Sustained Attention and Spatial Attention Distinctly
Influence Long-term Memory Encoding
Megan T. deBettencourt
, 1, Stephanie D. Williams1,2,
Edward K. Vogel1, and Edward Awh1
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
7
4
8
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Abstrakt
■ Our attention is critically important for what we remember.
Prior measures of the relationship between attention and
Erinnerung, Jedoch, have largely treated “attention” as a mono-
lith. Hier, across three experiments, we provide evidence for
two dissociable aspects of attention that influence encoding into
long-term memory. Using spatial cues together with a sensitive
continuous report procedure, we find that long-term memory
response error is affected by both trial-by-trial fluctuations of
sustained attention and prioritization via covert spatial attention.
Außerdem, using multivariate analyses of EEG, we track both
sustained attention and spatial attention before stimulus onset.
Intriguingly, even during moments of low sustained attention,
there is no decline in the representation of the spatially attended
location, showing that these two aspects of attention have
robust but independent effects on long-term memory encoding.
Endlich, sustained and spatial attention predicted distinct vari-
ance in long-term memory performance across individuals.
Das ist, the relationship between attention and long-term mem-
ory suggests a composite model, wherein distinct attentional
subcomponents influence encoding into long-term memory.
These results point toward a taxonomy of the distinct attentional
processes that constrain our memories. ■
EINFÜHRUNG
In our daily lives, we fail to remember many of the items
that we encounter, largely because we were not paying suf-
ficient attention. Although attention and long-term mem-
ory are clearly intertwined (Decker & Duncan, 2020;
Hannula, 2018; Aly & Turk-Browne, 2017; Chun & Turk-
Browne, 2007), past work investigating the relationship
between attention and long-term memory often treats at-
tention as a monolithic cognitive construct. Jedoch, bei-
tention has numerous subcomponents, any one of which
could underlie the relationship with memory (Hakim,
Adam, Gunseli, Awh, & Vogel, 2019; Robison & Brewer,
2019; Chun, Golomb, & Turk-Browne, 2011; Poole &
Kane, 2009; Fan, McCandliss, Fossella, Flombaum, &
Posner, 2005). Zum Beispiel, sustained attention fluctuates
from trial to trial, and spatial attention can be oriented to
different locations in space. Jedoch, studies investigat-
ing long-term memory have generally lacked the ability
to disentangle whether memory failures reflect poor sus-
tained attention, misallocated spatial attention, oder beides.
This raises two potential explanations for the relationship
between attention and long-term memory: In a unified
model of attention and memory, memory failures could
be attributable to convergent failures of multiple forms
of attention. Das ist, lapses of sustained attention coincide
with moments when spatial attention is oriented to the
1Universität von Chicago, 2Boston University
© 2021 Massachusetts Institute of Technology
wrong location and together influence memory encoding.
Alternativ, in a composite model of attention and mem-
ory, memory failures arise from failures of any particular
attentional subcomponent. Das ist, lapses of sustained
attention and misoriented spatial attention could each
account for distinct failures of memory encoding. Daher,
the goal of this study was to determine whether sustained
attention and spatial attention exert overlapping or dis-
tinct influences on long-term memory.
Spatial attention and sustained attention are each
important for long-term memory encoding. Spatial atten-
tion has been traditionally studied using spatial cues, In
which participants orient spatial attention to a particular
location. The influence of spatial cues can be measured
behaviorally and via multivariate analyses of brain data
(Foster, Sutterer, Serences, Vogel, & Awh, 2017; Sprague
& Serences, 2013). Spatially cueing an item has further
been shown to improve long-term memory (Ziman, Lee,
Martinez, & Manning, 2019; LaRocque et al., 2015; Turk-
Browne, Golomb, & Chun, 2013; Uncapher, Hutchinson,
& Wagner, 2011). A largely distinct literature has studied
the influence of sustained attention on behavior, tradition-
ally using continuous performance tasks in which partici-
pants are required to repeatedly respond (deBettencourt,
Keene, Awh, & Vogel, 2019; Rosenberg, Noonan, DeGutis,
& Esterman, 2013). The hallmark signature of sustained
attention is that behavioral and brain systems fluctuate
between advantageous and disadvantageous states across
trials over time (Rosenberg et al., 2016; deBettencourt,
Zeitschrift für kognitive Neurowissenschaften 33:10, S. 2132–2148
https://doi.org/10.1162/jocn_a_01748
Cohen, Lee, Norman, & Turk-Browne, 2015; Esterman,
Noonan, Rosenberg, & DeGutis, 2013). Trial-by-trial
fluctuations of sustained attention have been shown to
influence which items will be later remembered
(deBettencourt, Norman, & Turk-Browne, 2018).
Außerdem, sustained attention and working memory
are closely linked cognitive constructs, although not syn-
onymous (Hakim, deBettencourt, Awh, & Vogel, 2020;
Adam & deBettencourt, 2019; deBettencourt et al., 2019;
Hakim et al., 2019). Extant work, Jedoch, has not exam-
ined whether these fluctuations of sustained attention
could be explained by trial-by-trial differences in the de-
ployment of spatial attention. Daher, the central goal of
the present work was to determine whether sustained
and spatial attention have related or distinct impacts on
long-term memory performance.
In this study, we examine whether sustained and spatial
attention exert distinct or common influences on long-term
memory encoding using behavioral and neural signatures.
To measure long-term memory behavior with high sensi-
tivity, we employed a continuous report task, in which
participants report a particular dimension of a stimulus
along a continuous space (Tompary, Zhou, & Davachi,
2020; Xie, Park, Zaghloul, & Zhang, 2020; Biderman,
Luria, Teodorescu, Hajaj, & Goshen-Gottstein, 2019; Fan
& Turk-Browne, 2016; Richter, Cooper, Bays, & Simons,
2016; Sutterer & Awh, 2016). To resolve the moment-by-
moment influence of attention on long-term memory, Wir
analyze multivariate EEG signals during time intervals
before encoding. We manipulate spatial attention via
spatial cues and observe the influence of sustained atten-
tion via trial-by-trial fluctuations of working memory per-
formance for spatially attended and unattended items. In
Experiment 1, we present a behavioral paradigm that
captures how sustained and spatial attention distinctly in-
fluence long-term location memory on a continuous
report task. In Experiment 2, we identify a multivariate
EEG signature of sustained attention that predicts long-
term memory performance independent of variations in
spatial attention. In Experiment 3, we extend these find-
ings to show that spatial and sustained attention influence
color memory. Endlich, collapsing across all studies, Wir
show that individual differences of the influence of sus-
tained and spatial attention predict unique variance in
long-term memory performance.
EXPERIMENT 1
The goal of this experiment was to examine whether sig-
natures of sustained and spatial attention predict long-
term memory. We hypothesized that long-term memory
would reflect trial-by-trial fluctuations of sustained atten-
tion, as well as the prioritization of cued stimuli by spatial
attention. We obtained a sensitive measure of long-term
memory accuracy by asking participants to report their
memory for the spatial location of trial-unique objects
using a continuous report task.
Methoden
Teilnehmer
In Experiments 1a and 1b, a combined 52 adults partici-
pated for University of Chicago course credit or $20 pay- ment ($10/Std). In all studies, we targeted data collection
aus 25 participants before exclusion. In Experiment 1a,
25 Erwachsene (15 weiblich, Durchschnittsalter = 23.2 Jahre) participated,
and in Experiment 1b, 27 adults participated (17 weiblich,
Durchschnittsalter = 24.2 Jahre). We excluded any participants
whose performance exceeded 3 SDs from the population
mean (n = 2 in Experiment 1a; n = 1 in Experiment 1b)
and participants who were outliers in terms of study com-
pletion (n = 2 in Experiment 1b completed 50% of the
study in the allotted time). daher, the final sample of
participants was 23 for Experiment 1a and 24 für
Experiment 1b. All participants in this experiment and
the following experiments reported normal or corrected-
to-normal color vision and provided informed consent to a
protocol approved by the University of Chicago institu-
tional review board.
Apparatus
Participants were seated facing an LCD monitor (120-Hz
refresh rate) in a testing room. In Experiment 1a, partic-
ipants were approximately 70 cm from the monitor, Und
in Experiment 1b, they were approximately 88 cm from
the monitor, because of a reconfiguration of the behav-
ioral testing rooms. Stimuli were presented in Python
using PsychoPy (Peirce, 2007).
Stimuli
Trial-unique real-world object pictures were presented
on a gray background (Brodeur, Guérard, & Bouras,
2014; Brady, Konkle, Alvarez, & Oliva, 2008). At encod-
ing, these images (subtending 3° visual angle) were pre-
sented along a light gray ring (at 5° eccentricity). A black
fixation dot (0.5°) appeared at the center of the screen.
Peripheral spatial cues (black dots, 0.5°) appeared along
the gray ring.
Verfahren
In Experiment 1a, on each working memory trial, a pe-
ripheral spatial cue briefly appeared (250 ms) along
the ring and participants were instructed to covertly
attend to the cued spatial location (Figure 1A). After an
extended prestimulus interval (2–4 Sek), four items (trial-
unique object pictures) briefly appeared along the ring
(250 ms). Participants were instructed to hold the
items in mind over a retention interval (2 Sek). Dann,
one of the items reappeared at the center, und das
mouse cursor was initialized to a random position along
the ring. Participants reported the original location of the
probed item by clicking along the ring with the mouse.
deBettencourt et al.
2133
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
7
4
8
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 1. Experimentelles Design.
(A) In the working memory
Phase, each trial was composed
of a brief peripheral spatial cue,
a blank prestimulus interval, A
memory array of four real-world
trial-unique object pictures, Und
a blank retention interval. An
valid trials (75%), the location of
the cue matched the location of
the probed item. On invalid
Versuche (25%), one of the other
three uncued items was probed.
During the working memory
probe, participants reported the
item’s original location by
clicking along the ring. (B) In
the long-term memory phase,
we measured absolute response
Fehler (0–180°) for cued, uncued,
and unprobed items.
On valid trials (75%), the location of the cue matched the
location of the probed item. On invalid trials (25%), eins
of the other three uncued items was probed. Nach dem
response, there was a blank intertrial interval (1–2 sec).
Participants were instructed to maintain central fixation
throughout the trial. Participants completed 16 blocks,
each consisting of 24 working memory trials, und das
cue position was counterbalanced across trials within a
block.
In the long-term memory phase, we examined memory
items that had appeared in the working memory phase. In
each block, these items could be probed (18 cued items
from valid trials, 6 uncued items from invalid trials) or un-
probed (6 items from invalid trials). By definition, Dort
was no measure of working memory performance for un-
probed items. A balanced number of new object pictures
appeared (30 per block); these items otherwise never
appeared during the experiment. Erste, participants com-
pleted a recognition memory rating for each item. Below
the image, a 4-point confidence rating scale appeared
below the image. When participants made their responses
(using the keys 1–4), it was briefly displayed on the scale
(0.5 Sek). Dann, for all old items, participants completed a
location memory continuous report. The fixation dot
turned white, and the participant was instructed to
retrieve the original location of that item for 1 Sek. Dann,
the fixation dot turned black, the mouse cursor appeared
randomly along the ring, and the participant reported the
original location for that object along the ring. After each
item, there was a blank intertrial interval (0.5 Sek).
In Experiment 1b, half of the blocks were identical to
Experiment 1a with a single cue that was either valid or
invalid (cued/uncued ). In the other half of the blocks,
the encoding arrays were preceded by four dots (neu-
tral ). These four dots indicated the location of each of
the items but provided no information about which item
was most likely to be tested. Als solche, this condition con-
trolled for the visual presentation of peripheral dots and
provided the same temporal information about an up-
coming memory array. The order of single cued blocks
and neutral cued blocks were randomized, such that
there were two of each type every four blocks.
Participants performed this task for 2 hr or until they
completed 16 blocks in total (384 Versuche). In Experiment 1a,
participants completed, on average, 345 (89.95 %) von
the maximum 384 Versuche, ranging from 240 Zu 384. In
Experiment 1b, participants completed 360 (93.75 %) von
the maximum 384 Versuche, ranging from 312 Zu 384.
Behavioral Analysis
Location memory was measured via continuous report
and analyzed as response error, or the angular difference
between the original minus reported location (−180° to
180°). We primarily conducted subsequent memory anal-
yses, by focusing our analyses on long-term location
Erinnerung. We examined whether long-term memory re-
flected differences in spatial attention (cued vs. uncued
Artikel) and differences in sustained attention (Betrieb-
alized as trial-by-trial fluctuations, for cued, uncued, Und
unprobed items). For recognition memory, hoch-
confidence old responses were treated as remembered
and all other responses as forgotten to calculate item rec-
ognition memory hit rate. Recognition memory perfor-
mance was summarized as a single nonparametric
measure of sensitivity (A
0).
Statistics
Descriptive statistics are reported as the mean and 95%
confidence interval (CI) of the bootstrapped distribution.
If the hypothesis was directional, one-sided tests were
durchgeführt. Trial-wise correlations were computed using
the Pearson correlation coefficient for each participant.
The reliability of the correlations was assessed across
Teilnehmer. Because some of the data violated the
2134
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 10
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
J
Ö
C
N
_
A
_
0
1
7
4
8
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
assumption of normality, nonparametric statistics were
performed by resampling participants with replacement
100,000 mal. The p value corresponds to the propor-
tion of the iterations in which the bootstrapped mean
was in the opposite direction (Efron & Tibshirani,
1986). Any p values smaller than one in 1000 were ap-
proximated as p < .001.
Results
Experiment 1a
We obtained a sensitive measure of long-term location
memory as the absolute response error from the original
location (dltm = 61.00°, 95% CIs [54.36, 66.83]). We con-
ducted a subsequent memory analysis, sorting trials ac-
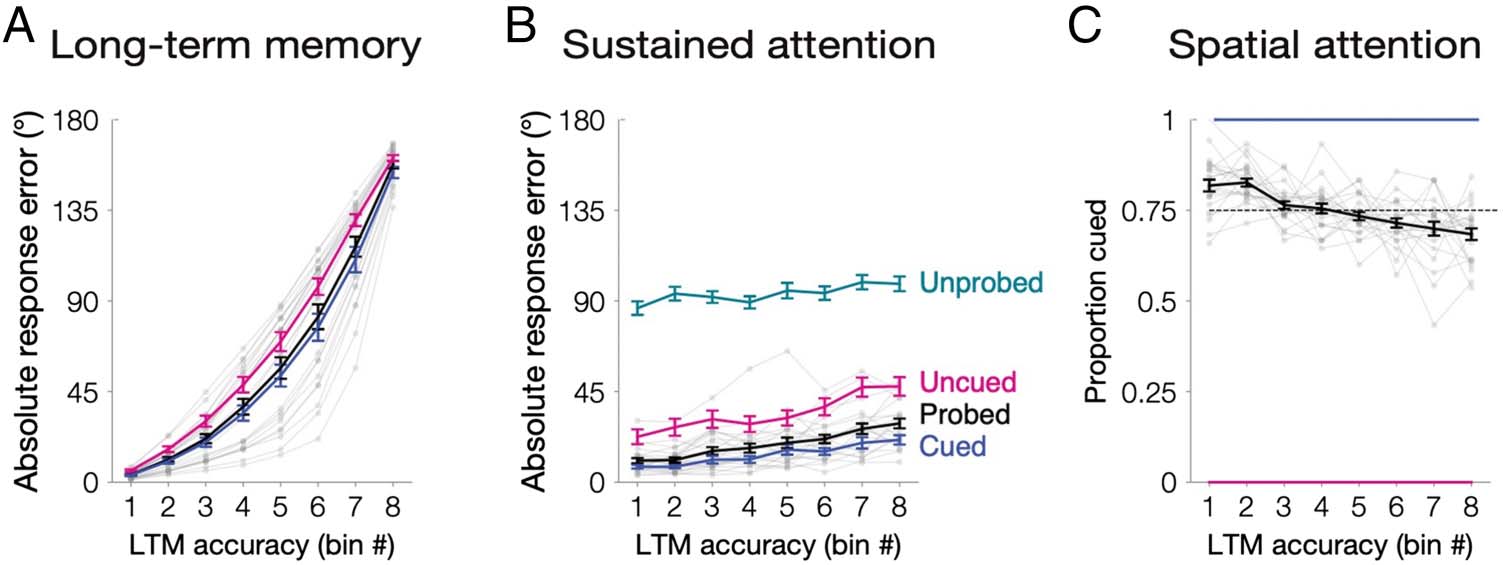
cording to long-term memory performance (Figure 2A).
We were interested in whether trial-by-trial fluctuations
of sustained attention influenced long-term memory per-
formance. We examined performance in the working
memory phase (dwm = 18.86°, 16.11, 23.17), as a more
proximal assay of attentional state. Across all probed trials,
working memory response error was reliably correlated
with long-term memory response error (r = .19, .15–.23,
one-tailed p < .001). To quantify this relationship, we cal-
culated a linear fit to relate long-term memory (bin num-
ber, 1–8) to working memory response error. We
observed a reliably positive slope relating long-term mem-
ory to working memory (m = 2.71, 2.11–3.46; one-tailed
p < .001; Figure 2B). This correlation between working
memory and long-term memory performance is consis-
tent with prior work that working memory serves as a
“gateway” that determines the bandwidth of encoding into
long-term memory (Fukuda & Vogel, 2019).
The correlation between working memory and long-
term memory could be explained in two distinct ways.
On the one hand, successful prioritization of the cued
item over the other items in the display (spatial atten-
tion) could influence long-term memory. On the other
hand, memory outcomes might reflect a broader fluctua-
tion of attentional state (sustained attention) that could
impact the quality of memory for all items, regardless of
whether they were spatially attended or not. We exam-
ined each of these factors in turn:
First, we examined whether long-term memory re-
flected differences in spatial attention. Spatial attention
benefitted long-term memory, as cued items were better
remembered (dcued = 57.95°, 50.69–64.40°; duncued =
70.14°, 65.03–74.78°; one-tailed p < .001). We also calcu-
lated a linear fit to relate long-term memory (bin number,
1–8) to the proportion of trials that had been cued in
each bin. Indeed, we observed a reliably negative slope
(m = −0.02, −.03 to −0.01; one-tailed p < .001;
Figure 2C). Thus, spatially attending to items enhanced
long-term memory fidelity.
To examine whether sustained attention shaped long-
term memory performance independently of spatial at-
tention, we separately analyzed cued and uncued items.
There were reliably positive correlations between trial-by-
trial fluctuations of working memory response error with
long-term memory, for cued items (r = .15, .12–.20,
one-tailed p < 001) and uncued items (r = .19, .13–.26,
one-tailed p < .001). We further quantified this relation-
ship as a positive slope between long-term memory bin
and working memory response error for cued items
and uncued items (mcued = 2.02, 1.49–2.78; muncued =
3.53, 2.24–4.72; one-tailed ps < .001; Figure 2B). This
revealed the influence of trial-by-trial fluctuations of
Figure 2. Experiment 1a
results. (A) Long-term memory
variability across trials. Location
memory absolute response
error (0–180°) was sorted and
binned into octiles within
participant for all probed items
(black line). To equate for
whether an item was cued to be
attended, we also repeated all
analyses separately for cued
items (blue) and uncued items
(pink). (B) Memory outcomes
reflect trial-by-trial fluctuations
of sustained attention.
Memory performance was
operationalized as absolute response error (0–180°) for items within each bin. For uncued (pink), probed (black), and cued (blue) items, absolute
response error was obtained during the working memory phase. Unprobed items (teal) were, by definition, not tested in the working memory phase.
Therefore, absolute response error for these items was obtained during the long-term memory phase (0–180°). The slope of each line is positive
across the bins ( ps < .001). (C) Memory outcomes reflect differences of spatial attention. In each bin, we calculated the proportion of trials that had
been cued. For all probed items (black), the proportion of trials that were cued decreases across the eight bins ( p < .001). The dashed line
represents the mean proportion cued (0.75). Cued items (blue) and uncued items (pink) controlled for whether an item was cued to be attended
(100% and 0% cued, respectively). Error bars depict the standard error of the mean. Data from each participant for all probed items are overlaid in
small gray dots connected with lines.
deBettencourt et al.
2135
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
sustained attention on long-term memory performance,
regardless of whether the items were spatially attended.
We further posited that sustained attention could
broadly impact memory for multiple items from the same
display, reflecting synchronous fluctuations among all
simultaneously displayed items. We generally measured
long-term memory for a single item per display, which is
unable to speak to these broader fluctuations. However,
on trials when the cue was invalid, we also measured
long-term location memory of the item that was initially
cued but unprobed during the working memory phase of
the task. As these items were unprobed, we did not obtain
a working memory response for these unprobed items. We
therefore examined whether the long-term memory for the
uncued item was correlated with long-term memory for the
unprobed item from the same display. For each participant,
we correlated the long-term memory response error for the
unprobed and uncued items from the same display. Across
participants, the correlation between unprobed and un-
cued items was reliably positive (r = .6, .03–.10, one-tailed
p < .001). We further quantified this relationship as a slope
relating the long-term memory bin for the uncued item to
long-term memory for the unprobed item (m = 1.49, 0.64–
2.49; one-tailed p < .001; Figure 2B). This provides evi-
dence that trial-by-trial fluctuations of sustained attention
have broad influences on long-term memory for multiple
items presented simultaneously.
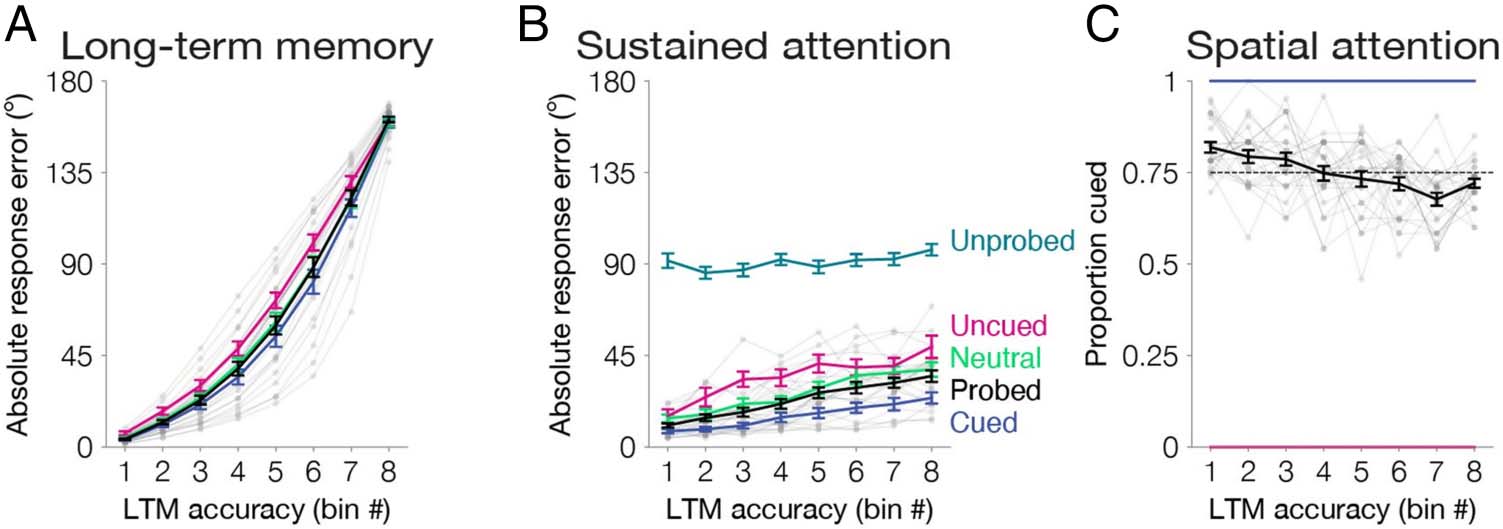
Experiment 1b
We conducted this experiment to replicate and extend
the previous findings. Half of the blocks were the same
as Experiment 1a (single cues, valid or invalid). In the
other half of the blocks (neutral blocks), all images were
preceded by four cues, which indicated the spatial loca-
tion of each item but provided no information about
which item was likely to be tested. We again operational-
ized performance as absolute response error in the long-
term memory phase (dltm = 63.72°, 58.43–68.52°) and in
the working memory phase (dwm = 23.22°, 19.79–
26.90°). We observed a reliably positive correlation be-
tween long-term memory and working memory response
error across all probed items (r = .19, .16–.23; one-tailed
p < .001). We further quantified this relationship as a
slope relating long-term memory bins to working memo-
ry response error (m = 3.54, 2.84–4.44; one-tailed p <
.001; Figure 3A and 3B). This replicated the finding of a
general correlation between working memory and long-
term memory performance.
We next examined whether this general correlation
reflected distinct contributions of spatial and sustained
attention. Indeed, spatial attention influenced long-term
memory: Neutrally cued items were remembered worse
than cued items (dcued = 60.39°, 54.60–65.78; dneutral =
64.32°, 58.66–69.27; one-tailed p < .001) and better than
uncued items (duncued = 71.12°, 66.87–75.82; one-tailed
p = .004). Moreover, we quantified the effect of spatial
attention on long-term memory, by repeating our bin-
ning analyses from Experiment 1a within the single cued
blocks. We replicated the finding from Experiment 1a
that the proportion cued decreased across bins (m =
−0.02, −0.01 to −0.02, one-tailed p < .001; Figure 3C).
These findings confirm the strong influence of spatial
attention on long-term memory.
We predicted that long-term memory would also reflect
trial-by-trial fluctuations of sustained attention, as
observed in Experiment 1a. First, we correlated long-
term memory and working memory response error for
all probed items within each condition (rcued = .18,
.14–.24; runcued = .21, .14–.29; rneutral = .23, .17–.28;
one-tailed ps < .001). Next, we calculated the slope that
related long-term memory (bin number, 1–8) to working
Figure 3. Experiment 1b
results. (A) Long-term location
memory variability binned
across trials. Absolute response
error (0–180°) was sorted and
binned into octiles within
participant for all probed items
(black line). To equate for
whether an item was cued to be
attended, we also repeated all
analyses separately for cued
items (blue), uncued items
(pink), and neutral items
(green). (B) Memory outcomes
reflect trial-by-trial fluctuations
of sustained attention. Memory performance was operationalized as absolute response error (0–180°) for items within each bin. For uncued (pink),
neutral (green), probed (black), and cued (blue) items, absolute response error was obtained during the working memory phase. Unprobed items
(teal) were not tested in the working memory phase. Therefore, absolute response error for unprobed items was obtained during the long-term
memory phase (0–180°). The slope of each line is positive across the bins ( ps < .05). (C) Memory outcomes reflect differences of spatial attention. In
each bin, we calculated the proportion of trials that had been cued. For all probed items (black), the proportion of trials that were cued decreases
across the eight bins ( p < .001). The dashed line represents the mean proportion cued (0.75). Cued items (blue) and uncued items (pink)
controlled for whether an item was cued to be attended (100% and 0% cued, respectively). Error bars depict the standard error of the mean.
Data from each participant for all probed items are overlaid in small gray dots connected with lines.
2136
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
memory response error (Figure 3B). We observed a reli-
ably positive slopes for cued, uncued, and neutral items
(mcued = 2.43, 1.74–3.29, muncued = 4.04, 2.91–5.29;
mneutral = 3.80, 2.94–4.90; one-tailed ps < .001). Finally,
we examined whether long-term memory was related
across items from the same display using the unprobed
items. Long-term memory was reliably correlated between
items from the same display (r = .05, 0.00–0.10, one-tailed
p = .03), which replicates our finding from Experiment 1a.
However, this correlation was reliable between the un-
cued and unprobed in the blocks with single cues (r =
.08, .02–.17, one-tailed p = .01), and not in the blocks with
neutral cues (r = .02, −.04–.08, one-tailed p = .26). We fur-
ther quantified this relationship between items from the
same display by calculating the slope across bins (m =
0.98, –0.15 to 2.22, one-tailed p = .05).
Recognition Memory
We designed this experiment to measure long-term loca-
tion memory via continuous report, in order to obtain a
sensitive measure of memory fidelity. However, we also
measured item recognition memory. Overall recognition
memory sensitivity was well above chance in Experiment 1
0 = 0.83; 0.80–0.85; one-tailed p < .001 vs. chance =
(A
0.5). We examined whether recognition memory corre-
lated with working memory performance. Working mem-
ory response error was lower for items that were later
recognized (drecog = 17.39, 15.61–19.42; dunrecog =
24.09, 21.29–27.22; one-tailed p < .001).
To examine the effect of spatial attention on long-term
item recognition memory, we calculated the proportion
of items that had been initially cued, separately for items
that were later recognized versus not. A greater propor-
tion of items that were later recognized were initially
cued (qrecog = 0.77, 0.76–0.78; qunrecog = 0.72, 0.71–
0.73; one-tailed p < 001).
To examine the effect of trial-by-trial fluctuations of sus-
tained attention on long-term item recognition memory,
we calculated the working memory response error for
items that were later recognized versus not. Working
memory response error was lower for items that were later
recognized, for cued items (drecog = 11.83, 10.40–13.45;
dunrecog = 17.04, 14.63–19.83; one-tailed p < .001) and un-
cued items (drecog = 31.16, 27.40–35.81; dunrecog = 35.90,
31.16–41.29; one-tailed p = .006). We also observed evi-
dence that recognition memory was correlated between
items from the same display: The memory hit rate for un-
probed items was higher when the item from the same dis-
play was recognized (hrecog = 0.18, 0.15–0.20; hunrecog =
0.13, 0.11–0.16; one-tailed p < .001). In summary, recog-
nition memory corroborated the findings from long-term
location memory obtained via continuous report.
Discussion
attention, on the encoding of visual information into
long-term memory. Long-term memory reflected whether
participants were spatially attending the memoranda, as
cued items were better remembered. We also found
evidence for strong trial-by-trial fluctuations of sustained
attention that influenced both spatially attended and spa-
tially unattended items. We also observed that long-term
memory was correlated for cued and uncued items with-
in the same display, which argues against spatial attention
as the source of these trial-to-trial fluctuations. Thus, this
experiment provided preliminary evidence for the com-
posite model of attention and long-term memory.
However, these behavioral results alone cannot speak
to whether these results are a function of stimulus encod-
ing (e.g., increased pattern separation of the memory array
items) or retrieval (e.g., stronger reinstatement of the
memory array item during the probe). Therefore, in
Experiment 2, we extended these findings by measuring
EEG activity while participants performed a similar task.
This provided the opportunity to identify neural signals
that tracked fluctuations in sustained attention and the
current locus of covert spatial attention. To anticipate
the findings, differences in sustained attention were de-
tectable based on neural activity even before stimulus on-
set, and this appears to be separate from fluctuations in
the quality of spatial orienting of attention.
EXPERIMENT 2
The goal of this experiment was to characterize the neural
signals of sustained and spatial attention that predict long-
term memory. We collected eye-tracking and EEG data
while participants performed the task from Experiment 1a.
Methods
Participants
Forty-two adults (23 female; mean = 23.5 years) completed
Experiment 2 for $60 payment ($15/hr). A larger number of
participants were chosen so as to have an adequate sample
size after excluding participants who had excessive EEG or
eye artifacts (six participants who all had fewer than half of
the trials remaining after artifact rejection of the prestimu-
lus period) or problems with EEG or eye-tracking equip-
ment during the recording session (six participants). The
final sample size was 30 participants. These exclusion cri-
teria were determined a priori and are consistent with
prior studies from our laboratory.
Apparatus
Participants were seated approximately 75 cm from an
LCD monitor (120-Hz refresh rate) in a shielded booth.
This experiment demonstrates a robust influence of two
distinct attentional factors, spatial attention and sustained
Stimuli and Procedure
Stimuli and procedure are the same as in Experiment 1a.
deBettencourt et al.
2137
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Eye Tracking
We monitored gaze position using a desk-mounted infra-
red eye-tracking system (EyeLink 1000 Plus, SR
Research). Gaze position was sampled at 1000 Hz, and
head position was stabilized with a chin rest.
EEG Recording
We recorded EEG activity using 30 active Ag/AgCl elec-
trodes mounted in an elastic cap (Brain Products
actiCHamp). We recorded from International 10–20 sites:
FP1, FP2, F3, F4, F7, F8, Fz, FC1, FC2, FC5, FC6, C3, C4,
Cz, CP1, CP2, CP5, CP6, P3, P4, P7, P8, Pz, PO3, PO4,
PO7, PO8, O1, O2, and Oz. Two electrodes were placed
on the left and right mastoids, and a ground electrode
was placed at position FPz. All sites were recorded with
a right-mastoid reference, and were rereferenced off-line
to the algebraic average of the left and right mastoids.
Eye movements and blinks were recorded with passive
electrodes using horizontal and vertical EOG. Data were
filtered on-line (0.01–250 Hz) and were digitized at 1000 Hz
using BrainVision Recorder.
Artifact Rejection
We extracted data relative to the onset of spatial attention
cues (−300 to 1500 msec relative to cue onset). We used
an automatic pipeline implemented in prior publications
by our laboratory following a suggested EEG artifact re-
jection procedure (Luck, 2014). Data were automatically
examined for EEG artifacts (amplifier saturation, drifting,
dropout, and excessive high-frequency noise) and EOG
artifacts (blinks and saccades). We also extracted the
same time period (−300 to 1500 msec relative to cue on-
set) from the eye-tracking data. Using the automatic
pipeline implemented in prior publications by our labo-
ratory, these data were automatically examined for eye
artifacts (blinks and eye movements exceeding 0.5° from
fixation). On average, 11% of trials were rejected because
of eye artifacts, labeled as blinks (8%) and/or eye move-
ments (10%). After artifact rejection, we manually in-
spected all trials using EEGLAB in MATLAB. Participants
were excluded if fewer than half of the trials remained
after discarding those with artifacts. On average, 295 (77%)
trials remained per participant after artifact rejection.
2009). To equate for power differences across frequency
bands without removing sustained prestimulus signals, we
demeaned the signal based on the global average power
within each band.
Each trial therefore provided a multifrequency vector as
an input to the multivariate classification. Each trial was as-
signed a label for multivariate classification, corresponding
to the binary categorization of whether that trial was accu-
rately or inaccurately remembered. Accurate trials were tri-
als where the long-term memory response error was below
the median response error in the long-term memory phase.
Inaccurate trials were trials where the long-term response
error was greater than the median response error in the
long-term memory phase. To control for spatial imbalances,
we calculated a separate median response error for each
quadrant. Therefore, accurate and inaccurate trials con-
tained the same number of items per quadrant. These
labels were not redundant with the accurate/inaccurate
labels that would have been obtained from the working
memory phase (only 56% of trials would have received
the same label).
We split all accurate trials into two bins and all inaccu-
rate trials into two bins. We averaged all trials within each
bin to maximize the signal to noise of our EEG data. One
accurate bin and one inaccurate bin served as training
data, and the other accurate and inaccurate bins served
as test data. The assignment of trials to bins was random,
and we repeated this random assignment 1000 times for
each participant. In the end, the classifier was trained on
two vectors of length 180 (30 electrodes, six frequencies)
with binary labels. We trained a multivariate classifier using
L2-penalized logistic regression (C = 1) with the scikit-
learn package in Python. We tested the classifier on two
vectors of length 180 (30 electrodes, six frequencies) with
binary labels.
To examine sustained attention over time, we trained
separate classifiers at different moments of time follow-
ing cue presentation. For each 100-msec time window,
we averaged the power in each frequency band for each
electrode. We repeated this entire procedure for 10-msec
time steps. We conducted statistical analyses across par-
ticipants to compare classification accuracy at each time
point to theoretical chance (50%) as well as a shuffled
null, for which we permuted the labels. Code that de-
scribes the entire multivariate decoding procedure of
sustained attention is available on-line.
Multivariate Classification of Sustained Attention
Multivariate Classification of Spatial Attention
For all artifact-free trials, we used multivariate pattern classi-
fication to predict long-term memory from the prestimulus
time window. We decomposed EEG ERPs into oscillatory
bands (4–7 Hz, 8–12 Hz, 13–16 Hz, 16–20 Hz, 20–25 Hz,
25–30 Hz) by bandpass filtering the data and applying the
Hilbert transform (Kikumoto & Mayr, 2020; Weidemann &
Kahana, 2020; Manning, Polyn, Baltuch, Litt, & Kahana,
2011; Guderian, Schott, Richardson-Klavehn, & Düzel,
For all artifact-free trials, we used multivariate pattern
classification to predict the quadrant where the cue had
appeared. We repeated the same general procedure as
what was used to decode sustained attention. We decom-
posed EEG ERPs into alpha (8–12 Hz) power. We chose
to focus our analysis on the alpha frequency band a priori
based on extensive work showing classification of spatial
attention in this band (e.g., Foster et al., 2017).
2138
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Each valid trial therefore provided a vector input to the
multivariate classification. Each trial was assigned a label
for multivariate classification, corresponding to the quad-
rant where the cue had appeared (1–4). We down-
sampled trials such that there was an equal number of
trials per quadrant.
We determined for each trial which quadrant was cued
and whether long-term memory for that trial was accu-
rate or inaccurate. We split all trials within each quadrant
according to the median response error for that quad-
rant. Within each quadrant, we split all accurate trials into
two bins and all inaccurate trials into bins. Therefore, in
total, we had 16 bins of data (8 bins of accurate trials,
8 bins of inaccurate trials). We averaged all trials within
each bin to maximize the signal to noise of our EEG data.
We trained a classifier using a combined training set (one
bin per quadrant of accurate trials, one bin per quadrant
of inaccurate trials). We tested the classifier separately on
accurate (one bin per quadrant) and inaccurate data (one
bin per quadrant). The assignment of trials to bins was
random, and we repeated this random assignment 1000
times for each participant. In the end, the classifier was
trained on eight vectors of length 30 (30 electrodes, one
frequency) with Labels 1–4. We trained a multivariate
classifier using L2-penalized logistic regression (C = 1)
with the scikit-learn package in Python. We tested the
classifier twice, each time on four vectors of length 30
(30 electrodes, one frequency) with Labels 1–4.
To examine spatial attention over time, we trained sep-
arate classifiers at different moments of time following cue
presentation. We averaged the alpha power in each elec-
trode over a 100-msec time window. We repeated this en-
tire procedure for 10-msec time steps. We conducted
statistical analyses across participants to compare classifi-
cation accuracy at each time point to theoretical chance
(25%) as well as a shuffled null, for which we permuted
the labels. Code that describes the entire multivariate de-
coding procedure of spatial attention is available on-line.
Statistics
Statistics are the same as in Experiment 1.
Results
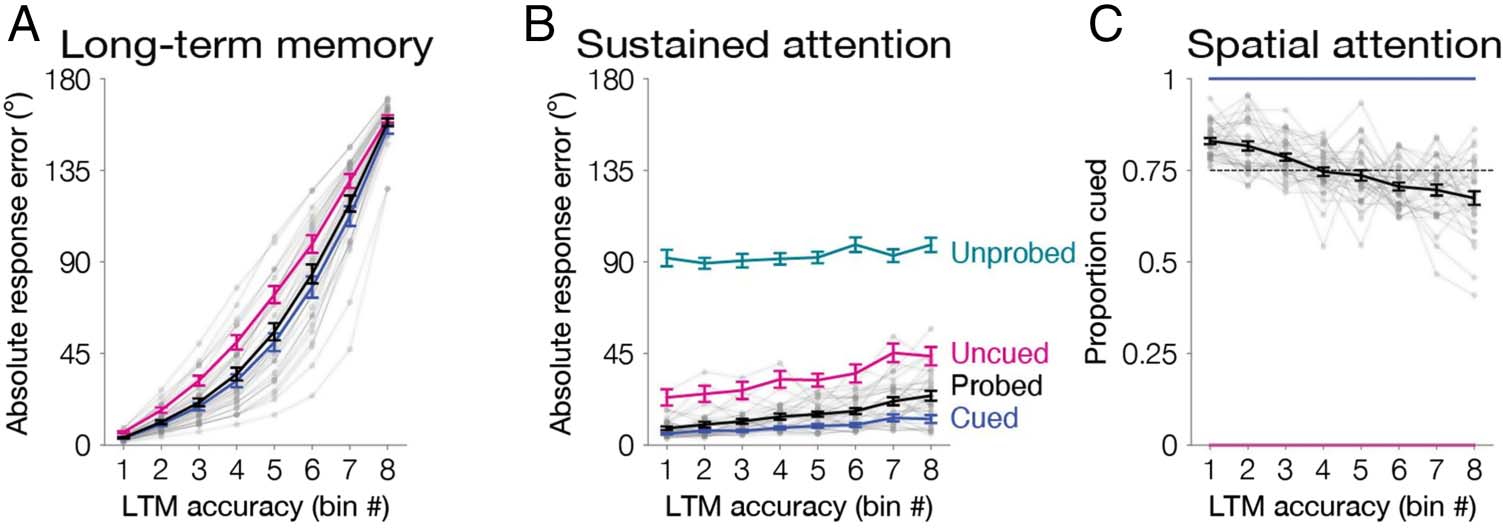
Behavioral Results
We again operationalized long-term memory as absolute
response error (dltm = 61.10°, 95% CIs [56.14, 65.92];
Figure 4A). We replicated our findings from Experiment
1 of a general correlation between working memory and
long-term memory. We observed a reliably positive corre-
lation between long-term memory and working memory
response error (r = .17, .13–.22, one-tailed p < 001). We
further quantified this as a positive slope relating long-
term memory (bin number, 1–8) to working memory re-
sponse error (m = 2.21, 1.70–2.89; one-tailed p < .001;
Figure 4B).
We next disentangled how long-term memory reflected
both spatial and sustained attention. We replicated the
finding that long-term memory reflected spatial attention,
as items that were cued to be spatially attended were better
remembered (dcued = 57.69°, 52.35–62.98; duncued =
71.34°, 66.19–75.86; one-tailed p < .001). This relationship
between spatial attention and long-term memory was fur-
ther quantified as a negative slope of the proportion cued
across bins (m = −0.023, −0.031 to −0.017; one-tailed
p < .001; Figure 4C).
We also replicated the finding that long-term memory
was affected by trial-by-trial fluctuations in sustained atten-
tion (Figure 4B). We observed a positive correlation be-
tween long-term memory and working memory response
Figure 4. Experiment 2
behavioral results. (A) Long-
term location memory
variability binned across trials.
Absolute response error
(0–180°) was sorted and binned
into octiles within participant
for all probed items (black line).
To equate for whether an item
was cued to be attended, we
also repeated all analyses
separately for cued items (blue)
and uncued items (pink). (B)
Memory outcomes reflect trial-
by-trial fluctuations of sustained
attention. Memory performance was operationalized as absolute response error (0–180°) for items within each bin. For uncued (pink), probed
(black), and cued (blue) items, absolute response error was obtained during the working memory phase. Unprobed items (teal) were not tested
in the working memory phase. Therefore, absolute response error for these items was obtained during the long-term memory phase. The slope
of each line is positive across the bins ( ps < .05). (C) Memory outcomes reflect differences of spatial attention. In each bin, we calculated the
proportion of trials that had been cued. Within all probed items (black), the proportion of trials that were cued decreases across the eight bins ( p <
.001). The dashed line represents the mean proportion cued (0.75). Cued items (blue) and uncued items (pink) controlled for whether an item
was cued to be attended (100% and 0% cued, respectively). Error bars depict the standard error of the mean. Data from each participant from
all probed items with a single cue are overlaid in small gray dots connected with lines.
deBettencourt et al.
2139
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
error for cued and uncued items (rcued = .11, .08–.16; run-
cued = .18, .13–.24; one-tailed ps < .001). We further
quantified this relationship as a slope relating long-term
memory (bin number, 1–8) to working memory response
error (mcued = 1.09, 0.74–1.77; muncued = 3.18, 2.27–4.26;
one-tailed ps < .001). Then, we examined the long-term
memory response error for unprobed items. We observed
a positive correlation between long-term memory for the
uncued items and long-term memory for the unprobed
items from the same display (r = .05, .00–.10, p = .02)
and a positive slope (m = 1.05, −0.23–2.17; one-tailed
p = .045). In summary, behavioral evidence replicated
the observation that long-term memory reflects distinct sig-
natures of spatial and sustained attention.
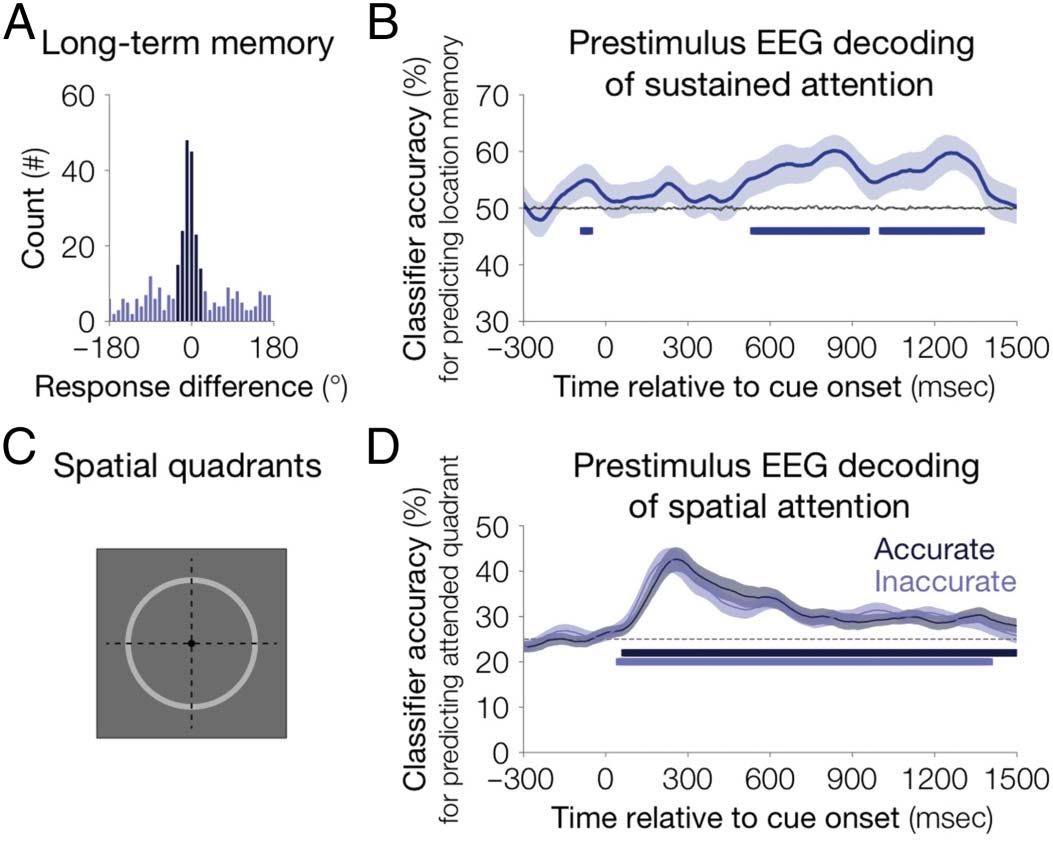
EEG
We observed from behavioral evidence the importance of
trial-to-trial fluctuations of sustained attention for later
memory. However, based on these behavioral findings,
we were unable to specify when and how these fluctua-
tions emerged. By examining EEG activity that tracks the
participants’ current attentional state, our goal was to in-
vestigate the role of prestimulus fluctuations of attention
on subsequent encoding into long-term memory. Thus,
we tested whether ongoing neural activity could predict
long-term memory success even before item presenta-
tion. Given that participants had to sustain attention over
2–4 sec following cue presentation, we targeted this pres-
timulus time window for analyses. We used multivariate
classification to decode whether a trial was remembered
accurately or inaccurately, relative to the median long-
term memory response error per quadrant (Figure 5A).
We reliably decoded long-term memory accuracy based
on EEG patterns in the time window following the cue
(mean accuracy = 57.23%, 95% CIs [52.87%, 62.58%];
n = 30, one-tailed p < .001, chance = 50%; t = 500–
1000 msec; Figure 5B). That is, we could predict whether
an upcoming item would be better remembered, even
before it appeared. These decoding results are consistent
with trial-by-trial fluctuations of sustained attention that
occur before stimulus presentation and influence memory
encoding.
The behavioral evidence suggested that sustained atten-
tion is distinct from spatial attention, as it broadly impacts
both cued and uncued items from the same display.
Therefore, we predicted that fluctuations of sustained
attention should be independent from fluctuations of
spatial attention. If this is the case, then our classifier of
sustained attention should be robust, even when we col-
lapse data across all spatial locations. Alternatively, fluctu-
ations of sustained attention could have reflected general
task disengagement. If so, fluctuations of sustained atten-
tion would predict worse spatial attention during low
moments of sustained attention. To explore these possi-
bilities, we investigated whether we could decode the
Figure 5. Multivariate decoding
of attentional processes from
EEG data. (A) Long-term
memory accuracy for a
representative participant.
A histogram of response
differences across trials, the
angular distance from the
original image location (−180°
to 180°). Labels for the
multivariate pattern classifier
reflected the median absolute
response error per quadrant,
either accurate (dark blue) or
inaccurate (light blue). For the
EEG decoding analyses, we
were interested in predicting
later accuracy from prestimulus
windows. To reduce the
influence of spatial attention on
our multivariate decoding, we
balanced the items from each
quadrant. (B) Prestimulus EEG
decoding of sustained attention. Long-term memory accuracy can be predicted before stimulus onset. A multivariate classifier was trained to predict
long-term location memory (accurate vs. inaccurate) based on multifrequency EEG patterns and tested on held out trials. Average classification
accuracy is depicted as the solid blue line, the gray line depicts empirical chance after shuffling the labels 1000 times. The shaded area is the standard
error of the mean. Blue squares highlight time points for which classification accuracy is above chance ( p < .05). (C) Spatially attended quadrants.
The display visually depicts the labels provided to the labels spatial attention classifier. Trials were split according the cued quadrant (1–4), four
quadrants illustrated by dashed lines. (D) Prestimulus EEG decoding of long-term memory is not explained by differences in spatial attention.
The cued quadrant was reliably decoded from both accurately and inaccurately remembered items. A multivariate classifier was trained to predict
cued location and tested on held out accurate or inaccurate trials. Average classification accuracy is depicted as solid lines for accurate trials
(dark blue) and inaccurate trials (light blue). Squares depict time points in which classification accuracy for either condition is above chance. The
shaded area is the standard error of the mean.
2140
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
cued quadrant (Figure 5C). During inaccurately remem-
bered trials, we reliably decoded the cued position (mean
accuracy = 31.82%, 95% CIs [29.99%, 33.98%]; n = 30;
one-tailed p < .001, chance = 25%; t = 0–1500 msec;
Figure 5D). We also reliably decoded the cued position
during accurately remembered trials (mean accuracy =
31.99%, 95% CIs [30.17%, 34.90%]; n = 30; one-tailed
p < .001, chance = 25%; t = 0–1500 msec; Figure 5D).
Critically, in both trials, we decoded the cued location,
consistent with the robust influence of spatial attention
on behavior. However, there was no reliable difference
in spatial attention decoding between accurate and inac-
curate trials (two-tailed p = .76). These decoding results
suggest that the ability to predict long-term memory
differences during prestimulus windows is not driven
by differences in spatial attention.
Recognition Memory
Overall recognition memory was above chance for all
0 = 0.87; 0.85–0.89; one-tailed p < .001 vs.
probed items (A
chance = 0.5). Recognized items exhibited a lower absolute
response error in the working memory phase (drecog =
14.30, 12.31–17.35; dunrecog = 16.48, 14.16–19.28; one-tailed
p < .001). We observed an effect of spatial attention on sub-
sequent item recognition, a greater proportion of items that
were later recognized had been cued (qrecog = 0.76, 0.75–
0.78; qunrecog = 0.73, 0.71–0.74; one-tailed p < .001). We
replicated our finding that items that were later recognized
exhibited better working memory response errors for
cued items (drecog = 8.73, 7.59–10.43; dunrecog = 10.13,
8.40–12.76; one-tailed p = .01), but not for uncued items
(drecog = 32.01, 25.95–40.75; dunrecog = 33.74, 27.70–
41.24; one-tailed p = .20) or unprobed items (hrecog =
0.16, 0.13–0.19; hunrecog = 0.15, 0.12–0.18; one-tailed
p = .15). Though item recognition results are largely
consistent with results obtained from continuous report,
these findings also suggest that continuous report may
provide a more sensitive assay of how spatial and sus-
tained attentional factors influence long-term memory.
reflect moments during which participants completely dis-
engaged from the task: EEG activity showed that partici-
pants maintained covert spatial attention at the cued
position, even when the cued stimulus was later inaccu-
rately remembered. Thus, multivariate analyses of EEG
data suggest that fluctuations in sustained attention can be
distinguished from the waxing and waning of spatial atten-
tion, and do not reflect episodes of global disengagement
with the task. Future studies could explore how prestimulus
attentional states, including both sustained attention and
spatial attention, influence stimulus encoding, mainte-
nance, and reinstatement. One possibility is that atten-
tional states (sustained and spatial attention) determine
a cascade of subsequent processes, including working
memory, that are all reflected in long-term memory.
EXPERIMENT 3
Thus far, we have focused primarily on recall of the spa-
tial position of the memoranda. Experiment 3 examined
whether distinct attentional subcomponents would also
influence long-term memory for other features that were
not shared with the spatial cue, namely, color. We hy-
pothesized that sustained and spatial attention would dis-
tinctly predict long-term memory for color.
Methods
Participants
In Experiment 3a and 3b, a combined 52 adults partici-
pated for University of Chicago course credit or $20 pay- ment ($10/hr). In Experiment 3a, 25 adults (15 female,
mean age = 23.0 years) participated. In Experiment 3b,
27 adults (20 female, mean age = 20.1 years) participated.
Two participants were excluded from Experiment 3 (one
from 3a and one from 3b) because of errors during data
collection, resulting in a final sample size of 50.
Apparatus
Apparatus used is the same as in Experiment 1b.
Discussion
Experiment 2 replicated and extended the behavioral
findings from Experiment 1, with concurrent eye tracking
to ensure spatial attention was maintained covertly. We
demonstrated that prestimulus multivariate EEG patterns
predicted later memory. These EEG decoding results are
consistent with a multivariate EEG representation of sus-
tained attentional state that preceded stimulus onset and
complement the behavioral evidence for trial-to-trial fluc-
tuations in behavior.
Furthermore, these behavioral and neural findings sup-
ported the composite model of attention and long-term
memory, as fluctuations of sustained attention were distinct
from spatial attention. These results also rule out the inter-
pretation that trial-by-trial fluctuations of sustained attention
Stimuli
A subset of the real-world object images from Experiments 1–
2 selected based on the relative uniqueness of the shape out-
lines was manipulated to be a one-dimensional color mask.
The color of each image was sampled randomly from a
360° Hue Saturation Value space, with saturation and value
of 1, and remapped to red, green, blue values for presentation
in PsychoPy.
Procedure
In the memory probes, participants reported the color
memory for each item instead of its spatial location
(Figure 6A). The item, colored dark gray, appeared at the
center of the screen, surrounded by a color wheel. The
deBettencourt et al.
2141
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
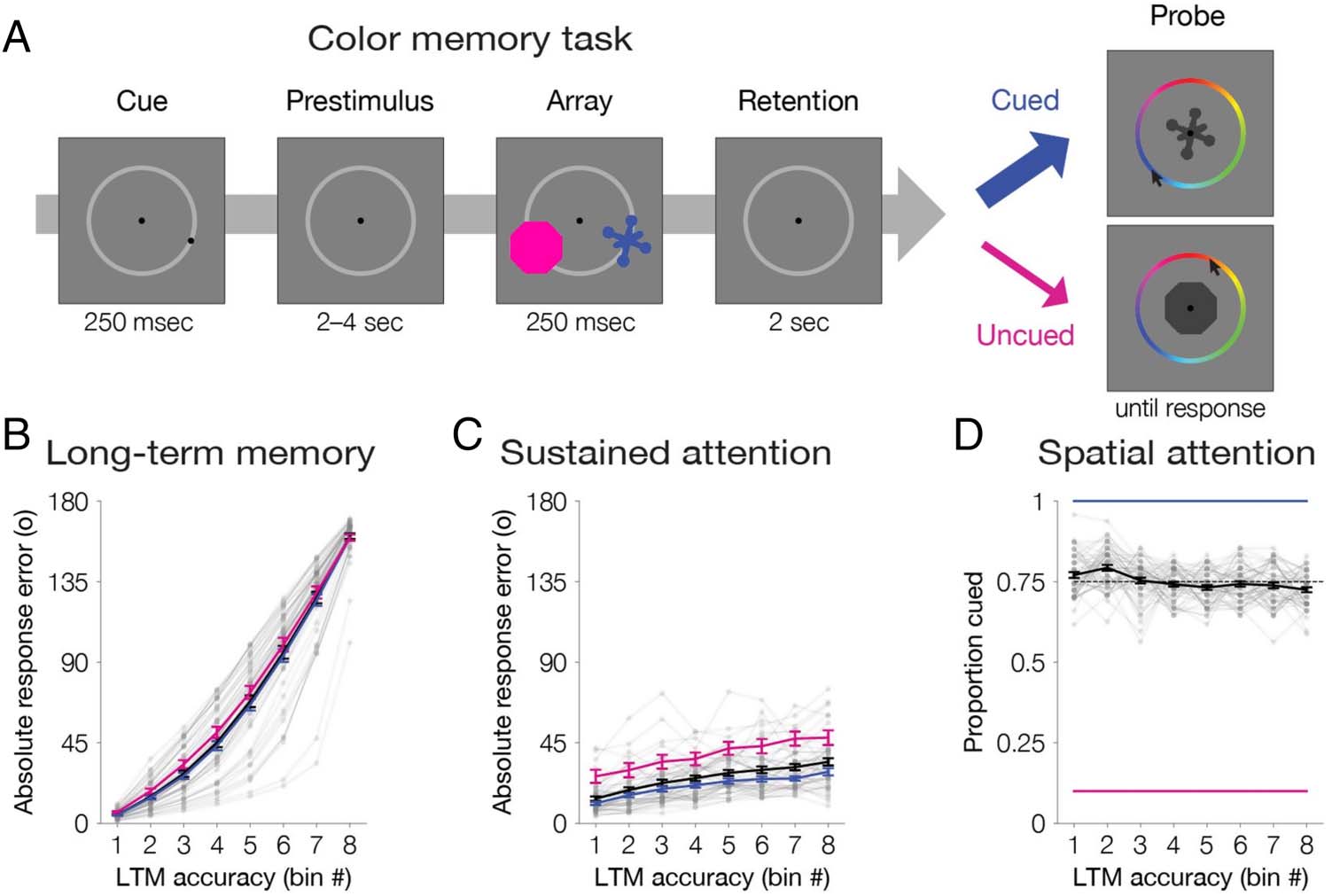
Figure 6. Experiment 3 color
memory results. (A) Task design
of the working memory phase.
Each trial was composed of
a brief peripheral spatial cue,
a blank prestimulus interval, a
memory array of two trial-
unique object pictures, and a
blank retention interval. The
object pictures were filled with
a color from a continuous color
wheel. For the memory probe,
one of the items (cued: 75%;
uncued 25%) reappeared at
the center in dark gray, and
participants reported its original
color by clicking along the
wheel. (B) Long-term memory
variability binned across trials.
Color memory absolute
response error (0–180°) was
sorted and binned into octiles
within participant for all probed
items (black line). To equate for
whether an item was cued to be
attended, we also repeated all
analyses separately for cued
items (blue) and uncued items (pink). (C) Memory outcomes reflect trial-by-trial fluctuations of sustained attention. Memory performance was
operationalized as absolute response error for items within each bin (0–180°). For uncued (pink), probed (black), and cued (blue) items, absolute
response error was obtained during the working memory phase. (D) Memory outcomes reflect differences of spatial attention. In each bin, we
calculated the proportion of trials that had been cued. For all probed items (black), the proportion of trials that were cued decreases across the
eight bins ( p < .001). The dashed line represents the mean proportion cued (0.75). Cued items (blue) and uncued items (pink) controlled for
whether an item was cued to be attended (100% and 0% cued, respectively). Error bars depict the standard error of the mean. Data from each
participant from all probed items with a single cue are overlaid in small gray dots connected with lines.
color wheel was randomly oriented for each trial, and this
random orientation was held consistent between working
memory and long-term memory tests. Based on piloting,
we reduced the working memory array to two items, which
were separated by a minimum distance of 40° in color
space. In the long-term memory phase, participants made
source memory judgments only for the probed images
from the working memory phase, both cued or uncued.
In Experiment 3a, participants completed 24 blocks of 16
trials, and participants completed, on average, 368 of the
maximum 384 trials (95.83%), ranging from 272 (70.83%)
to 384 (100%). In Experiment 3b, we eliminated the recog-
nition memory judgments and reduced the block length. In
Experiment 3b, participants completed 48 blocks of eight
trials and all participants completed all 384 trials.
Behavioral Analysis and Statistics
Behavioral analysis and statistics are the same as Experiments
1–2. All results were consistent for both Experiment 3a and
3b, and therefore we report the results as Experiment 3.
Results for each study separately are available on-line.
Results
The goal of this experiment was to replicate the findings
from Experiments 1 and 2 and extend them to the color
dimension. We operationalized long-term memory as ab-
solute response error from the original color (dltm =
67.90°, 95% CIs [63.44, 71.58]). For each participant,
we sorted and binned trials according to their long-term
color memory performance (Figure 6B). We observed
that long-term color memory bins reflected trial-by-trial
fluctuations of sustained attention, via a positive correla-
tion between long-term memory and working memory
(r = .18, .16–.21; one-tailed p < .001). We quantified this
relationship by relating long-term memory (bin number,
1–8) to working memory (m = 2.77, 2.38–3.26; one-
tailed p < .001; Figure 6C).
We next examined whether long-term color memory
also reflected distinct influences of spatial and sustained
attention. To examine the influence of spatial attention,
we observed that long-term color memory was enhanced
by cues (dcued = 66.75°, 62.37–70.41; duncued = 71.36°,
66.39–75.50; one-tailed p < .001). Furthermore, the pro-
portion of cued items within each bin declined across
bins (m = −0.008, −0.011 to −0.004; one-tailed p <
.001; Figure 6D).
To examine the influence of sustained attention, we re-
peated the binning analysis within each cueing condition
(Figure 6C). We observed that the long-term memory bin
was positively related to working memory response error
for cued items and uncued items (rcued = .17, 0.14–0.20;
runcued = .20, .16–.24; one-tailed ps < .001). We further
2142
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
quantified this relationship as a slope across long-term
memory bins (mcued = 2.25, 1.91–2.69, muncued = 3.23,
2.51–4.00; one-tailed ps < .001). In summary, behavioral
evidence confirms that both spatial attention and sus-
tained attention are critical in understanding long-term
color memory.
Recognition Memory
We only collected long-term recognition memory ratings
in Experiment 3a. Overall recognition memory sensitivity
0 = 0.78,
was well above chance for all probed items (A
0.74–0.82; one-tailed p < .001 vs. chance = 0.5). We also
observed lower working memory response error for
recognized versus unrecognized items (drecog = 23.52,
19.95–29.51; dunrecog = 29.51, 25.67–34.64; one-tailed
p < .001). We did not observe a reliable effect of spatial
attention: There was no difference in the proportion of cued
items for recognized versus unrecognized trials (qrecog =
0.76, 0.75–0.78; qunrecog = 0.75, 0.74–0.75; one-tailed p =
.08). However, trial-by-trial fluctuations of sustained atten-
tion influenced item recognition memory. Working memory
absolute response error was lower for items that were later
recognized, for cued items (drecog = 19.50, 16.66–24.47;
dunrecog = 23.69, 20.29–28.18; one-tailed p < .001) and
uncued items (drecog = 37.76, 29.59–51.29; dunrecog =
46.14, 38.02–58.34; one-tailed p < .001).
Discussion
These findings verified the influence of attention for long-
term memory of a nonspatial feature. Both sustained and
spatial attention distinctly related to long-term color mem-
ory. These results extend the findings from Experiment 1–
2, as the spatial location was a feature shared by the cue
and item. These results reaffirm that sustained attention
fluctuations are a broad and general influence for memo-
ries that can be clearly dissociated from the effects of
spatial attention.
Individual Differences
The goal of this study was primarily to explore attention and
long-term memory within individuals. However, we also ex-
amined between-subjects variations in long-term memory
performance and the underlying attentional factors that
drove those differences. We collapsed across data from all
experiments to boost the number of participants (n = 127).
We were interested in exploring the degree to which indi-
vidual differences in long-term memory reflected individual
differences in sustained or spatial attention.
Methods
Participants
We collapsed across data from all participants in
Experiments 1–3 (1a, 1b, 2, 3a, 3b) to maximize the
number of participants. We included all participants from
Experiment 1 (n = 47), Experiment 2 (n = 30), and
Experiment 3 (n = 50).
Behavioral Analysis
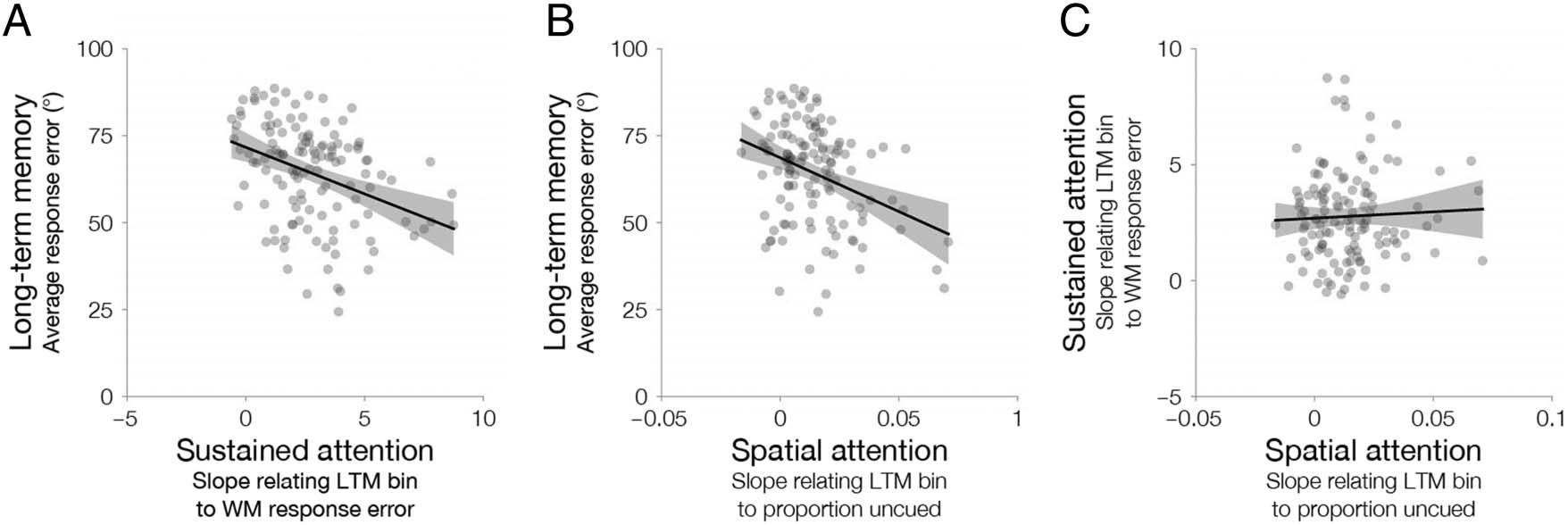
We examined measures of sustained and spatial attention
from the binning analysis of Experiments 1–3. For each
participant, we used our measure of the influence of sus-
tained attention from the previous studies, the slope that
related long-term memory to working memory absolute
response error. This operationalization of sustained at-
tention highlights the key influence of trial-by-trial fluctu-
ations of performance on later memory. To eliminate any
influence of spatial cues, we calculated the slope sepa-
rately for cued and uncued items and averaged across
conditions for a single measure of sustained attention.
We also used our measure of the influence of spatial
attention from the previous studies, the slope that related
long-term memory to the proportion of cued items within
each bin. So that sustained and spatial attention had the
same sign, we modified this value to be the slope that
related long-term memory to the proportion of uncued
items within each bin (1− proportion of cued items per
bin). This operationalization of spatial attention high-
lights the key influence of spatial cues on later memory.
We examined how measures of sustained and spatial
attention related to average long-term memory absolute
response error. We implemented two complementary
analyses, linear regression and nonparametric correla-
tions. We ran regression diagnostics to ensure linearity
and homoscedasticity. We also ensured our results were
robust after detecting and excluding any outliers. To ex-
amine the reliability of these measures of sustained atten-
tion, we replicated all of our results after subselecting
trials and after z-scoring within each of the five experi-
ments (Experiment 1a, 1b, 2, 3a, and 3b).
Statistics
Descriptive statistics are reported as the mean and 95% CI
of the bootstrapped distribution. Correlations and partial
correlations were computed using the nonparametric
Spearman’s rank-order correlation function. Linear models
were fit in R, and we report the adjusted r2 values. F statis-
tics evaluated different model fits. For the downsampling
analyses, we report the 95% CIs across downsampling
iterations.
Results
We obtained a measure of the influence of sustained and
spatial attention on performance from Experiments 1 to
3. We were interested in whether relying on either sus-
tained or spatial attention could be effective strategies
for long-term memory. As such, we quantified measures
of the strength of the influence of sustained and spatial
deBettencourt et al.
2143
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
attention on long-term memory behavior for each partic-
ipant. We also quantified a measure of overall long-term
memory performance for each participant as average
long-term memory response error.
Sustained Attention
Our measure of the influence of sustained attention was the
slope that related trial-by-trial fluctuations of working mem-
ory and long-term memory performance. A more positive
slope reflects a stronger influence of sustained attention
on memory. We were interested in whether a stronger in-
fluence of sustained attention correlated with overall long-
term memory performance. There was a reliably negative
relationship between the influence of sustained attention
on long-term memory and average long-term memory re-
sponse error (βsust = −2.68, −3.90 to −1.46; r2 = .12;
p < .001; Figure 7A). We also verified a relationship by cor-
relating the influence of sustained attention on long-term
memory and average long-term memory response error
(r = −.38, −.52 to −.22; p < .001). That is, a stronger in-
fluence of sustained attention correlates with better
average long-term memory performance.
Spatial Attention
Our measure of the influence of spatial attention was the
slope that related the proportion of items that had been
uncued within each long-term memory bin. A more posi-
tive slope reflects a stronger influence of spatial attention
on long-term memory. There was reliably negative relation-
ship between the influence of spatial attention and average
long-term memory response error (βspatial = −310, −459
to −161, r2 = .11, p < .001; Figure 7B). We also verified a
relationship by correlating the influence of spatial attention
on long-term memory and average long-term memory re-
sponse error (r = −.28, −0.43 to −0.11; p = .002). That is,
a stronger influence of spatial attention correlates with
better average long-term memory performance.
The critical question is whether these two signals, sus-
tained and spatial attention, reflect separate influences
on long-term memory. If so, we would predict that they
are uncorrelated with each other and explain a unique
portion of the variance. We found that individual differ-
ences in spatial attention and sustained attention were
uncorrelated with each other (r = .02, −.15 to .20; p =
.81; Figure 7C). In addition, they explained unique variance
in individual differences in long-term memory: The partial
correlation between sustained attention and long-term
memory (r = −.38, −0.52 to −0.23; p < .001) and the par-
tial correlation between spatial attention and long-term
memory (r = −.29, −.44 to −.12; p < .001) were both
reliable. The model using both signals as predictors ex-
plained more variance in individual differences in long-
term memory (adjusted r2 = .23) than a model that just
included sustained attention, F(1, 125) = 17.63, p < .001,
or spatial attention, F(1, 125) = 19.70, p < .001. In sum-
mary, individual differences for sustained attention and
spatial attention distinctly influence long-term memory.
Discussion
We examined the influence of sustained and spatial atten-
tion on long-term memory performance across participants
in the three experiments. These findings revealed that
Figure 7. Individual differences. (A) Individual differences in sustained attention relate to long-term memory. For each participant, we calculated a
measure of sustained attention, as the slope that related long-term memory (LTM) bin to working memory ( WM) response error. A more positive
value of sustained attention reflects a stronger influence of sustained attention on long-term memory. We observed that sustained attention was
negatively correlated with absolute response error in the long-term memory phase ( p < .001). That is, a stronger influence of sustained attention
correlates with better overall long-term memory. (B) Individual differences in spatial attention relate to long-term memory. For each participant, we
calculated a measure of spatial attention, as the slope that related long-term memory (LTM) bin to the proportion of uncued items per bin. A more
positive value of spatial attention reflects that long-term memory was especially related to whether an item was spatially cued. We observed a stronger
influence of spatial attention was negatively correlated with absolute response error in the long-term memory phase ( p < .001). That is, a stronger
influence of spatial attention correlates with better overall long-term memory. (C) Individual differences in sustained and spatial attention are
unrelated. We examined whether these measures of sustained attention was correlated with spatial attention across individuals. There was no reliable
relationship between sustained and spatial attention. Data from all participants in Experiments 1, 2, and 3 are overlaid in gray dots. The linear
relationship is depicted with a black line. Shaded area reflects 95% confidence limits.
2144
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
sustained and spatial attention predict distinct variance in
long-term memory performance.
GENERAL DISCUSSION
Over three experiments, we found that spatial and sus-
tained attention had robust and distinct effects on long-
term memory. Behaviorally, long-term memory was
superior when spatial attention was oriented toward the
item. Neurally, we decoded spatial attentional states from
multivariate analyses of EEG data before stimulus presen-
tation. We also observed robust behavioral influences of
trial-by-trial fluctuations of sustained attention, regardless
of whether items were spatially attended or not. In addi-
tion, trial-by-trial fluctuations of sustained attention were
correlated between multiple items from the same display.
We successfully decoded evidence for these sustained at-
tentional states, as we predicted future memory perfor-
mance from multivariate analyses of EEG data before
stimulus presentation. EEG measures of sustained atten-
tion were distinct from spatial attention, as they general-
ized across spatial positions, and the spatially attended
location was equally well decoded during lapses of sus-
tained attention. Moreover, this empirical pattern rules
out a general task engagement explanation of the fluctu-
ations associated with sustained attention. Finally, indi-
vidual differences analyses revealed that either spatial
attention or sustained attention were effective strategies
for overall long-term memory performance. In summary,
memory failures can be attributed to failures of distinct
spatial or sustained attentional processes, as shown by
these behavioral results, neural analyses, and individual
differences. Thus, our work provides key evidence that
begins to fractionate the monolithic relationship between
attention and long-term memory.
This work provides critical support for a composite
model of attention and long-term memory. That is, distinct
attentional subcomponents can explain memory failures.
We can conceptually illustrate the consequences of distinct
attentional processes with an addendum to the traditional
spotlight analogy: If spatial attention reflects the location of
the spotlight on the stage, sustained attention corresponds
to the number of audience members who are awake.
Critical to this analogy is the idea that sustained and
spatial attention can each vary independently. That is,
even when attention is oriented correctly, it could be
that sustained attention was deployed unsuccessfully.
Furthermore, either the spotlight or the audience’s
wakefulness can contribute to later memory of the per-
formance, as suggested by the composite model of atten-
tion and long-term memory. Notably, the composite
model does not require complete independence, and
there may be situations where these distinct processes
fluctuate in unison. Conversely, the unified model would
suggest that sustained attention and spatial attention
have completely overlapping influences (e.g., spotlight
location and brightness), which we did not observe.
Spatial versus Sustained Attention
This study sheds light on how two distinct attentional
processes impact encoding into long-term memory.
These findings align with prior attention research that
has shown spatial and sustained attention reflect distinct
cognitive processes, both within and between individuals
(Dowd & Golomb, 2019; Robison & Brewer, 2019; Poole
& Kane, 2009; Fan et al., 2005). Our study makes the key
contribution of extending the dissociable nature of sus-
tained and spatial attention to the domain of long-term
memory. In particular, this work advocates for a nuanced
approach of the relationship between multifaceted atten-
tional states and long-term memory. Traditionally, the re-
lationship between attention and long-term memory has
been investigated by manipulating top–down attention
(Aly & Turk-Browne, 2017). Here, we argue that it is im-
portant to distinguish between multiple forms of atten-
tion, and specifically consider fluctuations of sustained
attention, to understand how attention shapes memory.
Sustained Attention and Long-term Memory
This study also motivates further investigation of the links
between sustained attention and memory. In particular,
recent work has begun to explore the role of trial-by-trial
fluctuations of sustained attention as a gateway for work-
ing memory and long-term memory (Hakim et al., 2020;
deBettencourt et al., 2018, 2019). Sustained attention it-
self has represented a growing area within recent atten-
tion research (deBettencourt et al., 2015, 2018, 2019;
Rosenberg et al., 2016; Esterman, Rosenberg, &
Noonan, 2014; Esterman et al., 2013). However, memory
studies more rarely consider the role of sustained atten-
tion for memory formation (cf. Adam & deBettencourt,
2019). In part, sustained attention has been traditionally
measured via specific tasks, especially the sustained at-
tention to response task. This study extends our concep-
tualization of sustained attention from the sustained
attention to response task or highly similar tasks, to a
broader definition of trial-to-trial fluctuations of task per-
formance. This research characterizes sustained attention
as a sustained cognitive state, detectable pretrial, that im-
pacts encoding for multiple items from the display into
working memory and eventually long-term memory.
Future studies could examine how the sustained atten-
tional states in this study relate to those described in tra-
ditional sustained attention tasks.
Attention and Memory
This composite model of attention and memory reveals
how distinct attentional components influence long-term
memory. A key goal going forward will be to explore how
these distinct attentional processes influence memory
systems (Aly & Turk-Browne, 2016; LaRocque et al.,
2013). It also raises the possibility that previous
deBettencourt et al.
2145
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
demonstrations of prestimulus signals that predict mem-
ory may reflect trial-by-trial fluctuations of sustained
a n d / o r s p a t i a l a t t e n t i o n ( G ü n s e l i & A l y , 2 0 2 0 ;
Weidemann & Kahana, 2020; Adam, Mance, Fukuda, &
Vogel, 2015; Gruber & Otten, 2010; Guderian et al.,
2009; Otten, Quayle, Akram, Ditewig, & Rugg, 2006).
By carefully considering the role of distinct attentional
processes, specifically spatial and sustained attention, this
approach may reveal insight into the intricate and com-
plex relationship between attention and memory.
Memory Improvement
Finally, this work provides important suggestions for
strategies to improve or worsen memory. Cueing atten-
tion enhances long-term memories at the attended loca-
tion at the expense of memories elsewhere. In addition,
harnessing sustained attention fluctuations may be an-
other key for altering memory performance. Our findings
suggest that advantageous moments of sustained atten-
tion enhance memories for items distributed across spa-
tial locations. Thus, targeting optimally attentive states or
optimally attentive moments, or even improving general
sustained attention abilities (deBettencourt et al., 2015),
may be a key strategy for broadly improving memory.
Acknowledgments
We thank A. Gale for assistance with data collection for
Experiment 2, M. Bolouri for assistance with data collection
for Experiment 3, and K. C. S. Adam, N. Hakim, and A.
Tompary for comments on earlier versions of the paper.
Reprint requests should be sent to Megan T. deBettencourt,
Institute for Mind and Biology, University of Chicago, 940 E
57th St., Chicago, IL, or via e-mail: debetten@uchicago.edu.
Author Contributions
Megan T. deBettencourt: Conceptualization; Formal analysis;
Funding acquisition; Investigation; Methodology; Software;
Visualization; Writing—Original draft; Writing—Review &
editing. Stephanie D. Williams: Investigation; Methodology;
Writing—Review & editing. Edward K. Vogel: Conceptua-
lization; Funding acquisition; Resources; Supervision;
Writing—Review & editing. Edward Awh: Conceptua-
lization; Funding acquisition; Resources; Supervision;
Writing—Review & editing.
Funding Information
This research was supported by National Institutes of
Health (https://dx.doi.org/10.13039/100000002), grant
number: F32MH115597, the Office of Naval Research
(https://dx.doi.org/10.13039/100000006), grant number:
N00014-12-1-0972, and National
Institute of Mental
Health (https://dx.doi.org/10.13039/100000025), grant
number: R01MH087214 (M. T. dB.).
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/ W = .108, and W/ W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/ W = .102, and W/ W =
.076 (Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender
citation balance. The authors of this article report its pro-
portions of citations by gender category to be as follows:
M/M = .42, W/M = .36, M/W = .11, and W/W = .11.
REFERENCES
Adam, K. C. S., & deBettencourt, M. T. (2019). Fluctuations of
attention and working memory. Journal of Cognition, 2, 33.
https://doi.org/10.5334/joc.70, PubMed: 31440739
Adam, K. C. S., Mance, I., Fukuda, K., & Vogel, E. K. (2015). The
contribution of attentional lapses to individual differences
in visual working memory capacity. Journal of Cognitive
Neuroscience, 27, 1601–1616. https://doi.org/10.1162/jocn_a
_00811, PubMed: 25811710
Aly, M., & Turk-Browne, N. B. (2016). Attention promotes
episodic encoding by stabilizing hippocampal representations.
Proceedings of the National Academy of Sciences, U.S.A., 113,
E420–E429. https://doi.org/10.1073/pnas.1518931113, PubMed:
26755611
Aly, M., & Turk-Browne, N. B. (2017). How hippocampal
memory shapes, and is shaped by, attention. In The
hippocampus from cells to systems (pp. 369–403). Cham:
Springer. https://doi.org/10.1007/978-3-319-50406-3_12
Biderman, N., Luria, R., Teodorescu, A. R., Hajaj, R., & Goshen-
Gottstein, Y. (2019). Working memory has better fidelity than
long-term memory: The fidelity constraint is not a general
property of memory after all. Psychological Science, 30,
223–237. https://doi.org/10.1177/0956797618813538,
PubMed: 30589615
Brady, T. F., Konkle, T., Alvarez, G. A., & Oliva, A. (2008). Visual
long-term memory has a massive storage capacity for object
details. Proceedings of the National Academy of Sciences,
U.S.A., 105, 14325–14329. https://doi.org/10.1073/pnas
.0803390105, PubMed: 18787113
Brodeur, M. B., Guérard, K., & Bouras, M. (2014). Bank of
standardized stimuli (BOSS) phase II: 930 New normative
photos. PLoS One, 9, e106953. https://doi.org/10.1371/journal
.pone.0106953, PubMed: 25211489
Chun, M. M., Golomb, J. D., & Turk-Browne, N. B. (2011). A
taxonomy of external and internal attention. Annual Review
of Psychology, 62, 73–101. https://doi.org/10.1146/annurev
.psych.093008.100427, PubMed: 19575619
Chun, M. M., & Turk-Browne, N. B. (2007). Interactions
between attention and memory. Current Opinion in
Neurobiology, 17, 177–184. https://doi.org/10.1016/j.conb
.2007.03.005, PubMed: 17379501
deBettencourt, M. T., Cohen, J. D., Lee, R. F., Norman, K. A., &
Turk-Browne, N. B. (2015). Closed-loop training of attention
2146
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
with real-time brain imaging. Nature Neuroscience, 18,
470–475. https://doi.org/10.1038/nn.3940, PubMed: 25664913
deBettencourt, M. T., Keene, P. A., Awh, E., & Vogel, E. K.
(2019). Real-time triggering reveals concurrent lapses of
attention and working memory. Nature Human Behaviour,
3, 808–816. https://doi.org/10.1038/s41562-019-0606-6,
PubMed: 31110335
deBettencourt, M. T., Norman, K. A., & Turk-Browne, N. B.
(2018). Forgetting from lapses of sustained attention.
Psychonomic Bulletin & Review, 25, 605–611. https://doi.org
/10.3758/s13423-017-1309-5, PubMed: 28585055
Decker, A. L., & Duncan, K. (2020). Acetylcholine and the
complex interdependence of memory and attention. Current
Opinion in Behavioral Sciences, 32, 21–28. https://doi.org
/10.1016/j.cobeha.2020.01.013
Dowd, E. W., & Golomb, J. D. (2019). Object-feature binding
survives dynamic shifts of spatial attention. Psychological
Science, 30, 343–361. https://doi.org/10.1177/0956797618818481,
PubMed: 30694718
Efron, B., & Tibshirani, R. (1986). Bootstrap methods for
standard errors, confidence intervals, and other measures of
statistical accuracy. Statistical Science, 1, 54–75. https://doi
.org/10.1214/ss/1177013815
Esterman, M., Noonan, S. K., Rosenberg, M., & DeGutis, J.
(2013). In the zone or zoning out? Tracking behavioral and
neural fluctuations during sustained attention. Cerebral
Cortex, 23, 2712–2723. https://doi.org/10.1093/cercor/bhs261,
PubMed: 22941724
Esterman, M., Rosenberg, M. D., & Noonan, S. K. (2014). Intrinsic
fluctuations in sustained attention and distractor processing.
Journal of Neuroscience, 34, 1724–1730. https://doi.org/10
.1523/JNEUROSCI.2658-13.2014, PubMed: 24478354
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J. I., & Posner,
M. I. (2005). The activation of attentional networks.
Neuroimage, 26, 471–479. https://doi.org/10.1016/j
.neuroimage.2005.02.004, PubMed: 15907304
Fan, J. E., & Turk-Browne, N. B. (2016). Incidental biasing
of attention from visual long-term memory. Journal of
Experimental Psychology. Learning, Memory, and
Cognition, 42, 970–977. https://doi.org/10.1037/xlm0000209,
PubMed: 26618914
Foster, J. J., Sutterer, D. W., Serences, J. T., Vogel, E. K., &
Awh, E. (2017). Alpha-band oscillations enable spatially and
temporally resolved tracking of covert spatial attention.
Psychological Science, 28, 929–941. https://doi.org/10.1177
/0956797617699167, PubMed: 28537480
Fukuda, K., & Vogel, E. K. (2019). Visual short-term memory
capacity predicts the “bandwidth” of visual long-term
memory encoding. Memory & Cognition, 47, 1481–1497.
https://doi.org/10.3758/s13421-019-00954-0, PubMed:
31236821
Gruber, M. J., & Otten, L. J. (2010). Voluntary control over
prestimulus activity related to encoding. Journal of
Neuroscience, 30, 9793–9800. https://doi.org/10.1523
/JNEUROSCI.0915-10.2010, PubMed: 20660262
Guderian, S., Schott, B. H., Richardson-Klavehn, A., & Düzel, E.
(2009). Medial temporal theta state before an event predicts
episodic encoding success in humans. Proceedings of the
National Academy of Sciences, U.S.A., 106, 5365–5370.
https://doi.org/10.1073/pnas.0900289106, PubMed: 19289818
Günseli, E., & Aly, M. (2020). Preparation for upcoming
attentional states in the hippocampus and medial prefrontal
cortex. eLife, 9, e53191. https://doi.org/10.7554/eLife.53191,
PubMed: 32255423
Hakim, N., Adam, K. C. S., Gunseli, E., Awh, E., & Vogel, E. K.
(2019). Dissecting the neural focus of attention reveals
distinct processes for spatial attention and object-based
storage in visual working memory. Psychological Science,
30, 526–540. https://doi.org/10.1177/0956797619830384,
PubMed: 30817220
Hakim, N., deBettencourt, M. T., Awh, E., & Vogel, E. K. (2020).
Attention fluctuations impact ongoing maintenance of
information in working memory. Psychonomic Bulletin &
Review, 27, 1269–1278. https://doi.org/10.3758/s13423-020
-01790-z, PubMed: 32808159
Hannula, D. E. (2018). Chapter nine: Attention and long-term
memory: bidirectional interactions and their effects on
behavior. In K. D. Federmeier (Ed.), Psychology of learning
and motivation (Vol. 69, pp. 285–323). Academic Press.
https://doi.org/10.1016/bs.plm.2018.09.004
Kikumoto, A., & Mayr, U. (2020). Conjunctive representations
that integrate stimuli, responses, and rules are critical for
action selection. Proceedings of the National Academy of
Sciences, U.S.A., 117, 10603–10608. https://doi.org/10.1073
/pnas.1922166117, PubMed: 32341161
LaRocque, J. J., Eichenbaum, A. S., Starrett, M. J., Rose, N. S.,
Emrich, S. M., & Postle, B. R. (2015). The short- and long-
term fates of memory items retained outside the focus of
attention. Memory & Cognition, 43, 453–468. https://doi.org
/10.3758/s13421-014-0486-y, PubMed: 25472902
LaRocque, K. F., Smith, M. E., Carr, V. A., Witthoft, N., Grill-
Spector, K., & Wagner, A. D. (2013). Global similarity and
pattern separation in the human medial temporal lobe
predict subsequent memory. Journal of Neuroscience, 33,
5466–5474. https://doi.org/10.1523/JNEUROSCI.4293-12
.2013, PubMed: 23536062
Luck, S. J. (2014). An Introduction to the event-related
potential technique (2nd ed.). Cambridge, MA: MIT Press.
Manning, J. R., Polyn, S. M., Baltuch, G. H., Litt, B., & Kahana, M. J.
(2011). Oscillatory patterns in temporal lobe reveal context
reinstatement during memory search. Proceedings of the
National Academy of Sciences, U.S.A., 108, 12893–12897.
https://doi.org/10.1073/pnas.1015174108, PubMed: 21737744
Otten, L. J., Quayle, A. H., Akram, S., Ditewig, T. A., & Rugg, M. D.
(2006). Brain activity before an event predicts later recollection.
Nature Neuroscience, 9, 489–491. https://doi.org/10.1038
/nn1663, PubMed: 16501566
Peirce, J. W. (2007). PsychoPy—Psychophysics software in
Python. Journal of Neuroscience Methods, 162, 8–13. https://
doi.org/10.1016/j.jneumeth.2006.11.017, PubMed: 17254636
Poole, B. J., & Kane, M. J. (2009). Working-memory capacity
predicts the executive control of visual search among
distractors: The influences of sustained and selective
attention. Quarterly Journal of Experimental Psychology,
62, 1430–1454. https://doi.org/10.1080/17470210802479329,
PubMed: 19123118
Richter, F. R., Cooper, R. A., Bays, P. M., & Simons, J. S. (2016).
Distinct neural mechanisms underlie the success, precision,
and vividness of episodic memory. eLife, 5, e18260. https://
doi.org/10.7554/eLife.18260, PubMed: 27776631
Robison, M., & Brewer, G. (2019). Individual differences in
vigilance: Implications for measuring sustained attention and
its association with other cognitive abilities and psychological
constructs. PsyArXiv. https://doi.org/10.31234/osf.io/nc9b6
Rosenberg, M. D., Finn, E. S., Scheinost, D., Papademetris, X.,
Shen, X., Constable, R. T., et al. (2016). A neuromarker of
sustained attention from whole-brain functional connectivity.
Nature Neuroscience, 19, 165–171. https://doi.org/10.1038
/nn.4179, PubMed: 26595653
Rosenberg, M., Noonan, S., DeGutis, J., & Esterman, M. (2013).
Sustaining visual attention in the face of distraction: A novel
gradual-onset continuous performance task. Attention,
Perception, & Psychophysics, 75, 426–439. https://doi.org/10
.3758/s13414-012-0413-x, PubMed: 23299180
Sprague, T. C., & Serences, J. T. (2013). Attention modulates
spatial priority maps in the human occipital. parietal and
deBettencourt et al.
2147
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
frontal cortices. Nature Neuroscience, 16, 1879–1887.
https://doi.org/10.1038/nn.3574, PubMed: 24212672
Sutterer, D. W., & Awh, E. (2016). Retrieval practice enhances
the accessibility but not the quality of memory. Psychonomic
Bulletin & Review, 23, 831–841. https://doi.org/10.3758
/s13423-015-0937-x, PubMed: 26404635
Tompary, A., Zhou, W., & Davachi, L. (2020). Schematic
memories develop quickly, but are not expressed unless
necessary. Scientific Reports, 10, 16968. https://doi.org/10
.1038/s41598-020-73952-x, PubMed: 33046766
Turk-Browne, N. B., Golomb, J. D., & Chun, M. M. (2013).
Complementary attentional components of successful
memory encoding. Neuroimage, 66, 553–562. https://doi
.org/10.1016/j.neuroimage.2012.10.053, PubMed: 23108276
Uncapher, M. R., Hutchinson, J. B., & Wagner, A. D. (2011).
Dissociable effects of top–down and bottom–up attention
during episodic encoding. Journal of Neuroscience, 31,
12613–12628. https://doi.org/10.1523/JNEUROSCI.0152-11
.2011, PubMed: 21880922
Weidemann, C. T., & Kahana, M. J. (2020). Neural measures of
subsequent memory reflect endogenous variability in cognitive
function. BioRxiv, 576173. https://doi.org/10.1101/576173
Xie, W., Park, H.-B., Zaghloul, K. A., & Zhang, W. (2020).
Correlated individual differences in the estimated precision
of working memory and long-term memory: Commentary
on the study by Biderman, Luria, Teodorescu, Hajaj, and
Goshen-Gottstein (2019). Psychological Science, 31, 345–348.
https://doi.org/10.1177/0956797620903718, PubMed:
32049590
Ziman, K., Lee, M. R., Martinez, A. R., & Manning, J. R. (2019).
Feature-based and location-based volitional covert attention
are mediated by different mechanisms and affect memory at
different timescales [Preprint]. PsyArXiv. https://doi.org/10
.31234/osf.io/2ps6e
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
1
3
2
1
9
6
2
1
4
3
/
/
j
o
c
n
_
a
_
0
1
7
4
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
2148
Journal of Cognitive Neuroscience
Volume 33, Number 10