Stress Elevates Frontal Midline Theta in Feedback-based
Category Learning of Exceptions
Marcus Paul1, Marie-Christin Fellner1, Gerd T. Waldhauser1, John Paul Minda2,
Nikolai Axmacher1, Boris Suchan1, and Oliver T. Wolf1
Abstrakt
■ Adapting behavior based on category knowledge is a funda-
mental cognitive function, which can be achieved via different
learning strategies relying on different systems in the brain.
Whereas the learning of typical category members has been
linked to implicit, prototype abstraction learning, which relies
predominantly on prefrontal areas, the learning of exceptions
is associated with explicit, exemplar-based learning, which has
been linked to the hippocampus. Stress is known to foster im-
plicit learning strategies at the expense of explicit learning. Pro-
cedural, prefrontal learning and cognitive control processes are
reflected in frontal midline theta (4–8 Hz) oscillations during
feedback processing. In the current study, we examined the
effect of acute stress on feedback-based category learning of
typical category members and exceptions and the oscillatory
correlates of feedback processing in the EEG. A computational
modeling procedure was applied to estimate the use of abstrac-
tion and exemplar strategies during category learning. Wir
tested healthy, male participants who underwent either the
socially evaluated cold pressor test or a nonstressful control
procedure before they learned to categorize typical members
and exceptions based on feedback. The groups did not differ
significantly in their categorization accuracy or use of categori-
zation strategies. In the EEG, Jedoch, stressed participants re-
vealed elevated theta power specifically during the learning of
exceptions, whereas the theta power during the learning of typi-
cal members did not differ between the groups. Elevated frontal
theta power may reflect an increased involvement of medial pre-
frontal areas in the learning of exceptions under stress. ■
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
J
Ö
C
N
_
A
_
0
1
2
4
1
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
EINFÜHRUNG
Learning to categorize objects, Veranstaltungen, and people to dis-
crete classes is a fundamental cognitive ability, which en-
ables humans to make quick decisions in rewarding or
threatening situations. Physical and psychosocial threats
elicit stress responses, which are associated with endo-
crine changes in the body, such as the release of gluco-
corticoids. Stress has been shown to modulate feedback
processing and learning processes (Glienke, Wolf, &
Bellebaum, 2015; Lighthall, Gorlick, Schoeke, Frank, &
Mather, 2013; Cavanagh, Frank, & Allen, 2011; Ossewaarde
et al., 2011).
Different learning strategies have been identified in
feedback-based category learning, which are associated
with separate systems in the brain. Prototype-based pro-
cedural learning strategies (Reed, 1972; Posner & Keele,
1968), which rely on regions in PFC (Pan & Sakagami, 2012)
and the striatal learning system (Cincotta & Seger, 2007;
Ashby & Ennis, 2006), are distinguished from exemplar-
based declarative learning strategies (Schenk, Minda, Lech,
& Suchan, 2016; Nosofsky, 1986; Medin & Schaffer, 1978).
In prototype-based procedural learning, common
characteristics of all category members are abstracted
1Ruhr-University Bochum, 2University of Western Ontario
© 2018 Massachusetts Institute of Technology
from the prototype of this category and are used to form
a category representation. In exemplar-based learning,
categories are learned on the basis of each stimulus.
Whereas abstraction-based strategies are employed to
learn typical members of a category, exceptions have to
be learned by means of an exemplar-based strategy
(Lech, Güntürkün, & Suchan, 2016; Cook & Schmied, 2006).
The striatum is crucial for associative stimulus–response
learning based on reward prediction errors, welche sind
defined as violations to outcome predictions (Diederen,
Spencer, Vestergaard, Fletcher, & Schultz, 2016; Rolls,
McCabe, & Redoute, 2008). In feedback-based category
learning, stimulus-category associations are acquired
(Cincotta & Seger, 2007; Seger & Cincotta, 2005) by updat-
ing reward expectations after a reward prediction error was
experienced (Nasser, Calu, Schoenbaum, & Sharpe, 2017;
Seger, Peterson, Cincotta, Lopez-Paniagua, & Anderson,
2010; Sutton & Barto, 1981).
In electrophysiological studies with macaque monkeys,
it was found that the striatum acts jointly with PFC during
category learning, which was demonstrated by increases
in the functional connectivity between the striatum and
PFC during learning (Antzoulatos & Müller, 2011, 2014).
Medial PFC regions, such as the dorsal ACC (dACC) Und
the dorsomedial PFC, represent the value of response op-
tions and reward expectancies and code reward prediction
Zeitschrift für kognitive Neurowissenschaften 30:6, S. 799–813
doi:10.1162/jocn_a_01241
Fehler. The dACC receives reinforcement learning signals
(Alexander & Braun, 2011; Holroyd & Coles, 2002,
2008), and theta oscillations (4–8 Hz) are employed by
medial frontal regions in the communication with lateral
prefrontal and premotor regions to realize the adaptation
of behavior (Smith et al., 2015; Oehrn et al., 2014; van de
Vijver, Ridderinkhof, & Cohen, 2011).
In the scalp EEG, these frontal midline theta (FMT)
oscillations have been linked with cognitive control pro-
Prozesse (Cavanagh & Frank, 2014) and are involved in
the evaluation of feedback and errors based on reinforce-
ment learning signals. A larger FMT power is detected
after negative feedback compared with positive feedback
and after an error was made compared with a correct re-
sponse (Cavanagh & Frank, 2014; Cohen, 2014; Cohen,
Wilmes, & van de Vijver, 2011). The power of the FMT cor-
relates with size of the reward prediction errors and the
adaptation of behavior in subsequent trials (Mas-Herrero
& Marco-Pallarés, 2014, 2016; van de Vijver et al., 2011;
Cavanagh, Frank, Klein, & Allen, 2010). Inducing inphase
theta oscillations synchronously in the medial and lat-
eral PFC by means of transcranial alternating current
stimulation has been shown to increase the feedback-
based adaptation of behavior, whereas the induction
of antiphase theta oscillations impairs the adaptation
of behavior (Reinhart, 2017). Intracranial recordings in
humans demonstrated directionality in this connectivity
between medial and lateral prefrontal areas, such that
theta oscillations propagate from medial prefrontal
areas to the lateral PFC during feedback processing to
adapt behavior (Smith et al., 2015). These studies sug-
gest a causal role of the FMT in adaptive behavior. Theta
oscillations are therefore a promising target to investi-
gate the stress-related modulation of medial frontal pro-
cesses in associative learning, such as abstraction-based
categorization, with a high temporal resolution.
Stress and the stress-induced release of glucocorticoids
are associated with changes in the feedback processing
and learning (Lighthall et al., 2013; Cavanagh et al.,
2011; Ossewaarde et al., 2011). The influence of stress
on electrophysiological correlates of the feedback pro-
cessing has been tested by recent studies using probabi-
listic reinforcement learning tasks. It was demonstrated
that stress increases the amplitude of the feedback-related
negativity (FRN; Wirz, Wacker, Felten, Reuter, & Schwabe,
2017; Glienke et al., 2015), a negative potential, welches ist
more pronounced after negative feedback compared with
positive feedback and which is the time domain representa-
tion of the FMT (Cohen et al., 2011).
Stress-related changes have also been found to influ-
ence categorization. In category learning, stress causes
a switch between the relative use of learning strategies
from declarative to nondeclarative strategies (Schwabe
& Wolf, 2013). In a deterministic categorization task, dis-
sociating striatal, procedural from prefrontal, rule-based
learning, stress was found to enhance striatum-dependent
procedural learning (Ell, Cosley, & McCoy, 2011). So far, Es
is unknown whether stress influences the learning of
categories comprising typical, rule-following members
and exceptions to the rule, which often occur in natural-
istic categories.
The current study therefore tested the influence of
stress on category learning of typical category members
and exceptions. The analysis of frontal theta oscillations
in the EEG during the feedback processing allows us to
investigate the modulatory effects of stress on cortical
oscillations that are involved in learning and feedback
Verarbeitung. Außerdem, a computational modeling pro-
cedure was applied to assess the use of prototype abstrac-
tion and exemplar-based strategies during learning. To
investigate these questions, we exposed participants to
either a stressful situation or a nonstressful control situa-
tion before they conducted a feedback-based category
learning task composed of typical category members and
exceptions (Cook & Schmied, 2006). It was expected that
stress might impair the exemplar-based learning of excep-
tionen, whereas implicit, abstraction-based learning of typi-
cal stimuli should be unaffected. On the basis of previous
Studien (Schwabe & Wolf, 2012), it was expected that stress
would foster the use of the abstraction-based strategies
stattdessen. Außerdem, stress was hypothesized to increase
the FMT power reflecting changes in the frontal feedback
Verarbeitung. The FMT power increase may compensates for
reduced hippocampal contribution or an increased need
for cognitive control in the learning of exceptions.
METHODEN
Teilnehmer
Forty-seven men participated in the study after a short
screening for their general health status, with exclusion
criteria such as smoking, current or history of psychiatric
or neurological disorders, the intake of medication, A
body mass index below 18 or above 29 kg/m2, substance
abuse, and color blindness. Zusätzlich, participants had
to be naive to the socially evaluated cold pressor test
(SECPT; Schwabe, Haddad, & Schachinger, 2008). Zwanzig-
four participants were randomly assigned to the stress group
(Durchschnittsalter = 24.8 Jahre, SEM = 0.8 Jahre), Und 23 war
assigned to the control group (Durchschnittsalter = 25.6 Jahre,
SEM = 0.9 Jahre). Participants were excluded if their EEG
data consisted of fewer than 10 artifact-free EEG trials in any
of the experimental conditions (n = 11; see below for a
detailed description). Zusätzlich, cortisol nonresponders
(d.h., those not showing an increase in cortisol from
baseline to peak in response to the stress of larger than
1.5 nmol/L; n = 5; Müller, Plessow, Kirschbaum, & Stalder,
2013) were excluded (Glienke et al., 2015; McCullough,
Ritchey, Ranganath, & Yonelinas, 2015), because pharma-
cological work has repeatedly shown that glucocorticoids
are crucial mediators in the impact of stress on different
aspects of cognition (Vogel et al., 2017; Vogel, Fernández,
Joëls, & Schwabe, 2016; Schwabe, Tegenthoff, Höffken, &
800
Zeitschrift für kognitive Neurowissenschaften
Volumen 30, Nummer 6
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
J
Ö
C
N
_
A
_
0
1
2
4
1
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Wolf, 2013). The final sample consisted of 16 Teilnehmer
in the stress group and 16 in the control group.
The study was approved by the local ethics committee of
the Faculty of Psychology at the Ruhr-University Bochum
and is according to the Declaration of Helsinki. All partici-
pants gave written informed consent before participation
and were reimbursed with A20.
Verfahren
The experimental procedure was conducted between 2
Und 6 Uhr. to control for the diurnal cycle of endogenous
cortisol concentrations (Kalsbeek et al., 2012). Partici-
pants refrained from alcohol and excessive exercise the
day before the testing and refrained from eating and
drinking anything except for water 2 hr before the testing.
After giving written informed consent, electrodes were
prepared for the EEG recordings. Subsequently, the first
saliva sample was collected (−1 min) before the stress
group underwent the stressful SECPT, whereas the control
group was assigned to a nonstressful control procedure
(both procedures are described in detail in the following
section). Nachher, the second saliva sample (+1 min)
was collected, and subjective stress ratings were obtained
from the participants. Twenty minutes after the treatment,
a saliva sample was collected (+20 min) and participants
conducted the category learning task (siehe unten). Nach-
ward, a final saliva sample was collected (+55 min), Und
participants were shortly debriefed and reimbursed.
Stress Induction
Stress was induced by means of the SECPT, und der Betrug-
trol group was assigned to a nonstressful control proce-
dure. The stress group immersed their right hand in ice
Wasser (0–2°C) for up to 3 min. During the SECPT, Sie
were filmed with a video camera and received instruc-
tions from a reserved female experimenter to keep their
gaze fixated at the camera and to refrain from movement.
The control group immersed their hands in warm water
(35–37°C). They were neither filmed nor observed by a
demure experimenter. Subjective ratings of the difficulty
to keep the hand immersed, the discomfort, the pain,
and the stress felt during the treatment were given on
11-point Likert scales from 0 (not at all ) Zu 100 (sehr
viel). Systolic and diastolic blood pressure and heart
rate were measured before, während, and after the treat-
ment with the Dinamap system (Critikon, Tampa, FL).
At each time point, three measurements of blood pres-
sure and heart rate were obtained, which were used to
calculate an average for each time point. Saliva was col-
lected using Salivettes (Sarstedt, Nümbrecht, Deutschland).
Samples were kept at −18°C until analysis. Cortisol concen-
trations were determined in duplicates using a cortisol
enzyme-linked immunosorbent assay (Demeditec, Kiel,
Deutschland). Interassay and intra-assay coefficients of vari-
ance were below 10%.
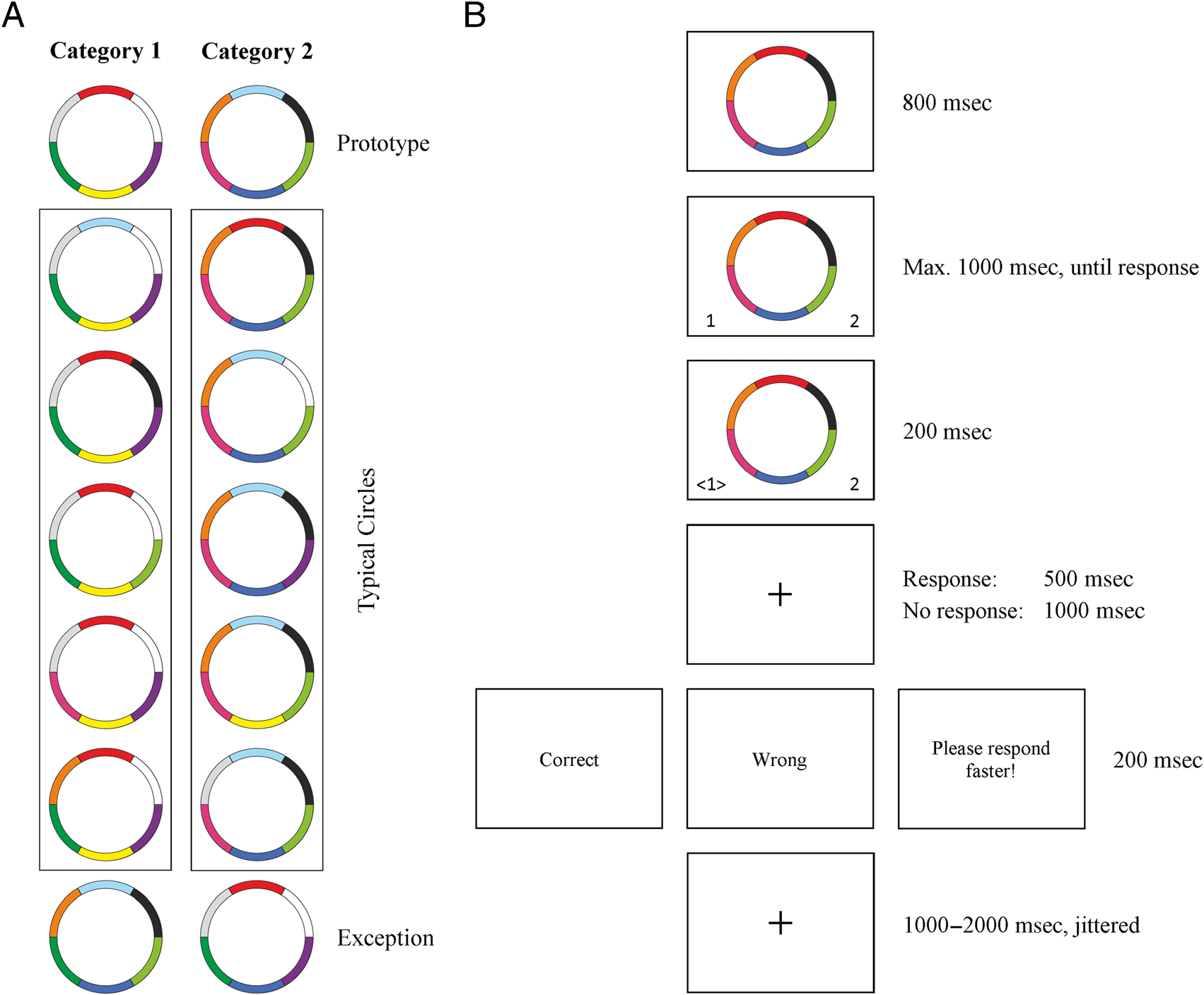
Category Learning Task
In the category learning task, participants learned, based
on feedback to their responses, to assign circular stimuli
(Figure 1A) to one of two categories (Schenk et al., 2016;
Cook & Schmied, 2006). Stimuli were constructed by system-
atically changing the colors of the six sections of a circle.
Typical stimuli were derived from the prototype stimuli
by changing the color of one section. Folglich, alle
typical stimuli in one category shared four of six colors
(66.7%). One circle in each category was constructed as
an exception. This exception shared zero to two colors
(0%, 16.7%, oder 33.3%) with the prototype and the typical
circles of its category but instead shared four to five colors
with the typical circles of the opposite category. In each
trial, participants had to categorize one circle by a left or
right button press and received a feedback about their
accuracy.
The circles were presented for 800 ms, before the
response options (Categories 1 Und 2) were displayed
concurrently with the stimulus for up to 1000 ms, until
a response was made (Figure 1B). The chosen category
was highlighted on the screen for 200 ms, followed by
a blank screen for 500 ms, and feedback was presented
für 1000 ms. As feedback, either “correct” or “wrong”
was presented, depending on whether the prior stimulus
was categorized correctly. If the response was too slow,
a reminder for fast responses was presented. Intertrial
intervals were jittered between 1000 Und 2000 ms. Par-
ticipants had to choose categories by button presses on
die linke (Category 1) or the right (Category 2) Ctrl keys
on a standard keyboard. The learning task was composed
von 420 trials in five blocks of 84 trials each, die Waren
intermitted by self-paced breaks. Each block encom-
bestanden 72 typical trials and 12 exceptions. Prototypes
and typical circles were pooled in the typical condition
for all analyses (Lech et al., 2016; Schenk et al., 2016).
To analyze behavioral performance, the accuracy for both
groups and for typical circles and exceptions was averaged
over each block. Darüber hinaus, we applied a computational
modeling procedure (described in detail in the following
section) to assess the use of abstraction- and exemplar-
based learning strategies in the categorization task.
Computational Modeling Procedures
To gain insight into how participants in each condition
learned these category sets, we fit two well-known com-
putational models to the data of stressed and control par-
ticipants. The first model was based on the prototype
model originally described by Minda and Smith (Minda
& Schmied, 2001, 2011; Schmied & Minda, 1998). This model
is ideally suited to model the current data as it was for-
mulated and tested with the same categories used in the
current study (Schmied & Minda, 1998).
In the prototype model, each category is represented by
a single prototype. Participants are assumed to abstract this
Paul et al.
801
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
J
Ö
C
N
_
A
_
0
1
2
4
1
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
J
Ö
C
N
_
A
_
0
1
2
4
1
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 1. Schematic illustration of stimulus categories and the sequence of events in each trial. (A) The paradigm consisted of two categories.
Typical circles were derived from the prototype of the respective category by changing one of the six colors. Exceptions were derived from the
prototype of the opposite category. (B) In jedem Versuch, participants had to categorize one circle by a button press. On the basis of feedback, which was
presented to the participants after their responses, they had to learn to categorize the circles correctly.
prototype from experience with individual exemplars, Und
they base a classification decision on the similarity of the
to-be-classified item to each prototype. The item is then
classified in accord with the most similar prototype. Der
prototype model predicts strong performance on proto-
typical and typical instances but poor performance on
exception items.
The model is formulated as follows. Erste, the distance
(D ) between the item i and the prototype P is calculated
by comparing the two stimuli along each weighted dimen-
sion k, as shown in Equation 1.
diP ¼
”
XN
k¼1
#1
R
wk xik − Pk
J
jr
(1)
The value of r corresponds to the distance metric.
When r = 1, the model uses a city-block distance metric,
which is appropriate for separable-dimension stimuli (Die
present case). When r = 2, the model uses a Euclidean dis-
tance metric, which is appropriate for integral-dimension
Reize. Attentional weights (w) vary between 0.0 (NEIN
attention) Und 1.0 (exclusive attention) and are con-
strained to sum to 1.0 across all the dimensions. The results
of these weighted comparisons are summed across the di-
mensions to get the distance between the item and the
prototype.
This distance (diP) between the item and the proto-
type is then converted into a measure of similarity
(ηiP), following Shepard (1987), by taking
η
iP ¼ e−cdiP
(2)
which gives a measure of similarity of an item i to proto-
type P. The exponent is distance (diP) multiplied by the
scaling or sensitivity parameter c, which is a freely esti-
mated parameter that reflects the steepness of the decay
of similarity around the prototype. Prototype A similarity
is then divided by the sum of Prototype A and Prototype
B similarity to generate the model’s predicted probability
of a Category A response P (RA) for stimulus (Si), als
shown in the probabilistic choice rule in Equation 3.
P RAð
jSiÞ ¼
η
η
iPA
þ η
iPA
iPB
(3)
802
Zeitschrift für kognitive Neurowissenschaften
Volumen 30, Nummer 6
We also fit an exemplar model to the data, which was
based on the Generalized Context Model of Nosofsky
(1986, 2011). Bei diesem Modell, the learner is assumed to
store representations of each category exemplar, Und
classification is based on the similarity of the to-be-
categorized item to the entire collection of each exemplar
in the category. The model is similar to the prototype
Modell, except that the comparisons are made between
exemplars. Erste, the distance between item i and exem-
plar j is calculated by comparing the two stimuli along
each weighted dimension k, as shown in Equation 4. Der
value of r again corresponds to 1 in diesem Fall.
dij ¼
”
XN
k¼1
(cid:2)
(cid:2)
wk xik − xjk
#1
R
(cid:2)
(cid:2)R
(4)
Attentional weights (w) vary between 0.0 (no attention)
Und 1.0 (exclusive attention) and are constrained to sum
Zu 1.0 across all the dimensions. The results of these
weighted comparisons are summed across the dimensions
to get the distance between the item and the exemplar.
This distance (dij) between the item and the exemplar
is then converted into a measure of similarity (ηij) von
taking
η
ij
¼ e−cdij
(5)
which gives a measure of similarity of an item i to exem-
plar j. As with the prototype model, c is the freely esti-
mated scaling parameter that reflects the steepness of
the decay of similarity among exemplars.
The similarity of the item to the summed similarity to
all the Category A exemplars is divided by the summed
similarity of the item to exemplars in Categories A and
B, as shown in the probabilistic choice rule in Equation 6.
X
PðRA Sij Þ ¼
X
η
ij
j2CA
η
ij
X
j2CA
þ
j2CB
(6)
η
ij
Although the models differ in terms of their repre-
sentational assumptions (abstracted prototype vs. stored
exemplars), they are equivalent in terms of free param-
eters, attentional assumptions, and decision rules. Jede
model has a set of attentional weight parameters that
correspond to the number of dimensions in a task (in diesem
Fall, six dimensions) along with a psychological scaling
parameter that corresponds to how close or distant the
prototypes or exemplars are in psychological space. Weil
the attentional weights are constrained to sum to 1.0 (d.h.,
full attention), it leaves five free attentional parameters
and the single, unconstrained scaling parameter for a
total of six free parameters for each model.
Our analyses were based on fitting of each model to
classification probabilities produced by each individual
participant in each condition and at each block. Der
models were fit with a hill-climbing algorithm that adjusted
the model’s parameters to minimize the root mean square
deviation (RMSD) between the observed data and the
model’s predictions (Minda & Schmied, 2001). To find the
best-fitting parameter settings of each model, a single
parameter configuration (six attention weights and one
scaling parameter) was chosen randomly and the pre-
dicted categorization probabilities for each of the 14 stim-
uli were calculated according to that configuration. Der
RMSD was calculated as shown in Equation 7.
S
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
X
Þ2
D
N
i¼1
Oi − Pi
N
RMSD ¼
(7)
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
J
Ö
C
N
_
A
_
0
1
2
4
1
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
The RMSD between the observed (Oi) and predicted
(Pi) probabilities was then minimized with an algorithm
that made a small adjustment to the provisional best-fitting
parameter settings and chose the new settings if they pro-
duced a better fit (d.h., a smaller RMSD between predicted
and observed performances). On each iteration of the algo-
rithm, a weight parameter was adjusted by 0.01 (bounded
by 0.00–1.00), or a scaling parameter was adjusted by 0.1
(bounded by 0.00–20.00). The algorithm continued to
adjust until the fit could not be minimized further. To
ensure that local minima were not a problem, the fitting
procedure was repeated five times by choosing different
random starting configurations of the model and hill-
climbing from there. We chose the best-fitting parameters
of the multiple fittings. The data for each block and each
participant were fit independently, and each model (proto-
type or exemplar) was fit separately. This method, origi-
nally devised by Smith and Minda (1998), provides a
static snapshot of performance, rather than an estimate of
learning.
The resulting fit index (RMSD) from the model-fitting
procedure provides information about how well (or not)
each model fits the data of participants in each condition.
On the basis of earlier work by Smith and Minda (1998)
and more recent work by Minda and Smith (2011) Und
Schenk et al. (2016), we assumed that both models
may fit moderately well early in learning but the exemplar
model should provide a better fit later in the learning
Phase, because participants should have learned to clas-
sify all the exemplars, including the exception items.
Jedoch, Smith and Minda (1998) also noted that the
prototype model often fits the data better than the exem-
plar model early on, because many participants find it
easy to learn to classify the prototype and typical items
and systematically misclassify the exception items. In
the present case, an advantage for the prototype model
over the exemplar model might indicate that the partici-
pants have not been able to commit sufficient cognitive
resources to learn the exceptions.
EEG Recording and Processing
Scalp EEG was recorded from 30 passive Ag/AgCl elec-
trodes, which were distributed according to the 10–20 sys-
tem (Pivik et al., 1993). Data were recorded at a sampling
Paul et al.
803
rate of 500 Hz by a 32-channel BrainAmp Standard AC
amplifier (Brain Products, Gilching, Deutschland), with a time
constant of 10 Sek. An electrode at the midfrontal position
FPz was affixed to ground the participants, and data were
referenced to linked mastoids. Impedances were kept
below 10 kΩ.
Eye blinks were removed from continuous data using an
independent component analysis as implemented in Brain
Vision Analyzer 2 (Brain Products, Gilching, Deutschland; Lee,
Girolami, & Sejnowski, 1999). For each participant, eins
independent component with a symmetrical frontal pos-
itive topography was removed, before the data were
back-transformed. The successful removal of eye blinks
was confirmed by visual inspection. Further analyses
were performed using the FieldTrip toolbox (Oostenveld,
Fries, Maris, & Schoffelen, 2011) and MATLAB 2016a
(The MathWorks, Inc., Natick, MA).
Data were filtered with a 0.5-Hz high-pass, zero-phase
Butterworth IIR filter and a band-stop filter from 48 Zu
52 Hz to eliminate line noise. Continuous data were
epoched from −1.5 to 3 sec around feedback presentation.
Epochs containing residual artifacts were removed during
careful visual inspection. Subsequently, time–frequency
decomposition was performed using 59 complex Morlet
wavelets from 1 Zu 30 Hz, each having a width of five
cycles. Power values were averaged across trials in each
of the four following conditions: typicals/negative feed-
back, typicals/positive feedback, exceptions/negative
Rückmeldung, and exceptions/positive feedback. Teilnehmer
with less than 10 trials in any condition were excluded
from further analyses. On average, typicals/negative feed-
back contains 63.44 (SEM = 5.29) Versuche, typicals/positive
feedback contains 244.34 (6.37) Versuche, exceptions/negative
feedback contains 26.75 (1.30) Versuche, and exceptions/
positive feedback contains 23.25 (1.16) Versuche. Nachher,
relative signal change to the condition-averaged base-
line period (500 Zu 100 msec before stimulus presenta-
tion) was calculated for each condition to quantify
feedback-related signal changes (Pfurtscheller & Aranibar,
1977). The baseline theta power did not differ between
groups. To analyze differences in theta power, we aver-
aged the power over the 200- to 600-msec postfeedback
time interval (Chen, Zheng, Han, Chang, & Luo, 2017;
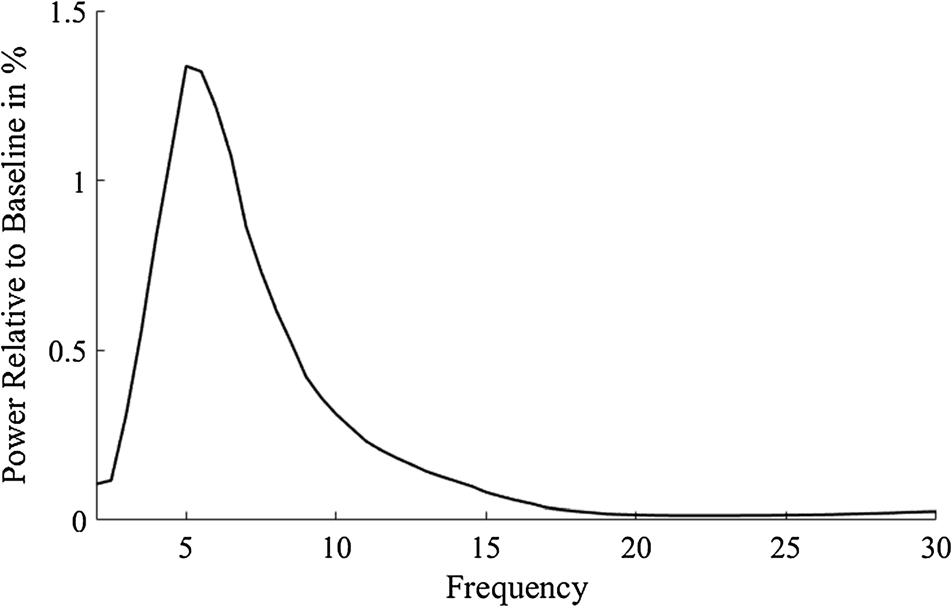
Cunillera et al., 2012) and over the frequencies of 5–
5.5 Hz. The frequencies of interest were determined
using the difference between negative and positive
feedback in averages across all groups and all conditions
(Figur 2; Leicht et al., 2013). Group and condition
differences were then analyzed in these peak frequencies,
which were determined from group and condition
averages.
Statistical Analyses
For the statistical analyses of the salivary cortisol concen-
trations and the behavioral data, repeated-measures
ANOVAs were applied. All ANOVAs included the
Figur 2. Power spectrum of the difference in the relative increase
(200–600 msec postfeedback interval) to baseline between negative and
positive feedback at electrode FCz. The frequencies of interest were
defined as the frequencies with the maximal difference between
negative and positive feedback pooled across both groups and both
Bedingungen.
between-participant factor Group (stress and control).
Group effects in salivary cortisol concentrations were an-
alyzed statistically using repeated-measures ANOVA with
the between-participant factor Group (stress and control)
and the within-participant factor Time (−1, +1, +20, Und
+55). The analysis of the behavioral data included the
within-participant factors Stimulus type (typical and
exceptional items) and Block (1–5). The Greenhouse–
Geisser correction was applied in cases of violations to
the assumption of sphericity, and ε are reported. Sig-
nificant interactions were resolved with Bonferroni-
corrected post hoc pairwise t tests.
Differences in the subjective stress ratings and the
cardiovascular measures of the stress response between
the groups were tested by means of a multivariate
ANOVA. Bonferroni-corrected post hoc pairwise compar-
isons were performed to test group differences on single
subjective scales. The alpha level of .05 was applied to all
parametric tests.
For the statistical analysis of the time–frequency data,
nonparametric cluster-based permutation tests were ap-
plied to account for alpha error accumulations in the con-
text of multiple comparison testing (Maris, 2012; Maris &
Oostenveld, 2007). This cluster-based permutation test
involves two steps: In the first step, spatially coherent
clusters of electrodes exceeding a first level t threshold
(α = .05) are identified, and summed t values in each
cluster are returned as test statistic. In the second step,
this cluster-based test statistic is compared with a null dis-
tribution of the test statistic obtained by repeating Step 1
on data with randomly permuted condition affiliations for
1,000 Iterationen. For clusters, which reached the cluster α
threshold, summed t values of the electrodes included in
the cluster (tsum) and cluster p values are reported. Addi-
tional to the whole electrode space covering cluster sta-
tistics, single-electrode t and p values of the frontocentral
804
Zeitschrift für kognitive Neurowissenschaften
Volumen 30, Nummer 6
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
J
Ö
C
N
_
A
_
0
1
2
4
1
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
electrode FCz are reported based on prior studies (z.B.,
Cavanagh, Figueroa, Cohen, & Frank, 2012; van de Vijver
et al., 2011). Significant interactions in the cluster-based
permutation tests were resolved using Wilcoxon signed-
rank tests.
ERGEBNISSE
Subjective Stress Response
The subjective ratings revealed a successful stress induction
by the SECPT (F(4, 27) = 31.34, P < .001, Wilk’s Λ = 0.177,
ηp
2 = .82). Stressed participants experienced more dis-
comfort ( p < .001, d = 2.55), more pain ( p < .001,
d = 3.54), and more stress ( p < .001, d = 2.15) and
reported more difficulty to keep the hand immersed
( p < .001, d = 3.38) during the treatment (Table 1).
Cardiovascular Stress Responses
Increases in the blood pressure and heart rate of stressed
participants during the stress induction revealed an acti-
vation of the sympathetic nervous system (F(6, 116) =
15.33, p < .001, Wilk’s Λ = 0.311, ηp
2 = .44). Elevations
of the systolic (F(2, 60) = 30.92, p < .001, ηp
2 = .508) and
diastolic (F(2, 60) = 39.06, p < .001, ηp
2 = .566) blood
pressure and of the heart rate (F(2, 60) = 14.75, p <
.001, ηp
2 = .330) were confirmed by Time × Group inter-
actions (Table 1).
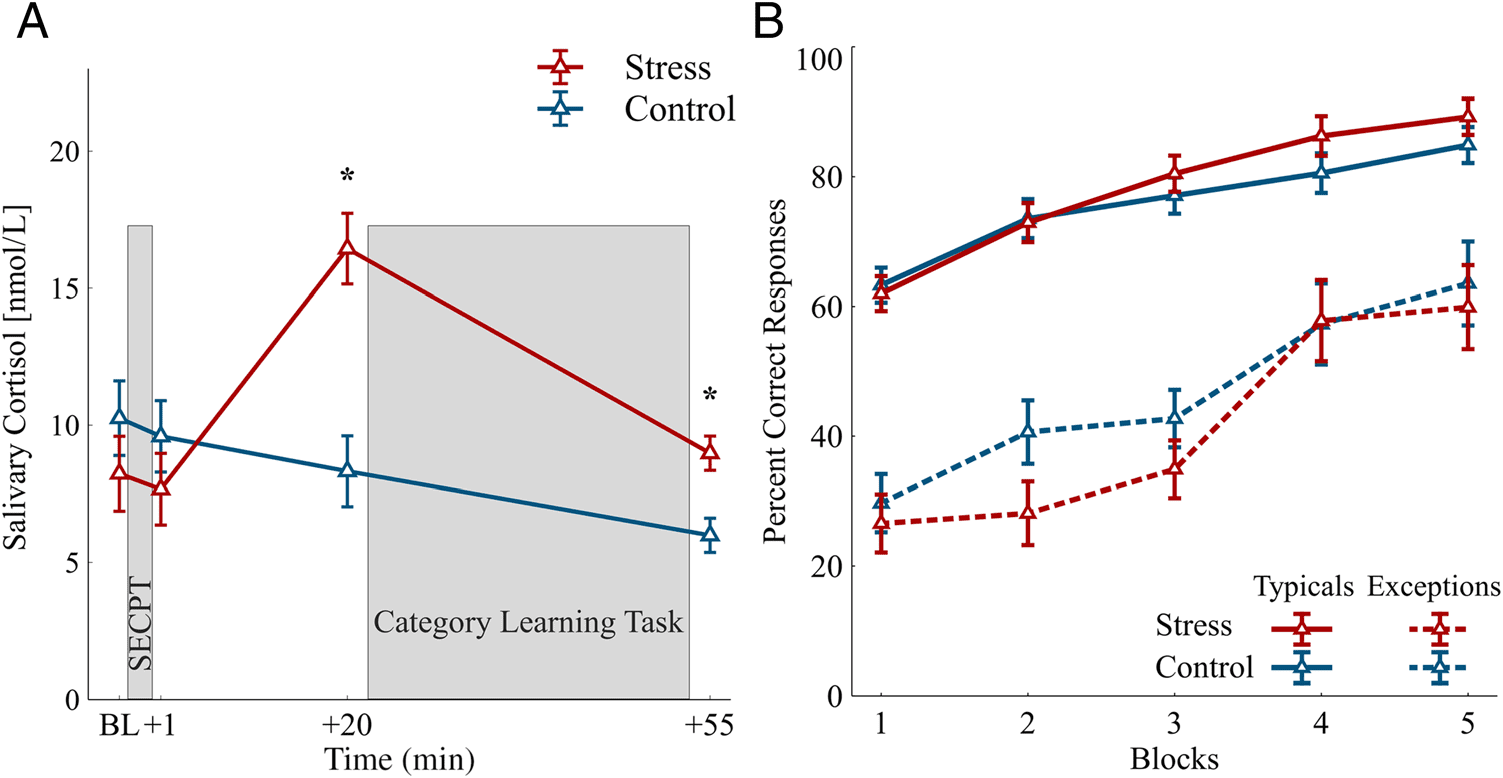
Salivary Cortisol Concentrations
The stress group shows an elevation of salivary cortisol
concentrations 20 min after the SECPT, whereas cortisol
concentrations decrease over the course of the experiment
in the control group (Figure 3A). The successful stress
induction was confirmed by a significant Group × Time
interaction (F(3, 90) = 26.3, p < .001, ηp
2 = .467) and a main
effect of Time (F(3, 90) = 19.86, p < .001, ηp
2 = .398).
The cortisol concentrations of the stress group were
higher compared with those of the control group 20 min
(t(30) = −4.45, p < .001, d = 1.57) and 55 min (t(30) =
−3.38, p = .002, d = 1.20) after the treatment.
Learning Performance
The learning performance was assessed by calculating the
percentage of correct responses of five blocks consisting
of 72 typical trials or 12 trials with exceptions. Both stim-
ulus classes, typical circles and exceptions, were success-
fully learned by the participants (Figure 3B), which was
confirmed by a main effect of Block (F(2.20, 66.01) =
2 = .596, ε = .550). The performance
44.21, p < .001, ηp
was better for typical circles throughout the whole task
(main effect Stimulus type: F(1, 30) = 192.15, p <
.001, ηp
2 = .865), but the increase in the performance
was larger for the exceptions (last block − first block =
Table 1. Differences in the Subjective and Cardiovascular
Stress Responses between the Stress and Control Groups
Subjective stress response
Discomfort
Pain
Stress
Control
Stress
4.38 (2.03)
48.13 (5.72)**
0.00 (0.00)
60.00 (5.99)**
5.00 (2.24)
45.00 (6.19)**
Difficulty to keep hand immersed
1.88 (1.88)
57.50 (5.52)**
Systolic blood pressure (mm Hg)
Pretreatment
123.27 (2.74) 125.92 (4.44)
During treatment
124.63 (2.58) 148.48 (4.05)**
Posttreatment
120.06 (2.65) 128.67 (3.73)
Diastolic blood pressure (mm Hg)
Pretreatment
71.20 (2.82)
66.04 (2.88)
During treatment
73.98 (2.62)
87.48 (2.24)**
Posttreatment
66.44 (2.71)
69.27 (2.97)
Heart rate (BPM)
Pretreatment
70.38 (2.65)
67.27 (3.30)
During treatment
69.46 (3.24)
75.19 (3.61)*
Posttreatment
68.48 (2.49)
65.46 (3.30)
Differences in subjective ratings were tested in planned t tests; differ-
ences in the cardiovascular responses were tested with Bonferroni-
corrected post hoc t tests. Values represent mean (±SEM ).
*p < .1.
** p < .001.
33.6%) compared with typical circles (24.44%; Stimulus
type × Block interaction: F(2.89, 86.80) = 3.960, p =
.012, ηp
2 = .117, ε = .723).
Stress, however, did not influence the learning perfor-
mance, and neither the main effect of Group (F(1, 30) =
0.20, p = .659, ηp
2 = .007) nor any interaction including
the factor Group revealed an influence of stress on the
performance (Stimulus type × Group: F(1, 30) = 2.58,
p = .119, ηp
2 = .079; Block × Group: F(2.20, 66.01) =
2 = .032, ε = .550; Stimulus type ×
1.00, p = .381, ηp
Block × Group: F(2.89, 86.80) = 0.36, p = .775, ηp
2 =
.012, ε = .723).
A post hoc power analysis using G*Power ( Version
3.1.9.2; Faul, Erdfelder, Lang, & Buchner, 2007) revealed
that a medium-sized Stimulus type × Group interaction
effect (ηp
2 = .10), which was expected based on stress-
induced changes in category learning in previous studies
(Schwabe & Wolf, 2012), would have been detectable
with a power of 1 − β = .81 (α = .05; the average
Paul et al.
805
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. (A) Mean salivary
cortisol concentrations at
baseline (BL) as well as +1, +20,
and +55 min after the SECPT/
control procedure. Cortisol
concentrations were elevated in
the stress group compared with
the control group 20 and 55 min
after the SECPT. *p < .01. Error
bars represent SEM. (B) Mean
percent correct responses over
the course of 30 blocks. The
performance improved over the
course of the experiments in
typical category members and in
exceptions as well as in both
groups. Learning performance
did not differ between the
groups. Error bars represent
SEM.
correlation between repeated measures in our sample
was r = .18).
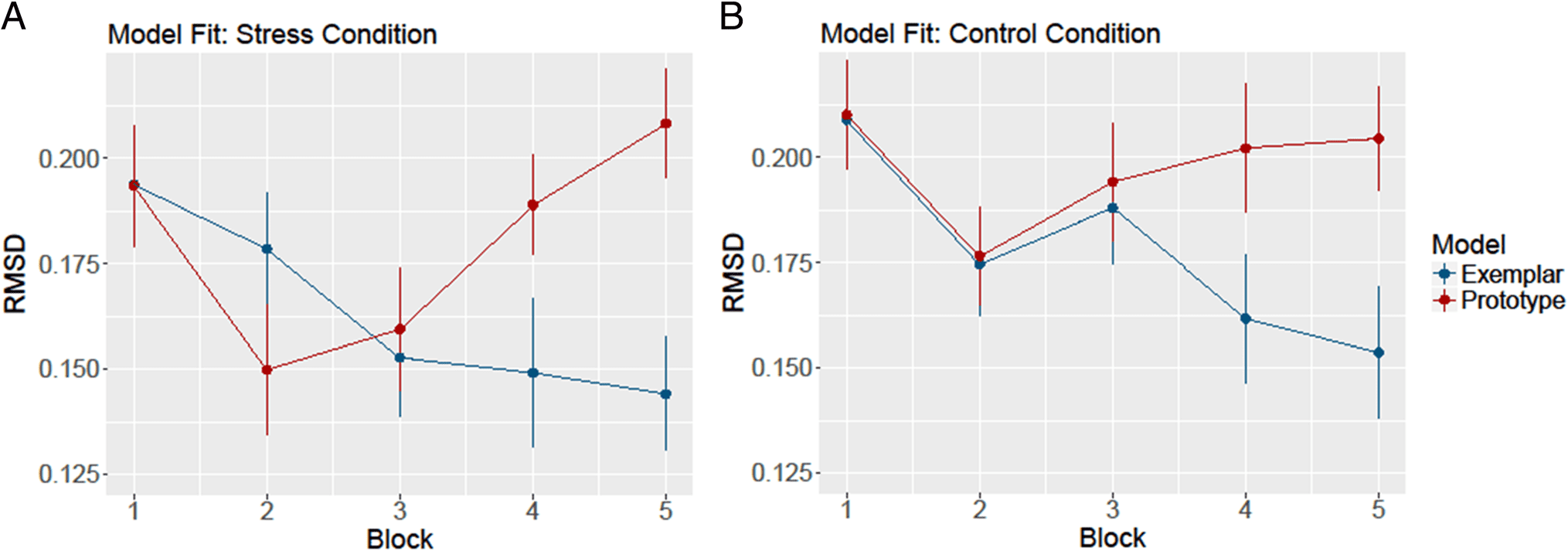
Computational Modeling
To examine the relative differences in model fit, we ob-
tained the best fit (the RMSD) for each model fitting the
data of each participant at each block, and we averaged
across participants to obtain the average fit of each
model. These average fits are plotted in Figure 4.
Figure 4A shows the relative fits of the prototype model
and the exemplar model for the participants in the stress
condition, and Figure 4B shows the relative fit of the
models when fitting data from the control condition. In
both cases, the exemplar model fits better (has a lower
RMSD) than the prototype model at the end of the learn-
ing phase. However, Figure 4A shows that the prototype
model had an early advantage over the exemplar model
at the second block for participants in the stress condi-
tion. This pattern was not observed in the data of the
participants in the control condition, suggesting no early
difficulty with exception items.
To examine these effects more directly, we conducted
a 2 (Model) × 5 (Block) repeated-measures ANOVA on
the RMSD values for each learning condition. For the
Stress condition, we found find a main effect for Model
(F(1, 15) = 6.90, p = .019, η2 = .315), but not for Block
(F(4, 60) = 1.95, p = .115, η2 = .115). We also observed
a significant interaction between Block and Model (F(4,
60) = 6.16, p < .001, η2 = .291). To explore this inter-
action, we conducted five paired t tests (prototype fit vs.
exemplar fit) at each block, using a Bonferroni correction
to adjust the alpha level to .01 for the five comparisons.
The models were not significantly different at the first to
fourth blocks (t(15) = −0.03, −1.78, 0.70, and 2.25; p =
.973, .095, .494, and .040, respectively), but the exemplar
Figure 4. A shows the average fit (RMSD) across participants for the prototype and exemplar model fitting the data of participants in the stress
condition. A lower RMSD indicates lower prediction error and a better fit for the model. B shows the average RMSD across participants for the
prototype and exemplar model fitting the data of participants in the control condition. Error bars denote SEM for the calculated means.
806
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
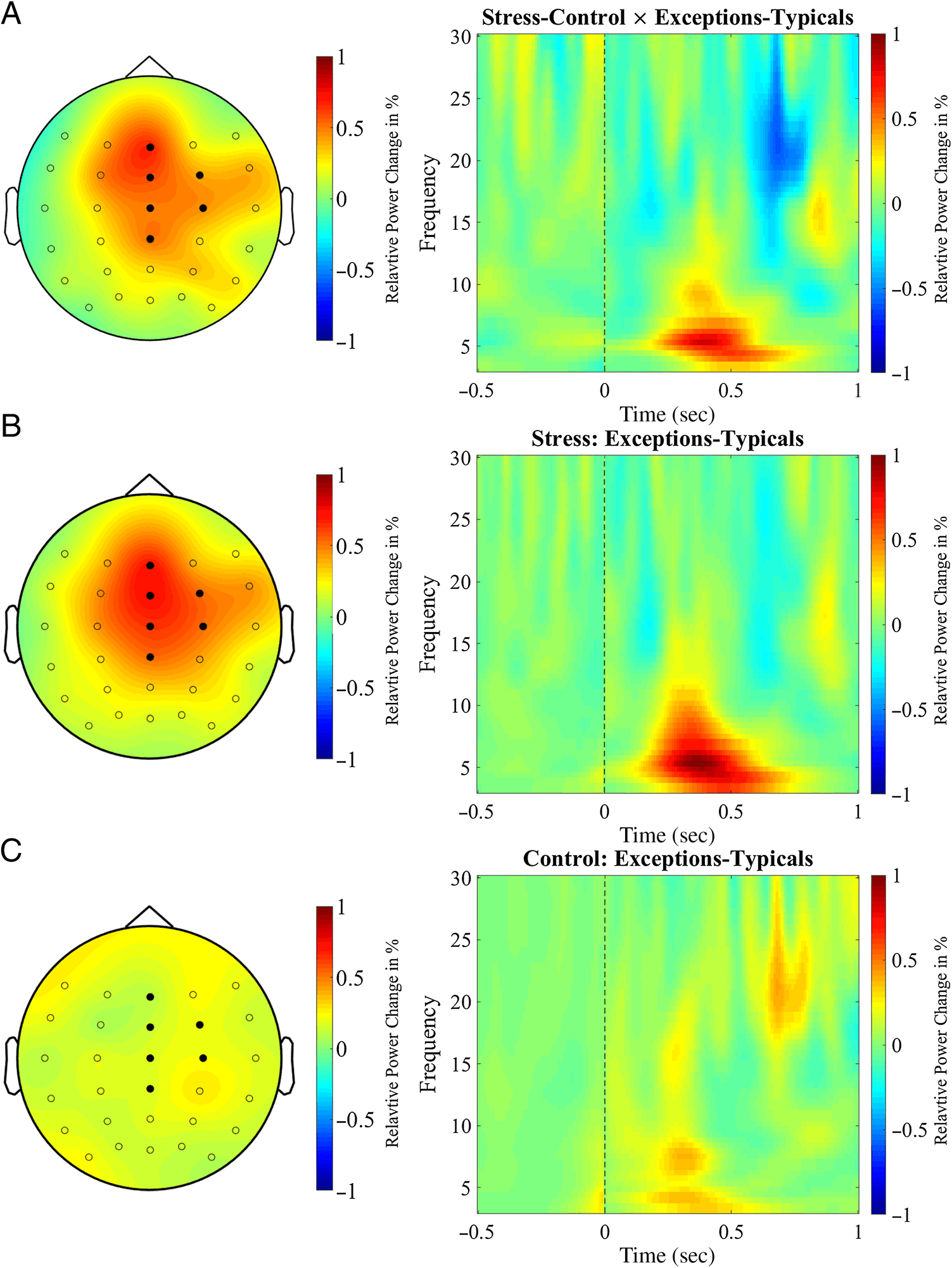
Figure 5. Topographical maps
of the relative theta power
(5–5.5 Hz, 200–600 msec
postfeedback interval) and
time–frequency plots at the
electrode FCz relative to the
presentation of negative
feedback. On the topographical
maps, filled circles represent
electrodes of the cluster that
revealed the significant Group ×
Stimulus type interaction. In
the time–frequency plots, the
prestimulus baseline interval
(left of the dotted line) and the
postfeedback activity (right of
the dotted line) are shown.
(A) A larger difference in
the theta power between
exceptions and typical circles
was found in the stress group
compared with the control
group. (B) In the stress group,
larger theta power was found
for exceptions than for typical
circles. (C) The control group
did not show a difference
in the theta power between
exceptions and typical circles.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
model fit significantly better than the prototype model in
the fifth block, t(15) = 3.57, p = .003. Thus, although the
prototype model appeared to have some advantage over
the exemplar model in the second block, the advantage
was not significant. The exemplar model did, however,
significantly outperform the prototype model at the end
of the study, and the main effect of model confirms that
the exemplar model was fitting better overall.
For the Control condition, we again found a main effect
for Model (F(1, 15) = 12.51, p = .003, η2 = .455), but not
for Block (F(4, 60) = 2.00, p = .106, η2 = .118). We also
observed a significant interaction between Block and
Model (F(4, 60) = 3.54, p = .012, η2 = .191). To explore
this interaction, we conducted five paired t tests (proto-
type fit vs. exemplar fit) at each block, using a Bonferroni
correction to adjust the alpha level to .01 for the five com-
parisons. The models were not significantly different at
the first to third blocks (t(15) = −0.14, 0.34, and 0.62;
p = .889, .738, and .543, respectively), but in the fourth
block, the exemplar model fits significantly better than
the prototype model (t(15) = 2.95, p < .010). In the fifth
block, the t test revealed a trend toward the same
direction (t(15) = 2.58, p = .021).
FMT
To assess whether the neural processes of feedback-
based learning differ between the groups and between
Paul et al.
807
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
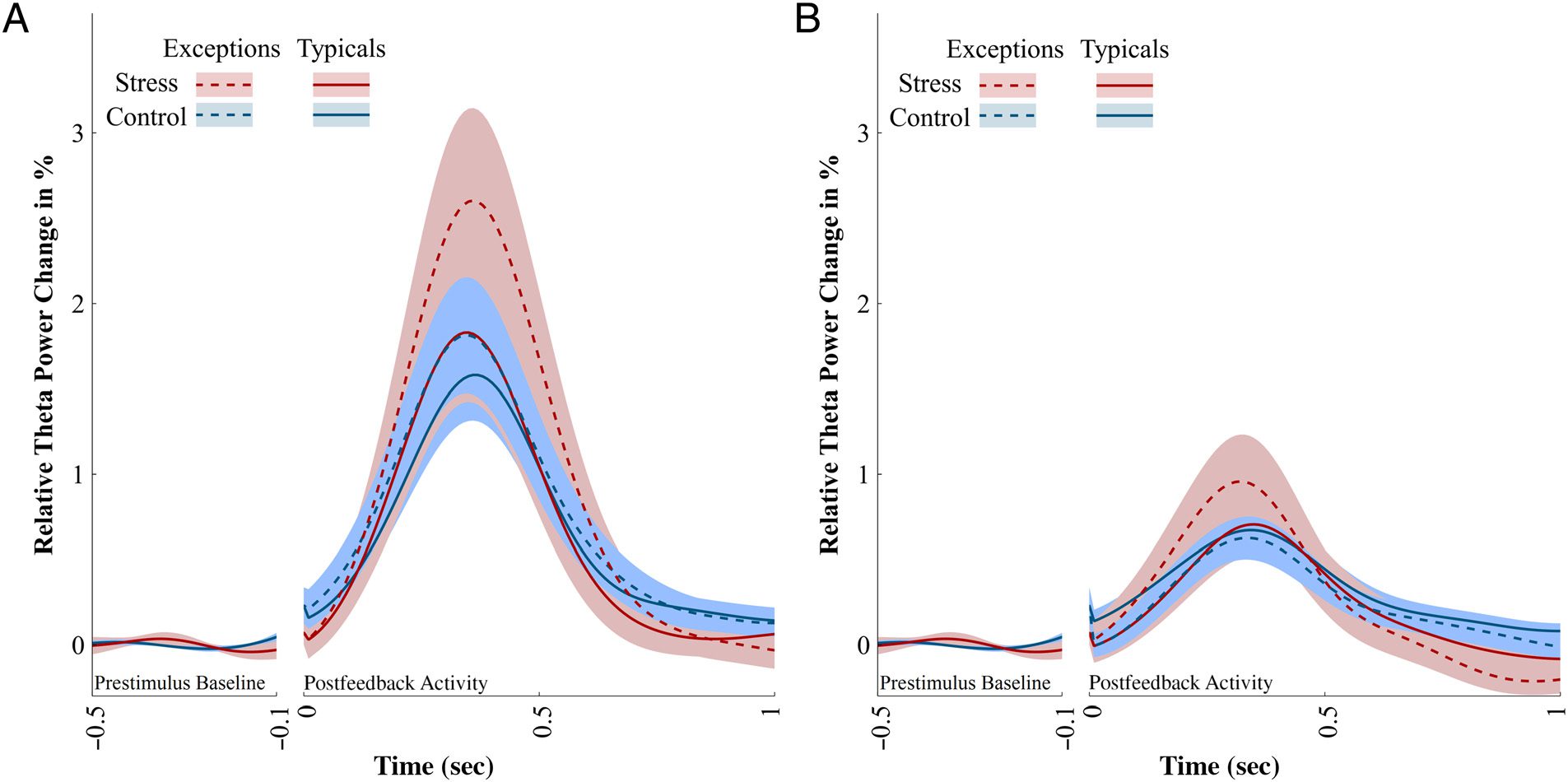
Figure 6. Theta power values (5–5.5 Hz) at electrode cluster Fz, FCz, FC4, Cz, C4, and CPz. The postfeedback theta power is plotted relative to
the condition-averaged prestimulus baseline. A shows theta power after the presentation of negative feedback, and B shows theta power after
positive feedback. Stress elevated the theta in exceptions after negative feedback. After positive feedback, theta power differs descriptively, but
nonsignificantly, between groups or between exceptions and typical circles. Shaded areas represent SEM.
the learning of typical category members and exceptions,
we analyzed the FMT power of feedback processing.
First, the frequency window of the feedback-related theta
power increase was determined by pooling the data of
both groups and both conditions (Figure 2) for each fre-
quency between 1 and 30 Hz and selecting the frequency
showing the highest feedback-related change relative to
baseline at electrode FCz. The highest feedback-related
increase in theta power was found at 5–5.5 Hz. To inves-
tigate whether learning of typical and exceptional stimuli
is related to different processes depending on stress
level, differences in theta power between typical and ex-
ceptional trials were contrasted between groups. A clus-
ter permutation test revealed that the Group × Stimulus
type × Feedback valence interaction was not significant
(all ts ≤ 1.6, all ps = 1). On the basis of previous studies,
which showed that stress influences predominantly
the use of negative feedback in reward learning and
decision-making (Park, Lee, & Chey, 2017; Petzold,
Plessow, Goschke, & Kirschbaum, 2010), we focused
the analysis on negative feedback trials, where an effect
of stress was expected. Indeed, a cluster permutation test
revealed a significant Group × Stimulus type interaction
(tsum(30) = 13.90, p = .042) of FMT power (5–5.5 Hz)
200–600 msec after negative feedback at a frontocentral
cluster of electrodes (Figure 5A), including the fronto-
central electrode FCz (t(30) = 2.25, p = .042). This result
reveals a difference in theta power during feedback pro-
cessing during learning of typical category members and
exceptions between the stress and control groups. To
characterize this effect in depth, Wilcoxon signed-rank
tests were used to contrast the theta power of this fron-
tocentral electrode cluster in typical and exceptional
stimuli separately for the stress and control groups.
These follow-up tests demonstrated that stressed partic-
ipants show an elevated theta power for trials with excep-
tions, compared with trials with typical circles (Z = 3.00,
p = .003, r = .750; Figure 5B). This effect of stronger in-
creases in theta power for exceptional compared with
typical stimuli was missing in the control group (Z =
0.88, p = .379, r = .220; Figure 5C). These results indi-
cate that the stress group shows an elevated FMT power
after negative feedback, which is specific to the learning
of exceptions (Figure 6A).
We focused the EEG analysis on negative feedback and
found differences between the groups and stimulus types
in frontal theta oscillations only in negative feedback.
After positive feedback, the Group × Stimulus type inter-
action did not reach significance at any electrode site or
cluster (all ts ≤ 1.88, all ps = 1; Figure 6B). Accordingly,
neither in the stress group (Z = 0.62, p = .535, r = .155)
nor in the control group (Z = −1.40, p = .163, r =
−.349) did the typical circles and exceptions differ with
respect to FMT oscillations after positive feedback.
DISCUSSION
In the current study, we examined the influence of acute
stress on neural correlates of feedback processing during
category learning of typical category members and excep-
tions. Participants were exposed to either a socially eval-
uative stress condition or a nonstressful control situation,
808
Journal of Cognitive Neuroscience
Volume 30, Number 6
before conducting a feedback-based category learning
task. The successful stress induction by the SECPT was
demonstrated by increases in blood pressure, heart rate,
and salivary cortisol concentrations. Both groups showed
no substantial differences in behavioral performance. The
stress and control groups successfully learned to assign
typical stimuli and exceptions to the two categories and
categorized typical category members more accurately
than exceptions. Computational modeling revealed
descriptively an increased use of the abstraction-based
learning strategy in the early learning phase after stress.
In the late learning phase, both groups made use of the
exemplar learning strategy. Moreover, FMT power in-
creases during negative feedback differed between the
stress and control groups. During the learning of excep-
tions, midfrontal theta power after negative feedback in-
creased in the stress group compared with the control
group. After positive feedback, there was a descriptive
but nonsignificant difference in the same direction.
Frontal theta oscillations are involved in the process-
ing of feedback and are related to behavioral adapta-
tion (Cohen et al., 2011; Cavanagh et al., 2010).
Accordingly, an elevation of frontal theta power after
stress suggests an increased involvement of frontal
feedback processing in the learning of exceptions. This
is in line with recent results of an increased FRN after
stress ( Wirz et al., 2017; Glienke et al., 2015), which is
the time-locked reflection of feedback processing
(Cavanagh, Zambrano-Vazquez, & Allen, 2012; Holroyd
& Coles, 2002).
Other previous results, however, demonstrated an
influence of anxiety on the FMT power (Mizuki et al.,
1992; Mizuki, Hashimoto, Tanaka, Inanaga, & Tanaka,
1983) and illustrate a role of FMT oscillations in work-
ing memory maintenance (Hsieh & Ranganath, 2014)
and cognitive control processes (Cavanagh & Frank,
2014; Cohen, 2014). Pastötter, Dreisbach, and Bäuml
(2013), for instance, reported the largest FMT power
in control-demanding incongruent task conditions.
The modeling analysis indicates that some participants
in the stress condition may have relied more heavily on
the abstracted prototypes early in the experiment relative
to participants in the control condition, although this
conclusion can only be tentative as the individual post hoc
t tests were not significant. This may suggest that
stressed participants had fewer cognitive resources avail-
able to learn the exceptions to the prototype, although
participants in the stress condition still showed a clear,
late advantage for exemplar learning, perhaps reflecting
a recovery later in the learning phase.
The notion that stress could affect the learning of cer-
tain kinds of categories is consistent with studies demon-
strating that performing a concurrent task that reduced
the available working memory (Miles & Minda, 2011) or
resource-depleting tasks (Minda & Rabi, 2015) impaired
learning categories that required hypothesis testing and a
rule selection, but not for tasks that relied on purely as-
sociative learning. The latter is assumed to be analogous
to the prototype abstraction process (Minda & Miles,
2010). It is reasonable that stress might have reduced
the availability of cognitive resources enough to allow
interference with exception learning but not interference
with prototype learning. Additional work is needed, how-
ever, with other category sets, such as those used by re-
searchers showing impaired performance by older adults
as a function of reduced working memory resources
(Rabi & Minda, 2016). Alternatively, future research could
also examine deterministic versus probabilistic respond-
ing and the possibility that the stress induction might
have altered the choice strategy. Certain formulations
of Nosofsky’s Generalized Context Model can model this
difference with a single additional parameter, but this
same change cannot be instantiated in the prototype
model without affecting the operation of the scaling pa-
rameter (Minda & Smith, 2001; Smith & Minda, 1998,
2000). Other possibilities for future work could examine
alternative models, such as the drift diffusion class of
models to make specific predictions about choice behav-
ior and response time (Pedersen, Frank, & Biele, 2017).
In the current study, stress did not influence learning
success. This is in line with previous studies, in which
stress did not influence category learning performance
per se but modulated the use of learning strategies and
their neuronal basis ( Wirz et al., 2017; Schwabe et al.,
2013; Schwabe & Wolf, 2012). In light of the behavioral
results, increases in theta power could reflect the in-
creased recruitment of PFC regions, which have been
linked with the abstraction of commonalities of category
members (Mack, Preston, & Love, 2017; Pan & Sakagami,
2012). Medial prefrontal regions, such as the dACC and
the dorsomedial PFC, have been shown to use theta os-
cillations to recruit regions in the lateral PFC in the real-
ization of learning (Smith et al., 2015; van de Vijver et al.,
2011). An alternative explanation could be that the early
advantage of abstraction learning after stress increases the
need of cognitive control, which has also been linked to
midfrontal theta oscillations (Cavanagh & Frank, 2014), to
switch to the exemplar learning strategy. Previous studies
demonstrated that successful learning of exceptions relies
on hippocampus activations during learning (Lech et al.,
2016; Davis, Love, & Preston, 2012). It is known that
stress impairs hippocampus-dependent memory pro-
cesses, such as long-term memory retrieval ( Wolf,
2017) as well as long-term memory encoding (Shields,
Sazma, McCullough, & Yonelinas, 2017). We suggest
that, after stress, an increased involvement of medial
prefrontal regions might be responsible for adequate
learning of exceptions in the current category learning
task compensating a potential decline in hippocampal
functionality.
Increases in electrophysiological correlates of the feed-
back processing after stress are thought to be related to
an increased availability of dopamine in the striatum and
PFC (Grace, 2016; Holly & Miczek, 2016), which is closely
Paul et al.
809
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
linked to the processing of feedback (Frank, Seeberger,
& O’Reilly, 2004; Holroyd & Coles, 2002; Schultz &
Dickinson, 2000). Other studies, however, found an im-
pairing effect of stress and glucocorticoids on the
feedback-related activity of the reward system (Porcelli
& Delgado, 2017; Montoya, Bos, Terburg, Rosenberger,
& van Honk, 2014; Ossewaarde et al., 2011) and on the
FRN and frontal theta power (Banis, Geerligs, & Lorist,
2014; Banis & Lorist, 2012). These inconsistent findings
of stress effects on electrophysiological correlates of
the feedback processing might depend on the timing
of the stress, because enhancing effects of stress on the
FRN ( Wirz et al., 2017; Glienke et al., 2015) and theta
power were found in the aftermath of stress, when corti-
sol levels are elevated (de Kloet, Joëls, & Holsboer,
2005), whereas attenuations were found when partici-
pants were stressed during the task (Banis et al., 2014;
Banis & Lorist, 2012). Future studies are needed, which
explicitly address the issue of timing of the stressor, sim-
ilar to previous experiments in the domain of decision-
making (e.g., Pabst, Brand, & Wolf, 2013).
We demonstrated an increase of the frontal theta
power by stress after negative feedback. This is in line
with the finding that FMT differentiates between positive
and negative feedback, such that negative feedback elicits
larger increases in theta power than positive feedback
(Cohen, Elger, & Ranganath, 2007). Accordingly, theta
oscillations are thought to reflect the processing of negative
feedback and to be related to learning based on negative
feedback (Andreou et al., 2017; Mas-Herrero, Ripollés,
HajiHosseini, Rodríguez-Fornells, & Marco-Pallarés, 2015;
van de Vijver et al., 2011).
In the current study, we could show that stress ele-
vates the feedback-related theta power. The findings
have implications for future fMRI or simultaneous EEG/
fMRI studies (Hauser et al., 2015), which have to eluci-
date the relationship and timing of the learning systems
involved in category learning. Importantly, future studies
have to investigate the neural source of the observed
increase in frontal theta oscillations and whether this
increase is accompanied by reduced hippocampal
activations.
Limitations
There are some limitations to the current study that need
to be addressed. First, 15 participants had to be excluded
from the study sample because of missing cortisol re-
sponses or too few trials for the EEG analysis. As a con-
sequence, the absence of a significant stress effect on
category learning performance and used learning strategy
might be due to the somewhat reduced statistical power.
Finally, the current study controlled for confounding gen-
der differences by testing only male participants. Former
studies reported sex differences in the responsiveness to
acute stressors (Reschke-Hernández, Okerstrom, Bowles
Edwards, & Tranel, 2017), in the effects of stress on emo-
tional learning (Merz & Wolf, 2017), and in the effects of
glucocorticoids on the reward system (Kinner, Wolf, &
Merz, 2016). Future studies have to elucidate possible
sex differences in the effects of stress on category learn-
ing and frontal theta power.
Conclusion
In summary, the current study reveals that stress has an
enhancing effect on frontal theta oscillations in the pro-
cessing of negative feedback during the category learning
of exceptions. These results illustrate that stress modu-
lates the neural basis of the learning of exceptions. The
enhanced frontal theta oscillations might reflect a com-
pensatory mechanism allowing preserved categorization
performance of exceptions in the immediate aftermath
of stress.
Acknowledgments
This research was supported by the Deutsche Forschungsge-
meinschaft Projects B4, B8, and B11 of the Collaborative Research
Centre (SFB) 874 “Integration and Representation of Sensory Pro-
cesses.” We thank Osman Akan, Alessa de Vries, Farina Helmke,
Eve Hessas, Alexander Quent, and Theresa Wortmeier for assis-
tance with data collection.
Reprint requests should be sent to Oliver T. Wolf, Department of
Cognitive Psychology, Institute of Cognitive Neuroscience, Ruhr-
University Bochum, Universitätsstr. 150, Bochum, Germany,
44780, or via e-mail: oliver.t.wolf@rub.de.
REFERENCES
Alexander, W. H., & Brown, J. W. (2011). Medial prefrontal
cortex as an action–outcome predictor. Nature
Neuroscience, 14, 1338–1344.
Andreou, C., Frielinghaus, H., Rauh, J., Mußmann, M., Vauth, S.,
Braun, P., et al. (2017). Theta and high-beta networks for
feedback processing: A simultaneous EEG–fMRI study in
healthy male subjects. Translational Psychiatry, 7, e1016.
Antzoulatos, E. G., & Miller, E. K. (2011). Differences between
neural activity in prefrontal cortex and striatum during
learning of novel abstract categories. Neuron, 71, 243–249.
Antzoulatos, E. G., & Miller, E. K. (2014). Increases in functional
connectivity between prefrontal cortex and striatum during
category learning. Neuron, 83, 216–225.
Ashby, F. G., & Ennis, J. M. (2006). The role of the basal ganglia
in category learning. In B. H. Ross (Ed.), The psychology of
learning and motivation (Vol. 46, pp. 1–36). New York:
Elsevier.
Banis, S., Geerligs, L., & Lorist, M. M. (2014). Acute stress
modulates feedback processing in men and women:
Differential effects on the feedback-related negativity and
theta and beta power. PLoS One, 9, e95690.
Banis, S., & Lorist, M. M. (2012). Acute noise stress impairs
feedback processing. Biological Psychology, 91, 163–171.
Cavanagh, J. F., Figueroa, C. M., Cohen, M. X., & Frank, M. J.
(2012). Frontal theta reflects uncertainty and unexpectedness
during exploration and exploitation. Cerebral Cortex, 22,
2575–2586.
Cavanagh, J. F., & Frank, M. J. (2014). Frontal theta as a
mechanism for cognitive control. Trends in Cognitive
Sciences, 18, 414–421.
810
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cavanagh, J. F., Frank, M. J., & Allen, J. J. B. (2011). Social stress
reactivity alters reward and punishment learning. Social
Cognitive and Affective Neuroscience, 6, 311–320.
Cavanagh, J. F., Frank, M. J., Klein, T. J., & Allen, J. J. B. (2010).
Frontal theta links prediction errors to behavioral adaptation
in reinforcement learning. Neuroimage, 49, 3198–3209.
Cavanagh, J. F., Zambrano-Vazquez, L., & Allen, J. J. B. (2012).
Theta lingua franca: A common mid-frontal substrate for
action monitoring processes. Psychophysiology, 49, 220–238.
Chen, X., Zheng, T., Han, L., Chang, Y., & Luo, Y. (2017). The neural
dynamics underlying the interpersonal effects of emotional
expression on decision making. Scientific Reports, 7, 46651.
Cincotta, C. M., & Seger, C. A. (2007). Dissociation between
striatal regions while learning to categorize via feedback and via
observation. Journal of Cognitive Neuroscience, 19, 249–265.
Cohen, M. X. (2014). A neural microcircuit for cognitive conflict
detection and signaling. Trends in Neurosciences, 37,
480–490.
Cohen, M. X., Elger, C. E., & Ranganath, C. (2007). Reward
expectation modulates feedback-related negativity and EEG
spectra. Neuroimage, 35, 968–978.
Cohen, M. X., Wilmes, K., & van de Vijver, I. (2011). Cortical
electrophysiological network dynamics of feedback learning.
Trends in Cognitive Sciences, 15, 558–566.
Cook, R. G., & Smith, J. D. (2006). Stages of abstraction and
exemplar memorization in pigeon category learning.
Psychological Science, 17, 1059–1067.
Cunillera, T., Fuentemilla, L., Periañez, J., Marco-Pallarès, J.,
Krämer, U. M., Càmara, E., et al. (2012). Brain oscillatory
activity associated with task switching and feedback
processing. Cognitive, Affective & Behavioral Neuroscience,
12, 16–33.
Davis, T., Love, B. C., & Preston, A. R. (2012). Learning the
exception to the rule: Model-based fMRI reveals specialized
representations for surprising category members. Cerebral
Cortex, 22, 260–273.
de Kloet, E. R., Joëls, M., & Holsboer, F. (2005). Stress and the
brain: From adaptation to disease. Nature Reviews
Neuroscience, 6, 463–475.
Diederen, K. M. J., Spencer, T., Vestergaard, M. D., Fletcher,
P. C., & Schultz, W. (2016). Adaptive prediction error coding
in the human midbrain and striatum facilitates behavioral
adaptation and learning efficiency. Neuron, 90, 1127–1138.
Ell, S. W., Cosley, B., & McCoy, S. K. (2011). When bad stress
goes good: Increased threat reactivity predicts improved
category learning performance. Psychonomic Bulletin &
Review, 18, 96–102.
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007).
G*Power 3: A flexible statistical power analysis program for
the social, behavioral, and biomedical sciences. Behavior
Research Methods, 39, 175–191.
Frank, M. J., Seeberger, L. C., & O’Reilly, R. C. (2004). By carrot
or by stick: Cognitive reinforcement learning in
parkinsonism. Science, 306, 1940–1943.
Glienke, K., Wolf, O. T., & Bellebaum, C. (2015). The impact of
stress on feedback and error processing during behavioral
adaptation. Neuropsychologia, 71, 181–190.
Grace, A. A. (2016). Dysregulation of the dopamine system in
the pathophysiology of schizophrenia and depression.
Nature Reviews Neuroscience, 17, 524–532.
Hauser, T. U., Hunt, L. T., Iannaccone, R., Walitza, S., Brandeis,
D., Brem, S., et al. (2015). Temporally dissociable
contributions of human medial prefrontal subregions to
reward-guided learning. Journal of Neuroscience, 35,
11209–11220.
Holly, E. N., & Miczek, K. A. (2016). Ventral tegmental area
dopamine revisited: Effects of acute and repeated stress.
Psychopharmacology, 233, 163–186.
Holroyd, C. B., & Coles, M. G. H. (2002). The neural basis
of human error processing: Reinforcement learning,
dopamine, and the error-related negativity. Psychological
Review, 109, 679–709.
Holroyd, C. B., & Coles, M. G. H. (2008). Dorsal anterior
cingulate cortex integrates reinforcement history to guide
voluntary behavior. Cortex, 44, 548–559.
Hsieh, L.-T., & Ranganath, C. (2014). Frontal midline theta
oscillations during working memory maintenance and
episodic encoding and retrieval. Neuroimage, 85, 721–729.
Kalsbeek, A., van der Spek, R., Lei, J., Endert, E., Buijs, R. M., &
Fliers, E. (2012). Circadian rhythms in the hypothalamo–
pituitary–adrenal (HPA) axis. Molecular and Cellular
Endocrinology, 349, 20–29.
Kinner, V. L., Wolf, O. T., & Merz, C. J. (2016). Cortisol alters
reward processing in the human brain. Hormones and
Behavior, 84, 75–83.
Lech, R. K., Güntürkün, O., & Suchan, B. (2016). An interplay of
fusiform gyrus and hippocampus enables prototype- and
exemplar-based category learning. Behavioural Brain
Research, 311, 239–246.
Lee, T. W., Girolami, M., & Sejnowski, T. J. (1999). Independent
component analysis using an extended infomax algorithm for
mixed subgaussian and supergaussian sources. Neural
Computation, 11, 417–441.
Leicht, G., Troschütz, S., Andreou, C., Karamatskos, E., Ertl, M.,
Naber, D., et al. (2013). Relationship between oscillatory
neuronal activity during reward processing and trait
impulsivity and sensation seeking. PLoS One, 8, e83414.
Lighthall, N. R., Gorlick, M. A., Schoeke, A., Frank, M. J., &
Mather, M. (2013). Stress modulates reinforcement learning
in younger and older adults. Psychology and Aging, 28,
35–46.
Mack, M. L., Preston, A. R., & Love, B. C. (2017). Medial
prefrontal cortex compresses concept representations
through learning. bioRxiv, 178145, 1–22.
Maris, E. (2012). Statistical testing in electrophysiological
studies. Psychophysiology, 49, 549–565.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190.
Mas-Herrero, E., & Marco-Pallarés, J. (2014). Frontal theta
oscillatory activity is a common mechanism for the
computation of unexpected outcomes and learning rate.
Journal of Cognitive Neuroscience, 26, 447–458.
Mas-Herrero, E., & Marco-Pallarés, J. (2016). Theta oscillations
integrate functionally segregated sub-regions of the medial
prefrontal cortex. Neuroimage, 143, 166–174.
Mas-Herrero, E., Ripollés, P., HajiHosseini, A., Rodríguez-
Fornells, A., & Marco-Pallarés, J. (2015). Beta oscillations and
reward processing: Coupling oscillatory activity and
hemodynamic responses. Neuroimage, 119, 13–19.
McCullough, A. M., Ritchey, M., Ranganath, C., & Yonelinas, A.
(2015). Differential effects of stress-induced cortisol
responses on recollection and familiarity-based recognition
memory. Neurobiology of Learning and Memory, 123, 1–10.
Medin, D. L., & Schaffer, M. M. (1978). Context theory of
classification learning. Psychological Review, 85, 207–238.
Merz, C. J., & Wolf, O. T. (2017). Sex differences in stress effects
on emotional learning. Journal of Neuroscience Research,
95, 93–105.
Miles, S. J., & Minda, J. P. (2011). The effects of concurrent
verbal and visual tasks on category learning. Journal of
Experimental Psychology: Learning, Memory, and
Cognition, 37, 588–607.
Miller, R., Plessow, F., Kirschbaum, C., & Stalder, T. (2013).
Classification criteria for distinguishing cortisol responders
from nonresponders to psychosocial stress: Evaluation of
Paul et al.
811
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
salivary cortisol pulse detection in panel designs.
Psychosomatic Medicine, 75, 832–840.
Minda, J., & Miles, S. (2010). The influence of verbal and
nonverbal processing on category learning. In B. H. Ross
(Ed.), Psychology of learning and motivation—Advances in
research and theory (Vol. 52, pp. 117–162). Burlington, VT:
Academic Press.
Minda, J. P., & Rabi, R. (2015). Ego depletion interferes with
rule-defined category learning but not non-rule-defined
category learning. Frontiers in Psychology, 6, 1–9.
Minda, J. P., & Smith, J. D. (2001). Prototypes in category
learning: The effects of category size, category structure, and
stimulus complexity. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 27, 775–799.
Minda, J. P., & Smith, J. D. (2011). Prototype models of
categorization: Basic formulation, predictions, and
limitations. In E. Pothos & A. J. Wills (Eds.), Formal
approaches in categorization (pp. 40–64). Cambridge:
Cambridge University Press.
Mizuki, Y., Hashimoto, M., Tanaka, T., Inanaga, K., & Tanaka, M.
(1983). A new physiological tool for assessing anxiolytic
effects in humans: Frontal midline theta activity.
Psychopharmacology, 80, 311–314.
Mizuki, Y., Kajimura, N., Kai, S., Suetsugi, M., Ushijima, I., &
Yamada, M. (1992). Differential responses to mental stress in
high and low anxious normal humans assessed by frontal
midline theta activity. International Journal of
Psychophysiology, 12, 169–178.
Montoya, E. R., Bos, P. A., Terburg, D., Rosenberger, L. A., &
van Honk, J. (2014). Cortisol administration induces global
down-regulation of the brain’s reward circuitry.
Psychoneuroendocrinology, 47, 31–42.
Nasser, H. M., Calu, D. J., Schoenbaum, G., & Sharpe, M. J.
(2017). The dopamine prediction error: Contributions to
associative models of reward learning. Frontiers in
Psychology, 8, 1–17.
Nosofsky, R. M. (1986). Attention, similarity, and the
identification–categorization relationship. Journal of
Experimental Psychology: General, 115, 39–57.
Nosofsky, R. M. (2011). The generalized context model: An
exemplar model of classification. In E. Pothos & A. J. Wills
(Eds.), Formal approaches in categorization (pp. 18–39).
Cambridge, UK: Cambridge University Press.
Oehrn, C. R., Hanslmayr, S., Fell, J., Deuker, L., Kremers, N. A.,
Do Lam, A. T., et al. (2014). Neural communication patterns
underlying conflict detection, resolution, and adaptation.
Journal of Neuroscience, 34, 10438–10452.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis of MEG,
EEG, and invasive electrophysiological data. Computational
Intelligence and Neuroscience, 2011, 156869.
Ossewaarde, L., Qin, S., Van Marle, H. J. F., van Wingen, G. A.,
Fernández, G., & Hermans, E. J. (2011). Stress-induced
reduction in reward-related prefrontal cortex function.
Neuroimage, 55, 345–352.
Pabst, S., Brand, M., & Wolf, O. T. (2013). Stress and decision
making: A few minutes make all the difference. Behavioural
Brain Research, 250, 39–45.
Pan, X., & Sakagami, M. (2012). Category representation and
generalization in the prefrontal cortex. European Journal of
Neuroscience, 35, 1083–1091.
Park, H., Lee, D., & Chey, J. (2017). Stress enhances model-free
reinforcement learning only after negative outcome. PLoS
One, 12, e0180588.
Pastötter, B., Dreisbach, G., & Bäuml, K.-H. T. (2013). Dynamic
adjustments of cognitive control: Oscillatory correlates
of the conflict adaptation effect. Journal of Cognitive
Neuroscience, 25, 2167–2178.
Pedersen, M. L., Frank, M. J., & Biele, G. (2017). The drift
diffusion model as the choice rule in reinforcement learning.
Psychonomic Bulletin & Review, 24, 1234–1251.
Petzold, A., Plessow, F., Goschke, T., & Kirschbaum, C. (2010).
Stress reduces use of negative feedback in a feedback-based
learning task. Behavioral Neuroscience, 124, 248–255.
Pfurtscheller, G., & Aranibar, A. (1977). Event-related cortical
desynchronization detected by power measurements of
scalp EEG. Electroencephalography and Clinical
Neurophysiology, 42, 817–826.
Pivik, R. T., Broughton, R. J., Coppola, R., Davidson, R. J., Fox,
N., & Nuwer, M. R. (1993). Guidelines for the recording and
quantitative analysis of electroencephalographic activity in
research contexts. Psychophysiology, 30, 547–558.
Porcelli, A. J., & Delgado, M. R. (2017). Stress and decision
making: Effects on valuation, learning, and risk-taking.
Current Opinion in Behavioral Sciences, 14, 33–39.
Posner, M. I., & Keele, S. W. (1968). On the genesis of abstract
ideas. Journal of Experimental Psychology, 77, 353–363.
Rabi, R., & Minda, J. P. (2016). Category learning in older
adulthood: A study of the Shepard, Hovland, and Jenkins
(1961) tasks. Psychology and Aging, 31, 185–197.
Reed, S. K. (1972). Pattern recognition and categorization.
Cognitive Psychology, 3, 382–407.
Reinhart, R. M. G. (2017). Disruption and rescue of interareal
theta phase coupling and adaptive behavior. Proceedings
of the National Academy of Sciences, U.S.A., 114,
11542–11547.
Reschke-Hernández, A. E., Okerstrom, K. L., Bowles Edwards,
A., & Tranel, D. (2017). Sex and stress: Men and women show
different cortisol responses to psychological stress induced
by the Trier social stress test and the Iowa singing social
stress test. Journal of Neuroscience Research, 95, 106–114.
Rolls, E. T., McCabe, C., & Redoute, J. (2008). Expected value,
reward outcome, and temporal difference error
representations in a probabilistic decision task. Cerebral
Cortex, 18, 652–663.
Schenk, S., Minda, J. P., Lech, R. K., & Suchan, B. (2016). Out of
sight, out of mind: Categorization learning and normal aging.
Neuropsychologia, 91, 222–233.
Schultz, W., & Dickinson, A. (2000). Neuronal coding of
prediction errors. Annual Review of Neuroscience, 23,
473–500.
Schwabe, L., Haddad, L., & Schachinger, H. (2008). HPA axis
activation by a socially evaluated cold-pressor test.
Psychoneuroendocrinology, 33, 890–895.
Schwabe, L., Tegenthoff, M., Höffken, O., & Wolf, O. T. (2013).
Mineralocorticoid receptor blockade prevents stress-induced
modulation of multiple memory systems in the human brain.
Biological Psychiatry, 74, 801–808.
Schwabe, L., & Wolf, O. T. (2012). Stress modulates the
engagement of multiple memory systems in classification
learning. Journal of Neuroscience, 32, 11042–11049.
Schwabe, L., & Wolf, O. T. (2013). Stress and multiple memory
systems: From “thinking” to “doing”. Trends in Cognitive
Sciences, 17, 60–68.
Seger, C. A., & Cincotta, C. M. (2005). The roles of the caudate
nucleus in human classification learning. Journal of
Neuroscience, 25, 2941–2951.
Seger, C. A., Peterson, E. J., Cincotta, C. M., Lopez-Paniagua, D.,
& Anderson, C. W. (2010). Dissociating the contributions of
independent corticostriatal systems to visual categorization
learning through the use of reinforcement learning modeling
and Granger causality modeling. Neuroimage, 50, 644–656.
Shepard, R. (1987). Toward a universal law of generalization for
psychological science. Science, 237, 1317–1323.
Shields, G. S., Sazma, M. A., McCullough, A. M., & Yonelinas,
A. P. (2017). The effects of acute stress on episodic memory:
812
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
A meta-analysis and integrative review. Psychological
Bulletin, 143, 636–675.
Smith, D. J., & Minda, J. P. (2000). Thirty categorization results
in search of a model. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 26, 3–27.
Smith, E. H., Banks, G. P., Mikell, C. B., Cash, S. S., Patel, S. R.,
Eskandar, E. N., et al. (2015). Frequency-dependent
representation of reinforcement-related information in the
human medial and lateral prefrontal cortex. Journal of
Neuroscience, 35, 15827–15836.
Smith, J. D., & Minda, J. P. (1998). Prototypes in the mist: The
early epochs of category learning. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 24, 1411–1436.
Sutton, R. S., & Barto, A. G. (1981). Toward a modern theory of
adaptive networks: Expectation and prediction.
Psychological Review, 88, 135–170.
van de Vijver, I., Ridderinkhof, K. R., & Cohen, M. X. (2011).
Frontal oscillatory dynamics predict feedback learning and
action adjustment. Journal of Cognitive Neuroscience, 23,
4106–4121.
Vogel, S., Fernández, G., Joëls, M., & Schwabe, L. (2016).
Cognitive adaptation under stress: A case for the
mineralocorticoid receptor. Trends in Cognitive Sciences, 20,
192–203.
Vogel, S., Klumpers, F., Schröder, T. N., Oplaat, K. T., Krugers,
H. J., Oitzl, M. S., et al. (2017). Stress induces a shift towards
striatum-dependent stimulus–response learning via the
mineralocorticoid receptor. Neuropsychopharmacology, 42,
1262–1271.
Wirz, L., Wacker, J., Felten, A., Reuter, M., & Schwabe, L. (2017).
A deletion variant of the α2b-adrenoceptor modulates the
stress-induced shift from “cognitive” to “habit” memory.
Journal of Neuroscience, 37, 2149–2160.
Wolf, O. T. (2017). Stress and memory retrieval: Mechanisms
and consequences. Current Opinion in Behavioral Sciences,
14, 40–46.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
7
9
9
1
7
8
7
4
3
7
/
j
o
c
n
_
a
_
0
1
2
4
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Paul et al.
813