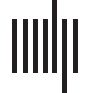The Temporal Modulation Structure of
Infant-Directed Speech
Victoria Leong1, Marina Kalashnikova2, Denis Burnham2, and Usha Goswami1
1Centre for Neuroscience in Education, Abteilung für Psychologie, University of Cambridge
2The MARCS Institute for Brain, Behaviour and Development, Western Sydney University
Schlüsselwörter: infant-directed speech, Phase, oscillations, Amplitudenmodulation
Keine offenen Zugänge
Tagebuch
ABSTRAKT
The temporal modulation structure of adult-directed speech (ADS) is thought to be encoded
by neuronal oscillations in the auditory cortex that fluctuate at different temporal rates.
Oscillatory activity is thought to phase-align to amplitude modulations in speech at
corresponding rates, thereby supporting parsing of the signal into linguistically relevant units.
The temporal modulation structure of infant-directed speech (IDS) is unexplored. Here we
compare the amplitude modulation (BIN) structure of IDS recorded from mothers speaking,
over three occasions, to their 7-, 9-, and 11-month-old infants, and the same mothers
speaking ADS. Analysis of the modulation spectrum in each case revealed that modulation
energy in the theta band was significantly greater in ADS than in IDS, whereas in the delta
band, modulation energy was significantly greater for IDS than ADS. Außerdem, Phase
alignment between delta- and theta-band AMs was stronger in IDS compared to ADS. Das
remained the case when IDS and ADS were rate-normalized to control for differences in
speech rate. These data indicate stronger rhythmic synchronization and acoustic temporal
regularity in IDS compared to ADS, structural acoustic differences that may be important for
early language learning.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
Zitat: Leong, V., Kalashnikova, M.,
Burnham, D., & Goswami, U. (2017).
The Temporal Modulation Structure of
Infant-Directed Speech. Open Mind:
Discoveries in Cognitive Science,
1(2), 78–90. https://doi.org/10.1162/
opmi_a_00008
DOI:
https://doi.org/10.1162/opmi_a_00008
Erhalten: 27 April 2016
Akzeptiert: 27 Januar 2017
Konkurrierende Interessen: The authors
declare no competing interests exist.
Korrespondierender Autor:
Usha Goswami
ucg10@cam.ac.uk
Urheberrechte ©: © 2017
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
EINFÜHRUNG
Human speech perception relies in part on neural tracking of the temporal modulation patterns
in speech at different timescales simultaneously: “multi-time resolution processing” (Chait,
Greenberg, Arai, Simon, & Kacke, 2015; Ghitza & Greenberg, 2009; Greenberg, 2006;
Luo & Kacke, 2007; Kacke, 2003). According to multi-time resolution models, cortical
oscillations entrain or phase-align their activity to modulations at corresponding timescales in
the signal, thereby encoding the different energy patterns, and binding the information together
in the final speech percept (Ghitza, 2011; Ghitza, Giraud, & Kacke, 2012; Giraud &
Kacke, 2012; Kacke, 2014). Exploration of the temporal characteristics of adult-directed
Rede (ADS) has revealed that accurate oscillatory phase alignment is mediated in part by
amplitude “rise times,” auditory “edges” associated with amplitude (Energie) modulations that
help to specify temporal modulation rates (Gross et al., 2013). Rise times in the theta band
appear to be particularly important for successful speech encoding, as acoustic “landmarks”
(amplitude rise times) in critical band envelopes in the theta range of ADS provide perceptual
markers that are critical for speech intelligibility (Doelling, Arnal, Ghitza, & Kacke, 2014).
When acoustic landmarks in the theta range (in this study, 2–9 Hz) were removed, Rede
became unintelligible, and when artificial landmarks of uniform height were inserted instead,
the speech became intelligible again. Entsprechend, Doelling et al. argued that auditory “edges”
Modulation Structure of Infant-Directed Speech Leong et al.
(amplitude rise times) in the theta band drive oscillatory activity to track and entrain to speech
at its syllabic rate. Vor allem, all experiments to date have investigated literate adult participants.
Im Gegensatz, in child populations, delta band information appears to be critical for suc-
cessful oscillatory entrainment. Studies of children with poor literacy (developmental dyslexia)
reveal impaired encoding of speech envelopes between 0–2 Hz (Molinaro et al., 2016; Power,
Colling, Mead, Barnes, & Goswami, 2016). Children with dyslexia also show atypical phase
entrainment to rhythmic speech in the delta band, but equivalent entrainment to control chil-
dren in the theta band (Power, Mead, Barnes, & Goswami, 2013). Weiter, studies of oscillatory
entrainment to speech by typically developing children show a significant relationship between
theta entrainment and learning to read (Power, Mead, Barnes, & Goswami, 2012). For child
Populationen, daher, it is possible that acoustic information in frequency bands other than
theta may be of equal or even greater importance for speech processing. To explore this pos-
sibility, information about the temporal modulation structure of child-directed speech (CDS)
and infant-directed speech (IDS) is required.
To explore CDS, we recently applied a probabilistic amplitude demodulation approach
to modeling the rhythm patterning in children’s nursery rhymes (z.B., capturing whether they
are iambic or trochaic). We compared its output to that of an engineered modulation filterbank
Ansatz (Leong, 2012; Leong, Stein, Turner, & Goswami, 2014; Turner, 2010), ultimately
deriving a Spectral-Amplitude Modulation Phase Hierarchy (S-AMPH) Modell (Leong, 2012;
Leong & Goswami, 2015). The S-AMPH modeling demonstrated that English nursery rhymes
contained amplitude modulations in three critical temporal rate bands (corresponding to delta,
theta, and beta/low gamma, for this speech corpus 0.9–2.5 Hz, 2.5–12 Hz, and 12–40 Hz; sehen
(Leong, 2012; Leong & Goswami, 2015). Amplitude modulations in these bands were hier-
archically nested in the nursery rhymes, and phase alignment between the slower amplitude
modulation (BIN) bands (delta and theta) played a key role in the perception of rhythmic pat-
terning (judging whether the nursery rhymes were trochaic or iambic). A strong syllable was
perceived when delta and theta modulation peaks were in alignment. Entsprechend, the strong
rhythmic character and acoustic temporal regularity of spoken nursery rhymes appears to be
influenced by amplitude modulations in the delta band.
Like nursery rhymes, IDS is a highly rhythmic and temporally regular signal (Fernald
et al., 1989;
Jusczyk, Cutler, & Redanz, 1993). Entsprechend, it is plausible that delta band
information and delta-theta phase alignment may also be important characteristics of IDS.
Infant-directed speech is classically described in terms of its higher pitch and greater pitch
range, slower rate and higher amplitude than ADS, with hyperarticulated vowels (Burnham,
Kitamura, & Vollmer-Conna, 2002; Kuhl et al., 1997). Vowel hyperarticulation in IDS has
been proposed to support phoneme discrimination (Liu, Kuhl, & Tsao, 2003) and word recog-
Nation (Song, Demuth, & Morgan, 2010), while high pitch and greater pitch range capture
infant attention (Fernald & Kuhl, 1987) and facilitate speech segmentation (Thiessen, Hill, &
Saffran, 2005). Neural multi-time resolution models of speech encoding (Giraud & Kacke,
2012) raise the possibility that the temporal modulation hierarchy may also provide acoustic
landmarks that support infant language learning, helping infants to locate word and phrase
boundaries. Currently, the temporal modulation structure of IDS is unexplored. Entsprechend,
here we applied the S-AMPH modelling approach (Leong & Goswami, 2015) to IDS.
The S-AMPH model is a low-dimensional representation of the speech signal (see also
Greenberg & Arai, 2001). The model generates a hierarchical representation of the domi-
nant spectral (acoustic frequency) and temporal (oscillatory rate) modulation patterns in the
OPEN MIND: Discoveries in Cognitive Science
79
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
speech envelope. Amplitude modulation patterns in three temporal rate bands (~delta, theta,
and beta/low gamma) form a three-tier nested hierarchy, thereby preserving AM patterning
across different speaker rates. Annotation of the nursery rhyme (CDS) corpus revealed that
for these nursery rhymes, this AM hierarchy (centered on ~2 Hz, ~5 Hz, and ~20 Hz AMs)
mirrored the linguistic phonological hierarchy of stressed syllables, syllables, and onset-rime
units in the spoken material (Leong & Goswami, 2015). Oscillatory cycles in each AM band
thus supported the identification of phonological units of different sizes. The number of bands
and their respective bandwidths used by the S-AMPH were originally determined using
principal component analysis (PCA) dimensionality reduction of original high-dimensional
spectral (29 equivalent rectangular bandwidth [ERB]-spaced frequency channels spanning 100–
7,250 Hz) and temporal (24 modulation channels spanning 0.9–40 Hz) envelope representa-
tionen (Leong, 2012; Leong & Goswami, 2015).
Here we applied the S-AMPH to IDS collected from a sample of 24 mother–infant dyads
bei 7, 9, Und 11 Monate alt, and we also analyzed ADS collected from the same mothers.
While application of the S-AMPH formulae to novel corpora cannot be assumed to identify the
same linguistic units, the methods nevertheless enable comparison of relative speech energy in
delta, theta, and beta/low-gamma bands and AM phase synchronization. We were interested
to see whether natural IDS has a different temporal modulation structure to ADS, and whether
the strength of phase alignment between the different AM bands (delta, theta, beta/low gamma)
might show a different patterning in IDS as compared to ADS.
METHODEN
Teilnehmer
Twenty-four mother-infant dyads (9 männlich, 15 female infants) participated in the study.1 None
of the families had any history of language or cognitive disorders. All mothers spoke Australian
English as their native language, and had normal (not > 0.5 SDs below the mean) nonverbal
IQ on a combined subtest score averaging block design and matrix reasoning subscales of
the Wechsler Adult Intelligence Scale IV (WAIS-IV; Wechsler, 2008) (M = 12.2, SD = 2.0,
compared with the population mean, 10.0, and SD, 3.0).
Design and Experimental Setup
There were four speaking conditions, three IDS, and one ADS. Infant-directed speech sam-
ples were collected longitudinally at three time-points, when the infants were 7 (Durchschnittsalter =
31 weeks, SD = 1.1), 9 (Durchschnittsalter = 40 weeks, SD = 1.2), Und 11 (Durchschnittsalter = 48 weeks,
SD = 1.2) Monate alt. Mothers were told that the purpose of the experiment was to cap-
ture their natural interactions with their infants during a brief play session and soft toys and
pictures were provided. Mothers were asked to talk to their infants for as long as they felt
appropriate. All mothers and infants were observed to be motivated and engaged in the task.
Mother and infant interacted alone in a laboratory room. The mother sat facing the infant,
who was sitting in a highchair. A video camera was mounted in each corner of the room to
allow for monitoring and video recording of the session, but only the audio data are analyzed
Hier. The mother wore a head-mounted microphone (AudioTechnica AT892), connected to
Adobe Audition CS6 software via an audio input/output device (MOTU Ultralite MK3). Speech
samples were digitally recorded at 16 kHz (or at 44.1 kHz and resampled to 16 kHz). For the
1 A subset of nine mother-infant dyads also contributed data to an earlier study piloting these analyses (Leong,
Kalashnikova, Burnham, & Goswami, 2014).
OPEN MIND: Discoveries in Cognitive Science
80
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
ADS recordings, each mother interacted with a female experimenter in the laboratory room
using the same recording apparatus, and the infant was not present. Mothers were interviewed
about the experimental play sessions. Adult-directed speech samples were collected when the
infant was approximately 12 months old (mean age: 53 weeks, SD = 8.2). Both IDS and ADS
recording sessions lasted between 5 Und 10 minutes.
Speech Data
The raw audio recordings were manually divided into short segments for analysis using Praat
Software (Boersma & Weenink, 2013). Each segment contained a complete phrase from the
original utterance. Portions of speech that were interrupted by the infant or adult addressee,
or that contained excessive background noise, were not used. In Summe, across the four speak-
ing conditions, mothers contributed 679 speech segments for analysis. On average, segments
were 9.13s in length (range 5.02s–16.92s), and mothers contributed an average of 7.8, 7.9, Und
7.1 segments at the infant ages of 7, 9, Und 11 months, jeweils. Mothers contributed
on average 6.1 segments in ADS. The mean length of speech segments for the different
speaking conditions was 9.2s (7 months IDS), 9.5S (9 months IDS), 9.1S (11 months IDS),
and 8.7s (ADS).
Speech Analysis Method
Each speech segment was
Schritt 1: The S-AMPH Representation of the Amplitude Envelope
z-scored (to standardize its mean and standard deviation), and a low-dimensional spectro-
temporal model (d.h., S-AMPH) of its amplitude envelope was extracted using a two-stage
filtering process, as described in Leong (2012) and Leong and Goswami 2015. Erste, Die
z-scored acoustic signal was band-pass filtered into five frequency bands (channel edge fre-
quencies: 100, 300, 700, 1,750, 3,900, Und 7,250 Hz) using a series of adjacent finite impulse
response (FIR) filters. Nächste, the Hilbert envelope was extracted from each band-filtered signal
and these five Hilbert envelopes were down-sampled to 1,050 Hz and passed through a second
series of band-pass filters to isolate the three different AM bands within the envelope
modulation spectrum.
These three AM bands corresponded to delta rate modulations
(0.9–2.5 Hz), theta rate modulations (2.5–12 Hz) and beta/low-gamma rate modulations,
jeweils (12–40 Hz). The result of this two-step filtering process was a 5 (frequency) X
3 (rate) spectro-temporal representation of the speech envelope, described as comprising
delta-,
theta-, and beta/low gamma-rate AM bands for each of the five spectral bands
(Leong & Goswami, 2015).
Schritt 2: Multi-timescale Synchronization Measure: Phase Synchronization Index (PSI) Nächste, a mea-
sure of multi-timescale phase synchronization, the Phase Synchronization Index (PSI), War
computed between adjacent modulation rate bands in the S-AMPH representation of each
speech segment (d.h., delta-theta phase synchronization and theta-beta/low-gamma phase syn-
chronization). This PSI computation was performed separately for each of the five spectral
bands in the S-AMPH representation. The n : m PSI was originally conceptualized to quantify
phase synchronization between two oscillators of different frequencies (z.B., muscle activity;
Tass et al., 1998), and was subsequently adapted for use in neural analyses of oscillatory
phase-locking (z.B., Schack & Weiss, 2005). This adaptation is applied to speech AMs in our
Modell. The PSI was computed as:
OPEN MIND: Discoveries in Cognitive Science
PSI = |(cid:2)e1(nθ
1
−mθ
)(cid:3)|
2
(1)
81
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
Figur 1. Hypothetical amplitude modulation (BIN) pairs (sinusoids) that yield Phase Synchroniza-
tion Index (PSI) scores of 1 (links) Und 0.07 (Rechts), jeweils. The red and blue curves represent
AMs with a frequency ratio of 1:2.
In Equation 1, n and m are integers describing the frequency relationship between the two
AMs being compared. Following previous studies (Leong & Goswami, 2014, 2016; Leong,
Stein, et al., 2014), for the delta-theta AM band analysis, an n : m ratio of 2:1 was used, while
for the theta-beta/low-gamma AM band analysis, an n : m ratio of 3:1 was used. The values
θ1 and θ2 refer to the instantaneous phase of the two AMs at each point in time. daher,
(nθ1–mθ2) is the generalized phase difference between the two AMs, which was computed
by taking the circular distance (modulus 2π) between the two instantaneous phase angles.
The angled brackets denote averaging of this phase difference over all time-points. The PSI is
the absolute value of this average, and can take values between 0 (no synchronization) Und
1 (perfect synchronization), as illustrated in Figure 1. A sound with a PSI of 1 is perceived as
being perfectly rhythmically regular (a repeating pattern of strong and weak beats), whereas a
sound with a PSI of 0 is perceived as being random in rhythm.
To assess the modulation spectrum of the speech samples, Die
Schritt 3: Modulation Spectrum
sub-band Hilbert envelopes of the stimuli (resulting from stage 1 of the previously-described
S-AMPH decomposition procedure) were individually passed through a modulation FIR filter-
bank with 24 channels logarithmically spaced between 0.9–40 Hz. For each speech sample,
and each frequency sub-band, the mean power across all modulation channels was computed,
and the relative power difference from this mean was computed for each modulation channel.
Endlich, the differenced modulation power spectrum was averaged across the five frequency
sub-bands for each speech sample, and this grand average was used for statistical analysis.
Control Analysis With Rate-Normalized Data
The speaking rate of IDS was slower than that of ADS. As this could introduce potential con-
founds, a rate-normalization procedure was also performed, rescaling the IDS data samples to
the same temporal rate as the ADS samples. Erste, each IDS and ADS speech sample was man-
ually annotated (by native English speakers) to ascertain the number of syllables and stressed
syllables that it contained. This provided a mean syllable rate and stressed syllable rate per
second for each participant in each speaking condition. For each IDS condition, and for each
participant, a scaling factor was then computed based on the ratio between their stressed sylla-
ble rate in the ADS condition, and their stressed syllable rate in the IDS conditions. This scaling
factor was then used to resample each IDS utterance to the appropriate length for that speaker
(z.B., if the stress rate for IDS was half that of ADS, then the IDS sample was compressed so
that it was half of its original length). Identical signal processing steps and statistical analyses
were then conducted with the rescaled data.
OPEN MIND: Discoveries in Cognitive Science
82
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
RESULT
Natural Speech
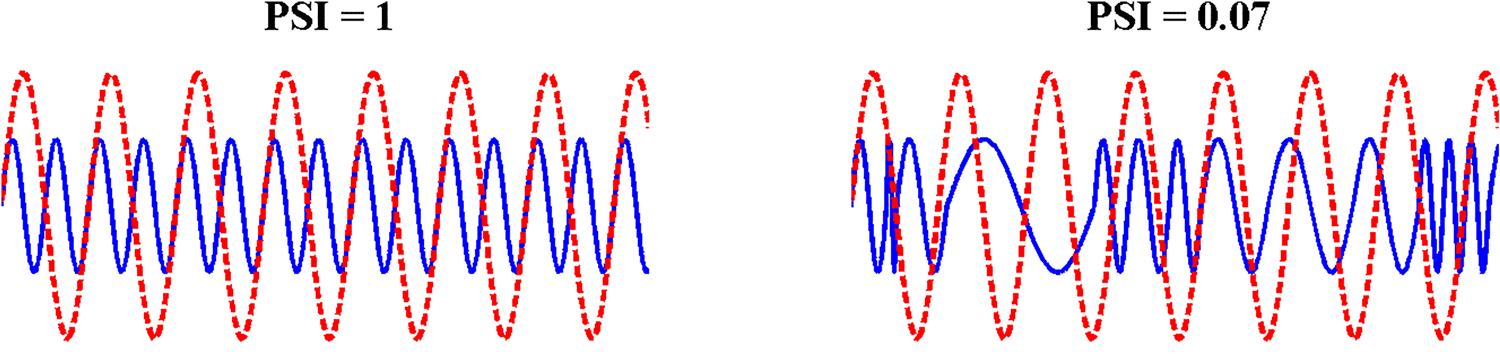
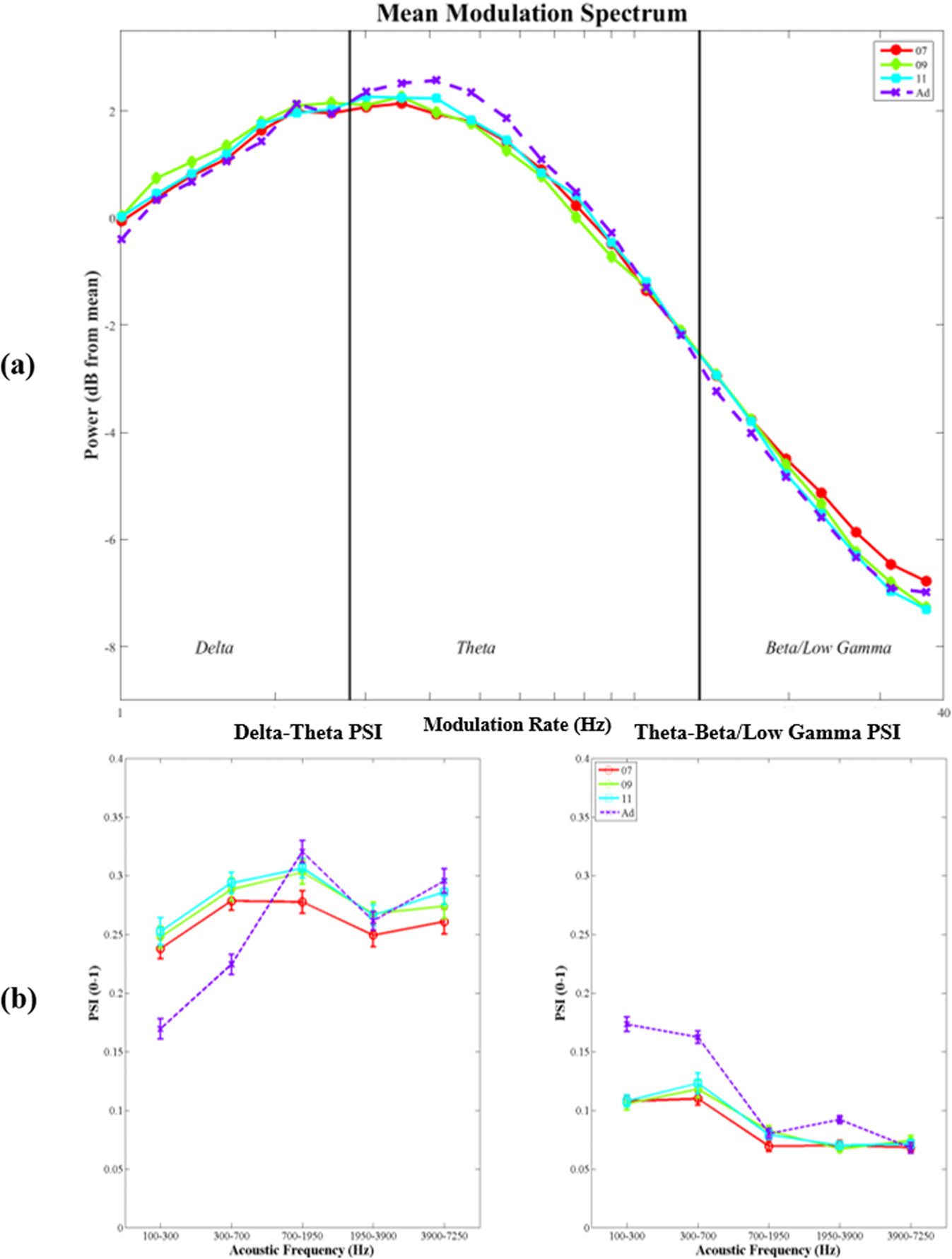
Figur 2 (panel a) shows the mean modulation spectrum of the speech
Modulation Spectrum
amplitude envelope for each speaking condition. Compared to ADS, the modulation spec-
trum of IDS contained more energy in the delta band (0.9–2.5 Hz), whereas the modulation
spectrum of ADS showed the typical profile of most energy in the theta band, as previously
reported (Greenberg, Carvey, Hitchcock, & Chang, 2003). To assess whether these differences
were statistically significant, the area under the curve corresponding to the three modulation
bands was computed (shown in Figure 2, panel b) and assessed using a repeated measures
ANOVA with modulation rate (three levels) and speaking condition (four levels) as within-
subjects factors. There was a significant interaction between Rate and Condition, F(6, 138) =
7.78, P < .0001, showing that the relative amount of modulation energy in the delta, theta
and beta/low-gamma bands differed between IDS and ADS. Tukey post-hoc analysis revealed
that all three IDS conditions contained significantly more modulation energy than ADS in
the delta band (p < .05 for all). Conversely, for the theta band, ADS contained significantly
more modulation energy than IDS at 7 and 9 months (p < .05 for both), but not at 11 months
(p = .059). In the beta/low-gamma band, ADS contained significantly more modulation energy
than 9- and 11-month IDS (p < .05 for both) but did not differ from 7-month IDS ( p = .81).
Finally, modulation energy did not differ between the three IDS conditions for any AM band
(p > .93 for all).
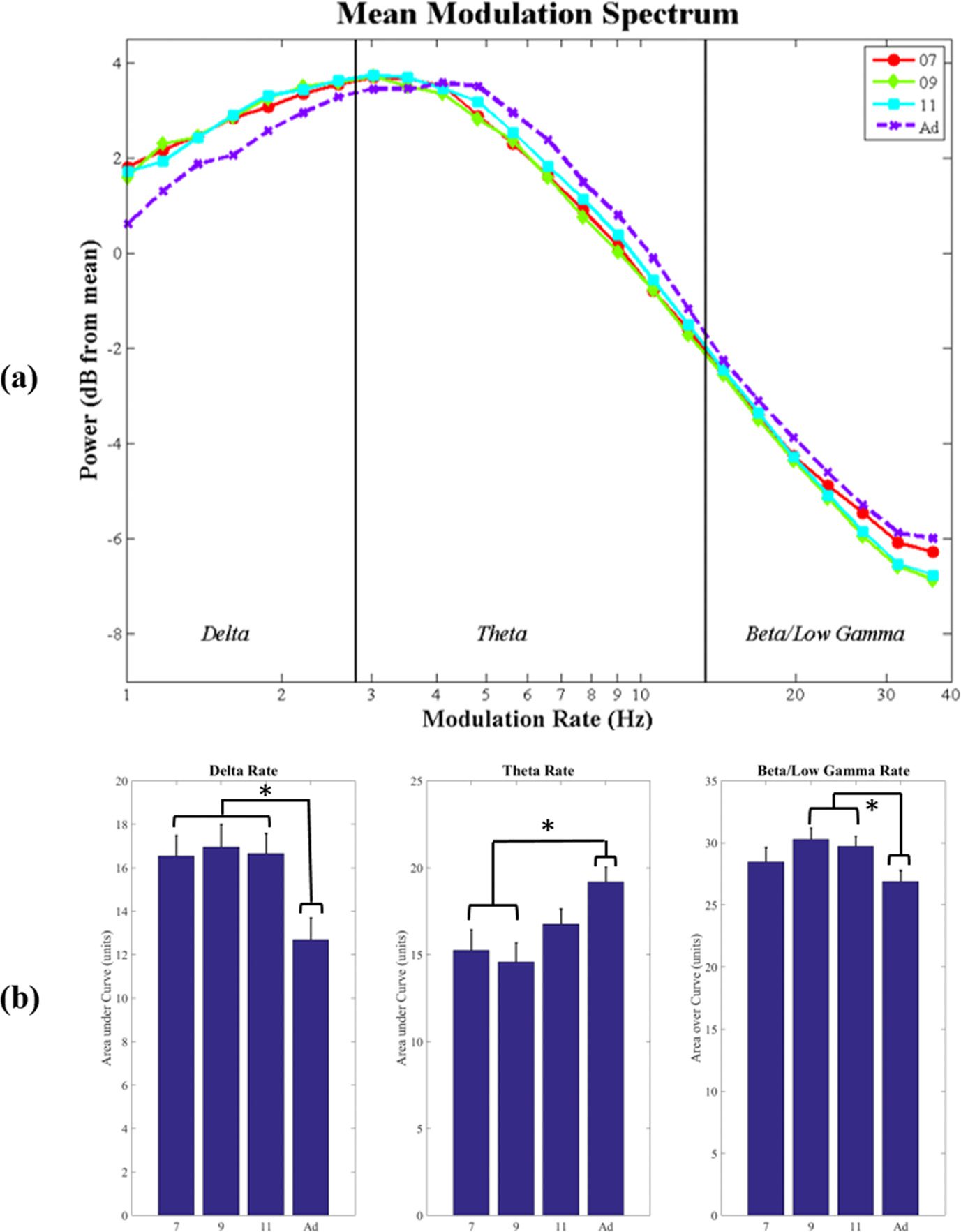
the phase synchronization between AM bands in the
Phase Synchronization (PSI) Nächste,
mothers’ IDS was assessed using the PSI between pairs of AM bands in the S-AMPH repre-
sentation of the speech envelope. Mean PSI values are shown in Figure 3. Each subplot shows
the PSI value (ranging from 0 [no synchronization] Zu 1 [perfect synchronization]) on the y-
axis, and acoustic frequency on the x-axis. The figure shows more synchronization in IDS than
ADS for delta-theta AM band synchronization in low-frequency spectral bands (100–300 Hz
and 300–700 Hz), but less synchronization for theta-beta/low-gamma AM bands. Darüber hinaus,
for delta-theta band phase synchronization in the middle spectral frequency band (700–
1,950 Hz), typically corresponding to vocalic energy, PSI values increase with increasing age
of the addressee (7M < 9m < 11m < ADS).
To compare the conditions statistically, two repeated measures ANOVAs were performed,
taking either delta-theta band PSIs or theta-beta/low-gamma band PSIs as the dependent vari-
able. In each ANOVA, speaking condition (four levels) and spectral frequency band (five levels)
were the within-subjects factors. For the delta-theta band ANOVA, there were significant main
effects of Condition, F(3, 69) = 3.07, p < .05, and Spectral Band, F(4, 92) = 27.76, p <
.001. The interaction between Condition and Spectral Band was also significant, F(12, 276) =
17.92, p < .001. Post-hoc analysis (Newman-Keuls) indicated that for the two lowest spec-
tral frequency bands (100–300 Hz; 300–700 Hz), PSI scores for all three IDS conditions were
significantly higher than for ADS (p < .01 for all comparisons). By contrast, for the middle
and highest spectral frequency bands (700–1,950 Hz; 3,900–7,250 Hz), PSI scores for all IDS
conditions were significantly lower than for ADS (p < .05 for all comparisons). The PSI scores
for the 1,950–3,950 spectral frequency band did not differ between IDS and ADS. Finally, for
the middle spectral frequency band (700–1,750 Hz), there was a graded effect between IDS
at the different ages. Delta-theta AM band phase synchronization in IDS to 7-month-olds was
lower (less synchronized) than in IDS to 11-month-olds (p < .05). However, it did not differ
OPEN MIND: Discoveries in Cognitive Science
83
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
o
p
m
_
a
_
0
0
0
0
8
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
o
p
m
_
a
_
0
0
0
0
8
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Panel (a). Mean modulation spectrum for infant-directed speech (IDS) (“07,” “09,” and
“11” months) and adult-directed speech (ADS) (“Ad”) conditions, computed for each of five spectral
frequency bands (100–300 Hz; 300–700 Hz; 700–1,750 Hz; 1,750–3,900 Hz; 3,900–7,250 Hz)
and then averaged across bands and speakers. The x-axis shows the modulation rate, and the
y-axis the power in dB (normalized with respect to the mean power of each frequency band in
each sample). Vertical lines indicate the boundaries used to delineate the area under the curve for
Spectral-Amplitude Modulation Phase Hierarchy (S-AMPH) delta-, theta-, and beta/low-gamma-rate
amplitude modulation (AM) bands. Panel (b). Area under the curve computed for the modulation
spectrum in the delta, theta, and beta/low-gamma ranges for each speaking condition (“7” = 7 month
IDS, “9” = 9 month IDS, “11” = 11 month IDS, “Ad” = ADS). Note that for the beta/low-gamma
range, as the curve falls below zero, the area over the curve is reported instead. Here, a smaller
numerical value denotes greater power in the beta/low-gamma range.
statistically from delta-theta AM band phase synchronization in IDS to 9-month-olds (p = .14).
This suggests that the rhythmic regularity of IDS increases as a function of infant age.
For the theta-beta/low-gamma AM band ANOVA, there were again significant main
effects of Condition, F(3, 69) = 45.84, p < .001, and Spectral Band, F(4, 92) = 119.88,
OPEN MIND: Discoveries in Cognitive Science
84
Modulation Structure of Infant-Directed Speech Leong et al.
Figure 3. Phase Synchronization Index (PSI) values computed for the delta-theta amplitude mod-
ulation (AM) bands (left) and the theta-beta/low-gamma AM bands (right), for infant-directed speech
(IDS) collected at each different age (different-colored solid lines) and adult-directed speech (ADS)
(dotted line). The x-axis indicates the frequency (spectral) band from which the AMs originate. The
y-axis shows the PSI value. Error bars indicate the standard error of the mean.
p < .001, and a significant interaction between Condition and Spectral Band, F(12, 276) =
18.21, p < .001. However, Newman-Keuls post-hoc analysis revealed a reverse symmetry to
the results obtained for the delta-theta AM band comparisons. For the majority of spectral fre-
quency bands (first, second, and fourth; 100–300 Hz; 300–700 Hz; 1,950–3,900 Hz), the IDS
conditions showed lower PSI scores than ADS (p < .001 for all comparisons). However, PSI
scores in the middle and highest spectral frequency bands (700–1,950 Hz; 3,900–7,250 Hz)
did not differ between speaking conditions (p > .05 for all comparisons).
Control Analysis: Rate-Normalized Data
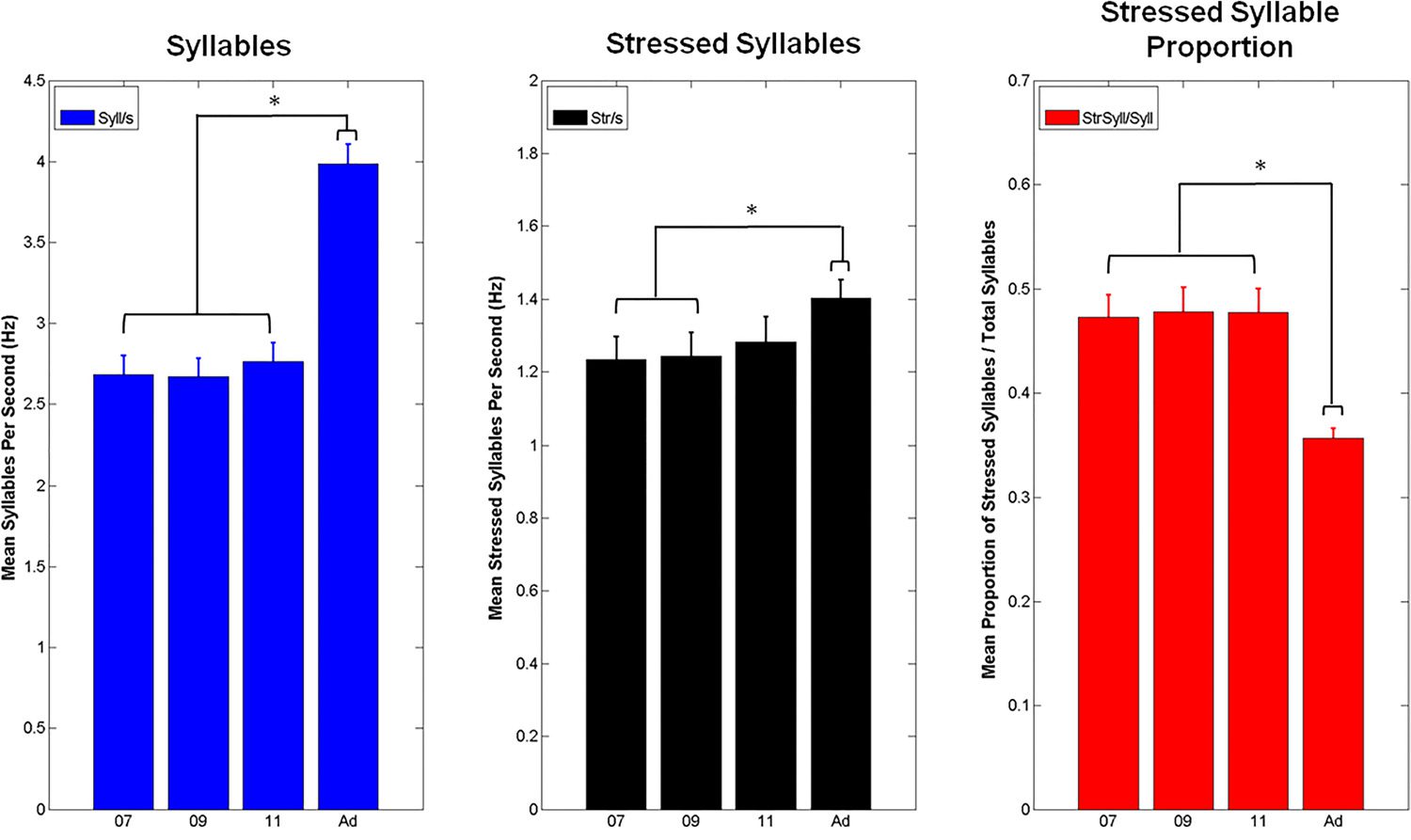
Figur 4 shows the mean number of syllables and
Syllables and Stressed Syllables per Second
stressed syllables produced per second for IDS and ADS, along with the mean proportion of
stressed syllables as a function of the total number of syllables produced. We used repeated
measure ANOVAs to compare speaking conditions in each case. For the number of syllables
produced, the ANOVA showed a significant main effect of speaking condition, F(3, 69) =
74.42, P < .0001). Tukey post-hoc analysis showed that ADS had a significantly greater num-
ber of syllables per utterance than the three IDS conditions ( p < .001), which did not differ
significantly from each other. For the number of stressed syllables produced, the ANOVA
again showed a significant main effect of speaking condition, F(3, 69) = 3.42, p < .05. Tukey
post-hoc analysis revealed significantly more stressed syllables per second in ADS compared
to IDS at 7 months and 9 months ( p < .05), but not IDS at 11 months. Finally, the ANOVA for
the proportion of stressed syllables produced in the different speaking conditions also showed
a significant effect of condition, F(3, 69) = 28.33, p < .00001. Here, post-hoc tests showed
that ADS contained a significantly lower proportion of stressed syllables than IDS at any age
(p < .001 for all comparisons). Therefore, although ADS contained more syllables and more
stressed syllables than IDS within a given period of time (i.e., a quantitative rate effect), there
was a qualitative difference between the speaking styles. Infant-directed speech contained a
OPEN MIND: Discoveries in Cognitive Science
85
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
o
p
m
_
a
_
0
0
0
0
8
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
Figure 4. Mean number of syllables per second, stressed syllables per second, and proportion
of syllables that were stressed for the infant-directed speech (IDS) and adult-directed speech (ADS)
data for each speaking condition (“07” = 7 month IDS, “09” = 9 month IDS, “11” = 11 month IDS,
“Ad” = ADS).
higher proportion of stressed to unstressed syllables. In IDS, nearly half (~48%) of all syllables
were stressed, whereas in ADS, only around one-third (~36%) of syllables were stressed.
Modulation Spectrum
The modulation spectra for the rate-normalized control data are shown in Figure 5 (panel a).
Comparison with Figure 2 suggests that rate-normalization has not changed the pattern of
IDS/ADS difference. A 3 × 4 ANOVA (Modulation Rate x Condition) showed the predicted
interaction between Rate and Condition, F(6, 138) = 2.16, p < .05, one-tailed. Post-hoc anal-
ysis (Fisher LSD) showed that IDS at 9 months contained significantly more modulation energy
in the delta band than in ADS (p < .05). For the theta band, ADS contained significantly more
energy than IDS at 7 and 9 months (p < .05 for both), but not at 11 months (p = .077). Finally,
in the beta/low-gamma band, ADS contained significantly more energy than IDS at 7 months
(p < .05). Therefore, the patterns observed with natural speech were broadly conserved after
the rate-normalization procedure.
The PSI values obtained for the rate-normalized data are
Phase synchronization (PSI)
also shown in Figure 5 (panel b). Two 4 x 5 (Speaking Condition x Spectral Band) ANOVAs
were again performed, again taking either delta-theta band PSIs or theta-beta/low-gamma band
PSIs as the dependent variable. For the delta-theta band ANOVA, there were significant main
effects of Condition, F(3, 69) = 5.82, p < .01, and Spectral Band, F(4, 92) = 41.41, p < .0001,
and a significant interaction, F(12, 276) = 9.88, p = .0001. Tukey post-hoc analysis indicated
that ADS had significantly lower PSI scores (p < .001) than all three IDS conditions for the
two lowest spectral frequency bands (100–300 Hz; 300–700 Hz). By contrast, for the middle
spectral frequency band (700–1,950 Hz), PSI scores for ADS were significantly higher than for
IDS, but at 7 months only (p < .01).
OPEN MIND: Discoveries in Cognitive Science
86
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
o
p
m
_
a
_
0
0
0
0
8
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
o
p
m
_
a
_
0
0
0
0
8
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Panel (a). Mean modulation spectrum for rate-normalized infant-directed speech (IDS)
(“07”, “09,” and “11” months) and adult-directed speech (ADS) (“Ad”) conditions, computed for
each of five spectral frequency bands (100–300 Hz; 300–700 Hz; 700–1,750 Hz; 1,750–3,900 Hz;
3,900–7,250 Hz) and then averaged across bands and speakers. The x-axis shows the modula-
tion rate, and the y-axis the power in dB (normalized with respect to the mean power of each
frequency band in each sample). Vertical lines indicate the boundaries used to delineate the area
under the curve for Spectral-Amplitude Modulation Phase Hierarchy (S-AMPH) delta-, theta- and
beta/low-gamma-rate amplitude modulation (AM) bands. Panel (b). Phase Synchronization Index
(PSI) values computed for the rate-normalized data, for delta-theta AM bands (left) and the theta-
beta/low-gamma AM bands (right), for IDS collected at each different age (different-colored solid
lines) and ADS (dotted line). The x-axis indicates the frequency (spectral) band from which the AMs
originate. The y-axis shows the PSI value. Error bars indicate the standard error of the mean.
For the theta-beta/low-gamma AM band ANOVA, there were again significant main
effects of Condition, F(3, 69) = 27.76, p < .00001, and Spectral Band, F(4, 92) = 96.69,
p < .0001, and a significant interaction, F(12, 276) = 15.30, p < .0001. Tukey post-hoc anal-
ysis revealed that ADS scores were significantly higher (p < .01) than IDS scores at all ages in
the first, second, and fourth spectral bands (100–300 Hz; 300–700 Hz; 1,950-3,900 Hz), but
OPEN MIND: Discoveries in Cognitive Science
87
Modulation Structure of Infant-Directed Speech Leong et al.
did not differ for the third and fifth spectral bands (700–1,950 Hz; 3,900-7,250 Hz).Therefore,
the results of the PSI analysis with rate-normalized data were again consistent with the PSI
analysis conducted with natural speech.
DISCUSSION
The temporal modulation structure of IDS differs significantly from that of ADS. First, there
is significantly greater modulation energy in the delta band in IDS relative to ADS, while
ADS shows greater energy in the theta band compared to IDS. This difference is not simply
a consequence of IDS typically being spoken more slowly than ADS, as demonstrated by the
rate-normalized control analysis. The higher delta power in IDS was accompanied by a higher
proportion of syllables being stressed in IDS as compared to ADS, providing a more rhythmic
input. Second, IDS showed significantly greater phase synchronization between the slower
delta- and theta-rate AM bands compared to ADS. This finding was also mirrored in the con-
trol analysis with rate-normalized data. Accordingly, there are clear differences in the pattern
of rhythmic synchronization between IDS and ADS. Greater phase synchronization between
delta- and theta-rate AM bands accompanies greater rhythmic regularity in IDS: syllables are
stressed more regularly.
These findings have important implications for the neural basis of language acquisition,
at least for English. The temporal rate bands at these different timescales can be assumed to
entrain neuronal oscillatory activity (Giraud & Poeppel, 2012). For adult speech, oscillations
in the delta (1–3 Hz), theta (4–8 Hz), and beta/low-gamma bands (15–30 Hz / >30 Hz) bzw-
tively have been shown to be critical for encoding (frequency ranges from Poeppel, 2014).
One proposal is that oscillatory entrainment to amplitude modulations in these bands helps to
parse the speech signal into linguistically relevant units (delta—syllable stress patterns, theta—
syllables, beta—onset-rime units, low gamma—phonetic information). Our data suggest that
IDS is acoustically structured to facilitate neural entrainment to delta band (prosodic) informa-
tion by the infant brain. In der Tat, a large infant behavioral literature attests to the importance
of stressed syllables in early word segmentation (z.B., Cutler & Norris, 1988; Echols, 1996).
Weiter, at least two studies of infant entrainment to nonspeech AMs have revealed significant
neuronal entrainment in this frequency range for German-learning infants, including new-
borns (Telkemeyer et al., 2009; Telkemeyer et al., 2011). The stronger phase synchronization
between delta- and theta-rate AM bands that characterizes IDS would further facilitate the ex-
traction of rhythmic patterning, Zum Beispiel, in distinguishing trochaic versus iambic prosodic
Struktur (Leong, Stein, et al., 2014). The infant electrophysiological literature shows that in-
fants are encoding the difference between trochees and iambs by 4 Monate alt (Weber et al.,
2004). These differences in phase synchronization strength between slower (IDS) versus faster
(ADS) timescales could also be related to recent demonstrations that phonetic contrasts in
IDS are less clear than in ADS (Martin et al., 2015; McMurray, Kovack-Lesh, Goodwin, &
McEchron, 2013).
In der Tat, phase synchronization at faster modulation timescales was greater in ADS, Und
this effect was also preserved in the rate-normalized data. Perceptually, this would increase the
rhythmic regularity of the acoustic relationship between syllables and phonemes, which could
potentially reflect the acquisition of literacy. A more regular temporal patterning of phonemes
within syllables may reflect the explicit awareness of phonemes in syllables that is a cognitive
consequence of learning to read (see Ziegler & Goswami, 2005, for data across languages).
Entsprechend, ADS may show stronger acoustic patterning at faster temporal rates because most
ADS speakers are literate. Experimental investigation of the temporal modulation structure of
OPEN MIND: Discoveries in Cognitive Science
88
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
ADS as produced by literate versus illiterate populations offers one method for exploring this
possibility further (Araújo, Flanagan, Castro-Caldas, & Goswami, 2016).
ACKNOWLEDGMENTS
We thank Maria Christou-Ergos for preparation of data for analysis. This work was supported by
funding from the Australian Research Council (DP110105123) to DB and UG. Victoria Leong
is now in the Division of Psychology, Nanyang Technological University, Singapur.
BEITRÄGE DES AUTORS
VL, DB, and UG conceived and designed the study, MK and DB collected the data, VL analyzed
the data, VL, MK, DB, and UG wrote the article.
VERWEISE
Araújo, J., Flanagan, S., Castro-Caldas, A., & Goswami, U. (2016,
Juni). The temporal modulation structure of illiterate versus liter-
ate speech. Poster presented at the 9th National Symposium on
Research in Psychology, Portuguese Psychological Association,
Faro, Portugal.
Boersma, P., & Weenink, D. (2013). Praat: Doing phonetics by com-
puter. Version 5.3.51, http://www.praat.org/
Burnham, D., Kitamura, C., & Vollmer-Conna, U. (2002) What’s
New Pussycat? On talking to babies and animals. Wissenschaft, 296,
1435.
Chait, M., Greenberg, S., Arai, T., Simon, J. Z., & Kacke, D.
(2015). Multi-time resolution analysis of speech: Evidence from
Psychophysics. Frontiers in Neuroscience, 9, 214.
Cutler, A., & Norris, D. (1988). The role of strong syllables in seg-
mentation for lexical access. Journal of Experimental Psychology:
Human Perception and Performance, 14, 113–121.
Doelling, K. B., Arnal, L. H., Ghitza, O., & Kacke, D.
(2014). Acoustic landmarks drive delta–theta oscillations to en-
able speech comprehension by facilitating perceptual parsing.
NeuroImage, 85, 761–768.
Echols, C. H. (1996). A role for stress in early speech segmentation.
In J. L. Morgan & K. Demuth (Hrsg.), Signal to syntax: Bootstrap-
ping from speech to grammar in early acquisition (S. 151–170).
Mahwah, NJ: Lawrence Erlbaum.
Fernald, A., & Kuhl, P. (1987). Acoustic determinants of infant pref-
erence for motherese speech. Infant Behavior and Development,
10, 279–293.
Fernald, A., Taeschner, T., Dunn, J., Papousek, M., De Boysson-
Bardies, B., & Fukui, ICH. (1989). A cross-language study of pro-
sodic modifications in mothers’ and fathers’ speech to preverbal
infants. Journal of Child Language, 16(3), 477–501.
Ghitza, Ö. (2011). Linking speech perception and neurophysiology:
Speech decoding guided by cascaded oscillators locked to the
input rhythm. Grenzen in der Psychologie, 2, 130.
Ghitza, O., Giraud, A.-L., & Kacke, D.
(2012). Neuronal
oscillations and speech perception: Critical-band temporal
envelopes are the essence. Grenzen der menschlichen Neurowissenschaften, 6,
340.
Ghitza, O., & Greenberg, S. (2009). On the possible role of brain
rhythms in speech perception: Intelligibility of time-compressed
speech with periodic and aperiodic insertions of silence. Phonet-
ica, 66, 113–126.
Giraud, A. L., & Kacke, D. (2012). Cortical oscillations and speech
Verarbeitung: Emerging computational principles and operations.
Naturneurowissenschaften, 15, 511–517.
Greenberg, S. (2006). A multi-tier framework for understanding
spoken language. In S. Greenberg & W. Ainsworth (Hrsg.), Un-
derstanding speech: An auditory perspective (S. 411–434).
Mahwah, NJ: Lawrence Erlbaum.
Greenberg, S., & Arai, T. (2001). The relation between speech intel-
ligibility and the complex modulation spectrum. Proceedings of
the 7th Eurospeech Conference on Speech Communication and
Technologie (Eurospeech-2001), 473–476.
Greenberg, S., Carvey, H., Hitchcock, L., & Chang, S. (2003). Tem-
poral properties of spontaneous speech: A syllable-centric per-
spective. Journal of Phonetics, 31, 465–485.
Brutto, J., Hoogenboom, N., Thut, G., Schyns, P., Panzeri, S., Belin,
P., & Garrod, S. (2013). Speech rhythms and multiplexed oscil-
latory sensory coding in the human brain. PLOS Biology, 11(12),
e1001752.
Jusczyk, P. W., Cutler, A., & Redanz, N.J. (1993). Infants’ prefer-
ence for the predominant stress patterns of English words. Child
Development, 64, 675–687.
Kuhl, P. K., Andruski, J. E., Chistovich, ICH. A., Chistovich, L. A.,
Kozhevnikova, E. V., Ryskina, V. L., Stolyarova, E. ICH., . . . Lacerda,
F. (1997). Cross-language analysis of phonetic units in language
addressed to infants. Wissenschaft, 277, 684–686.
Leong, V. (2012). Prosodic rhythm in the speech amplitude enve-
lope: Amplitude modulation phase hierarchies (AMPHs) Und
AMPH models (Doctoral dissertation). University of Cambridge.
Retrieved August 20, 2014, from http://www.cne.psychol.cam.
ac.uk/pdfs/phds/vleong
Leong, V., & Goswami, U. (2014). Assessment of rhythmic entrain-
ment at multiple timescales in dyslexia: Evidence for disruption
to syllable timing. Hörforschung, 308(100), 141–161.
OPEN MIND: Discoveries in Cognitive Science
89
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Modulation Structure of Infant-Directed Speech Leong et al.
Leong, V., & Goswami, U. (2015). Acoustic-emergent phonology
in the amplitude envelope of child-directed speech. PLOS ONE,
10(12), e0144411.
Leong, V., & Goswami, U. (2016). Difficulties in auditory organi-
sation as a cause of reading backwardness? An auditory neu-
roscience perspective. Developmental Science. Advance online
Veröffentlichung. doi:10.1111/desc.12457
Leong, V., Kalashnikova, M., Burnham, D., & Goswami, U. (2014,
September). Infant-directed speech enhances rhythmic structure
in the envelope. Paper presented at Interspeech, Singapur.
Leong, V., Stein, M., Turner, R., & Goswami, U. (2014). A role
for amplitude modulation phase relationships in speech rhythm
perception. Journal of the Acoustical Society of America, 136,
366–381.
Liu, H. M., Kuhl, P., & Tsao, F. M. (2003). An association between
mothers’ speech clarity and infants’ speech discrimination skills.
Developmental Science, 6(3), F1–F10.
Luo, H., & Kacke, D. (2007). Phase patterns of neuronal responses
reliably discriminate speech in human auditory cortex. Neuron,
54, 1001–1010.
Martin, A., Schatz, T., Versteegh, M., Miyazawa, K., Mazuka, R.,
Dupoux, E., & Cristia, A. (2015). Mothers speak less clearly to in-
fants: A comprehensive test of the hyperarticulation hypothesis.
Psychological Science, 26(3), 341–347.
McMurray, B., Kovack-Lesh, K. A., Goodwin, D., & McEchron, W.
(2013). Infant directed speech and the development of speech
perception: Enhancing development or an unintended conse-
quence? Cognition, 129(2), 362–378.
Molinaro, N., Lizarazu, M., Lallier, M., Bourguignon, M., &
Carreiras, M. (2016). Out-of-synchrony speech entrainment in
developmental dyslexia. Kartierung des menschlichen Gehirns. Advance online
Veröffentlichung. doi:10.1002/hbm.23206
Kacke, D. (2003). The analysis of speech in different temporal
lateralization as “asymmetric
integration windows: Zerebral
sampling in time.” Speech Communication, 41, 245–255.
Kacke, D. (2014). The neuroanatomic and neurophysiological in-
frastructure for speech and language. Current Opinion in Neuro-
biology, 28C, 142–149.
Power, A. J., Colling, L. C., Mead, N., Barnes, L., & Goswami, U.
(2016). Neural encoding of the speech envelope by children with
developmental dyslexia. Brain and Language, 160, 1–10. doi:10.
1016/j.bandl.2016.06.006
Power, A. J., Mead, N., Barnes, L., & Goswami, U. (2012). Neu-
ral entrainment to rhythmically-presented auditory, visual and
audio-visual speech in children. Grenzen in der Psychologie, 3, 216.
Power, A. J., Mead, N., Barnes, L., & Goswami, U. (2013). Neuronal
entrainment to rhythmic speech in children with developmental
dyslexia. Grenzen der menschlichen Neurowissenschaften, 7, 777.
Schack, B., & Weiss, S. (2005). Quantification of phase synchro-
nization phenomena and their importance for verbal memory
processes. Biological Cybernetics, 92, 275e287.
Song, J. Y., Demuth, K., & Morgan, J. (2010). Effects of the acoustic
properties of infant-directed speech on infant word recognition.
Journal of the Acoustical Society of America, 128(1), 389–400.
Tass, P., Rosenblum, M. G., Weule, J., Kurths, J., Pikovsky, A.,
Volkmann, J., Schnitzler, A., & Freund, H. J. (1998). Detection
of n:m phase locking from noisy data: Application to magne-
toencephalography. Physical Review Letters, 81, 3291.
Telkemeyer, S., Rossi, S., Koch, S. P., Nierhaus, T., Steinbrink, J.,
Kacke, D., . . . Wartenburger, ICH. (2009). Sensitivity of newborn
auditory cortex to the temporal structure of sounds. Zeitschrift für
Neurowissenschaften, 29, 14726–14733.
Telkemeyer, S., Rossi, S., Nierhaus, T., Steinbrink, J., Obrig, H.,
& Wartenburger, ICH. (2011). Acoustic processing of temporally-
modulated sounds in infants: Evidence from a combined NIRS
and EEG study. Grenzen in der Psychologie, 2, 62.
Thiessen, E. D., Hill, E. A., & Saffran, J. R. (2005). Infant-directed
speech facilitates word segmentation. Infancy, 7(1), 53–71.
Turner, R. E. (2010). Statistical models for natural sounds (Doctoral
dissertation). University College, London. Retrieved from http://
www.gatsby.ucl.ac.uk/~turner/Publications/Thesis.pdf
Weber, A., Hahne, A., Friedrich, M., & Friederici, A. D. (2004).
Discrimination of word stress in early infant perception: Electro-
physiological evidence. Kognitive Gehirnforschung, 18, 149–161.
Wechsler, D. (2008). WAIS IV administration and scoring manual.
San Antonio, TX: The Psychological Corporation.
Ziegler,
J. C., & Goswami, U. (2005). Reading acquisition, von-
velopmental dyslexia, and skilled reading across languages: A
psycholinguistic grain size theory. Psychological Bulletin,
131, 3–29.
OPEN MIND: Discoveries in Cognitive Science
90
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
e
D
u
Ö
P
M
ich
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
/
1
2
7
8
1
8
6
8
2
8
2
Ö
P
M
_
A
_
0
0
0
0
8
P
D
.
ich
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3