Veto and Vacillation: A Neural Precursor
of the Decision to Withhold Action
Erman Misirlisoy and Patrick Haggard
Abstrakt
■ The capacity to inhibit a planned action gives human behavior
its characteristic flexibility. How this mechanism operates and
what factors influence a decision to act or not act remain relatively
unexplored. We used EEG readiness potentials (RPs) to examine
preparatory activity before each action of an ongoing sequence, In
which one action was occasionally omitted. We compared RPs be-
tween sequences in which omissions were instructed by a rule
(z.B., “omit every fourth action”) and sequences in which the
participant themselves freely decided which action to omit. RP
amplitude was reduced for actions that immediately preceded a
voluntary omission but not a rule-based omission. We also used
the regular temporal pattern of the action sequences to explore
brain processes linked to omitting an action by time-locking EEG
averages to the inferred time when an action would have oc-
curred had it not been omitted. When omissions were instructed
by a rule, there was a negative-going trend in the EEG, recalling
the rising ramp of an RP. No such component was found for
voluntary omissions. The results are consistent with a model in
which spontaneously fluctuating activity in motor areas of the
brain could bias “free” decisions to act or not. ■
EINFÜHRUNG
In everyday life, our initial impulses do not always produce
optimal actions. Most people recognize the experience
of deciding against saying what one really feels to avoid
offending a friend. In such cases, the decision to inhibit
the action is intentional, self-generated, and often very
wise. Jedoch, most previous studies of action inhibition
have focused on external “stop” signals (Schmied, Johnstone,
& Barry, 2008; Verbruggen & Logan, 2008; Falkenstein,
Hoormann, & Hohnsbein, 1999). These tasks capture only
a part of self-control and cannot capture the familiar case
of an endogenous decision to cancel an action, referred to
as “intentional inhibition” (Filevich, Kühn, & Haggard,
2012). Recent studies have explored intentional inhibition
by asking people to prepare an action and then decide for
themselves whether to execute or inhibit it. Intentional
inhibition produced distinct fMRI activations in medial
pFC (Kühn, Haggard, & Brass, 2009; Brass & Haggard,
2007) and changes in EEG power around the time of the
Entscheidung (Walsh, Kühn, Brass, Wenke, & Haggard, 2010).
Although intentional inhibition may seem closer to
human self-control than external stop signals, das Gehirn
mechanisms involved remain unclear. Most neurocom-
putational models of voluntary action have focused on
action generation rather than inhibition. Zum Beispiel,
several models of frontal cortex are based on hierarchies
(z.B., Kouneiher, Charron, & Koechlin, 2009; Koechlin,
Ody, & Kouneiher, 2003; Brass & Von Cramon, 2002)
University College London
© 2013 Massachusetts Institute of Technology Published under a Creative
Commons Attribution-NonCommercial 3.0 Unportiert (CC BY-NC 3.0) Lizenz.
with anterior areas generating abstract aspects of a plan
and posterior motor areas executing them or generating
stimulus-driven responses. Jedoch, the mechanisms that
initiate plans at the highest level areas remain unexplained.
One could instead think of the voluntary motor system
as a loop, in which each action depends on a preceding
Aktion, rather than as a linear process with an unexplained
initiation. Frontal cortico- BG loops (Alexander & Crutcher,
1990; Alexander, DeLong, & Strick, 1986) could produce
internally generated action sequences (Boecker et al.,
1998; Tanji & Shima, 1994; Brotchie, Iansek, & Horne,
1991) by chaining each action to the one before. On this
view, there need not be any obvious hierarchical starting
point for voluntary behaviors. Thinking of voluntary action
as an iterating loop with a characteristic activation level
may also explain the strong relation between cognitive
resources and action inhibition. Inhibition of action be-
comes difficult and self-control may break down when a
drive to act is sustainedly present (Baumeister, Vohs, &
Tice, 2007; Mischel & Ebbesen, 1970). Here we examine
the opposite possibility, whether action inhibition emerges
at points where the level of activation in a repeated
behavior is momentarily reduced.
We have considered how voluntary decisions to act
or not to act arise within a continuous action sequence.
Participants were asked to omit one particular action
within a repetitive manual motor sequence, based either
on an endogenous voluntary choice or an external in-
struction. Incorporating intentional inhibition into a regu-
lar action sequence provides a background of prepotent
Aktion. Infolge, it may be necessary to truly inhibit an
Zeitschrift für kognitive Neurowissenschaften 26:2, S. 296–304
doi:10.1162/jocn_a_00479
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
Ö
0
C
4
N
6
_
8
A
/
_
J
0
Ö
0
C
4
N
7
9
_
A
P
_
D
0
0
B
4
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
.
T
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Aktion, as opposed to merely failing to initiate it (Filevich
et al., 2012; Kühn et al., 2009). Außerdem, studying a
regular sequence of actions helps to fix action timing,
allowing us to infer when inhibition of action should occur,
if it is present.
We investigated the relation between preparation of
the next element in a continuous motor sequence and
the voluntary decision to omit an action by examining the
spät, lateralized component of the readiness potential
(RP; Shibasaki & Hallett, 2006). Because this component
immediately precedes voluntary actions (Matsuhashi &
Hallett, 2008; Sirigu et al., 2004; Libet, Gleason, Wright,
& Pearl, 1983), its magnitude offers a valuable neural sig-
nature of the generation versus inhibition of voluntary
Aktion. We described above a nonhierarchical loop model
for voluntary chaining of sequences of motor actions. Solch
models predict that activation states are passed around
successive iterations of the loop. On this view, the genera-
tion or inhibition of a current action may depend on the
level of system activity associated with a previous action.
Zum Beispiel, a falling level of activation over successive
loop iterations may eventually lead to an inhibition or
failure to generate the next action.
METHODEN
Teilnehmer
Twenty-six right-handed participants (15 men, 11 Frauen)
were tested. Eight participants were excluded (three had
excessive blink/ EOG artifacts, one could not produce a
regular action sequence, one could not avoid tapping
their foot in addition to their finger, one made excessive
finger movements between actions, and two made too
few voluntary omissions for ERP analysis to be possible),
leaving 18 participants with usable data. All had normal
or corrected-to-normal vision, and none had a history
of neurological or psychiatric disorders.
Design
In all tasks, participants pressed the space button on a
keyboard with their right index finger once every 2 Sek
in a self-paced manner. In the rule-based omission task,
participants used a rule given by the experimenter to omit
every fourth or sixth keypress in the sequence. In the volun-
tary omission task, participants were instructed to omit a
keypress when they themselves chose. Participants were
asked to be as spontaneous as possible in voluntary omis-
sions and to decide at the very last moment prior to action.
They were asked to avoid preplanning omissions or fol-
lowing a rule. Rule-based and voluntary tasks had 20 Versuche
jede. The order of these two tasks was alternated across
Teilnehmer. Individual trials ended once 30 keypresses
were made, resulting in trial durations of approximately
1 min.
For the first nine participants, the instruction for the
rule-based condition was to omit every fourth keypress.
Jedoch, preliminary inspection of the data showed that
voluntary omissions tended to occur more rarely than
Das, producing a confound between condition and motor
Aktivität. daher, the remaining nine participants were
instructed to omit every sixth keypress. This successfully
balanced overall action and omission frequency across
Bedingungen.
Verfahren
Participants first practiced synchronizing keypresses to a
0.5-Hz auditory metronome, allowing them to learn the re-
quired rhythm. In the subsequent tasks, no external pacing
stimulus was given. Participants were asked to avoid count-
ing seconds in their timing and simply to follow the rhythm
they had learned in the synchronization phase. In all trials,
participants were first presented with an instruction to
“Press space to begin the trial.” Following the keypress,
a white fixation cross on a black background appeared
centrally on the monitor, which participants were asked
to remain fixated on while continuing keypresses at an
internally paced rate of approximately 0.5 Hz.
In the practice task, participants produced a sequence
von 30 keypresses at 0.5 Hz in each trial. Then the experi-
mental tasks began. In the rule-based omission condition,
participants were instructed to omit every fourth or every
sixth keypress (see Design). In the voluntary omission
condition, participants freely chose when to omit key-
presses. Precise timing between keypresses was incenti-
vized in the experimental session. Participants received
feedback about their performance at the end of each trial.
The mean interval between their actions had to be within
1700–2300 msec, and the standard deviation of these in-
tervals had to be below 200. Omitted actions required an
extended average interval of 3500–4500 msec between
consecutive actions and standard deviation below 500.
Fifteen pence per trial was gained for passing all criteria.
Optimal performance could potentially increase participant
income by £6.
EEG
Twenty-seven EEG channels (FT8, FC6, FC4, FC2, FCz,
FC1, FC3, FC5, FT7, T8, C6, C4, C2, Cz, C1, C3, C5, T7,
TP8, CP6, CP4, CP2, CPz, CP1, CP3, CP5, TP7) were re-
corded from sensorimotor areas. The ground electrode
was at scalp position AFz, and the reference electrode
was attached to the right earlobe. EOG electrodes were
attached to the external canthi of each eye and the supra
and suborbital areas of the right eye. A bandpass filter
zwischen 0.1 Und 30 Hz and a notch filter of 50 Hz were
applied, and the sampling rate was 256 Hz. Daten waren
preprocessed and analyzed in EEGLAB v10.2.5.6b running
in Matlab 7.10.
Misirlisoy and Haggard
297
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
Ö
0
C
4
N
6
_
8
A
/
_
J
0
Ö
0
C
4
N
7
9
_
A
P
_
D
0
0
B
4
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Epochs were defined from 1000 msec before to 500 ms
after each keypress and baseline-corrected at −1000 to
−800 msec. The baseline correction served to remove
effects of very slow EEG drifts and isolate the component
of the EEG related specifically to each action. Omission
events were inferred using the temporal interval between
immediately preceding actions. If the omitted action is
labeled n and the preceding actions n − 1 and n − 2,
then whenever an extended interkeypress interval indi-
cated an omission, the temporal interval between n − 1
and n − 2 was repeated to insert an omission event in
the data. Although this was only an estimate of the
omitted event timing, its accuracy was assumed to be
similar for rule-based and voluntary omissions.
Epochs were discarded if the potential from any EOG
electrode fell outside +80 to −80 μV or any other electrode
fell outside +100 to −100 μV. Improbable epochs that con-
tained EEG signal amplitudes exceeding five standard devia-
tions of the mean probability distribution or the mean
kurtosis value were removed (Delorme, Sejnowski, &
Makeig, 2007). Linear detrending was applied over the re-
cording period to identify and remove drift (Matsuhashi
& Hallett, 2008). Participants were discarded if the num-
ber of events for any condition fell below 50 after arti-
fact rejection (see Participants).
To investigate action preparation, the EEG signal for
the last 500 msec before action onset was averaged from
the C3 electrode (the late bereitschaftspotential (or late
RP) over the contralateral motor area; Shibasaki & Hallett,
2006). Given the short intervals of approximately 2 Sek
between each action event, earlier components risked con-
tamination from previous action related activity. daher,
only the late RP found at C3 was analyzed. The same
procedure was used for inferred omission onsets.
ERGEBNISSE
Inspection of EEG traces and statistical comparisons
showed no differences between the groups that omitted
every fourth and every sixth action in the rule-based
condition. daher, the data were pooled across groups.
We defined “omission − 1” actions as those that immedi-
ately precede omissions and “omission + 1” actions as
those that follow omissions. All other actions were classified
as “standard actions.”
Behavioral Data
Behavioral data are shown in Table 1.
There was no difference between the rule-based and
voluntary conditions in the number of rewarded, Das
Ist, accurately timed trials, T(17) = 0.41, p > .05. The tem-
poral intervals between successive actions tended to be
shorter in the rule-based than the voluntary condition,
although this effect did not reach significance t(17) =
−1.93, p = .07. There was no difference in the duration
of omission intervals, T(17) = −1.18, p > .05. This sug-
Tisch 1. Rewards (d.h., Appropriately Timed Actions, See Text)
and Mean Temporal Intervals in Rule-based and Voluntary
Conditions
Rule-based
Voluntary
Rewards (%)
70 (23.3)
68.3 (24.4)
Action intervals (ms)
1977.2 (109.0)
2045.5 (110.7)
Inhibition intervals (ms)
3870.5 (198.5)
3935.9 (206.8)
gests that the conditions did not differ substantially in
difficulty and participants maintained broadly similar
timing across both. In the voluntary condition, Teilnehmer
made 5.54 actions on average between omissions with a
standard deviation of 1.1. In the rule-based condition, von
Kontrast, the number of actions between omissions was
instructed to be 3 oder 5 (for “omit every fourth action”
and “omit every sixth action,” respectively; see Design).
EEG Data
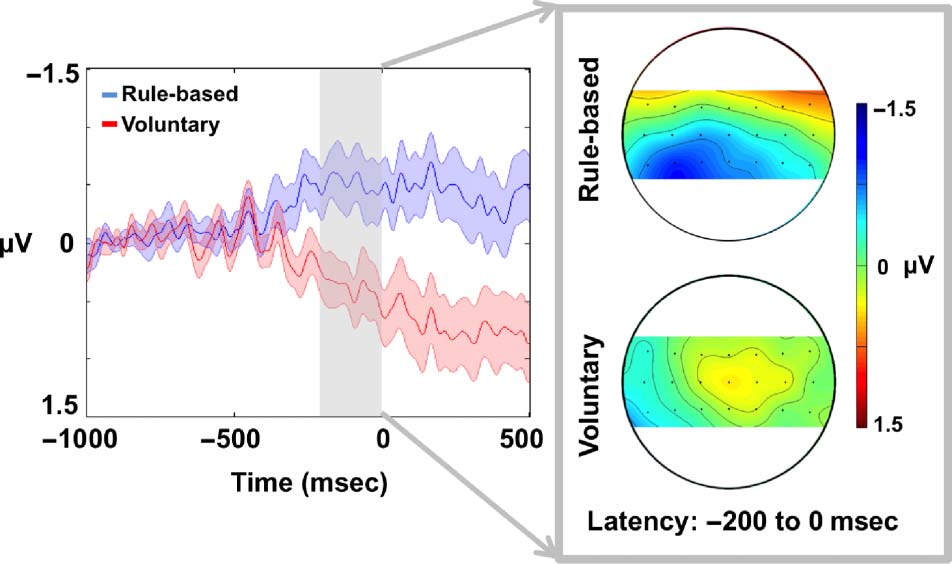
Figur 1 shows ERPs for the different types of action at
electrode C3.
RP amplitudes were calculated prior to actions in the late
RP period (mean of the signal during the period −500
Zu 0 ms). We separately averaged actions immediately
before and after the omission and all other “standard”
Aktionen. A 2 × 3 ANOVA with factors of Omission Type
(rule-based/voluntary) and Action Position (standard
actions/omission − 1 actions/omission + 1 Aktionen) showed
no main effects of either Omission Type, F(1, 17) = 1.34,
p > .05, or Action Position, F(2, 34) = 1.15, p > .05.
Jedoch, a significant interaction was found between
these factors, F(2, 34) = 6.68, P < .01.
To explore this interaction, simple effects tests were
used to compare voluntary and rule-based RPs for each
action. There were no differences in RP amplitudes be-
tween rule-based and voluntary conditions for standard
actions across the conditions, t(17) = 0.45, p > .05, oder
omission + 1 Aktionen, T(17) = 1.61, p > .05, but a significant
difference was found for omission − 1 Aktionen, T(17) =
−4.01, p = .001 (scalp map plots directly showing the
difference between conditions can also be seen in the
supplementary data). The crucial difference between
conditions therefore lies in preparatory activity for actions
that immediately precede an omission. Simple effects
were also tested by comparing each RP within the rule-
based and voluntary conditions. RP amplitude for omission
− 1 actions was significantly reduced compared with stan-
dard actions in the voluntary condition, T(17) = 2.56,
P < .05, but no difference was found in the rule-based
condition, t(17) = −1.36, p > .05. Omission − 1 RPs were
reduced relative to omission + 1 RPs in the voluntary
condition, T(17) = 2.89, p = .01, but not in the rule-
based condition, T(17) = −0.32, p > .05. Omission + 1
potentials did not differ from standard actions in either
298
Zeitschrift für kognitive Neurowissenschaften
Volumen 26, Nummer 2
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
Ö
0
C
4
N
6
_
8
A
/
_
J
0
Ö
0
C
4
N
7
9
_
A
P
_
D
0
0
B
4
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
T
F
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
Ö
0
C
4
N
6
_
8
A
/
_
J
0
Ö
0
C
4
N
7
9
_
A
P
_
D
0
0
B
4
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
.
/
T
F
Figur 1. ERP data (n = 18) for standard, omission − 1, and omission + 1 actions at electrode C3—the position of the omitted action in the
sequence is indicated by the dots. Shaded colors around ERPs show standard error. Data are time-locked to action onset (keypress). Note difference
between conditions in RPs for actions immediately prior to omission.
the voluntary condition, T(17) = 1.58, p > .05, or the rule-
based condition, T(17) = 0.37, p > .05.
slopes significantly differed between the two conditions:
T(17) = −5.02, P < .001.
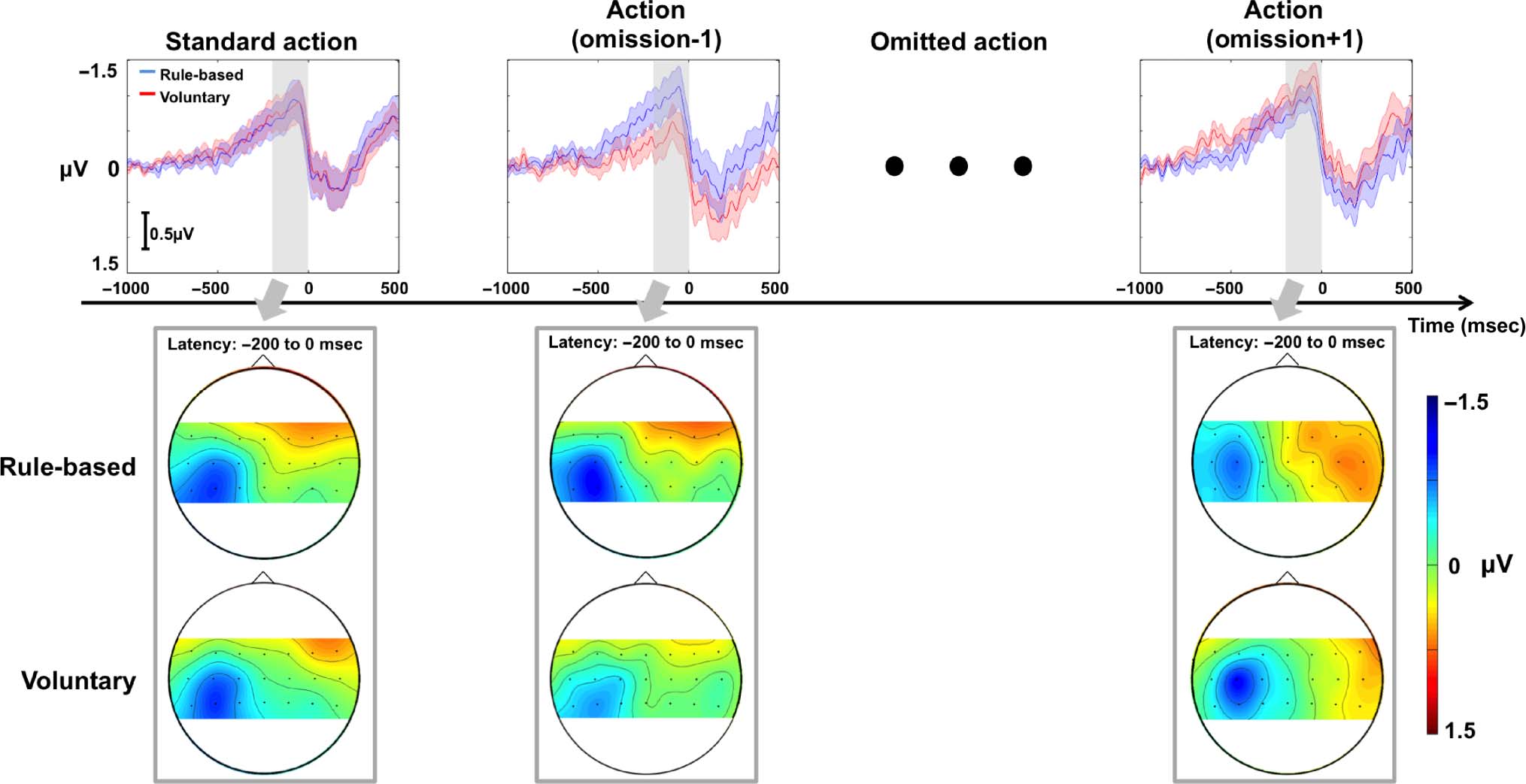
We also explored the EEG activity related to action
omission by time-locking to the expected time of action.
We had no strong prior hypothesis about the scalp loca-
tion or form of omission potentials. However, we note
that response inhibition has been frequently localized
to medial frontal areas (Simmonds, Pekar, & Mostofsky,
2008; Picton et al., 2007; Mostofsky et al., 2003) and that
departures and omissions from regular sequences of
events are typically measured by vertex potentials (e.g.,
Nordby, Hammerborg, Roth, & Hugdahl, 1994). We there-
fore compared the omission potential at Cz using the same
time window as for RPs preceding action. The data are
shown in Figure 2. In the period before rule-based omis-
sions, the trace showed a negative-going deflection. This
deflection was maximal over the contralateral sensorimotor
cortex and had a similar form to an RP, though a somewhat
smaller amplitude. Furthermore, the abrupt shift to positiv-
ity just before movement onset that marks the end of the
classical RP was not present for rule-based omission trials.
Because the classical RP is characterized by a negative-going
ramp-like form, we used linear fits to the averaged EEG in
the −500 to 0 msec time window corresponding to the late
RP to investigate whether a component similar to RP might
be present on omission trials. We found a trend toward
a negative slope for rule-based omissions, t(17) = −1.89,
p = .08. In contrast, we found a positive-going slope
before voluntary omissions. This slope was significantly
greater than zero: t(17) = 3.4, p < .01. Furthermore, the
Finally, we compared mean amplitudes during the time
window of the late RP for omissions in the same way as we
previously did for actions. This showed a significantly
greater amplitude for voluntary than for rule-based omis-
sion potentials, t(17) = 2.99, p < .01.
DISCUSSION
We measured event-related EEG associated with actions
and with decisions to omit actions in a regular sequence.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. ERP data (n = 18) for omissions at electrode Cz (left)
and across the scalp (right). Shaded colors around ERPs show
standard error. Zero milliseconds indicates the inferred time of
action omission.
Misirlisoy and Haggard
299
Our results indicated that the “voluntary” decision to
omit a particular action was prefigured by a reduced RP
for the immediately preceding action in the sequence.
The motor system seems primed in advance to omit an
action voluntarily. This was not the case in a rule-based
condition, where the particular action to be omitted was
specified beforehand by a rule. Therefore, the decrease
in action-related processing prior to voluntary action is
unlikely to reflect simple advance planning for the forth-
coming omission, because such advance planning should
be even more evident when using a rule than when
choosing “freely.” Any difference in task difficulty across
the rule-based and voluntary tasks also seems unable to
account for the findings. Although a difference in diffi-
culty could explain a main effect of task, it cannot readily
explain the interaction between task and action type (i.e.,
standard/preomission/postomission actions) found in
our RPs. One might suggest that the effect of task difficulty
could be temporally focused on the period just prior to
omission, but this seems implausible, because scalp topog-
raphies show the main modulation during these periods to
be over motor areas, rather than the distributed network
associated with task difficulty (e.g., Sunaert, Van Hecke,
Marchal, & Orban, 2000). Instead, these results are consis-
tent with a loop model for action generation, in which
the level of activation in an iterating motor loop influ-
ences decisions to act or not. In particular, background
fluctuations in the activation level of such a loop could
affect a high-level choice process. A purely hierarchical,
feedforward model,
in which decisions originate at
high levels and cascade unidirectionally to lower levels
for execution (e.g., Kouneiher et al., 2009), cannot easily
explain the association between the activation associ-
ated with execution for one action and the decision to
omit the next. Our result does not rule out hierarchical
models per se, but it does imply that the higher levels in
the hierarchy are influenced by feedback from lower-level
execution processes.
Because our sequential action task had regular timing,
we were also able to calculate ERPs associated with actions
that are inhibited by time-locking to the moment when the
action would have been expected to occur. The latencies
of ERP components for these averages should be inter-
preted with caution, because the time-locking point is only
inferred, rather than measured. Any inaccuracy in this
inference will produce a temporal smearing of ERP com-
ponents and a reduction in ERP amplitude. Therefore,
comparing amplitudes between omission-related poten-
tials and action-related potentials may be problematic.
However, we can compare omission-related potentials
between rule-based and voluntary conditions. Both condi-
tions should be equally affected by smearing, and indeed
temporal intervals for actions and omissions did not differ
between the conditions (see Results). We found a trend
toward a negative-going, ramp-like component, similar
in some ways to the form of an RP, prior to instructed,
rule-based omissions. Voluntary omissions, however, were
preceded by a positive-going potential. We cannot exclude
the possibility that this prolonged positive shift may include
other components. For example, studies of stimulus-locked
no-go potentials found a no-go-P3 component (Bokura,
Yamaguchi, & Kobayashi, 2001; Falkenstein et al., 1999),
directly related to response inhibition. A temporally
smeared version of these components could contribute
to our omission-related potentials, but the difference be-
tween conditions nevertheless remains. Alternatively, the
negative-going omission-related potential in the rule-based
condition could conceivably represent a “simulated” but
not executed action. For example, participants might
represent the rule-based action sequence as a rhythmic
pattern. They might then simulate the “missed beat” of
this rhythm to follow the omission rule.1 In any case, this
component was not present prior to voluntary omissions,
suggesting an important difference between voluntary
and rule-based omissions.
Previous studies of intentional inhibition have been
hampered by the difficulty of assessing whether a failure
to act was because of a predecision to not initiate any
action processing or because of a specific last moment
process of inhibition (Filevich et al., 2012). Because there
is no clear behavioral marker to indicate the time of in-
hibition, it is difficult to distinguish whether inhibition
occurs early or late relative to action preparation. This
distinction is important, because an early inhibition effec-
tively reduces the decision to inhibit to action selection
(Mostofsky & Simmonds, 2008), whereas late inhibition
implies a specialized cognitive process reminiscent of self-
control. The design of our task controls this aspect. In
particular, rule-based omission implies predecision about
inhibition. In contrast, our voluntary condition invited
participants to decide spontaneously to omit an action, as
a result of their own real-time choice. If participants had
in fact predecided which action to omit in the voluntary
condition, then no difference would be expected be-
tween voluntary and rule-based conditions. The results
showed that voluntary omission was associated with a re-
duced RP for preceding actions, whereas rule-based
omission was not. Reduction of preceding RPs may reflect
a mechanism underlying voluntary decisions to inhibit
which is independent of conscious prior decision and
which could potentially explain the spontaneous nature
of some action choices (Libet et al., 1983). In essence, free
decisions may capitalize on the momentary state of activa-
tion in motor circuits.
Competition between Action and Omission
Recent models of action selection have proposed that plans
for multiple response alternatives compete simultaneously
in the motor system (Klaes, Westendorff, Chakrabarti, &
Gail, 2011; Cisek & Kalaska, 2010; Cisek, 2007). Accumulat-
ing information and biasing influences from the BG and
pFC contribute to resolving the competition and to select-
ing a particular response.
300
Journal of Cognitive Neuroscience
Volume 26, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
o
0
c
4
n
6
_
8
a
/
_
j
0
o
0
c
4
n
7
9
_
a
p
_
d
0
0
b
4
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Ongoing fluctuations in the level of activity of the cortico-
BG-thalamocortical circuit (Alexander & Crutcher, 1990;
Alexander et al., 1986) might provide the basis for binary
decisions about action and inhibition behavior. Low activity
would make omission more likely, whereas higher activ-
ity would favor action. Progressively reducing motor ac-
tivity would lead to the “voluntary” decision to omit, as
shown by our RPs for actions immediately preceding
omission.
The lack of preparatory motor activity or excitability
during voluntary omissions is consistent with the idea
that low spontaneous motor activity biases the choice
to omit an action. At electrode Cz, voluntary omissions
showed an increasing positivity replacing the ramp-like
negativity of the normal RP. EEG provides only a weak
indication about the location of underlying generators,
which may not correspond to the maximal location of the
component on the scalp. However, the central distribution
of this positive-going potential could reflect activity in pre-
SMA or other medial frontal areas involved in inhibition
(Simmonds et al., 2008; Brass & Haggard, 2007; Picton
et al., 2007). In any case, our results suggest a specific
inhibitory process associated with voluntary inhibition
and marked by a positive-going potential at the scalp.
Although both voluntary and rule-based action selections
employ a prefrontal control component in the planning
of the temporal structures of events and the control of goal-
directed action behaviors (Tanji & Hoshi, 2008), a clear
external rule appears to override the biasing effects of
spontaneous activity in the motor systems. Thus, we
found no reduction in RP for actions prior to omission
in the rule-based condition. In fact, we found a trend to-
ward a negative-going shift even before rule-based omis-
sions themselves. We speculate that this may correspond to
an internal preparation or simulation process, which occurs
even when action execution is omitted (Osman, Albert,
Ridderinkhof, Band, & van der Molen, 2006) —indeed sev-
eral fMRI studies confirm that primary motor cortex is often
activated during simulated or imagined action (Lacourse,
Orr, Cramer, & Cohen, 2005; Gerardin et al., 2000; Lotze
et al., 1999). Interestingly, this component was not present
for voluntary decisions to omit. This finding is consistent
with our hypothesis that voluntary omissions are asso-
ciated with a reducing level of motor activation.
Another possible account for the present findings is
based on changing levels of uncertainty about action in
the voluntary condition. Greater probability (certainty)
of an impending action has been associated with larger
preparatory motor potentials (Scheibe, Schubert, Sommer,
& Heekeren, 2009; Low & Miller, 1999). In the voluntary
task of this study, there is ongoing uncertainty about
whether one should act or omit action in each moment.
Given that actions should be omitted at some point in
the sequence, voluntary decisions to omit action may
involve a hazard function. Each successive action in the
sequence up until the omission involves greater uncer-
tainty than the one before, because it is increasingly likely
that the omission will shortly occur. This growing uncer-
tainty might progressively reduce RP amplitudes and
could therefore account for the reduced RP we found for
omission − 1 actions. In contrast, there is never uncertainty
about action in the rule-based task. An account based on
uncertainty therefore makes two clear predictions. First,
the voluntary task should have globally smaller RPs com-
pared with the rule-based task. The near-identical RPs
found for standard actions across the voluntary and rule-
based conditions goes against this prediction. Second, in
the voluntary task only, RPs should progressively decrease
in amplitude as the omission approaches, following a
hazard function. In principle, this second prediction could
be tested by comparing RPs for omission − 1, omission −
2, …, omission − n actions. However, our design gave too
few trials at omission − 2 and earlier actions to calculate
reliable ERPs for testing this prediction. A study with more
trials and longer sequences of actions prior to omission
would be required to test the uncertainty hypothesis
in detail.
The Influential Role of Spontaneous Motor Activity
in Voluntary Decisions
There is strong evidence indicating that ongoing sponta-
neous activity in neural systems influences motor behaviors
(Mazaheri, Nieuwenhuis, van Dijk, & Jensen, 2009; Fox,
Snyder, Vincent, & Raichle, 2007; Churchland, Afshar, &
Shenoy, 2006; Connolly, Goodale, Goltz, & Munoz, 2005).
In fact, the RP itself may reflect spontaneously fluctuating
neural activity. If the fluctuation is sufficient to cross a
threshold, a movement may be triggered, whereas sub-
threshold fluctuations spontaneously decay without caus-
ing a movement (Schurger, Sitt, & Dehaene, 2012).
According to this model, the ramp-like shape of the RP
may simply reflect averaging those fluctuations, which
eventually succeed in crossing the threshold level for
action. Several researchers emphasize the functional
relevance of spontaneous brain activity and the risk of
dismissing such activity as irrelevant noise (Sadaghiani,
Hesselmann, Friston, & Kleinschmidt, 2010; Fox &
Raichle, 2007). Such fluctuating activity is typically shown
to affect behavior when it is examined immediately pre-
stimulus (Sadaghiani et al., 2010). Our results extend this
view to the case of action inhibition.
The relation between the EEG signal and these hypothe-
sized fluctuating activation levels is an important issue.
The EEG time series contains low-frequency components,
including “slow drifts,” whose physiological significance
is unclear. As a result, there will be trivial dependence
between EEG amplitudes at successive time points. Base-
line correction removes these low-frequency components.
Our analysis is based on ERPs during brief, nonoverlapping
epochs, baseline-corrected at the start of each epoch. This
approach reduces the probability of detecting a spurious
association between the activity levels for successive
actions/omissions because of both events riding upon
Misirlisoy and Haggard
301
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
o
0
c
4
n
6
_
8
a
/
_
j
0
o
0
c
4
n
7
9
_
a
p
_
d
0
0
b
4
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the same slow drift. Thus, our findings imply fluctuation in
signals related to processing successive action events
rather than long-lasting shifts in global brain state. For
example, Figure 1 shows a greater postmovement positivity
for the action just prior to omission in the voluntary con-
dition relative to the rule-based condition. However, this
cannot be assumed to continue into the positive-going
shift seen during subsequent voluntary omissions, be-
cause a baseline correction intervenes between the two
components. That is, our results demonstrate a relation
between event-related processing for successive events
rather than a trivial continuity of the EEG time series.
The focus on event-related processing rather than EEG
level across interevent intervals is methodologically ap-
propriate for removing artifactual correlations because
of global slow drifts. It is also scientifically appropriate,
given that our task was constructed around action and
omission events, with no instruction regarding the rela-
tively long 2-sec interval in between.
Inhibition, Cognitive Control, and Consciousness
Many theories of inhibition accord it special status, as a
specific cognitive control function with a privileged link
to consciousness (Hughes, Velmans, & De Fockert,
2009; Dehaene et al., 2003). Our data suggest a rather dif-
ferent view. Instead of intervention by a high-level pro-
cess, intentional inhibition may also reflect fluctuating
activity in low-level motor circuits. In the real world, when
the external motivations for acting or withholding an ac-
tion are clear-cut and an easy decision between competing
options can be made—as is the case in rule-based action
selection—spontaneous fluctuations in the motor system
may have little influence on voluntary decisions. However,
in situations where there are competing motivations to
act or not to act, the action selection system must re-
solve the conflict by choosing between equally attractive
alternatives. Our results suggest that such “decisions”
could simply capitalize on the preceding state of the motor
system. Many scientists have suggested that inhibition is a
necessarily conscious top–down override (Dehaene &
Naccache, 2001; Jack & Shallice, 2001; Merikle, Joordens,
& Stolz, 1995) or “free wonʼt” veto mechanism (Libet et al.,
1983). Indeed, legal systems assume that the capacity to
refrain from inappropriate action is the basis of human
moral responsibility. Our results suggest a less homuncular
view, namely that “decisions” to inhibit may be conse-
quences of the ongoing state of the cortical and subcortical
motor systems. Some theories of volition have involved
the very strong contention that “free will” simply reduces
to random fluctuations in neural processes (Carpenter,
1999; Eccles, 1985). However, these theories could not
convincingly identify the locus of randomness. Our findings
suggest that intentional inhibition, like other cognitive
processes, indeed interacts with background fluctua-
tions of neural activity. However, we add two important
caveats: Our study cannot provide direct evidence that
these fluctuations are random, and our study does not
preclude other processes contributing to intentional in-
hibition. However, the design of our task does offer
some hints about the mechanism where this fluctuating
influence acts. Specifically, intentional inhibition may be
determined by the interaction between cognitive, pre-
frontal decision processes and levels of activation in
an iterative motor execution loop that links each action
to the next (Marsden, 1984).
In conclusion, endogenous decisions to spontaneously
inhibit an action are influenced by lower-level motor ac-
tivity. Neural activity related to actions preceding and lead-
ing up to intentional inhibition can be predictive of its
upcoming occurrence. In situations where we are making
a difficult decision about whether to act or not, sponta-
neous levels of activity in our motor system may feed into
our decision, biasing us to go one way or the other.
Acknowledgments
We would like to thank Dr. Mathew Salvaris for his technical
help and advice throughout the study and the ESRC for funding
this project as part of an ESF ECRP “Intentional Inhibition of
Human Action” (grant RES-062-23-2183). P. H. was additionally
supported by a Research Fellowship from the Leverhulme
Trust, an ESRC Professorial Fellowship, and ERC Advanced
Grant HUMVOL. We also thank the reviewers of this paper
for their useful comments and suggestions.
Reprint requests should be sent to Erman Misirlisoy, Institute
of Cognitive Neuroscience, University College London, 17
Queen Square, London WC1N 3AR, United Kingdom, or via
e-mail: e.misirlisoy.11@ucl.ac.uk.
Note
1. We are grateful to an anonymous reviewer for suggesting
this interpretation.
REFERENCES
Alexander, G. E., & Crutcher, M. D. (1990). Functional
architecture of basal ganglia circuits: Neural substrates of
parallel processing. Trends in Neurosciences, 13, 266–271.
Alexander, G. E., DeLong, M. R., & Strick, P. L. (1986). Parallel
organization of functionally segregated circuits linking basal
ganglia and cortex. Annual Review of Neuroscience, 9,
357–381.
Baumeister, R. F., Vohs, K. D., & Tice, D. M. (2007). The
strength model of self-control. Current Directions in
Psychological Science, 16, 351–355.
Boecker, H., Dagher, A., Ceballos-Baumann, A. O., Passingham,
R. E., Samuel, M., Friston, K. J., et al. (1998). Role of the
human rostral supplementary motor area and the basal
ganglia in motor sequence control: Investigations with H2
15O PET. Journal of Neurophysiology, 79, 1070–1080.
Bokura, H., Yamaguchi, S., & Kobayashi, S. (2001).
Electrophysiological correlates for response inhibition in a
Go/NoGo task. Clinical Neurophysiology, 112, 2224–2232.
Brass, M., & Haggard, P. (2007). To do or not to do: The neural
signature of self-control. The Journal of Neuroscience, 27,
9141–9145.
302
Journal of Cognitive Neuroscience
Volume 26, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
o
0
c
4
n
6
_
8
a
/
_
j
0
o
0
c
4
n
7
9
_
a
p
_
d
0
0
b
4
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Brass, M., & Von Cramon, D. Y. (2002). The role of the frontal
cortex in task preparation. Cerebral Cortex, 12, 908–914.
Brotchie, P., Iansek, R., & Horne, M. K. (1991). Motor function
of the monkey globus pallidus. 2. Cognitive aspects of
movement and phasic neuronal activity. Brain, 114, 1685–1702.
Carpenter, R. H. S. (1999). A neural mechanism that randomises
behaviour. Journal of Consciousness Studies, 6, 13–22.
Churchland, M. M., Afshar, A., & Shenoy, K. V. (2006). A central
source of movement variability. Neuron, 52, 1085–1096.
Cisek, P. (2007). Cortical mechanisms of action selection:
The affordance competition hypothesis. Philosophical
Transactions of the Royal Society, Series B, Biological
Sciences, 362, 1585–1599.
Cisek, P., & Kalaska, J. F. (2010). Neural mechanisms for
interacting with a world full of action choices. Annual Review
of Neuroscience, 33, 269–298.
Connolly, J. D., Goodale, M. A., Goltz, H. C., & Munoz, D. P.
(2005). fMRI activation in the human frontal eye field is
correlated with saccadic reaction time. Journal of
Neurophysiology, 94, 605–611.
Dehaene, S., Artiges, E., Naccache, L., Martelli, C., Viard, A.,
Schürhoff, F., et al. (2003). Conscious and subliminal conflicts
in normal subjects and patients with schizophrenia: The
role of the anterior cingulate. Proceedings of the National
Academy of Sciences, 100, 13722–13727.
Dehaene, S., & Naccache, L. (2001). Towards a cognitive
neuroscience of consciousness: Basic evidence and a
workspace framework. Cognition, 79, 1–37.
Delorme, A., Sejnowski, T., & Makeig, S. (2007). Enhanced
detection of artifacts in EEG data using higher-order statistics
and independent component analysis. Neuroimage, 34,
1443–1449.
Eccles, J. C. (1985). Mental summation: The timing of voluntary
intentions by cortical activity. Behavioral and Brain
Sciences, 8, 542–543.
Falkenstein, M., Hoormann, J., & Hohnsbein, J. (1999). ERP
components in go/no-go tasks and their relation to
inhibition. Acta Psychologica, 101, 267–291.
Filevich, E., Kühn, S., & Haggard, P. (2012). Intentional
inhibition in human action: The power of “no”. Neuroscience
& Biobehavioral Reviews, 36, 1107–1118.
Fox, M. D., & Raichle, M. E. (2007). Spontaneous fluctuations in
brain activity observed with functional magnetic resonance
imaging. Nature Reviews Neuroscience, 8, 700–711.
Fox, M. D., Snyder, A. Z., Vincent, J. L., & Raichle, M. E. (2007).
Intrinsic fluctuations within cortical systems account for
intertrial variability in human behavior. Neuron, 56, 171–184.
Gerardin, E., Sirigu, A., Lehéricy, S., Poline, J.-B., Gaymard, B.,
Marsault, C., et al. (2000). Partially overlapping neural networks
for real and imagined hand movements. Cerebral Cortex, 10,
1093–1104.
Hughes, G., Velmans, M., & De Fockert, J. (2009). Unconscious
priming of a no-go response. Psychophysiology, 46, 1258–1269.
Jack, A. I., & Shallice, T. (2001). Introspective physicalism as an
approach to the science of consciousness. Cognition, 79,
161–196.
Klaes, C., Westendorff, S., Chakrabarti, S., & Gail, A. (2011).
Choosing goals, not rules: Deciding among rule-based action
plans. Neuron, 70, 536–548.
Koechlin, E., Ody, C., & Kouneiher, F. (2003). The architecture
of cognitive control in the human prefrontal cortex. Science,
302, 1181–1185.
Kouneiher, F., Charron, S., & Koechlin, E. (2009). Motivation
and cognitive control in the human prefrontal cortex. Nature
Neuroscience, 12, 939–945.
Kühn, S., Haggard, P., & Brass, M. (2009). Intentional inhibition:
How the “veto-area” exerts control. Human Brain Mapping,
30, 2834–2843.
Lacourse, M. G., Orr, E. L. R., Cramer, S. C., & Cohen, M. J.
(2005). Brain activation during execution and motor imagery
of novel and skilled sequential hand movements.
Neuroimage, 27, 505–519.
Libet, B., Gleason, C. A., Wright, E. W., & Pearl, D. K. (1983).
Time of conscious intention to act in relation to onset of
cerebral activity (readiness-potential). Brain, 106, 623–642.
Lotze, M., Montoya, P., Erb, M., Hülsmann, E., Flor, H., Klose,
U., et al. (1999). Activation of cortical and cerebellar motor
areas during executed and imagined hand movements: An
fMRI study. Journal of Cognitive Neuroscience, 11, 491–501.
Low, K. A., & Miller, J. (1999). The usefulness of partial
information: Effects of go probability in the choice/nogo
task. Psychophysiology, 36, 288–297.
Marsden, C. D. (1984). Which motor disorder in Parkinsonʼs
disease indicates the true motor function of the basal ganglia?
Ciba Foundation Symposium, 107, 225–241.
Matsuhashi, M., & Hallett, M. (2008). The timing of the
conscious intention to move. European Journal of
Neuroscience, 28, 2344–2351.
Mazaheri, A., Nieuwenhuis, I. L. C., van Dijk, H., & Jensen, O.
(2009). Prestimulus alpha and mu activity predicts failure
to inhibit motor responses. Human Brain Mapping, 30,
1791–1800.
Merikle, P. M., Joordens, S., & Stolz, J. A. (1995). Measuring the
relative magnitude of unconscious influences. Consciousness
and Cognition, 4, 422–439.
Mischel, W., & Ebbesen, E. B. (1970). Attention in delay of
gratification. Journal of Personality and Social Psychology,
16, 329–337.
Mostofsky, S. H., Schafer, J. G. B., Abrams, M. T., Goldberg, M. C.,
Flower, A. A., Boyce, A., et al. (2003). fMRI evidence that the
neural basis of response inhibition is task-dependent. Brain
Research. Cognitive Brain Research, 17, 419–430.
Mostofsky, S. H., & Simmonds, D. J. (2008). Response
inhibition and response selection: Two sides of the same
coin. Journal of Cognitive Neuroscience, 20, 751–761.
Nordby, H., Hammerborg, D., Roth, W. T., & Hugdahl, K.
(1994). ERPs for infrequent omissions and inclusions of
stimulus elements. Psychophysiology, 31, 544–552.
Osman, A., Albert, R., Ridderinkhof, K. R., Band, G., &
van der Molen, M. (2006). The beat goes on: Rhythmic
modulation of cortical potentials by imagined tapping.
Journal of Experimental Psychology: Human Perception
and Performance, 32, 986–1005.
Picton, T. W., Stuss, D. T., Alexander, M. P., Shallice, T.,
Binns, M. A., & Gillingham, S. (2007). Effects of focal
frontal lesions on response inhibition. Cerebral Cortex,
17, 826–838.
Sadaghiani, S., Hesselmann, G., Friston, K. J., & Kleinschmidt, A.
(2010). The relation of ongoing brain activity, evoked
neural responses, and cognition. Frontiers in Systems
Neuroscience, 4, 20.
Scheibe, C., Schubert, R., Sommer, W., & Heekeren, H. R.
(2009). Electrophysiological evidence for the effect of prior
probability on response preparation. Psychophysiology, 46,
758–770.
Schurger, A., Sitt, J. D., & Dehaene, S. (2012). An accumulator
model for spontaneous neural activity prior to self-initiated
movement. Proceedings of the National Academy of
Sciences, 109, E2904–E2913.
Shibasaki, H., & Hallett, M. (2006). What is the
Bereitschaftspotential? Clinical Neurophysiology, 117,
2341–2356.
Simmonds, D. J., Pekar, J. J., & Mostofsky, S. H. (2008).
Meta-analysis of Go/No-go tasks demonstrating that fMRI
activation associated with response inhibition is task-
dependent. Neuropsychologia, 46, 224–232.
Misirlisoy and Haggard
303
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
o
0
c
4
n
6
_
8
a
/
_
j
0
o
0
c
4
n
7
9
_
a
p
_
d
0
0
b
4
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Sirigu, A., Daprati, E., Ciancia, S., Giraux, P., Nighoghossian, N.,
Posada, A., et al. (2004). Altered awareness of voluntary
action after damage to the parietal cortex. Nature
Neuroscience, 7, 80–84.
Smith, J. L., Johnstone, S. J., & Barry, R. J. (2008). Movement-
related potentials in the go/nogo task: The P3 reflects both
cognitive and motor inhibition. Clinical Neurophysiology,
119, 704–714.
Sunaert, S., Van Hecke, P., Marchal, G., & Orban, G. A. (2000).
Attention to speed of motion, speed discrimination,
and task difficulty: An fMRI study. Neuroimage, 11,
612–623.
Tanji, J., & Hoshi, E. (2008). Role of the lateral prefrontal cortex in
executive behavioral control. Physiological Reviews, 88, 37–57.
Tanji, J., & Shima, K. (1994). Role for supplementary motor
area cells in planning several movements ahead. Nature,
371, 413–416.
Verbruggen, F., & Logan, G. D. (2008). Response inhibition in
the stop-signal paradigm. Trends in Cognitive Sciences, 12,
418–424.
Walsh, E., Kühn, S., Brass, M., Wenke, D., & Haggard, P. (2010).
EEG activations during intentional inhibition of voluntary
action: An electrophysiological correlate of self-control?
Neuropsychologia, 48, 619–626.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
2
2
6
2
/
9
2
6
/
1
2
9
9
4
6
6
/
4
1
5
7
9
8
o
0
c
4
n
6
_
8
a
/
_
j
0
o
0
c
4
n
7
9
_
a
p
_
d
0
0
b
4
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
304
Journal of Cognitive Neuroscience
Volume 26, Number 2