“Virus and Epidemic”: Causal Knowledge
Activates Prediction Error Circuitry
Daniela B. Fenker1, Mircea A. Schoenfeld1, Michael R. Waldmann2,
Hartmut Schuetze1, Hans-Jochen Heinze1, and Emrah Duezel3,4
Abstrakt
■ Knowledge about cause and effect relationships (z.B., virus–
epidemic) is essential for predicting changes in the environment
and for anticipating the consequences of events and oneʼs own ac-
tionen. Although there is evidence that predictions and learning from
prediction errors are instrumental in acquiring causal knowledge, Es
is unclear whether prediction error circuitry remains involved in the
mental representation and evaluation of causal knowledge already
stored in semantic memory. In an fMRI study, participants assessed
whether pairs of words were causally related (z.B., virus–epidemic)
or noncausally associated (z.B., emerald–ring). In a second fMRI
Studie, a task cue prompted the participants to evaluate either the
causal or the noncausal associative relationship between pairs of
Wörter. Causally related pairs elicited higher activity in OFC, amyg-
dala, striatum, and substantia nigra/ventral tegmental area than non-
causally associated pairs. These regions were also more activated by
the causal than by the associative task cue. This network overlaps
with the mesolimbic and mesocortical dopaminergic network
known to code prediction errors, suggesting that prediction error
processing might participate in assessments of causality even under
conditions when it is not explicitly required to make predictions. ■
EINFÜHRUNG
The knowledge of the specific relationship between a
cause and an effect (z.B., virus–epidemic) and its disso-
ciation from noncausally associated events (z.B., virus–
bacteria) enables predicting consequences of events and
Aktionen. Since the pioneering work of Pavlov (1927), an ex-
tensive range of theoretical and experimental work has
addressed the question how animals and humans learn
to associate a cue with an outcome (Cobos, López, Caño,
Almaraz, & Shanks, 2002; Shanks & López, 1996) or link a
cause to an effect (Blaisdell, Sawa, Leising, & Waldmann,
2006; Waldmann, 1996, 2000, 2001; Waldmann, Holyoak,
& Fratianne, 1995; Waldmann & Holyoak, 1992), enabling
them to guide their behavior accordingly. There is con-
verging evidence that processing prediction errors is one
candidate mechanism driving such learning of causality
(Pearce & Hall, 1980; Rescorla & Wagner, 1972). Learning
is enabled because expectations regarding outcomes (z.B.,
reward or punishment) are updated following predic-
tion errors until expectations and outcomes eventually
converge.
Evidence from animal studies indicates that a desig-
nated neural network is critical for the ability to predict
reward or punishment, to compute prediction errors,
and to adjust response selection and goal-directed behav-
1Otto-von-Guericke University, Magdeburg, Deutschland, 2Universität
of Göttingen, Deutschland, 3Institute of Cognitive Neuroscience Uni-
versity College London, 4Institute of Cognitive Neurology and
Dementia Research, Magdeburg, Deutschland
ior on the basis of such errors. This network includes
the medial pFC (Ostlund & Balleine, 2005; Matsumoto
& Tanaka, 2004; Matsumoto, Suzuki, & Tanaka, 2003),
the OFC (Wallis, 2007; Izquierdo, Suda, & Murray, 2004),
the amygdala (Balleine, Killcross, & Dickinson, 2003), Die
striatum/nucleus accumbens (Balleine, Delgado, & Hikosaka,
2007; Cromwell, Hassani, & Schultz, 2005; de Borchgrave,
Rawlins, Dickinson, & Balleine, 2002; Schultz, Tremblay, &
Holland, 1998), and the thalamus (Corbit, Muir, & Balleine,
2003) and is closely linked to dopaminergic (DA) neuro-
modulation (Schultz, 2002, 2006; Andrzejewski, Spencer, &
Kelley, 2005). In humans, components of this network have
been implicated in making predictions for desired outcomes
and computing prediction errors (Knutson & Wimmer, 2007;
Knutson, Taylor, Kaufman, Peterson, & Glover, 2005; Breiter,
Aharon, Kahneman, Dale, & Shizgal, 2001; Berns, McClure,
Pagnoni, & Montague, 2001), in classification learning
(Rodriguez, Aron, & Poldrack, 2006), in associative causal
learning mechanisms (Corlett et al., 2004; Turner et al.,
2004; Fletcher et al., 2001), and in encoding causal effects
of actions (Tanaka, Balleine, & OʼDoherty, 2008). Darüber hinaus,
mesencephalic midbrain structures known to harbor DA
Neuronen (substantia nigra/ventral tegmental area, SN/ VTA)
are activated during the encoding of a stimulus predicting
monetary reward. Such an activation pattern has recently
been shown to facilitate long-term memory for that stimulus
(Wittmann et al., 2005).
The functional anatomical circuitry underlying predic-
tion error learning has been most intensively studied for
reward prediction errors (Schultz, 2004). During unexpected
© 2009 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 22:10, S. 2151–2163
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
e
1
0
–
P
2
D
1
F
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
Ö
5
C
1
N
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
3
8
0
7
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
/
T
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
primary (z.B., food) or secondary (z.B., money) rewards, Die
nucleus accumbens receives DA signals from the SN/ VTA
of the midbrain. Once a reward becomes predictable by a
conditioned (d.h., reward-predicting) stimulus (CS+), Die
DA signal in the nucleus accumbens shifts forward from
the time of reward outcome to the presentation of the
CS+ (OʼDoherty, Hampton, & Hackjin, 2007; OʼDoherty,
Dayan, Friston, Critchley, & Dolan, 2003). Umgekehrt,
omission of a reward after presentation of a CS+ leads
to a decrease of DA signaling below baseline. It has thus
been hypothesized that the DA signal in the SN/ VTA and
in the nucleus accumbens code both reward prediction
and prediction error.
Jedoch, not only reinforcement learning paradigms
provide valuable insights into the mechanisms underlying
the acquisition of causal knowledge; associative causal
learning studies have shown that prediction error mecha-
nisms are also at play during the acquisition of cause–effect
relationships even in the absence of apparent reward
Manipulation (Corlett et al., 2004).
Both human and nonhuman animals store a rich data-
base of causal relationships. Knowledge of such causal re-
lationships is instrumental in selecting appropriate goals
and actions and, in a much broader perspective, in guiding
social interactions as well as in shaping our understanding
of natural phenomena. Given that the computation of pre-
diction error seems to be an important component in the
learning of causal contingencies, our study addresses the
questions whether brain regions associated with predic-
tion error processing participate in the representation of
causal knowledge in semantic memory.
To date, only a few studies have examined how already
acquired causal knowledge is represented and how it can
be dissociated from noncausal associative relationships in
human semantic memory (Fenker, Waldmann, & Holyoak,
2005; Satpute et al., 2005). These studies have shown that
causal relations are stored and accessed differently from
noncausal associative relations. Given the fact that we es-
tablish causal knowledge through life experience and that
prediction error processing is involved during the acquisi-
tion of causal knowledge (Corlett et al., 2004; Turner et al.,
2004), it is plausible that cause–effect relationships stored
in semantic memory are represented differently from non-
causal associations, in a way that allows updating cause–
effect contingency changes, hence allowing modification
by experience. According to this possibility, every time
we encounter a causal relationship, prediction error cir-
cuitry is activated by default, allowing the modification of
cause–effect relationships stored in semantic memory if
a prediction error occurs due to changed contingencies.
Im Gegensatz, noncausal associative relations (z.B., emerald–
ring) should not engage prediction error circuitry.
The key question addressed here is whether prediction
error circuitry is a part of the representation of causal se-
mantic memories. An alternative possibility is that causal
Beziehungen, once acquired and part of our semantic
Wissen, are stored and represented just like any other
associative relationship (hence in a more “static” fashion),
thereby reflecting the fact that we have learned extensively
that certain causes determine specific effects. Updating
this knowledge may thus not be necessary, and hence it
may not be the case that prediction error circuitry is en-
gaged whenever we encounter virus and epidemic to-
gether, thus making the representations of well-learned
causal knowledge undistinguishable from well-learned
noncausal associative relationships.
The dissociation of causal and noncausal associative rela-
tionships stored in semantic memory was investigated in two
event-related fMRI experiments. We examined the potential
involvement of prediction error circuitry in the retrieval of
already existing causal in contrast to noncausal associative
semantic knowledge. In our design, daher, we deliber-
ately avoided standard learning tasks that are already
known to recruit prediction error circuitry. Somit, partici-
pants were not explicitly required to make predictions.
In the first experiment, word pairs denoting a cause–
effect relationship were simply contrasted with pairs de-
noting a noncausal associative relationship. In dieser Sekunde
Experiment, we compared the task-related preparation for
the retrieval of causal and noncausal associative relation-
ships. The words of each pair were shown one after the other
and they could either be unrelated (z.B., door–pinball), nicht-
causal associatively related (related in the absence of a causal
relationship e.g., emerald–ring), or causally related (z.B.,
virus–epidemic). In Experiment 1, participants had to evalu-
ate the presented word pairs for a causal relationship. Im
Experiment 2, the word pairs had to be evaluated either for a
causal relationship or for a noncausal associative relationship
indicated by a verbal cue presented before each pair.
If causal relationships are represented in a similar fash-
ion as noncausal associative relationships, the retrieval of a
cause–effect relationship from semantic memory should
only differ in terms of the semantic meaning and would
be reflected mainly in semantic memory regions, wie zum Beispiel
prefrontal (Noppeney, Phillips, & Price, 2004; Nyberg
et al., 2003), zeitlich (Patterson, Nestor, & Rogers, 2007;
Rogers et al., 2006; Noppeney et al., 2004), and parietal cor-
tices (Rogers et al., 2006; Wiggs, Weisberg, & Martin, 1999).
Im Gegensatz, if cause–effect relationships are represented as
contingency-based relationships, their retrieval and evalua-
tion could involve also updating processes that would be
reflected in neural activation going beyond semantic pro-
cessing of noncausal associations by involving mesolimbic
and mesocortical circuitry known to compute prediction
errors during learning.
METHODEN
Experiment 1
Teilnehmer
Fifteen participants (age range between 19 Und 30 Jahre
alt, 12 Frauen) with normal or corrected-to-normal vision
participated in the study. They were paid A12 and gave
2152
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
e
1
0
–
P
2
D
1
F
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
Ö
5
C
1
N
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
3
8
0
7
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
.
F
.
.
T
/
Ö
N
1
8
M
A
j
2
0
2
1
their written consent. All subjects were right-handed ac-
cording to self-report and had no history of neurological ill-
ness. The study was approved by the ethics committee of
the Otto-von-Guericke University, Magdeburg.
Stimuli
The stimuli were 210 German word pairs of 4- to 13-letter
Wörter. These word pairs were translated into German from
the English word pairs used by Fenker et al. (2005). The orig-
inal English causal and noncausal associative word pairs
were selected from the University of South Florida (USF)
Word Association Norm list (Nelson, McEvoy, & Schreiber,
1998) with a forward and backward strength <0.01. In ad-
dition, a norming study was used to select causally related
item pairs equated in both directions in terms of the
strength of statistical relations (e.g., frequency of occur-
rence of epidemic, given a virus versus frequency of occur-
rence of a virus given epidemic; Fenker et al., 2005). The
German words were presented in white Arial 28-point font
on black background via a custom projection system.
Eighty-four (84) word pairs were causally related (e.g.,
virus–epidemic), 42 noncausal associatively related (e.g.,
ring–emerald) and 84 unrelated (e.g., door–pinball).
After ensuring that participants understood the instruc-
tions, they were placed in the scanner. Participants were
asked to fixate on a central dot. In each trial, the first word
replaced the fixation dot for 1 sec and then was replaced by
the second word for 1 sec followed by fixation. The ISIs
between trials were jittered between 4 and 8 sec in 2-sec
steps (mean ISI = 4.67 sec). If a word pair was causally re-
lated, participants pressed a button with the right index
finger, and if a word pair was noncausal associatively re-
lated or unrelated, they responded with the left index
finger. The response hands were counterbalanced over
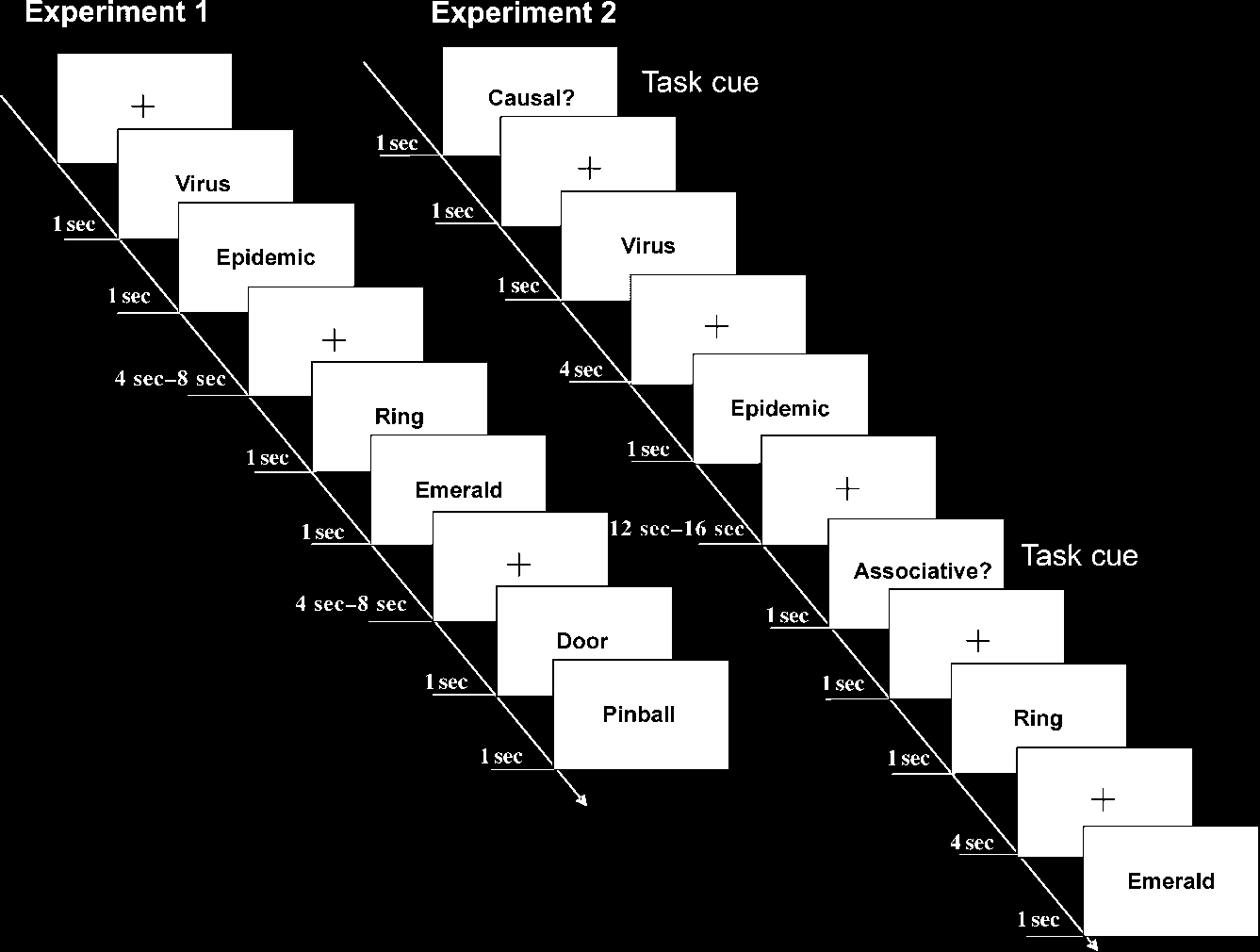
participants (Figure 1, left panel).
D
o
w
n
l
o
a
d
e
d
fMRI Image Acquisition
Images were acquired on a neurooptimized General Electric
Signa LX 1.5T system. Whole-head fMRI data were acquired,
with 23 slices (matrix = 64 × 64; field of view = 22 cm; slice
thickness = 5 mm; 1 mm gap; orientation AC–PC), using an
echo-planar gradient-echo sequence (repetition time/echo
time/flip angle = 2000 msec/35 msec/80°, ramp sampling
off ). The data were collected over two runs each lasting
approximately 8.5 min (253 volumes).
Procedure
Participants were given written instructions before scan-
ning. A causal relation was defined as follows: “the event
described by the first word causes or is caused by the event
described by the second word.” A noncausal associative
relation was defined as follows: “meaningful relationship
between the two events, but not a causal relationship.”
f MRI Image Analysis
Data were time sliced, realigned, normalized to the Mon-
treal Neurological Institute (MNI) template, and spatially
smoothed (Gaussian kernel, 8 mm). Statistical analysis
used the standard hemodynamic response function and
the movement parameters serving as regressors in the
event-related design for each subject (SPM99, Wellcome
Department of Imaging Neuroscience, London). For group
Figure 1. Examples of trials
and timing. Left: Experiment 1,
causal trial followed by a
noncausal associative trial.
Right: Experiment 2, causal
task cue followed by a
causally related word pair and
associative task cue followed
by a noncausal associative
word pair.
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
e
1
0
-
p
2
d
1
f
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
o
5
c
1
n
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
8
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
.
.
f
t
.
o
n
1
8
M
a
y
2
0
2
1
Fenker et al.
2153
analyses, we entered contrast images into one-sample
t tests, treating subjects as a random variable. SPM contrasts
were calculated using a threshold of p < .001 and a cluster
size criterion of >10 voxels. All reported coordinates refer
to MNI space. SPM group contrast images are depicted on
single-subject T1-weighted slices from MRIcro (www.cla.sc.
edu/psyc/faculty/rorden/mricro.html). Zusätzlich, to assess
differences between the three conditions (d.h., type of rela-
tionship), we performed an ROI analysis for the left amyg-
dala, left SN/ VTA, and bilateral nucleus accumbens. Der
ROIs were functionally defined on the basis of the compar-
ison causal versus noncausal associative word pairs. Der
beta values of the peak voxel were extracted for each partic-
ipant and the three conditions (causally related, noncausal
associatively related, and unrelated word pairs; Abbildung 3B).
Experiment 2
Teilnehmer
Eighteen healthy right-handed participants (14 Frauen),
ranging in age from 19 Zu 27 Jahre, took part in the experi-
ment and were paid A24. They all had to fulfill the same re-
quirements as the participants in the Experiment 1. Der
samples of Experiments 1 Und 2 were independent.
Stimuli
The stimuli were 252 German word pairs of length be-
zwischen 4 Und 13 letters and were presented in white Arial
28-point font on black background via a custom projection
System. The word pairs were the same as in Experiment 1,
Aber 42 noncausal associatively related word pairs were
added to account for stimulus balancing.
Verfahren
Participants were given written instructions before scan-
ning. In the scanner, each trial began with the presenta-
tion of the cue, which was either the word “Associative?”
or the word “Causal?.” The cue was presented for 1 Sek,
followed by a 1-sec fixation and the first word at fixation for
1 Sek. Between the presentation of the first and the second
word, fixation was shown for 4 Sek. The ISI between the
cues ranged from 12 Zu 16 Sek (in steps of 2 Sek) with a
mean ISI of 12.9 Sek ( Figur 1, right panel). Teilnehmer
were instructed to determine if the relationship between
the two words match the cue, das ist, if the word pairs de-
scribe a noncausal associative relationship (associative
cue) or a causal relationship (causal cue). The definition
of the relationships was identical to Experiment 1. Half
of the pairs were presented with the associative cue and
the other half with the causal cue. This combination was
counterbalanced across participants; das ist, word pairs
presented with the associative cue for a subject were pre-
sented with the causal cue for another subject and vice versa.
Participants responded by pressing a button with the left or
right index finger for matching and nonmatching, bzw-
aktiv. The response fingers were counterbalanced across
Teilnehmer. Response classes for correct responses were
“causal–causal” for causal cue and causal relation ( yes re-
sponse), “associative–associative” for associative cue and
noncausal associative relation ( yes response), “causal–
associative” for causal cue and noncausal associative rela-
tion (no response), “associative–causal” for associative cue
and causal relation ( yes response), “causal–unrelated” for
causal cue and unrelated word pairs (no response), Und
“associative–unrelated” for associative cue (no response).
fMRI Image Acquisition
The parameters of the image acquisition were the same
as in Experiment 1. The data were collected over six sep-
arate runs each lasting approximately 9.5 min, ergebend
the acquisition of 285 volumes.
fMRI Image Analysis
The functional data of Experiment 2 were analyzed using
SPM99 ( Wellcome Department of Imaging Neuroscience,
London) using the same parameters as in Experiment 1.
To assess brain activation separately for “type of cue” and
“type of relationship,” we performed a multiple regres-
sion analysis for each single subject (Postle, 2005; Schon,
Hasselmo, Lopresti, Tricarico, & Stern, 2004; Postle, Berger,
Taich, & DʼEsposito, 2000). We created the following three
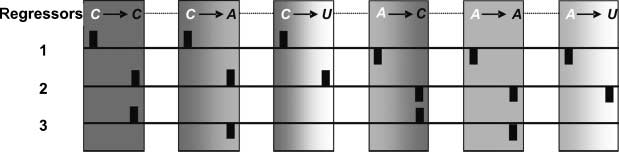
regressors (Figur 2): Regressor 1 assessed preparatory dif-
ference induced by the cues, Regressor 2 differentiated the
influence of the causal cue and the associative cue on rela-
tionship processing, and Regressor 3 assessed the differ-
ence between the causal relationship and the noncausal
associative relationship, regardless of the previous cue.
Given the fact that the results of a regressor cannot be inter-
preted independently from another regressor if they share a
significant amount of variance, we orthogonalized our three
regressors to minimize the shared variance. Erste, we as-
signed weight coefficients for each regressor such that
the internal sum of the products of the weights was zero.
Zweite, we made sure that the sum of the products of
the weight coefficients (for cue and word) of each regressor
pair was also zero (Bortz, 2005). For group analyses, we en-
tered the regressor images into one-sample t tests, treating
subjects as a random variable. SPM group contrast images
are depicted on single-subject T1-weighted slices from
MRIcro (www.cla.sc.edu/psyc/faculty/rorden/mricro.html)
with a threshold of p < .001, >10 voxels and p < .005,
>10 voxels, jeweils.
ERGEBNISSE
Experiment 1: Verhaltensergebnisse
We computed two separate ANOVAs over RTs and re-
sponse accuracy using the type of relationship (causal,
2154
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
e
1
0
–
P
2
D
1
F
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
Ö
5
C
1
N
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
3
8
0
7
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
/
T
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
Figur 2. Regressors of
Experiment 2. Regressor 1
assessed preparatory difference
induced by the cues. Regressor 2
differentiated the influence of
the causal and the associative
cue at the time of relationship
evaluation, and Regressor 3
assessed the relationship
evaluation between the causal
and the noncausal associative word pairs regardless of the cue. The white letters refer to the type of cue (c = causal; a = associative), and the black letters
refer to the type of relationship (c = causal; a = noncausal associative; u = unrelated). The three regressors were orthogonalized with respect to
each other; das ist, the sum of the products of the weight coefficient (for cue and word) of each regressor pair was zero.
noncausal associative, and unrelated) as within-subjects
factor. The RTs for the three conditions differed signifi-
cantly, F(2, 28) = 13.05, P < .01. A post hoc least significant
difference (LSD) test showed that participants responded
significantly slower to noncausal associative word pairs
than to causal word pairs and unrelated word pairs (all
ps < .01). Their response times did not differ between
the causal and the unrelated word pairs ( p > .05; Tisch 1).
Response accuracy did also differ significantly across con-
ditions, F(2, 28) = 18.67, P < .01. Participants gave signifi-
cantly more correct responses to the unrelated word pairs
than to the causal and to the noncausally associated word
pairs (all ps < .01). There was no significant difference in
response accuracy between the causally related and the
noncausally associated word pairs ( p > .05; Tisch 1).
fMRI Results
During the evaluation of noncausally associated word
pairs in contrast to unrelated word pairs, a network of
areas predominantly in the left hemisphere was activated.
Significant activations were observed in bilateral frontal
(left Brodmannʼs area [BA] 8, 46, 47 and right BA 9, 47),
left temporal (BA 37) and parietal areas (BA 40), right oc-
cipital regions (BA 18, 19), left thalamus, and left and right
caudate nucleus (Tisch 2). The presence of a causal rela-
tionship between the word pairs compared with unrelated
word pairs also elicited activity in a widespread network of
regions including bilateral frontal (left BA 8, 46 and right
BA 9, 47), left temporal (BA 37), left parietal (BA 40), links
posterior cingulate gyrus (BA 31), right occipital (BA 18)
and left and right caudate nucleus, and left amygdala
(Tisch 2). The differences of both evaluation processes
were substantiated by exclusively masking ( p = .05, für
the mask) the activity elicited by the comparison causal
versus unrelated with the comparison noncausal associa-
tive versus unrelated word pairs. During this masking pro-
cedure, all voxels that reach the default level of significance
in the masking contrast will be removed. The remaining
Aktivierung (causal vs. unrelated word pairs) is devoid of
the activation found in the masking contrast (noncausal
associative vs. unrelated word pairs) and therefore dem-
onstrates the distinct processing of causal relations.
daher, significant activations reflected those regions
that were exclusively activated by the presence of a causal
relationship resulting in a network of areas predominantly
located in left hemisphere including in middle frontal
(BA 6, 46), superior/medial frontal (BA 9) Regionen, superior
temporal regions (BA 22), left and right nucleus accumbens,
posterior cingulate gyrus (BA 31), amygdala, and SN/ VTA
(Figure 3A and B).
To differentiate the evaluation of cause–effect relation-
ships from the evaluation of noncausal associative relation-
ships, we directly contrasted the activation for causally
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
e
1
0
–
P
2
D
1
F
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
Ö
5
C
1
N
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
3
8
0
7
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
.
.
F
/
T
.
Tisch 1. Behavioral Data of the Two Experiments
Experiment 1
RTs
Accuracy
Causal
Relationship
1359 (73)
87 (2)
Noncausal Associative
Relationship
1510 (93)
83 (3)
Ö
N
1
8
M
A
j
2
0
2
1
Unrelated Word Pairs
1293 (83)
99 (1)
Experiment 2
Causal–Causal
Causal–Noncausal
Associative
Causal–
Unrelated
Associative–
Causal
Associative–Noncausal
Associative
Associative–
Unrelated
RTs
Accuracy
1949 (112)
2272 (122)
1560 (72)
1751 (92)
1945 (112)
86 (2)
66 (4)
97 (1)
93 (1)
80 (3)
1708 (81)
95 (1)
Mean RTs in milliseconds and accuracy in percent for all participants. Data in parentheses refer to SEMs. For Experiment 2, the top word refers to the
type of cue and the bottom word refers to the type of relationship.
Fenker et al.
2155
Tisch 2. Significant Activation Found in Experiments 1 Und 2
BA
z Value
X
j
z
MNI coordinates
Experiment 1
Causal vs. unrelated
Left caudate nucleus
Left inferior frontal
Left inferior parietal
Left insula
Left middle temporal
Left orbito-frontal
Left posterior cingulate
Left superior frontal/medial part
Left cerebellum
Right caudate
Right inferior frontal
Right lingual gyrus
Right middle frontal
Noncausal associative vs. unrelated
Left caudate
Left inferior frontal
Left inferior parietal
Left middle temporal
Left superior frontal/medial part
Left thalamus
Right caudate
Right inferior frontal
Right lingual gyrus
Right middle frontal
46
40
37
31
8
47
18
9
46
47
40
37
8
47
18
19
9
5.18
4.47
5.46
4.43
4.21
4.06
4.48
5.21
3.81
4.42
4.28
5.07
3.46
5.03
5.47
3.8
5.03
3.58
5.45
4.65
4.62
4.5
5.38
4.1
3.47
Causal vs. unrelated exclusively masked ( p = .05) by noncausal associative vs. unrelated
Left amygdala
Left caudate/nucleus accumbens
Left middle frontal
Left posterior cingulate
Left SN/ VTA
Left superior frontal/medial part
Left superior temporal
Right putamen/nucleus accumbens
6
46
31
9
22
5.44
6.2
5.72
4.83
5.71
3.57
5.06
6.33
4.85
−9
−39
−39
−51
−57
−12
−6
−3
−30
12
36
9
51
−9
−45
−30
−45
−60
−3
−3
12
54
6
21
51
−18
−6
−39
−45
−6
−9
−6
−54
15
9
39
−66
15
−51
6
−48
30
−69
12
24
−81
24
6
42
24
−57
−51
39
−27
12
21
−78
−51
24
0
−12
3
36
−45
−15
54
−60
12
6
6
45
0
−9
−18
30
54
−39
3
0
−18
42
6
9
0
45
−12
48
3
3
−6
−12
−3
42
−18
−3
36
24
30
−12
36
15
−3
2156
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
e
1
0
–
P
2
D
1
F
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
Ö
5
C
1
N
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
3
8
0
7
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
.
/
.
F
T
.
Ö
N
1
8
M
A
j
2
0
2
1
Tisch 2. (Fortsetzung )
Causal vs. noncausal associative
Left amygdala
Left inferior frontal
Left inferior parietal lobe
Left nucleus accumbens
Left posterior cingulate gyrus
Left SN/ VTA
Left superior frontal gyrus/medial part
Right inferior frontal
Right nucleus accumbens
Right orbito-frontal
Experiment 2
44
45
40
31
9
44
11
Cue: causal vs. associative, at cue presentation (Regressor 1)
Left amygdala/posterior orbito-frontal
Left anterior cingulate
Left caudate nucleus
Left cerebellum
Left insula/putamen
32
Left parahippocampal gyrus
35/36
Left thalamus
Right inferior frontal/posterior orbito-frontal
47/25
Right insula
Right parahippocampal gyrus
28/35
Right precuneus
Right putamen/globus pallidus
Right thalamus
Left SN/ VTA (cluster size >5 voxels)
Cue: causal vs. associative, at relational evaluation (Regressor 2)
Left amygdala/posterior orbito-frontal
Left inferior frontal
Left middle frontal
47
44
46
10
10
6/8
BA
z Value
X
5.06
3.83
3.69
4.01
4.53
4.33
4.86
4.67
3.92
4.05
3.53
4.43
3.83
3.73
3.61
3.53
3.71
4.13
3.65
3.61
3.56
3.94
3.38
3.91
4.06
4.02
3.75
3.62
3.39
3.29
3.64
3.73
−18
−54
−51
−60
−6
−3
−6
−6
63
12
3
−18
−6
−15
−15
−27
−27
−9
24
45
24
15
15
6
−9
−21
−39
−54
−42
−33
−42
−42
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
e
1
0
–
P
2
D
1
F
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
Ö
5
C
1
N
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
3
8
0
7
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
.
F
T
/
.
.
Ö
N
1
8
M
A
j
2
0
2
1
MNI coordinates
j
−3
6
30
−36
12
−15
−27
51
12
12
45
0
39
12
−45
18
−30
−6
12
6
−21
−66
3
−12
−15
3
39
12
42
51
51
15
z
−15
27
12
42
−6
45
−15
39
15
−6
−12
−12
15
3
−24
0
−18
0
−18
0
−21
24
6
12
−18
−15
−6
24
15
12
−6
48
Fenker et al.
2157
Tisch 2. (Fortsetzung )
Relational evaluation: causal vs. noncausal associative (Regressor 3)
BA
z Value
X
Cingulate gyrus
Left caudate nucleus
Left inferior parietal
Left middle frontal
Left middle temporal
Left precuneus
Left superior frontal
Left thalamus
Right cerebellum
Left orbito-frontal ( p = .005)
31
40
9
46
10
21
7
8
8
4.95
3.85
5.26
5
3.67
3.58
4.38
4.06
4.14
3.92
3.94
4.07
3.64
0
−9
−48
−42
−45
−42
−63
−6
−21
−15
−3
36
−15
MNI coordinates
j
−33
3
−66
18
36
51
−51
−78
33
45
−27
−72
6
z
45
6
42
42
18
9
0
45
54
45
3
−45
−18
If not stated otherwise in the table, the significance level was p = .001, uncorrected at a cluster size of at least 10 voxels. BA = Brodmannʼs area.
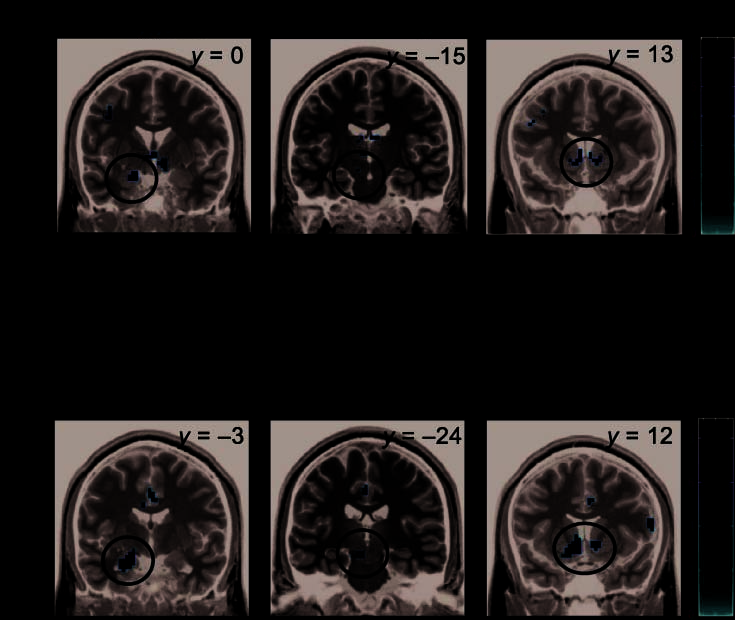
Figur 3. Activation found
during relationship evaluation
for Experiment 1. (A) SPM
contrast images of the contrast
causal versus unrelated word
pairs exclusively masked with
the contrast of noncausal
associative word pairs versus
unrelated word pairs. (B)
Results of the ROI analysis. Der
bars depict the difference of
the mean beta values, das ist,
causal minus unrelated (c − u),
noncausal associative minus
unrelated (a − u), and causal
minus noncausal associative
word pairs (c − a). Error bars
refer to the SEM. The gray bars
below the bilateral nucleus
accumbens activation refer to
the right and the black bars
refer to the left nucleus
accumbens activation. (C) SPM
contrast images for causal
versus noncausal associative
word pairs. The scales refer to
SPM t values.
2158
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
T
F
R
P
Ö
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
H
.
M
A
ich
R
e
.
D
C
u
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
e
1
0
–
P
2
D
1
F
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
Ö
5
C
1
N
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
3
8
0
7
7
.
S
P
e
D
P
F
e
M
B
j
B
e
R
G
u
2
0
e
2
S
3
T
/
J
F
T
/
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
versus noncausally associated word pairs. This comparison
revealed a higher activation for the presence of a causal re-
lationship in the left medial (BA 9) and inferior frontal gyri
(BA 44, BA 45), right inferior frontal gyrus (BA 44), Rechts
orbito-frontal (BA 11), left amygdala, left and right nucleus
accumbens, left posterior part of the SN/ VTA, left parietal
lobe (BA 40), and left cingulate gyrus (BA 31) ( Tisch 2,
Figure 3C).
Experiment 2: Verhaltensergebnisse
Behavioral data were analyzed with two separate repeated
measures ANOVAs applying type of cue (causal vs. associa-
tiv) and type of relationship (causal, noncausal associa-
tiv, and unrelated) to measures of RTs and response
accuracy. For the RTs, there was a significant main effect
of factor Type of Cue, F(1, 17) = 10.58, P < .01. The re-
sponses were faster for the associative cue. Factor Type
of Relationship also showed a significant main effect,
F(2, 34) = 42.12, p < .01. Participants gave the fastest re-
sponse to unrelated word pairs and slowest response to
noncausal associative word pairs. The interaction of the
two factors was also significant, F(2, 34) = 12.42, p < .01.
Post hoc (LSD) testing revealed that for the causal cue, the
responses to unrelated word pairs were the fastest and
the responses to noncausal associative word pairs were
the slowest ( ps < .05). For the associative cue, the re-
sponses to unrelated word pairs and causal word pairs
were significantly faster than the responses to noncausal
associated word pairs, ps < .05 (Table 1). The ANOVA
for the accuracy data revealed a significant main effect of
Type of Cue, F(1, 17) = 32.14, p < .01. Participants gave
more correct responses for the associative cue than for the
causal cue. The factor Type of Relationship was also signifi-
cant, F(2, 34) = 50.66, p < .01. Participants were most ac-
curate if the word pairs were unrelated, and their accuracy
was worst for noncausal associative word pairs. Finally, the
Type of Cue × Type of Relationship interaction was signifi-
cant as well, F(2, 34) = 6.81, p < .01. Post hoc testing (LSD)
showed for the causal cue that participants had the highest
accuracy for unrelated word pairs and the lowest for non-
causal associative word pairs, ps < .05. They also gave sig-
nificantly more correct response for causal and unrelated
word pairs presented after the associative cue in compari-
son to noncausal associative word pairs, ps < .05 (Table 1).
f MRI Results
In Experiment 2, statistical analyses for Regressors 2 and 3
were calculated to separate task-related and meaning-related
aspects of activity elicited by the word pairs themselves.
With Regressor 3, we contrasted causal with noncausal as-
sociative word pairs (Figure 2) compatible with the fMRI
analysis of Experiment 1. This showed a higher BOLD re-
sponse for causal word pairs in the left inferior parietal
lobe (BA 40), left posterior cingulate gyrus (BA 31), left
middle frontal (BA 9), left inferior frontal (BA 46), left mid-
dle temporal (BA 21), left superior frontal (BA 8), left
caudate nucleus, left thalamus, and at a more lenient thresh-
old ( p = .005) left OFC, regardless of the preceding cue
(Table 2, Figure 4A). To differentiate task-related aspects
of causal versus noncausal associative processing of word
pairs independent of type of word pair (collapsed over
causal, noncausal associative, and unrelated word pairs),
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
e
1
0
-
p
2
d
1
f
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
o
5
c
1
n
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
8
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
t
/
.
f
.
o
n
1
8
M
a
y
2
0
2
1
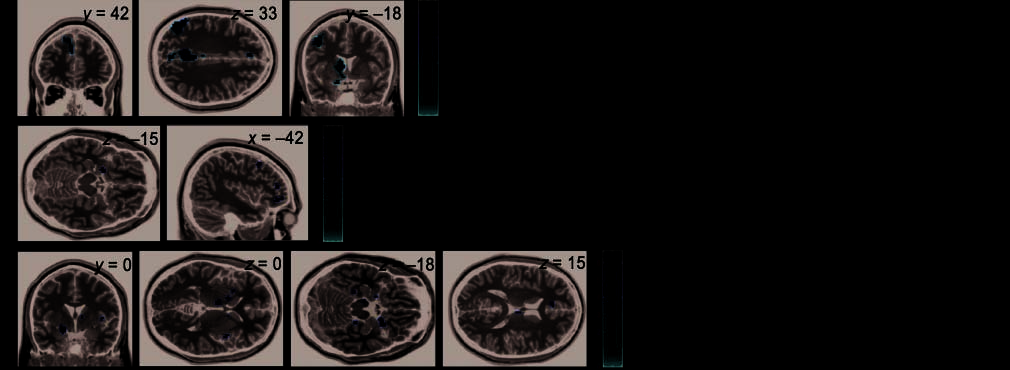
Figure 4. Significant activations of Experiment 2. Left side, activations found; right side, corresponding regressor. (A) SPM images for Regressor 3 (causal
vs. noncausal associative word pairs) show a significant activation in the left superior/medial/frontal cortex, left inferior parietal cortex, posterior cingulate
gyrus, left middle frontal cortex, left caudate nucleus, and left OFC ( p < .005). (B) Regressor 2 (influence of causal vs. associative cue on relationship
evaluation) yielded significant activation in the left amygdala/orbito-frontal region and left inferior and middle frontal gyri. The scales refer to SPM t values.
(C) Higher preparatory activation (Regressor 1: causal vs. associative cue) was found in the left amygdala, bilateral insula, bilateral thalamus, bilateral
caudate nucleus, bilateral parahippocampal gyrus, bilateral orbito-frontal, and left anterior cingulate. The scales refer to SPM t values.
Fenker et al.
2159
we compared the activation following the causal cue with
activation following the associative cue at the time when
the second word was presented (Figure 2, Regressor 2).
There was a stronger BOLD response following the causal
cue in the left amygdala, left inferior (BA 10, 44, 46, 47),
and middle frontal gyrus (BA 6, 8, 10), right middle frontal
(BA 8, 10), and left caudate nucleus (Table 2, Figure 4B).
Preparatory Activity
Experiment 1 investigated the dissociation between the
evaluation of a causal relationship and a noncausal associa-
tive relationship. Experiment 2 was designed to assess to
what extent activity patterns observed in Experiment 1
were related to the task demands of assessing causal rela-
tionships (Regressor 1). Therefore, activation elicited by
the causal cue was compared with the activation elicited
by the associative cue (Figure 2, Regressor 1) to investigate
different activations in terms of preparing for a causal or a
noncausal associative judgment. The preparation for a causal
judgment revealed higher activation in the left amygdala and
OFC, left and right caudate nucleus, left and right insula, left
thalamus, right OFC (BA 25/47), left SN/ VTA, left and right
parahippocampal gyrus (left BA 35, 36 and right BA 28/35),
left anterior cingulate gyrus (BA 32), right precuneus, and
the cerebellum (Table 2, Figure 4C).
DISCUSSION
In two event-related fMRI experiments, we sought to deter-
mine whether representations of cause–effect relationships
are distinct from noncausal associative relational representa-
tions. We focused on the retrieval of preexisting, well-learned
semantic causal knowledge. We show that the evaluation
of cause–effect relationships engages a mesolimbic and
mesocortical circuitry known to mediate prediction error
learning. This indicates that prediction error circuitry is en-
gaged when well-known causal associations are retrieved
from semantic memory even under conditions when pre-
diction error learning is not explicitly required. Such default
engagement of prediction error circuitry in the representa-
tion of well-known cause–effect relationships that are al-
ready stored in semantic memory may allow for efficient
updating to keep causal knowledge accurate.
Common Networks of Causal and Noncausal
Semantic Processing
Experiment 1 gave us the opportunity to investigate com-
monalities of semantic processing of noncausal associative
and causal relationships compared with semantic processing
of unrelated word pairs. These common networks in-
cluded areas, mainly left lateralized, in inferior parietal, in-
ferior and superior frontal, middle temporal, thalamus,
caudate nuclei, and occipital regions. These results are
compatible with previous studies which have found that
several of these areas are activated during semantic mem-
ory retrieval (Patterson et al., 2007; McDermott, Petersen,
Watson, & Ojemann, 2003; Nyberg et al., 2003; Cabeza &
Nyberg, 2000; Wiggs et al., 1999), selection of semantic in-
formation among competing alternatives (Thompson-
Schill, DʼEsposito, Aguirre, & Farah, 1997), and verbal
working memory maintenance or manipulation (Veltman,
Rombouts, & Dolan, 2003). It is unlikely that these activa-
tion differences between related (causal and noncausal as-
sociative) and unrelated word pairs were driven solely by
difficulty because there was no systematic RT or response
accuracy difference between related and unrelated pairs
across both experiments.
Dissociating Causal Relationships from
Noncausal Associative Relationships beyond
Semantic Processing
To derive a clear distinction between causal and noncau-
sal associative knowledge processing, we directly com-
pared the evaluation of causal and noncausal associative
word pairs. Activation in bilateral inferior, right orbito-
frontal, left superior frontal regions, left parietal regions,
left amygdala, and bilateral nucleus accumbens exclu-
sively differentiated causal relationships from noncausal
associative relationships. Activations in the OFC and the
left amygdala as well as bilateral nucleus accumbens con-
siderably overlap with regions from studies investigating
prediction error learning for rewards (Schultz, 2006;
Berns et al., 2001), contingency learning (Balleine et al.,
2003), and decision making processes (Hampton,
Adolphs, Tyszka, & OʼDoherty, 2007; Wallis, 2007; Yang
& Shadlen, 2007; Bechara, Damasio, & Damasio, 2003;
Krawczyk, 2002). As in Experiment 1, causal versus non-
causal associative relationships activated OFC and cau-
date (Regressor 3 in Experiment 2; however, unlike in
Experiment 1, we observed no amygdala activation for
this contrast in Experiment 2). This suggests that the ac-
tivity difference between causal and noncausal associative
relationships in both the orbito-frontal and the caudate
holds independent of task cues. The analysis of task
cue effect, on the other hand, showed that word pair pro-
cessing following a causal cue activated posterior left
OFC and amygdala irrespective of whether the word pairs
denoted causal, noncausal associative, or unrelated rela-
tionships. Therefore, the data from Regressors 2 and 3 in
Experiment 2 together with the data from Experiment 1
suggest that word pairs activate OFC and caudate more
strongly when they denote causal as opposed to noncausal
associative relationships. Irrespective of their meaning, word
pairs activate posterior OFC and amygdala more strongly
when word processing is directed toward causality rather
than noncausal associative relationships.
Thus, the contribution of Experiment 2 was that it al-
lowed us to investigate the pure task-related, prepara-
tory aspects of retrieving a cause–effect relationship by
2160
Journal of Cognitive Neuroscience
Volume 22, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
e
1
0
-
p
2
d
1
f
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
o
5
c
1
n
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
8
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
.
t
/
f
.
o
n
1
8
M
a
y
2
0
2
1
comparing activation specifically found at the time of the
task cue. Cues indicating to prepare to retrieve a causal
relationship as opposed to a noncausal associative rela-
tionship (Regressor 1) again elicited activation of a network
including mesolimbic and mesocortical structures (SN/ VTA
and amygdala), the caudate nucleus, and the OFC. This
cue-related finding effectively rules out the possibility that
the differences between causal and noncausal associative
relationships were driven solely by stimulus differences
on a semantic level.
Causal Knowledge and Prediction Error Circuitry
Our data show that the retrieval of cause–effect relationships
is distinguished from noncausal associative relationships by
engaging activity of brain regions such as the SN/ VTA and
ventral striatum, which are known to code prediction errors
during stimulus-reward learning (Schultz, 2007) and associa-
tive causal learning (Corlett et al., 2004). The amygdala and
OFC, key players in contingency learning and decision mak-
ing (Hampton et al., 2007), can also cooperatively contribute
to learning from prediction errors. The OFC can signal the
value of expected outcomes (Wallis, 2007), and these out-
come expectancies are held to permit the rapid recognition
of unexpected outcomes and prediction errors, thereby driv-
ing new learning through facilitation of associative flexibility
in downstream regions, such as the amygdala. Hence, this
set of structures together permits the representation of out-
comes, prediction errors and allows associative flexibility for
the updating of existing contingencies.
Involving this prediction error circuitry by default, that
is, in the absence of an explicit requirement to make pre-
dictions or learn from outcomes, during retrieval of well-
known causal relationships is a plausible mechanism that
would allow the updating of stored causal knowledge on
the basis of potential alterations of the cause–effect con-
nections. Such a mechanism seems suitable for keeping
causal knowledge accurate and maintaining knowledge
adaptable in changing environments. The results of Ex-
periment 2 also support this possibility and furthermore
suggest that even a potential encounter with a cause–
effect relationship involves activation of prediction error
circuitry.
A striking aspect of our data is that both experiments
did not actually allow for differences in explicit predic-
tions to contribute to our findings. That is in both experi-
ments, we did not explicitly manipulate prediction error
processing. In Experiment 1, the first word of each pair
was unpredictive as to whether the second word was re-
lated causally, noncausal associatively, or unrelated, and it
is therefore implausible to explain the findings here by a dif-
ferential engagement of participants in making explicit pre-
dictions about the second word of each pair. Likewise, in
Experiment 2, the cue itself did not allow for any explicit
predictions regarding the specific content of the upcoming
word pair. Hence, the finding that only causal relationships
in contrast to noncausal associative relationships engage
prediction error circuitry was unrelated to explicit differen-
tial engagement in making predictions regarding cause
and effect. Rather, the data suggest that, independent of
actual explicit predictions, the prediction error network
is activated by default when causality is an actual (Experi-
ment 1) or an expected (Experiment 2) part of stored se-
mantic relationships. One interesting difference between
the two is the involvement of the ventral striatum with
actual causality and the dorsal striatum with expected
causality.
DA neuromodulation is critical for the coding of pre-
diction errors of reward (Schultz, 2002, 2006; Schultz &
Dickinson, 2000; Schultz et al., 1998; Schultz, Dayan, &
Montague, 1997; Schultz, Apicella, Scarnati, & Ljungberg,
1992) but has not yet been implicated in the ability to
update already acquired causal knowledge in the absence
of any apparent reward-related reinforcement. Our data,
particularly the finding that SN/ VTA is activated by causal-
ity, raise the possibility that DA circuitry may indeed play
a role also in updating existing causal knowledge. This
leads to the interesting possibility that neurodegenerative
epidemics that are characterized by dysfunction of DA cir-
cuitry, such as schizophrenia and Parkinsonʼs epidemic,
might be associated with impaired updating of causal
knowledge leaving these patients less adaptive in changing
environments. Hence, the reported reasoning impairments
in schizotypy (Sellen, Oaksford, & Gray, 2005) as well as
schizophrenia (Moore & Sellen, 2006), but also prediction
error-dependent delusion formation in psychosis (Murray
et al., 2008; Corlett et al., 2007), and the recently demon-
strated problems of patients with Parkinsonʼs epidemic to
learn from prediction errors of rewards (Schott et al., 2007)
and their problems in counterfactual reasoning (McNamara,
Durso, Brown, & Lynch, 2003) may extend to a more gen-
eral impairment of modifying existing causal relationships
within semantic memory.
To summarize, we have shown that the representation
of causal knowledge is different from noncausal associative
knowledge and involves mesolimbic and mesocortical
structures that are part of a prediction error circuitry. Thus,
prediction error circuits are not only recruited during learn-
ing but also play a role in the representation of knowledge
that has already been acquired earlier. These findings shed
light on mechanisms that allow for a flexible updating of
already acquired causal knowledge. In more general terms,
they redefine the functional architecture of brain regions
that are likely to contribute to updating semantic memories.
Acknowledgments
This study was supported by grants from the Deutsche For-
schungsgemeinschaft (Klinische Forschergruppe “Kognitive
Kontrolle,” TP4) and the Volkswagen Foundation from the Uni-
versity of Magdeburg.
Reprint requests should be sent to Daniela B. Fenker, Department
of Neurology II, Otto-von-Guericke University, Leipziger Strasse 44,
39120 Magdeburg, Germany, or via e-mail: daniela.fenker@med.
ovgu.de.
Fenker et al.
2161
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
e
1
0
-
p
2
d
1
f
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
o
5
c
1
n
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
8
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
f
t
/
.
.
o
n
1
8
M
a
y
2
0
2
1
REFERENCES
Andrzejewski, M. E., Spencer, R. C., & Kelley, A. E. (2005).
Instrumental learning, but not performance, requires
dopamine D1-receptor activation in the amygdala.
Neuroscience, 135, 335–345.
Balleine, B. W., Delgado, M. R., & Hikosaka, O. (2007). The role
of the dorsal striatum in reward and decision-making.
Journal of Neuroscience, 27, 8161–8165.
Balleine, B. W., Killcross, A. S., & Dickinson, A. (2003). The
effect of lesions of the basolateral amygdala on instrumental
conditioning. Journal of Neuroscience, 23, 666–675.
Bechara, A., Damasio, H., & Damasio, A. R. (2003). Role of the
amygdala in decision-making. Annals of the New York
Academy of Sciences, 985, 356–369.
Berns, G. S., McClure, S. M., Pagnoni, G., & Montague, P. R.
(2001). Predictability modulates human brain response to
reward. Journal of Neuroscience, 21, 2793–2798.
Blaisdell, A. P., Sawa, K., Leising, K. J., & Waldmann, M. R.
(2006). Causal reasoning in rats. Science, 311, 1020–1022.
Bortz, J. (2005). Statistik für Human- und Sozialwissenschaftler
(6th ed.). Heidelberg: Springer Medizin Verlag.
Breiter, H. C., Aharon, I., Kahneman, D., Dale, A., & Shizgal, P.
(2001). Functional imaging of neural responses to expectancy
and experience of monetary gains and losses. Neuron, 30,
619–639.
Cabeza, R., & Nyberg, L. (2000). Imaging cognition II: An
empirical review of 275 PET and fMRI studies. Journal of
Cognitive Neuroscience, 12, 1–47.
Cobos, P. L., López, F. J., Caño, A., Almaraz, J., & Shanks, D. R.
(2002). Mechanisms of predictive and diagnostic causal
induction. Journal of Experimental Psychology: Animal
Behavior Processes, 28, 331–346.
Corbit, L. H., Muir, J. L., & Balleine, B. W. (2003). Lesions of
mediodorsal thalamus and anterior thalamic nuclei produce
dissociable effects on instrumental conditioning in rats.
European Journal of Neuroscience, 18, 1286–1294.
Corlett, P. R., Aitken, M. R. F., Dickinson, A., Shanks, D. R.,
Honey, G. D., Honey, R. A. E., et al. (2004). Prediction error
during retrospective revaluation of causal associations in
humans: fMRI evidence in favor of an associative model of
learning. Neuron, 44, 877–888.
Corlett, P. R., Murray, G. K., Honey, G. D., Aitken, M. R. F.,
Shanks, D. R., Robbins, T. W., et al. (2007). Disrupted
prediction-error signal in psychosis: Evidence for an
associative account of delusions. Brain, 130, 2387–2400.
Cromwell, H. C., Hassani, O. K., & Schultz, W. (2005). Relative
reward processing in primate striatum. Experimental Brain
Research, 162, 520–525.
de Borchgrave, R., Rawlins, J., Dickinson, A., & Balleine, B.
(2002). Effects of cytotoxic nucleus accumbens lesions on
instrumental conditioning in rats. Experimental Brain
Research, 144, 50–68.
Fenker, D. B., Waldmann, M. R., & Holyoak, K. J. (2005).
Accessing causal relations in semantic memory. Memory &
Cognition, 33, 1036–1046.
Fletcher, P. C., Anderson, J. M., Shanks, D. R., Honey, R., Carpenter,
T. A., Donovan, T., et al. (2001). Responses of the human frontal
cortex to surprising events are predicted by formal associative
learning theory. Nature Neuroscience, 4, 1043–1048.
Hampton, A. N., Adolphs, R., Tyszka, M. J., & OʼDoherty, J. P.
(2007). Contributions of the amygdala to reward expectancy
and choice signals in human prefrontal cortex. Neuron, 55,
545–555.
Izquierdo, A., Suda, R. K., & Murray, E. A. (2004). Bilateral
orbital prefrontal cortex lesions in rhesus monkeys disrupt
choices guided by both reward value and reward
contingency. Journal of Neuroscience, 24, 7540–7548.
Knutson, B., Taylor, J., Kaufman, M., Peterson, R., & Glover, G.
(2005). Distributed neural representation of expected value.
Journal of Neuroscience, 25, 4806–4812.
Knutson, B., & Wimmer, G. E. (2007). Splitting the difference:
How does the brain code reward episodes? Annals of the
New York Academy of Sciences, 1104, 54–69.
Krawczyk, D. C. (2002). Contributions of the prefrontal cortex
to the neural basis of human decision making. Neuroscience
& Biobehavioral Reviews, 26, 631–664.
Matsumoto, K., Suzuki, W., & Tanaka, K. (2003). Neuronal
correlates of goal-based motor selection in the prefrontal
cortex. Science, 301, 229.
Matsumoto, K., & Tanaka, K. (2004). The role of the medial
prefrontal cortex in achieving goals. Current Opinion
Neurobiology, 14, 178–185.
McDermott, K. B., Petersen, S. E., Watson, J. M., & Ojemann,
J. G. (2003). A procedure for identifying regions preferentially
activated by attention to semantic and phonological relations
using functional magnetic resonance imaging.
Neuropsychologia, 41, 293–303.
McNamara, P., Durso, R., Brown, A., & Lynch, A. (2003).
Counterfactual cognitive deficit in persons with Parkinsonʼs
epidemic. Journal of Neurology, Neurosurgery and
Psychiatry, 74, 1065–1070.
Moore, S. C., & Sellen, J. L. (2006). Jumping to conclusions:
A network model predicts schizophrenic patientsʼ
performance on a probabilistic reasoning task. Cognitive,
Affective, & Behavioral Neuroscience, 6, 261–269.
Murray, G. K., Corlett, P. R., Clark, L., Pessiglione, M., Blackwell,
A. D., Honey, G., et al. (2008). Substantia nigra//ventral
tegmental reward prediction error disruption in psychosis.
Molecular Psychiatry, 13, 267–276.
Nelson, D. L., McEvoy, C. L., & Schreiber, T. A. (1998). The
University of South Florida word association, rhyme, and
word fragment norms. Retrieved from http://w3.usf.edu/
FreeAssociation/.
Noppeney, U., Phillips, J., & Price, C. (2004). The neural areas
that control the retrieval and selection of semantics.
Neuropsychologia, 42, 1269–1280.
Nyberg, L., Marklund, P., Persson, J., Cabeza, R., Forkstam, C.,
Petersson, K. M., et al. (2003). Common prefrontal activations
during working memory, episodic memory, and semantic
memory. Neuropsychologia, 41, 371–377.
OʼDoherty, J. P., Dayan, P., Friston, K., Critchley, H., & Dolan,
R. J. (2003). Temporal difference models and reward-related
learning in the human brain. Neuron, 38, 329–337.
OʼDoherty, J. P., Hampton, A., & Hackjin, K. (2007). Model-based
fMRI and its application to reward learning and decision making.
Annals of the New York Academy of Sciences, 1104, 35–53.
Ostlund, S. B., & Balleine, B. W. (2005). Lesions of medial
prefrontal cortex disrupt the acquisition but not the
expression of goal-directed learning. Journal of
Neuroscience, 25, 7763–7770.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you
know what you know? The representation of semantic
knowledge in the human brain. Nature Reviews
Neuroscience, 8, 976–987.
Pavlov, I. P. (1927). Conditioned reflexes: An investigation of
the physiological activity of the cerebral cortex. Translated
and Edited by G. V. Anrep. London: Oxford University Press.
Pearce, J. M., & Hall, G. (1980). A model for Pavlovian
conditioning: Variations in the effectiveness of conditioned but
not of unconditioned stimuli. Psychological Review, 87, 532.
Postle, B. R. (2005). Delay-period activity in the prefrontal
cortex: One function is sensory gating. Journal of Cognitive
Neuroscience, 17, 1679–1690.
Postle, B. R., Berger, J. S., Taich, A. M., & DʼEsposito, M. (2000).
Activity in human frontal cortex associated with spatial
2162
Journal of Cognitive Neuroscience
Volume 22, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
e
1
0
-
p
2
d
1
f
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
o
5
c
1
n
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
8
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
/
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
working memory and saccadic behaviour. Journal of
Cognitive Neuroscience, 12(Suppl. 2), 2–14.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian
conditioning: Variations in the effectiveness of
reinforcement and non-reinforcement. New York:
Appleton-Century-Crofts.
Rodriguez, P. F., Aron, A. R., & Poldrack, R. A. (2006). Ventral-striatal/
nucleus-accumbens sensitivity to prediction errors during
classification learning. Human Brain Mapping, 27, 306–313.
Rogers, T. T., Hocking, J., Noppeney, U., Mechelli, A.,
Gorno-tempini, M., Patterson, K., et al. (2006). Anterior
temporal cortex and semantic memory: Reconciling findings
from neuropsychology and functional imaging. Cognitive,
Affective, & Behavioral Neuroscience, 6, 201–213.
Satpute, A. B., Fenker, D. B., Waldmann, M. R., Tabibnia, G.,
Holyoak, K. J., & Lieberman, M. D. (2005). An fMRI study of
causal judgments. European Journal of Neuroscience, 22,
1233–1238.
Schon, K., Hasselmo, M. E., Lopresti, M. L., Tricarico, M. D., &
Stern, C. E. (2004). Persistence of parahippocampal
representation in the absence of stimulus input enhances
long-term encoding: A functional magnetic resonance
imaging study of subsequent memory after a delayed match-
to-sample task. Journal of Neuroscience, 24, 11088–11097.
Schott, B. H., Niehaus, L., Wittmann, B. C., Schutze, H.,
Seidenbecher, C. I., Heinze, H.-J., et al. (2007). Ageing and
early-stage Parkinsonʼs epidemic affect separable neural
mechanisms of mesolimbic reward processing. Brain, 130,
2412–2424.
Schultz, W. (2002). Getting formal with dopamine and reward.
Neuron, 36, 241–263.
Schultz, W. (2004). Neural coding of basic reward terms of
animal learning theory, game theory, microeconomics and
behavioural ecology. Current Opinion in Neurobiology, 14,
139–147.
Schultz, W. (2006). Behavioral theories and neuophysiology of
reward. Annual Review of Psychology, 57, 87–115.
Schultz, W. (2007). Behavioral dopamine signals. Trends in
Neurosciences, 30, 203–210.
Schultz, W., Apicella, P., Scarnati, E., & Ljungberg, T. (1992).
Neuronal activity in monkey ventral striatum related to the
expectation of reward. Journal of Neuroscience, 12, 4595.
Schultz, W., Dayan, P., & Montague, R. R. (1997). A neural
substrate of prediction and reward. Science, 275, 1593.
Schultz, W., & Dickinson, A. (2000). Neuronal coding of
prediction errors. Annual Review of Neuroscience, 23, 473.
Schultz, W., Tremblay, L., & Holland, A. J. (1998). Reward
prediction in primate basal ganglia and frontal cortex.
Neuropharmacology, 37, 421–429.
Sellen, J. L., Oaksford, M., & Gray, N. S. (2005). Schizotypy and
conditional reasoning. Schizophrenia Bulletin, 31, 105–116.
Shanks, D. R., & López, F. J. (1996). Causal order does not affect
cue selection in human associative learning. Memory and
Cognition, 25, 125–134.
Tanaka, S. C., Balleine, B. W., & OʼDoherty, J. P. (2008).
Calculating consequences: Brain systems that encode the causal
effects of actions. Journal of Neuroscience, 28, 6750–6755.
Thompson-Schill, S. L., DʼEsposito, M., Aguirre, G. K., & Farah,
M. J. (1997). Role of left inferior prefrontal cortex in retrieval
of semantic knowledge: A reevaluation. Proceedings of the
National Academy of Sciences, U.S.A., 94, 14792–14797.
Turner, D. C., Aitken, M. R., Shanks, D. R., Sahakian, B. J.,
Robbins, T. W., Schwarzbauer, C., et al. (2004). The role of
the lateral frontal cortex in causal associative learning:
Exploring preventative and super-learning. Cerebral Cortex,
14, 872–880.
Veltman, D. J., Rombouts, S. A., & Dolan, R. J. (2003).
Maintenance versus manipulation in verbal working memory
revisited: An fMRI study. Neuroimage, 18, 247–256.
Waldmann, M. (1996). Knowledge-based causal induction:
Vol. 34. Causal learning. San Diego, CA: Academic Press.
Waldmann, M., Holyoak, K. J., & Fratianne, A. (1995). Causal
models and the acquisition of category structure. Journal of
Experimental Psychology: General, 124, 181–206.
Waldmann, M. R. (2000). Competition among causes but not
effects in predictive and diagnostic learning. Journal of
Experimental Psychology: Learning, Memory, & Cognition,
26, 53–76.
Waldmann, M. R. (2001). Predictive versus diagnostic causal
learning: Evidence from an overshadowing paradigm.
Psychonomic Bulletin & Review, 8, 600–608.
Waldmann, M. R., & Holyoak, K. J. (1992). Predictive and
diagnostic learning within causal models: Asymmetries in cue
competition. Journal of Experimental Psychology: General,
121, 222–236.
Wallis, J. D. (2007). Orbitofrontal cortex and its contribution
to decision-making. Annual Review of Neuroscience, 30,
31–56.
Wiggs, C. L., Weisberg, J., & Martin, A. (1999). Neural
correlates of semantic and episodic memory retrieval.
Neuropsychologia, 37, 103–118.
Wittmann, B. C., Schott, B. H., Guderian, S., Frey, J. U., Heinze,
H. J., & Duzel, E. (2005). Reward-related fMRI activation of
dopaminergic midbrain is associated with enhanced
hippocampus-dependent long-term memory formation.
Neuron, 45, 459–467.
Yang, T., & Shadlen, M. N. (2007). Probabilistic reasoning by
neurons. Nature, 447, 1075–1080.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
e
1
0
-
p
2
d
1
f
5
/
1
2
1
2
9
/
3
1
9
0
7
/
1
2
8
1
o
5
c
1
n
/
2
1
0
7
0
7
9
0
4
2
1
1
3
8
8
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
8
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
.
.
/
f
t
o
n
1
8
M
a
y
2
0
2
1
Fenker et al.
2163