tDCS Modulation of Visually Induced Analgesia
Flavia Mancini1, Nadia Bolognini2,3, Patrick Haggard1,
and Giuseppe Vallar2,3
Abstrakt
■ Multisensory interactions can produce analgesic effects. Im Par-
besonders, viewing oneʼs own body reduces pain levels, perhaps be-
cause of changes in connectivity between visual areas specialized
for body representation, and sensory areas underlying pain per-
ception. We tested the causal role of the extrastriate visual cortex
in triggering visually induced analgesia by modulating the excitabil-
ity of this region with transcranial direct current stimulation
(tDCS). Anodal, cathodal, or sham tDCS (2 mA, 10 min) was admin-
istered to 24 healthy participants over the right occipital or over
the centro-parietal areas thought to be involved in the sensory
processing of pain. Participants were required to rate the intensity
of painful electrical stimuli while viewing either their left hand or
an object occluding the left hand, both before and immediately
after tDCS. We found that the analgesic effect of viewing the body
was enhanced selectively by anodal stimulation of the occipital
Kortex. The effect was specific for the polarity and the site of stim-
ulation. The present results indicate that visually induced analgesia
may depend on neural signals from the extrastriate visual cortex. ■
EINFÜHRUNG
The experienced level of pain strongly depends on the
context in which nociceptive stimuli occur. Attention, ex-
pectations, and motivation are well-known examples of
contextual modulation (Wiech, Ploner, & Tracey, 2008).
In addition to modulation of pain by psychological con-
texts, multisensory perceptual contexts may also modu-
late pain. Zum Beispiel, simply looking at oneʼs own
body can be analgesic. In neurologically unimpaired par-
ticipants, passive vision of the hand reduces pain ratings
and the N2/P2 complex of laser-evoked potentials for
the same stimuli (Longo, Betti, Aglioti, & Haggard, 2009).
Außerdem, viewing oneʼs own hand increases contact
heat pain thresholds by 3.2°C, relative to viewing a neutral
Objekt (Mancini, Longo, Kammers, & Haggard, 2011). Das
“visually induced analgesia” can be considered a form of
multisensory modulation in which the visual representa-
tion of the body has effects on pain processing.
Interactions between visual and somatosensory areas
seem to mediate this modulatory effect of viewing the
Körper. A recent functional MRI experiment demonstrated
that changes in the effective connectivity between visual
and pain centers in the cerebral cortex underlie visually
induced analgesia (Longo, Iannetti, Mancini, Driver, &
Haggard, 2012). Insbesondere, that study showed increased
functional coupling between visual and parietal areas that
are activated by the visual perception of oneʼs own body,
including the “extrastriate body area” (EBA) in the lateral
occipital cortex (Downing, Jiang, Schumann, & Kanwisher,
1University College London, 2University of Milano-Bicocca,
3IRCCS Istituto Auxologico Italiano
© 2012 Massachusetts Institute of Technology Published under a
Creative Commons Attribution 3.0 Unportiert (CC BY 3.0) Lizenz
2001) and areas activated by painful stimuli. In this study,
we tested the causal role of the extrastriate visual cortex in
triggering the cross-modal modulation of pain perception.
To achieve this aim, we modulated the level of excitability
of the extrastriate visual cortex by transcranial direct cur-
rent stimulation (tDCS) and examined the resulting effect
on visually induced analgesia.
tDCS is a noninvasive technique of brain stimulation
that can modulate cortical excitability by polarizing brain
tissue in a polarity-dependent fashion, with anodal stim-
ulation generally increasing excitability, and cathodal stim-
ulation generally reducing excitability (Brunoni et al., 2012;
Paulus, 2011; Nitsche et al., 2003; Nitsche & Paulus, 2001).
When delivered to specific cortical areas, tDCS can alter
physiological, perceptual, and higher-order cognitive pro-
Prozesse (Vallar & Bolognini, 2011; Antal, Nitsche, & Paulus,
2001; Nitsche & Paulus, 2001). tDCS alters activity of corti-
cal areas situated under the electrodes, but also of distant
Bereiche, probably through interconnections of the primary
stimulated area with these structures (Polania, Nitsche, &
Paulus, 2011).
The behavioral task was to rate the intensity of a pain-
ful sensation elicited by trains of electrical shocks applied
to the left hand in two different visual contexts: Teilnehmer
were required to look at their own left hand (“hand-view”
condition) or at an object occluding that hand (a wooden
block, “object-view” condition). In two separate experi-
gen, we investigated the effect of anodal (Experiment 1)
and cathodal (Experiment 2) tDCS over the occipital cortex
(extrastriate visual) and the centro-parietal cortex, welche
contributes to sensory processing of nociceptive stimuli
(Valentini et al., 2012; Liang, Mouraux, & Iannetti, 2011;
Tracey, 2011). We expected that tDCS over the occipital
Zeitschrift für kognitive Neurowissenschaften 24:12, S. 2419–2427
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
Ö
8
C
4
N
7
_
1
A
/
_
J
0
Ö
0
C
2
N
9
3
_
A
P
_
D
0
0
B
2
j
9
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
cortex would not modulate pain generally, but specifically
influence the analgesic effect of viewing the body (increas-
ing or decreasing analgesia, depending on the tDCS polar-
ität). Umgekehrt, tDCS over the centro-parietal cortex might
induce analgesia independent of the visual context, accord-
ing to previous reports that demonstrate analgesic effects
of both anodal and cathodal stimulation of the motor cor-
tex without any explicit visual task (for reviews, see Vallar &
Bolognini, 2011; Lefaucheur et al., 2008).
METHODEN
Teilnehmer
Twenty-four naive right-handed (Oldfield, 1971) partici-
Hose (Durchschnittsalter = 23.4 Jahre, SD = 4.4 Jahre; 17 Frauen)
took part in the two experiments (n = 12 jede). All were
free of medical disorders, substance abuse or depen-
dence, CNS effective medication, and psychiatric and neu-
rological disorders (Poreisz, Boros, Antal, & Paulus, 2007)
and participated on the basis of informed consent. Guide-
lines of the ethical committees of the University of Milano-
Bicocca (Milan, Italien) and the Declaration of Helsinki (BMJ
1991; 302: 1194) were followed.
Stimuli
For stimulation, we used a custom planar concentric elec-
trode, consisting of a central metal cathode (diameter =
0.5 mm), an isolation insert (diameter = 5 mm), und ein
external anode ring (diameter = 6 mm), and providing a
stimulation area of 19.6 mm2. The electrode was applied
along the digital nerve path, approximately on the second
metacarpal space of the left hand. In jedem Versuch, a 500-msec
train of electrical shocks at 10 Hz was generated by a Digi-
timer DS7A electrical stimulator (www.digitimer.com/)
under computer control.
By virtue of its concentric design and small anode–
cathode distance, this somatosensory stimulating electrode
produces high current density at low current intensities. Es
can therefore depolarize the superficial layer of the dermis
containing nociceptive A-delta fibers (Kaube, Katsarava,
Kaufer, Diener, & Ellrich, 2000); Jedoch, A-beta fibers also
might be concomitantly stimulated (de Tommaso et al.,
2011). Pinprick-like painful sensation is generally produced
at currents between 0.6 Und 1.6 mA. Mean onset latencies
of blink reflexes and pain-related evoked potentials for
such stimulation were found to be compatible with con-
duction velocities of A-delta fibers (Katsarava et al., 2006;
Katsarava, Ellrich, Diener, & Kaube, 2002).
At the beginning of each session, the individual thresh-
old for painful pinprick sensations was identified by two
ascending and descending stimulation sequences in 0.1 mA
Schritte. The mean threshold was 0.52 mA (SD = 0.26 mA).
Two different intensities (+0.20 Und +0.70 mA above
individual pain thresholds) were then selected and used
in the main experiments (low-intensity stimulus: mean =
0.70 mA, SD = 0.26 mA; high-intensity stimulus: mean =
1.18 mA, SD = 0.32 mA).
Verfahren
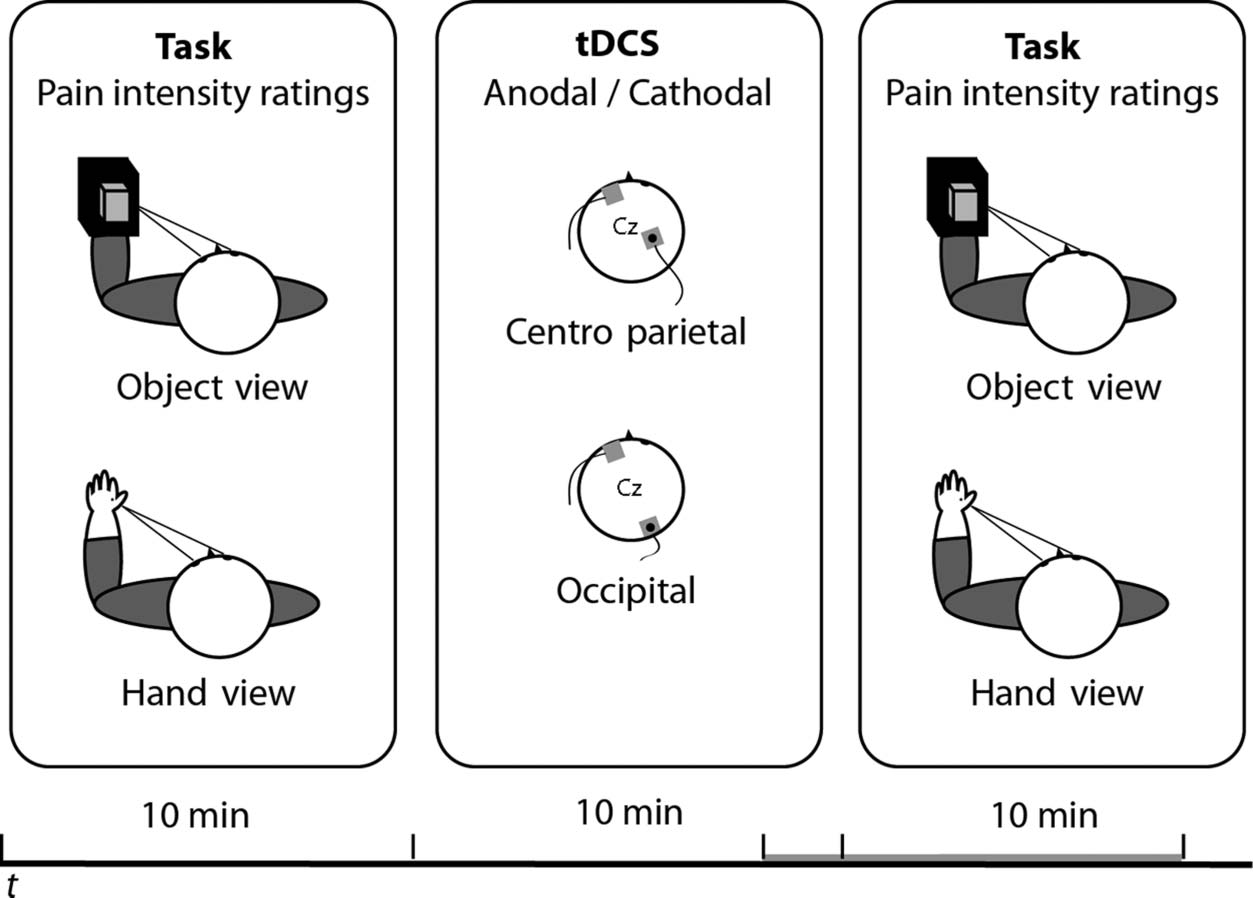
Participants sat at a table with their hands resting palm
down on the desktop, gazing toward their left hand
(Figur 1). A black cape hid from sight their arms and
right hand, so that participants could see only their left
Hand. Two visual conditions, “hand-view” and “object-
view,” were presented in different blocks in an ABBA
Befehl (initial condition counterbalanced across partici-
pants and sessions). In the “hand-view” condition, partici-
pants gazed toward their left hand. In the “object-view”
condition, the left hand was occluded by a box, and par-
ticipants looked at a hand-sized wooden block placed on
Figur 1. Stimuli and
procedure. Two sessions of
the same behavioral task were
administered before and after
10 min of 2 mA tDCS over
either the right occipital or the
centro-parietal cortex. Während
the behavioral task, Teilnehmer
were required to look at their
own left hand or at an object in
the same spatial location while
verbally rating the intensity of
electrical shocks delivered on
their left hand.
2420
Zeitschrift für kognitive Neurowissenschaften
Volumen 24, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
Ö
8
C
4
N
7
_
1
A
/
_
J
0
Ö
0
C
2
N
9
3
_
A
P
_
D
0
0
B
2
j
9
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
top of it (etwa 3 cm above the hand). Partici-
pants were instructed to fixate the hand/object continu-
ously. In jedem Versuch, a 500-msec train of five shocks at
either low or high intensity was administered. Teilnehmer
were asked to verbally rate the intensity of the stimulus
using a pain scale, aus 0 ( just noticeable) Zu 100 (worst
pain imaginable; Ohnhaus & Adler, 1975). The intensity
of the stimulus was randomized within each block. Jede
condition was repeated four times per block, for a total of
eight repetitions per behavioral test (32 total trials, taking
etwa 10 min). The test was repeated before and
after each of the three tDCS sessions (siehe unten).
tDCS
In two different experiments, either anodal (Experiment 1)
or cathodal (Experiment 2) stimulation (2 mA/35 cm2) War
administered. The tDCS stimulation was delivered by a
battery-driven constant current stimulator (www.eldith.
de/products/stimulator), using a pair of surface saline-
soaked sponge electrodes. A constant current of 2 mA
intensity was applied, complying with current safety guide-
lines (Poreisz et al., 2007). The stimulating current was
ramped up during a 10-sec fade-in phase, then held con-
stant at 2 mA for 10 min, and then ramped down during
a 10-sec fade-out phase. The duration of the tDCS stimula-
tion was chosen on the basis of previous literature, mit
effects on cortical excitability sufficiently enduring to cover
the duration of our experimental task (Nitsche & Paulus,
2001). The experimental task was initiated in the last 2 min
of tDCS, wie in der Abbildung gezeigt 1.
Each participant performed three sessions of tDCS
(Hinterhaupt, centro-parietal, and sham stimulation), vorgeführt
in counterbalanced order across participants, and separated
by at least 90 min to avoid carryover effects and to guaran-
tee a sufficient washout of the effects of the previous run
(z.B., Bolognini, Rossetti, Casati, Mancini, & Vallar, 2011;
Bolognini, Fregni, Casati, Olgiati, & Vallar, 2010; Bolognini,
Olgiati, Rossetti, & Maravita, 2010; Boggio et al., 2009;
Sparing et al., 2009; Ragert, Vandermeeren, Camus, &
Cohen, 2008). Während der 90 min of break, Teilnehmer
were free to leave the laboratory. In different sessions,
the active electrode (to which polarity refers) was placed
over one of the targeted areas in the right hemisphere,
according to the 10–20 system for EEG electrode place-
ment. Wichtig, participants were naive to all the stim-
ulation conditions, being not informed as to which they
had been assigned to and as to whether the stimulation
was real or sham.
For occipital stimulation (Ö), the active electrode was
placed between O2 and PO8 to stimulate the extrastriate
visual cortex including visual body-specific regions
(Downing et al., 2001).
For centro-parietal stimulation (CP), the active elec-
trode was placed between CP4 and C4, in proximity to
the primary somatosensory cortex (Overduin & Servos,
2004; Geyer, Schleicher, & Zilles, 1999). For both areas,
the reference electrode was placed over a contralateral
supraorbital region (Fp3), as this montage has proven
to be effective in previous tDCS experiments (Dasilva,
Volz, Bikson, & Fregni, 2011; Vallar & Bolognini, 2011).
For sham stimulation, the electrodes were placed over
one of the target areas, the same parameters of stimulation
were employed, but the stimulator was turned off after
30 Sek. This ensured that participants could feel the initial
itching sensation at the beginning of tDCS but was as-
sumed not to produce any effective modulation of cortical
excitability by tDCS (Gandiga, Hummel, & Cohen, 2006).
In-house software switched the tDCS on and off without
intervention from the participant or experimenter, erlauben-
ing for a successful blinding of both of them. For each
Experiment, six participants received sham stimulation at
the occipital site and six at the centro-parietal site.
Multiple testing sessions were used to allow tDCS effects
to wash out. A corollary of this design, Jedoch, ist der
session-to-session variability in pain ratings (Rosier, Iadarola,
& Coghill, 2002; Yarnitsky & Sprecher, 1994). Trotzdem,
because we were interested in the modulation of context
and time within sessions, rather than main effects of ses-
sion, changes in overall pain levels between sessions were
not problematic.
Analyses
Statistical analyses were performed separately for each
Experiment. Erste, planned comparisons were conducted
to assess visually induced analgesia; we averaged the raw
pain ratings across all three pre-tDCS conditions and
compared the view-hand and view-object conditions with
a paired t test.
Zweite, we explored the effect of tDCS submitting raw
pain ratings into repeated-measures ANOVAs with the main
within-subject factors of Stimulation Session (active tDCS
over occipital cortex, active tDCS over centro-parietal
Kortex, sham tDCS), Time (pre- and post-tDCS), and Visual
Context (hand-view, object-view). Bonferroni correction
was used to adjust p values of post hoc tests.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
Ö
8
C
4
N
7
_
1
A
/
_
J
0
Ö
0
C
2
N
9
3
_
A
P
_
D
0
0
B
2
j
9
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
ERGEBNISSE
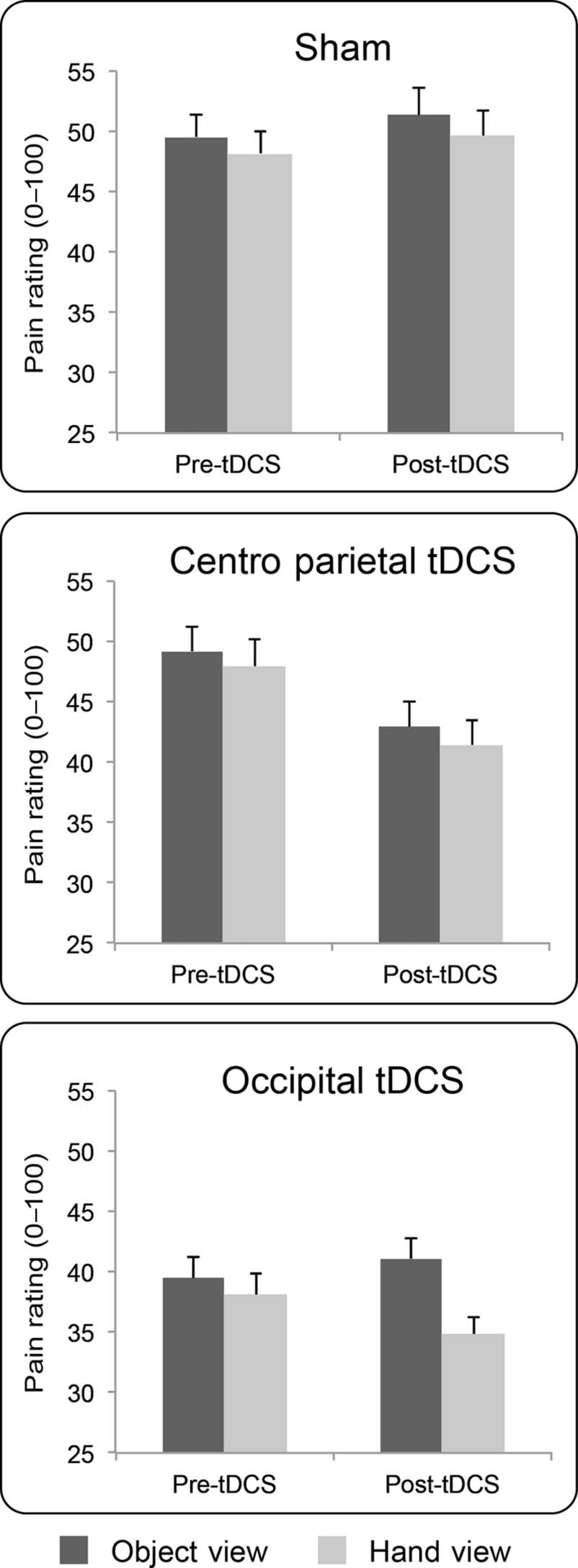
Experiment 1: Anodal/Excitatory tDCS
The results are shown in Figure 2. In the pre-tDCS sessions,
pain ratings were reduced in the hand-view condition in
comparison with the object-view condition, T(11) = 2.88,
P < .015, confirming visually induced analgesia for electri-
cal nociceptive stimulation.
Omnibus ANOVA on raw pain ratings showed a signifi-
cant main effect of Visual Context, F(1, 11) = 19.18, p <
.001, ηp
2 = 0.64: pain ratings were reduced in the hand-
view condition, in comparison with the object-view condi-
tion, replicating the finding that viewing oneʼs own body is
analgesic (Mancini, Longo, et al., 2011; Longo et al., 2009).
The main effects of Session, F(2, 22) = 2.89, p = .08,
Mancini et al.
2421
and session by time by context, F(2, 22) = 4.68, p < .02,
ηp
2 = 0.30, were all significant, suggesting that tDCS mod-
ulations were specific for the visual context and the session
(Figure 2).
We explored this significant three-way interaction by
submitting raw pain ratings to three 2 × 2 ANOVAs, one
for each session, with main factors of Context and Time.
Ratings in the sham session were not modulated by time
(F < 1). There was a near-significance main effect of
Context, F(1, 11) = 4.13, p = .067, ηp
2 = 0.27, and no
significant Time × Context interaction (F < 1).
For centro-parietal tDCS, the ANOVA revealed a signifi-
cant main effect of Time, F(1, 11) = 9.91, p = .009, ηp
2 =
0.47, because pain ratings decreased after anodal centro-
parietal tDCS (Figure 2). The main effect of Context, F(1,
11) = 3.23, p = .10, ηp
2 = 0.23, and the Time × Context
interaction (F < 1) were not significant.
For occipital tDCS, the main effect of Time was not sig-
nificant (F < 1), but there was a significant main effect of
Context, F(1, 11) = 22.27, p = .001, ηp
2 = 0.67, and a sig-
nificant Time × Context interaction, F(1, 11) = 10.78, p =
.007, ηp
2 = 0.49. This occurred because the difference in
ratings between view-object and view-hand contexts was
bigger (mean difference = 6.28, SEM = 1.5, p = .002) after
occipital tDCS, as compared with before stimulation (mean
difference = 1.35, SEM = 0.5; p = .046). In other words,
occipital tDCS enhanced the analgesic pain modulation
caused by viewing the hand (Figure 2).
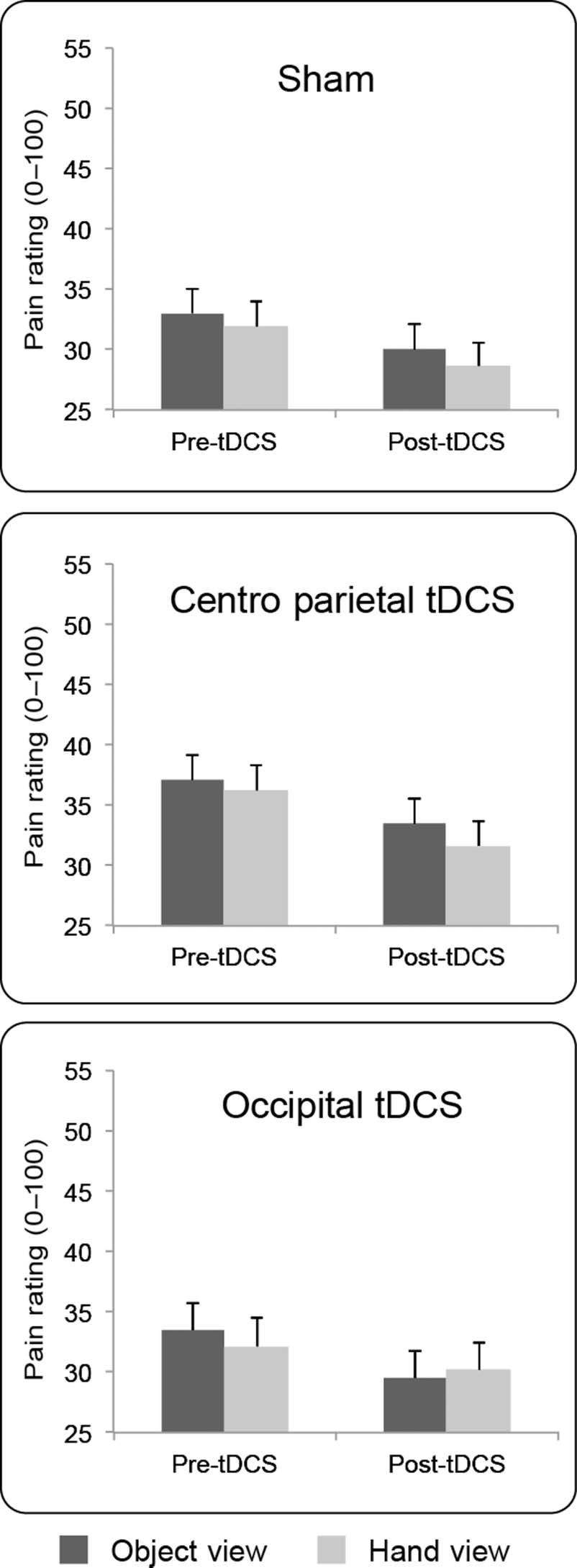
Experiment 2: Cathodal/Inhibitory tDCS
The results are shown in Figure 3. Visually induced
analgesia for electrical stimuli was again replicated in
the pre-tDCS sessions of Experiment 2, because pain rat-
ings were reduced during vision of the hand in compar-
ison with the object, t(11) = 2.22, p < .048.
Omnibus ANOVA on raw pain ratings showed a signifi-
cant main effect of Context, F(1, 11) = 8.15, p < .02, ηp
2 =
0.43, indicating a visually induced analgesia. The main
effect of Session, F(2, 22) = 1.12, p = .34, ηp
2 = 0.09,
was not significant, but a significant effect of Time was
found, F(1, 11) = 5.19, p < .04, ηp
2 = 0.32. The inter-
actions were not significant (Context × Session: F(2, 22) =
1.43, p = .26, ηp
2 = 0.11; Time × Context: F < 1; Session ×
Time: F < 1; Session × Time × Context: F(2, 22) = 2.04,
p = .15, ηp
2 = 0.16). Therefore, cathodal stimulation of the
occipital and centro-parietal cortices did not modulate pain
levels differently from sham. There was a significant pain
reduction in the poststimulation session in comparison
with the prestimulation session, independently of the
tDCS session, possibly because of nociceptor habituation
(Milne, Kay, & Irwin, 1991).
DISCUSSION
Viewing oneʼs own body reduces the level of acute pain
caused by an electrocutaneous stimulus, relative to viewing
Figure 2. Experiment 1. Anodal stimulation: Group results. Mean
(± SEM ) pain intensity ratings (0–100) by stimulation session, visual
context, and time.
ηp
2 = 0.21, and of Time (F < 1) were not significant, nor
was their interaction, F(2, 22) = 2.26, p = .13, ηp
2 =
0.17, indicating that anodal tDCS over either the contra-
lateral occipital or centro-parietal cortex did not modu-
late overall pain levels. Importantly, the interactions of
session by context, F(2, 22) = 3.62, p < .04, ηp
2 = 0.25,
time by context, F(1, 11) = 6.01, p < .03, ηp
2 = 0.35,
2422
Journal of Cognitive Neuroscience
Volume 24, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
o
8
c
4
n
7
_
1
a
/
_
j
0
o
0
c
2
n
9
3
_
a
p
_
d
0
0
b
2
y
9
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
enhanced the analgesic effect of viewing the body. This ef-
fect is specific for stimulation site, because it was not found
for anodal stimulation of the centro-parietal cortex. Anodal
centro-parietal stimulation produced an overall reduction
in pain levels, but no interaction with visual context. The
occipital tDCS effect is also specific for the polarity of
stimulation, because cathodal tDCS on either occipital or
centro-parietal cortex did not differ from sham.
Extrastriate Visual Cortex
The finding that viewing the body reduces acute pain
may seem counterintuitive, given that acute pain is often
associated with the sight of stimuli threatening the body
and given that attention to threatening stimuli increases
pain, rather than reducing it (e.g., Hofle, Hauck, Engel, &
Senkowski, 2012; Van Damme, Legrain, Vogt, & Crombez,
2010; Legrain, Guerit, Bruyer, & Plaghki, 2002). Impor-
tantly, however, in our study, participants viewed their
body but did not see any visual event corresponding to
the painful stimulus. The painful electrode shocks in our
experiment were entirely invisible. The mechanisms un-
derlying inhibitory pain modulation from viewing the body
may differ from those involved in facilitatory pain modula-
tion from viewing nociceptive or potentially nociceptive
stimulation events, such as threatening objects approach-
ing the body. In particular, the latter process involves ex-
pectation of pain, which can be triggered either by viewing
a threatening stimulus (Hofle et al., 2012) or by almost any
other stimulus that has previously been associated with
pain (Atlas, Bolger, Lindquist, & Wager, 2010; Koyama,
McHaffie, Laurienti, & Coghill, 2005).
Our results suggest that pain reduction induced by
viewing the body may involve an inhibitory interaction
between visual areas and pain networks. Interestingly,
Longo et al. (2012) have recently shown that visual analge-
sia for laser pain is associated with increased effective con-
nectivity between areas activated when viewing the body
and areas activated by noxious stimuli. Our result is
consistent with theirs and further shows that this visual-
nociceptive coupling plays a causal role in visual analgesia.
Other examples of visual-somatosensory links have been
suggested from behavioral experiments. For example,
viewing the body enhances tactile spatial acuity (Kennett,
Taylor-Clarke, & Haggard, 2001) and vibrotactile ampli-
tude discrimination but reduces vibrotactile detection
(Harris, Arabzadeh, Moore, & Clifford, 2007).
Moreover, the effect is specifically triggered by viewing
the body, because occipital tDCS does not modulate
overall pain levels, but only the analgesic effect of viewing
the body. We suggest that visual areas specific for repre-
senting the body are likely to mediate these effects. The
spatial resolution of tDCS is relatively poor, and we did not
assess the precise location of body-specific visual areas in
our participants. Therefore, this conclusion remains tenta-
tive. However, extensive neuroimaging evidence shows
that occipital-temporal areas, including EBA (Downing
Mancini et al.
2423
Figure 3. Experiment 2. Cathodal stimulation: Group results. Mean
(± SEM ) pain intensity ratings (0–100) by stimulation session, visual
context, and time.
an object. This replicates previous findings obtained using
different nociceptive stimuli (radiant heat in Longo et al.,
2009; contact heat in Mancini, Longo, et al., 2011). The pre-
sent novel finding is that visually induced analgesia can be
modulated by the level of excitability in the contralateral
extrastriate occipital cortex. Specifically, experimentally in-
creasing excitability of this area by means of anodal tDCS
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
o
8
c
4
n
7
_
1
a
/
_
j
0
o
0
c
2
n
9
3
_
a
p
_
d
0
0
b
2
y
9
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
et al., 2001) and the fusiform body area (Peelen & Downing,
2005), are preferentially activated by the view of bodies and
body parts (Orlov, Makin, & Zohary, 2010). Anodal tDCS
may boost neural responses to viewing the hand in these
body-specific visual areas, producing a neural drive that
inhibits processing in pain networks. This connection is
unlikely to be direct. Rather, visual information about the
body may reach the multisensory areas of the posterior
parietal cortex (Vallar & Maravita, 2009). These areas may
then modulate the somatosensory regions of the pain
matrix, including the somatosensory and insular cortices
(Longo et al., 2012).
Interestingly, the analgesic effect of viewing the body
may require recognition of personal identity. A previous
study found no modulation of pain ratings and laser-evoked
potentials when viewing the hand of another person (Longo
et al., 2009). It is still controversial whether activity in EBA
varies with the identity of the body that is viewed and
whether this region might contribute to discriminating the
self from the other (Hodzic, Muckli, Singer, & Stirn, 2009;
Myers & Sowden, 2008; Saxe, Jamal, & Powell, 2006). Lamm
and Decety (2008) did not find evidence for EBA involve-
ment in empathy for pain in others. Recognition of identity
seems also to involve other multisensory associative regions
in the posterior parietal cortex, including the inferior parietal
lobule and the inferior parietal sulcus (Hodzic, Kaas, Muckli,
Stirn, & Singer, 2009). These regions are connected with
visual body areas and are also associated with visually in-
duced analgesia (Longo et al., 2012).
It is worth noting that EBA itself may be a multisensory
area, because it is activated by haptic and visual body per-
ception (Kitada, Johnsrude, Kochiyama, & Lederman,
2009) and by motor commands (Astafiev, Stanley, Shulman,
& Corbetta, 2004). This suggests that the body represen-
tations formed in EBA might be partly multisensory, as
indeed are representations of nonbody objects in other
visual areas (Mancini, Bolognini, Bricolo, & Vallar, 2011;
Lacey, Tal, Amedi, & Sathian, 2009).
Cathodal stimulation of the occipital cortex did not mod-
ulate visual analgesia. The analgesic effect induced by the
view of the body was resistant to excitability-reducing
cathodal tDCS. Sensorimotor effects of tDCS are frequently
limited to one polarity of stimulation (for a recent review,
see Vallar & Bolognini, 2011). This may be because of addi-
tional factors such as orientation of the electric field (e.g.,
Nitsche & Paulus, 2000) and the background level of activ-
ity in the system when tDCS is applied. As a result, some
features of task-related activation may interact with the
physiological state of the cortex and polarity of tDCS stimu-
lation (Vallar & Bolognini, 2011; Antal & Paulus, 2008; Antal
et al., 2004). Further experiments are required to address
the potential role of these additional factors with respect to
the absence of effects of cathodal stimulation.
Finally, it is worth mentioning that tDCS may even alter
the activity of the areas below the reference electrode, in
this case, the supraorbital region. This area has many con-
nections with the other brain structures that are involved
in pain modulation (Mendonca et al., 2011; Boggio, Zaghi,
Lopes, & Fregni, 2008). The fact that in this study the two
sites of anodal tDCS (occipital and centro-parietal) had
different effects on perception, despite the same cathodal
position, argues against a significant effect of the fronto-
polar return current. Instead, our results suggest that the
critical factor for influencing neural activity with tDCS is
anodal stimulation, in this case, over occipital areas.
Centro-parietal Cortex
Our study also demonstrates that the tDCS effects on
visual modulation of pain are specific for anodal stimu-
lation of the occipital cortex. Anodal stimulation of the
centro-parietal cortex reduced pain levels independently
of the visual condition, whereas cathodal stimulation
over any site failed to modulate pain levels. Previous
evidence suggests that centro-parietal stimulation might
influence activity in relevant areas of the pain matrix, in-
cluding somatosensory cortex (Valentini et al., 2012; Liang
et al., 2011).
The current literature does not provide a clear view of
the modulation of pain by centro-parietal stimulation. A
previous report shows that cathodal, but not anodal, stim-
ulation of contralateral primary somatosensory cortex (SI)
can reduce both pain ratings and the N2 component of
nociceptive laser evoked potentials (Antal et al., 2008).
On the other hand, Grundmann et al. (2011) report that
neither anodal nor cathodal tDCS over contralateral SI
modulate pain thresholds, whereas cathodal tDCS over
the same region increases cold and warm detection
thresholds. Finally, there is also evidence of analgesic ef-
fects of cathodal (Csifcsak et al., 2009; Terney et al., 2008)
and anodal (Boggio et al., 2008) stimulation of the contra-
lateral primary motor cortex in healthy participants (for
reviews, see Vallar & Bolognini, 2011; Lefaucheur et al.,
2008). Reduced pain perception is mostly associated with
anodal stimulation of the primary motor cortex, whereas
the effects of tDCS stimulation of SI remain unclear both
in healthy participants (Grundmann et al., 2011; Antal
et al., 2008) and in patients (Tracey, 2011; Zaghi, Thiele,
Pimentel, Pimentel, & Fregni, 2011; Zaghi, Heine, & Fregni,
2009; Lima & Fregni, 2008).
Our finding of analgesia after anodal centro-parietal
tDCS is in line with previous evidence, but several caveats
need to be kept in mind. In the experiment with cathodal
tDCS, conducted on a different group of participants, pain
levels were reduced after every tDCS session, including
sham (but independently of the visual context). The sim-
plest explanation is pain habituation (Milne et al., 1991);
however, it is also possible that weak inhibitory effects
of cathodal tDCS have been masked by pain habituation
mechanisms. Likewise, also the analgesia induced by
anodal tDCS over the centro-parietal cortex may include
an element of pain habituation independent of specific
brain stimulation and needs to be confirmed in future
studies.
2424
Journal of Cognitive Neuroscience
Volume 24, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
o
8
c
4
n
7
_
1
a
/
_
j
0
o
0
c
2
n
9
3
_
a
p
_
d
0
0
b
2
y
9
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Several methodological and anatomical factors should
also be considered in interpreting our centro-parietal re-
sults. First, our current intensity and tDCS duration were
different from those of some previous studies (e.g., 1 mA
for 15 min: Antal et al., 2008; 2 mA for min in our study).
Electrode placement may also be important (e.g., Mendonca
et al., 2011), because tDCS effects on pain may be not
based on modulations of focal activity, but on connectiv-
ity changes (Dieckhofer et al., 2006; Matsunaga, Nitsche,
Tsuji, & Rothwell, 2004). Furthermore, no single primary
nociceptive- or pain-specific cortex has been found so
far, and the specific role of centro-parietal areas in coding
pain levels is not fully clear (Oertel et al., 2011; Iannetti &
Mouraux, 2010). Finally, different types of nociceptive
stimulation and different pain measures (e.g., thresholds
vs. ratings) have been used in previous studies, complicat-
ing comparisons between different tDCS results.
Conclusion
To conclude, we show that multisensory interactions can
be facilitated by anodal occipital tDCS. In particular, cortical
excitability shifts induced by tDCS can modulate visual–
nociceptive interactions, enhancing visual analgesia. tDCS
is becoming popular clinically in the treatment of neuro-
pathic and chronic pain (Brunoni et al., 2012; Zaghi
et al., 2009; Lefaucheur et al., 2008; Fregni, Freedman,
& Pascual-Leone, 2007). Mounting evidence suggests
that other sensory modalities including touch (Drew &
MacDermott, 2009; Inui, Tsuji, & Kakigi, 2006) and vision
(Mancini, Longo, et al., 2011) can significantly modulate
pain. The present findings might motivate research on
tDCS pain therapies beyond the range of nociceptive brain
regions currently targeted by tDCS.
Acknowledgments
F. M. was supported by a doctoral program of the University of
Milano-Bicocca. P. H. was supported by a Research Fellowship
from the Leverhulme Trust and by EU FP7 project VERE. N. B.
and G. V. were supported by FAR grants from the University of
Milano-Bicocca, Milano, Italy. G. V. was supported by Ricerca
Corrente grants from the Istituto Auxologico Italiano, Milano, Italy.
Reprint requests should be sent to Flavia Mancini, Institute
of Cognitive Neuroscience, University College London, London
WC1N 3AR, United Kingdom, or via e-mail: f.mancini@ucl.ac.uk.
REFERENCES
Antal, A., Brepohl, N., Poreisz, C., Boros, K., Csifcsak, G., &
Paulus, W. (2008). Transcranial direct current stimulation
over somatosensory cortex decreases experimentally induced
acute pain perception. Clinical Journal of Pain, 24, 56–63.
Antal, A., Nitsche, M. A., Kruse, W., Kincses, T. Z., Hoffmann,
K. P., & Paulus, W. (2004). Direct current stimulation over
V5 enhances visuomotor coordination by improving motion
perception in humans. Journal of Cognitive Neuroscience,
16, 521–527.
Antal, A., Nitsche, M. A., & Paulus, W. (2001). External modulation
of visual perception in humans. NeuroReport, 12, 3553–3555.
Antal, A., & Paulus, W. (2008). Transcranial direct current
stimulation and visual perception. Perception, 37, 367–374.
Astafiev, S. V., Stanley, C. M., Shulman, G. L., & Corbetta, M.
(2004). Extrastriate body area in human occipital cortex
responds to the performance of motor actions. Nature
Neuroscience, 7, 542–548.
Atlas, L. Y., Bolger, N., Lindquist, M. A., & Wager, T. D. (2010).
Brain mediators of predictive cue effects on perceived pain.
Journal of Neuroscience, 30, 12964–12977.
Boggio, P. S., Fregni, F., Valasek, C., Ellwood, S., Chi, R., Gallate, J.,
et al. (2009). Temporal lobe cortical electrical stimulation
during the encoding and retrieval phase reduces false
memories. PLoS One, 4, e4959.
Boggio, P. S., Zaghi, S., Lopes, M., & Fregni, F. (2008).
Modulatory effects of anodal transcranial direct current
stimulation on perception and pain thresholds in healthy
volunteers. European Journal of Neurology, 15,
1124–1130.
Bolognini, N., Fregni, F., Casati, C., Olgiati, E., & Vallar, G.
(2010). Brain polarization of parietal cortex augments
training-induced improvement of visual exploratory and
attentional skills. Brain Research, 1349, 76–89.
Bolognini, N., Olgiati, E., Rossetti, A., & Maravita, A. (2010).
Enhancing multisensory spatial orienting by brain
polarization of the parietal cortex. European Journal of
Neuroscience, 31, 1800–1806.
Bolognini, N., Rossetti, A., Casati, C., Mancini, F., & Vallar, G.
(2011). Neuromodulation of multisensory perception:
A tDCS study of the sound-induced flash illusion.
Neuropsychologia, 49, 231–237.
Brunoni, A. R., Nitsche, M. A., Bolognini, N., Bikson, M.,
Wagner, T., & Merabet, L. (2012). Clinical research with
transcranial direct current stimulation (tDCS): Challenges
and future directions. Brain Stimulation, 5, 175–195.
Csifcsak, G., Antal, A., Hillers, F., Levold, M., Bachmann, C. G.,
Happe, S., et al. (2009). Modulatory effects of transcranial
direct current stimulation on laser-evoked potentials.
Pain Medicine, 10, 122–132.
Dasilva, A. F., Volz, M. S., Bikson, M., & Fregni, F. (2011).
Electrode positioning and montage in transcranial direct
current stimulation. Journal of Visualized Experiments,
23, 2744.
de Tommaso, M., Santostasi, R., Devitofrancesco, V., Franco, G.,
Vecchio, E., Delussi, M., et al. (2011). A comparative study
of cortical responses evoked by transcutaneous electrical vs
CO(2) laser stimulation. Clinical Neurophysiology, 122,
2482–2487.
Dieckhofer, A., Waberski, T. D., Nitsche, M., Paulus, W.,
Buchner, H., & Gobbele, R. (2006). Transcranial direct
current stimulation applied over the somatosensory
cortex—Differential effect on low and high frequency
SEPs. Clinical Neurophysiology, 117, 2221–2227.
Downing, P. E., Jiang, Y., Shuman, M., & Kanwisher, N. (2001).
A cortical area selective for visual processing of the human
body. Science, 293, 2470–2473.
Drew, L. J., & MacDermott, A. B. (2009). Neuroscience:
Unbearable lightness of touch. Nature, 462, 580–581.
Fregni, F., Freedman, S., & Pascual-Leone, A. (2007). Recent
advances in the treatment of chronic pain with non-invasive
brain stimulation techniques. Lancet Neurology, 6,
188–191.
Gandiga, P. C., Hummel, F. C., & Cohen, L. G. (2006).
Transcranial DC stimulation (tDCS): A tool for double-blind
sham-controlled clinical studies in brain stimulation. Clinical
Neurophysiology, 117, 845–850.
Geyer, S., Schleicher, A., & Zilles, K. (1999). Areas 3a, 3b, and 1
of human primary somatosensory cortex. Neuroimage, 10,
63–83.
Mancini et al.
2425
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
o
8
c
4
n
7
_
1
a
/
_
j
0
o
0
c
2
n
9
3
_
a
p
_
d
0
0
b
2
y
9
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Grundmann, L., Rolke, R., Nitsche, M. A., Pavlakovic, G., Happe, S.,
Treede, R. D., et al. (2011). Effects of transcranial direct current
stimulation of the primary sensory cortex on somatosensory
perception. Brain Stimulation, 4, 253–260.
Harris, J. A., Arabzadeh, E., Moore, C. A., & Clifford, C. W.
(2007). Noninformative vision causes adaptive changes in
tactile sensitivity. Journal of Neuroscience, 27, 7136–7140.
Hodzic, A., Kaas, A., Muckli, L., Stirn, A., & Singer, W. (2009).
Distinct cortical networks for the detection and identification
of human body. Neuroimage, 45, 1264–1271.
Hodzic, A., Muckli, L., Singer, W., & Stirn, A. (2009). Cortical
responses to self and others. Human Brain Mapping, 30,
951–962.
Hofle, M., Hauck, M., Engel, A. K., & Senkowski, D. (2012).
Viewing a needle pricking a hand that you perceive as yours
enhances unpleasantness of pain. Pain, 153, 1074–1081.
Iannetti, G. D., & Mouraux, A. (2010). From the neuromatrix to
the pain matrix (and back). Experimental Brain Research,
205, 1–12.
Inui, K., Tsuji, T., & Kakigi, R. (2006). Temporal analysis of
cortical mechanisms for pain relief by tactile stimuli in
humans. Cerebral Cortex, 16, 355–365.
Katsarava, Z., Ayzenberg, I., Sack, F., Limmroth, V., Diener, H. C.,
& Kaube, H. (2006). A novel method of eliciting pain-related
potentials by transcutaneous electrical stimulation. Headache,
46, 1511–1517.
Katsarava, Z., Ellrich, J., Diener, H. C., & Kaube, H. (2002).
Optimized stimulation and recording parameters of human
“nociception specific” blink reflex recordings. Clinical
Neurophysiology, 113, 1932–1936.
Kaube, H., Katsarava, Z., Kaufer, T., Diener, H., & Ellrich, J.
(2000). A new method to increase nociception specificity
of the human blink reflex. Clinical Neurophysiology, 111,
413–416.
Kennett, S., Taylor-Clarke, M., & Haggard, P. (2001).
Noninformative vision improves the spatial resolution of
touch in humans. Current Biology, 11, 1188–1191.
Kitada, R., Johnsrude, I. S., Kochiyama, T., & Lederman, S. J.
(2009). Functional specialization and convergence in the
occipito-temporal cortex supporting haptic and visual
identification of human faces and body parts: An fMRI study.
Journal of Cognitive Neuroscience, 21, 2027–2045.
Koyama, T., McHaffie, J. G., Laurienti, P. J., & Coghill, R. C. (2005).
The subjective experience of pain: Where expectations become
reality. Proceedings of the National Academy of Sciences,
U.S.A., 102, 12950–12955.
Lacey, S., Tal, N., Amedi, A., & Sathian, K. (2009). A putative
model of multisensory object representation. Brain
Topography, 21, 269–274.
Lamm, C., & Decety, J. (2008). Is the extrastriate body area
(EBA) sensitive to the perception of pain in others?
Cerebral Cortex, 18, 2369–2373.
Lefaucheur, J. P., Antal, A., Ahdab, R., Ciampi de Andrade, D.,
Fregni, F., Khedr, E. M., et al. (2008). The use of repetitive
transcranial magnetic stimulation (rTMS) and transcranial
direct current stimulation (tDCS) to relieve pain. Brain
Stimulation, 1, 337–344.
Lima, M. C., & Fregni, F. (2008). Motor cortex stimulation for
chronic pain: Systematic review and meta-analysis of the
literature. Neurology, 70, 2329–2337.
Longo, M. R., Betti, V., Aglioti, S. M., & Haggard, P. (2009).
Visually induced analgesia: Seeing the body reduces pain.
Journal of Neuroscience, 29, 12125–12130.
Longo, M. R., Iannetti, G. D., Mancini, F., Driver, J., & Haggard, P.
(2012). Linking pain and the body: Neural correlates of
visually induced analgesia. Journal of Neuroscience, 32,
2601–2607.
Mancini, F., Bolognini, N., Bricolo, E., & Vallar, G. (2011).
Cross-modal processing in the occipito-temporal cortex:
A TMS study of the Muller-Lyer illusion. Journal of
Cognitive Neuroscience, 23, 1987–1997.
Mancini, F., Longo, M. R., Kammers, M. P., & Haggard, P.
(2011). Visual distortion of body size modulates pain
perception. Psychological Science, 22, 325–330.
Matsunaga, K., Nitsche, M. A., Tsuji, S., & Rothwell, J. C. (2004).
Effect of transcranial DC sensorimotor cortex stimulation on
somatosensory evoked potentials in humans. Clinical
Neurophysiology, 115, 456–460.
Mendonca, M. E., Santana, M. B., Baptista, A. F., Datta, A.,
Bikson, M., Fregni, F., et al. (2011). Transcranial DC
stimulation in fibromyalgia: Optimized cortical target
supported by high-resolution computational models.
Journal of Pain, 12, 610–617.
Milne, R. J., Kay, N. E., & Irwin, R. J. (1991). Habituation
to repeated painful and non-painful cutaneous stimuli:
A quantitative psychophysical study. Experimental
Brain Research, 87, 438–444.
Myers, A., & Sowden, P. T. (2008). Your hand or mine? The
extrastriate body area. Neuroimage, 42, 1669–1677.
Nitsche, M. A., Liebetanz, D., Antal, A., Lang, N., Tergau, F., &
Paulus, W. (2003). Modulation of cortical excitability by weak
direct current stimulation-technical, safety and functional
aspects. Supplements to Clinical Neurophysiology, 56,
255–276.
Nitsche, M. A., & Paulus, W. (2000). Excitability changes
induced in the human motor cortex by weak transcranial
direct current stimulation. Journal of Physiology, 527,
633–639.
Nitsche, M. A., & Paulus, W. (2001). Sustained excitability
elevations induced by transcranial DC motor cortex
stimulation in humans. Neurology, 57, 1899–1901.
Oertel, B. G., Preibisch, C., Martin, T., Walter, C., Gamer, M.,
Deichmann, R., et al. (2011). Separating brain processing
of pain from that of stimulus intensity. Human Brain
Mapping, 33, 883–894.
Ohnhaus, E. E., & Adler, R. (1975). Methodological problems in
the measurement of pain: A comparison between the verbal
rating scale and the visual analogue scale. Pain, 1, 379–384.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologia,
9, 97–113.
Orlov, T., Makin, T. R., & Zohary, E. (2010). Topographic
representation of the human body in the occipitotemporal
cortex. Neuron, 68, 586–600.
Legrain, V., Guerit, J. M., Bruyer, R., & Plaghki, L. (2002).
Overduin, S. A., & Servos, P. (2004). Distributed digit
Attentional modulation of the nociceptive processing into
the human brain: Selective spatial attention, probability
of stimulus occurrence, and target detection effects on
laser evoked potentials. Pain, 99, 21–39.
Liang, M., Mouraux, A., & Iannetti, G. D. (2011). Parallel
processing of nociceptive and non-nociceptive
somatosensory information in the human primary and
secondary somatosensory cortices: Evidence from dynamic
causal modeling of functional magnetic resonance imaging
data. Journal of Neuroscience, 31, 8976–8985.
somatotopy in primary somatosensory cortex. Neuroimage,
23, 462–472.
Paulus, W. (2011). Transcranial electrical stimulation (tES - tDCS;
tRNS, tACS) methods. Neuropsychological Rehabilitation,
21, 602–617.
Peelen, M. V., & Downing, P. E. (2005). Selectivity for the
human body in the fusiform gyrus. Journal of Neurophysiology,
93, 603–608.
Polania, R., Nitsche, M. A., & Paulus, W. (2011). Modulating
functional connectivity patterns and topological functional
2426
Journal of Cognitive Neuroscience
Volume 24, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
o
8
c
4
n
7
_
1
a
/
_
j
0
o
0
c
2
n
9
3
_
a
p
_
d
0
0
b
2
y
9
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
organization of the human brain with transcranial direct
current stimulation. Human Brain Mapping, 32,
1236–1249.
Poreisz, C., Boros, K., Antal, A., & Paulus, W. (2007). Safety
aspects of transcranial direct current stimulation concerning
healthy subjects and patients. Brain Research Bulletin, 72,
208–214.
Ragert, P., Vandermeeren, Y., Camus, M., & Cohen, L. G. (2008).
Improvement of spatial tactile acuity by transcranial direct
current stimulation. Clinical Neurophysiology, 119,
805–811.
Rosier, E. M., Iadarola, M. J., & Coghill, R. C. (2002).
Reproducibility of pain measurement and pain perception.
Pain, 98, 205–216.
Saxe, R., Jamal, N., & Powell, L. (2006). My body or yours? The
effect of visual perspective on cortical body representations.
Cerebral Cortex, 16, 178–182.
Sparing, R., Thimm, M., Hesse, M. D., Kust, J., Karbe, H., & Fink,
G. R. (2009). Bidirectional alterations of interhemispheric
parietal balance by non-invasive cortical stimulation. Brain,
132, 3011–3020.
Terney, D., Bergmann, I., Poreisz, C., Chaieb, L., Boros, K.,
Nitsche, M. A., et al. (2008). Pergolide increases the efficacy
of cathodal direct current stimulation to reduce the amplitude
of laser-evoked potentials in humans. Journal of Pain and
Symptom Management, 36, 79–91.
Tracey, I. (2011). Can neuroimaging studies identify pain
endophenotypes in humans? Nature Reviews Neurology,
7, 173–181.
Valentini, E., Hu, L., Chakrabarti, B., Hu, Y., Aglioti, S. M., &
Iannetti, G. D. (2012). The primary somatosensory cortex
largely contributes to the early part of the cortical response
elicited by nociceptive stimuli. Neuroimage, 59, 1571–1581.
Vallar, G., & Bolognini, N. (2011). Behavioural facilitation
following brain stimulation: Implications for neurorehabilitation.
Neuropsychological Rehabilitation, 21, 618–649.
Vallar, G., & Maravita, A. (2009). Personal and extra-personal
spatial perception. In G. G. Berntson & J. T. Cacioppo (Eds.),
Handbook of neuroscience for the behavioral sciences.
( Vol. 1, pp. 322–336). New York: Wiley.
Van Damme, S., Legrain, V., Vogt, J., & Crombez, G. (2010).
Keeping pain in mind: A motivational account of attention to
pain. Neuroscience and Biobehavioral Reviews, 34, 204–213.
Wiech, K., Ploner, M., & Tracey, I. (2008). Neurocognitive
aspects of pain perception. Trends in Cognitive Sciences,
12, 306–313.
Yarnitsky, D., & Sprecher, E. (1994). Thermal testing: Normative
data and repeatability for various test algorithms. Journal of
Neurological Sciences, 125, 39–45.
Zaghi, S., Heine, N., & Fregni, F. (2009). Brain stimulation
for the treatment of pain: A review of costs, clinical effects,
and mechanisms of treatment for three different central
neuromodulatory approaches. Journal of Pain Management,
2, 339–352.
Zaghi, S., Thiele, B., Pimentel, D., Pimentel, T., & Fregni, F.
(2011). Assessment and treatment of pain with non-invasive
cortical stimulation. Restorative Neurology and Neuroscience,
29, 439–451.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
2
4
/
2
1
4
2
1
/
9
2
1
4
9
1
4
9
4
/
6
1
6
5
7
7
o
8
c
4
n
7
_
1
a
/
_
j
0
o
0
c
2
n
9
3
_
a
p
_
d
0
0
b
2
y
9
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Mancini et al.
2427