No me detengas ahora: Bases neuronales del aumento
Impulsivity to Temporally Predictable Events
, Boris Burle1, Jennifer T. Coull1, Halszka Ogińska2,
Inga Korolczuk1,2,3
Michał Ociepka2, Magdalena Senderecka2* , and Kamila Śmigasiewicz1*
Abstracto
■ Although the benefit of temporal predictability for behavior
is long-established, recent studies provide evidence that know-
ing when an important event will occur comes at the cost of
greater impulsivity. Aquí, we investigated the neural basis of
inhibiting actions to temporally predictable targets using an
EEG–EMG method. In our temporally cued version of the
stop-signal paradigm (two-choice task), participants used tem-
poral information delivered by a symbolic cue to speed their
responses to the target. In a quarter of the trials, an auditory sig-
nal indicated that participants had to inhibit their actions. Behav-
ioral results showed that although temporal cues speeded RTs,
they also impaired the ability to stop actions as indexed by lon-
ger stop-signal reaction time. In line with behavioral benefits of
temporal predictability, EEG data demonstrated that acting at
temporally predictable moments facilitated response selection
at the cortical level (reduced frontocentral negativity just before
the response). Asimismo, activity of the motor cortex involved in
suppression of incorrect response hand was stronger for tempo-
rally predictable events. De este modo, by keeping an incorrect response
in check, temporal predictability likely enabled faster implemen-
tation of the correct response. En tono rimbombante, there was no effect
of temporal cues on the EMG-derived index of online, dentro-
trial inhibition of subthreshold impulses. This result shows that
although participants were more prone to execute a fast
response to temporally predictable targets, their inhibitory con-
trol was, En realidad, unaffected by temporal cues. Altogether, nuestro
results demonstrate that greater impulsivity when responding
to temporally predictable events is paralleled by enhanced neu-
ral motor processes involved in response selection and imple-
mentation rather than impaired inhibitory control. ■
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
oh
C
norte
_
a
_
0
1
9
7
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
INTRODUCCIÓN
In the temporal prediction literature, the behavioral ben-
efits of acting to temporally predictable events are usually
emphasized. The vast majority of studies have investigated
the effects of temporal predictability using simple detec-
tion or discrimination tasks and have repeatedly demon-
strated that responses are faster and more accurate when
the time of target onset could be predicted in advance
(Nobre & van Ede, 2018; Correa, Lupiáñez, & Tudela,
2006; Coull & Nobre, 1998). Sin embargo, temporal predict-
ability does not always serve an adaptive function. De hecho,
when an already initiated response needs to be inhibited
or when responses require conflict resolution, temporal
predictability might actually be detrimental to performance
(Korolczuk, Burle, & Coull, 2018; Correa, Cappucci, Nobre,
& Lupiáñez, 2010). Por ejemplo, behavioral studies have
shown that when participants knew when a target would
appear, both correct and incorrect responses were more
likely to be co-activated, making it harder to resolve any
response conflict (Menceloglu, suzuki, & Song, 2021).
Online recordings of muscle activity have further demon-
strated that whenever a temporally predictable target
1Aix-Marseille University & CNRS, Francia, 2Jagiellonian Univer-
sity, Kraków, Poland, 3Medical University of Lublin, Poland
*Shared last authorship.
induces potentially conflicting responses, there were a
greater number of fast activations of the incorrect response
muscle. These erroneous muscle activations included both
fully executed suprathreshold responses as well as sub-
threshold response impulses (“twitches”; Korolczuk, Burle,
Coull, & Smigasiewicz, 2020). Sin embargo, temporal predict-
ability did not affect the ability to successfully suppress
subthreshold erroneous twitches, allowing the participant
to eventually execute the correct response. En otra
palabras, acting to temporally predictable yet conflicting
events exacerbates the urge to act impulsively but does
not weaken the corrective inhibitory processes.
In a recent EEG study, we identified the neural bases of
the costs and benefits of temporal predictability for con-
flicting actions (Korolczuk, Burle, Coull, & Śmigasiewicz,
2022). By investigating the cortical markers of correct
response activation and incorrect response inhibition
before the response had even been initiated, we found
that an EEG marker of incorrect response inhibition
(Burle, Vidal, Tandonnet, & Hasbroucq, 2004; Vidal,
Grapperon, Bonnet, & Hasbroucq, 2003) was differentially
modulated depending on response choice complexity.
For conflicting responses, this inhibitory activity was
weaker for temporally predictable targets, which suggests
that the behavioral costs of temporal predictability (p.ej.,
more fast errors) are because of insufficient suppression
© 2023 Instituto de Tecnología de Massachusetts. Published under a
Creative Commons Attribution 4.0 Internacional (CC POR 4.0) licencia.
Revista de neurociencia cognitiva 35:5, páginas. 885–899
https://doi.org/10.1162/jocn_a_01978
of the incorrect action. Strikingly, activity in the very same
inhibitory circuit was stronger when temporally predict-
able targets induced nonconflicting responses, indicando
that the behavioral benefits of temporal predictability
(p.ej., speeded RT) are achieved by keeping an incorrect
response in check. De este modo, in the context of competing
response alternatives, temporal predictability utilizes a
parsimonious cortical inhibitory mechanism that operates
right before the response is even initiated.
Todavía, efficient adaptation not only requires suppression
of the inappropriate action in favor of more goal-directed
unos, but might also require suppression of any action at
todo (Ridderinkhof, Forstman, Wylie, Burle, & van den
Wildenberg, 2011; Mostofsky & Simmonds, 2008). En efecto,
inhibiting the action in general (es decir., global response inhi-
bition) has been demonstrated to be conceptually and
empirically different than to suppressing a competing
response alternative (Duque, Greenhouse, Labruna, &
Ivry, 2017; Mostofsky & Simmonds, 2008; Verbruggen &
logan, 2008; Burle et al., 2004). Primero, suppression of the
competing motor response is usually measured with
stimulus–response incompatibility tasks, como el
Simón, Stroop, and flanker tasks. All of them require the
ability to inhibit the processing of irrelevant information
and to select the correct response (Beppi, Violante,
Hampshire, Grossman, & Sandrone, 2020; Ridderinkhof
et al., 2011). A diferencia de, global response inhibition is most
frequently studied using the stop signal task. The task
requires the ability to attend stop signals and to efficiently
counteract the preplanned motor response ( Verbruggen
& logan, 2008). All these paradigms undoubtedly share
some common features, but they also tap into different
subprocesses of response inhibition. Segundo, a recent
meta-analysis of brain imaging studies of action control
has revealed that selective response inhibition recruits dis-
tinct anatomical substrates than global response inhibition
(zhang, Geng, & Sotavento, 2017). Más precisamente, suppression
of the competing motor response, relative to action
withholding/cancellation, elicits stronger activation in
the left supplementary area, precentral gyrus, and supe-
rior parietal gyrus. This suggests its close association with
the response selection process. A diferencia de, acción
withholding/cancellation relies more pronouncedly on
the fronto-striatal network, which implies it as a late phase
of inhibitory process. Tercero, although psychopharmaco-
logical studies on different forms of inhibition are scarce,
an emerging body of research suggests that inhibitory pro-
cesses can be modulated by different neurotransmitter
sistemas (Lamar et al., 2009; Eagle, Barí, & robbins,
2008). More specifically, selective response inhibition
appears more sensitive to serotonin, whereas action can-
cellation to noradrenaline (apart from dopamine).
As already mentioned, this more global form of inhibi-
tion is often studied with the so-called stop-signal para-
digm ( Verbruggen & logan, 2008; logan, Cowan, & davis,
1984), in which participants perform a discrimination task
y, in some of the trials, an auditory signal is presented to
inform participants that they need to inhibit their res-
ponses completely (es decir., stop trials). The estimated time
taken to stop a response, termed the stop-signal reaction
tiempo (SSRT), provides an index of the ability to stop
actions that are no longer appropriate in a given context.
In a recent behavioral study investigating the effects of
temporal predictability on the ability to suppress inappro-
priate actions, we showed that SSRT was prolonged
when participants knew in advance the time of target
occurrence. In parallel, temporal cues led to faster RT
(Korolczuk et al., 2018). Such results suggest that tempo-
ral predictability increases overall response activation,
which leads to excessive response readiness, which might,
Sucesivamente, indirectly hinder the ability to inhibit the response.
Sin embargo, the exact neural mechanisms that could explain
the detrimental effects of temporal predictability on
stopping unwanted actions remain unknown.
The goal of the current study was to examine the periph-
eral and cortical bases of impulsivity triggered by active
prediction of the onset time of events. En tono rimbombante, nosotros
were interested in understanding how temporal predict-
ability affects online action control occurring within the
time-course of the action (after target presentation),
rather than anticipatory action regulation (occurring
before target presentation). These two aspects of action
control, often used interchangeably, are distinct in terms
of their neural bases, dinámica, and other factors like indi-
vidual differences or task characteristics (Ridderinkhof
et al., 2011). The vast majority of previous studies examin-
ing the neural bases of temporal predictability used simple
RT tasks, in which participants could prepare their
response in advance (p.ej., Volberg & Thomaschke, 2017;
Van Elswijk, Kleine, Overeem, & Stegeman, 2007; Miniussi,
Wilding, Coull, & Nobre, 1999). A diferencia de, our temporally
cued version of the stop-signal task allowed us to study the
modulatory mechanism of temporal prediction involved in
choosing the correct action and stopping responses that
are no longer appropriate. Específicamente, in this EEG–EMG
investigación, we studied several action control mecha-
nisms involved in both the selection and implementation
of responses as well as inhibiting actions in general.
Primero, to reveal the effects of temporal predictability
on neural response selection, we analyzed an electro-
physiological marker of response selection, known as
the N-40 component (Carbonnell et al., 2013; Vidal, Burle,
Grapperon, & Hasbroucq, 2011; Vidal et al., 2003). Este
frontomedial negative activity peaks around 40 mseg
before EMG onset and is modulated by the difficulty of
response choice demands (Burle, van den Wildenberg,
Spieser, & Ridderinkhof, 2016; Carbonnell et al., 2013).
More specifically, N-40 amplitude is greater for more dif-
ficult responses. Although previous neurophysiological
data revealed no effect of temporal cues on the N-40
component in a Simon task (Korolczuk et al., 2022), nosotros
sought to further clarify whether temporal predictability
might affect cortical response selection in the context of
a bimanual choice task.
886
Revista de neurociencia cognitiva
Volumen 35, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
oh
C
norte
_
a
_
0
1
9
7
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
EMG recording allowed us also to measure peripheral
markers of response activation, as indexed by EMG bursts
in the muscles involved in the response. Sucesivamente, EEG
additionally allowed us to study central markers of
activation of the correct response hand and inhibition of
the incorrect response hand—in choice RT tasks, este
“activation/inhibition” pattern is observed over primary
motor cortices (M1). Shortly before EMG onset, a negative
wave develops over the motor cortex contralateral to the
response agonist (activation of the correct response) y un
positive wave is observed over the motor cortex ipsilateral
to the response agonist ( Vidal et al., 2003, 2011) eso
reflects inhibition of the incorrect response (Burle,
Possamaï, Vidal, Bonnet, & Hasbroucq, 2002; Hasbroucq,
Akamatsu, Burle, Bonnet, & Possamaï, 2000; see Burle
et al., 2004, for a discussion). En tono rimbombante, the “activation/
inhibition” pattern over the M1 cannot be equated to the
lateralized readiness potential (LRP), a component known
to reflect motor preparation. The LRP is calculated as a dif-
ference between the left and right motor areas of the brain
(graton, Coles, Sirevaag, Eriksen, & Donchin, 1988), y
thus does not allow the activity of the contralateral and ipsi-
lateral motor cortex to be separated ( Vidal et al., 2003;
Eimer, 1999; graton, 1998). Además, the LRP is based
on the monopolar data and so the activities recorded
may stem from nonmotor remote areas making the
motor interpretation of the LRP questionable. En cambio,
the “activation/inhibition” pattern is based on the current
source density (CSD)-transformed signal (a través de
Laplacian estimation). The CSD increases the spatial res-
olution of the EEG signal as if electrodes were placed on
the surface of the cortex, and thus allows the sources of
the signal to be successfully segregated (Kayser & Tenke,
2015; Gevins, 1989). Además, by separating the activ-
ity of distinct neural generators, the CSD also improves
the temporal resolution of the signal of interest (Burle
et al., 2015; Law, Rohrbaugh, Adams, & Eckardt, 1993).
In terms of the timing of the “activation/inhibition” pat-
tern, the activities over M1 follow the N-40 negativity
(Burle et al., 2016; Vidal et al., 2003), which would indi-
cate the hierarchical organization of these areas involved
en el control de motores. En otras palabras, the M1 “activation/
inhibition” pattern would be situated downstream of
the SMA within a motor command hierarchy (Orgogozo
& Larsen, 1979). Alternativamente, the SMA and motor cortex
might work in parallel during response selection (Woolsey
et al., 1952). In this investigation, we aimed to examine the
role of cortical selection of the response as well as motor
activation and inhibition when acting to temporally pre-
dictable events.
EMG recordings can enhance the temporal resolution of
cortical markers of interest by allowing us to identify brain
activity right before the motor response is even initiated.
En tono rimbombante, EMG can also be effectively utilized to mea-
sure motor processes directly at the peripheral level.
En efecto, overt errors are only the tip of the iceberg and
it is critical to also study covert indices of impulsive
comportamiento. In the context of the stop-signal task, one can
quantify subthreshold muscle activations in the stop trials,
also called “partial responses,” which have been sup-
pressed and are thus not detectable in behavioral investi-
gaciones ( Van Boxtel, Van der Molen, Jennings, & Brunia,
2001; De Jong, Coles, logan, & graton, 1990). Besides
revealing covert response activations, such partial
responses can be also used to reveal the correction pro-
cesses directly at the peripheral level by computing the
partial response correction ratio. It is calculated as the pro-
portion of stop trials containing a partial response (es decir., a
subthreshold activation of the correct hand) comparado
with all successfully stopped trials (including a partial
EMG response or not). The correction ratio allows one
to measure how often initial impulses to act are subse-
quently suppressed. Aquí, we aimed to investigate
whether the increased impulsivity induced by temporal
cues is indeed because of an impaired ability to suppress
these partial responses by measuring the online inhibi-
tory mechanisms that act to stop covert subthreshold
impulses.
We formulated the following hypotheses. If temporal
predictability leads to greater impulsivity by impairing
global inhibitory processes, we would expect to see its
effects on the direct index of the within-trial inhibition of
subthreshold EMG activations that are no longer appropri-
ate. Específicamente, we would predict a lower partial response
correction ratio in temporal versus neutral condition.
Alternativamente, the detrimental effects of temporal predict-
ability on stopping impulsive responses could originate
from an increased urge to act. In the context of a discrim-
ination task, in which one cannot prepare a response in
advance, the facilitative effects of temporal cues would
be observed primarily within the time-course of the action
(after target presentation). Such motor facilitation could
stem from an easier selection and/or execution of the
respuesta. We would thus predict that at the brain level,
the effects of temporal predictability would be reflected
in easier response selection, empirically observed as atten-
uated N-40 activity. We would also predict that temporal
predictability would affect execution of the selected
respuesta. De este modo, right before response initiation, temporal
predictability would either lead to increased activation of
the correct response agonist and/or stronger suppression
of the incorrect response agonist, resulting in less inter-
ference from the incorrect hand and, por lo tanto, faster
implementation of the correct action. These mechanisms
would allow for rapid responding at precise moments in
time but might increase the difficulty of stopping actions
en general.
MÉTODOS
Participantes
We tested thirty-six participants (Mage = 22.1 años, DE =
2.8 años, 27 women) in the study approved by the
Korolczuk et al.
887
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
oh
C
norte
_
a
_
0
1
9
7
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
research ethics committee at the Institute of Applied Psy-
chology at the Jagiellonian University (Kraków, Poland).
The sample size was based on previous work (Korolczuk
et al., 2018, 2022). All participants had normal or
corrected-to-normal vision and no history of neurological
or psychiatric disorders. All participants gave written
informed consent. Data from seven participants were
discarded from the analysis because of excessive artifacts
(±2 SDs of the group average) in EEG recordings (two
individuals) or noisy or “flat” EMG recordings (five indi-
viduals). The final sample consisted of 29 Participantes.
Experimental Task
Participants performed a temporally cued version of the
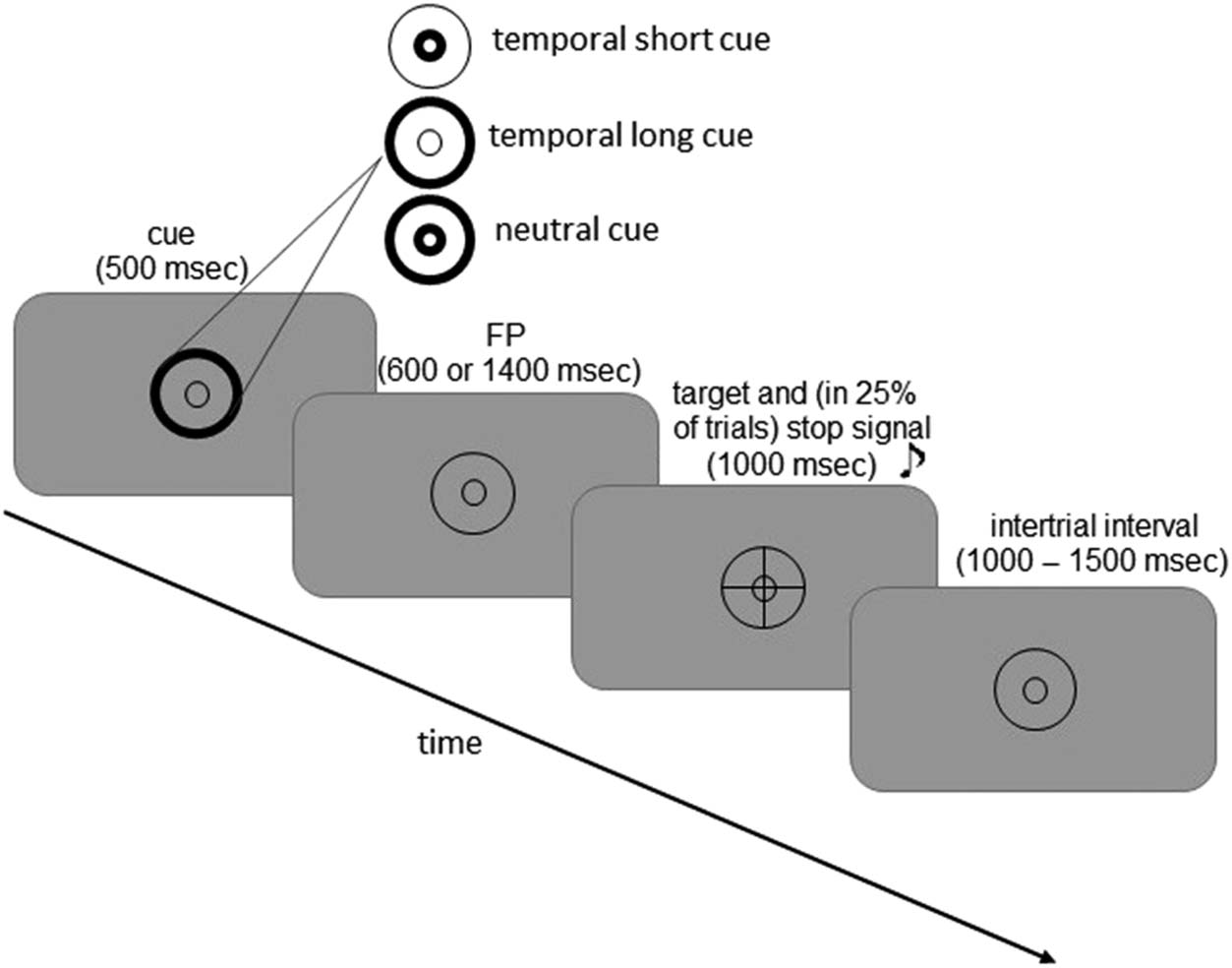
stop-signal task (Korolczuk et al., 2018; Cifra 1) estafa-
trolled by PsychoPy (Peirce et al., 2019; Peirce, 2007). Todo
stimuli were black, presented centrally on a gray back-
ground. Two concentric circles (1° eccentricity) eran
always present in the center of the screen (as a back-
ground display). Targets (“×” or “+”) were 1° × 1° stimuli
and appeared within the background display.
There were two cue conditions. In the temporal (t)
condición, thickening of the line forming the smaller
(inner) circle informed participants that a target would
occur after a short delay or “foreperiod” (FP; 600 mseg),
whereas thickening of the larger (outer) circle informed
participants that a target would occur after a longer inter-
vale (1400 mseg). Temporal cues were always valid. En el
neutral (norte) condición, the lines forming both circles were
thickened, thereby providing no temporally precise infor-
formación, and targets occurred randomly after either short
or long FPs. The cue (T or N) was presented for 500 mseg,
followed by presentation of the background display for
600 mseg (short FP) o 1400 mseg (long FP) and the target
para 1000 mseg.
Participants were encouraged to use the information
provided by the temporal cue to speed their RTs to targets
(“×” or “+”). In the neutral condition, they were encour-
aged to respond as quickly as possible to the targets
although they could not predict when it would appear.
Half of the participants responded with their left thumb
to “×” and with their right thumb to “+” on a standard
QWERTY keyboard (left “ctrl” and right “+” keys). Estos
target–response pairings were reversed for the remaining
Participantes. The target appeared within the circles and
remained there for 1000 mseg. During this time, participar-
pants gave their lateralized response according to target
forma. The trial ended with presentation of the back-
ground display for a duration between 1000 y 1500 mseg
(random jitter of 100 mseg).
En 25% of trials, an auditory stop signal (750 Hz, 50 mseg)
was presented a very short time after the target appeared,
instructing participants to withhold their response (stop
ensayos). There were never two stop-signal trials presented
consecutively. The stop-signal delay (SSD) between target
onset and the auditory beep was initially set at 100 mseg
and was adjusted continually using a staircase procedure.
If the participant successfully suppressed their response,
the SSD increased by 50 msec on the next stop trial. En
doblar, if the participant failed to inhibit their response,
the SSD decreased by 50 msec on the next stop trial. Estos
adjustments were made separately for temporal and neu-
tral cues, and for short and long FPs, thus allowing the
effects of cue and FP to be effectively disentangled. El
SSD ranged from 50 a 400 msec across trials with a jitter
de 50 mseg.
The two cue conditions (T and N) were presented
in two consecutive blocks in an alternating manner
Cifra 1. Temporally cued
version of the stop-signal task. A
cue (500 mseg) either predicted
(temporal condition) or not
(neutral condition) the time of
target onset. A background
display was then presented
for one of two FPs: corto
(600 mseg) or long (1400 mseg).
Entonces, the target (“×” or “+”)
appeared centrally for 1000 mseg
during which participants gave
their lateralized response
depending on the shape of
the target. En 25% of trials,
an auditory stop signal was
presented right after the target
with a variable SSD, informing
participants that they had
to withhold their response.
The intertrial interval was
randomized between 1000
y 1500 mseg.
888
Revista de neurociencia cognitiva
Volumen 35, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
oh
C
norte
_
a
_
0
1
9
7
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(TT-NN-TT-NN or NN-TT-NN-TT), which allowed us to bal-
ance training effects and fatigue across the two cuing con-
ditions. Había 128 trials per block, which resulted in
1024 trials altogether. In each block, the proportion of
short and long FPs was 50:50, and the proportion of go
to stop trials was 75:25. De este modo, Había 192 trials for
each of the four combinations of cue and FP in the go
ensayos, y 64 trials for each of the four combinations of
cue and FP in the stop trials. During an initial training
session, participants performed 30 temporal and 30 neu-
tral trials to familiarize themselves with the task.
EMG and EEG Recordings
We recorded electrophysiological data from 64 Ag/AgCl
active pre-amplified electrodes (Biosemi Inc.) at a rate of
1024 Hz (analogue bandwidth limit: from direct current to
268 Hz, −3 dB at one fifth of the sampling rate). The elec-
trodes were positioned in accordance with the extended
10–20 convention. Two electrodes lateral to the external
canthi were used to record the EOG and measure horizon-
tal eye movements. To measure vertical eye movements
and blinks, we recorded activity from an electrode beneath
the left eye and subtracted this activity from the FP1 elec-
trode. Además, we recorded the bipolar electromyo-
graphic activity of the flexor pollicis brevis from each hand
also using Ag/AgCl active electrodes positioned 2 cm apart
on the thenar eminence.
EMG and EEG Preprocessing
All the preprocessing steps and analysis of EMG and EEG
data were conducted using BrainVision Analyzer 2.0 (Cerebro
Products GmbH), MNE Python toolbox (Gramfort et al.,
2013), and customized Python scripts (www.python.org).
To detect the onset and offset of EMG activity, we used a
customized Python program (Spieser & Burle, 2022),1
which is based on a combination of two algorithms: “inte-
grated profile” (Liu & Liu, 2016; Santello & McDonagh,
1998) and a variance comparison (Hodges & Bui, 1996).
Entonces, a naive observer, unaware of the trial type, manually
corregido (if needed) the EMG onsets detected by the
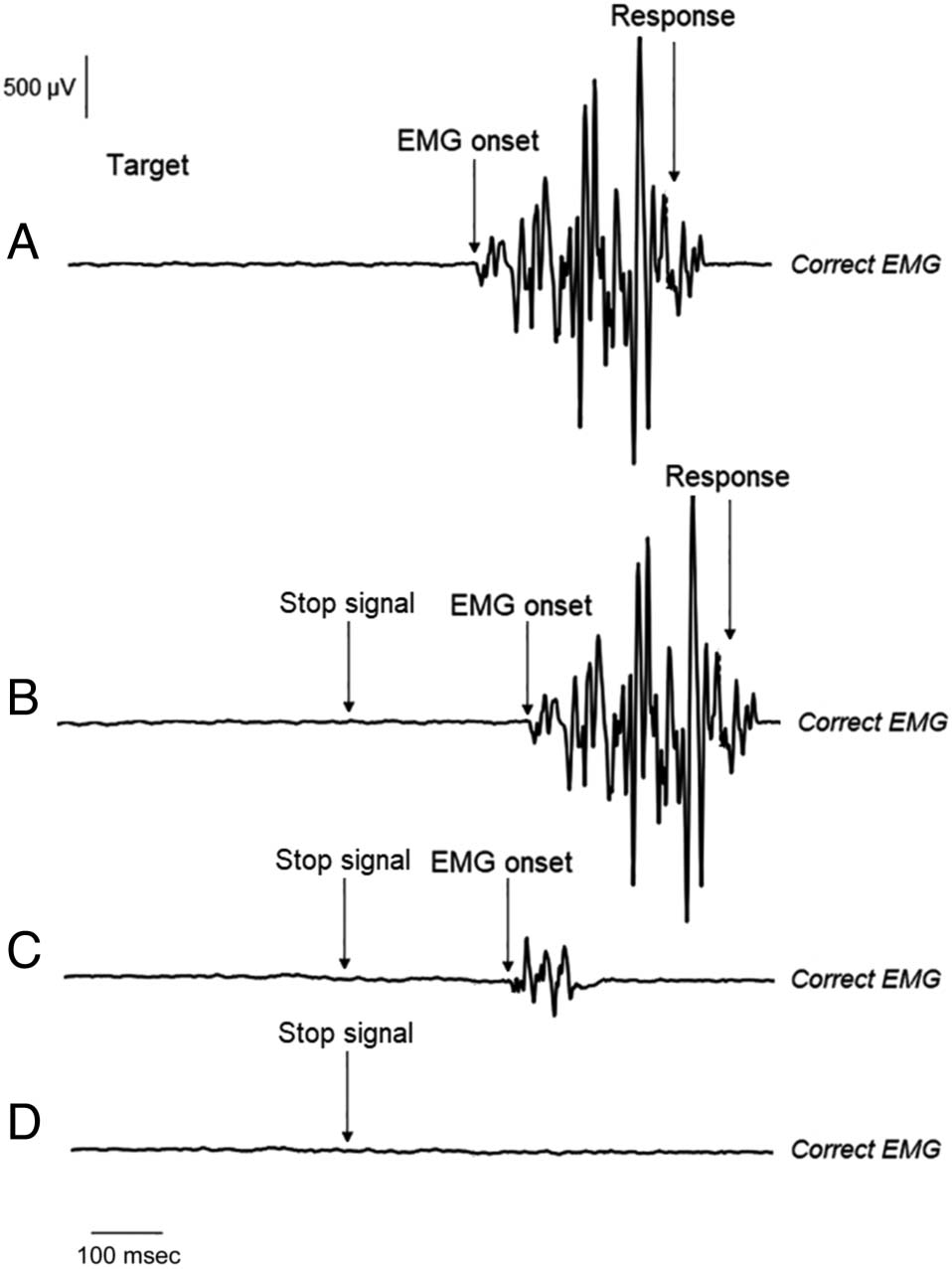
script. Based on this procedure, we distinguished and ana-
lyzed four types of trials: (1) pure correct go trials (es decir., go
trials with a single suprathreshold EMG activation for the
correct hand), (2) failed stop trials (es decir., stop trials with an
overt behavioral response and a single suprathreshold
EMG activation for the correct hand), (3) partial response
stop trials (es decir., stop trials without an overt behavioral
response but with a single subthreshold EMG activation
after a stop signal for the correct hand), y (4) pure stop
ensayos (es decir., stop trials without EMG activation). Partial
response stop trials in which subthreshold EMG activity
started before and finished after the stop signal were not
analyzed because of an insufficient number of trials. simí-
mucho, partial responses made with the incorrect hand were
not analyzed because of an insufficient number of trials.
Cifra 2. EMG trial types. (A) A pure correct response in a go trial. El
EMG activity appeared only in the correct hand and resulted in an overt
correct response. (B) A failed stop trial. The EMG activity appeared only
in the correct hand and resulted in an overt unsuccessfully stopped
respuesta. (C) A partial response stop trial. The subthreshold EMG
activity appeared only in the correct hand stop trials without an overt
behavioral response. (D) A successfully stopped trial. No EMG activity
was observed.
Cifra 2 presents an example of each trial type used in
the analysis.
The EEG data were rereferenced to the average of the
right and left mastoids, and the signal was band-pass
filtered between 0.01 y 100 Hz using a second-order
infinite impulse response Butterworth digital filter (slope:
12 dB/Oct). The MNE Python toolbox (Gramfort et al.,
2013; Uusitalo & Ilmoniemi, 1997) was used to correct
ocular artifacts. Data were then visually inspected for any
remaining noise and artifacts. All electrodes were rejected
even if only a small local artifact was present to allow for
subsequent use of the CSD computation, which is partic-
ularly sensitive to local artifacts.
Análisis de los datos
Behavioral Data Analysis
The mean RT from correct go trials was calculated sepa-
rately for each cue (temporal/neutral) and FP (short/long)
condición. Similarmente, the error rate in go trials (3%) was cal-
culated for cue and FP conditions. The omission rate
Korolczuk et al.
889
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
oh
C
norte
_
a
_
0
1
9
7
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
(3.8%) was not further analyzed. The SSD was quantified
for each cue and FP condition as the average delay
between the target onset and the auditory stop signal
onset for both successful and failed stop trials. The SSRT
(the mean time to inhibit a response) was estimated using
the integration method (logan, 1994). Primero, the RTs from
correct-only trials were rank ordered for each participant
and for each of the four conditions (temporal/neutral cue;
short/long FP). Entonces, the number of all responses in a given
condition was multiplied by the probability of responding
to a stop signal at a given delay [pag(respond|señal)] a favor-
duce the critical RT. Subtracting the SSD from this RT pro-
vides an estimate of the SSRT. En tono rimbombante, this integration
method of calculating the SSRT does not require the
assumption of 50% inhibition (es decir., participants inhibit
their responses in approximately half of stop trials), y
so it provides a reliable measure of the ability to inhibit
actions even when participants’ probability of responding
to a stop signal deviates from 50% (logan, 1994).
To measure the effects of temporal predictability on per-
rendimiento, we conducted a series of two-way repeated-
measures ANOVA involving Cue (temporal, neutral) y
FP (corto, largo). We examined the effects of temporal
predictability on go trial RT, go trial error rate, SSRT, SSD,
SRRT (signal-response RT, es decir., RTs on failed stop trials), y
[pag(respond|señal)]. Based on our previous findings
(Korolczuk et al., 2018), we expected to observe faster RT
paralleled by longer SSRT in temporal versus neutral trials.
Along with shorter SSD after temporal rather than neutral
señales, these results would indicate greater impulsivity to
temporally predictable targets. We further predicted that
these findings would be strongest in short FP trials.
EMG Data Analysis
To investigate the covert mechanisms for inhibiting
actions to temporally predictable targets, we measured
the partial response rate and the partial response correc-
tion ratio (the equivalent of the correction ratio in choice
RT tasks) in stop trials. The partial response was computed
as the proportion of stop trials containing a partial
respuesta (es decir., a subthreshold activation of the correct
mano) to all successfully stopped trials. The partial
response correction ratio was computed by dividing the
number of trials with partial responses by the overall num-
ber of incorrect activation trials (both failed stop trials and
partial response trials). It indexes the ability to inhibit a
response after it has been initiated.
Our paradigm included only two FPs and no catch trials.
Por lo tanto, if a target had not been presented at the short
FP in the neutral condition, the participant knew it would
necessarily have to appear at the long FP (Coull, Frith,
Büchel, & Nobre, 2000). This is because of the influence
of the hazard function, which is the increasing conditional
probability of target appearance over time given that it has
not already appeared (Luce, 1986; Durup & Requin, 1970;
Elithorn & lorenzo, 1955). Because targets presented at
the long FP were therefore 100% predictable in neutral as
well as temporal conditions, temporal and neutral cues
induced differential levels of temporal predictability at
the short FP only. Por lo tanto, the effects of temporal cue-
ing on partial response rate and the correction ratio were
evaluated by paired-samples t tests comparing temporal
and neutral conditions at the short FP only (van Ede,
Rohenkohl, Gould, & Nobre, 2020; Griffin, Miniussi, &
Nobre, 2002).
EEG Data Analysis
The analysis of EEG data was conducted on short FP trials
solo. In go trials, we first analyzed the effects of temporal
predictability on the frontocentral negativity known as the
N-40 component, which has been shown to vary with the
difficulty in response selection (Carbonnell et al., 2013;
Vidal et al., 2003, 2011). Activity over the FCz electrode
was segmented from −500 msec to 500 msec time-locked
to EMG onset, and baseline correction (from −500 msec
to −300 msec time-locked to EMG onset) was performed.
Próximo, data for individual participants were averaged for
each cue condition (temporal/neutral) for short FP trials
solo. We then performed the CSD computation using
BrainVision Analyzer 2.0. The signal was interpolated using
the spherical spline interpolation procedure (Perrin,
Pernier, & Bertrand, 1989), setting the degree of spline
to three. The second derivatives in two dimensions of space
were calculated with a maximum of 15° for the Legendre
polinomio. With the assumption of a head radius of
10 cm, the unit of EEG activity was μV/cm2. Individual
participants’ peak values (es decir., the most negative values)
were then extracted for the two cueing conditions, en un
time window from −100 msec to 0 msec relative to the
onset of the EMG. The statistical evaluation of these
peak values was performed using paired-samples t tests
(temporal short vs. neutral short). Además, nosotros estafamos-
ducted a between-participants Spearman’s rho correlation
analysis to explore the relationship between N-40 negativ-
ity and behavioral performance across participants. Más
específicamente, we sought to determine the relationship
between N-40 activity in temporal relative to neutral con-
ditions (T-N) and the RT benefit of temporal cues (N-T).
To investigate whether temporal predictability modu-
lated the motor cortex involved in activating the correct
hand and inhibiting the incorrect one in go trials, we seg-
mented the data separately for right- and left-hand
responses in a time window from −500 msec to 500 mseg
time-locked to the EMG onset. This was done separately
for the two cue conditions (temporal/neutral) for short
FP trials only. Entonces, the averaged and CSD-transformed
signal data were “collapsed” across the two hemispheres:
Data from left hemisphere C3 electrodes during (contra-
lateral) right-hand responses were averaged with data from
right hemisphere C4 electrodes during (contralateral)
left-hand responses (weighted average) to reflect the
activity of the cortex involved in producing the correct
890
Revista de neurociencia cognitiva
Volumen 35, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
oh
C
norte
_
a
_
0
1
9
7
8
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
respuesta. These averaged contralateral responses were
attributed to the C3 electrode for visualization purposes.
Similarmente, data from left hemisphere C3 electrodes during
(ipsilateral) left-hand responses were averaged with data
from right hemisphere C4 electrodes during (ipsilateral)
right-hand responses (weighted average) to reflect the
activity of the cortex involved in producing an incorrect
respuesta. These averaged ipsilateral responses were
attributed to the C4 electrode for visualization purposes.
We analyzed activity shortly preceding muscle activation in
a time window from −100 msec to 50 msec relative to the
EMG onset. To obtain a baseline-independent index of
phasic activity, we calculated the slopes of neural activity
with a customized Python script by fitting a linear regres-
sion to the data in the time window of interest. Finalmente,
statistical analysis of the slopes was conducted using
paired-samples t tests (temporal short vs. neutral short).
We also performed a Spearman’s rho correlation analysis
to test the relationship between ipsilateral motor cortex
inhibition and behavioral performance across participants.
More specifically, we investigated the relationship between
the inhibitory motor cortex activity in temporal relative to
neutral conditions (T-N) and the RT benefit of temporal
señales (N-T/N). For EMG and EEG analyses, one-tailed tests
were used whenever the directional hypotheses were
drawn based on our previous findings. For the remaining
contrasts, two-tailed tests were implemented.
RESULTADOS
Behavioral Results
Go Trials
We first aimed to establish whether participants used tem-
poral predictions to speed their motor responses by ana-
lyzing RTs in go trials. A two-way repeated-measures
ANOVA comprising Cue (temporal, neutral) and FP (corto,
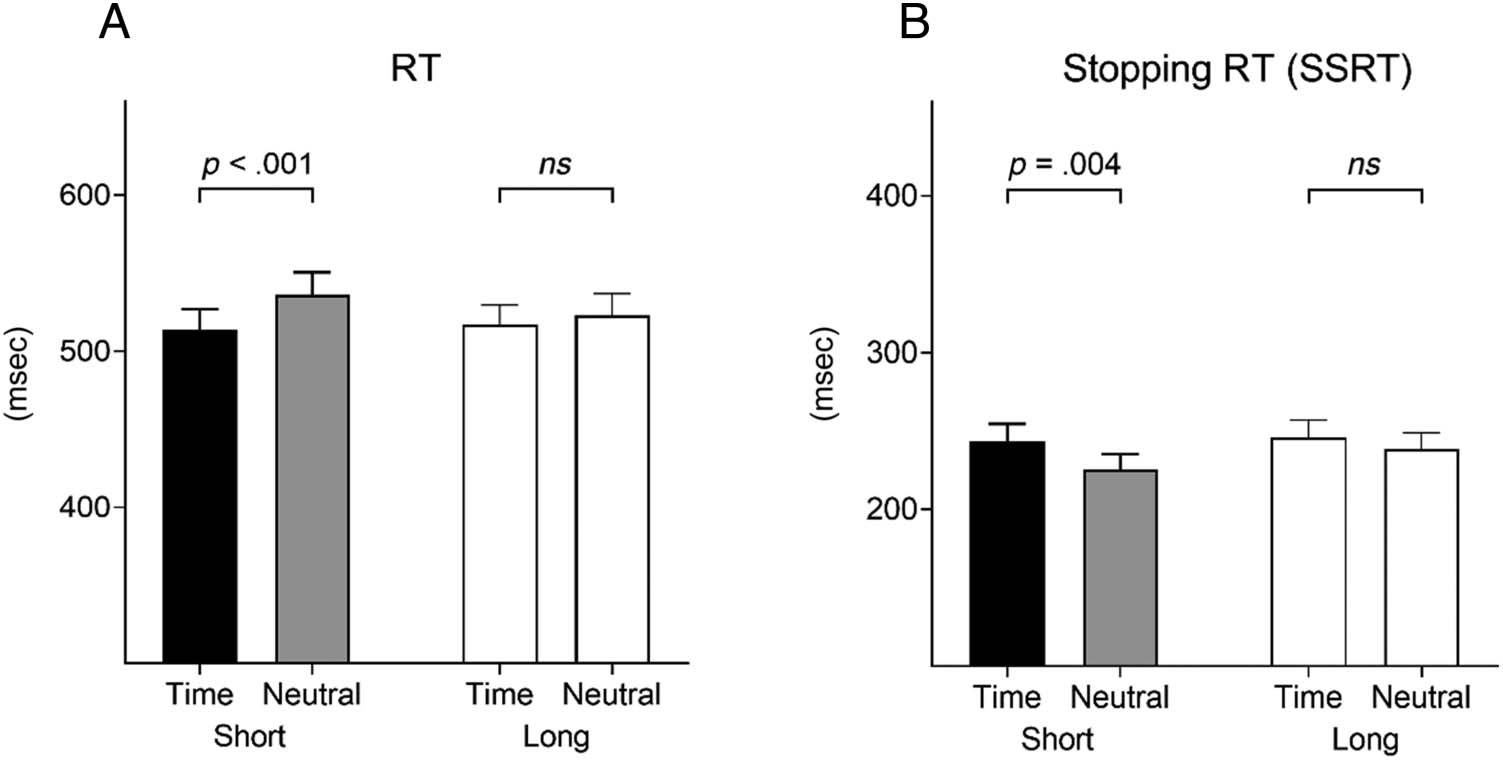
largo) revealed a main effect of Cue, F(1, 28) = 9.76, pag =
.004, ηp
2 = .26, and a main effect of FP, F(1, 28) = 5.25, pag =
.03, ηp
2 = .16, which were further qualified by a significant
Cue × FP interaction, F(1, 28) = 26.02, pag < .001, ηp
2 = .48.
Post hoc comparisons showed the typical pattern of
results: RTs were faster after temporal rather than neutral
cues in short FP trials ( p < .001), but not in long FP trials
( p = .206; Figure 3). This replicates previous findings
(Correa et al., 2006; Nobre, 2001; Coull & Nobre, 1998)
and confirms that effects of temporal cueing on response
speed are most pronounced at the short FP. Participants
made more errors in go trials after temporal than neutral
cues, F(1, 28) = 11.68, p = .002, ηp
2 = .29.
Stop Trials
Replicating our previous results (Korolczuk et al., 2018),
we found that temporal cueing made it harder for partici-
pants to inhibit their actions. The estimated RT to stop an
already activated response (SSRT) was longer in temporal
versus neutral trials, F(1, 28) = 5.43, p = .027, ηp
2 = .16.
Again, there was a Cue × FP interaction, F(1, 28) = 4.61,
p = .04, ηp
2 = .14. Temporal cueing led to longer SSRT in
short FP trials ( p = .004), but not long FP trials ( p = .227).
In parallel, the analysis of the SSD revealed a main effect of
Cue, F(1, 28) = 9.61, p = .004, ηp
2 = .26, and FP, F(1, 28) =
4.99, p = .034, ηp
2 = .15, which was explained by a signif-
icant Cue × FP interaction, F(1, 28) = 8.86, p = .006, ηp
2 =
.24. The SSD was shorter after temporal cues only in short
FP trials ( p < .001), whereas long FP trials cancelled out
this effect ( p = .178). The analysis of the RT in failed stop
trials further revealed main effects of Cue, F(1, 28) = 9.22,
p = .005, ηp
2 = .25, and FP, F(1, 28) = 5.26, p = .029,
ηp
2 = .16. As previously, these main effects were qualified
by a significant Cue × FP interaction, F(1, 28) = 6.73,
p = .015, ηp
2 = .19. RT in failed stop trials (SRRT) was
Figure 3. The effects of temporal cueing on RT in go trials and stopping RT (SSRT) in stop trials. (A) Temporal cueing speeded RTs in go trials. (B) In
parallel, temporal cues led to slower SSRT in stop trials. As expected, these effects were most pronounced in short FP condition but not long FP
condition. Error bars reflect standard errors.
Korolczuk et al.
891
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Behavioral and EMG Results
Measure
Behavioral
EMG
Go RT
SSRT
SSD
SRRT
% Partial response (stop trials)
Partial response correction (%)
Time
514 (13)
243 (11)
247 (16)
570 (15)
22.4 (2.0)
31.2 (2.8)
Neutral
536 (14)
225 (10)
275 (15)
597 (13)
21.2 (1.8)
29.7 (2.3)
Behavioral measures include: go RT, SSRT, SSD, and SRRT. EMG measures include: percentage of partial response and partial response correction
rate in stop trials. Indices are provided with standard errors (msec) for short FP trials.
shorter in the temporal cue condition than in the neutral
condition in short FP trials ( p < .001) but not in long FP
trials ( p = .131). Finally, the analysis of the mean per-
centage of the failure to stop a response [p(respond|sig-
nal)] showed main effects of Cue, F(1, 28) = 6.06, p =
2 = .18, and FP, F(1, 28) = 5.55, p = .026, ηp
.02, ηp
2 =
.17. Again, these effect were explained by a Cue × FP
interaction, F(1, 28) = 5.66, p = .024, ηp
2 = .17. The per-
centage of failures to stop a response was higher for the
temporal versus neutral cue condition at the short FP
( p = .003), but not at the long FP ( p = .209).
Overall, the consistent pattern of Cue × FP interactions
confirms that the differential effects of temporal predict-
ability can be measured at the short FP only. Therefore,
in all subsequent EMG and EEG analyses, we examined
the effects of temporal cueing on short FP trials only. More
importantly, behavioral results revealed a complementary
influence of temporal predictability in go versus stop
trials. Although temporal cueing facilitated responding
as demonstrated by faster RTs in go trials, it also led to
greater impulsivity as revealed by slower SSRT in stop
trials (Figure 3).
EMG Results
Partial Response Rate
Table 1 shows the effects of temporal predictability on
EMG-derived measures. The analysis of the partial
response rate showed that on approximately 22% of suc-
cessfully stopped trials, participants emitted a subthresh-
old muscle activation in the correct response hand that
was subsequently suppressed. However, temporal pre-
dictability did not affect the number of these activations,
t(28) = 0.61, p = .28, one-tailed, Cohen’s d = 0.11. Given
that we have previously shown that temporal cues led to a
greater likelihood of subthreshold muscle activations that
were later inhibited in the context of the Simon conflict
task (see Korolczuk et al., 2020), we ran an additional
Bayesian paired-samples t test, to interpret the current null
effect more confidently. A BF01 (i.e., an exclusion BF,
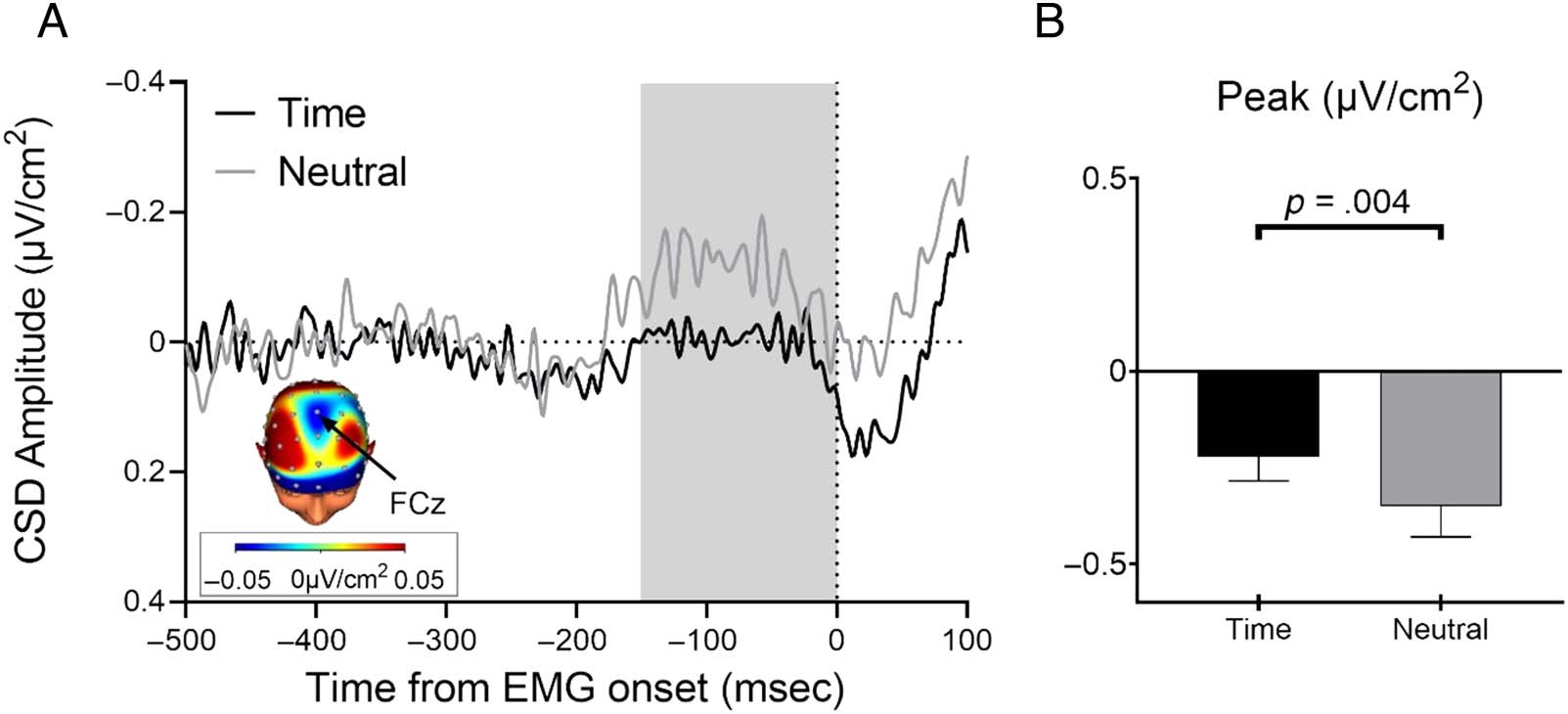
Figure 4. The frontocentral negativity indexing response selection (i.e., N-40 component) in go trials, CSD-transformed, time-locked to EMG onset.
(A) The N-40 component was less pronounced for the temporal cue condition (black) than the neutral cue condition (gray), indicating that temporal
predictability made it easier to select a response. Topography (CSD-transformed) was recorded over the FCz electrode. (B) The statistical analysis of
the peak revealed a significant Cue effect.
892
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
indicating the probability ratio between H0 and H1
models) was 4.28, which indicated that there was substan-
tial evidence for an absence of effect of temporal predict-
ability on partial response rate.
Altogether, the EMG findings indicate that although
temporal cueing makes it more difficult to stop a response
by speeding response initiation, it does not impair the
ability to interrupt a response once it has been initiated.
Partial Response Correction Ratio
To examine whether temporal cues impaired the ability
to suppress an already initiated response (i.e., partial
response) in stop trials, we compared the partial response
correction rate in temporal and neutral conditions. Impor-
tantly, and in line with our previous results (see Korolczuk
et al., 2020), there was no difference between the two cue
conditions, t(28) = 0.53, p = .60, two-tailed, Cohen’s d =
0.1. To determine the evidence in favor of this null effect,
we also conducted a Bayesian paired-samples t test. A BF01
was 4.45, which indicated that there was substantial evi-
dence for the lack of an effect of temporal cueing on the
partial response correction ratio.
EEG Results
Response Selection
To examine the effects of temporal predictability on
response selection, we measured the frontocentral nega-
tive activity (N-40 component), which occurs shortly
before EMG onset in choice RT tasks. The N-40 has been
found to be more pronounced (i.e., more negative) for
more difficult response choice demands (Carbonnell
et al., 2013; Vidal et al., 2003, 2011). We hypothesized
that acting at predictable moments in time would facili-
tate response selection, which should be observed
empirically as a smaller N-40 in temporal versus neutral
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
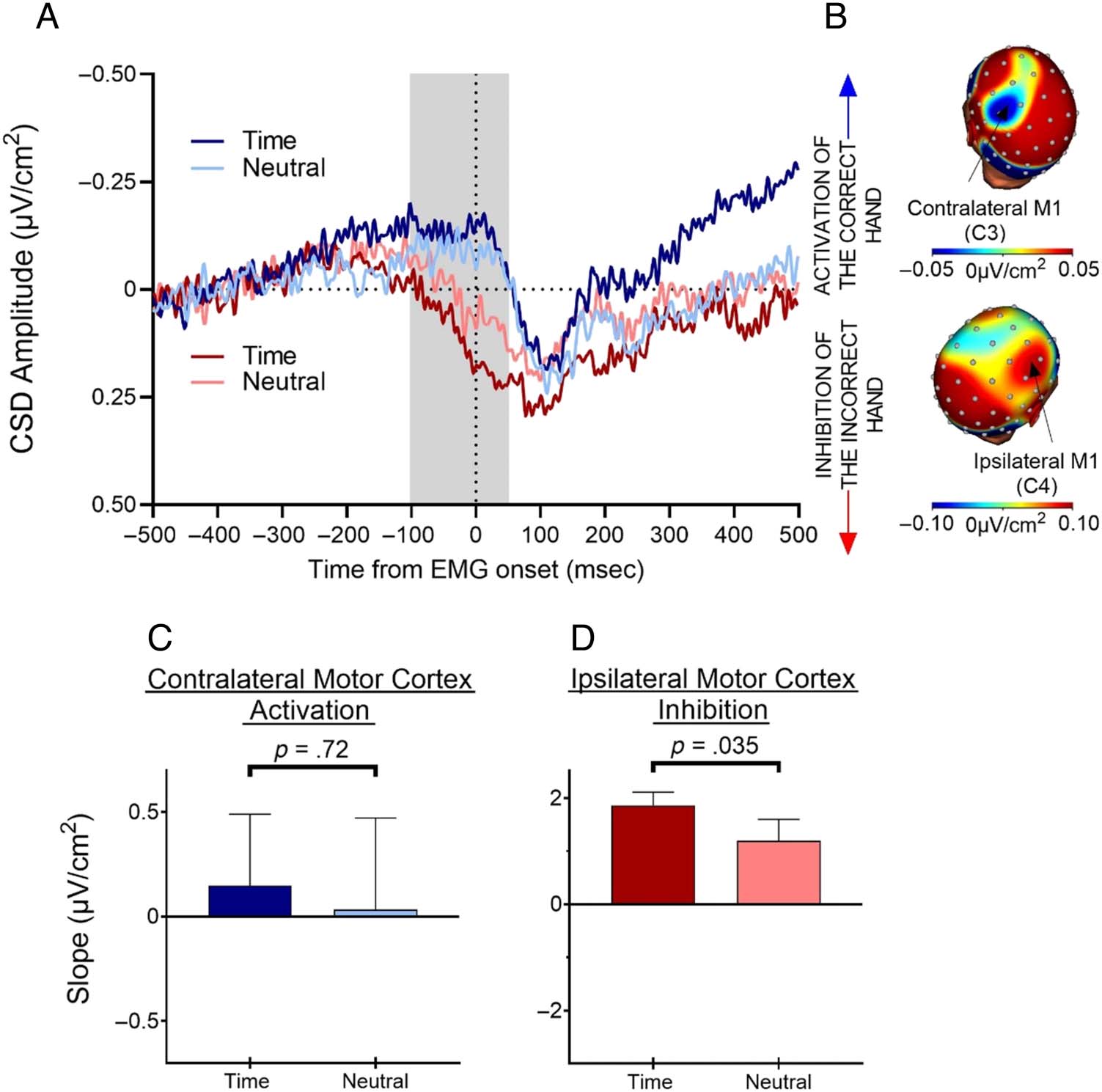
Figure 5. Motor cortex activation of the correct hand and inhibition of the incorrect hand shortly before EMG onset in go trials. (A) The negative-
going slope indexes correct response activation in contralateral motor cortex for temporal (dark red) and neutral (light red) conditions, whereas the
positive-going slope indexes incorrect response inhibition in ipsilateral motor cortex for temporal (dark blue) and neutral (light blue) conditions. (B)
Topographies (CSD-transformed) around EMG onset for motor cortex activation (recorded over the C3 electrode) and motor cortex inhibition
(recorded over the C4 electrode). (C) Temporal predictability did not affect motor cortex activation of the correct hand. (D) In contrast, motor cortex
inhibition of the incorrect hand was stronger in the temporal condition, as demonstrated by steeper slopes following temporal than neutral cues.
Error bars reflect standard errors.
Korolczuk et al.
893
cue conditions. We conducted peak analysis in the time
window from −150 msec to 0 msec (time-locked to the
EMG onset) on CSD-transformed data from the go trials.
The N-40 activity was less negative following temporal
than neutral cues, t(28) = 3.02, p = .005, two-tailed,
Cohen’s d = 0.56 (Figure 4). Thus, being able to predict
when a target will occur results in more efficient cortical
response selection within the time course of the action.
The facilitative effects of temporal predictability on
response selection are in line with temporal performance
benefits such as faster RTs and premotor times.
Correct Response Activation
We then tested whether temporal predictability acts by
modulating cortical activation of the correct hand as
indexed by the negativity developing over the motor cor-
tex contralateral to the response agonist immediately
before EMG onset (Burle et al., 2004; Vidal et al., 2003).
The slope analysis was conducted on the CSD-
transformed data from go trials in a time window from
−100 msec to 50 msec, time-locked to the EMG onset.
Replicating previous results using the Simon response
conflict paradigm (Korolczuk et al., 2022), there was no
effect of temporal predictability on motor cortex activa-
tion, t(28) = 0.36, p = .72, two-tailed, Cohen’s d = 0.07
(Figure 5).
Incorrect Response Inhibition
We next analyzed the effects of temporal predictability on
cortical inhibition of the incorrect hand as indexed by the
positivity developing over the motor cortex ipsilateral to
the response agonist (i.e., contralateral to the incorrect
response hand) before EMG onset. Again, slope analysis
was performed in the time window of −100 msec to
50 msec, time-locked to the EMG onset, on the CSD-
transformed data from go trials.
Given our previous results using the Simon response
conflict task (Korolczuk et al., 2022), we expected to
observe stronger motor cortex inhibition of the incorrect
hand (more positive-going neural activity) when reacting
to temporally predictable targets. Confirming our hypoth-
esis, the slopes were more positive-going for temporal
than neutral trials, t(28) = 1.89, p = .035, one-tailed,
Cohen’s d = 0.35 (Figure 5). These findings indicate that
temporal predictability recruits a cortical inhibitory mech-
anism that keeps an incorrect response in check to ensure
rapid initiation of the appropriate response.
Brain–Behavior Correlations
Finally, we correlated task performance and cortical motor
control indices across participants. We hypothesized that
the behavioral benefits of temporal predictability such as
faster RT are linked to improved cortical response selec-
tion and stronger inhibition of the incorrect hand in the
temporal, relative to neutral, condition.
We thus correlated the RT benefit of temporal cues (N-T,
with higher values reflecting greater temporal benefits),
with (1) the relative attenuation of the negative activity
indexing response selection processes at the cortical level
(N-40) for temporal cues (T-N, with more positive values
reflecting easier response selection in temporal than
neutral condition), and (2) the relative increase of cortical
inhibition of the incorrect response hand for temporal
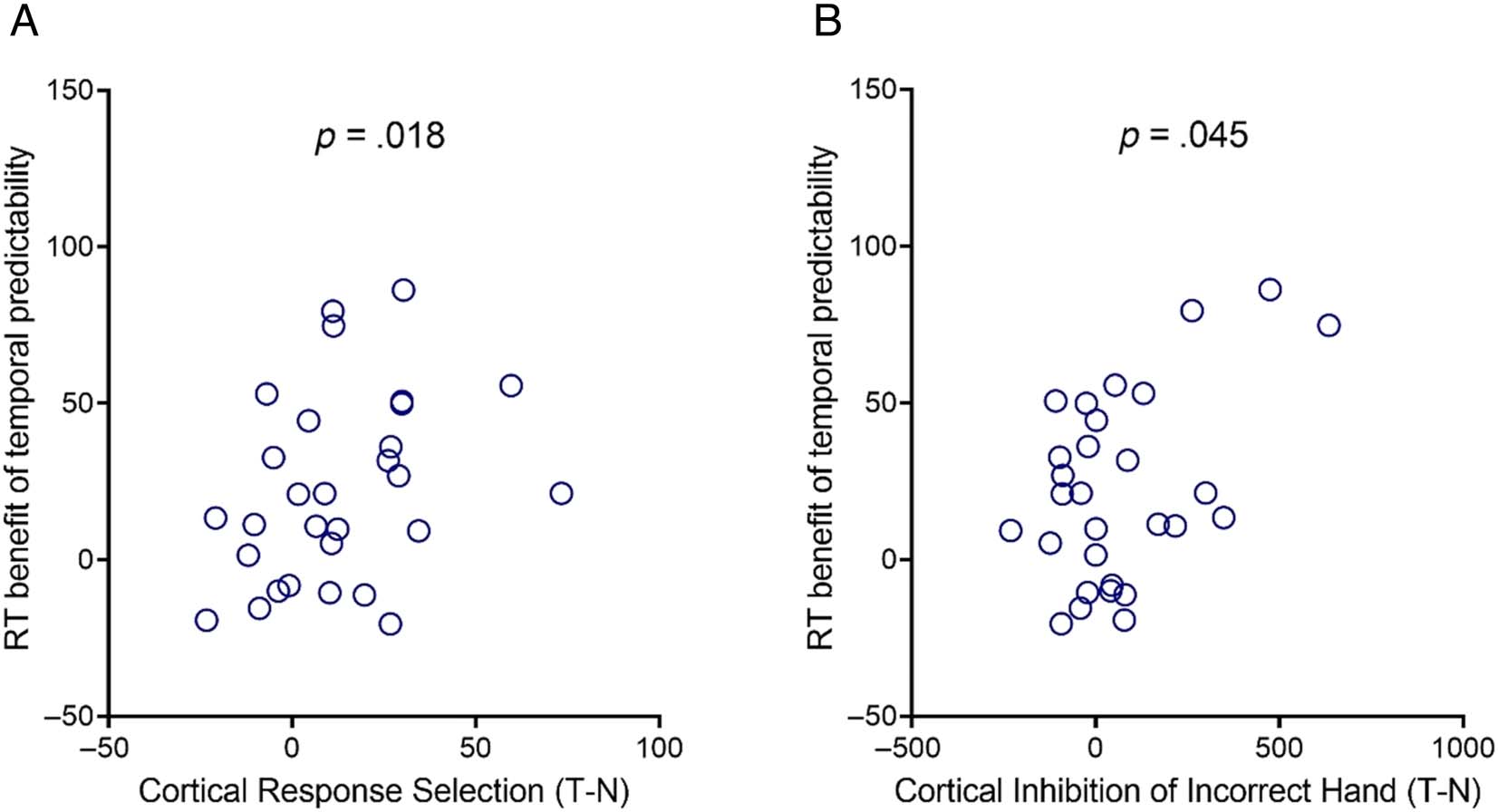
Figure 6. Brain–behavior correlations. Each point represents an individual participant. (A) Improvements in cortical response selection following
temporal cues (more positive values for temporal/T than neutral/N condition) correlated positively with RT benefits of temporal predictability. (B)
Similarly, stronger cortical inhibition of the incorrect hand in temporal, relative to neutral, trials, correlated positively with the RT benefit of temporal
predictability.
894
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
cues (T-N, with more positive values reflecting stronger
inhibition of the incorrect response hand in temporal
than neutral condition).
In line with our interpretation of the findings, there was
a positive correlation between the reduction in response
selection negativity (N-40 component) in temporal, versus
neutral, trials, and the RT benefit of temporal cues, r(27) =
.392, p = .018, one-tailed (Figure 6A). In other words, if
a participant had a greater difference in amplitude of the
N-40, they also had a greater difference in performance.
Similarly, there was also a positive correlation between
the increase in the strength of motor cortex inhibition in
the temporal, relative to neutral, condition, and the RT
benefit of temporal predictability, r(27) = .322, p =
.045, one-tailed (Figure 6B). In other words, if a participant
had a greater difference in the strength of cortical inhibi-
tion of the incorrect response hand, they also had a greater
difference in performance.
DISCUSSION
Being able to predict when an event is going to occur opti-
mizes motor processes. However, temporal predictability
can also increase impulsivity when a prepotent response
needs to be inhibited. We used a temporally cued version
of the stop-signal task to reveal cortical and peripheral
mechanisms of both reacting and stopping those reactions
at predictable moments in time. First, we confirmed that
temporal cues both speeded RT and exacerbated impul-
sive behavior, with the latter being indexed by the longer
time needed to inhibit a response (SSRT). To identify the
neural bases of impulsive responding to temporally pre-
dictable events, we examined cortical activity right before
the response was even initiated. EEG results showed that
temporal predictability facilitated response selection. In
parallel, inhibition of motor cortex involved in the incor-
rect response agonist was stronger following temporal
cues. Importantly, however, EMG data demonstrated that
temporal predictability did not impede the ability to with-
hold the to-be-stopped response once it has started to be
executed (partial false alarm).
Behavioral Costs and Benefits of
Temporal Predictability
As predicted, RT was faster after temporal than neutral
cues, which demonstrates once again the behavioral ben-
efits of temporal predictability. This was, however, accom-
panied by an increased number of incorrect responses in
go trials, revealing a speed-accuracy trade-off. On the
other hand, temporal predictability made it harder to stop
responses, as indexed by longer stopping RT (SSRT).
Altogether, these results demonstrate that temporal pre-
diction exacerbates the urge to act, which increases impul-
sivity in tasks requiring a flexible adjustment of actions.
One facet of the impulsive behavior triggered by temporal
prediction depends on the specific motor context. In
response conflict tasks, in which one response needs to
be inhibited in favor of another, temporal predictability
increases the tendency to initiate a fast and incorrect
response (Korolczuk et al., 2020, 2022; Menceloglu et al.,
2021; Correa et al., 2010). In turn, in tasks that require the
response to be withheld entirely, such as the stop-signal
task, a priori temporal expectancies make it harder to flex-
ibly stop actions (Korolczuk et al., 2018). Importantly, this
behavioral pattern (i.e., longer SSRT along with shorter
RT) appears to be specific to explicit temporal prediction
induced by temporal cues rather than any form of prepa-
ration in general. In fact, higher motor preparation has
been demonstrated to correlate negatively with both RT
and SSRT such that the higher the motor preparation,
the shorter the RT and SSRT ( Wang et al., 2018).
Notably, although one may argue that temporal predict-
ability impairs inhibitory processes, previous results have
not supported this hypothesis (Korolczuk et al., 2020). On
the contrary, it appears that a link between timing and
impulsivity comes from the effects of temporal predict-
ability on response activation rather than impairment of
corrective inhibitory processes. Likewise, in the current
study, both accelerated RT and slower SSRT following
temporal cues were likely underlined by an excessive
response readiness. Thus, the inability to stop prepotent
responses can be explained by the increased level of acti-
vation caused by the prediction of the time of the event.
The neural correlates of such increased activation are
discussed below.
Temporal Predictability Enhances Response
Selection as Indexed by the Frontocentral
N-40 Component
EEG analyses revealed that the N-40 component, which
covaries with response selection difficulty (Carbonnell
et al., 2013), was modulated by temporal predictability.
More specifically, the negative activity became less pro-
nounced when participants were about to make a tempo-
rally guided response, which might indicate easier
response selection after temporal cues. The N-40 compo-
nent, reflecting response selection process, arises before
the “activation/inhibition” pattern (Burle et al., 2016; Vidal
et al., 2003); the facilitative effects of temporal cues are sit-
uated upstream in the motor command hierarchy (at least
in the context of a discrimination task; Orgogozo & Larsen,
1979).
Incidentally, at first glimpse, these findings are at odds
with previous EEG data suggesting that temporal predict-
ability does not act by modulating the selection of
responses (Korolczuk et al., 2022). In the prior investiga-
tion using a temporally cued Simon conflict task, we found
that although N-40 amplitude varied as a function of
response choice difficulty with more pronounced activity
for conflicting rather than nonconflicting responses, it
remained insensitive to temporal characteristics of the
task. Importantly, however, the current study employed
Korolczuk et al.
895
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
a nonconflict choice task. It could be that increased activity
because of conflict induces a ceiling effect preventing a
genuine cue effect to appear. Alternatively, although not
mutually exclusively, spatial certainty might be necessary
for the beneficial effects of temporal cues to be observed.
Indeed, previous EEG data have demonstrated that the
effects of temporal predictability are more pronounced
when the location of the target is known in advance
(Rohenkohl, Gould, Pessoa, & Nobre, 2014; Doherty, Rao,
Mesulam, & Nobre, 2005). Hence, in a spatially certain
stop-signal task, the consequences of temporal predictabil-
ity for response selection might be stronger than in a spa-
tially uncertain Simon task. Our data support this notion
and link the behavioral benefits of temporal cues to more
effective cortical response selection.
The Motor Cortex Involved in Inhibiting
the Incorrect Hand Is Modulated by
Temporal Prediction
Following response selection, a correct response is acti-
vated over contralateral motor cortex and an incorrect
response is inhibited over ipsilateral motor cortex (Burle
et al., 2004). Prior EEG results showed that temporal
predictability does not act by increasing the activation of
the correct response hand in a response conflict task
(Korolczuk et al., 2022). The current data replicated and
extended this observation: Activation over the motor cor-
tex involved in generating the correct action remained
insensitive to the temporal structure of the task at hand
also in a nonconflict choice RT paradigm. It should be
noted, however, that this finding does not contradict prior
investigations, indicating that temporal predictability
increases motor activation. Whereas most of the EEG
reports examined the effects of temporal predictability
using a simple RT task ( Van Elswijk et al., 2007; Miniussi
et al., 1999) or when a response hand was known in
advance ( Volberg & Thomaschke, 2017), in our task, par-
ticipants could not prepare a motor response in advance.
Consequently, our results provide insight into the neural
mechanisms by which temporal prediction modulates
neural motor processes after target presentation. Within
this context, we demonstrate that the modulatory effects
of temporal predictability do not include cortical activation
of the correct action.
More importantly, however, temporal predictability
modulated the motor cortex involved in suppressing the
incorrect action. Right before the action was even initi-
ated, the positive activity in the ipsilateral motor cortex
associated with an incorrect response hand was more pro-
nounced following temporal cues, indicating stronger
inhibition of incorrect response (Burle et al., 2004; Vidal
et al., 2003) when acting to temporally predictable events.
Such an exclusive effect of temporal cues on cortical inhi-
bition lends further support to the notion that perfor-
mance benefits are achieved by stronger inhibition of
the incorrect hand (Korolczuk et al., 2022), possibly by
ensuring faster initiation and execution of the correct
action. Indeed, the strength of the inhibition of the ipsilat-
eral motor cortex involved in the suppression of errone-
ous actions correlated positively with the RT benefit of
temporal cues. Taken as a whole, these data indicate that
in the context of choice RT tasks (both conflicting and
nonconflicting), temporal prediction utilizes inhibitory
circuits over the motor cortex involved in keeping an
incorrect response in check to ensure a timely and rapid
response.
Temporal Predictability Leaves the Ability to
Correct Subthreshold Impulses Intact
To complement the EEG data, we used EMG recordings to
obtain a direct measure of peripheral processes involved
in suppressing actions at temporally precise moments.
The partial response correction ratio—a direct, online
marker of response inhibition—was unaffected by tempo-
ral prediction. This null effect is an important aspect of our
findings, which demonstrates that impulsive behavior fol-
lowing temporal cues does not originate from impaired
inhibitory processes per se. Instead, our results showed
that increased motor readiness prompted more rapid
responding, which exacerbated the difficulty in stopping
actions in general. Finally, it might seem contradictory that
temporal predictability did not increase the number of
subthreshold responses but, at the same time, it made it
harder to stop a response completely. These findings can
be explained in terms of all-of-none type of impulsivity,
which is triggered by temporal predictions. In other
words, temporal predictability leads to more overt false
alarms (makes it harder to stop the error completely),
but it does not increase the number of subthreshold
responses nor it affects the ability to correct an impulsive
error for a more appropriate response.
To conclude, our results provide compelling evidence
for the dual nature of temporal predictability on action
control. On one hand, acting at temporally predictable
moments enhanced cortical facilitation of response selec-
tion and led to stronger suppression of the incorrect
response hand. Yet, temporal predictability led to perfor-
mance costs when activated responses needed to be
stopped. Importantly, however, the online inhibition of
these impulsive activated actions was not impaired by
temporal predictability. Taken together, our results
demonstrate that costs of temporal predictability for
stopping unwanted actions are paralleled by enhanced
neural motor processes rather than impaired response
inhibition.
Reprint requests should be sent to Inga Korolczuk, Laboratoire
des Neurosciences Cognitives UMR 7291, Federation 3C, Aix-
Marseille University & CNRS, 3 Place Victor Hugo, 13331 Marseille
cedex 3, France, or via e-mail: inga.korolczuk@univ-amu.fr.
896
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Data Availability Statement
Materials, data, and analysis script will be made available
upon request to the lead author.
Funding Information
This work was supported by the National Science Centre
(Narodowe Centrum Nauki) of Poland (https://dx.doi.org
/10.13039/501100004281), grant number: 2018/31/N/HS6/
00633; and by a postdoctoral fellowship from the Fyssen
Foundation (Fondation Fyssen; https://dx.doi.org/10
.13039/501100003135), awarded to Inga Korolczuk. Inga
Korolczuk was also supported by the Foundation for
Polish Science (Fundacja na rzecz Nauki Polskiej; https://
dx.doi.org/10.13039/501100001870). The funding source
had no impact on any part of the present study.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .64; W/M = .08;
M/ W = .08; W/ W = .2.
Note
1. Soon to be released under open source license.
REFERENCES
Beppi, C., Violante, I. R., Hampshire, A., Grossman, N., &
Sandrone, S. (2020). Patterns of focal- and large-scale
synchronization in cognitive control and inhibition: A review.
Frontiers in Human Neuroscience, 14, 196. https://doi.org
/10.3389/fnhum.2020.00196, PubMed: 32670035
Burle, B., Possamaï, C. A., Vidal, F., Bonnet, M., & Hasbroucq,
T. (2002). Executive control in the Simon effect: An
electromyographic and distributional analysis. Psychological
Research, 66, 324–336. https://doi.org/10.1007/s00426-002
-0105-6, PubMed: 12466929
Burle, B., Spieser, L., Roger, C., Casini, L., Hasbroucq, T., &
Vidal, F. (2015). Spatial and temporal resolutions of EEG:
Is it really black and white? A scalp current density view.
International Journal of Psychophysiology, 97, 210–220.
https://doi.org/10.1016/j.ijpsycho.2015.05.004, PubMed:
25979156
Burle, B., van den Wildenberg, W. P. M., Spieser, L., &
Ridderinkhof, K. R. (2016). Preventing (impulsive) errors:
Electrophysiological evidence for online inhibitory control
over incorrect responses. Psychophysiology, 53, 1008–1019.
https://doi.org/10.1111/psyp.12647, PubMed: 27005956
Burle, B., Vidal, F., Tandonnet, C., & Hasbroucq, T. (2004).
Physiological evidence for response inhibition in choice
reaction time tasks. Brain and Cognition, 56, 153–164. https://
doi.org/10.1016/j.bandc.2004.06.004, PubMed: 15518932
Carbonnell, L., Ramdani, C., Meckler, C., Burle, B., Hasbroucq,
T., & Vidal, F. (2013). The N-40: An electrophysiological
marker of response selection. Biological Psychology, 93,
231–236. https://doi.org/10.1016/j.biopsycho.2013.02.011,
PubMed: 23454519
Correa, Á., Cappucci, P., Nobre, A. C., & Lupiáñez, J. (2010).
The two sides of temporal orienting: Facilitating perceptual
selection, disrupting response selection. Experimental
Psychology, 57, 142–148. https://doi.org/10.1027/1618-3169
/a000018, PubMed: 20178944
Correa, Á., Lupiáñez, J., & Tudela, P. (2006). The attentional
mechanism of temporal orienting: Determinants and
attributes. Experimental Brain Research, 169, 58–68. https://
doi.org/10.1007/s00221-005-0131-x, PubMed: 16273403
Coull, J. T., Frith, C. D., Büchel, C., & Nobre, A. C. (2000).
Orienting attention in time: Behavioural and
neuroanatomical distinction between exogenous and
endogenous shifts. Neuropsychologia, 38, 808–819. https://
doi.org/10.1016/S0028-3932(99)00132-3, PubMed: 10689056
Coull, J. T., & Nobre, A. C. (1998). Where and when to pay
attention: The neural systems for directing attention to spatial
locations and to time intervals as revealed by both PET and
fMRI. Journal of Neuroscience, 18, 7426–7435. https://doi
.org/10.1523/jneurosci.18-18-07426.1998, PubMed: 9736662
De Jong, R., Coles, M. G. H., Logan, G. D., & Gratton, G. (1990).
In search of the point of no return: The control of response
processes. Journal of Experimental Psychology: Human
Perception and Performance, 16, 164–182. https://doi.org/10
.1037/0096-1523.16.1.164, PubMed: 2137517
Doherty, J. R., Rao, A., Mesulam, M. M., & Nobre, A. C. (2005).
Synergistic effect of combined temporal and spatial
expectations on visual attention. Journal of Neuroscience,
25, 8259–8266. https://doi.org/10.1523/JNEUROSCI.1821-05
.2005, PubMed: 16148233
Duque, J., Greenhouse, I., Labruna, L., & Ivry, R. B. (2017).
Physiological markers of motor inhibition during human
behavior. Trends in Neurosciences, 40, 219–236. https://doi
.org/10.1016/j.tins.2017.02.006, PubMed: 28341235
Durup, H., & Requin, J. (1970). Hypothèses Sur le rôle des
probabilités conditionnelles du signal d’exécution dans le
temps de réaction simple. Psychologie Française, 15, 37–46.
Eagle, D. M., Bari, A., & Robbins, T. W. (2008). The
neuropsychopharmacology of action inhibition:
Cross-species translation of the stop-signal and go/no-go
tasks. Psychopharmacology, 199, 439–456. https://doi.org/10
.1007/s00213-008-1127-6, PubMed: 18542931
Eimer, M. (1999). Facilitatory and inhibitory effects of masked
prime stimuli on motor activation and behavioural
performance. Acta Psychologica, 101, 293–313. https://doi
.org/10.1016/s0001-6918(99)00009-8, PubMed: 10344189
Elithorn, A., & Lawrence, C. (1955). Central inhibition—Some
refractory observations. Quarterly Journal of Experimental
Psychology, 7, 116–127. https://doi.org/10.1080
/17470215508416684
Gevins, A. (1989). Dynamic functional topography of cognitive
tasks. Brain Topography, 2, 37–56. https://doi.org/10.1007
/BF01128842, PubMed: 2641474
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A.,
Strohmeier, D., Brodbeck, C., et al. (2013). MEG and EEG
Korolczuk et al.
897
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
data analysis with MNE-python. Frontiers in Neuroscience,
7, 267. https://doi.org/10.3389/fnins.2013.00267, PubMed:
24431986
Gratton, G. (1998). The contralateral organization of visual
memory: A theoretical concept and a research tool.
Psychophysiology, 35, 638–647. https://doi.org/10.1017
/S0048577298980453, PubMed: 9844426
Gratton, G., Coles, M. G. H., Sirevaag, E. J., Eriksen, C. W., &
Donchin, E. (1988). Pre- and Poststimulus activation of
response channels: A psychophysiological analysis. Journal
of Experimental Psychology: Human Perception and
Performance, 14, 331–344. https://doi.org/10.1037/0096-1523
.14.3.331, PubMed: 2971764
Griffin, I. C., Miniussi, C., & Nobre, A. C. (2002). Multiple
mechanisms of selective attention: Differential modulation
of stimulus processing by attention to space or time.
Neuropsychologia, 40, 2325–2340. https://doi.org/10.1016
/S0028-3932(02)00087-8, PubMed: 12417462
Hasbroucq, T., Akamatsu, M., Burle, B., Bonnet, M., & Possamaï,
C. A. (2000). Changes in spinal excitability during choice
reaction time: The H-reflex as a probe of information
transmission. Psychophysiology, 37, 385–393. https://doi.org
/10.1017/S0048577200982052, PubMed: 10860416
Hodges, P. W., & Bui, B. H. (1996). A comparison of computer-
based methods for the determination of onset of muscle
contraction using electromyography. Electroencephalography
and Clinical Neurophysiology, 101, 511–519. https://doi.org
/10.1016/s0013-4694(96)95190-5, PubMed: 9020824
Kayser, J., & Tenke, C. E. (2015). Issues and considerations for
using the scalp surface Laplacian in EEG/ERP research: A
tutorial review. International Journal of Psychophysiology,
97, 189–209. https://doi.org/10.1016/j.ijpsycho.2015.04.012,
PubMed: 25920962
Korolczuk, I., Burle, B., & Coull, J. T. (2018). The costs and
benefits of temporal predictability: Impaired inhibition of
prepotent responses accompanies increased activation of
task-relevant responses. Cognition, 179, 102–110. https://doi
.org/10.1016/j.cognition.2018.06.006, PubMed: 29935426
Korolczuk, I., Burle, B., Coull, J. T., & Smigasiewicz, K. (2020).
Mechanisms of impulsive responding to temporally
predictable events as revealed by electromyography.
Neuroscience, 428, 13–22. https://doi.org/10.1016/j
.neuroscience.2019.12.022, PubMed: 31918009
Korolczuk, I., Burle, B., Coull, J. T., & Śmigasiewicz, K. (2022).
Time for action: Neural basis of the costs and benefits of
temporal predictability for competing response choices.
Journal of Cognitive Neuroscience, 34, 273–289. https://doi
.org/10.1162/jocn_a_01795, PubMed: 34813659
Lamar, M., Cutter, W. J., Rubia, K., Brammer, M., Daly, E. M.,
Craig, M. C., et al. (2009). 5-HT, prefrontal function and
aging: fMRI of inhibition and acute tryptophan depletion.
Neurobiology of Aging, 30, 1135–1146. https://doi.org/10
.1016/j.neurobiolaging.2007.09.013, PubMed: 18061310
Law, S. K., Rohrbaugh, J. W., Adams, C. M., & Eckardt, M. J. (1993).
Improving spatial and temporal resolution in evoked EEG
responses using surface Laplacians. Electroencephalography
and Clinical Neurophysiology/Evoked Potentials, 88,
309–322. https://doi.org/10.1016/0168-5597(93)90055-T,
PubMed: 7688286
Liu, J., & Liu, Q. (2016). Use of the integrated profile for
voluntary muscle activity detection using EMG signals with
spurious background spikes: A study with incomplete spinal
cord injury. Biomedical Signal Processing and Control, 24,
19–24. https://doi.org/10.1016/j.bspc.2015.09.004
Logan, G. D. (1994). On the ability to inhibit thought and
action: A user’s guide to the stop-signal paradigm. In
Inhibitory processes in attention, memory, and language,
(pp. 135–140). Academic Press.
Logan, G. D., Cowan, W. B., & Davis, K. A. (1984). On the ability
to inhibit simple and choice reaction time responses: A
model and a method. Journal of Experimental Psychology:
Human Perception and Performance, 10, 276–291.
https://doi.org/10.1037/0096-1523.10.2.276, PubMed: 6232345
Luce, R. D. (1986). Response times: Their role in inferring
elementary mental organization. New York: Oxford
University Press. https://doi.org/10.1093/acprof:oso
/9780195070019.001.0001
Menceloglu, M., Suzuki, S., & Song, J. H. (2021). Revealing the
effects of temporal orienting of attention on response
conflict using continuous movements. Attention, Perception,
and Psychophysics, 83, 1463–1478. https://doi.org/10.3758
/s13414-020-02235-4, PubMed: 33442827
Miniussi, C., Wilding, E. L., Coull, J. T., & Nobre, A. C. (1999).
Orienting attention in time: Modulation of brain potentials.
Brain, 122, 1507–1518. https://doi.org/10.1093/brain/122.8
.1507, PubMed: 10430834
Mostofsky, S., & Simmonds, D. (2008). Response inhibition and
response selection: Two sides of the same coin. Journal of
Cognitive Neuroscience, 20, 751–761. https://doi.org/10.1162
/jocn.2008.20500, PubMed: 18201122
Nobre, A. C. (2001). Orienting attention to instants in time.
Neuropsychologia, 39, 1317–1328. https://doi.org/10.1016
/S0028-3932(01)00120-8, PubMed: 11566314
Nobre, A. C., & van Ede, F. (2018). Anticipated moments:
Temporal structure in attention. Nature Reviews
Neuroscience, 19, 34–48. https://doi.org/10.1038/nrn.2017
.141, PubMed: 29213134
Orgogozo, J. M., & Larsen, B. (1979). Activation of the
supplementary motor area during voluntary movement in
man suggests it works as a supramotor area. Science, 206,
847–850. https://doi.org/10.1126/science.493986, PubMed:
493986
Peirce, J. W. (2007). PsychoPy—Psychophysics software in
Python. Journal of Neuroscience Methods, 162, 8–13.
https://doi.org/10.1016/j.jneumeth.2006.11.017, PubMed:
17254636
Peirce, J., Gray, J. R., Simpson, S., MacAskill, M., Höchenberger,
R., Sogo, H., et al. (2019). PsychoPy2: Experiments in
behavior made easy. Behavior Research Methods, 51,
195–203. https://doi.org/10.3758/s13428-018-01193-y,
PubMed: 30734206
Perrin, F., Pernier, J., & Bertrand, O. (1989). Spherical splines
for scalp potential and current density mapping.
Electroencephalography and Clinical Neurophysiology, 72,
184–187. https://doi.org/10.1016/0013-4694(89)90180-6,
PubMed: 2464490
Ridderinkhof, R. K., Forstmann, B. U., Wylie, S. A., Burle, B., &
van den Wildenberg, W. P. M. (2011). Neurocognitive
mechanisms of action control: Resisting the call of the sirens.
Wiley Interdisciplinary Reviews: Cognitive Science, 2,
174–192. https://doi.org/10.1002/wcs.99, PubMed: 26302009
Rohenkohl, G., Gould, I. C., Pessoa, J., & Nobre, A. C. (2014).
Combining spatial and temporal expectations to improve
visual perception. Journal of Vision, 14, 8. https://doi.org/10
.1167/14.4.8, PubMed: 24722562
Santello, M., & McDonagh, M. J. (1998). The control of timing
and amplitude of EMG activity in landing movements in
humans. Experimental Physiology, 83, 857–874. https://doi
.org/10.1113/expphysiol.1998.sp004165, PubMed: 9782194
Uusitalo, M. A., & Ilmoniemi, R. J. (1997). Signal-space
projection method for separating MEG or EEG into
components. Medical and Biological Engineering and
Computing, 35, 135–140. https://doi.org/10.1007
/BF02534144, PubMed: 9136207
Van Boxtel, G. J. M., Van der Molen, M. W., Jennings, J. R., &
Brunia, C. H. M. (2001). A psychophysiological analysis of
898
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
inhibitory motor control in the stop-signal paradigm.
Biological Psychology, 58, 229–262. https://doi.org/10.1016
/S0301-0511(01)00117-X, PubMed: 11698116
van Ede, F., Rohenkohl, G., Gould, I., & Nobre, A. C. (2020).
Purpose-dependent consequences of temporal expectations
serving perception and action. Journal of Neuroscience,
40, 7877–7886. https://doi.org/10.1523/JNEUROSCI.1134-20
.2020, PubMed: 32900836
Van Elswijk, G., Kleine, B. U., Overeem, S., & Stegeman, D. F. (2007).
Expectancy induces dynamic modulation of corticospinal
excitability. Journal of Cognitive Neuroscience, 19, 121–131.
https://doi.org/10.1162/jocn.2007.19.1.121, PubMed: 17214569
Verbruggen, F., & Logan, G. D. (2008). Response inhibition in
the stop-signal paradigm. Trends in Cognitive Sciences, 12,
418–424. https://doi.org/10.1016/j.tics.2008.07.005, PubMed:
18799345
Vidal, F., Burle, B., Grapperon, J., & Hasbroucq, T. (2011). An
ERP study of cognitive architecture and the insertion of
mental processes: Donders revisited. Psychophysiology, 48,
1242–1251. https://doi.org/10.1111/j.1469-8986.2011.01186.x,
PubMed: 21342207
Vidal, F., Grapperon, J., Bonnet, M., & Hasbroucq, T. (2003).
The nature of unilateral motor commands in between-hand
choice tasks as revealed by surface Laplacian estimation.
Psychophysiology, 40, 796–805. https://doi.org/10.1111/1469
-8986.00080, PubMed: 14696733
Volberg, G., & Thomaschke, R. (2017). Time-based expectations
entail preparatory motor activity. Cortex, 92, 261–270.
https://doi.org/10.1016/j.cortex.2017.04.019, PubMed:
28544941
Wang, W., Hu, S., Ide, J. S., Zhornitsky, S., Zhang, S., Yu, A. J.,
et al. (2018). Motor preparation disrupts proactive control
in the stop signal task. Frontiers in Human Neuroscience,
12, 151. https://doi.org/10.3389/fnhum.2018.00151, PubMed:
29780308
Woolsey, C. N., Settlage, P. H., Meyer, D. R., Spencer, W.,
Hamuy, T. P., & Travis, A. M. (1952). Patterns of localisation
in precentral and “supplementary” motor areas and their
relation to the concept of a premotor area. Research
Publications—Association for Research in Nervous and
Mental Disease, 30, 238–264. PubMed: 12983675
Zhang, R., Geng, X., & Lee, T. M. C. (2017). Large-scale
functional neural network correlates of response inhibition:
An fMRI meta-analysis. Brain Structure and Function, 222,
3973–3990. https://doi.org/10.1007/s00429-017-1443-x,
PubMed: 28551777
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
8
8
5
2
0
7
7
7
2
8
/
j
o
c
n
_
a
_
0
1
9
7
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Korolczuk et al.
899