Der Fall der Vortäuschung: Beobachten von Aktionen
und Ableiten von Zielen
Ricarda I. Schubotz1 und D. Yves von Cramon2
Abstrakt
& Wenn wir eine Aktion beobachten, wir wissen es fast sofort
welches Ziel der Schauspieler verfolgt. Auffallend, das gilt auch für
Handlung vortäuschen (Pantomime), die relevante Informationen liefert-
Dabei geht es um die Manipulation selbst, nicht aber um das Manipulierte
Objekte. Die vorliegende fMRT-Studie befasste sich mit der Zielfrage
Schlussfolgerung aus vorgetäuschter Handlung im Vergleich zu tatsächlicher Handlung. Wir
fanden sowohl Unterschiede als auch Gemeinsamkeiten für das Gehirnkorre-
Methoden, aus beiden Handlungsarten auf Ziele zu schließen. Sie unterschieden sich
im Hinblick auf die Gewichte der zugrunde liegenden Handlungsbeobachtung
Netzwerk, Angabe der Ausnutzung von Objektinformationen in der
bei realen Handlungen und Manipulationsinformationen im Falle von
Vorwand. Jedoch, Zielschlussfolgerungen aus Manipulationsinformationen-
tion führte zu einem gemeinsamen Netzwerk für reale und vorgetäuschte
Aktion. Interessant, Dieses letztgenannte Netzwerk umfasste auch Bereiche, die
werden durch die Aktionsbeobachtung nicht identifiziert und das könnte daran liegen
zur Verarbeitung des Szeneninhalts und zur Evaluierung der Passung
mutmaßliche Handlungsziele. Diese Ergebnisse legen nahe, dass die Beobachtung von
Vortäuschung betont die Anforderung, das intern zu simulieren
beobachtete Handlung, schließen aber grundsätzliche Unterschiede in der Art und Weise aus, wie ob-
Server bewältigen reale und vorgetäuschte Aktionen. &
EINFÜHRUNG
Wenn wir Zeuge einer vorgetäuschten Handlung werden, das ist, Pantomime,
Wir haben sofort eine Vorstellung davon, welches Ziel verfolgt wird
vom Schauspieler. Wie wird dies erreicht?? Obwohl mehrere im-
Alterungsstudien haben die Leistung von Pfannen untersucht-
tomime (Hermsdörfer, Linde, Mühlau, Goldenberg,
& Wohlschläger, 2007; Imazu, Sugio, Tanaka, & Hassen,
2007; Kro´liczak, Cavina-Pratesi, Guter Mann, & Culham,
2007; Buxbaum, Kyle, & Menon, 2005; Ohgami, Matsuo,
Uchida, & Nakai, 2004; Moll et al., 2000) und der Beobachter-
vation echten Handelns (z.B., Newman-Norlund, von Schie,
van Zuijlen, & Bekkering, 2007; Calvo-Merino, Gre`zes,
Glaser, Passingham, & Haggard, 2006; Costantini et al.,
2005; Schubotz & von Cramon, 2004; Johnson-Frey et al.,
2003; Manthey, Schubotz, & von Cramon, 2003; Wellhornschnecke
et al., 2001; für einen aktuellen Rückblick, vgl. Vogt & Thomaschke,
2007), Wir wissen immer noch nichts über die neuronalen Korrelate
des Verstehens von Zielen in der beobachteten Pantomime. Der
Studieren Sie nur die Umsetzung der Beobachtung vorgetäuschter Handlungen
(Deutsch, Niehaus, Roarty, Giesbrecht, & Müller, 2004)
eine verdeckte Anweisung verwendet, das ist, Teilnehmer waren
nicht darauf ausgerichtet, die Absicht oder das Ziel der Akteure zu berücksichtigen
sondern vielmehr auf das Auftreten einer Bildschirmunterbrechung
die Aktion.
In der vorliegenden Studie, Wir haben die Hypothese getestet, dass
Das Gehirn korreliert mit dem Ableiten von Zielen aus Vorgabe und
1Max-Planck-Institut für neurologische Forschung, Köln, Ger-
viele, 2Max-Planck-Institut für menschliche Kognition und Gehirn
Wissenschaften, Leipzig, Deutschland
reale Action sind teilweise vergleichbar, wie in mehr angegeben
Einzelheiten dazu im Folgenden. Anscheinend gibt es gute Gründe-
Söhne, die sowohl Unterschiede als auch Gemeinsamkeiten annehmen
Das Gehirn korreliert mit dem Ableiten von Zielen aus Vorgabe und
echte Aktion, jeweils.
Einerseits, obwohl wir es unterscheiden können
real aus vorgetäuschter Aktion, es ist nicht plausibel anzunehmen
dass neuronale Prozesse der Interpretation dienen
vorgetäuschte Handlungen sollten sich grundsätzlich von unterscheiden
diejenigen, die der Interpretation realer Handlungen dienen. Das
liegt nicht nur daran, dass beide eine Vielzahl von Wahrnehmungen mit sich bringen-
tual, Eselsbrücke, und kognitive Prozesse, die trigonometrischer Natur sind-
wird durch einen komplexen und gesellschaftlich relevanten Reiz ausgelöst, Aber
Genauer gesagt, weil beides von entscheidender Bedeutung ist, unter
natürliche Bedingungen, die intensive Analyse der Hände
Haltung und Bewegungen, das ist, Manipulationsinfor-
mation. Modelle aus der Motorsteuerungstheorie wurden
wird kürzlich verwendet, um zu beschreiben, wie wir Beobachtetes analysieren
Aktionen, um auf die Ziele unserer Artgenossen zu schließen (Grush, 2004;
Miall, 2003; Wolpert & Flanagan, 2001). Diese Modelle,
Entwickelt, um zu beschreiben, wie wir unsere kontinuierlich anpassen
Bewegungen an sich ändernde Umweltbedingungen und
Online-Fehlerkorrektur, Geben Sie an, dass mehrere nach vorne gerichtet sind
Modelle werden eingerichtet, um bevorstehende Ereignisse vorherzusagen
sich entfaltende Handlung, egal ob von uns durchgeführt-
selbst oder nur beobachtet. Entsprechend, die Prozesse
zugrunde liegendes Ziel
Die rechnerische Schlussfolgerung beträgt
das Ausführen einer Simulation mehrerer Aktionsskripte in
parallel, bis das am besten passende Drehbuch gewinnt. Zum Beispiel,
Beobachten eines Schauspielers, der eine Tasse ergreift, wir sagen es voraus
entweder führe es zu seinem Mund, oder reinigen, oder verschieben Sie es weiter
D 2008 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 21:4, S. 642–653
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
F
.
T
.
/
.
Ö
N
1
8
M
A
j
2
0
2
1
ein Regal, oder geben Sie es an jemand anderen weiter, und so weiter. Testen
Hypothesen über aktuell gültige Zieloptionen, egal
ob auf realen oder vorgetäuschten Handlungen basiert, so ruft
zur sensomotorischen Transformation (zur internen Simulation
von Action-Skripten), Arbeitsgedächtnis (für die Auswahl von
aktuell gültige Skripte), und interne Belohnungsbewertung
(für die motivierende Führung der laufenden Schätzung
der Skriptanpassung und schrittweise Reduzierung der aktuellen
getestete Zieloptionen).
Andererseits, obwohl die Analyse beobachtet
Eine Aktion, egal ob real oder vorgetäuscht, erfordert Tests
Hypothesen über aktuell gültige Ziele, diese Hypothe-
ses stammen aus zumindest teilweise unterschiedlichen Quellen
für echte und vorgetäuschte Action. B. Objektinformationen, können-
nicht ausgenutzt werden, um aus einer vorgetäuschten Handlung auf das Ziel zu schließen,
Man würde erwarten, dass Komponenten der Aktion Beobachter sind-
vationsnetzwerk, das sich mit der Analyse von Hand beschäftigt
Körperhaltungen und Bewegungen, das ist, Manipulation, sollte sein
Besonders verbessert in der vorgetäuschten Aktion. Umgekehrt,
diejenigen, die mit der Verarbeitung von Objektinformationen befasst sind
sollte für echte Aktionen aktiver sein. Auch wenn iso-
verspätet, beide Informationsquellen, Manipulationen und
Objekte, sind dafür bekannt, hervorragende Handlungshinweise zu geben
Ziele, wie das frühe Auftauchen von Pre zeigt-
tendenzielles oder symbolisches Spiel in der kindlichen Entwicklung zum einen
Hand (Fein, 1981) und experimentell
Untersuchungen von
Objekt-Affordanz andererseits (Helbig, Graf, &
Kiefer, 2006).
Die vorliegende fMRT-Studie befasste sich mit der Zielfrage
Schlussfolgerung aus vorgetäuschten Handlungen im Vergleich zu realen
Aktion. Zu diesem Zweck, Dazu haben wir kurze Videoclips präsentiert
zeigte entweder vorgetäuschte oder reale Handlungen (zweistufiger Faktor,
Typ). Wir erwarteten, dass es sowohl vorgetäuschte als auch echte Action geben würde-
messen Sie das Aktionsbeobachtungsnetzwerk, in dem berichtet wird
Literatur, umfassend, unter anderen, der ventrale Prä-
motorischer Kortex (PMv), der vordere intraparietale Sulcus
(aIPS), und der hintere obere Schläfensulcus (pSTS)
(Rizzolatti & Craighero, 2004). Jedoch, aufgrund der
unterschiedliche Bedeutung von Manipulation und Objektinformation-
tion in der Analyse von vorgetäuschter und realer Handlung, bzw-
aktiv, Wir erwarteten höhere Signale im Extrastriate-Körper
Bereich (EBA; Taylor, Wiggett, & Downing, 2007), das Hu-
Mann bewegungsselektiver Bereich (hMT; Greenlee, 2000; vgl. Auch
Peuskens, Vanrie, Verfaillie, & Orban, 2005), und das pSTS
(Chip & Perrett, 2003) für vorgetäuscht im Vergleich zur Realität
Aktion. Umgekehrt, der laterale Hinterhauptskomplex (LOC;
Grill-Spector, Kourtzi, & Kanwisher, 2001) wurde erwartet
im Vergleich zur vorgetäuschten Handlung erhöht werden.
Obwohl dieses Aktionsbeobachtungsnetzwerk erwartet wurde
für vorgetäuschte und reale Handlungen unterschiedlich gewichtet werden,
Das Netzwerk spiegelt vielfältige Wahrnehmungen wider, Eselsbrücke, Und
kognitive Prozesse, die nicht alle unbedingt dazu beitragen
zum Zielschluss. Zum Beispiel, die aufmerksame Analyse von
Es wird erwartet, dass die beobachtete Aktion auch danach anhält
Das Ziel wurde erfolgreich anerkannt. daher, In
ein Versuch, ausschließlich Zielschlussprozesse anzuzapfen, Wir
ein Schaltprotokoll implementiert. Die Begründung dafür
Das Protokoll bezog sich auf die sogenannte Wiederholungsabschwächung-
tions- oder Unterdrückungseffekt. Wiederholungsunterdrückung bezieht sich
darauf, dass die Wiederholung eines Reizes zu einem De führt-
Gefaltetes Fettgedrucktes Signal in Bereichen, die diesen Reiz kodieren
(Hamilton & Grafton, 2006; Grill-Spector & Malach, 2001;
Naccache & Dehaene, 2001; Thompson-Schill, D'Esposito,
& Kann, 1999). Hier haben wir Versuche mit neuen Informationen verglichen-
mation (Schalterversuche, Jenseits) mit denen, die Nr. enthalten
neue Informationen (Wiederholungsversuche, Jenseits), relativ zu
der vorangegangene Prozess. Bereiche, die an der Verarbeitung beteiligt sind
Eine bestimmte Art von Informationen sollte ansprechender sein
bei Wechselversuchen als bei Wiederholungsversuchen.
Wir verwendeten drei Arten von Versuchen (der dreistufige Faktor,
Schalten): (A) „Zielwechsel“-Versuche (G) darin, dass sowohl die
Manipulationen und die Gegenstände der dargestellten Handlung
im aktuellen Versuch unterschied sich n von denen im vorangegangenen-
ing probe n (cid:1) 1; (B) „Objektwechsel“-Versuche (Ö) nur darin
Der Gegenstand der vorgelegten Klage in Versuch n unterschied sich von
das im vorangegangenen Versuch verwendete Objekt n (cid:1) 1 (während die
In beiden Fällen kam es zu wiederholten Manipulationen); Und (C)
„Manipulationsschalter“-Versuche (M) darin nur die Manipulation-
Die Ergebnisse der vorgestellten Aktion im Versuch n unterschieden sich von
die Manipulation im vorangegangenen Prozess n (cid:1) 1 (während die
Die verwendeten Gegenstände wurden in beiden Fällen wiederholt). Durch con-
Vergleich von Zielwechselversuchen mit Objektwechselversuchen (G >
Ö), Unser Ziel war es, Gehirnbereiche zu identifizieren, die dazu beitragen
Schlussfolgerung auf der Grundlage von Manipulationsinformationen-
zum Ziel
mation. Beachten Sie, dass G-Studien beides neu lieferten
Manipulation und neue Objektinformationen, die Kontraste
G > O (and G > M, siehe unten) hat keinen Verwandten identifiziert
Unterschied zwischen zwei verschiedenen Arten von Informationen
sondern eher der relative Unterschied zwischen neu und alt
(=wiederholt) Informationen der gleichen Art.
Beachten Sie, dass wir diesen Kontrast als Hervorhebungsziel betrachten
Inferenzprozesse gemäß der experimentellen Op-
Erationalisierung der Zielinferenz, die wir in der wählen
vorliegende Studie. Insbesondere, Wir haben den Ansatz gewählt
Die Zielinferenz besteht aus einer Reihe unterschiedlicher kognitiver Fähigkeiten
Teilprozesse, nicht zu einem Moment des einheitlichen Aha-Erlebnisses-
enz. Daher, zur Ableitung von Zielen auf der Grundlage von Manipu-
lationsinformationen, Egal ob von echt oder von
Vortäuschen von Handlungen, wir haben PMv erwartet, aIPS, und angrenzend
supramarginaler Gyrus (SMG) als relevante Bereiche für sen-
sorimotorische Transformation (Rizzolatti & Luppino, 2001),
lateraler präfrontaler Kortex (lPFC) Untergebener Arbeitsmem-
Theorie im adaptiven zielgerichteten Verhalten (Watanabe, 2007;
Petride, 2005), und OFC als ein bekanntermaßen engagierter Bereich
bei der Belohnungsbewertung (Wallis, 2007).
Manipulationsschalter (M) Versuche wurden durchgeführt, um
Gleichgewicht der Wahrscheinlichkeit, dass neuartige Informationen pro sind-
entweder durch Objekte oder durch Manipulationen erzeugt. Objekt drin-
Formation, im Gegensatz zu Manipulationsinformationen, War
voraussichtlich nur in realen Aktionen und weitgehend ausgenutzt werden
wird bei vorgetäuschten Aktionen ignoriert. daher, wir erwartet
the contrast G > M that reflected goal inference on
Die Grundlage der Objektinformationen ergibt keinen gemeinsamen Wechselstrom-
Motivationen für reale und vorgetäuschte Handlungen.
Schubotz and von Cramon
643
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
.
F
.
/
.
T
Ö
N
1
8
M
A
j
2
0
2
1
METHODEN
Teilnehmer
Achtzehn Rechtshänder, gesunde Freiwillige (acht Frauen;
Altersspanne = 21–32 Jahre; Durchschnittsalter = 26.4 Jahre) Par-
an der Studie beteiligt. Nachdem ich über Poten informiert wurde-
tielle Risiken untersucht und von einem Arzt der Einrichtung untersucht,
Die Probanden gaben vor der Teilnahme eine Einverständniserklärung ab.
Die experimentellen Standards wurden von der örtlichen Behörde genehmigt
Ethikkommission der Universität Leipzig. Daten waren
anonym behandelt.
Anregungen und Aufgaben
Den Probanden wurden Filme präsentiert, die Aktionen zeigten
und mit kurzen verbalen Handlungsbeschreibungen, die sich darauf beziehen
diese Aktionen. Jeder Versuch (6 Sek) begann mit einem Film
(2 Sek) Es folgt eine Fixierungsphase. Die Länge der
Fixierungsphase (2.5–4 Sek) hing von der Variablen jit ab-
ter mal (0, 500, 1000, oder 1500 ms) die eingefügt wurden
vor dem Film, um die zeitliche Auflösung zu verbessern
die BOLD-Antwort. Aktionen wurden entweder ausgeführt
entsprechende Gegenstände (z.B., Wasser aus einer Flasche gießen
in ein Glas) oder auf ungeeigneten Gegenständen (z.B., Herstellung
die gleichen Bewegungen mit einem Mülleimer und einem Schlüssel). Diese beiden
Die einzelnen Klageklassen werden im Folgenden als „real“ bezeichnet
„Handlungen“ und „Handlungen vortäuschen“.,'' jeweils. Beachten Sie, dass
um reichhaltige Informationsinhalte aus beiden Ma zu generieren-
Manipulationen und Objekte, jeden von uns präsentierten Filmclip
zeigte eine Kette spezifischer Manipulationen (z.B., begreifen,
drehen, und Eröffnung) und Kombinationen von zwei Objekten
(z.B., eine Tasse und ein Löffel).
Die Probanden wurden angewiesen, sich um die Präsentation zu kümmern
Filme. Sie wurden darüber informiert, dass einige der Filme
Es folgte ein Prozess, der mit einer Klage begann-
Beschreibung, die entweder mit der Beschreibung übereinstimmte oder nicht-
Zelt des vorherigen Films. Es wurde betont, dass es
Es spielte keine Rolle, ob Aktionen zu dieser Aktion durchgeführt wurden
Bei den Beschreibungen, auf die Bezug genommen wurde, handelte es sich um reale oder vorgetäuschte Handlungen.
Für den Fall, dass ein Versuch mit einer Handlungsbeschreibung vorlag-
gesendet, Die Probanden gaben ihre Antworten sofort ab
mit dem Zeigefinger auf ein Antwortfeld mit zwei Tasten tippen
für positive Antworten und ihren Mittelfinger für re-
Einwürfe. Fünfzig Prozent der Handlungsbeschreibungen lauteten darauf
bekräftigt werden und 50% abzulehnen.
Zusätzlich zum zweistufigen Reizfaktor Typ [real
Aktion (R) und vorgeben, etwas zu tun (P)], ein dreistufiger Faktor
Schalten [Ziel (G), Objekte (Ö), und Manipulationen (M)]
wurde umgesetzt. Die Probenachfolge wurde umgesetzt
so dass es sich bei den Versuchen entweder um Wechselversuche oder um Wiederholungsversuche handelte
in Bezug auf die Manipulationen, zu den physischen Objekten
im Einsatz, oder beides (siehe Abbildung 1). Alle Kombinationen davon
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
F
T
.
.
.
/
Ö
N
1
8
M
A
j
2
0
2
1
Figur 1. Experimentelles Design. Beispiele für Ziele (z.B., „Schreiben mit Stift“) sind in farbigen Kästchen angegeben, in denen Fotos auf das Physische hinweisen
Objekt, das tatsächlich im entsprechenden Movieclip dargestellt wird. Stufen des experimentellen Faktors Switch (Objekt, Manipulation, und Ziel) entsprechen
zu Spalten, mit den Spalten 1–3 (solide Rahmen) und 4–6 (gestrichelte Rahmen) entsprechend den beiden Ebenen des experimentellen Faktortyps (echt und
vorgeben). Die erste Kästchenreihe repräsentiert das Ziel in einem Versuch n (cid:1) 1, wohingegen die restlichen Zeilen Beispiele für Aktionen darstellen, im Prozess n, entweder
wiederholen (zweite Reihe) oder wechseln (dritte Reihe) die manipulierten Objekte (erste und vierte Spalte), Manipulationen (zweite und fünfte Spalte), oder Ziele
(dritte und sechste Spalte). Die Felder für die Zielwiederholung sind leer, da dieser Versuchstyp nicht Teil des Versuchsdesigns war.
644
Zeitschrift für kognitive Neurowissenschaften
Volumen 21, Nummer 4
Bis auf eine identische Wiederholung waren zwei Faktoren möglich
des Prozesses n (cid:1) 1. Darüber hinaus, die Übergangsfrequenzen von
echtes und vorgetäuschtes Handeln wurden ausgeglichen. Zwanzig-
fünf Prozent der Filme (d.h., 21 von 84 echte Handlungen und
21 von 84 Vortäuschen von Handlungen) Es folgte eine Aktion de-
Skript, das die Länge eines regulären Prozesses hatte (2 trocken von-
Skript, inklusive Reaktionsphase, Plus 4 Sekundenfixierung
Phase), ergebend 42 zusätzliche Versuche. Jede Aktion de-
Auf die Anmeldung folgte ein regulärer Scheinversuch
Film mit entweder einer realen oder einer vorgetäuschten Handlung, aber keinem
weder ein regulärer Wechsel noch ein Wiederholungsversuch. Entsprechend, diese
Scheinversuche (n = 42) trat in den Analysekontrast ein-
reale und vorgetäuschte Handlungen ausführen (summiert sich zu 84 + 21 =
105 Versuche für echte Handlungen und 84 + 21 = 105 Versuche für
Vortäuschen von Handlungen) nicht jedoch die Analysen zum Wechseln oder Wiederholen-
tionseffekte von Manipulationen und Objekten. Endlich, 20 leer
Versuche (Ruhezustand) wurden gemischt mit dem präsentiert
experimentelle Versuche.
Insgesamt, 272 Versuche wurden vorgestellt: 84 echte Taten
Plus 21 echte Action-Dummies, 84 Vortäuschen von Aktionen plus
21 Stellen Sie sich Actionpuppen vor, 21 Aktionsbeschreibungen fol-
Lowing reale Aktionen, 21 Aktionsbeschreibungen folgen vor-
neigende Handlungen, Und 20 leere Versuche.
MRT-Datenerfassung
Die Bildgebung wurde auf einem 3-T Bruker durchgeführt (Ettlingen,
Deutschland) Medspec 30/100 System ausgestattet mit
Standard-Birdcage-Kopfspule. Die Teilnehmer wurden platziert
auf dem Scannerbett in Rückenlage mit der rechten Seite
Zeige- und Mittelfinger liegen auf der rechten Seite
Antwortschaltflächen einer Antwortbox. Formschlüssiges Kissen-
Ionen wurden verwendet, um Kopf zu verhindern, Arm, und Hand
Bewegungen. Dafür wurden den Teilnehmern Ohrstöpsel zur Verfügung gestellt
Scannergeräusche würden gedämpft. Zweiundzwanzig axial
Scheiben (192 mm Sichtfeld; 64 (cid:2) 64 Pixelmatrix; 4 mm
Dicke; 1 mm Abstand; Auflösung in der Ebene von 3 (cid:2)
3 mm) parallel zur bikommissuralen Linie (AC–PC) Abdeckung
Das gesamte Gehirn wurde mit einem Single-Shot-Gradi erfasst-
ent EPI-Sequenz (2000 ms Wiederholungszeit; 30 ms
Echozeit; 908 Flipwinkel; 100 kHz-Erfassungsbandbreite)
empfindlich gegenüber BOLD-Kontrast. Vor dem Funktionsbild-
ing, 22 anatomische T1-gewichtete MDEFT-Bilder (Norris,
2000; Ugurbil et al., 1993) Und 22 T1-gewichteter EPI
Bilder mit der gleichen räumlichen Ausrichtung wie die Funktion-
Es wurden zusätzliche Daten erfasst. In einer separaten Sitzung, hoch-
Von jedem wurden hochauflösende Bilder des gesamten Gehirns aufgenommen
Vorbehaltlich einer Verbesserung der Lokalisierung von Aktivierungsherden
unter Verwendung einer T1-gewichteten 3D-segmentierten MDEFT-Sequenz
das gesamte Gehirn abdeckt.
MRT-Datenanalyse
Die Datenverarbeitung erfolgte mit dem Softwarepaket LIPSIA
(Lohmann et al., 2001). Funktionsdaten standen an erster Stelle
bewegungskorrigiert unter Verwendung einer passenden Metrik basierend auf Lin-
Ohrkorrelation. Der zeitliche Versatz soll korrigiert werden-
Zwischen den in einem Bild erfassten Schichten wechseln, ein kubischer Spline
Es wurde Interpolation eingesetzt. Niederfrequenzsignal
Änderungen und Abweichungen von der Grundlinie wurden mit einem Tem entfernt-
Poraler Hochpassfilter mit einer Grenzfrequenz von 1/85 Hz.
Die räumliche Glättung wurde mit einem Gauß-Filter durchgeführt
5.65 mm FWHM. Zum Ausrichten der Funktionsdaten-Slices
ein dreidimensionales stereotaktisches Koordinatenreferenzsystem, eine starre Linie-
Ohrregistrierung mit sechs Freiheitsgraden (drei Rota-
tional, drei translatorisch) durchgeführt wurde. Die Rotation
und die Translationsparameter wurden am erfasst
Basis der MDEFT- und EPI-T1-Slices, um eine zu erreichen
optimale Übereinstimmung zwischen diesen Scheiben und dem Individuum
3-D-Referenzdatensatz. Der MDEFT-Volumendatensatz mit
160 Scheiben und eine Scheibendicke von 1 mm wurden standardisiert
der stereotaktische Raum von Talairach (Talairach & Tournoux,
1988). Die Rotations- und Translationsparameter
wurden anschließend durch lineare Skalierung in einen Stan transformiert-
Dard-Größe. Die resultierenden Parameter wurden dann verwendet
Transformieren Sie die Funktionsschnitte mithilfe der trilinearen Interpolation,
so dass die resultierenden Funktionsschnitte ausgerichtet waren
das stereotaktische Koordinatensystem, und erzeugt so Output
Daten mit einer räumlichen Auflösung von 3 (cid:2) 3 (cid:2) 3 mm (27 mm3).
Die statistische Auswertung basierte auf der Methode der kleinsten Quadrate
Schätzung mit dem Allgemeinen
lineares Modell für seriell
autokorrelierte Beobachtungen (Friston et al., 1995; Worsley
& Friston, 1995). Die Designmatrix wurde mit erstellt
eine Güterwagenfunktion, mit der Hämodynamik verwickelt
Antwortfunktion und ihre erste Ableitung. Gehirnaktivierungen
wurden zeitbezogen zum Beginn der Filme analysiert, Und
die analysierte Epoche umfasste die gesamte Dauer (2 Sek)
der präsentierten Filme. Die Modellgleichung, inkl-
ing der Beobachtungsdaten, die Designmatrix, und der Fehler
Begriff, wurde mit einem Gaußschen Dispersionskern gefaltet
von 4 sec FWHM, um die zeitliche Autokorrela zu berücksichtigen-
tion (Worsley & Friston, 1995). Im Folgenden, Kontrast
Bilder, das ist, Beta-Wert-Schätzungen des Raw-Score-Dif-
Bezüge zwischen bestimmten Bedingungen, wurden generiert
für jeden Teilnehmer. Wie alle einzelnen Funktionsdatensätze
wurden auf den gleichen stereotaktischen Referenzraum ausgerichtet,
Die Einzelsubjekt-Kontrastbilder wurden in ein eingegeben
Zufallseffektanalyse der zweiten Ebene für jeden der Nachteile-
Bünde. Für die Gruppe wurden T-Tests mit einer Stichprobe durchgeführt
analysiert die Kontrastbilder aller Probanden
gab an, ob Unterschiede zwischen den Bedingungen beobachtet wurden-
Die Werte unterschieden sich deutlich von Null. Die t-Werte
wurden anschließend in Z-Scores umgewandelt. Korrigieren
für falsch-positive Ergebnisse, in einem ersten Schritt, ein anfänglicher voxelweiser
Z-Schwelle wurde auf Z = gesetzt 2.33 ( p = .01, unkorrigiert). In
ein zweiter Schritt, Die Ergebnisse wurden mehrfach korrigiert
Vergleiche unter Verwendung von Clustergröße und Clusterwert-Thresh-
Olds, die durch Monte-Carlo-Simulationen mit einer Signifikanz erhalten wurden
Niveau von p = .005, das ist, Die gemeldeten Aktivierungen sind
deutlich aktiviert bei p < .005, corrected for multiple
comparisons at the cluster level.
To investigate more thoroughly the comparability of
brain responses in the areas identified by contrasts, per-
centage signal change analyses of the BOLD response
Schubotz and von Cramon
645
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
4
e
-
6
p
4
d
2
f
1
/
9
2
3
1
7
/
5
4
0
/
2
6
o
4
c
2
n
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
4
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
t
/
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
were carried out where the mean signal change over a
6-sec epoch, starting 4 sec after movie onset, were ex-
tracted from selected voxels within significantly acti-
vated brain areas for the experimental and the resting
baseline conditions. The mean signal change of a voxel
for each condition was calculated in relation to the mean
signal intensity of that voxel across all time steps.
mentary motor area (pre-SMA), and the left superior
intraparietal sulcus. Extensive activation was also found
in the fusiform gyrus (FG) and in the occipital gyri in-
cluding probably the LOC as well as an area we will
hereafter refer to as EBA/hMT, as hMT overlaps closely
with EBA (Downing, Wiggett, & Peelen, 2007; Figure 2,
Table 1).
RESULTS
Behavioral Results
Performance was assessed by error rates and reaction
times. Repeated measures ANOVAs were performed for
each of these measures with the two-level factor Type
(pretend and real) and the three-level factor Switch
(goal, objects, and manipulations). Regarding reaction
times, a main effect for the factor Type [F(1,17) =
13.791, p < .005] and an interaction Type (cid:2) Switch
[F(2,34) = 4.344, p < .05] was found. These effects re-
flected that responses to the action descriptions were
slower for pretend (mean ± standard error, 421 ±
32 msec) as compared with real actions (393 ± 26 msec).
The t tests showed that in case of pretend actions,
responses to trials in that only objects switched were
faster (386 ± 26 msec) than when only manipulations
were switched (446 ± 32 msec; t18 = 5.059, p < .001)
and were marginally faster than when both manipula-
tions and objects were switched (431 ± 39 msec; t18 =
1.858, p = .081). For error rates, a significant main effect
was found for the factor Switch [F(2,34) = 5.023, p < .01]
and for the factor Type [F(1,17) = 9.305, p < .05]. In par-
ticular, subjects made more errors when matching action
descriptions with pretend actions (7.4 ± 3.3%) than with
real actions (1.1 ± 0.8%). The t tests showed that action
descriptions for trials in that only objects switched were
easier (1.4 ± 1.1% errors) than for trials in that only
manipulations (5.3 ± 2.2%, t18 = 3.487, p < .005) or both
manipulations and objects (6.0 ± 2.8%, t18 = 2.955, p <
.01) switched. Overall, behavioral performance implicated
that inferring goals was slightly more demanding from
pretend than from real actions, and that the inspection of
manipulation information was more demanding than the
inspection of object information. However, as all effects
remained below differences of 60 msec and about 6%
errors, we did not expect them to account for activation
differences in our BOLD contrasts.
fMRI Results
Observing Real and Pretend Action
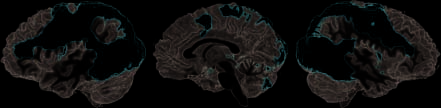
The network commonly activated by either the obser-
vation of real action and that of pretend action [conjunc-
tion (real > ausruhen) \ (pretend > rest)] umfasste die
bilaterales PMv [Brocas Gegend (BA) 6/44], die linke untere
Stirnfurche (IFS; BA 9/46), das aIPS, die linke Maschinenpistole, Die
Linker dorsaler prämotorischer Kortex (PMd), das linke Vorsupplement-
Beobachten von vorgetäuschter versus realer Handlung, und umgekehrt
Unter diesen Bereichen, die Beobachtung vorgetäuschter Handlungen
erbrachte deutlich mehr Aktivierung als echte Aktion
((pretend > real) \ ((real > rest) \ (pretend > rest)))
im linken PMv, das linke aIPS erstreckt sich bis ins SMG, die linke
IFS (BA 9/46), das linke pSTS, und das richtige EBA/hMT
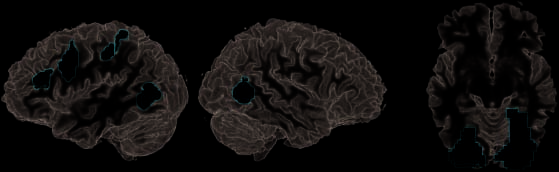
(Abbildung 3A, Tisch 2). Im Gegensatz, die Beobachtung des Realen
Handeln führte zu deutlich mehr Aktivierung als Vortäuschen
Aktion ((real > pretend) \ ((real > rest) \ (pretend >
ausruhen))) im FG und/oder LOC bilateral (im Folgenden LOC),
bestehend aus vorderen und hinteren Fächern im
rechter oberer Parietallappen (BA 7) und im rechten
postzentraler Gyrus (Abbildung 3B, Tisch 2).
Ableiten von Zielen
Auf dieser Grundlage können aus beobachtetem Handeln Ziele abgeleitet werden
von Manipulation und Objektinformationen. Zu identifizieren
Gehirnbereiche, die an der Zielinduktion beteiligt sind, auf der Grundlage von
Manipulationen, Wir haben die Auswirkung der Bereitstellung von Sub analysiert-
Objekte mit neuen Manipulationsinformationen (Manipulation
switch effect), and subsequently to identify brain areas
involved in goal inference on the basis of object infor-
mation, we analyzed the effect of providing subjects with
new object information (object switch effect). Beachten Sie, dass
in a direct comparison between the M and the O trials,
the effects of manipulation switches and object repeti-
tion would have been inextricably confounded. Ac-
cordingly, manipulation switch effects were tested
while controlling for object switches by contrasting G
with O trials and object switch effects by contrasting G
with M trials.
Figur 2. The network conjointly activated by the observation of
real and pretend actions (as compared with rest) umfasste die
areas that are typically seen for action observation, including PMv
extending from BA 6 into BA 44, anterior parietal regions (aIPS and
SMG) as well as the pSTS. For further abbreviations, see Results section.
646
Zeitschrift für kognitive Neurowissenschaften
Volumen 21, Nummer 4
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
.
.
/
F
T
.
Ö
N
1
8
M
A
j
2
0
2
1
Tisch 1. Gemeinsames Aktionsbeobachtungsnetzwerk für Pretend
und echte Action: Konjunktion der Beobachtung realer Handlungen
im Vergleich zu Ruhe und Beobachtung vorgetäuschter Handlungen
im Vergleich zur Ruhe
Bereich
X
j
z
Z
Konjunktion von realer Aktion versus Ruhe und vorgetäuschter Aktion
versus Ruhe
PMv
PMd
vor SMA
IFS (BA 9/46)
Vorderes IPS
Überlegenes IPS
SMG (BA 40)
EBA/hMT/pSTS
FG
LOC
37
(cid:1)47
(cid:1)26
(cid:1)5
(cid:1)38
(cid:1)35
31
(cid:1)29
(cid:1)59
46
(cid:1)41
43
(cid:1)35
31
6
6
(cid:1)8
3
21
(cid:1)35
(cid:1)35
(cid:1)71
(cid:1)23
(cid:1)54
(cid:1)63
(cid:1)45
(cid:1)84
(cid:1)81
30
33
51
51
24
42
45
27
34
3
(cid:1)3
(cid:1)6
3
3
5.25
5.9
5.16
4.14
5.05
5.862
5.178
5.984
6.127
6.789
6.93
6.536
5.98
6.622
Anatomische Spezifikation, Talairach-Koordinaten, maximaler Z-Wert
(Das Volumen ist nicht angegeben, da alle Aktivierungen lokale Maxima eines gemeinsamen Wertes waren
Aktivierung).
Abkürzungen: PMv = ventraler prämotorischer Kortex; PMd = dorsaler Prämotor
Kortex; Prä-SMA = präsupplementärer motorischer Bereich; IFS = inferior frontal
Sulkus; IPS = intraparietaler Sulcus; SMG = supramarginaler Gyrus; EBA/
hMT = extrastriatischer Körperbereich/menschlicher bewegungsselektiver Bereich; pSTS =
Sulcus temporalis posterior superior; FG = Gyrus fusiformis; LOC = seitlich
Hinterhauptskomplex.
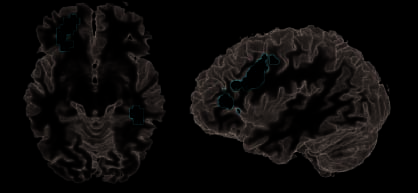
Ableitung von Zielen durch Analyse neuer Manipulationen
Zur Beobachtung von Handlungen in beiden Manipula-
tionen und vertauschte Objekte im Vergleich zu denen in
dass nur Objekte geschaltet wurden (G > O, Manipulationsschalter
Wirkung), Aktivierungen befanden sich im linken zentralen OFC
Tisch 2. Unterschiedliche Gewichtungen der Aktionsbeobachtung
Netzwerk: Direkte Kontraste zwischen der Beobachtung des Realen
und vorgetäuschte Aktion ((Pretend > Real) \ ((Real > Rest) \
(Pretend > Rest))) Und ((Real > Pretend) \ ((Real >
Ausruhen) \ (Pretend > Rest)))
Bereich
X
j
z
Z
mm3
Beobachtung von vorgetäuschter versus realer Handlung
PMv
IFS (BA 9/46)
aIPS
SMG
(cid:1)45
(cid:1)42
3
33 4.35
3672
30
15 3.50
1296
(cid:1)42 (cid:1)39
54 2.99
2025
(cid:1)53 (cid:1)23
36 2.95
l.m.
EBA/hMT/pSTS
(cid:1)54 (cid:1)54
12 3.32
3078
EBA/hMT
48 (cid:1)57
3 3.80
1728
Beobachtung realer und vorgetäuschter Handlungen
LOC
(cid:1)27 (cid:1)66 (cid:1)9 4.12
9504
(cid:1)29 (cid:1)92
3 4.05
l.m.
21 (cid:1)69 (cid:1)6 4.82 18738
22 (cid:1)83 (cid:1)3 4.63
l.m.
Oberer Parietallappen (SPL)
21 (cid:1)48
66 3.77
2322
Postzentraler Gyrus (GEBEN)
63 (cid:1)12
27 4.32
3294
l.m. = lokales Maximum.
(BA 11/10), linkes vorderes IFS (BA 9/46), links PMv (BA 6),
erstreckt sich leicht in den operkulären Teil des Unterleibs
frontaler Gyrus (BA 44), und in einem rechten inferioren temporalen Re-
Gion um den Kollateralsulcus, spiegelt wahrscheinlich das wider
parahippocampaler Ortsbereich (PPA; Epstein & Kanwisher,
1998) (Figur 4, Tisch 3). Eine Signaländerungsanalyse in der
Die identifizierten Bereiche bestätigten diesen Manipulationsschalter
Die Effekte unterschieden sich statistisch gesehen nicht zwischen real und vorher-
neigende Handlungen. Einen Haupteffekt gab es für den Faktor Typ
(echte Action und vorgetäuschte Action) aufgrund höherer Signaleingänge
vorgeben im Vergleich zu realen Handlungen in PMv [F(2,34) =
29.332, P < .000], BA 44 [F(2,34) = 6.113, p < .05], and
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
4
e
-
6
p
4
d
2
f
1
/
9
2
3
1
7
/
5
4
0
/
2
6
o
4
c
2
n
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
4
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
.
f
.
t
.
Figure 3. Direct contrasts
between observation of real
and pretend actions revealed
different weights of the action
observation network. (A) Areas
elevated by the observation
of pretend as compared with
real action included left IFS
(BA 9/46), left PMv, left aIPS,
and EBA/hMT extending into
pSTS in the left hemisphere.
(B) Conversely, areas more
engaged in processing real
as compared with pretend
actions were primarily found
in LOC bilaterally.
o
n
1
8
M
a
y
2
0
2
1
Schubotz and von Cramon
647
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
4
e
-
6
p
4
d
2
f
1
/
9
2
3
1
7
/
5
4
0
/
2
6
o
4
c
2
n
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
4
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
/
.
t
f
.
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Manipulation switch. Contrast between trials showing new versus repeated manipulations (G > Ö) und entsprechende Signaländerungen
In (1) OFC, (2) PPA, (3) anterior, (4) operkulärer Sulcus frontalis inferior (BA 44), Und (5) PMv. Diese Bereiche wurden erhöht, wenn Ziele erreicht werden sollten
aufgrund geänderter Manipulationsinformationen abgeleitet werden. Balkendiagramme zeigen prozentuale Signaländerungen aufgrund neuer und alter Objekte an
(Blau), Manipulationen (Gelb), oder beides (d.h., Ziele; Rot). Weiße Balken zeigen das Signal im Ruhezustand.
IFS [F(2,34) = 10,016, P < .01]. However, there were no
significant interactions of Type (cid:2) Switch, underlining that
the considered brain areas showed a comparable ma-
nipulation switch effect in both real as well as pretend
Table 3. Inferring Goals: Contrasts between New and
Repeated/Old Information about Manipulations (G > Ö)
und Objekte (G > M)
Bereich
X
j
z
Z
mm3
Neue versus wiederholte Manipulationen (Contrast G > O)
OFC (BA 11/10)
PMv
(cid:1)24
(cid:1)42
IFS pars opercularis (BA 44) (cid:1)50
45 (cid:1)3
4.80
2619
27
21
3.71
3132
3.36
l.m.
IFG/IFS
PPA
(cid:1)41
9
3.83
36 (cid:1)33 (cid:1)6
5.12
864
756
6
18
33
Neue versus wiederholte Objekte (Contrast G > M)
LOC
(cid:1)33 (cid:1)66 (cid:1)3
3.31
1188
33 (cid:1)81 (cid:1)3
3.72
3375
(cid:1)33 (cid:1)48 (cid:1)9
4.33
7101
33 (cid:1)48 (cid:1)9
3.58
2781
Sensomotorischer Kortex
33 (cid:1)18
54
3.83
6939
Aktionen. Einen Haupteffekt haben wir für den Faktor Switch gefunden
(Objekt, Manipulation, oder beides) im OFC [F(2,34) = 10.325,
P < .005] due to significant differences between G > Ö
(t18 = 6.304, P < .001) and G > M (t18 = 2.485, P < .05);
the same was true for BA 44 [Switch, F(2,34) = 10.708,
p < .001; G > Ö, t18 = 3.977, P < .001; G > M, t18 =
3.169, P < .01], for IFS [Switch, F(2,34) = 11.414, p <
.001; G > Ö, t18 = 5.203, P < .001; G > M, t18 = 3.708,
P < .005], and for PMv [Switch, F(2,34) = 6.598, p < .01;
G > Ö, t18 = 4.22, P < .001; G > M, t18 = 2.876 P < .01];
only for PPA, G and M did not differ though showing the
same trend [Switch, F(2,34) = 5.15, p < .05; G > Ö, t18 =
4.018 P < .001].
We finally also calculated the interaction contrast G >
Ö (cid:2) Pretend > Real. Passend zum Signalwechsel
Analysen, Dieser Kontrast erbrachte keine nennenswerten Aktiva-
tionen, Dadurch wird diese vorgetäuschte und tatsächliche Handlung bestätigt-
tionen waren mit dem gleichen Aktivierungsmuster verbunden
in Bezug auf den Wechsel.
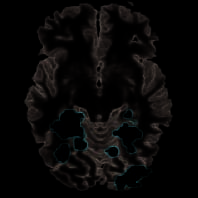
Ableiten von Zielen durch Analysieren neuer Objekte
Wie eingangs gesagt, Wir haben nicht mit Gemeinsamkeiten gerechnet
Aktivierungen für reale und vorgetäuschte Aktionen für vorhandene Prüfungen-
ing neue Objekte im Vergleich zu wiederholten Objekten, als Objekt in-
Es wurde erwartet, dass die Bildung weitgehend ignoriert wird
Aktionen. Unerwartet, Jedoch, für Objektwechsel versus
648
Zeitschrift für kognitive Neurowissenschaften
Volumen 21, Nummer 4
Objektwiederholungsversuche (G > M), Aktivierung wurde gefunden
bilateral in vier Teilregionen des LOC (Figur 5,
Tisch 3). Um dieses Ergebnis weiter zu untersuchen, ein Signalwechsel
Die Analyse wurde berechnet, enthüllte ein Muster, das war
für alle vier analysierten Bereiche vergleichbar: der Signalwechsel
war für alle Bedingungen außer ma vergleichsweise hoch-
Nipulationswechselversuche in vorgetäuschter Aktion, die dazu führte
ein niedrigeres Signal. Für linken und rechten vorderen LOC (Zuweisungen)
und rechter hinterer LOC (pLOC), Es gab einen Haupteffekt-
Perfekter Schalter [verließ aLOC, F(2,34) = 7.272, P < .01; right
aLOC, F(2,34) = 12,177, p < .001; right pLOC: F(2,34) =
6.802, p < .005] but also an interaction Switch (cid:2)
Type [left aLOC, F(2,34) = 4.008, p < .05; right aLOC,
F(2,34) = 10.597, p < .001; tendency in right pLOC,
F(2,34) = 3.187, p = .054] that was due to a higher
signal in all pretend actions that contained an object
switch [left aLOC: OP > Abgeordneter (t18 = 4.215, P < .001), GP >
Abgeordneter (t18 = 3.933, P < .001); right aLOC: OP > Abgeordneter (t18 =
6.815, P < .001), GP > Abgeordneter (t18 = 4.149, P < .001); right
pLOC: OP > Abgeordneter (t18 = 5.403, P < .001), GP > Abgeordneter (t18 =
4.106, P < .001)]. Left pLOC showed also a main effect
Switch [F(2,34) = 7.453, p < .005] that was due to a
higher signal in all actions that contained an object switch
[O > M (t18 = 3.558, P < .005), G > M (t18 = 3.104, P <
.01)] and a main effect for Type due to a higher signal in
real as compared with pretend action [F(2,34) = 18.327,
p < .001] but no interaction Switch (cid:2) Type.
DISCUSSION
The present fMRI study investigated goal inference from
pretend action as compared with real action. The in-
spection of real and pretend actions was considered to
differ with regard to the weighting of specific compo-
nents of the action observation network, pointing to-
ward an attentional focus on manipulation information
for pretend relative to real actions and an attentional fo-
cus on object information for real relative to pretend ac-
tions. In contrast, goal inference as an internal simulation
of the currently observed manipulations was expected to
yield comparable activations for real and pretend actions
in regions related to sensorimotor integration, working
memory, and internal reward evaluation.
Observing Pretend versus Real Actions: Relying
on Manipulation Information
Relative to the resting baseline, observation of real and
pretend actions yielded highly similar brain responses
in the typical action observation network, emphasizing
commonalities rather than fundamental differences in
the way we cope with real and pretend action. How-
ever, among the areas that were activated for obser-
vation of real and pretend actions, four areas showed
relatively enhanced response to pretend as compared
with real actions: left PMv, left lPFC, left aIPS (extend-
ing into SMG), and right and left EBA/hMT, the latter
extending into left pSTS as well (for anatomical con-
nections, cf. Schmahmann et al., 2007). As the two fron-
tal areas,
left PMv and left lPFC, were also found to
respond to goal switches, they will be considered sepa-
rately below.
Enhanced activation in EBA/hMT and pSTS had been
hypothesized for pretend versus real action observation
due to their function in body, motion, and biological mo-
tion processing, respectively (Downing et al., 2007; Taylor
et al., 2007; Peelen, Wiggett, & Downing, 2006; Puce &
Perrett, 2003; Downing, Jiang, Shuman, & Kanwisher,
2001). Because the presence of biological motion as well
as body parts was balanced between pretend and real
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
4
e
-
6
p
4
d
2
f
1
/
9
2
3
1
7
/
5
4
0
/
2
6
o
4
c
2
n
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
4
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
f
t
.
.
.
Figure 5. Object switch.
Contrast between trials
showing new versus repeated
objects (G > M) nachgegeben
erweiterte Aktivierung in der
LOC (aLOC = vorher, pLOC
hintere). Signaländerungen in
die vier lokalen Maxima zeigen
diese Wiederholung von Objekten
verursachte nur eine Dämpfung
Vortäuschen von Handlungen, wohingegen
Objektinformationen waren
in realen Aktionen verarbeitet nein
Egal ob wiederholt
(gelbe Balken) oder neu (Rot
und blaue Balken).
Ö
N
1
8
M
A
j
2
0
2
1
Schubotz and von Cramon
649
Aktionen, Wir schlagen vor, dass die Inspektion der Bewegung und
Körperinformationen wurden bei vorgetäuschten Handlungen intensiviert,
das ist, als die Zielschlussfolgerung ausschließlich auf Manipulation beruhen musste-
ulationsinformationen, während in realen Aktionen, Objektinformationen-
Für den gleichen Zweck könnte auch die Nutzung genutzt werden.
Jedoch, die Tatsache, dass das linke aIPS/SMG mehr war
aktiv für vorgetäuschtes Verhalten im Vergleich zum tatsächlichen Handlungsbeobachter-
vation fügt eine sehr interessante Facette hinzu, da diese Region war
Berichten zufolge relevant für die Leistung von Panto-
Mime ( Johnson-Frey, 2004; Ohgami et al., 2004; Moll
et al., 2000) und wurde zum expliziten Abruf vorgeschlagen
von werkzeugbezogenen Handbewegungen für unterschiedliche Verhaltensweisen
Zwecke (Imazu et al., 2007). Darüber hinaus, Hamilton und
Grafton (2006) gemeldete Aktivierung in IPS an Systemati-
werden durch die Wiederholung erreichter Objekte gezielt abgeschwächt (Aber
Flugbahnen nicht erreichen) bei der Aktionsbeobachtung.
Unsere Ergebnisse bestätigen insbesondere die Ergebnisse von pa-
Patienten mit Läsionen des unteren Parietals links
eine starke Beziehung zwischen der Anerkennung und dem
Nachahmung (Leistung) der objektbezogenen Pantomime
(Buxbaum, Kyle, et al., 2005) das kann man in Betracht ziehen
spiegeln Ausdrücke von Defiziten in internen Modellen wider
Planung objektbezogener Aktionen (Buxbaum, Johnson-
Frey, & Bartlett-Williams, 2005). In diesem Sinne, erhöht-
tion der inferioren parietalen Aktivierung zur Beobachtung vorgeben
im Vergleich zur realen Handlung kann als Mani interpretiert werden-
Festigungen höherer Anforderungen an diese interne Modellierung
wegen fehlendem (extern) Objektinformationen.
Vor allem,
Die inferiore parietale Aktivierung umfasste beides
SMG und aIPS in der vorliegenden Studie. Makakenforschung
impliziert zwei ziemlich unterschiedliche Prozesse in der Puta-
tive Homologe von SMG (Makakengebiet PF) und aIPS
(Makakengebiet AIP; vgl. Ausschuss et al., 2007; McGeoch,
Brang, & Ramachandran, 2007). Ersteres enthält
parietale Spiegelneuronen und vermittelt zwischen PMv und
pSTS in einem Netzwerk sowohl für Aktionsbeobachtung als auch für Aktion
Ausführung (Keyser & Perrett, 2004); Letzteres ist sugger-
Es wurde darum gebeten, dem PMv eine pragmatische Beschreibung zu geben
von Objekten (Fagg & Arbib, 1998). Funktional, es ist mehr
plausibel, unsere Ergebnisse im Sinne von pa zu interpretieren-
rietale Spiegelneuronen und die Analyse der beobachteten Aktion.
Jedoch, zusätzlich
Die Beteiligung von aIPS könnte z. B. sein-
auf zwei Arten erklärt. Als Objektinformationen, auf die verwiesen wird-
Das aktuell gültige Ziel war im Vorfeld nicht verfügbar
Aktion, aIPS kann entweder die getesteten Bilder widerspiegeln
Klassen von Objekten, die mit der aktuell beobachteten ma übereinstimmen-
Nipulationen, angetrieben durch Top-Down-Modulation von ventral
Es ist bekannt, dass Bilder dazu führen
prämotorische Bereiche. In der Tat,
In vielen Fällen sind die BOLD-Antworten höher als die Wahrnehmung
(z.B., Imazu et al., 2007). Alternativ, aIPS kann reflektieren
die Unterdrückung des derzeit ungültigen pragmatischen Objekts in-
Formation, die sich aus den dargestellten Wildcard-Objekten ergibt
in der vorgetäuschten Aktionsbedingung. Beachten Sie in diesem Zusammenhang, dass
man könnte vermuten, dass das Vorhandensein unangemessen ist
Objekte im vorgetäuschten Zustand können zu einer Aktivierung führen
von Gehirnregionen, die am Umgang mit Inkongruenzen beteiligt sind.
Jedoch, Die beiden Bereiche werden meist in erweiterter Form vorgeschlagen
kognitive Kontrolle während der Auflösung von Inkongruenzen, das ist,
der ACC- und der DLPF-Kortex (Fuhrmann & van Veen, 2007),
waren nicht Teil des Vergleichs zwischen vorgetäuschter und realer Handlung.
daher, Es scheint, dass Inkongruenzen auftreten und resultieren-
Die Steigerung der kognitiven Kontrolle spielte keine signifikante Rolle
Rolle für die vorgetäuschte Aktionsbedingung.
Beobachten realer und vorgetäuschter Handlungen: Sich verlassen
zu Objektinformationen
Der Kontrast zwischen realen und vorgetäuschten Handlungsbeobachtungen-
Es wurde erwartet, dass die Ausbeutung von Objekten im Vordergrund steht
Information. Es zeigte sich eine erweiterte LOC-Aktivität, ein Gebiet
Es ist bekannt, dass es eine wichtige Rolle bei der menschlichen Objektaufnahme spielt-
Erkenntnis. Im Rahmen der vorliegenden Studie, LOC ist da-
Engagement ist als ein repräsentierender Bereich höchst plausibel
die Formen von Objekten unabhängig von visuellen Hinweisen auf niedriger Ebene
wie zum Beispiel Farbe, Bewegung, oder Textur (Grill-Spector et al.,
2001). Als die Form eines Objekts und nicht als seine Farbe oder Textur
bestimmt/regelt die Handhaltung und Bewegung während der Aktion,
Die Erkennung der Objektform war von primärer Aufgabenrelevanz.
Ableiten von Zielen: Überprüfung neuer Manipulationen
Verwendung eines Switching-Protokolls, Wir haben uns genauer darauf eingelassen
Untersuchen Sie Zielschlussfolgerungen auf der Grundlage von Manipulation
Information. Obwohl diese Informationen berücksichtigt wurden
besonders relevant für das Verständnis von Pretend sein
Aktionen, Es wurde erwartet, dass es zur Zielschlussfolgerung ausgenutzt wird
sowohl in realen als auch in vorgetäuschten Handlungen, und es wurde angenommen
Besonders erhöht in Studien, da diese Art von Informationen-
tion wurde im Vergleich zum vorherigen Versuch geändert. The G >
O-Kontrast zeigte eine verstärkte Aktivierung in vier Bereichen zwei
davon waren Teil des Aktionsbeobachtungsnetzwerks und
auch verbessert für vorgetäuschte versus echte Action, nämlich, Die
linker ventraler prämotorischer Kortex (PMv), hier erstreckt sich hinein
die Pars opercularis des unteren Frontalgyrus (BA 44)
und der linke lPFC. Im Gegensatz, der linke OFC (BA 11/10, OFC
Jenseits) und das richtige PPA wurden nicht primär identifiziert
im Aktionsbeobachtungsnetzwerk aber ausschließlich gefunden
by contrasting G > O.
Zu den dem OFC zugeschriebenen Funktionen, ein aktueller
Die Überprüfung legt nahe, dass OFC Informationen darüber besitzt
Wert der Belohnungsergebnisse im Arbeitsgedächtnis, wann
Wir formulieren Aktionspläne und prognostizieren und überwachen Ex-
erwartete Ergebnisse ( Wallis, 2007). Um dies voll und ganz zu würdigen
Interpretation im Hinblick auf die vorliegenden Erkenntnisse, es ist
wichtig, das zu bedenken, zuerst, Wir haben OFC gefunden (als
sowie PPA) nur für Manipulationsschalter, aber nicht für
Objektschalter (siehe unten), und zweitens, diese Aktivität
wurde bei Vergleichsversuchen beobachtet, bei denen beide ma-
Manipulation und Objekte wurden vertauscht (Torwechsel) mit
Versuche, bei denen nur Objekte gewechselt wurden, dadurch auszuschließen
unspezifische Schalteffekte. In enger Übereinstimmung mit Wallis
(2007), Wir schlagen vor, dass das OFC die Beurteilung übernimmt-
ment von Kompromissen, wenn ein Szenario Abwechslung zulässt-
tive Handlungsziele und ermittelt, wie gut diese tatsächlich sind
650
Zeitschrift für kognitive Neurowissenschaften
Volumen 21, Nummer 4
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
F
T
/
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Das beobachtete Ergebnis erfüllt die aktuell getestete Zukunft
Modelle. Gleichzeitig mit OFC, Manipulationsschalter
verstärkte Aktivität im rechten PPA und in zwei Bereichen des
Aktionsbeobachtungsnetzwerk: links lPFC (BA 9/46) und ging
PMv. Diese Bereiche sind miteinander verbunden: OFC hat einige
Verbindungen zu PM (Morecraft, Geula, & Mesulam, 1992)
und intensive Verbindungen mit dem lPFC (siehe Wallis, 2007)
das wiederum hat Verbindungen zum PMv (Lu, Preston,
& Strick, 1994; Widerhaken & Pandya, 1987; Matelli, Camarda,
Glickstein, & Rizzolatti, 1986); darüber hinaus, PPA-Projekte
an beide OFC (Widerhaken, 1988) und lPFC (Goldman-Rakic,
Selemon, & Schwartz, 1984). Wie funktionieren diese Bereiche?-
im Rahmen von Zielwechseln interagieren?
Über die Aktivierung unseres Motors besteht weitgehend Einigkeit
System während der Aktionsbeobachtung ist auf einen internen zurückzuführen
Simulation der beobachteten Aktion; das ist, Das System ist
ähnlich aktiviert, als ob wir das Beobachtete ausführen würden
selbst handeln ( Jeannerod, 2001). Rechnerisch,
die Vorstellung mehrerer Vorwärtsmodelle, die parallel laufen-
Allel wurde verwendet, um die Effizienz zu erklären
Wir engagieren uns für das Ziel
Schlussfolgerung bei der Beobachtung von Handlungen
(Miall, 2003; Wolpert & Flanagan, 2001). Diese setzen
Perspektiven zusammen, das Bild, das aus dem hervorgeht
Aktuelle Erkenntnisse sind, dass die OFC-Aktivierung den Cal widerspiegelt-
Berechnung des Wertes eines Belohnungsergebnisses des Cur-
kürzlich getestete Vorwärtsmodelle der beobachteten Aktion.
Wallis (2007) führt aus, dass die lPFC die Belohnung verwendet
Signal vom OFC, Verhalten in Richtung Erhalt zu planen
das Ziel. Im Kontext der aktuell verwendeten Aktion
Beobachtungsaufgabe, Der lPFC kann daher aktuell auswählen
potenziell relevante Ziele einer Belohnung zu unterziehen
Analyse von OFC. Dazu gehört auch das Halten dieser Altäre-
native Pläne im Arbeitsgedächtnis. Potenziell relevanter Wechselstrom-
Die Ziele werden von unten nach oben durch den Input von PMv vorgegeben
das im Zusammenhang damit sensomotorische Simulationen durchführt
seine parietalen Projektionsstellen. Auf der Grundlage der Belohnung
Signale von OFC, lPFC kann wiederum die Auswahl ändern
aktuell getestete Handlungsziele (simuliert) in PMv. Notiz-
würdig, die parietale Projektionsstelle des PMv, das aIPS,
was missing in the G > O contrast. Daher, Aktivität in aIPS
wurde durch die Beobachtung von nicht wesentlich verbessert
neu im Vergleich zu wiederholten Manipulationen, möglicherweise
weil die Neuheit der Objektinformationen kontrolliert wurde
in diesem Gegensatz.
Im Hinblick auf den funktionalen Beitrag des PPA
in diesem Zusammenhang, Darauf bezieht sich eine spannende Erklärung
Die Rolle des Bereichs bei der Verarbeitung des Szenenkerns (Epstein, 2005). Nur
Was OFC betrifft, PPA wurde jedoch nur für Torwechseleffekte gesehen
nicht für die Handlungsbeobachtung an sich, was darauf hinweist, dass OFC
und PPA kann einen transienten Eingang bereitstellen, möglicherweise sein
Top-Down im Falle von OFC und Bottom-Up im Falle von
PPA, zu lPFC, Dadurch moduliert es seinen Einfluss auf PMv.
Ableiten von Zielen: Inspektion neuer Objekte
Im Gegensatz zur Prüfung von Neu versus Wiederholt
Manipulationen, was zum gleichen Wechselstrommuster führte-
Motivationen für reale und vorgetäuschte Handlungen, die Inspektion von
Es wurde festgestellt, dass neue Objekte im Vergleich zu wiederholten Objekten die LOC modulieren
ausschließlich für vorgetäuschte Aktionen (siehe Abbildung 5). Das Muster
von Aktivierungen impliziert, dass obwohl die Verarbeitung von
Die Objektform wurde unterdrückt oder vorzeitig abgebrochen
Objektwiederholung in vorgetäuschten Aktionen, neue Manipulationen
löste eine Neubewertung der Objektinformationen aus, sogar
bei Wiederholung, im Falle realer Handlungen.
„Theory of Mind“ versus „Mirror Neuron System“
Gegenüberstellung der Beobachtung einer vorgetäuschten Handlung mit einer tatsächlichen Handlung,
Wir haben eine Aktivierung in den besprochenen Bereichen festgestellt
gehören zum sogenannten „Spiegelneuronensystem“ (MNS)
Netzwerk (Rizzolatti & Craighero, 2004). Im Gegensatz, Die
einzige andere Studie, die die Beobachtung von Vortäuschen vergleicht
und echte Action (Deutsch et al., 2004) Aktiva gemeldet-
tionen, die sich auf die Zuschreibung mentaler Zustände beziehen, was auch so ist
wird als „Theorie des Geistes“ bezeichnet (ToM) (vgl. Gallagher &
Frith, 2003). Darüber gibt es eine anhaltende Debatte-
Beziehung zwischen MNS und ToM und ihren jeweiligen
Rollen beim Verständnis beobachteter Handlungen (vgl. Sachsen, 2005).
Allerdings sind wir nicht in der Lage, das Problem zu überbrücken-
Große Lücke zwischen den neuronalen Korrelaten der Zielinferenz
und Gedankenlesen im Allgemeinen, erhebliche Unterschiede
im experimentellen Design von German et al. (2004)
Studie und unsere müssen berücksichtigt werden, das kann ac-
zählen für abweichende Ergebnisse. zuerst, der Anblick des Ganzen
Personen können ToM-Prozesse viel stärker provozieren als
der Anblick von Händen auf Gegenständen. Zweitens, weil Objekte
fehlten in den eingesetzten vorgetäuschten Aktionen gänzlich
von German et al., Die Probanden waren möglicherweise engagierter
in aktiven Überlegungen und Schlussfolgerungen über die Klasse
von Objekten, bei denen eine Manipulation vorgetäuscht wurde. Fi-
schließlich, Kurzfilme könnten eine direkte Zuordnung verzerrt haben
der beobachteten Manipulationen auf das eigene Handeln
Repertoire, wohingegen bei längeren Beobachtungszeiten,
Es können sich weitere kognitive Prozesse entwickeln, sogar automatisch-
automatisch, die sich auf die Absichten und Männer des Schauspielers beziehen-
tal-Staaten.
Aus Vortäuschung auf Ziele schließen: Allgemein
Abschließende Bemerkungen
Das Bild, das sich aus den vorliegenden Erkenntnissen ergibt, ist verwirrend
Neues Licht auf die Mechanismen, die die Schlussfolgerung steuern
Ziele aus beobachtetem Schein sowie beobachteter Realität
Aktion. Wir haben klare Belege für beide Gemeinsamkeiten gefunden
und Unterschiede zwischen den neuronalen Korrelaten von unter-
Stehen, vorgeben und echte Handlungen. Auffallend, unser Fund-
ings zeigen, dass nicht alle Komponenten der Aktion
Beobachtungsnetzwerke beschäftigen sich mit der Schlussfolgerung von
neue Ziele in beobachteten Handlungen und so weiter, umgekehrt, nicht
alle Bereiche, die sich mit der Schlussfolgerung eines neuen AC befassen-
tionsziel sind integraler Bestandteil der Handlungsbeobachtung
Netzwerk. Diese Dissoziation könnte darauf zurückzuführen sein
Inferenzprozesse sind subtiler und phasischer
Ziel
Schubotz and von Cramon
651
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
.
.
F
.
T
/
Ö
N
1
8
M
A
j
2
0
2
1
als die massive und tonische Aktivität, die durch das Ob ausgelöst wird-
Bedienung eines laufenden Geschehens mit all seinen Facetten pro-
Konzeptuelle Analyse und mnemonische Anforderungen.
Die vorliegenden Ergebnisse implizieren, dass die Anforderung besteht
Die Ableitung eines neuen Ziels stützt sich unter anderem auf eine Auswahl dieser Ziele
Bereiche, die eher der Manipulationsanalyse dienen
als Objektinformationen. Die Tendenz zur Manipulation
hängt wahrscheinlich damit zusammen, dass wir es operativ gemacht haben
Ziele als Manipulationsketten von Objekten und nicht, als
wäre sicherlich möglich gewesen, als Objektziele
(z.B., eine Diskette, ein Keks), räumliche Ziele (z.B., links, Rechts),
oder Kombinationen aus beidem (vgl. Hamilton & Grafton, 2006).
Unsere Erkenntnisse deuten auf diese Ziele hin, wenn dies operationalisiert wird
Weg, hängen eher damit zusammen, wie sich jemand bewegt-
auf Objekte gerichtet als auf die Objekte selbst, Pos-
offensichtlich, weil es sich um eine Kette von Handhaltungen und -bewegungen handelt
ist im Hinblick auf das angestrebte Ziel weniger eindeutig,
wohingegen Objekte vielfältige Möglichkeiten bieten
Zielgerichtete Manipulationen.
Danksagungen
Wir danken Anna Abraham herzlich, Uta Wolfensteller, Und
Kirsten Volz für ihre sehr hilfreichen Kommentare zum Handbuch-
Skript, Gaby Lohmann und Karsten Müller für Unterstützung im MRT
Statistiken, Andrea Gast-Sandmann and Kerstin Flake for support
in grafischem Layout und Anregungsmaterialien, und Marcel Mücke
für experimentelle Unterstützung.
Nachdruckwünsche sind an Ricarda I zu richten. Schubotz, Motor
Erkenntnisgruppe, Max-Planck-Institut für neurologische Forschung-
suchen, Gleueler Str. 50, 50931 Köln, Deutschland, oder per E-Mail:
schubotz@nf.mpg.de.
VERWEISE
Widerhaken, H. (1988). Anatomische Organisation von basoventralen und
mediodorsale präfrontale Regionen des visuellen Empfängers im Rhesus
Affe. Zeitschrift für Vergleichende Neurologie, 276, 313–342.
Widerhaken, H., & Pandya, D. N. (1987). Architektur und Frontal
kortikale Verbindungen des prämotorischen Kortex (Bereich 6) In
der Rhesusaffen. Zeitschrift für Vergleichende Neurologie,
256, 211–228.
Wellhornschnecke, G., Binkofski, F., Fink, G. R., Ermüdung, L., Fogassi, L.,
Walisisch, V., et al. (2001). Aktionsbeobachtung wird aktiviert
prämotorische und parietale Bereiche auf somatotopische Weise:
Eine fMRT-Studie. Europäisches Journal für Neurowissenschaften, 13,
400–404.
Buxbaum, L. J., Johnson-Frey, S. H., & Bartlett-Williams, M.
(2005). Unzureichende interne Modelle zur Planung von Handobjekten
Wechselwirkungen bei Apraxie. Neuropsychologie, 43, 917–929.
Buxbaum, L. J., Kyle, K. M., & Menon, R. (2005). Im Jenseits
Spiegelneuronen: Interne Darstellungen dienen
Nachahmung und Erkennung geschickter objektbezogener Handlungen
beim Menschen. Gehirnforschung. Kognitive Gehirnforschung,
25, 226–239.
Calvo-Merino, B., Gre`zes, J., Glaser, D. E., Passingham, R. E.,
& Haggard, P. (2006). Sehen oder tun? Einfluss des Visuellen
und motorische Vertrautheit bei der Aktionsbeobachtung. Aktuelle Biologie,
16, 1905–1910.
Fuhrmann, C. S., & van Veen, V. (2007). Vorderer cingulärer Kortex und
Konflikterkennung: Ein Update von Theorie und Daten. Kognitiv,
Affektiv & Verhaltensneurowissenschaften, 7, 367–379.
Sich verpflichten, G., Pitzalis, S., Galata, G., Patria, F., Haut, G.,
Sabatini, U., et al. (2007). Neuronale Grundlagen persönlicher und
außerpersönliche Vernachlässigung beim Menschen. Gehirn, 130, 431–441.
Konstantin, M., Galata, G., Ferretti, A., Caulo, M., Zahnstein, A.,
Roma, G. L., et al. (2005). Neuronale Systeme zugrunde liegen
Beobachtung menschlich unmöglicher Bewegungen: Ein fMRT
Studie. Hirnrinde, 15, 1761–1767.
Downing, P. E., Jiang, Y., Schumann, M., & Kanwisher, N.
(2001). Ein kortikaler Bereich, der selektiv für die visuelle Verarbeitung von ist
der menschliche Körper. Wissenschaft, 293, 2470–2473.
Downing, P. E., Wiggett, A., & Peeling, M. (2007). fMRT
Untersuchung überlappender lateraler okzipitotemporaler Bewegungen
Aktivierungen mittels Multi-Voxel-Musteranalyse. Zeitschrift
der Neurowissenschaften, 27, 226–233.
Epstein, R. A. (2005). Die kortikale Basis der visuellen Szene
Verarbeitung. Visuelle Wahrnehmung, 12, 954–978.
Epstein, R. A., & Kanwisher, N. (1998). Ein kortikaler
Darstellung der lokalen visuellen Umgebung. Natur,
392, 598–601.
Fagg, A., & Arbib, M. (1998). Modellierung parietal-prämotorisch
Interaktionen bei der Kontrolle des Greifens durch Primaten. Neuronal
Netzwerke, 11, 1277–1303.
Fein, G. G. (1981). Rollenspiele in der Kindheit: Ein Integrativ
Rezension. Kinderentwicklung, 52, 1095–1118.
Friston, K. J., Holmes, A. P., Worsley, K. J., Polina, J. P., Frith,
C. D., & Frackowiak, R. S. J. (1995). Statistische Parameter
Karten in der funktionellen Bildgebung: Ein allgemeiner linearer Ansatz.
Kartierung des menschlichen Gehirns, 2, 189–210.
Gallagher, H. L., & Frith, C. D. (2003). Funktionelle Bildgebung
der „Theorie des Geistes“. Trends in den Kognitionswissenschaften, 7,
77–83.
Deutsch, T. P., Niehaus, J. L., Roarty, M. P., Giesbrecht, B.,
& Müller, M. B. (2004). Neuronale Korrelate des Erkennens
Vorwand: Automatisches Eingreifen des Vorsatzes
Haltung unter verdeckten Bedingungen. Zeitschrift für Kognition
Neurowissenschaften, 16, 1805–1817.
Goldman-Rakic, P. S., Selemon, L. D., Schwartz, M. L. (1984).
Duale Bahnen, die den dorsolateralen präfrontalen Kortex verbinden
mit der Hippocampusformation und dem Parahippocampus
Kortex beim Rhesusaffen. Neurowissenschaften, 12, 719–743.
Greenlee, M. W. (2000). Menschliche kortikale Bereiche liegen zugrunde
die Wahrnehmung des optischen Flusses: Untersuchungen zur Bildgebung des Gehirns.
International Review of Neurobiology, 44, 269–292.
Grill-Spector, K., Kourtzi, Z., & Kanwisher, N. (2001). Der
lateraler Hinterhauptskomplex und seine Rolle bei der Objekterkennung.
Sehforschung, 41, 1409–1422.
Grill-Spector, K., & Malach, R. (2001). FMR-Anpassung: Ein Werkzeug
zur Untersuchung der funktionellen Eigenschaften der menschlichen Kortikalis
Neuronen. Acta Psychologica, 107, 293–321.
Grush, R. (2004). Die Emulationstheorie der Darstellung:
Motorsteuerung, Bilder, und Wahrnehmung. Verhaltens- und
Gehirnwissenschaften, 27, 377–442.
Hamilton, A. F., & Grafton, S. T. (2006). Zieldarstellung
im vorderen intraparietalen Sulcus des Menschen. Zeitschrift für
Neurowissenschaften, 25, 1133–1137.
Helbig, H. B., Graf, M., & Kiefer, M. (2006). Die Rolle von
Aktionsdarstellung bei der visuellen Objekterkennung. Gehirn
Forschung. Experimentelle Hirnforschung, 174, 221–228.
Hermsdörfer, J., Linde, G., Mühlau, M., Goldenberg, G.,
& Wohlschläger, A. M. (2007). Neuronale Darstellungen
von pantomimischem und tatsächlichem Werkzeuggebrauch: Beweise aus einem
ereignisbezogene fMRT-Studie. Neurobild, 36, T109–T118.
Imazu, S., Sugio, T., Tanaka, S., & Hassen, T. (2007). Unterschiede
zwischen tatsächlichem und imaginärem Gebrauch von Essstäbchen: Ein
fMRT-Studie. Kortex, 43, 301–307.
Jeannerod, M. (2001). Neuronale Handlungssimulation: A
vereinheitlichender Mechanismus für die motorische Wahrnehmung. Neurobild, 14,
S103–S109.
652
Zeitschrift für kognitive Neurowissenschaften
Volumen 21, Nummer 4
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
T
.
.
/
.
F
Ö
N
1
8
M
A
j
2
0
2
1
Johnson-Frey, S. H. (2004). Die neuronalen Grundlagen komplexer Werkzeuge
Anwendung beim Menschen. Trends in den Kognitionswissenschaften, 8, 71–78.
Johnson-Frey, S. H., Maluof, F. R., Newman-Norlund, R.,
Farrer, C., Es sagte, S., & Grafton, S. T. (2003). Aktionen bzw
Hand-Objekt-Interaktionen? Menschlicher unterer frontaler Kortex
und Handlungsbeobachtung. Neuron, 39, 1053–1058.
Keyser, C., & Perrett, D. ICH. (2004). Soziales entmystifizieren
Erkenntnis: Eine hebräische Perspektive. Trends im kognitiven Bereich
Wissenschaften, 8, 501–507.
Kro´liczak, G., Cavina-Pratesi, C., Guter Mann, D. A., & Culham,
J. C. (2007). Was macht das Gehirn, wenn man es vortäuscht??
Eine fMRT-Studie zum pantomimischen und realen Greifen. Zeitschrift
der Neurophysiologie, 97, 2410–2422.
Lohmann, G., Müller, K., Bosch, V., Mentzel, H., Heßler, S.,
Chen, L., et al. (2001). LIPSIA – Ein neues Softwaresystem für
die Auswertung funktioneller Magnetresonanzbilder von
das menschliche Gehirn. Computergestützte medizinische Bildgebung und
Grafik, 25, 449–457.
Lu, M., Preston, J., & Strick, P. (1994). Verbindungen
zwischen dem präfrontalen Kortex und den prämotorischen Bereichen
der Frontallappen. Zeitschrift für Vergleichende Neurologie, 341,
375–392.
Manthey, S., Schubotz, R. ICH., & von Cramon, D. Y. (2003).
Prämotorischer Kortex bei der Beobachtung fehlerhafter Handlungen: Ein fMRT
Studie. Gehirnforschung. Kognitive Gehirnforschung, 15,
296–307.
Matelli, M., Camarda, R., Glickstein, M., & Rizzolatti, G.
(1986). Afferente und efferente Projektionen des Unterkiefers
Bereich 6 beim Makaken. Zeitschrift für Vergleichende
Neurologie, 251, 281–298.
McGeoch, P. D., Brang, D., & Ramachandran, V. S. (2007).
Apraxie, Metapher und Spiegelneuronen. Medizinische Hypothesen,
69, 1165–1168.
Miall, R. C. (2003). Spiegelneuronen verbinden und weiterleiten
Modelle. NeuroReport, 14, 2135–2137.
Moll, J., de Oliveira-Souza, R., Passmann, L. J., Cunha, F. C.,
Souza-Lima, F., & Andreiuolo, P. A. (2000). Funktional
MRT-Korrelate realer und imaginärer Werkzeuggebrauchspantomimen.
Neurologie, 54, 1331–1336.
Morecraft, R. J., Geula, C., & Mesulam, M. M. (1992).
Zytoarchitektur und neuronale Afferenzen des orbitofrontalen Kortex
im Gehirn des Affen. Zeitschrift für Vergleichende
Neurologie, 323, 341–358.
Naccache, L., & Dehaene, S. (2001). Die Priming-Methode:
Die Bildgebung durch unbewusstes Wiederholungspriming bringt eine Zusammenfassung zum Vorschein
Darstellung der Zahl in den Parietallappen. Zerebral
Kortex, 11, 966–974.
Newman-Norlund, R. D., von Schie, H. T., van Zuijlen, A. M.,
& Bekkering, H. (2007). Das Spiegelneuronensystem ist mehr
aktiv während komplementär im Vergleich zu nachahmend
Aktion. Naturneurowissenschaften, 10, 817–818.
Norris, D. G. (2000). Multiskalen-MDEFT mit reduzierter Leistung
Bildgebung. Zeitschrift für Magnetresonanztomographie, 11,
445–451.
Ohgami, Y., Matsuo, K., Uchida, N., & Nakai, T. (2004). Ein
fMRT-Studie zu Gesten bei der Verwendung von Werkzeugen: Körperteil als Objekt und
Pantomime. NeuroReport, 15, 1903–1906.
Peeling, M. V., Wiggett, A. J., & Downing, P. E. (2006). Muster
der fMRT-Aktivität dissoziieren überlappende funktionelle Gehirnfunktionen
Bereiche, die auf biologische Bewegung reagieren. Neuron, 49,
815–822.
Petride, M. (2005). Lateraler präfrontaler Kortex: Architektonisch
und funktionale Organisation. Philosophische Transaktionen
der Royal Society of London, Serie B, Biologisch
Wissenschaften, 360, 781–795.
Peuskens, H., Vanrie, J., Verfaillie, K., & Orban, G. A.
(2005). Spezifität von Regionen, die biologische Bewegung verarbeiten.
Europäisches Journal für Neurowissenschaften, 21, 2864–2875.
Chip, A., & Perrett, D. (2003). Elektrophysiologie und Gehirn
Abbildung biologischer Bewegung. Philosophische Transaktionen
der Royal Society of London, Serie B, Biologisch
Wissenschaften, 358, 435–445.
Rizzolatti, G., & Craighero, L. (2004). Das Spiegelneuron
System. Jahresrückblick auf die Neurowissenschaften, 27, 169–192.
Rizzolatti, G., & Luppino, G. (2001). Der kortikale Motor
System. Neuron, 31, 889–901.
Sachsen, R. (2005). Gegen Simulation: Das Argument von
Fehler. Trends in den Kognitionswissenschaften, 9, 174–179.
Schmahmann, J. D., Pandya, D. N., Wang, R., Dai, G., D'Arceuil,
H. E., de Crespigny, A. J., et al. (2007). Assoziationsfaser
Bahnen des Gehirns: Parallele Beobachtungen aus der Diffusion
Spektralbildgebung und Autoradiographie. Gehirn, 130,
630–653.
Schubotz, R. ICH., & von Cramon, D. Y. (2004). Sequenzen von
abstrakte nichtbiologische Reize teilen sich den ventralen Prämotor
Kortex mit Aktionsbeobachtung und Bildern. Zeitschrift für
Neurowissenschaften, 24, 5467–5474.
Talairach, J., & Tournoux, P. (1988). Koplanare Stereotaxie
Atlas des menschlichen Gehirns. New York: Thieme.
Taylor, J. C., Wiggett, A. J., & Downing, P. E. (2007). fMRT
Analyse von Körper- und Körperteildarstellungen im
extrastriatische und fusiforme Körperbereiche. Zeitschrift für
Neurophysiologie, 98, 1626–1633.
Thompson-Schill, S. L., D'Esposito, M., & Kann, ICH. P. (1999).
Auswirkungen von Wiederholung und Wettbewerb auf die Aktivität der linken Seite
Präfrontaler Kortex während der Wortbildung. Neuron, 23,
513–522.
Ugur-Auto, K., Garwood, M., Ellermann, J., Heinrich, K., Wanderung,
R., Hu, X., et al. (1993). Bildgebung bei hohen Magnetfeldern:
Erste Erfahrungen bei 4 T. Magnetresonanz vierteljährlich,
9, 259–277.
Vogt, S., & Thomaschke, R. (2007). Von der Visuomotorik
Interaktionen bis hin zum Nachahmungslernen: Verhalten und Gehirn
Bildgebungsstudien. Zeitschrift für Sportwissenschaften, 25, 497–517.
Wallis, J. D. (2007). Orbitofrontaler Kortex und sein Beitrag
zur Entscheidungsfindung. Jahresrückblick auf die Neurowissenschaften, 30,
31–56.
Watanabe, M. (2007). Rolle der erwarteten Belohnung im kognitiven Bereich
Verhaltenskontrolle. Aktuelle Meinung in der Neurobiologie,
17, 213–219.
Wolpert, D. M., & Flanagan, J. R. (2001). Motorvorhersage.
Aktuelle Biologie, 11, R729–R732.
Worsley, K. J., & Friston, K. J. (1995). Analyse von fMRT
Zeitreihen erneut betrachtet – noch einmal. Neurobild, 2, 173–181.
Schubotz and von Cramon
653
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
1
l
4
e
–
6
P
4
D
2
F
1
/
9
2
3
1
7
/
5
4
0
/
2
6
Ö
4
C
2
N
/
1
2
0
7
0
6
9
0
0
2
0
1
7
0
4
/
9
J
Ö
P
C
D
N
.
B
2
j
0
G
0
u
9
e
.
S
T
2
Ö
1
N
0
4
0
9
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
e
0
2
S
3
T
/
J
/
.
.
F
T
.
Ö
N
1
8
M
A
j
2
0
2
1