, 18) = 1.17, p < .287]; post hoc
comparisons revealed that accuracy was reduced for faces
compared to scenes, only in the nonoverlapped tasks (SM:
92.9%, FM: 89.5%, p < .01). There was no main effect of
stimulus for RT, and no interaction between stimulus and
overlap for RT. Accuracy in the passive view (PV-O) task
was 99.3%; RTs to arrow direction averaged 593 msec.
Results of the surprise postexperiment recognition test
revealed that participants remembered the previously
0: nonoverlap = 0.58, SE =
seen stimuli in the long term (d
±0.08; relevant overlap = 0.39, SE = ±0.08; irrelevant
overlap = 0.35, SE = ±0.06). The recognition strength
reported by the participants (indexed by confidence rat-
ings 1 through 4) revealed that relevant stimuli from both
nonoverlapped and overlapped tasks were rated signifi-
cantly higher than irrelevant stimuli from overlapped tasks
( p < .05 and p < .05, respectively) and stimuli from the
Rutman et al.
1227
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
6
e
-
1
p
2
d
2
f
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
o
2
c
4
n
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
2
5
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
.
/
f
.
o
n
1
8
M
a
y
2
0
2
1
EEG Results
P100 Component
P100 peak latency and amplitude from posterior EOIs
were subjected to separate 2 × 2 ANOVA with the type
of stimulus attended (face vs. scene) and overlap status
(overlapped vs. nonoverlapped) as factors. P100 measures
of peak latency were not significantly different between
stimulus type or overlap [ANOVA: main effect of stimu-
lus, F(1, 18) = 0.86, p = .34; overlap, F(1, 18) = 0.21, p =
.66; mean latency across participants: FM, 110 msec; FM-O,
113 msec; PV-O, 113 msec; SM-O, 114 msec; SM, 115 msec;
all p > .17 for all two-tailed comparisons]. Sin embargo, cosa-
sures of P100 amplitude showed significant differences
[main effect of overlap: F(1, 18) = 11.36, pag < .005; main
effect of stimulus type: F(1, 18) = 32.28, p < .0001; and
an interaction between overlap and stimulus type: F(2,
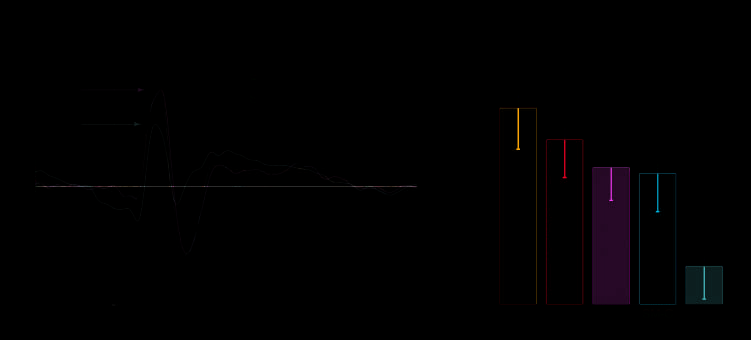
18) = 16.06, p < .001]. Post hoc comparisons revealed that
the amplitude of the P100 was significantly greater for the
FM task than for the SM task (FM vs. SM, p < .0001; all
participants exhibited greater P100 amplitude in FM vs.
SM) (Figure 3A and B), revealing a differential response
in the P100 component for faces compared to scenes, as re-
ported by others (i.e., bottom–up effect) (Herrmann et al.,
2005; Liu et al., 2002). Importantly, we report that for spa-
tially overlapped images of faces and scenes with equivalent
bottom–up information, attention to one stimulus while
ignoring the other resulted in significant attentional modu-
lation at this early time point in visual processing (i.e., top–
down effect) (FM-O vs. SM-O, p < .01; 15 of 19 participants
exhibited greater P100 amplitude in FM-O vs. SM-O) (Fig-
ure 3A and B). The P100 component of the FM-O task was
significantly different from the SM-O task at 97–129 msec
(paired two-tailed t tests across time points, p < .05). P100
amplitude in the FM-O task was closer to that of the FM
task, whereas the P100 amplitude in the SM-O task was
closer to that of the SM task (FM vs. FM-O, p = .10; SM-O
vs. SM, p < .01). Although P100 amplitude in the passive
view task (PV-O) was between FM-O and SM-O, it was not
significantly different from either overlap task (PV-O vs.
FM-O, p = .11; PV vs. SM-O, p = .72).
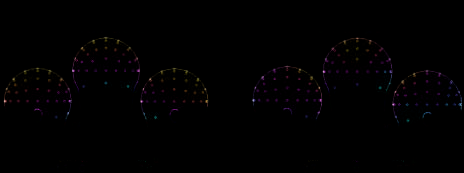
Topography maps of the P100 difference between pairs
of tasks are shown in Figure 4. The lateralized posterior
topography of the nonoverlapped face and scene differ-
ence (FM vs. SM: bottom–up contrast) is comparable to
the overlapped face and scene difference (FM-O vs. SM-O:
top–down contrast), revealing that top–down modulation
occurs in approximately the same visual cortical regions
that distinguish the stimuli based on bottom–up stimulus-
driven differences.
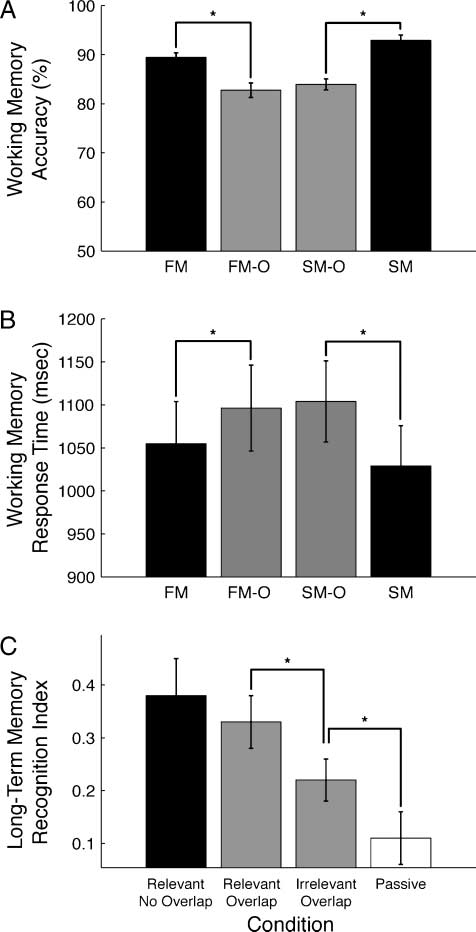
Figure 2. Behavioral results. (A) WM accuracy. Tasks utilizing
overlapped images showed significantly reduced WM recognition
accuracy. (B) WM RT. Overlapped tasks showed significant increases
in RT relative to the nonoverlapped task counterparts. (C) Long-term
memory recognition index. A postexperiment recognition test revealed
significantly better recognition of relevant images in the overlap
tasks (faces in FM-O and scenes in SM-O) than irrelevant images
from the overlap tasks (faces from SM-O and scenes from FM-O),
as well as images from the passive view task (PV-O). Error bars
represent standard error of the mean. Asterisks denote significant
differences ( p < .05). FM-O = face memory-overlap; SM-O =
scene memory-overlap; FM = face memory; SM = scene memory.
passive view task ( p < .01 and p < .01 respectively) (Fig-
ure 2C). These data confirm that participants were per-
forming the experiment as instructed, such that they were
selectively directing their attention to the relevant stimuli
and ignoring the irrelevant stimuli.
N170 Component
An ANOVA showed a significant effect of overlap and
stimulus type for N170 latency [main effect of stimulus
type: F(1, 18) = 10.93, p < .005; main effect of overlap:
F(1, 18) = 21.97, p < .0005]. Post hoc t tests revealed
1228
Journal of Cognitive Neuroscience
Volume 22, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
6
e
-
1
p
2
d
2
f
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
o
2
c
4
n
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
2
5
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
/
t
f
.
o
n
1
8
M
a
y
2
0
2
1
Figure 3. Top–down
modulation of the P1
component. (A) Grand-average
waveform of P1 EOIs (n = 19).
(B) P100 peak amplitudes
(n = 19). All peak amplitudes of
memory tasks show significant
differences across tasks (PV-O
is not significantly different
than FM-O or SM-O). Error
bars represent standard
error of the mean. Asterisks
denote significant difference
(single, p < .05; double,
p < .01; triple, p < .0001).
FM-O = face memory-overlap;
SM-O = scene memory-overlap;
PV-O = passive view-overlap;
FM = face memory;
SM = scene memory.
that the mean N170 latencies significantly differ between
isolated faces and scenes (FM, 174 msec vs. SM, 157 msec,
p < .01), but were not significantly different for over-
lapped tasks (FM-O, 184 msec vs. SM-O, 176 msec, p =
.13). The N170 peaked significantly later in the presence
of distraction (FM-O later than FM, p < .01; SM-O later
than SM, p < .01). However, there was no interaction be-
tween stimulus type and overlap [F(1, 1) = 0.73, p = .40].
Analysis of N170 amplitude reveals the classic finding
of face selectivity (N170 face-selective effect; Bentin et al.,
1996), with an ANOVA across tasks showing a main effect
of stimulus type [F(1, 18) = 13.9, p < .005], and post hoc
t tests revealing a significantly more negative N170 com-
ponent for isolated faces than scenes (FM vs. SM, p <
.01). However, the N170 amplitude was not modulated
by top–down attention in this experiment; that is, N170
amplitudes in the overlapped tasks were not significantly
different from each other [main effect of overlap: F(1,
18) = 0.31, p = .58; FM-O vs. SM-O, p = .81]. N170 am-
plitude in the PV-O task was not significantly different
from the other overlap tasks (vs. FM-O, p = .58; vs. SM-O,
p = .23).
Neural–Behavioral Correlations
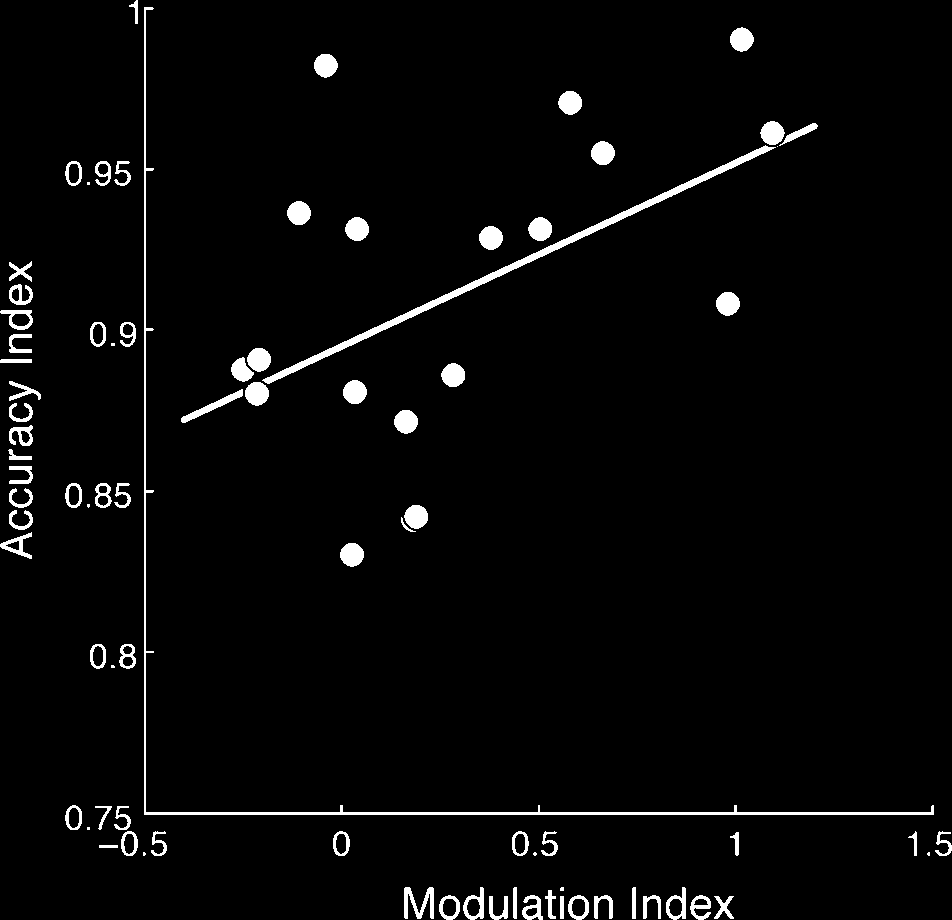
We report a significant across-participant correlation be-
tween the P100 modulation index and WM accuracy in-
dex (r = .45, p < .05; Figure 5); that is, the degree to
which a participant selectively modulates activity in the
first 100 msec of encoding a stimulus is a significant pre-
dictor of their ability to accurately recognize the stimulus
after a 4-sec delay. We found this critical correlation by
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
6
e
-
1
p
2
d
2
f
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
o
2
c
4
n
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
2
5
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
.
f
t
.
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Topographic ERP difference maps at 95–130 msec (P100
component). (A) The lateralized posterior scalp topography of the
nonoverlapped face and scene difference (FM minus SM: bottom–up
contrast) is comparable to (B) the topography of the overlapped face
and scene difference (FM-O minus SM-O: top–down contrast).
Figure 5. Neural–behavioral correlation. Measures of attentional
modulation (P1 modulation index) significantly correlate with WM
recognition (accuracy index). Subjects with greater attentional
modulation of P100 amplitude (∼100 msec poststimulus presentation)
show greater ability to subsequently remember encoded stimuli
after a delay period of WM maintenance (4 sec poststimulus
presentation), r = .45, p < .05.
Rutman et al.
1229
developing indices that allowed for a comparison of neu-
ral activity and behavior. First, to generate an index of
top–down modulation of the P100 amplitude, we com-
puted a P100 modulation index for each participant as
the difference in P100 amplitude in the overlap tasks,
corrected by the difference in P100 amplitude in non-
overlap tasks:
Modulation index ¼ FM-Oamp (cid:1) SM-Oamp
FMamp (cid:1) SMamp
This index allowed us to normalize for individual differences
in bottom–up sensory processing. Second, to generate an
index of WM recognition performance, we computed a
WM accuracy index for each participant, composed of the
participantʼs average accuracy in overlap tasks, corrected
by their average accuracy in nonoverlap tasks:
Accuracy index ¼
ðFM-OACC þ SM-OACCÞ=2
ðFMACC þ SMACCÞ=2
This index allowed us to normalize for individual differ-
ences in WM abilities. There was no comparable finding
of significant correlation between P100 modulation and
long-term memory measures, perhaps due to sparse sam-
pling of long-term memory measures.
Eye-movement Control
To investigate the possibility that a condition-dependent
shift in eye position either before or within 100 msec
after stimulus presentation may have resulted in the
reported P1 effect (as opposed to covert selective at-
tention), we performed an additional experiment with
eye tracking alone under identical conditions and instruc-
tions to the EEG experiment. Analysis revealed that there
were no condition-specific differences in eye position at
any time point. Furthermore, the median eye position
prior to stimulus onset (−200 to 0 msec) and immedi-
ately after stimulus onset (0 to 100 msec) showed no
dependence on condition in the vertical or horizontal di-
rections [two-way repeated measures ANOVA—vertical-
pre: F(3, 4) = 1.35, p = .26; vertical-post: F(3, 4) =
2.17, p = .09; horizontal-pre: F(3, 4) = 2.08, p = .11;
horizontal-post: F(3, 4) = 0.09, p = .96; post hoc
t tests—vertical-pre: FM vs. SM, p = .85; FM-O vs. SM-O,
p = .59; vertical-post: FM vs. SM, p = .28; FM-O vs. SM-O,
p = .49; horizontal-pre: FM vs. SM, p = .41; FM-O vs.
SM-O, p = .79; horizontal-post: FM vs. SM, p = .99;
FM-O vs. SM-O, p = .97]. In addition, measures of WM ac-
curacy for each participant in the eye-tracking experiment
were within 2 standard deviations of the mean WM accu-
racy measures for participants in the main experiment.
Although this experiment cannot definitively demon-
strate that eye position was not an influence on the re-
ported P1 effect and behavioral correlation (because
eye-tracking data were not obtained for the EEG ses-
sions), these results reveal that participants do not seem
to rely on a consistent and differential shift in eye gaze
to perform the experiment. Furthermore, reports from
participants in the EEG experiment do not suggest that
a strategy of fixating their eyes at a particular location
was utilized (e.g., repositioning gaze above the center of
the screen prior to stimulus onset to more easily detect
featural information from the faces, such as the eyes).
DISCUSSION
This study investigated top–down modulation of early vi-
sual processing and the influence of such modulation on
subsequent WM recognition performance. We capitalized
on the presence of well-described EEG signal differences
associated with bottom–up processing of isolated face
and scene stimuli (Herrmann et al., 2005; Liu et al., 2002;
Bentin et al., 1996) to explore attentional influences on
sensory cortical processing in the context of interfering
information (i.e., overlapped stimuli). By maintaining
bottom–up, sensory information constant and manipulat-
ing task goals, we were able to isolate the influence of
top–down modulation on visual processing. We found that
significant modulation of visual cortical activity begins as
early as 97 msec after stimulus presentation (P100 com-
ponent). Importantly, we found that at this early time point
the extent to which participants selectively modulate neu-
ral representations of task-relevant information, when
distracted by irrelevant information, correlates with their
ability to successfully recognize the relevant stimuli after
a period of WM maintenance. This provides a direct cor-
relative link between neural activity in early visual cortex
during selective encoding and behavioral measures of
WM performance.
Early Visual Cortex Modulation
Modulation of early ERP components have been well doc-
umented during covert spatial-based attention (Hillyard,
Vogel, & Luck, 1998), and more recently in feature-based
attention tasks (Schoenfeld et al., 2007). In contrast to
spatial- and feature-based attention, object-based attention
involves the integration of spatial and feature aspects of
an object to yield a holistic representation. In the current
study, the use of spatially superimposed faces and scenes
minimizes spatial-based mechanisms (Furey et al., 2006;
Yi & Chun, 2005; Serences et al., 2004; OʼCraven et al.,
1999), and the task goals of successfully recognizing the
relevant object after a delay period reduces reliance solely
on feature information. Although the task design in the
current study minimizes both spatial- and feature-based
attentional mechanisms, there may still be an influence of
feature and spatial information during WM encoding. For
example, a shift in covert spatial attention to an anticipated
location, such as that containing salient facial features,
may occur during or prior to the cue period, although
1230
Journal of Cognitive Neuroscience
Volume 22, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
6
e
-
1
p
2
d
2
f
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
o
2
c
4
n
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
2
5
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
none of the participants reported relying on a consistent
feature or spatial strategy. Moreover, the eye-tracking
control experiment revealed that overt eye movements
were not likely a confounding factor in the reported neural
results.
We report significant modulation of the P100 compo-
nent in a selective attention task for complex real-world
objects. This finding is consistent with several previously
published reports of object-based attention, but is at odds
with others. Object-based studies using illusory surface
paradigms have documented significant modulation of
the P100 (Valdes-Sosa et al., 1998), and even the earlier
C1 component (Khoe et al., 2005; Valdes-Sosa et al.,
1998). However, some studies have found modulatory
changes that begin slightly later in the time course of visual
processing, at the N170 component, ∼170 msec (Martinez,
Ramanathan, Foxe, Javitt, & Hillyard, 2007; Martinez,
Teder-Salejarvi, Vazquez, et al., 2006; He, Fan, Zhou, &
Chen, 2004; Pinilla et al., 2001); these studies utilized
either the discrimination of illusory surfaces defined by
transparent motion or the detection of luminance/shape
changes at one end of an object.
Also, the current findings are in contrast to the results of
an MEG study that utilized similar stimuli (superimposed
faces and houses), but in a 1-back repetition detection
task. This study showed modulation only at later time
points (>190 mseg) (Furey et al., 2006). Our results may
have revealed earlier modulation due to greater task de-
mands imposed by a two-item delayed-recognition task;
it has been shown that increasing task difficulty results
in enhanced activity modulation (Spitzer, Desimone, &
Moran, 1988).
It is important to note that unlike several other EEG
studies that did not find P100 selectivity for faces, nosotros
observed a P100 amplitude preference for faces versus
scenes both in the main experiment and in an inde-
pendent localizer task where faces and scenes were pre-
sented in separate blocks. Although the current study
and several others (Herrmann et al., 2005; Itier & taylor,
2002; Linkenkaer-Hansen et al., 1998) have revealed
P100 selectivity to faces, others that have used face stim-
uli have found the P100 to reflect more domain-general
aspects of visual processing (rossión, Joyce, Cottrell, &
Tarr, 2003; Rossion et al., 1999). Although all P100 find-
ings likely represent early visual processing, it is possible
that our results and those of studies that did not reveal
P100 face selectivity may not reflect exactly the same type
of processing, potentially as a result of differences in
task design. Sin embargo, the current study was intended
to capitalize on the observed face selectivity of the P100
in the functional localizer task only to serve as an early
marker of attentional control processes.
In a recently published study, we utilized face and scene
stimuli in a similar two-item delayed-recognition task, pero
instead of using simultaneously presented overlapped
estímulos, the face and scene images were presented sequen-
inicialmente, without overlap (Gazzaley et al., 2005). Curiosamente,
the study revealed significant N170 modulation, pero no
significant P100 amplitude modulation by attentional goals.
Sin embargo, we recently increased the number of research
participants in the sequential design version of this task
and revealed significant top–down modulation of the P100
amplitude for sequentially presented relevant versus irrele-
vant faces (Gazzaley et al., 2008), thus paralleling the cur-
rent study findings of very early object-based modulation.
Because it has been postulated that early bottom–up face
processing is rapid and largely automatic (Heisz, Cual, &
derramado, 2006), it is especially significant that top–down
modulation can occur at such an early phase in processing
these stimuli.
Several studies have suggested that early face process-
En g (P100/M100 component) is a reflection of face cate-
gorization/holistic perception (Itier & taylor, 2004; Liu
et al., 2002), whereas later processing (N170 component)
reflects configural information of faces (Latinus & taylor,
2006; Goffaux, Gauthier, & rossión, 2003; Liu et al., 2002;
Rossion et al., 2000). En ese caso, it follows that P100 modula-
tion observed in the overlap tasks might represent early
successful categorization of a face as being distinct from
a scene, perhaps based on low-level feature analysis
(Latinus & taylor, 2006). Sin embargo, this raises the ques-
tion as to why the N170 component was not modulated
by attention in the current study (es decir., no significant dif-
ference between FM-O and SM-O). One potential reason
is based on previous findings that configural face process-
ing requires extraction of low spatial frequency (LSF)
información (Goffaux, Hault, Michel, Vuong, & rossión,
2005). En el estudio actual, the application of a trans-
parency filter and an overlapped image obscures LSF in-
formación, while largely preserving high spatial frequency
(HSF) información. When Goffaux et al. (2003) aplicado
a filter to face stimuli that eliminated LSF and retained
HSF information, face-selective N170 perceptual effects
were abolished. It is thus possible that the bottom–up
perceptual modifications to the faces, introduced by our
experimental design, resulted in less LSF information and
interfered with top–down influences at this stage. En
support of this notion, it has also been revealed that the
projection of LSF information to prefrontal cortex influ-
ences top–down modulation of visual cortical areas at
∼180 msec (Bar et al., 2006). This may thus explain why
the present results differ from those previously reported
using the sequential version of this paradigm, eso es, el
same task with preserved LSF information resulted in
significant top–down modulation of the N170 (Gazzaley
et al., 2005).
Sin embargo, this explanation does not account for the fact
that several studies in which LSF information was present
have also not revealed N170 modulation as a function of
atención (Carmel & Bentín, 2002; Cauquil, Edmonds, &
taylor, 2000). It is possible that N170 modulation was
not observed in the current study and these other studies
because the salience of face stimuli was already too high
to benefit from additional perceptual modulation at this
Rutman et al.
1231
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
6
mi
–
1
pag
2
d
2
F
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
oh
2
C
4
norte
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
5
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
.
t
/
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
stage of encoding. En efecto, it has been argued that relative
to stimuli with high salience, stimuli with low salience are
more likely to benefit from additional attentional modula-
ción (Hawkins, Shafto, & Richardson, 1988).
In considering how activity modulation can occur so
early in the processing of the overlapped visual stimuli
(es decir., 100 msec after stimulus presentation), it is important
to recognize that participants were cued to the relevant
información, such that they were aware of the stimulus
to be remembered prior to presentation. This aspect of
the current study parallels that used in most spatial at-
tention tasks, which also report modulation of the P100
amplitude. En otras palabras, anticipatory gain modula-
tion may preactivate sensory cortical areas to enhance
the efficiency of subsequent sensory processing, as de-
scribed by others (Kastner, Pinsk, De Weerd, Desimone,
& Ungerleider, 1999; Luck et al., 1997).
Neural–Behavioral Correlation
It is well established that selective attention confers a
behavioral performance advantage for a variety of per-
ceptual tasks, such as visual detection (posner, Snyder, &
Davidson, 1980), discriminación (Carrasco & McElree,
2001), and categorization (Heekeren, Marrett, Bandettini,
& Ungerleider, 2004). In a comparable manner, failure to
selectively direct attentional resources negatively impacts
memory performance in both young (Zanto & Gazzaley,
2009) and older adults (Gazzaley et al., 2008; Gazzaley,
Cooney, Rissman, & DʼEsposito, 2005). The behavioral
advantage mediated by selective attention is presumably
the result of reduced interference from irrelevant infor-
mation in a system with limited capacity (Hasher, Lustig,
& zacks, 2008; Vogel, McCollough, & Machizawa, 2005),
likely mediated via top–down control mechanisms origi-
nating from prefrontal cortex (para una revisión, see Gazzaley
& DʼEsposito, 2007). Sin embargo, only recently have direct
correlations between the magnitude of visual cortex ac-
tivity modulation and behavioral measures of perceptual
and memory performance been established (Gazzaley,
Cooney, Rissman, et al., 2005; Vogel & Machizawa, 2004;
Persona, Kastner, & Ungerleider, 2002; rees, Friston, &
Koch, 2000; Cervecero, zhao, Desmond, guantero, & Gabrieli,
1998).
By revealing a significant correlation between very
early measures of visual cortex activity during selective
stimulus encoding and subsequent WM recognition ac-
curacy, our results contribute to a growing literature
describing the relationship between visual activity mod-
ulation and behavioral performance. Específicamente, the de-
gree to which participants modulate the P100 amplitude
in overlap tasks predicts their subsequent recognition
exactitud. This finding suggests that robust and early mod-
ulation generates higher fidelity stimulus representations,
which translates to improved maintenance of relevant in-
formation across a delay period, resulting in superior
recognition ability.
Conclusión
Consistency of goal-directed activity modulation occurring
so early in the processing of spatial-, feature- and object-
based information suggests that domain-general mech-
anisms of top–down modulation are targeted on early
cortical regions of the visual processing stream. The in-
fluence of such early top–down modulation of neural re-
presentations for real-world objects on WM recognition
performance is consistent with a growing appreciation of
the dynamic relationship of attention and WM (Awh &
En Jonia, 2001; de Fockert et al., 2001).
Expresiones de gratitud
This work was supported by National Institutes of Heath Grant
K08-AG025221, R01-AG030395, and the American Federation of
Aging Research (AFAR). We thank Nick Planet and Derek Wu for
their assistance in EEG data acquisition and pre-processing.
Reprint requests should be sent to Adam Gazzaley, Universidad de
California, San Francisco, 600 16th Street, Genentech Hall, Room
N472J, San Francisco, California 94158, o por correo electrónico: adam.gazzaley@
ucsf.edu.
REFERENCIAS
Awh, MI., & En Jonia, j. (2001). Overlapping mechanisms of
attention and spatial working memory. Tendencias en
Cognitive Sciences, 5, 119–126.
Bar, METRO., Kassam, k. S., Ghuman, A. S., Boshyan, J., Schmid,
A. METRO., Valle, A. METRO., et al. (2006). Top–down facilitation of
visual recognition. Proceedings of the National Academy
of Sciences, EE.UU., 103, 449–454.
Bentín, S., alison, T., Chip, A., Pérez, MI., & McCarthy, GRAMO.
(1996). Electrophysiological studies of face perception in
humanos. Revista de neurociencia cognitiva, 8, 551–565.
Cervecero, j. B., zhao, Z., Desmond, j. MI., guantero, GRAMO. h., &
Gabrieli, j. D. (1998). Making memories: Brain activity that
predicts how well visual experience will be remembered.
Ciencia, 281, 1185.
Carmel, D., & Bentín, S. (2002). Domain specificity versus
expertise: Factors influencing distinct processing of faces.
Cognición, 83, 1–29.
Carrasco, METRO., & McElree, B. (2001). Covert attention
accelerates the rate of visual information processing.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
98, 5363–5367.
Cauquil, A. S., Edmonds, GRAMO. MI., & taylor, METRO. j. (2000). Es
the face-sensitive N170 the only ERP not affected by
selective attention? NeuroReport, 11, 2167–2171.
de fockert, j. w., rees, GRAMO., Frith, C. D., & La vida, norte. (2001).
The role of working memory in visual selective attention.
Ciencia, 291, 1803–1806.
Desimone, R. (1996). Neural mechanisms for visual memory
and their role in attention. Actas del Nacional
Academia de Ciencias, EE.UU., 93, 13494–13499.
Desimone, r., & Duncan, j. (1995). Neural mechanisms
of selective visual attention. Annual Review of
Neurociencia, 18, 193–222.
Di Russo, F., Martínez, A., Sereno, METRO., Pitzalis, S., & Hillyard, S.
(2002). Cortical sources of the early components of the
visual evoked potential. Mapeo del cerebro humano, 15, 95–111.
Furey, METRO. l., Tanskanen, T., Beauchamp, METRO. S., Avikainen, S.,
Uutela, K., Hari, r., et al. (2006). Dissociation of
1232
Revista de neurociencia cognitiva
Volumen 22, Número 6
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
6
mi
–
1
pag
2
d
2
F
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
oh
2
C
4
norte
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
5
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
.
.
t
F
/
oh
norte
1
8
METRO
a
y
2
0
2
1
face-selective cortical responses by attention. Actas
of the National Academy of Sciences, EE.UU., 103,
1065–1070.
Gazzaley, A., Clapp, w., kelly, J., McEvoy, K., Caballero, R. T.,
& DʼEsposito, METRO. (2008). Age-related top–down
suppression deficit in the early stages of cortical
visual memory processing. Actas del Nacional
Academia de Ciencias, EE.UU., 105, 13122–13126.
Gazzaley, A., Cooney, j. w., McEvoy, K., Caballero, R. T., &
DʼEsposito, METRO. (2005). Top–down enhancement and
suppression of the magnitude and speed of neural
actividad. Revista de neurociencia cognitiva, 17, 507–517.
Gazzaley, A., Cooney, j. w., Rissman, J., & DʼEsposito, METRO.
(2005). Top–down suppression deficit underlies
working memory impairment in normal aging.
Neurociencia de la naturaleza, 8, 1298–1300.
Gazzaley, A., & DʼEsposito, METRO. (2007). Unifying prefrontal
cortex function: Executive control, neural networks and
top–down modulation. In J. Cummings & B. Molinero (Editores.),
The human frontal lobes (2y ed., páginas. 187–206). Nueva York:
The Guildford Press.
Goffaux, v., Gauthier, I., & rossión, B. (2003). Spatial scale
contribution to early visual differences between face
and object processing. Cognitive Brain Research, 16,
416–424.
Goffaux, v., Hault, B., Michel, C., Vuong, q. C., & rossión, B.
(2005). The respective role of low and high spatial
frequencies in supporting configural and featural
processing of faces. Percepción, 34, 77–86.
Gomez Gonzalez, C. METRO., clark, V. PAG., Admirador, S., Luck, S. J., &
Hillyard, S. A. (1994). Sources of attention-sensitive visual
event-related potentials. Brain Topography, 7, 41–51.
Hasher, l., Lustig, C., & zacks, j. METRO. (2008). Inhibitory
mechanisms and the control of attention. In A. Conway, C.
Jarrold, METRO. kane, A. Miyake, & j. Towse (Editores.), Variation
in working memory (páginas. 227–249). Nueva York: Oxford
Prensa universitaria.
Hawkins, h. l., Shafto, METRO. GRAMO., & Richardson, k. (1988).
Effects of target luminance and cue validity on the latency
of visual detection. Percepción & Psicofísica, 44,
484–492.
Él, X., Admirador, S., zhou, K., & Chen, l. (2004). Cue validity and
object-based attention. Revista de neurociencia cognitiva,
16, 1085–1097.
Heekeren, h. r., Marrett, S., Bandettini, PAG. A., & Ungerleider,
l. GRAMO. (2004). A general mechanism for perceptual
decision-making in the human brain. Naturaleza, 431,
859–862.
Heisz, j. J., Cual, S., & derramado, j. METRO. (2006). Progressive
N170 habituation to unattended repeated faces. Vision
Investigación, 46, 47–56.
Herrmann, METRO. J., Ehlis, A. C., Ellgring, h., & Fallgatter, A. j.
(2005). Early stages (P100) of face perception in humans
as measured with event-related potentials (ERPs). Diario
of Neural Transmission, 112, 1073–1081.
Hillyard, S. A., & Anllo-Vento, l. (1998). Event-related brain
potentials in the study of visual selective attention.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
95, 781–787.
Hillyard, S. A., Vogel, mi. K., & Luck, S. j. (1998). Sensory
gain control (amplification) as a mechanism of selective
atención: Electrophysiological and neuroimaging
evidencia. Philosophical Transactions of the Royal Society
of London, Serie B, Ciencias Biologicas, 353, 1257–1270.
Itier, R. J., & taylor, METRO. j. (2002). Inversion and contrast polarity
reversal affect both encoding and recognition processes
of unfamiliar faces: A repetition study using ERPs.
Neuroimagen, 15, 353–372.
Itier, R. J., & taylor, METRO. j. (2004). N170 or N1? Spatiotemporal
differences between object and face processing using
ERPs. Corteza cerebral, 14, 132–142.
Jha, A. PAG. (2002). Tracking the time-course of attentional
involvement in spatial working memory: An event-related
potential investigation. Brain Research, Cognitive Brain
Investigación, 15, 61–69.
Kastner, S., Pinsk, METRO. A., De Weerd, PAG., Desimone, r., &
Ungerleider, l. GRAMO. (1999). Increased activity in human
visual cortex during directed attention in the absence
of visual stimulation. Neurona, 22, 751–761.
Kastner, S., & Ungerleider, l. GRAMO. (2000). Mechanisms of
visual attention in the human cortex. Annual Review
of Neuroscience, 23, 315–341.
Khoe, w., mitchell, j. F., Reynolds, j. h., & Hillyard, S. A.
(2005). Exogenous attentional selection of transparent
superimposed surfaces modulates early event-related
potentials. Vision Research, 45, 3004–3014.
Latinus, METRO., & taylor, METRO. j. (2006). Face processing stages:
Impact of difficulty and the separation of effects. Cerebro
Investigación, 1123, 179–187.
Linkenkaer-Hansen, K., Palva, j. METRO., Sams, METRO., Hietanen,
j. K., Aronen, h. J., & Ilmoniemi, R. j. (1998).
Face-selective processing in human extrastriate cortex
alrededor 120 ms after stimulus onset revealed by
magneto- and electroencephalography. Neurociencia
Letras, 253, 147–150.
Liu, J., harris, A., & Kanwisher, norte. (2002). Stages of
processing in face perception: An MEG study. Naturaleza
Neurociencia, 5, 910–916.
López, METRO., Rodríguez, v., & Valdés-Sosa, METRO. (2004).
Two-object attentional interference depends on attentional
colocar. International Journal of Psychophysiology, 53,
127–134.
Luck, S. J., Chelazzi, l., Hillyard, S. A., & Desimone, R.
(1997). Neural mechanisms of spatial selective attention
in areas V1, V2, and V4 of macaque visual cortex. Diario
of Neurophysiology, 77, 24–42.
Martinez, A., Ramanathan, D. S., Foxe, j. J., Javitt, D. C.,
& Hillyard, S. A. (2007). The role of spatial attention in
the selection of real and illusory objects. Diario de
Neurociencia, 27, 7963–7973.
Martinez, A., Teder-Salejarvi, w., Vazquez, METRO., Molholm, S.,
Foxe, j. J., Javitt, D. C., et al. (2006). Objects are
highlighted by spatial attention. Revista de Cognitivo
Neurociencia, 18, 298–310.
OʼCraven, k. METRO., Downing, PAG. MI., & Kanwisher, norte. (1999).
fMRI evidence for objects as the units of attentional
selección. Naturaleza, 401, 584–587.
Persona, l., Kastner, S., & Ungerleider, l. GRAMO. (2002).
Attentional control of the processing of neural and
emotional stimuli. Brain Research, Cognitive Brain
Investigación, 15, 31–45.
Pinilla, T., Cobo, A., Torres, K., & Valdés-Sosa, METRO. (2001).
Attentional shifts between surfaces: Effects on detection
and early brain potentials. Vision Research, 41, 1619–1630.
posner, METRO. I., Snyder, C. r., & Davidson, B. j. (1980).
Attention and the detection of signals. Diario de
Psicología experimental, 109, 160–174.
rees, GRAMO., Friston, K., & Koch, C. (2000). A direct quantitative
relationship between the functional properties of human
and macaque V5. Neurociencia de la naturaleza, 3, 716–723.
rossión, B., Delvenne, j. F., Debatisse, D., Goffaux, v.,
Bruyer, r., Crommelinck, METRO., et al. (1999). Spatio-temporal
localization of the face inversion effect: An event-related
potentials study. Biological Psychology, 50, 173–189.
rossión, B., Gauthier, I., Tarr, METRO. J., Despland, PAG., Bruyer, r.,
Linotte, S., et al. (2000). The N170 occipito-temporal
Rutman et al.
1233
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
6
mi
–
1
pag
2
d
2
F
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
oh
2
C
4
norte
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
5
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
/
F
.
.
.
t
oh
norte
1
8
METRO
a
y
2
0
2
1
component is delayed and enhanced to inverted faces
but not to inverted objects: An electrophysiological
account of face-specific processes in the human brain.
NeuroReport, 11, 69–74.
rossión, B., Joyce, C. A., Cottrell, GRAMO. w., & Tarr, METRO. j.
(2003). Early lateralization and orientation tuning for
rostro, palabra, and object processing in the visual cortex.
Neuroimagen, 20, 1609–1624.
Schoenfeld, METRO. A., Hopf, j. METRO., Martinez, A., & Mai, h. METRO.
(2007). Spatio-temporal analysis of feature-based attention.
Corteza cerebral, 17, 2468–2477.
serencias, j. T., Schwarzbach, J., Courtney, S. METRO., Golay, X.,
& Yantis, S. (2004). Control of object-based attention in
human cortex. Corteza cerebral, 14, 1346–1357.
Spitzer, h., Desimone, r., & Moran, j. (1988). Increased
attention enhances both behavioral and neuronal
actuación. Ciencia, 240, 338–340.
Sreenivasan, k. K., & Jha, A. PAG. (2007). Selective attention
supports working memory maintenance by modulating
perceptual processing of distractors. Diario de
Neurociencia Cognitiva, 19, 32–41.
Sreenivasan, k. K., katz, J., & Jha, A. PAG. (2007). Temporal
characteristics of top–down modulations during working
memory maintenance: An event-related potential
study of the N170 component. Revista de Cognitivo
Neurociencia, 19, 1836–1844.
Talsma, D., Mulckhuyse, METRO., Slagter, h. A., & Theeuwes, j.
(2007). Faster, more intense! The relation between
electrophysiological reflections of attentional orienting,
sensory gain control, and speed of responding. Cerebro
Investigación, 1178, 92–105.
Thut, GRAMO., Nietzel, A., Brandt, S. A., & Pascual Leone, A. (2006).
Alpha-band electroencephalographic activity over occipital
cortex indexes visuospatial attention bias and predicts visual
target detection. Revista de neurociencia, 26, 9494–9502.
Valdes-Sosa, METRO., Bobes, METRO. A., Rodriguez, v., & Pinilla, t.
(1998). Switching attention without shifting the spotlight
object-based attentional modulation of brain potentials.
Revista de neurociencia cognitiva, 10, 137–151.
Vogel, mi. K., & Machizawa, METRO. GRAMO. (2004). Neural activity
predicts individual differences in visual working memory
capacity. Naturaleza, 428, 748–751.
Vogel, mi. K., McCollough, A. w., & Machizawa, METRO. GRAMO. (2005).
Neural measures reveal individual differences in controlling
access to working memory. Naturaleza, 438, 500–503.
Hacer, D. J., & Chun, METRO. METRO. (2005). Attentional modulation of
learning-related repetition attenuation effects in human
parahippocampal cortex. Revista de neurociencia, 25,
3593–3600.
Zanto, t. PAG., & Gazzaley, A. (2009). Neural suppression of
irrelevant information underlies optimal working memory
actuación. Revista de neurociencia, 29, 3059–3066.
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
2
yo
6
mi
–
1
pag
2
d
2
F
4
/
1
2
9
2
3
/
9
6
4
/
7
1
3
2
oh
2
C
4
norte
/
1
2
0
7
0
6
9
9
5
2
2
1
8
2
5
/
7
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
9
mi
.
s
t
2
oh
1
norte
2
5
0
7
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
.
.
/
.
t
oh
norte
1
8
METRO
a
y
2
0
2
1
1234
Revista de neurociencia cognitiva
Volumen 22, Número 6