Social and Nonsocial Relational Reasoning
in Adolescence and Adulthood
Lucía Magis-Weinberg1, Sarah-Jayne Blakemore1, and Iroise Dumontheil2
Abstrakt
■ Reasoning during social interactions requires the individual
manipulation of mental representations of one’s own traits and
those of other people as well as their joint consideration (rela-
tional integration). Research using nonsocial paradigms has
linked relational integration to activity in the rostrolateral PFC.
Hier, we investigated whether social reasoning is supported by
the same general system or whether it additionally relies on
regions of the social brain network, such as the medial PFC. Wir
further assessed the development of social reasoning. Im
social task, participants evaluated themselves or a friend, or com-
pared themselves with their friend, on a series of traits. In the non-
social task, participants evaluated their hometown or another
town or compared the two. In a behavioral study involving
325 Teilnehmer (11–39 years old), we found that integrating rela-
tionen, compared with performing single relational judgments,
improves during adolescence, both for social and nonsocial infor-
mation. Thirty-nine female participants (10–31 years old) took part
in a neuroimaging study using a similar task. Activation of the
relational integration network, including the rostrolateral PFC,
was observed in the comparison condition of both the social
and nonsocial tasks, whereas the medial PFC showed greater acti-
vation when participants processed social as opposed to non-
social information across conditions. Developmentally, the right
anterior insula showed greater activity in adolescents compared
with adults during the comparison of nonsocial versus social infor-
mation. This study shows parallel recruitment of the social brain
and the relational reasoning network during the relational inte-
gration of social information in adolescence and adulthood. ■
EINFÜHRUNG
Is London more expensive than Cambridge? Answering
this question entails at least two levels of relational rea-
soning. At the first level, one needs to judge the prices
in each city independently (evaluation of single relations,
z.B., how much do houses in London cost?). At the sec-
ond level, one needs to simultaneously consider mental
representations of both cities and to integrate the single
judgments into a higher-order comparison (relational
Integration, in diesem Fall, comparing the house prices in
London and Cambridge). Relational integration has typi-
cally been studied in nonsocial contexts, insbesondere,
using the Raven’s Progressive Matrices (Raven, 1998).
Jedoch, relational integration also occurs in the social
Domain, Zum Beispiel, when comparing people on per-
sonality traits (z.B., are you more patient than your
friend?). The neural processes supporting this kind of
social reasoning and the way it develops are not well
verstanden.
Previous fMRI research has identified the lateral PFC
and lateral parietal cortex as involved in relational integra-
tion (Dumontheil, 2014; Bunge, Helskog, & Wendelken,
2009; Wendelken, Nakhabenko, Donohue, Fuhrmann, &
Bunge, 2008; Schmied, Keramatian, & Christoff, 2007),
whereas medial prefrontal cortex (MPFC) has been asso-
1University College London, 2University of London
ciated with the processing and manipulation of social
Information ( Van Overwalle, 2009; Gilbert et al., 2006;
Holz & Grafman, 2003). The current study aimed to
bring together these separate strands of research to in-
vestigate domain-general and social domain-specific pro-
cesses that support the relational integration of social
Information. Both relational reasoning and social cogni-
tion and their underlying neural substrates undergo sig-
nificant reorganization during adolescence (Kilford,
Garrett, & Blakemore, 2016; Dumontheil, 2014). Dort-
Vordergrund, a second aim of the current study was to compare
social reasoning in adolescents and adults. We employed
a paradigm that allows the investigation and comparison
of relational integration of both social and nonsocial infor-
mation (Raposo, Vicens, Clithero, Dobbins, & Huettel,
2011). In a large behavioral study, we investigated the
development of relational integration of social and non-
social information from late childhood until adulthood.
In a follow-up fMRI study, we studied the neural cor-
relates of these cognitive processes in adolescence and
Erwachsensein.
Neural Bases of Relational Integration and
Social Cognition
Relational reasoning research suggests a central role of
rostrolateral prefrontal cortex (RLPFC), which corresponds
© 2017 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 Unportiert (CC BY 3.0) Lizenz.
Zeitschrift für kognitive Neurowissenschaften 29:10, S. 1739–1754
doi:10.1162/jocn_a_01153
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
to the lateral aspect of the anterior, or rostral, prefrontal
Kortex (Brodmann’s areas [BAs] 10/46 Und 10/47), In
relational integration compared with processing single
Beziehungen. Imaging studies using the Raven’s Progressive
Matrices in adults have shown RLPFC involvement in
the joint manipulation of visuospatial patterns (Kroger
et al., 2002; Christoff et al., 2001) as well as in the in-
tegration of relations in analogical reasoning tasks
( Wendelken et al., 2008; Bunge, Wendelken, Badre, &
Wagner, 2005) and in the integration of multiple relations
to reach a logical conclusion (Wendelken & Bunge, 2009).
A study comparing visuospatial and semantic variants of a
relational matching task found considerable activation
overlap within the left RLPFC, suggesting a domain-general
role for RLPFC in relational integration ( Wendelken,
Chung, & Bunge, 2012).
Social cognitive research suggests a role of the MPFC,
which corresponds to BA 8/BA 9/BA 10, in the processing
of social information (see Van Overwalle, 2009, for a meta-
Analyse). Studies with adults have shown that this region
is involved in considering one’s thoughts and feelings
(Rameson, Satpute, & Lieberman, 2010; Gusnard, 2005;
Zysset, Huber, Samson, Ferstl, & von Cramon, 2003) Und
in perspective taking (PT; David et al., 2008; D’Argembeau
et al., 2007; Aichhorn, Perner, Kronbichler, Staffen, &
Ladurner, 2006; David et al., 2006; Ruby & Decety, 2001,
2004; Vogeley et al., 2004). The MPFC is also recruited dur-
ing tasks that require mentalizing, das ist, the consideration
of other people’s mental states (Amodio & Frith, 2006;
Decety & Sommerville, 2003; Frith & Frith, 2003).
Development during Adolescence
Both relational integration and social cognition show pro-
tracted development in terms of improved performance
and associated brain activity between adolescence and
Erwachsensein (Blakemore, 2012; Crone & Dahl, 2012;
Dumontheil & Blakemore, 2012; Dumontheil, Hillebrandt,
Apperly, & Blakemore, 2012; Crone et al., 2009;
Dumontheil, Bürger, & Blakemore, 2008). The RLPFC un-
dergoes structural and functional development with age,
with evidence that its activity during relational integra-
tion tasks becomes increasingly specialized during child-
hood and adolescence (Dumontheil, 2014; Wendelken,
O’Hare, Whitaker, Ferrer, & Bunge, 2011; Dumontheil,
Houlton, Christoff, & Blakemore, 2010; Crone et al., 2009;
Ferrer, O’Hare, & Bunge, 2009; Dumontheil et al., 2008).
Zusätzlich, a complex pattern of developmental changes
in functional connectivity related to reasoning ability has
been identified, including changes in connectivity between
the RLPFC and the parietal cortex ( Wendelken, Ferrer,
Whitaker, & Bunge, 2016; Bazargani, Hillebrandt, Christoff,
& Dumontheil, 2014). Bazargani et al. (2014) observed a
decrease in short-range (fronto-insular) connectivity with
stable long-range connectivity (frontoparietal) und ein
increase of modulatory connections with age. Wendelken
et al. (2016) found a pattern of developmental changes
suggestive of increasing communication between pre-
frontal regions and specific targets.
Key regions of the social brain, including the MPFC,
undergo structural and functional changes during adoles-
cence. Cortical thickness and gray matter volume in the
MPFC decrease between late childhood and the early 20s
(Mills, Lalonde, Clasen, Giedd, & Blakemore, 2014). In
parallel, several fMRI studies have shown that MPFC activ-
ity during mentalizing tasks decreases between early
adolescence and adulthood (Blakemore, 2008, 2012). In
a previous study investigating the development of the
neural correlates of mentalizing, Teilnehmer waren
required either to take someone else’s perspective or
to use symbolic cues to select an appropriate action in
a communicative context. We found that adolescents
showed hypoactivation of domain-general cognitive con-
trol regions in the parietal cortex and PFC and hyperacti-
vation of parts of the social brain network (Dumontheil
et al., 2012). This study thus demonstrated the engage-
ment of cognitive control and social brain regions within
a single paradigm and that the engagement of these
regions changes as a function of age.
Relational integration within the social domain has been
investigated in adults using a task that combined both
mentalizing and relational integration (Raposo et al.,
2011). Participants judged how pleasant they found a cer-
tain word, how pleasant a friend would find the word, Und
how their rating of pleasantness would compare with that
of their friend. Behaviorally, RTs were higher when partic-
ipants were comparing themselves with their friend relative
to the two single-relation conditions. MPFC activation was
higher during the friend judgment compared with the
self-judgment, whereas RLPFC activation was higher
when contrasting the relational integration comparison
and self-judgment conditions. The study did not include
a nonsocial relational reasoning condition, preventing
the conclusion that the activation patterns are specific to
relational integration of social information per se or reflec-
tive of relational integration more generally.
This Study
Hier, we adapted the paradigm designed by Raposo et al.
(2011) to investigate behavioral development of social
reasoning (Study 1) and its neural development between
adolescence and adulthood (Study 2). We compared first-
order judgments (1-REL) of traits associated with oneself
or with another individual (z.B., How patient are you?
[Self condition]; How patient is your friend? [Other con-
dition]) with second-order judgments (2-REL) about how
these judgments related to each other (How much more
patient are you than your friend? [Comparison condi-
tion]). Our paradigm also included a control nonsocial
Aufgabe, in which participants were asked to rate character-
istics of towns. Our aim was to assess (1) how perfor-
mance on a task requiring relational integration of social
or nonsocial traits develops between late childhood and
1740
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Erwachsensein, (2) how neural activity underlying these pro-
cesses develops between early adolescence and adult-
hood, Und (3) whether there is domain-specific activation
for the relational integration of social versus nonsocial
Information.
In terms of behavior, we predicted improvements in
relational integration with age, both in terms of RT and
the consistency of participants’ responses between
1-REL and 2-REL judgments. In terms of BOLD signal,
we expected domain-general activations associated with
relational integration in the RLPFC, dorsolateral PFC,
and parietal cortex. We also predicted that there would
be additional domain-specific activations in parts of the
social brain network associated with the people task, spe-
cifically regions involved in processing social information
and mentalizing (Meyer, Taylor, & Lieberman, 2015;
Dumontheil & Blakemore, 2012; Meyer, Spunt, Berkman,
Taylor, & Lieberman, 2012; Raposo et al., 2011). Endlich,
we predicted that the RLPFC would show increased spec-
ificity of activation for 2-REL versus 1-REL judgments in
adults compared with adolescents (Dumontheil, 2014)
and that the MPFC would show greater activation in ad-
olescents than adults in the social versus nonsocial task
(Blakemore & Robbins, 2012; Blakemore, 2008).
STUDY 1: BEHAVIORAL STUDY
Teilnehmer
The data analyzed here are part of a larger project in male
and female children, adolescents, and adults who per-
formed a set of six tasks and provided saliva samples
for genetic analyses (Kilford, Dumontheil, Holz, &
Blakemore, 2015; Dumontheil et al., 2014). The present
analysis focused on the social and nonsocial comparison
task and the Wechsler Abbreviated Scale of Intelligence
( WASI; Wechsler, 1999) assessment. The data presented
here are from 325 participants aged between 11 Und
39 Jahre (n = 160 Erwachsene, n = 165 children and adoles-
cents). From an original sample of 399 Teilnehmer, eins
was excluded because of a diagnosis of Turner syndrome,
one was excluded because of a diagnosis of Asperger
syndrome, four were excluded because of a task program-
ming error, two were excluded because they interrupted
the task early, one participant did not have time to com-
plete this task, and data were lost from 15 Teilnehmer. In
addition, as only four male participants were younger than
11 Jahre (vs. 26 female participants), all participants youn-
ger than 11 years were excluded from further analyses.
Children and adolescents were recruited from schools in
and around London and were tested in their school, Und
adults were recruited from the University College London
(UCL) Psychology Department volunteer database and
word of mouth and tested in the laboratory. Written in-
formed consent was obtained from participants or from
the parent/guardian of participants under 18 Jahre alt.
Adult participants were remunerated for their time. Der
study was approved by the UCL ethics committee.
Child and adolescent participants were divided accord-
ing to age into three groups spanning 2 oder 3 Jahre, Und
there were four adult groups (Tisch 1). Verbal ability
Tisch 1. Demographics of Participants in the Behavioral (Study 1) and fMRI (Study 2) Studien
Age Group
N
Behavioral study (Study 1)
11–12 years
66
13–14 years
15–17 years
20–22 years
23–25 years
26–28 years
29–39 years
fMRT-Studie (Study 2)
10–16 years
22–31 years
57
42
43
50
39
28
19
20
Sex
F/M
39/27
30/27
20/27
20/22
28/22
20/19
10/18
Alter
Verbal IQ
Range
Mean (SD)
Range
Mean (SD)
11.10–12.98
13.06–14.97
15.01–17.99
20.34–22.97
23.01–25.96
26.06–28.68
29.00–39.39
12.05 (0.51)
13.92 (0.58)
16.18 (0.92)
21.93 (0.69)
24.55 (0.92)
27.30 (0.81)
33.22 (2.80)
87–137
84–140
84–134
97–137
81–137
84–129
94–129
114.4 (12.0)
113.1 (12.7)
114.0 (12.4)
118.1 (9.5)
113.7 (13.7)A
107.6 (12.2)B
115.5 (9.5)
Female only
Female only
10.98–16.83
22.22–31.67
14.10 (1.89)
25.89 (2.76)
93–134
107–131
116.8 (11.4)
119.9 (6.7)C
aWASI data were missing for one participant.
bWASI data were missing for three participants.
cWASI data were missing for one participant.
Magis-Weinberg, Blakemore, and Dumontheil
1741
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
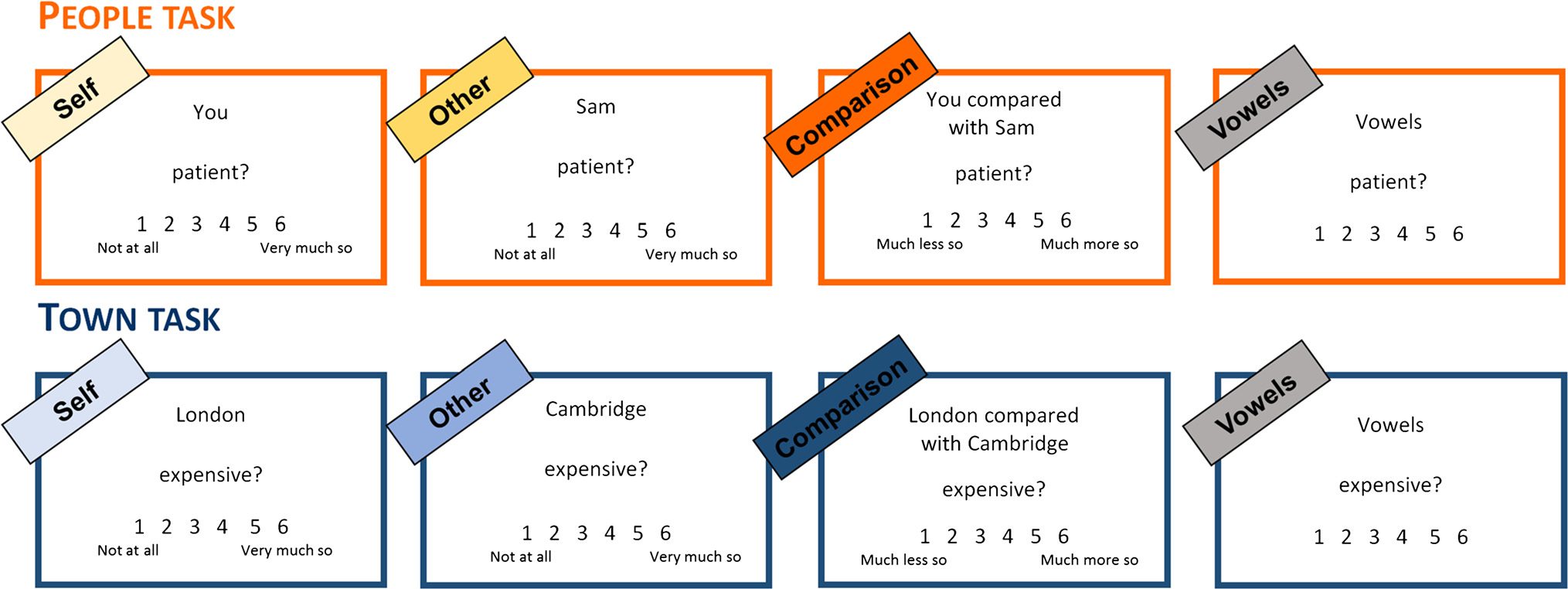
Figur 1. Example of stimuli for each task and condition. The Vowels condition was only included in Study 2. In the People task, Teilnehmer waren
asked: “How much do you think the following words apply to you/your friend?” (Self/Other) or “How much do you think the following words
apply to you compared with your friend?” (Comparison). On the rating scale, 1 indicated “not at all” and 6 indicated “very much so” in the
Self and Other conditions, wohingegen 1 indicated “much less so” and 6 indicated “much more so” in the Comparison condition. All text was presented
in white on a black background.
was measured using the vocabulary subtest of the WASI
( Wechsler, 1999). A two-way (Age group, Sex) ANOVA
indicated that there was a significant difference in verbal
IQ between age groups (F(1, 307) = 2.91, p = .009).
Paired post hoc comparisons demonstrated that the
age groups of 11–12 and 13–14 years had lower mean
verbal IQ than the age group of 20–22 years ( ps < .05)
and that the age group of 26–28 years had lower mean
verbal IQ than all other age groups ( ps < .05; Table 1).
Only the difference between the age groups of 20–22 and
26–28 years survived Bonferroni correction ( p = .001).
There was no main effect of Sex on IQ (F(1, 307) =
2.34, p = .127), but there was a significant Sex × Age
group interaction (F(6, 307) = 3.08, p = .006). Post hoc
comparisons of male and female participants in each age
group indicated that 11- to 12-year-old male participants
had lower mean verbal IQ (109.1, SD = 12.2) than female
participants (118.1, SD = 10.5, p = .002), whereas 23- to
25-year-old men had higher mean verbal IQ (118.6, SD =
11.1) than women (109.7, SD = 14.5, p = .023).
Design and Stimulus Material
The task had two within-participant factors (Task: people
or town; Condition: self, other, or comparison) and one
between-participant factor (Age group: seven levels), re-
sulting in a 2 × 3 × 7 mixed factorial design. The task
was computer based and adapted from the fMRI study by
Raposo et al. (2011). The task was administered as part of
a single individual testing session of approximately 45–
50 min. It was the third task administered in the task set,
and the WASI was administered as the sixth (and last) task.
The experimenter started by asking participants to
think of someone whom they knew quite well but who
was quite different from them and to give his or her
name. If participants did not respond, the experimenter
suggested that they consider a close friend or a sibling
who was quite different from them. Second, participants
were asked to name the town where they lived (typically,
London) and then pick a town that they knew quite well
but that was quite different from London. Again, if partic-
ipants did not respond, the experimenter suggested that
they consider a town where they go on a holiday or
where their grandparents live. Instructions were then
presented on the screen and read aloud to the partici-
pants, explaining the different types of judgment they
would make during the task and the rating scale. Partici-
pants used the index, middle, and ring fingers of both
hands to respond. The task was programmed in Cogent
(www.vislab.ucl.ac.uk/cogent_graphics.php) running in
MATLAB (The MathWorks, Inc., Natick, MA) on a Dell
12-in. laptop or similar.
Judgments were blocked according to Task and Condi-
tion, and the order of the blocks was counterbalanced
within and between participants. Each block started with
an instruction screen indicating to participants what type
of judgment they should make during that block. On
each trial, this information was repeated at the top
(e.g., “You”), an adjective was presented in the middle
of the screen, and a rating scale from 1 to 6 was provided
at the bottom of the scale (Figure 1).
Once participants had pressed a key to indicate their
response, the corresponding number on the scale (1–6)
was highlighted in red for 200 msec, followed by a
200-msec blank screen, and then a new trial started. There
were 10 trials per block and 12 blocks in total, with two
blocks of each of the six conditions (People or Town ×
Self, Other, or Comparison), that is, 20 trials in total per
1742
Journal of Cognitive Neuroscience
Volume 29, Number 10
condition. A list of 20 adjectives was used for this study
(fabulous, weird, loud, charming, romantic, crazy, pleasant,
lovely, wild, perfect, busy, unique, friendly, cool, unusual,
boring, dull, rich, quiet, and popular). All adjectives were
presented once in each condition of each task.
Data Analysis
Data were analyzed with SPSS 21 (IBM Corp., Armonk,
NY ). Median RTs were calculated for each Task and
Condition. A 2 (Task) × 3 (Condition) × 7 (Age group) ×
2 (Sex) mixed-model repeated-measures ANOVA (rmANOVA)
was performed on the median RT data. Because of the
nature of the task, a measure for accuracy is not avail-
able (we do not know objectively how “friendly” a par-
ticipant or their friend is). Instead, we ran Spearman
correlations to test whether participants were con-
sistent in their response, that is, whether there was a
positive correlation between their ratings in the Com-
parison judgment and the difference in ratings between
the Self and Other judgments, across the set of 20
adjectives. For example, if a participant rated herself
as “6” and her friend as “3” for “friendly,” a rating of
“5” in the Comparison judgment (“How much more
friendly do you think you are compared with your
friend?”) would be consistent with the individual judg-
ments. Spearman r correlation values obtained for each
participant in each task were transformed using Fisher’s z
transformation (z = 0.5 × ln ((1 + r)/(1 − r)), resulting in
consistency scores. A 2 (Task) × 7 (Age group) × 2 (Sex)
mixed-model rmANOVA was performed on the consistency
data. Post hoc comparisons investigating pairwise differ-
ences between age groups were Bonferroni corrected.
Greenhouse–Geisser correction for nonsphericity was
applied when necessary. Estimated means and standard
errors from the rmANOVAs are reported in the text and
in the figures.
Results
RTs
A 2 (Task) × 3 (Condition) × 7 (Age group) × 2 (Sex)
mixed rmANOVA showed a main effect of Condition (F(1.4,
435.6) = 500.00, p < .001, ηp
2 = 0.62), whereby partici-
pants responded faster in the Self (M = 1944 msec, SD =
36 msec) than the Other (M = 1989 msec, SD = 35 msec)
condition ( p = .042) and in both Self and Other conditions
than in the Comparison condition (M = 2811 msec, SD =
56 msec; ps < .001); a main effect of Task (F(1, 311) =
12.04, p < .001, ηp
2 = 0.04), with slower RTs in the People
(M = 2292 msec, SD = 44 msec) than the Town (M =
2203 msec, SD = 39 msec) task; and a main effect of
Age group (F(6, 311) = 2.48, p = .024, ηp
2 = 0.05) but
no main effect of Sex (F(1, 311) = 0.05, p = .818). Pairwise
post hoc comparisons indicated that the 11- to 12-year
age group responded more slowly than the 20- to 22-year
age group ( p = .019) and marginally slower than the 26- to
28-year age group ( p = .081). No other pairwise com-
parison was significant.
The only significant interaction was between Condition
and Age group (F(8.4, 435.6) = 3.64, p < .001, ηp
2 =
0.07). This was followed up by first entering the two
1-REL judgments in a 2 (Task) × 7 (Age group) × 2
(Sex) rmANOVA. This analysis showed no main effect of
Age group (F(6, 311) = 1.54, p = .165) and no Age group ×
Condition interaction (F(6, 311) = 1.35, p = .233), in-
dicating that the interaction was driven by the 2-REL,
Comparison condition. Investigated separately, this con-
dition showed a main effect of Age group (F(6, 311) =
3.61, p = .002, ηp
2 = 0.07). Post hoc comparisons indi-
cated that the 11- to 12-year age group responded slower
on average than the 20- to 22- and 23- to 25-year age
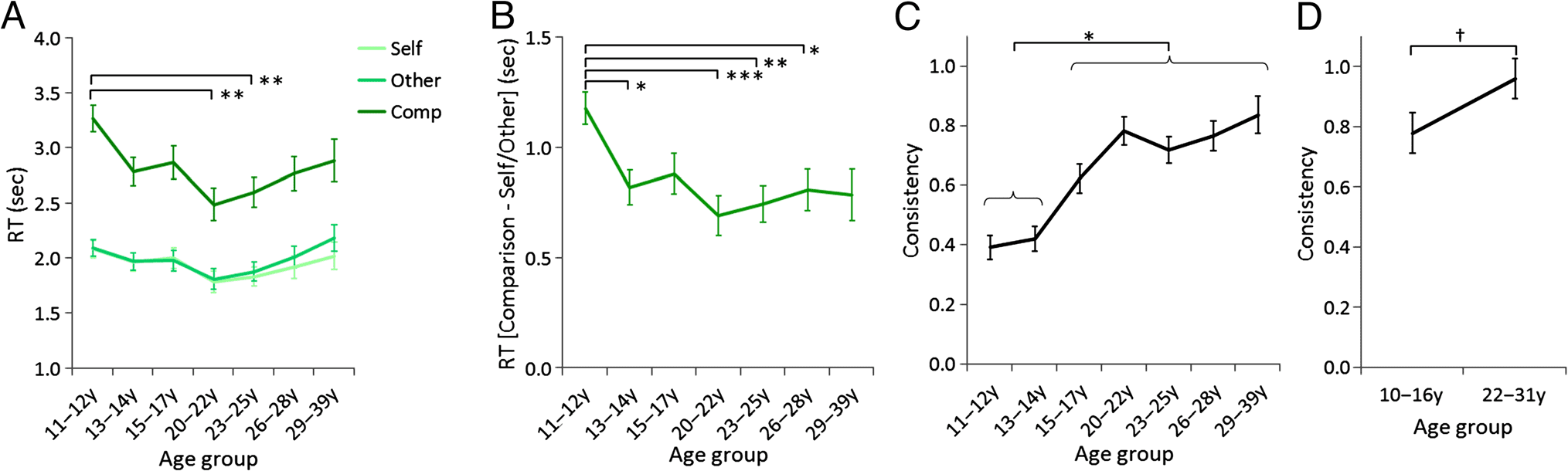
groups ( ps < .01; Figure 2A). To further investigate the inter-
action, the difference in median RT between the Comparison
condition (2-REL, relational integration) and Self and
Other conditions (1-REL judgments) was calculated. A 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
o
6
c
7
n
1
_
1
a
/
_
j
0
o
1
c
1
n
5
3
_
a
p
_
d
0
1
b
1
y
5
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Mean RT and consistency scores as a function of age group. (A) Study 1: mean RTs as a function of Age group and Condition. (B) Study 1:
mean difference in RTs between the Comparison and Self and Other conditions as a function of Age group. (C) Study 1: mean consistency
scores as a function of Age group. (D) Study 2: mean consistency scores as a function of Age group. Error bars represent SE. †
**p < .01, ***p < .001 (Bonferroni corrected). y = years.
p < .1, *p < .05,
Magis-Weinberg, Blakemore, and Dumontheil
1743
(Task) × 7 (Age group) × 2 (Sex) rmANOVA with post hoc
pairwise comparisons indicated that the 11- to 12-year
age group was relatively slower in 2-REL than 1-REL trials
than the 13- to 14-, 20- to 22-, 23- to 25-, and 26- to 28-year
age groups ( ps < .05; Figure 2B).
Consistency
A 2 (Task) × 7 (Age group) × 2 (Sex) mixed rmANOVA
was performed on consistency, which is the Fisher z-
transformed Spearman correlation between participants’
answers in the Comparison condition and the difference
between their answers in the Self and Other conditions.
The main effect of Task was significant (F(1, 311) =
21.74, p < .001, ηp
2 = 0.07), with greater consistency in
the Town (M = 0.713, SD = 0.025) than in the People
(M = 0.586, SD = 0.021) task. In both tasks, mean con-
sistency was positive, indicating some degree of consis-
tency in participants’ responses across conditions. The
main effect of Age group was also significant (F(6, 311) =
15.15, p < .001, ηp
2 = 0.23). No other main effect and no
interaction were significant. Post hoc comparisons indi-
cated that the 11- to 12- and 13- to 14-year age groups
did not differ from each other and were less consistent
than all older age groups ( ps < .05), which did not differ
from each other (Figure 2C).
In summary, participants were slightly slower in the
Other than in the Self condition and in the People than
in the Town task. However, there was a considerable dif-
ference in RT between 2-REL and 1-REL judgments,
which decreased between 11–12 and 13–14 years old.
Consistency was also higher in the Town than in the
People task and improved between 13–14 and 15–
17 years old. There was no interaction between Task and
Age group, suggesting no evidence of a developmental
difference in relational integration between the social and
nonsocial tasks.
STUDY 2: NEUROIMAGING STUDY
Methods
Participants
Thirty-nine female participants aged between 10 and
31 years (n = 20 adults, n = 19 children and adolescents)
took part in the neuroimaging study (Table 1). Seven ad-
olescents and two adults had taken part in Study 1, with
an interval of between 4 and 10 months between the test-
ing sessions. Only female participants were included to
reduce variability in the sample due to sex differences
in brain development (Herting, Maxwell, Irvine, & Nagel,
2012; Raznahan et al., 2011). Sex differences were not
found in Study 1 or in a previous behavioral and neuro-
imaging study of visuospatial relational reasoning devel-
opment ( Wendelken et al., 2011). However, other
behavioral studies have reported sex differences in men-
talizing (e.g., Charman, Ruffman, & Clements, 2002) and
in relational reasoning (Lynn & Irwing, 2004). Because we
were unable to collect a sample large enough to investi-
gate sex differences, we chose to maximize the homoge-
neity of our sample by only including female participants.
Participants were reimbursed £20 and their travel ex-
penses for taking part in the study. The study was
approved by the UCL research ethics committee. Partici-
pants were divided into two groups, adolescents and
adults. Adolescents were combined into a single group
because of the sample size, with a focus on investigating
the development of the neural correlates of relational
reasoning, whereas Study 1 had focused on developmen-
tal changes in performance. The two groups were
matched on estimated IQ (t(36) = 1.02, p = .314), which
was assessed using the vocabulary and matrices subtests
of the WASI ( Wechsler, 1999).
Design and Stimulus Material
The fMRI task had two within-participant factors (Task:
People or Town; Condition: Self, Other, Comparison,
or Vowels) and one between-participant factor (Age
group: adults, adolescents), resulting in a 2 × 4 × 2
mixed design. Participants were first trained on the task
outside the scanner. After going through the four types
of question for each Task, participants performed one
block of three trials for each Task and Condition. Partic-
ipants then performed four scanning runs as well as a
structural scan between the second and third task runs.
After scanning, participants completed the Interpersonal
Reactivity Index (IRI; Davis, 1980), which provides mea-
sures of four components of empathy (empathic concern
[EC], fantasy, personal distress, and PT), and were as-
sessed on the WASI. The IRI was included as it has been
used in previous neuroimaging studies as a measure of
individual differences in social cognition in everyday life
(Meyer et al., 2012; Raposo et al., 2011). Meyer et al.
(2012) found an association between memory load-
dependent activity within mentalizing regions and scores
on the PT scale. We therefore aimed to relate activity
within mentalizing regions in Study 2 with this everyday
life measure of PT.
The paradigm was similar to the task described in
Study 1, adapted from Raposo et al. (2011), with three
differences. First, the fMRI task included a Vowels condi-
tion in which participants were required to count the
number of vowels in the adjective presented on the
screen (“Control” condition used by Raposo et al.,
2011). This condition matched visual and verbal process-
ing and motor execution demands while minimizing rela-
tional reasoning demands and, as such, was used as a
baseline in the fMRI analyses. Regions of the social brain
often show high activation at rest or during fixation
phases, as part of the default mode network. Using an
active baseline allowed the identification of activation of
these brain regions, which was important for the People
task. In addition, the words presented in the People and
1744
Journal of Cognitive Neuroscience
Volume 29, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
o
6
c
7
n
1
_
1
a
/
_
j
0
o
1
c
1
n
5
3
_
a
p
_
d
0
1
b
1
y
5
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Town tasks differed, and using the Vowels condition as a
baseline allowed us to control for BOLD signal differ-
ences elicited by the presentation of these words. Finally,
using a similar baseline to Raposo et al. (2011) facilitates
comparison of the results of the two studies. Second,
traits were different for the People and Town tasks in an
attempt to make them more relevant to each category.
Third, a fixed ISI of 100 msec was used.
Participants performed two scanning runs of the Peo-
ple and Town task, alternating in an ABAB or BABA order,
counterbalanced across participants. Each run was com-
posed of five blocks of each condition (Self, Other, Com-
parison, and Vowels). The order of the conditions was
fixed within a run and counterbalanced across runs and
participants. After every Vowels block, there was a
fixation block. Task blocks were preceded by a 1-sec
instruction that specified the condition of the next block
(e.g., “You,” “Sam,” “You compared with Sam,” “Vowels,”
“London,” “Cambridge,” “London compared with
Cambridge”) and was composed of three trials each.
Participants had a maximum of 6.05 sec to input their
response on each trial, during which time the stimulus
remained on the screen. When participants responded,
the number they pressed turned red, and the stimulus
remained on the screen until 6.1 sec after the onset of
the trial presentation. A blank screen was displayed
during the ISI. Stimuli consisted of lists of 30 adjectives
in each task, which were matched for number of letters,
number of vowels, frequency, and familiarity (see Appen-
dix). Each adjective was presented once in each of the
four conditions. Half of the adjectives were presented
in the first scanning run of a task, half in the second run.
fMRI Acquisition
Multislice T2-weighted echo-planar volumes with BOLD
contrast (35 axial slices with a voxel resolution of 3 ×
3 × 3 mm covering most of the cerebrum, repetition time =
2.975 sec, echo time = 50 msec, acquisition time =
2.925 sec) were obtained using a 1.5-T MRI scanner
(Siemens TIM Avanto, Erlangen, Germany). Functional im-
ages were acquired in four scanning runs lasting approxi-
mately 8 min 40 sec each in which 174 volumes were
obtained. The first four volumes of each run were
discarded to allow for T1 equilibrium effects. A 3-D T1-
weighted fast-field echo anatomical image lasting 5 min
30 sec was acquired after the first two functional runs for
each participant.
Data Analysis
Behavioral data. A 2 (Task) × 4 (Condition) × 2 (Age
group) mixed rmANOVA was performed on median RT
data. A 2 (Task) × 2 (Age group) mixed rmANOVA was
employed to analyze mean consistency, which was cal-
culated in the same way as in Study 1.
MRI data. MRI data were preprocessed and analyzed
using SPM8 ( Wellcome Trust Centre for Neuroimaging,
London, United Kingdom; www.fil.ion.ucl.ac.uk/spm/).
Images were realigned to the first analyzed volume with
a second-degree B-spline interpolation to correct for
movement during the session. The bias-field-corrected
structural image was coregistered to the mean, realigned
functional image and segmented on the basis of Montreal
Neurological Institute (MNI)-registered International
Consortium for Brain Mapping tissue probability maps.
Resulting spatial normalization parameters were applied
to the realigned images to obtain normalized functional
images with a voxel size of 3 × 3 × 3 mm, which were
smoothed with an 8-mm FWHM Gaussian kernel.
Realignment estimates were used to calculated frame-
wise displacement (FD) for each volume, which is a com-
posite, scalar measure of head motion across the six
realignment estimates (Siegel et al., 2014). Volumes with
an FD > 0.9 mm were censored and excluded from gen-
eral linear model estimation by including a regressor of
no interest for each censored volume. Scanning sessions
with more than 10% of volumes censored or a root mean
square (RMS) movement over the whole session greater
als 1.5 mm (one session for three participants, two ses-
sions for one participant) were excluded from the analy-
Schwester. Adolescent and adult participants significantly differed
in the number of overall censored volumes (Madolescents =
3.39, SD = 3.61; Madults = 0.19, SD = 0.38; P < .001),
mean RMS translational movement (Madolescents = 0.32 mm,
SD = 0.11 mm; Madults = 0.24 mm, SD = 0.07 mm; p =
.005), and mean FD (Madolescents = 0.18 mm, SD = 0.08 mm;
Madults = 0.10 mm, SD = 0.02 mm; p < .001). There was
no difference between groups in terms of mean RMS rota-
tional movement (Madolescents = 0.23 mm, SD = 0.12 mm;
Madults = 0.17 mm, SD = 0.08 mm; p = .088).
Scanning runs were treated as separate time series,
and each series was modeled by a set of regressors in
the general linear model. Runs of the People or Town
Task were each modeled by six box-car regressors: four
regressors corresponding to each Condition (Self, Other,
Comparison, and Vowels), with a duration of 18.6 sec; In-
structions, with a duration of 1 sec; and Fixation blocks,
with a duration of 18.6 sec except for the last block,
which had a duration of 39 sec. All regressors were con-
volved with a canonical hemodynamic response function
and, together with the separate regressors representing
each censored volume and the mean over scans, com-
prised the full model for each session. The data and
model were high-pass filtered to a cutoff of 1/128 Hz.
The second-level whole-brain analysis focused on rela-
tional integration, that is, the main effect of Comparison
(2-REL) versus Self and Other (1-REL) conditions, and on
differences between social and nonsocial
tasks. The
1-REL conditions, Self and Other, were thus combined
within each task. Four first-level contrasts were calculated
using the Vowels condition as a baseline within each task:
People (Self, Other) − People Vowels (People SO),
Magis-Weinberg, Blakemore, and Dumontheil
1745
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
o
6
c
7
n
1
_
1
a
/
_
j
0
o
1
c
1
n
5
3
_
a
p
_
d
0
1
b
1
y
5
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
People Comparison − People Vowels (People Comp),
Town (Self, Other) − Town Vowels (Town SO), Town
Comparison − Town Vowels (Town Comp). These con-
trasts were then entered into a random effects analysis
using a Participant × Age group (2) × Block type (4) flex-
ible factorial design, modeling Participant as a main effect
(to account for the repeated-measure nature of the data)
and the Age group × Block type interaction.
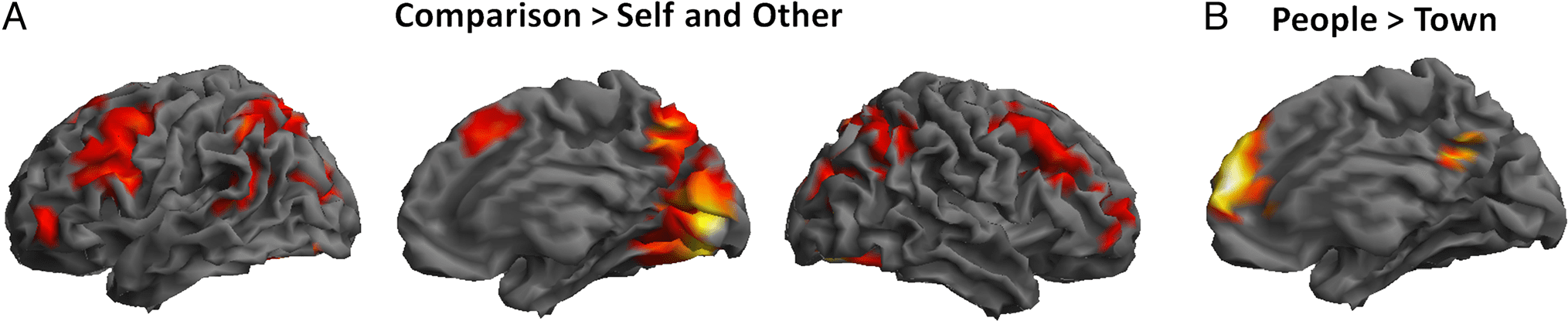
Main effects of Condition (Comparison > SO) Und
Task (People > Town and Town > People) and the inter-
action between the two factors and with Age group were
determined using the t statistic on a voxel-by-voxel basis.
Statistical contrasts were used to create SPMs thresholded
at p < .001 at the voxel level and at family-wise error
(FWE) corrected p < .05 at the cluster level (correspond-
ing to a minimum cluster size of 77 voxels determined
with SPM8). Activations that survived whole-brain FWE
correction at p < .05 at the voxel level are indicated. All
coordinates are given in MNI space. Significant inter-
actions were followed up by extracting the mean signal
across all voxels of significant clusters with MarsBar (Brett,
Anton, Valabregue, & Poline, 2002) and analyzing simple
effects in SPSS using t tests (with Bonferroni correction
for multiple comparisons).
We performed exploratory correlation analyses be-
tween consistency in ratings and individual differences
in activation in the tasks. Relevant task contrasts were
entered in a two-sample t test design modeling the two
age groups separately, with the behavioral measure as
a single covariate of interest. Correlations were run be-
tween the contrasts [Comparison > SO] Und [Comparison/
Self/Other > Vowels] and the mean consistency across
tasks, between the contrast [People Comparison > SO]
and consistency in the People task, and between the
Kontrast [Town Comparison > SO] and consistency in the
Town task. In the same manner, we explored correlations
between individual differences in the [People > Town] Und
[People Comparison > SO] contrasts and the PT scale of
the IRI (see Meyer et al., 2015, for a similar approach).
Ergebnisse
Verhaltensergebnisse
RTs. There was a main effect of Task (F(1, 37) = 13.51,
p = .001, ηp
2 = 0.27). In contrast to Study 1, Teilnehmer
were slower in the Town (M = 2419 ms, SE = 69 ms)
relative to the People (M = 2282 ms, SE = 63 ms) Aufgabe.
There was a main effect of Condition (F(1.8, 66.8) = 29.17,
P < .001, ηp
2 = 0.44; Table 2). Pairwise comparisons with
Bonferroni correction revealed that participants were
slowest in the Comparison condition relative to all other
conditions (MComparison = 2638 ± 79 msec, MSelf = 2180 ±
67 msec, MOther = 2196 ± 62 msec, MVowels = 2388 ± 77 msec;
all ps < .005). The Vowels condition was the next slowest
(all ps < .05). Self and Other conditions did not differ sig-
nificantly from one another ( p > .05). In contrast to Study 1,
there was no main effect of Age group (F(1, 37) = 2.21).
There was a significant interaction between Task and
Condition (F(3, 111) = 3.14, p = .028, ηp
2 = 0.08). Das
was followed up by analyzing the data in the People and
Town tasks separately. In both the People and Town
tasks, participants were slower in Comparison than Self
and Other trials (all ps < .001). In the People task, par-
ticipants were slower in Vowels than Self and Other trials
(all ps < .05). The two-way interaction is driven by a
greater difference between Comparison and Vowels trials
in the Town task (MComp-Vowels = 342 msec) than in the
People task (MComp-Vowels = 157 msec, p = .008), whereas
the difference between Comparison and Self and Other
trials did not differ between tasks ( ps > .05). Im Gegensatz
to Study 1, there was no significant interaction between
Condition and Age group (F(1.8, 66.8) = 0.58). In line
with Study 1, there were no significant interactions
between Task and Age group (F(1, 37) = 1.43) or among
Condition, Task, and Age group (F(2.7, 98.8) = 1.76).
Consistency. As in Study 1, participants were consistent
overall, with positive correlations between the 1-REL and
2-REL ratings. Similar to the pattern in Study 1, there was
a trend effect of Age group (F(1, 37) = 3.46, p = .071, ηp
2 =
0.09; Figure 2D), with lower consistency in adoles-
cents than adults. There was also a main effect of Task
(F(1, 37) = 18.04, P < .001, ηp
2 = 0.33): Consistency was
lower for People than for Town (MPeople = 0.74 ± 0.05,
MTown = 0.96 ± 0.07). As in Study 1, the interaction between
Task and Age group was not significant (F(1, 37) = 0.18).
IRI. Analyses of the subscales of the IRI revealed signif-
icantly higher PT and EC scores for adults (MPT = 19.5,
SE = 0.91; MEC = 20.9, SE = 1.1) than for adolescents
Table 2. Mean RT and SE (msec) for Each Task (People, Town) and Condition (Self, Other, Comparison, Vowels) in Study 2
Self
Other
Comparison
Vowels
People
Town
Adolescents
2215 (110)
2123 (94)
2570 (108)
2503 (112)
Adults
2057 (107)
2112 (92)
2460 (106)
2212 (109)
Adolescents
2333 (100)
2378 (96)
2904 (136)
2529 (120)
Adults
2115 (98)
2171 (94)
2618 (133)
2309 (117)
1746
Journal of Cognitive Neuroscience
Volume 29, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
o
6
c
7
n
1
_
1
a
/
_
j
0
o
1
c
1
n
5
3
_
a
p
_
d
0
1
b
1
y
5
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 3. Summary of Neuroimaging Results
Main effect of condition (Comparison > SO)
L/R
BA
MNI (X, j, z)
Z Score
Cluster Size
Lingual gyrus
Calcarine gyrus
Lingual gyrus
Lingual gyrus
Precuneus
Middle occipital gyrus
Middle occipital gyrus
Middle temporal gyrus
Inferior parietal gyrus
Fusiform gyrus
Inferior frontal gyrus
Precentral and middle frontal gyri
Middle cingulate cortex and pre-SMA
Inferior frontal gyrus
Middle frontal gyrus
Inferior frontal gyrus
Middle frontal gyrus
Middle and superior frontal gyri
Precentral gyrus
Inferior and middle frontal gyri
Main effect of task (People > Town)
MPFC
ACC
ACC
Anterior insula
Precuneus and posterior cingulate cortex
Main effect of task (Town > People)
Lingual gyrus
Calcarine gyrus
Calcarine gyrus
Middle occipital gyrus
Middle occipital gyrus
Superior occipital gyrus
Superior occipital gyrus
Fusiform and parahippocampal gyri
Fusiform gyrus
L
L
R
R
R
L
L
L
R
L
L
R
L
R
R
R
R
R
L
R
L
L
R
R
R
R
L
L
R
L
R
L
R
18
17
18
18
7
19
19
21
40
19
48
6
32
48
44
44
8
10
6
47
10
32
10
48
23
30
30
30
19
19
23
7
37
37
−9, −85, −14
0, −85, 1
−21, −79, −14
18, −79, −14
3, −61, 46
30, −79, 19
−30, −76, 22
−48, −46, 10
−45, −46, 43
27, −64, −5
−48, 14, 25
−39, −1, 55
9, 20, 46
−36, 20, 22
30, 14, 43
48, 26, 31
30, 23, 52
30, 59, 7
30, −4, 46
−39, 47, −2
6, 53, 13
−3, 53, 13
−6, 44, 1
30, 17, −17
6, −52, 28
9, −49, 4
15, −52, 13
−12, −55, 10
−33, −76, 28
36, −70, 37
−21, −64, 28
24, −76, 46
>8a
>8a
>8a
>8a
>8a
7.31A
6.53A
6.22A
6.13A
5.90A
6.59A
5.78A
5.77A
5.68A
5.55A
5.32A
5.03A
4.56A
4.44
4.91A
7.22A
7.11A
5.95A
5.74A
5.26A
4,959B
1,351B
819B
95B
1,649B
69
132B
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
.
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
>8a
2,164B
7.65A
7.17A
6.08A
5.90A
5.25A
4.62A
−30, −37, −14
>8a
30, −31, −17
6.97A
192B
302B
Magis-Weinberg, Blakemore, and Dumontheil
1747
MNI (X, j, z)
Z Score
Cluster Size
Tisch 3. (Fortsetzung )
Inferior temporal gyrus
Parahippocampal gyrus
Middle cingulate cortex
Precentral gyrus and inferior frontal operculum
Postcentral and precentral gyri
L/R
R
R
R
L
R
BA
20
35
23
44
43
54, −46, −11
21, −13, −20
9, −34, 34
−42, 8, 28
60, −10, 31
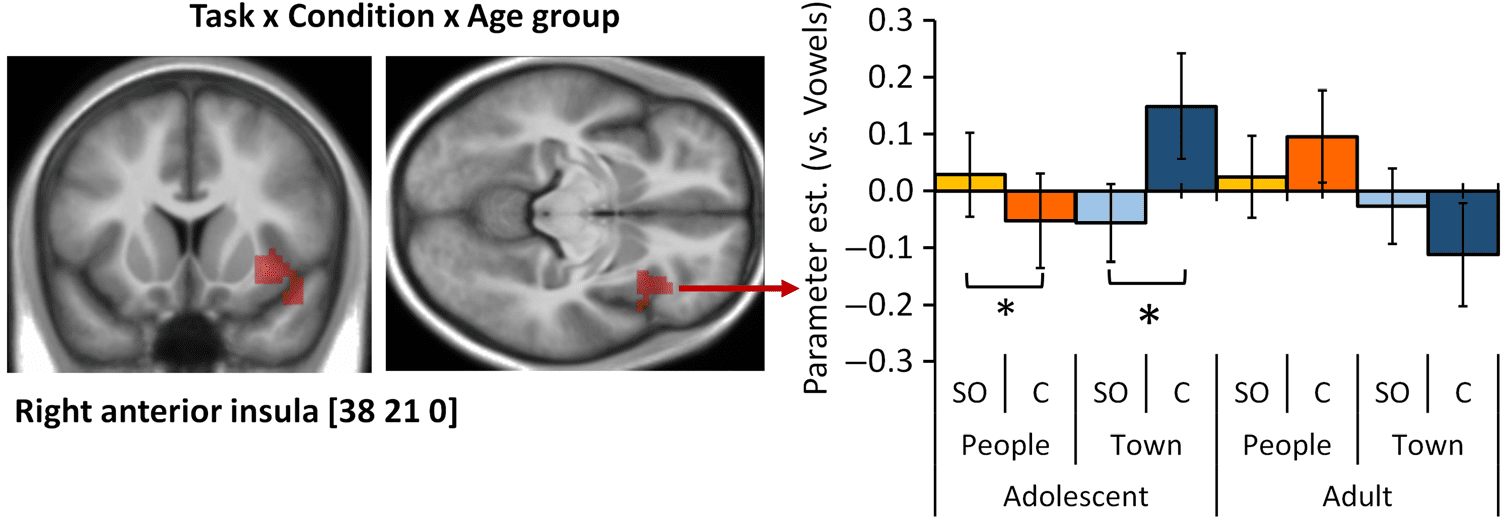
Interaction adolescents > adults [(Town Comparison > SO) > [People (Comparison > SO)]
Anterior insula
Anterior insula
Superior temporal pole
R
R
R
48
47
38
33, 17, −8
39, 26, 4
45, 14, −20
4.13
3.73
4.78A
4.58A
3.93
4.26
3.71
3.59
60
181B
108B
178B
Coordinates and Z scores are listed for regions showing a significant difference in BOLD signal for the main effect of Condition [Comparison > SO],
the main effect of Task [People > Town] oder [Town > People], and the interaction between Condition, Task, and Age Group [(Adolescents > Adults
[(Town Comparison > SO) > [People (Comparison > SO)]]. Region labeling was done using automatic anatomical labeling (Tzourio-Mazoyer et al.,
2002). BA labeling of peak of activations was done using MRIcron. L/R = left/right hemisphere.
aVoxels where pFWE < .05 at the voxel level. bClusters where pFWE < .05 at the cluster level, with a cluster-defining threshold of p < .001 uncorrected at the voxel level. (MPT = 14.1, SE = 0.94; MEC = 16.8, SE = 1.1; all ps < .05). Pearson correlation coefficients were computed to assess the relationship between PT and performance in the task. There was no correlation between PT and Con- sistency in the People task (r = −.187, p = .255). There was also no correlation between PT and mean median RT for the Self (r = −.021, p = .897), Other (r = .006, p = .973), or Comparison (r = .068, p = .680) conditions in the People task. Relational integration. A broad bilateral network of frontoparietal, temporal, and occipital regions, including bilateral RLPFC, showed increased BOLD signal in Com- parison versus SO conditions (Table 3 and Figure 3A), that is, in 2-REL as opposed to 1-REL processing. In- creases in BOLD signal were observed in a large posterior cluster extending into bilateral occipital and lingual gyri, calcarine sulcus, and inferior parietal lobule and in ante- rior clusters in the precentral gyrus, pre-SMA, and inferior and middle frontal gyri. fMRI Results Whole-brain analyses contrasted the Comparison condi- tion to the combined Self and Other (SO) conditions in the People and Town tasks. The Vowels condition served as an active baseline. Social Information Processing When comparing the People task with the Town task, that is, when the information to be processed was of social (traits of participant or a friend) versus nonsocial Figure 3. fMRI results across age groups. (A) Main effect of Condition. Regions showing increased BOLD signal in Comparison (2-REL) compared with Self and Other (1-REL) conditions are rendered on the SPM8 surface mesh template. From left to right: lateral view of the left hemisphere, medial and lateral views of the right hemisphere. (B) Main effect of Task. Regions showing increased BOLD signal in the People compared with the Town task are rendered on the SPM8 surface mesh template (medial view of the right hemisphere). 1748 Journal of Cognitive Neuroscience Volume 29, Number 10 D o w n l o a d e d f r o m l l / / / / j t t f / i t . : / / h t t p : / D / o m w i n t o p a r d c e . d s f i r o l m v e h r c p h a d i i r r e . c c t . o m m / j e o d u c n o / c a n r a t r i t i c c l e e - p - d p d 2 f 9 / 1 2 0 9 / 1 1 7 0 3 / 9 1 1 7 9 3 5 9 3 / 0 1 5 4 7 8 o 6 c 7 n 1 _ 1 a / _ j 0 o 1 c 1 n 5 3 _ a p _ d 0 1 b 1 y 5 g 3 u . e p s t d o f n b 0 y 7 S M e I p T e m L i b b e r r a 2 r 0 2 i 3 e s / j / . f t u s e r o n 1 7 M a y 2 0 2 1 Figure 4. fMRI results of the interaction between Task, Condition, and Age group. On the left, the right anterior insula cluster showing a three-way interaction between Task, Condition, and Age group is shown on an average structural scan of all participants in the study (left: y = 21, right: z = 0). The contrast was thresholded at p < .001 uncorrected at the voxel level, pFWE < .05 at the cluster level. On the right, parameter estimates extracted from this cluster are plotted in a bar chart to illustrate the three-way interaction. Error bars represent SE. † p < .1, *p < .05, **p < .01, ***p < .001. (characteristics of towns) nature, increased BOLD signal was observed in clusters in the MPFC, insula, and precuneus (Table 3 and Figure 3B). The reverse contrast revealed increased BOLD signal in a large cluster extend- ing into bilateral calcarine gyri, middle and superior oc- cipital gyri, and bilateral clusters in the fusiform and parahippocampal gyri extending into the medial tempo- ral gyri, middle cingulate cortex, bilateral precentral and postcentral gyri, and left inferior frontal cortex (Table 3). Age group differences. There was no two-way interac- tion between Task and Age group or between Condition and Age group; however, whole-brain analyses showed a significant three-way interaction between Task, Condi- tion, and Age group in BOLD signal in the right insula (see Table 3 and Figure 4). The mean parameter estimates in this cluster were calculated, and the interac- tion was followed up by running 2 (Condition) × 2 (Age group) mixed rmANOVAs in each Task separately. There was an interaction between Condition and Age group in the People task (F(1, 37) = 7.20, p = .011, ηp 2 = 0.16) and in the Town task (F(1, 37) = 8.64, p = .006, ηp 2 = 0.19). These interactions were further explored by compar- ing SO and Comparison conditions within each age group. In the People task, adolescents showed greater ac- tivation in the SO than the Comparison condition ( p = .026), whereas the conditions did not differ in adults ( p = .132). In the Town task, adolescents reversely showed greater activation in the Comparison than the SO condition ( p = .018), whereas again, there was no difference in adults ( p = .176). Covariate Analyses Whole-brain analyses were performed to investigate cor- relations between the behavioral measure of Consistency and BOLD signal during the task. No correlation between Consistency scores and individual differences in BOLD signal in the contrasts [Comparison > SO], [Comparison/
Self/Other > Vowels], [People Comparison > People SO],
Und [Town Comparison > Town SO]) was observed.
Further whole-brain analyses showed that BOLD signal in
Die [People > Town] Und [People Comparison > People
SO] contrasts was not significantly related to the PT scale
of the IRI.
DISKUSSION
We performed separate behavioral and fMRI studies aim-
ing to disentangle general and specific processes under-
lying relational integration of social information between
late childhood and adulthood. We found behavioral evi-
dence for general development of social and nonsocial
relational reasoning. We discovered similar patterns of
neural activity for adolescents and adults showing do-
main general involvement of the frontoparietal cortex
areas associated with relational integration for both
social and nonsocial relations and domain-specific in-
volvement of the social brain for the manipulation of
social information.
Behavioral Findings
Relational Integration
In Study 1, the large behavioral study, we found earlier
improvements in performance with age for RT and later
improvements in consistency from late childhood to
Erwachsensein. Across age groups, RTs were slower in the
Comparison condition, which required relational integra-
tion, than the Self and Other conditions, which required
processing of a single relation. These results are in line
with the robust processing speed costs observed in par-
adigms comparing relational integration with simpler re-
lational processing (Dumontheil et al., 2010; Crone et al.,
2009). In terms of relational integration performance, Re-
sults show a pattern of early improvement in RT between
ages of 11–12 and 13–14 years, with no further changes at
older ages (Figure 2B), and improvements between ages
Magis-Weinberg, Blakemore, and Dumontheil
1749
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
of 13–14 and 15–17 years for the consistency measure,
with no improvements at older ages (Figure 2C). Notiz
that verbal IQ was not matched across age groups;
Jedoch, the differences in verbal IQ did not directly
map onto the observed age effects. Insbesondere, Dort
was no difference in IQ between 11- Zu 12-, 13- Zu 14-,
Und 15- to 17-year age groups, the age range where the
key developmental changes were observed.
Ähnlich, in Study 2, the fMRI study, RTs were slower
in the Comparison than in the Self and Other conditions.
Relational integration was slower in adolescents than
Erwachsene. There was also a trend for poorer consistency in
the adolescents, which fits with the findings of the behav-
ioral study. The less robust behavioral results in Study 2
compared with Study 1 were likely due to the smaller
sample size in the imaging study.
Gesamt, we observed that late childhood and early ad-
olescence are associated with poorer relational integra-
tion performance than adulthood (Figure 2B). Das
pattern fits with previous findings from visuospatial rela-
tional integration tasks, which indicate poorer relational
integration accuracy in 8- to 12-year-olds than in adults
(Crone et al., 2009), improvements in accuracy between
9- and 19-year-olds (Rosso, Jung, Femia, & Yurgelun-
Todd, 2004), and poorer combined accuracy and RT in
7- to 9-year-olds than in 14- to 17-year-olds (Dumontheil
et al., 2010; reanalyzed in Dumontheil, 2014), although
note that Wendelken et al. (2011) did not find age differ-
ences in 7- to 18-year-olds (see Dumontheil, 2014, for a
Rezension). This study is consistent with a previous investi-
gation of the development of the integration of semantic
information using an analogical reasoning task, welche dm-
onstrated poorer performance in 6- to 13-year-olds than
bei Erwachsenen (Wright, Matlen, Baym, Ferrer, & Bunge, 2008).
Social Information Processing
Developmental improvements in RT for relational inte-
gration did not vary as a function of the type of informa-
tion. Jedoch, main effects of task across age were
observed. In Study 1, RTs were faster overall for Town
than People, whereas the opposite pattern was observed
in Study 2 in which RTs were faster overall for the People
task regardless of the level of relational processing,
which is in line with previous findings of benefits in per-
formance when stimuli are social rather than symbolic
(Dumontheil, Hillebrandt, Apperly, & Blakemore, 2012;
den Ouden, Frith, Frith, & Blakemore, 2005).
Both adolescents and adults showed increased speed
for social information. Ähnlich, although consistency
was overall greater in the nonsocial task, domain-general,
not social-specific, developmental changes were ob-
serviert. This is at odds with studies showing increased
sensitivity to social stimuli during adolescence (Foulkes
& Blakemore, 2016). This might be due to the low arousal
and/or affective demands of the current task and should
be explored in further studies. Außerdem, poorer over-
all consistency in the social task may be due to the greater
complexity and variability of people’s traits compared
with towns’ characteristics; this difference would apply
to both adolescents and adults. Zum Beispiel, my friend
Sam may be funny sometimes, but at other times, he is
quite grumpy, while London is always busy.
Slight discrepancies between the findings from Studies
1 Und 2 may be explained by methodological differences.
Study 1 had a larger sample than Study 2, reducing the
power needed to detect developmental changes in per-
Form. In Study 1, the adjectives were the same for
both the People and Town tasks. In Study 2, to maximize
the mentalizing requirements of the People task, we used
different adjectives for each task, which were more di-
rectly applicable to people or towns. The fMRI task was
not self-paced. These factors may have affected the be-
havioral results.
Our behavioral findings provide some evidence for dif-
ferential performance for social information, regardless
of relational level, for both adolescents and adults. Fur-
thermore, we provide evidence for domain-general
development of relational integration of simple relations,
which does not differ as a function of the social or
nonsocial nature of the semantic information being
processed.
Neuroimaging Findings
Relational Integration
Relational integration was associated with greater activa-
tion in a large bilateral frontoparietal network including
the RLPFC in both the People and Town tasks. A similar
pattern of activation was observed by Raposo et al. (2011)
in the Comparison versus Other contrast. These results
further support the involvement of RLPFC and the infe-
rior parietal cortex in relational integration ( Wendelken
et al., 2012; Crone et al., 2009). By adapting the task by
Raposo et al., we are able to directly compare manipula-
tion of social and nonsocial information and provide evi-
dence for domain-general recruitment of the RLPFC
through adolescence and adulthood across social and
nonsocial domains.
Gesamt, we did not find evidence of an interaction be-
tween relational integration and social versus nonsocial
Aufgabe: Both networks were recruited in parallel for rela-
tional integration and social demands. This parallel
recruitment of the two networks is similar to that ob-
served by Meyer et al. (2012, 2015) in their social working
memory task. Jedoch, these studies showed that MPFC
activation associated with social working memory was
modulated by working memory load (Meyer et al.,
2012, 2015), whereas in this study, MPFC activation was
not modulated by the number of relations participants
had to consider. Ähnlich, although Meyer et al. (2012)
found that PT on the IRI scale was positively associated
with social working memory load-dependent activity
1750
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
.
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
within the MPFC and posterior cingulate cortex in adults,
we found no association between self-reported PT on this
questionnaire and behavior or brain activity in our task. A
possible source of this difference in findings is that social
cognitive load in the Meyer et al. (2012) study was higher
than in this study, with the requirement to compare two,
three, or four individuals, whose names were maintained
in working memory, on given personality traits. In diesem
Studie, our participants only compared two individuals,
and the relevant information remained on the screen
over the duration of the trial. These differences may
have contributed to the lack of observed association be-
tween performance, MPFC activation, and self-reported
real-life PT.
We did not observe developmental differences in acti-
vation in the RLPFC during relational integration across
tasks in whole-brain analyses. Developmental differences
have been reported in this brain region (z.B., Dumontheil
et al., 2010; Crone et al., 2009). Jedoch, in the study by
Crone et al., the age effects were mainly due to differ-
ences in time course of activations evident in their
event-related design, which our block design did not
allow us to test. In our previous study, age group effects
were observed in ROI versus whole-brain analyses only
(Dumontheil et al., 2010). Relational reasoning studies
have traditionally involved demanding visuospatial rea-
soning tasks, such as the Raven Progressive Matrices. Es
might be that semantic reasoning about traits of people
and towns is not demanding enough to tax adolescents
in the same way.
Our results suggest domain-general recruitment of the
RLPFC through adolescence and adulthood, independent
of whether the information being manipulated is social or
nonsocial. These results are therefore in line with the
finding that the integration of visuospatial or semantic re-
lations elicits similar activation of the relational integra-
tion network ( Wendelken et al., 2012).
Social vs. Nonsocial Information Processing
Social information was associated with greater activation
in the precuneus and MPFC in both adolescents and
Erwachsene. These results are in line with a large body of
literature that documents the involvement of these re-
gions when processing social information (z.B., see Van
Overwalle, 2009, for a meta-analysis). Beachten Sie, dass, although
towns can be considered social to some extent, as one
can imagine the population of individuals living there,
characteristics of towns were considered to be less social
than traits of people.
Similar to this study, the investigation of social rela-
tional reasoning in adults by Raposo and colleagues
(2011) reported greater MPFC activation in Self, Other,
and Relational (Comparison) conditions when contrasted
to a Vowel judgment condition. Jedoch, in contrast to
our findings, Raposo et al. (2011) observed activation
across ventral and dorsal MPFC in the Other versus Self
contrast and no activation in the Self > Other contrast.
This difference between the two studies may be due to
the precise question participants were asked to answer.
In this study, participants simply rated how funny (oder
other adjectives) sie waren, or their friend was, wohingegen
in the Raposo et al. (2011) Studie, participants always
rated how pleasant or unpleasant they found a concept
(z.B., tower) or how pleasant or unpleasant they thought
their friend would find this concept. It is possible that
this latter question elicited greater mentalizing by asking
participants to put themselves in their friend’s shoes
rather than asking their own opinion. Gesamt, our results
suggest social-specific recruitment of the MPFC during
adolescence and adulthood for social information for
both simple relations and integration across levels.
Interaction between Task, Condition, and Age Group
One neural difference between age groups was observed.
At the cluster-corrected level, the right anterior insula
showed a significant three-way interaction between Con-
dition, Task, and Age group, driven by differential recruit-
ment according to the domain (social vs. nonsocial) Und
the relational integration requirements with age. Obwohl
these results were not significant with an FWE-corrected
threshold at the voxel level, they replicate the pattern of
decreased activation with age in the anterior insula observed
in a visuospatial relational reasoning task (Dumontheil
et al., 2010). Functional changes in the anterior insula
might reflect the maturation of neurocognitive strategies,
which possibly include changes in task-specific connec-
tivity between brain regions (z.B., see Bazargani et al.,
2014; Dumontheil, 2014; and Dumontheil et al., 2010, für
discussions). This neuroimaging finding does not directly
map onto behavioral differences between the age groups.
In a previous study, we similarly observed that develop-
mental changes in anterior insula activation during rela-
tional reasoning were not accounted for by individual
differences in performance on the task (Dumontheil
et al., 2010). Beyond differences in the sensitivity of be-
havioral and brain imaging measures (z.B., evidenced by
greater sensitivity of neuroimaging data than behavioral
data to genetic differences; Dumontheil et al., 2011), dif-
ferences may be due to the fact that behavior reflects a
large combination of factors beyond the block-related acti-
vations measured in the current fMRI paradigm, wie zum Beispiel
event-related activations, which may have had compen-
satory effects on performance.
Abschluss
We aimed to investigate the development of perfor-
mance in social and nonsocial relational reasoning and
their associated neural substrates. The paradigm required
participants to make first- and second-order relational
judgments about social and nonsocial information. Data
from a behavioral study and an fMRI study demonstrated
Magis-Weinberg, Blakemore, and Dumontheil
1751
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
.
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
development of social and nonsocial relational reasoning
in adolescence. These behavioral results with semantic
stimuli are in line with previous research using mostly
visuospatial relational reasoning tasks. We did not find
evidence of differential development of relational inte-
gration of social versus nonsocial information in our
behavioral studies. Ähnlich, the fMRI data showed that,
in both adolescents and adults, relational integration
of social and nonsocial information recruited a similar
frontoparietal network. The processing of social informa-
tion additionally engaged the MPFC and precuneus regions
of the social brain, regardless of the order of reasoning.
These findings provide further evidence that relational
integration is a domain-general process ( Wendelken
et al., 2012).
APPENDIX: LIST OF STIMULI FOR EACH TASK
IN STUDY 2
In the People task, participants were asked: “How much
do you think the following words apply to you/your
friend?” or “How much do you think the following words
apply to you compared with your friend?” In the Town
Aufgabe, participants were asked: “How much do you think
the following words apply to your town/other town?” or
“How much do you think the following words apply to
Adjectives
People
Town
arrogant, jealous, timid,
selfish, careless, witty,
cheerful, thoughtful,
stubborn, ambitious,
confident, aggressive,
clever, smart, bold, brave,
generous,helpful, mature,
wise, tough, funny, curious,
honest, sensitive, friendly,
fair, patient, bright, happy
run-down, shabby, boring,
rainy, noisy, sleepy, vibrant,
picturesque, polluted, quaint,
historic, lively, dull, exciting,
romantic, dirty, urban, expensive,
dangerous, ländlich, cultural, safe,
unusual, clean, quiet, traditional,
famous, amazing, flat, ancient
Number of letters
M = 6.7, SD = 1.8
Number of vowels
M = 2.5, SD = 1.1
M = 6.6, SD = 1.9
M = 2.6, SD = 1.1
Familiarity
M = 541.6, SD = 48.6
M = 561.6, SD = 53.6
Brown frequency
M = 9.26, SD = 10.6
M = 7.70, SD = 11.0
Kucera–Francis frequency
M = 32.0, SD = 26.9
M = 35.3, SD = 27.2
T(58) = 0.14, p = .89
T(58) = 0.36, p = .73
T(40) = 1.2, p = .23
T(41) = 0.47, p = .64
T(58) = 0.47, p = .64
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
1752
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 10
your town compared with the other town?” Familiarity
and frequency measures were included to ensure that
all adjectives were commonly used English words and
that their occurrence was comparable between Tasks.
Danksagungen
Sarah K. G. Jensen assisted with data collection. L. M. W. Ist
funded by the Mexican National Council on Science and Tech-
nology (CONACYT). S. J. B. is funded by a Royal Society Univer-
sity Research Fellowship, the Wellcome Trust, and the Jacobs
Foundation. This research was supported by the Leverhulme
Trust.
Reprint requests should be sent to Dr. Iroise Dumontheil,
Department of Psychological Sciences, Birkbeck, Universität
London, Malet Street, London WC1E 7HX, Großbritannien,
oder per E-Mail: i.dumontheil@bbk.ac.uk.
VERWEISE
Aichhorn, M., Perner, J., Kronbichler, M., Staffen, W., &
Ladurner, G. (2006). Do visual perspective tasks need theory
of mind? Neurobild, 30, 1059–1068.
Amodio, D. M., & Frith, C. D. (2006). Meeting of minds: Der
medial frontal cortex and social cognition. Nature Reviews
Neurowissenschaften, 7, 268–277.
Bazargani, N., Hillebrandt, H., Christoff, K., & Dumontheil, ICH.
(2014). Developmental changes in effective connectivity
associated with relational reasoning. Menschliches Gehirn
Mapping, 35, 3262–3276.
Blakemore, S.-J. (2008). The social brain in adolescence. Natur
Reviews Neuroscience, 9, 267–277.
Blakemore, S.-J. (2012). Development of the social brain in
adolescence. Journal of the Royal Society of Medicine,
105, 111–116.
Blakemore, S.-J., & Robbins, T. W. (2012). Decision-making in
the adolescent brain. Naturneurowissenschaften, 15, 1184–1191.
Brett, M., Anton, J.-L., Valabregue, R., & Polina, J.-B. (2002).
Analyse des interessierenden Bereichs mithilfe einer SPM-Toolbox. Paper
presented at the 8th International Conference on Functional
Mapping of the Human Brain, Sendai, Japan.
Bunge, S. A., Helskog, E. H., & Wendelken, C. (2009). Links, but not
Rechts, rostrolateral prefrontal cortex meets a stringent test of
the relational integration hypothesis. Neurobild, 46, 338–342.
Bunge, S. A., Wendelken, C., Badre, D., & Wagner, A. D. (2005).
Analogical reasoning and prefrontal cortex: Evidence for
separable retrieval and integration mechanisms. Zerebral
Kortex, 15, 239–249.
Charman, T., Ruffman, T., & Clements, W. (2002). Is there a
gender difference in false belief development?. Sozial
Development, 11, 1–10.
Christoff, K., Prabhakaran, V., Dorfman, J., Zhao, Z., Kroger,
J. K., Holyoak, K. J., et al. (2001). Rostrolateral prefrontal
cortex involvement in relational integration during reasoning.
Neurobild, 14, 1136–1149.
Crone, E. A., & Dahl, R. E. (2012). Understanding adolescence
as a period of social-affective engagement and goal flexibility.
Nature Reviews Neurowissenschaften, 13, 636–650.
Crone, E. A., Wendelken, C., van Leijenhorst, L., Honomichl,
R. D., Christoff, K., & Bunge, S. A. (2009). Neurocognitive
development of relational reasoning. Developmental
Wissenschaft, 12, 55–66.
D’Argembeau, A., Ruby, P., Collette, F., Degueldre, C., Balteau,
E., Luxen, A., et al. (2007). Distinct regions of the medial
prefrontal cortex are associated with self-referential
processing and perspective taking. Zeitschrift für Kognition
Neurowissenschaften, 19, 935–944.
David, N., Aumann, C., Santos, N. S., Bewernick, B. H., Eickhoff,
S. B., Newen, A., et al. (2008). Differential involvement
of the posterior temporal cortex in mentalizing but not
perspective taking. Social Cognitive and Affective Neuroscience,
3, 279–289.
David, N., Bewernick, B. H., Cohen, M. X., Newen, A., Lux, S.,
Fink, G. R., et al. (2006). Neural representations of self versus
andere: Visual-spatial perspective taking and agency in a
virtual ball-tossing game. Zeitschrift für kognitive Neurowissenschaften,
18, 898–910.
Davis, M. H. (1980). A multidimensional approach to individual
differences in empathy. JSAS Catalog of Selected Documents
in Psychology, 85.
Decety, J., & Sommerville, J. A. (2003). Shared representations
between self and other: A social cognitive neuroscience view.
Trends in den Kognitionswissenschaften, 7, 527–533.
den Ouden, H. E., Frith, U., Frith, C., & Blakemore, S.-J. (2005).
Thinking about intentions. Neurobild, 28, 787–796.
Dumontheil, ICH. (2014). Development of abstract thinking during
Kindheit und Jugend: The role of rostrolateral
prefrontal cortex. Developmental Cognitive Neuroscience,
10C, 57–76.
Dumontheil, ICH., & Blakemore, S.-J. (2012). Social cognition and
abstract thought in adolescence: The role of structural and
functional development in rostral prefrontal cortex. British
Journal of Educational Psychology Monograph Series II, 8,
99–113.
Dumontheil, ICH., Bürger, P. W., & Blakemore, S.-J. (2008).
Development of rostral prefrontal cortex and cognitive and
behavioural disorders. Developmental Medicine and Child
Neurologie, 50, 168–181.
Dumontheil, ICH., Hillebrandt, H., Apperly, ICH., & Blakemore, S.-J.
(2012). Developmental differences in the control of action
selection by social information. Zeitschrift für Kognition
Neurowissenschaften, 24, 2080–2095.
Dumontheil, ICH., Houlton, R., Christoff, K., & Blakemore, S.-J.
(2010). Development of relational reasoning during
adolescence. Developmental Science, 13, F15–F24.
Dumontheil, ICH., Jensen, S. K. G., Holz, N. W., Meyer, M. L.,
Lieberman, M. D., & Blakemore, S. J. (2014). Preliminary
investigation of the influence of dopamine regulating
genes on social working memory. Social Neuroscience, 9,
437–451.
Dumontheil, ICH., Roggeman, C., Ziermans, T., Peyrard-Janvid, M.,
Matsson, H., Kere, J., et al. (2011). Influence of the COMT
genotype on working memory and brain activity changes
during development. Biologische Psychiatrie, 70, 222–229.
Ferrer, E., O’Hare, E. D., & Bunge, S. A. (2009). Fluid reasoning
and the developing brain. Frontiers in Neuroscience, 3,
46–51.
Foulkes, L., & Blakemore, S-J. (2016). Is there heightened
sensitivity to social reward in adolescence? Current Opinion
in Neurobiology, 40, 81–85.
Frith, U., & Frith, C. D. (2003). Development and
neurophysiology of mentalizing. Philosophische Transaktionen
der Royal Society of London, Serie B, Biological Sciences,
358, 459–473.
Gilbert, S. J., Spengler, S., Simmons, J., Steele, J., Lawrie, S.,
Frith, C. D., et al. (2006). Functional specialization within
rostral prefrontal cortex (Bereich 10): A meta-analysis. Zeitschrift für
Cognitive Neuroscience, 18, 932–948.
Gusnard, D. A. (2005). Being a self: Considerations from functional
Bildgebung. Consciousness and Cognition, 14, 679–697.
Herting, M. M., Maxwell, E. C., Irvine, C., & Nagel, B. J. (2012).
The impact of sex, puberty, and hormones on white matter
microstructure in adolescents. Hirnrinde, 22, 1979–1992.
Magis-Weinberg, Blakemore, and Dumontheil
1753
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
F
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Kilford, E. J., Dumontheil, ICH., Holz, N. W., & Blakemore, S.-J.
(2015). Influence of COMT genotype and affective distractors
on the processing of self-generated thought. Social Cognitive
und affektive Neurowissenschaften, 10, 777–782.
Kilford, E. J., Garrett, E., & Blakemore, S.-J. (2016). Der
development of social cognition in adolescence: Ein
integrated perspective. Neurowissenschaften & Biobehavioral
Rezensionen, 70, 106–120.
Kroger, J. K., Sabb, F. W., Fales, C. L., Bookheimer, S. Y., Cohen,
M. S., & Holyoak, K. J. (2002). Recruitment of anterior
dorsolateral prefrontal cortex in human reasoning: A
parametric study of relational complexity. Hirnrinde,
12, 477–485.
Lynn, R., & Irwing, P. (2004). Sex differences on the progressive
matrices: A meta-analysis. Intelligence, 32, 481–498.
Meyer, M. L., Spunt, R. P., Berkman, E. T., Taylor, S. E., &
Lieberman, M. D. (2012). Evidence for social working memory
from a parametric functional MRI study. Verfahren der
Nationale Akademie der Wissenschaften, USA., 109, 1883–1888.
Meyer, M. L., Taylor, S. E., & Lieberman, M. D. (2015). Sozial
working memory and its distinctive link to social cognitive
ability: Eine fMRT-Studie. Social Cognitive and Affective
Neurowissenschaften, 10, 1338–1347.
Mills, K. L., Lalonde, F., Clasen, L. S., Giedd, J. N., & Blakemore,
S. J. (2014). Developmental changes in the structure of the
social brain in late childhood and adolescence. Sozial
Cognitive and Affective Neuroscience, 9, 123–131.
Rameson, L. T., Satpute, A. B., & Lieberman, M. D. (2010).
The neural correlates of implicit and explicit self-relevant
Verarbeitung. Neurobild, 50, 701–708.
Raposo, A., Vicens, L., Clithero, J. A., Dobbins, ICH. G., & Huettel,
S. A. (2011). Contributions of frontopolar cortex to
judgments about self, others and relations. Social Cognitive
und affektive Neurowissenschaften, 6, 260–269.
Raven, J., Raven, J. C., & Court, J. H. (1998). Manual for
Raven’s progressive matrices and vocabulary scales.
Oxford: Oxford Psychologists Press.
Raznahan, A., Shaw, P., Lalonde, F., Stockman, M., Wallace,
G. L., Greenstein, D., et al. (2011). How does your cortex
grow? Zeitschrift für Neurowissenschaften, 31, 7174–7177.
Rosso, ICH. M., Jung, A. D., Femia, L. A., & Yurgelun-Todd, D. A.
(2004). Cognitive and emotional components of frontal lobe
functioning in childhood and adolescence. Annals of the
New York Academy of Sciences, 1021, 355–362.
Ruby, P., & Decety, J. (2001). Effect of subjective perspective
taking during simulation of action: A PET investigation of
Agentur. Naturneurowissenschaften, 4, 546–550.
Ruby, P., & Decety, J. (2004). How would you feel versus how
do you think she would feel? A neuroimaging study of
perspective-taking with social emotions. Zeitschrift für Kognition
Neurowissenschaften, 16, 988–999.
Siegel, J. S., Power, J. D., Dubis, J. W., Vogel, A. C., Church, J. A.,
Schlaggar, B. L., et al. (2014). Statistical improvements in
functional magnetic resonance imaging analyses produced by
censoring high-motion data points. Kartierung des menschlichen Gehirns,
35, 1981–1996.
Schmied, R., Keramatian, K., & Christoff, K. (2007). Localizing the
rostrolateral prefrontal cortex at the individual level.
Neurobild, 36, 1387–1396.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neurobild, 15, 273–289.
Van Overwalle, F. (2009). Social cognition and the brain: A
meta-analysis. Kartierung des menschlichen Gehirns, 30, 829–858.
Vogeley, K., Mai, M., Ritzl, A., Falkai, P., Zilles, K., & Fink, G. R.
(2004). Neural correlates of first-person perspective as one
constituent of human self-consciousness. Zeitschrift für
Cognitive Neuroscience, 16, 817–827.
Wechsler, D. (1999). Wechsler Abbreviated Scale of Intelligence
( WASI). San Antonio, TX: Psychological Corporation.
Wendelken, C., & Bunge, S. A. (2009). Transitive inference:
Distinct contributions of rostrolateral prefrontal cortex and
the hippocampus. Zeitschrift für kognitive Neurowissenschaften, 22,
837–847.
Wendelken, C., Chung, D., & Bunge, S. A. (2012). Rostrolateral
prefrontal cortex: Domain-general or domain-sensitive?
Kartierung des menschlichen Gehirns, 33, 1952–1963.
Wendelken, C., Ferrer, E., Whitaker, K. J., & Bunge, S. A. (2016).
Fronto-parietal network reconfiguration supports the
development of reasoning ability. Hirnrinde, 26,
2178–2190.
Wendelken, C., Nakhabenko, D., Donohue, S. E., Fuhrmann, C. S.,
& Bunge, S. A. (2008). “Brain is to thought as stomach
is to ??”: Investigating the role of rostrolateral prefrontal
cortex in relational reasoning. Zeitschrift für Kognition
Neurowissenschaften, 20, 682–693.
Wendelken, C., O’Hare, E. D., Whitaker, K. J., Ferrer, E., &
Bunge, S. A. (2011). Increased functional selectivity over
development in rostrolateral prefrontal cortex. Zeitschrift für
Neurowissenschaften, 31, 17260–17268.
Holz, J. N., & Grafman, J. (2003). Human prefrontal cortex:
Processing and representational perspectives. Natur
Reviews Neuroscience, 4, 139–147.
Wright, S. B., Matlen, B. J., Baym, C. L., Ferrer, E., & Bunge, S. A.
(2008). Neural correlates of fluid reasoning in children and
Erwachsene. Grenzen der menschlichen Neurowissenschaften, 1, 8.
Zysset, S., Huber, O., Samson, A., Ferstl, E. C., & von Cramon, D. Y.
(2003). Functional specialization within the anterior medial
prefrontal cortex: A functional magnetic resonance imaging
study with human subjects. Neurowissenschaftliche Briefe, 335, 183–186.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
0
9
/
1
1
7
0
3
/
9
1
1
7
9
3
5
9
3
/
0
1
5
4
7
8
Ö
6
C
7
N
1
_
1
A
/
_
J
0
Ö
1
C
1
N
5
3
_
A
P
_
D
0
1
B
1
j
5
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
1754
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 10