Slow Endogenous Fluctuations in Cortical fMRI
Signals Correlate with Reduced Performance
in a Visual Detection Task and Are
Suppressed by Spatial Attention
David W. Bressler, Ariel Rokem*, and Michael A. Silver
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
1
e
8
–
5
P
D
1
9
F
3
/
1
3
1
2
4
/
4
1
/
Ö
8
C
5
N
/
_
A
1
_
8
0
6
1
1
4
4
7
0
0
0
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
4
N
7
0
0
8
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
T
F
/
.
Ö
N
0
5
M
A
j
2
0
2
1
Abstrakt
■ Spatial attention improves performance on visual tasks, In-
creases neural responses to attended stimuli, and reduces cor-
related noise in visual cortical neurons. In addition to being
visually responsive, many retinotopic visual cortical areas exhibit
very slow (<0.1 Hz) endogenous fluctuations in functional mag-
netic resonance imaging signals. To test whether these fluctua-
tions degrade stimulus representations, thereby impairing visual
detection, we recorded functional magnetic resonance imaging
responses while human participants performed a target detec-
tion task that required them to allocate spatial attention to either
a rotating wedge stimulus or a central fixation point. We then
measured the effects of spatial attention on response amplitude
at the frequency of wedge rotation and on the amplitude of en-
dogenous fluctuations at nonstimulus frequencies. We found
that, in addition to enhancing stimulus-evoked responses,
attending to the wedge also suppressed slow endogenous fluc-
tuations that were unrelated to the visual stimulus in topograph-
ically defined areas in early visual cortex, posterior parietal
cortex, and lateral occipital cortex, but not in a nonvisual cortical
control region. Moreover, attentional enhancement of response
amplitude and suppression of endogenous fluctuations were
dissociable across cortical areas and across time. Finally, we
found that the amplitude of the stimulus-evoked response was
not correlated with a perceptual measure of visual target detec-
tion. Instead, perceptual performance was accounted for by the
amount of suppression of slow endogenous fluctuations. Our re-
sults indicate that the amplitude of slow fluctuations of cortical
activity is influenced by spatial attention and suggest that these
endogenous fluctuations may impair perceptual processing in
topographically organized visual cortical areas. ■
INTRODUCTION
Visual processing and perception are enhanced at at-
tended locations (Carrasco, 2011), and in many visual
cortical areas, stimulus-evoked responses of individual
neurons are larger when spatial attention is directed to the
neuron’s receptive field (Maunsell, 2015). In human fMRI
studies, the BOLD (blood oxygenation level-dependent) re-
sponse to a stimulus in visual cortex is greater when atten-
tion is directed to the stimulus location (Gandhi, Heeger,
& Boynton, 1999).
Spatial attention can also improve visual processing by
suppressing brain activity. There is a sustained decrease
in BOLD signal in portions of early visual cortex that rep-
resent unattended visual field locations (Silver, Ress, &
Heeger, 2007; Müller & Kleinschmidt, 2004). Moreover,
electrophysiology studies in animal models have shown
that spatial attention can decrease variability of stimulus-
evoked responses (Mitchell, Sundberg, & Reynolds, 2007)
as well as the shared variability (noise correlations) in
University of California, Berkeley
*A. R. is currently at the University of Washington eScience
Institute.
© 2019 Massachusetts Institute of Technology
populations of neurons (Ruff & Cohen, 2014; Herrero,
Gieselmann, Sanayei, & Thiele, 2013; Cohen & Maunsell,
2009; Mitchell, Sundberg, & Reynolds, 2009).
In a previous study, we used a periodic visual stimulus, a
spatial attention task, and fMRI to study the effects of at-
tention on the reliability of stimulus-evoked responses in
many topographically organized areas in early visual, ven-
tral occipital, lateral occipital, and posterior parietal cortex
(Bressler & Silver, 2010). Participants performed a target
detection task that required them to maintain spatial at-
tention either within a rotating wedge stimulus or at a cen-
tral fixation point. We found that, relative to attending to
the fixation point, continuously directing attention to the
rotating wedge improves the reliability of fMRI responses
evoked by the wedge (Bressler & Silver, 2010).
In this study, we used Fourier decomposition to simul-
taneously and independently estimate both stimulus-
evoked and endogenous fMRI signals in the same time
series. Because the rotating wedge stimulus generated
a periodic response in voxels in cortical areas containing
a topographic map of the visual field, we could distin-
guish this evoked response from ongoing endogenous
fluctuations in the frequency domain (Engel et al.,
Journal of Cognitive Neuroscience 32:1, pp. 85–99
https://doi.org/10.1162/jocn_a_01470
1994). We found that the improved reliability of fMRI re-
sponses to an attended periodic visual stimulus (Bressler
& Silver, 2010) is due to both an enhancement of the
stimulus-evoked response and a reduction in the ampli-
tude of endogenous slow fluctuations that are unrelated
to the stimulus. In addition, the relative contributions of
response enhancement and suppression of endogenous
fluctuations to improved response reliability varied
across topographically organized cortical areas, and the
amplitudes of enhancement and suppression were not
correlated across 5-min fMRI runs.
We also correlated performance on the visual detec-
tion task with the amplitude of both stimulus-evoked
responses and endogenous fluctuations. In multiple cor-
tical areas, larger fMRI responses predict detection of a
threshold-level stimulus (Imamoglu, Heinzle, Imfeld, &
Haynes, 2014; Ress, Backus, & Heeger, 2000). However,
attentional suppression of endogenous fluctuations could
also improve the brain’s representation of the attended
stimulus, thereby facilitating perception of the attended
stimulus. Perceptual performance is affected by intrinsic
neural activity immediately before stimulus presentation
(Busch, Dubois, & VanRullen, 2009; Mathewson, Gratton,
Fabiani, Beck, & Ro, 2009; Supèr, van der Togt, Spekreijse,
& Lamme, 2003), including very slow (<0.1 Hz) fluctua-
tions in activity (Monto, Palva, Voipio, & Palva, 2008). In
addition, ongoing fMRI signals before initiation of visual
perceptual tasks predict behavior (Coste & Kleinschmidt,
2016; Wohlschläger et al., 2016; Hesselmann, Kell, &
Kleinschmidt, 2008; Weissman, Roberts, Visscher, &
Woldorff, 2006).
We found that the magnitude of slow endogenous fluc-
tuations predicted participants’ performance on a visual
target detection task in all 11 of the occipital and parietal
cortical areas that we studied. Specifically, target detec-
tion performance was greatest when the strength of sup-
pression of slow endogenous fluctuations was largest.
Surprisingly, we did not find a relationship between the
amplitude of the stimulus-evoked response and behav-
ioral performance in any cortical area. These results sug-
gest that endogenous fluctuations in neural activity are
modulated by spatial attention and that successful sup-
pression of this endogenous activity facilitates visual
perception.
METHODS
Visual stimuli, task design, fMRI data collection and pre-
processing, and definition of topographically organized
cortical areas have been described (Bressler & Silver,
2010) and will only be summarized here.
fMRI Data Acquisition and Preprocessing
fMRI experiments were conducted for five participants
with a 4T Varian INOVA MR scanner and for the other five
participants with a 3T Siemens MAGNETOM Trio MR
scanner. In the 4T scanner, a transmit/receive surface
radiofrequency coil was used to maximize the contrast-
to-noise ratio in occipital cortex, and in the 3T scanner,
a 12-channel receive-only head coil was used. Functional
echo-planar images were acquired using a gradient-echo
EPI sequence. The field of view was 180 × 180 (4T) or
200 × 200 mm (3T), and the matrix size was 64 × 64
(4T) or 96 × 96 (3T), resulting in an in-plane voxel res-
olution of 2.81 (4T) or 2.08 mm (3T). The repetition time
was 1.067 (4T) or 2.133 sec (3T), and the echo time was
28 (4T) or 26 msec (3T). Twenty (4T) or 22 (3T) slices
were prescribed with an interslice gap of 0.3 mm and a
slice thickness of 3 mm (4T) or 2 mm (3T).
The duration of each run was 281.6 sec, and the first
8.53 sec of the fMRI time series were discarded. Head
movements were corrected offline using a 3-D image
registration algorithm (MCFLIRT; Jenkinson, Bannister,
Brady, & Smith, 2002). For each voxel and fMRI run,
the mean fMRI signal across all time points in the run
(DC component) was subtracted from each time point
in the motion-corrected time series. As we were inter-
ested in studying a wide range of temporal frequencies,
no additional preprocessing steps (such as transforma-
tion to Fisher Z scores, conversion to percent signal
change, high-pass filtering, or temporal detrending) were
applied to the data.
Experimental Design and Statistical Analysis
Participants
In addition to the eight participants described in Bressler
and Silver (2010), two additional participants completed
this study. All 10 participants (7 female, 3 male) com-
pleted one session to acquire high-resolution whole-
brain anatomical MR images and one fMRI scanning
session. Experiments were undertaken with the under-
standing and written consent of each participant, and
all experimental procedures were approved by the
Committee for the Protection of Human Subjects at the
University of California, Berkeley.
Visual Stimuli and Task
A checkerboard wedge stimulus rotating about a central
fixation point (Sereno et al., 1995; Engel et al., 1994) was
continuously presented during acquisition of each fMRI
time series (Figure 1).
The wedge subtended 45° and extended from 0.5° (in-
ner radius) to 10.9° (outer radius) of visual angle. Each
check within the wedge reversed contrast at a rate of
7.5 Hz, and the stimulus contrast was 100%. The wedge
was presented for 2.13 sec in each location, and the sub-
sequent wedge location was displaced 22.5° in a clock-
wise direction. Therefore, there were a total of 16
wedge positions, and each position overlapped 50% with
the neighboring positions. The wedge completed a
86
Journal of Cognitive Neuroscience
Volume 32, Number 1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
0
5
M
a
y
2
0
2
1
independent and were based on a 50% probability of pre-
sentation for each wedge position). The attention-to-
wedge and attention-to-fixation runs always occurred in
pairs, with one member of the pair immediately following
the other, and any changes to the target sizes were ap-
plied to both runs in the pair. Thus, the only difference
between the two conditions was that the participants re-
sponded to wedge targets in the attention-to-wedge task
and to fixation targets in the attention-to-fixation task.
Eye movements were not recorded during the fMRI ex-
periments. However, all participants were highly trained
in maintaining fixation through participation in numer-
ous prior psychophysical experiments. Significant de-
viations from fixation would have reduced our ability to
retinotopically define cortical areas, but we defined many
areas in each participant in a single scanning session
(exact numbers of defined areas per participant are pre-
sented below).
Before fMRI data collection, each participant practiced
the two target detection tasks for a total of 4 hr in a be-
havioral testing room, allowing behavioral performance
to reach asymptotic levels. In addition, behavioral data
from the practice sessions were used to determine the
target sizes for each participant that resulted in equiva-
lent performance of the two tasks in the fMRI experi-
ment. During behavioral practice sessions, the size of
the fixation target was adjusted to insure that the task dif-
ficulty (percentage of fixation targets correctly detected)
was equal to that of each of the three eccentricity bands
in the attention-to-wedge task. Similar to the eight partic-
ipants described in Bressler and Silver (2010), there was
no significant difference in performance between the
attention-to-wedge and attention-to-fixation tasks for
either of the additional two participants reported in this
study (Participant 9: p = .09, n = 5 pairs of runs;
Participant 10: p = .37, n = 6 pairs of runs; two-tailed
t tests; see Table 1).
Definition of Topographic Cortical Areas and ROIs
The boundaries of visual cortical areas V1, V2, V3, V3A/B,
V4, LO1, LO2, and VO1 and posterior parietal areas IPS0,
IPS1, and IPS2 were defined using well-established phase-
encoded retinotopic mapping methods (Silver, Ress, &
Heeger, 2005; Sereno et al., 1995; Engel et al., 1994).
Table 1. Behavioral Performance
Participant No. Wedge
Fixation
Near Middle
Far
9
10
76
71
64
63
74
67
75
69
81
77
Values are percentage of targets correctly detected for all trials of the
attention-to-wedge task, the attention-to-fixation task, and for each of
the three eccentricity bands in the attention-to-wedge task for the
two participants not included in Bressler and Silver (2010).
Bressler, Rokem, and Silver
87
Figure 1. Stimulus and tasks. Participants viewed a wedge-shaped
stimulus that rotated around the central fixation point while
continuously maintaining fixation, and they pressed a button whenever
they detected a contrast decrement target. In the attention-to-wedge
condition, participants detected targets presented at random locations
within the wedge, and in the attention-to-fixation condition,
participants detected targets within the fixation point.
full rotation once every 34.13 sec. Participants were in-
structed to continuously maintain fixation on a central
fixation point (0.25° of visual angle) throughout each
scan.
In the attention-to-wedge task, participants were in-
structed to maintain fixation on the central point and to
press a button whenever they detected a brief (0.27 sec)
presentation of a square region of zero contrast within
the wedge (Figure 1). There was a 50% probability of target
presentation at each wedge position, and the target could
appear anywhere within the wedge stimulus at unpredict-
able times. Uncertainty regarding the time and location
of target presentation encouraged participants to continu-
ously allocate their attention over the entire wedge. In ad-
dition, to avoid the possibility that differences in difficulty
of target detection as a function of eccentricity could affect
the allocation of spatial attention over the wedge, the
target sizes in three eccentricity bands (0.5°–4.0°, 4.0°–
7.4°, and 7.4°–10.9° of visual angle) were adjusted for each
participant to equate the percentage of targets correctly
detected in each of these bands. The boundaries be-
tween these eccentricity bands were not visible to the
participants.
In the attention-to-fixation task, participants were in-
structed to maintain fixation on the central square and
to press a button when they detected a square region
of zero contrast within the fixation square. The targets
in the attention-to-fixation and attention-to-wedge tasks
had identical durations, contrasts, and probabilities of
presentation. To equate sensory stimulation in the two
tasks, the square zero-contrast regions were presented
in both the fixation point and the wedge for both atten-
tion conditions (although the temporal sequences of pre-
sentation within the fixation point and the wedge were
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
/
.
o
n
0
5
M
a
y
2
0
2
1
Each participant completed between five and seven attention-
to-wedge runs and an equal number of attention-to-
fixation runs; the order of runs was interleaved for the
two attention conditions. The time series from each voxel
were converted to units of percent signal change and av-
eraged across all runs (including both attention-to-wedge
and attention-to-fixation). Note that we convert to per-
cent signal change and average across runs only for the
purpose of defining the boundaries of topographically
organized areas; in our main time-series analysis (see
below), there was no conversion to percent signal change.
The duration of the stimulus cycle was 34.13 sec,
resulting in a modulation of fMRI signals in visually
responsive voxels of 1/34.13 sec = 0.0293 Hz. Any voxels
that responded to the visual stimulus in a spatially spe-
cific manner therefore exhibited modulations of activity
at this stimulus frequency. We computed the discrete
Fourier transform for each time series from each voxel,
and response phases at the stimulus frequency (relative
to the cycle of the rotating wedge) were spatially trans-
formed and overlaid on computationally flattened cortical
patches. The phase of the Fourier-transformed time
series at the frequency band containing the stimulus
frequency corresponds to the angular component (in po-
lar coordinates) of the visual field location that is repre-
sented by a given voxel. The spatial distribution of these
response phases on the cortical surface was then used to
define the locations and boundaries of topographically
organized cortical areas.
Using these procedures, the boundaries of V1, V2, V3,
V4, IPS1, and IPS2 were successfully defined in both
hemispheres of all participants (total of 20 hemispheres).
Of the 20 hemispheres in our sample, V3A/B, IPS0, and
LO1 (Larsson & Heeger, 2006) were defined in 19 of
these, LO2 (Larsson & Heeger, 2006) and VO1 (Brewer,
Liu, Wade, & Wandell, 2005) were defined in 18 hemi-
spheres, and each topographic area was defined in at
least one hemisphere for each participant. The locations
and boundaries of topographically organized areas in an
example hemisphere from a participant not reported in
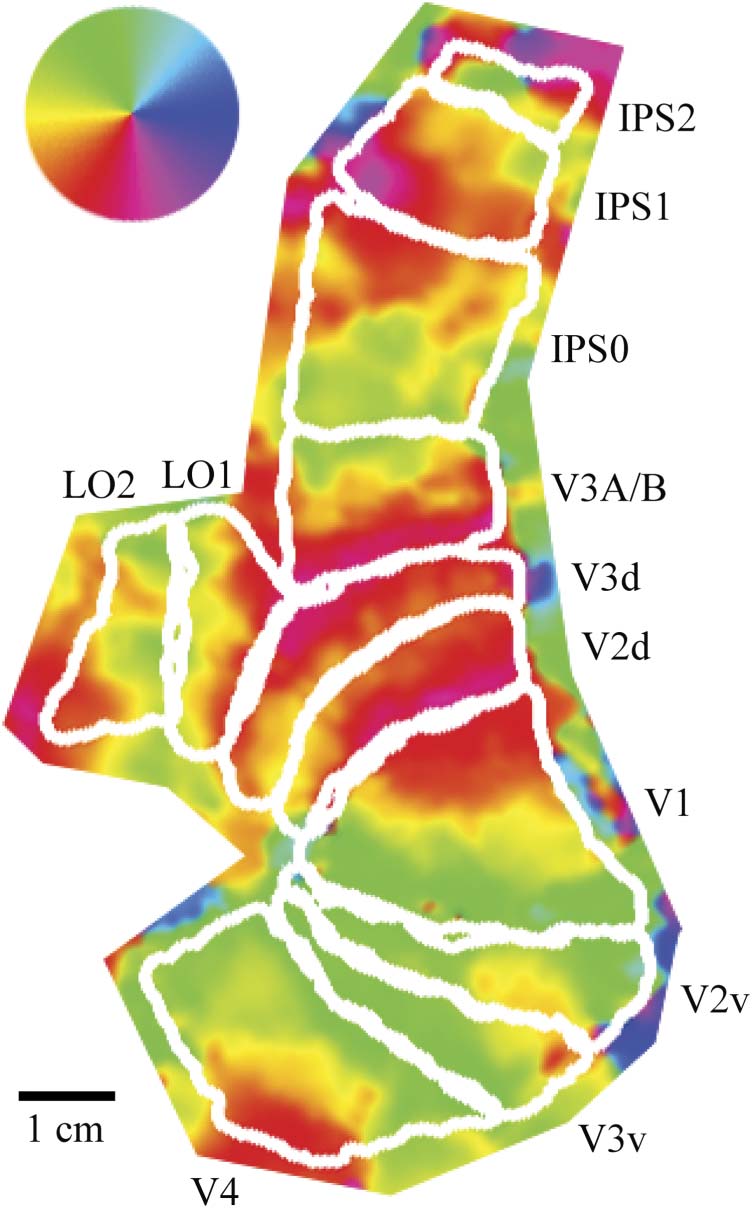
Bressler and Silver (2010) is shown in Figure 2.
In addition to these topographic areas, we also defined
a bilateral ROI centered in the posterior cingulate cortex
(PCC)/precuneus (Talairach coordinates [−2, −51, 27],
[2, −51, 27]; Greicius, Krasnow, Reiss, & Menon, 2003).
For each participant, the PCC/precuneus ROI was ex-
panded isotropically from the Talairach-defined center
within the cortical gray matter ( Wandell, Chial, &
Backus, 2000) until its volume was equal to the average
volume of the defined topographic cortical areas for that
participant.
Time-series Analysis
We applied Fourier decomposition to the time series
for each run to compute amplitude at the stimulus fre-
quency and nonstimulus frequencies. The strength of
Figure 2. Locations and boundaries of topographically organized
areas in occipital and parietal cortex. The color wheel indicates the
relationship between response phase at the frequency of wedge
rotation and the angular component of visual field location. In this right
hemisphere example, each area represents the contralateral left visual
field.
fluctuation at the stimulus frequency was defined as the
amplitude at the frequency band centered on 0.0293 Hz,
corresponding to the rate of wedge rotation. We defined
the strength of fluctuation at nonstimulus frequencies as
the average of amplitudes of frequency components with
ranges of 0.0073–0.0220 Hz, 0.0366–0.0513 Hz, 0.0659–
0.0806 Hz, and 0.0952–0.0989 Hz (i.e., all frequency
components below 0.1 Hz, not including 0 Hz (DC),
the stimulus frequency, harmonics of the stimulus fre-
quency, and frequency bands immediately adjacent to
these components).
For each voxel, attentional modulation of the ampli-
tudes at stimulus and nonstimulus frequencies was ex-
pressed as a contrast index: (w − f )/(w + f ), where w
is attention-to-wedge and f is attention-to-fixation. This
contrast index is similar to the percent change relative
to the attention-to-fixation baseline, but unlike percent
change values, contrast indices are symmetric for in-
creases and decreases. Contrast indices were computed
for every pair of attention-to-wedge and attention-to-
fixation runs, generating unbiased estimates of atten-
tional modulation for every voxel. These contrast index
values were averaged across all voxels within each cortical
area to obtain a total of 60 attentional modulation values
88
Journal of Cognitive Neuroscience
Volume 32, Number 1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
for the group of 10 participants. For each cortical area,
the mean of these 60 contrast index values was compared
with a value of zero (indicating no effect of spatial
attention) with a two-tailed t test. All statistical tests that
were applied to each of the 12 cortical areas were
corrected for multiple comparisons using the false dis-
covery rate (FDR) method (Genovese, Lazar, & Nichols,
2002).
In addition, we estimated the relationship between
amplitude at the stimulus frequency and that of nonsti-
mulus frequencies across attention-to-wedge runs. Each
5-min run generated an amplitude value at the stimulus
frequency and at nonstimulus frequencies in each voxel,
and the correlation of these two amplitudes was com-
puted across runs. These correlation coefficients were
then converted to normally distributed z scores by
Fisher transformation and then averaged across all voxels
within a given cortical area for each participant. For each
area, we assessed whether the mean correlation coeffi-
cient across the 10 participants was significantly different
from zero using a two-tailed t test.
We also conducted this correlation analysis across runs
using a normalized estimate of stimulus-evoked activity.
This normalization was applied to minimize the contribu-
tion of endogenous activity to the estimate of stimulus-
evoked responses. Endogenous fluctuations occurred at
all temporal frequencies we studied, so the amplitude at
the stimulus frequency includes both stimulus-evoked ac-
tivity and endogenous activity. We therefore subtracted
the mean of the amplitude at nonstimulus frequency
bands immediately adjacent to the stimulus frequency
(0.0256 and 0.0330 Hz, providing an estimate of the
amount of activity that is in the stimulus frequency band
but unrelated to the stimulus) from the amplitude at the
stimulus frequency band (0.0293 Hz) for each voxel. This
normalized stimulus frequency amplitude was then cor-
related with nonstimulus frequency amplitudes across
runs as described above.
This normalization procedure was not applied to the
calculation of the contrast index values described above
([wedge − fix]/[wedge + fix]; see Figures 3–5). We
found that subtracting the mean of the amplitude at non-
stimulus frequency bands immediately adjacent to the
stimulus frequency from the amplitude at the stimulus
frequency band sometimes generated very small differ-
ence values, resulting in small values of the denominator
of the contrast index and variable and unstable contrast
index values. The decision to not apply normalization to
contrast index values means that some portion of the am-
plitude at the stimulus frequency is due to endogenous
fluctuations in that frequency band. However, this would
only reduce differences between stimulus and nonstimu-
lus frequencies with respect to attentional modulation,
and our results clearly demonstrate opposing effects of
spatial attention on amplitudes at stimulus and nonstimu-
lus frequencies in the majority of studied topographically
organized areas (Figure 3).
Correlation with Perceptual Performance
For each attention-to-wedge run, behavioral performance
was correlated with amplitude at both stimulus and
nonstimulus frequencies. Although target size was occa-
sionally adjusted during the fMRI session to maintain
equivalent difficulty in the attention-to-fixation and
attention-to-wedge tasks, 9 of 10 participants had at least
five attention-to-wedge runs with the same target size
distribution (with the remaining participant having four
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
t
f
/
o
n
0
5
M
a
y
2
0
2
1
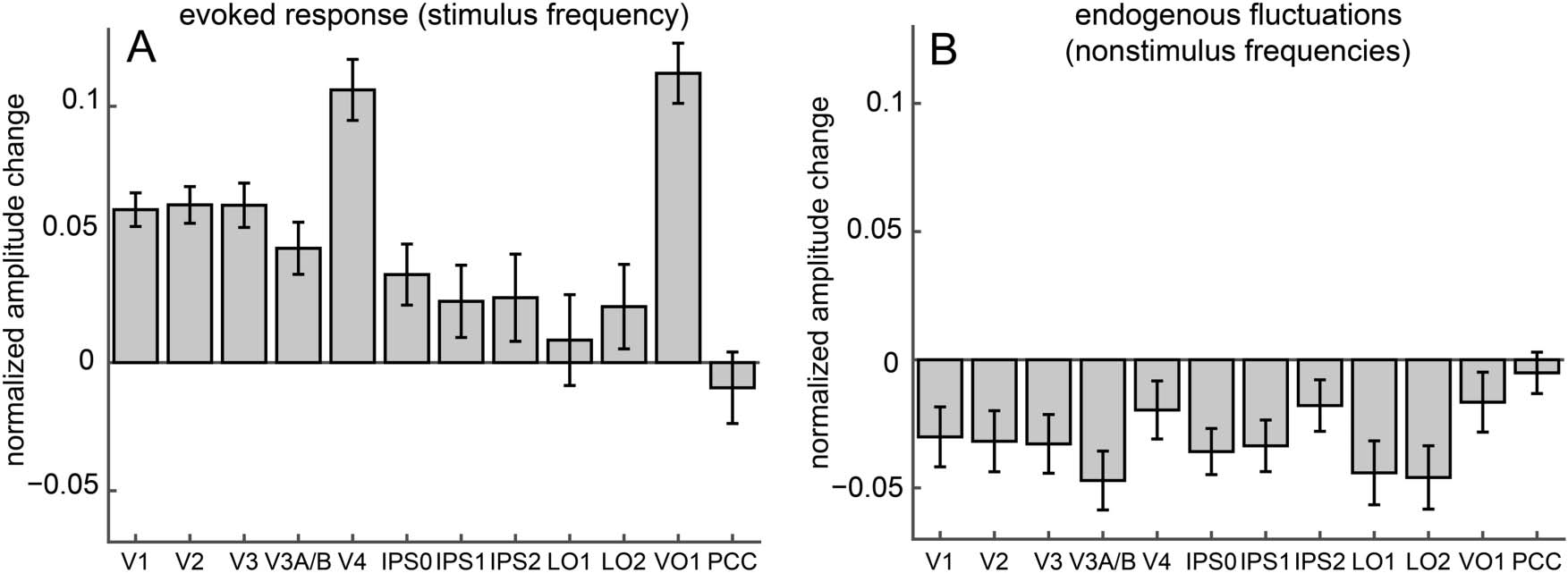
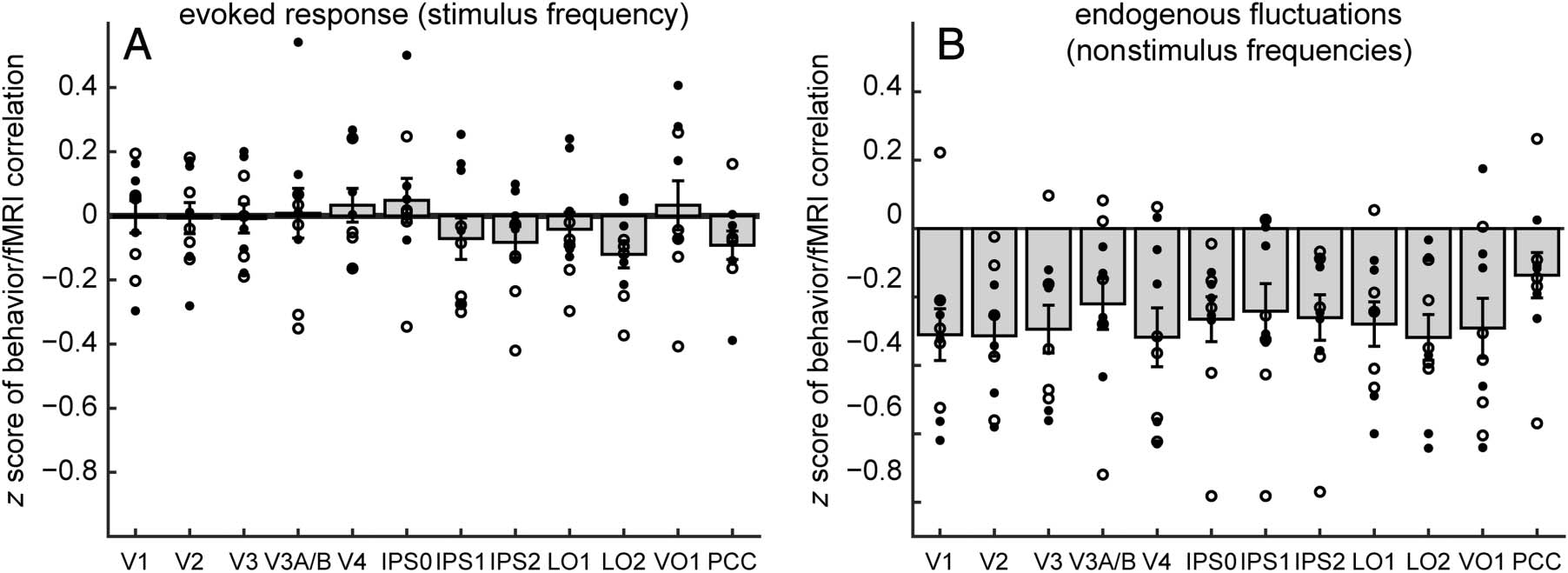
Figure 3. Attentional modulation of amplitude of the evoked response and fluctuations at nonstimulus frequencies. Effects of attention were
quantified as a contrast index of fMRI signal amplitudes for the two attention conditions: (wedge − fixation)/(wedge + fixation). (A) Attending
to the rotating wedge significantly increased the amplitude of the visual response (stimulus frequency, 0.0293 Hz) in early and ventral visual cortical
areas, but not in IPS1, IPS2, LO1, or LO2. (B) In contrast, attending to the wedge decreased the strength of endogenous nonstimulus fluctuations
(0.0073–0.1 Hz, excluding the stimulus frequency and its harmonics) in every measured topographically organized area except IPS2, V4, and
VO1. These attention effects were not observed in a PCC control region. Error bars represent SEMs across pairs of attention-to-wedge and
attention-to-fixation runs.
Bressler, Rokem, and Silver
89
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
f
t
o
n
0
5
M
a
y
2
0
2
1
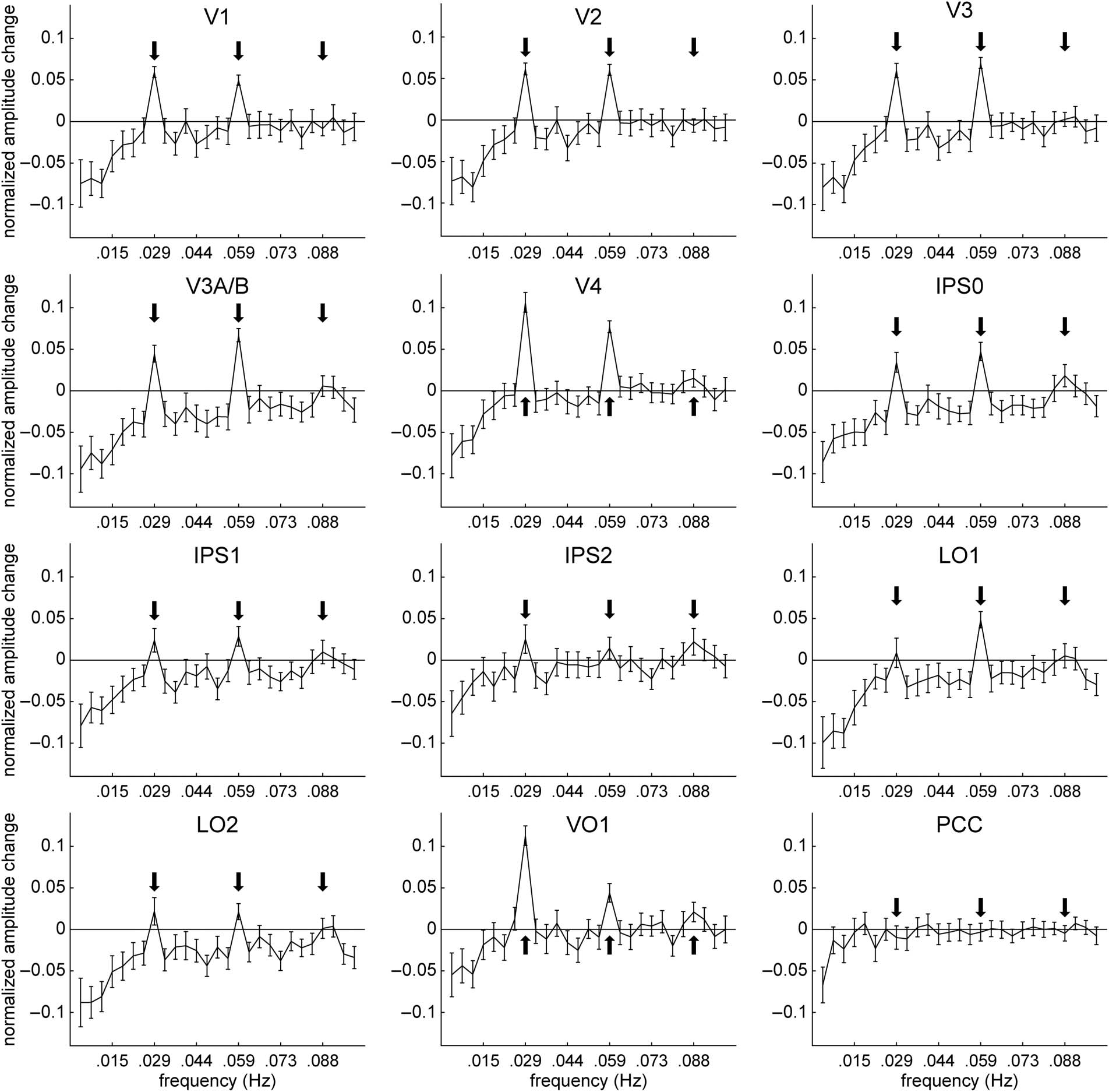
Figure 4. Attentional modulation of amplitude of fluctuations across the frequency spectrum. As in Figure 3, effects of attention were quantified as a
contrast index of fMRI signal amplitudes for the two attention conditions: (wedge − fixation)/(wedge + fixation). In all topographically organized
cortical areas, attending to the wedge stimulus caused suppression of nonstimulus frequency fluctuations that was greatest for the lowest frequencies
(approximately less than 0.03 Hz), whereas only a subset of areas showed a significant reduction in higher nonstimulus frequencies. Stimulus
frequency (0.029 Hz) and associated harmonics (0.059 and 0.088 Hz) are indicated with arrows. Error bars represent SEMs across pairs of
attention-to-wedge and attention-to-fixation runs.
attention-to-wedge runs with the same target size distri-
bution). Only sets of runs for which target size distribu-
tion did not change were included in the correlation
analysis, and this required excluding one run from the
data for four participants.
For each participant, we tested whether behavioral per-
formance across attention-to-wedge runs was significantly
different from a normal distribution using Kolmogorov–
Smirnov tests, and we found that no participant showed
significant deviations from normality (all ps > .59).
There was also no significant deviation from normality
for the distribution of behavioral performance for data
pooled across all participant ( p = .97). Correlation coef-
ficients were converted to normally distributed z scores
by Fisher transformation and were averaged across
all voxels within a given cortical area for each partic-
ipant. The mean of the z scores of the correlation co-
efficients across the 10 participants was then compared
with a value of zero for each cortical area using a two-
tailed t test.
90
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 1
response to the stimulus, and the frequency of this mod-
ulation was equal to the frequency of wedge rotation
(0.0293 Hz). A Fourier transform was computed for each
voxel’s time series to quantify the amplitude of the evoked
response at the stimulus frequency (0.0293 Hz) and at
nonstimulus frequencies between 0.0037 Und 0.1 Hz. Wir
measured attentional modulation of both of these quantities
and found that, relative to attending to fixation, attending to
the wedge significantly increased the amplitude of the
stimulus-evoked response in cortical areas V1 (t = 9.10,
p = 4.6 × 10−12), V2 (t = 8.59, p = 1.7 × 10−11), V3
(t = 7.10, p = 4.5 × 10−9), V3A/B (t = 4.39, p = 9.5 ×
10−5), V4 (t = 8.94, p = 5.9 × 10−12), IPS0 (t = 2.88,
p = 9.4 × 10−3), and VO1 (t = 9.61, p = 1.3 × 10−12),
but not in posterior parietal (IPS1: t = 1.70, p = .14; IPS2:
t = 1.49, p = .19) or lateral occipital (LO1: t = 0.49, p =
.62; LO2: t = 1.32, p = .23) Kortex (Abbildung 3A; two-tailed
t tests, FDR-corrected, df = 59).
Attending to the wedge also significantly decreased the
amplitude of slow endogenous fluctuations at nonstimu-
lus frequencies in all topographically organized cortical
areas that we tested ( V1: t = −2.57, p = .019; V2: t =
−2.67, p = .017; V3: t = −2.87, p = .012; V3A/B: t =
−4.11, p = 1.1 × 10−3; IPS0: t = −3.98, p = 1.1 × 10−3;
IPS1: t = −3.33, p = 3.6 × 10−3; LO1: t = −3.54, p = 2.4 ×
10−3; LO2: t = −3.71, p = 1.8 × 10−3), except V4, IPS2, Und
VO1 (V4: t = −1.73, p = .11; IPS2: t = −1.78, p = .11; VO1:
t = −1.42, p = .18; Abbildung 3B; two-tailed t tests, FDR-
corrected, df = 59).
Examination of power spectra of attentional modula-
tion in individual cortical areas (Figur 4) indicates that
attentional suppression was strongest for the lowest fre-
quencies (etwa <0.03 Hz), although this sup-
pression extended to higher frequencies for some
cortical areas (e.g., V3A/B, IPS0, LO1, and LO2).
Relative Contributions of Attentional Enhancement
and Suppression to Improved Response Reliability
Vary across Topographic Cortical Areas and
across Time
We previously showed that directing attention to the ro-
tating wedge increased the reliability of the fMRI re-
sponse to the wedge in many topographically organized
areas in occipital and parietal cortex (Bressler & Silver,
2010). This improved response reliability could be due
to increased amplitude of the stimulus-evoked response
and/or suppression of endogenous activity unrelated to
the representation of the stimulus. To determine the rel-
ative contributions of attentional enhancement and sup-
pression to improved response reliability, we correlated
the amount of attentional enhancement at the stimulus
frequency to the amount of attentional suppression at
nonstimulus frequencies across topographic cortical
areas. This analysis revealed that lateral occipital and pos-
terior parietal areas generally showed the strongest sup-
pression and weakest enhancement by spatial attention,
Bressler, Rokem, and Silver
91
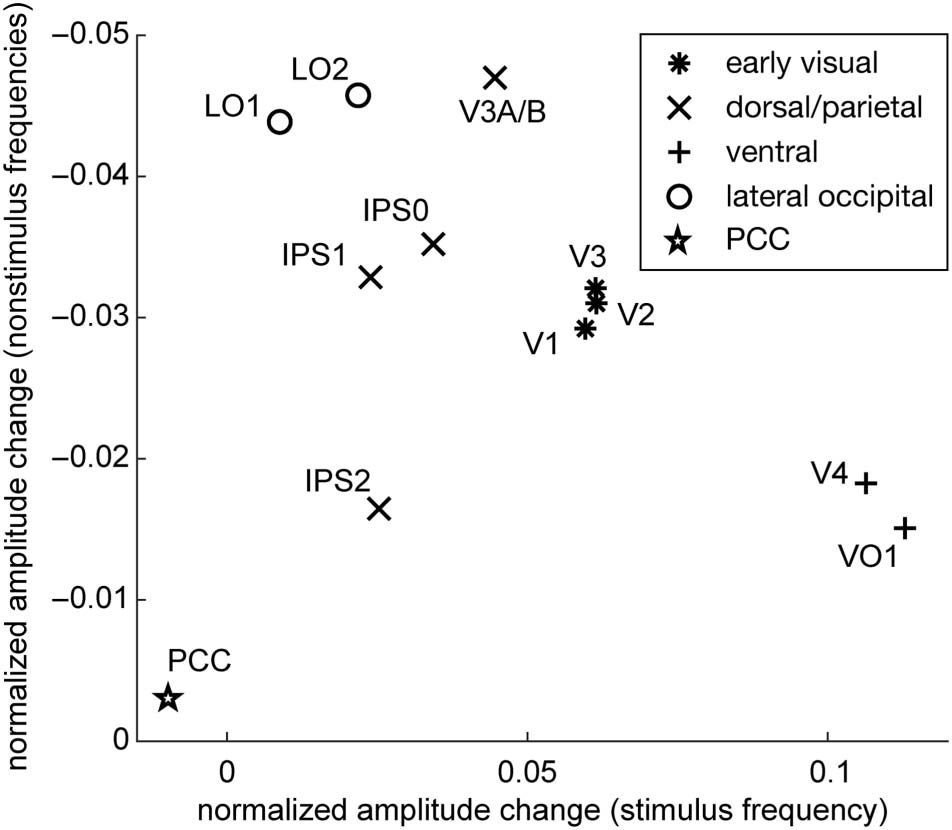
Figure 5. Correlation between attentional modulation of amplitude of
stimulus and nonstimulus frequencies across topographic areas. Ventral
visual (+) areas showed the strongest enhancement of visual responses
but the weakest suppression at nonstimulus frequencies, lateral
occipital (o) and posterior parietal (X) areas generally showed the
opposite pattern of results, and early visual cortical areas (*) exhibited
moderate levels of both enhancement of the visual response and
suppression at nonstimulus frequencies.
Availability of Data and Analysis Code
All the data presented here and associated analysis code are
available at https://osf.io/vznur/.
RESULTS
Participants maintained central fixation while viewing a
wedge-shaped visual stimulus that rotated around the
screen once every 34.13 sec (Figure 1). On separate runs,
attention was either maintained at the central fixation point
or directed toward the rotating wedge stimulus. During
attention-to-fixation runs, participants detected targets that
were presented within the fixation point, and in the
attention-to-wedge runs, they detected targets that were
presented at random locations within the rotating wedge.
Participants pressed a button whenever they detected a
low-contrast target in the attended region. The difficulty
of the task was controlled by adjusting target size for each
participant so that they detected approximately 70% of the
targets in each attention condition. fMRI responses were
recorded from topographically organized cortical areas
V1, V2, V3, V3A/B, V4, IPS0, IPS1, IPS2, LO1, LO2, and
VO1 as well as a control region in PCC that does not con-
tain a topographic map of the visual field.
Effects of Spatial Attention on Strength of
Fluctuations in fMRI Signals at Stimulus and
Nonstimulus Frequencies
The rotating wedge evoked periodic modulation of activ-
ity in any brain location that exhibited a spatially specific
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
f
t
/
o
n
0
5
M
a
y
2
0
2
1
ventral visual areas had the strongest enhancement and
weakest suppression, and early visual areas exhibited in-
termediate levels of both enhancement and suppression
(Figure 5). In addition, across cortical areas, there is an
inverse relationship between the amount of signal en-
hancement and suppression of endogenous frequencies
by spatial attention (r = −.65; p = .029; Figure 5).
Given that attending to the wedge generally increased
the amplitude at the stimulus frequency but decreased
the amplitude at nonstimulus frequencies, we also de-
termined whether periods of high stimulus-evoked
responses were associated with lower amplitudes of non-
stimulus frequencies within a given area across runs. For
each voxel, we correlated the amplitude of the evoked
response at the stimulus frequency with the amplitude
of endogenous fluctuations at nonstimulus frequencies
across the 5-min attention-to-wedge runs. These correla-
tion coefficients were significantly greater than zero in all
studied topographic areas ( V1: t = 8.56, p =6 .2 × 10−5;
V2: t = 9.56, p = 6.2 × 10−5; V3: t = 8.36, p = 6.2 × 10−5;
V3A/B: t = 4.42, p = 1.8 × 10−3; V4: t = 4.30, p = 2.0 × 10−3;
IPS0: t = 4.47, p = 1.8 × 10−3; IPS1: t = 7.51, p = 8.8 × 10−5;
IPS2: t = 5.88, p = 4.0 × 10−4; LO1: t = 7.78, p = 8.3 × 10−5;
LO2: t = 7.01, p = 1.3 × 10−4; VO1: t = 4.89, p = 1.1 × 10−3;
two-tailed t tests, FDR-corrected, df = 9). However, some
of this large positive correlation between amplitudes at
stimulus and nonstimulus frequencies across runs is due
to the fact that endogenous fluctuations of fMRI signal
occur simultaneously at many frequencies (Zarahn,
Aguirre, & D’Esposito, 1997), including the stimulus fre-
quency in our data. That is, amplitude at the stimulus fre-
quency represents a combination of stimulus-evoked
activity and endogenous fluctuations that are in the same
frequency band as the wedge rotation. In addition, very
slow fluctuations or drift in overall fMRI power across
multiple 5-min runs will lead to positive correlations of
amplitude in stimulus and nonstimulus frequency bands.
Thus, to more directly test for correlations between
enhancement of response amplitude and suppression
of endogenous frequencies across runs, we subtracted
the mean of the amplitude in nonstimulus frequency
bands on either side of the stimulus frequency from
the amplitude at the stimulus frequency on each run.
This generated a normalized amplitude at the stimulus
frequency that more accurately reflects the magnitude
of the stimulus-evoked response while minimizing contri-
bution from endogenous fluctuations. We then com-
puted correlation coefficients between this normalized
amplitude at the stimulus frequency and the mean ampli-
tudes of the nonstimulus frequencies across runs. None
of the topographic cortical areas exhibited a significant
correlation across runs ( V1: t = −1.16, p = .66; V2:
t = −2.07, p = .64; V3: t = −0.84, p = .84; V3A/B: t =
−0.13, p = .90; V4: t = −0.15, p = .90; IPS0: t = −0.26,
p = .90; IPS1: t = 1.19, p = .66; IPS2: t = 0.29, p = .90;
LO1: t = −0.47, p = .90; LO2: t = 0.19, p = .90; VO1: t =
1.28, p = .66; Figure 6; two-tailed t tests, FDR-corrected,
df = 9), indicating that, for a given cortical area, the am-
plitude of stimulus-evoked responses does not predict
the strength of endogenous fluctuations in that area over
the course of a 5-min fMRI run.
Our full data set includes experiments conducted at
both a 3T Siemens scanner and a 4T Varian scanner.
We therefore separately analyzed data collected at the
two scanners. This analysis shows that the correlation
between amplitudes at the stimulus and nonstimulus fre-
quencies across runs was significantly greater for partici-
pants in the 3T scanner in all topographically organized
areas except IPS0, IPS1, LO1, and VO1 (Figure 6; all sig-
nificant p values = .04, two-tailed t tests, FDR-corrected,
df = 8). However, neither the 3T nor the 4T participant
group exhibited values for the correlation between ampli-
tudes at stimulus and nonstimulus frequencies that were
significantly different from zero in any topographic area
(all ps >.05; two-tailed t tests; FDR-corrected, df = 4).
Visual Target Detection Is Negatively Correlated
with Amplitude of Endogenous Fluctuations but
not with Amplitude of the Stimulus-Evoked
Response
Our results demonstrate that both attentional enhance-
ment of response amplitude and suppression of endoge-
nous fluctuations contributed to improved response
reliability in topographic cortical areas. To determine
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
1
e
8
–
5
P
D
1
9
F
3
/
1
3
1
2
4
/
4
1
/
Ö
8
C
5
N
/
_
A
1
_
8
0
6
1
1
4
4
7
0
0
0
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
4
N
7
0
0
8
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
F
T
/
Ö
N
0
5
M
A
j
2
0
2
1
Figur 6. Amplitude of stimulus-evoked responses does not correlate
with amplitude of slow endogenous fluctuations across 5-min fMRI
runs. The amplitude at the stimulus frequency was normalized by
subtracting the amplitude of nonstimulus frequency bands on either
side of the stimulus frequency band to estimate the strength of the
stimulus-evoked response on each attention-to-wedge run. Across runs,
this measure of stimulus-evoked response amplitude was not
significantly correlated with the magnitude of slow endogenous
fluctuations at nonstimulus frequencies in any cortical area. Open
circles: individual participant data from 3T scanner; filled circles:
individual participant data from 4T scanner. Error bars represent SEMs
across participants.
92
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 1
the relative contributions of enhancement and suppres-
sion to visual target detection, we correlated the percent-
age of targets correctly detected in the attention-to-wedge
task with fMRI amplitude at both stimulus and nonstimu-
lus frequencies. Each 5-min run generated a single be-
havioral measure and fMRI measures of the amplitude
of both the stimulus-evoked response and endogenous
fluctuations at nonstimulus frequencies.
Surprisingly, target detection across attention-to-
wedge runs was not correlated with stimulus-evoked re-
sponse amplitude in any cortical area ( V1: t = −0.06, p =
.95; V2: t = −0.15, p = .95; V3: t = −0.18, p = .95;
V3A/B: t = .10, p = .95; V4: t = .63, p = .94; IPS0: t =
0.70, p = .94; IPS1: t = −1.10, p = .90; IPS2: t = −1.69,
p = .50; LO1: t = −0.80, p = .94; LO2: t = −2.86, p =
.23; VO1: t = .43, p = .95; Abbildung 7A; two-tailed t tests,
FDR-corrected, df = 9). This was also true for the corre-
lation between target detection and the normalized am-
plitude at the stimulus frequency ( V1: t = 2.07, p = .32;
V2: t = 1.89, p = .32; V3: t = 1.52, p = .33; V3A/B: t =
0.70, p = .67; V4: t = 1.96, p = .32; IPS0: t = 1.66, p =
.32; IPS1: t = −0.37, p = .86; IPS2: t = −0.05, p = .96;
LO1: t = 1.00, p = .51; LO2: t = −0.13, p = .96; VO1: t =
1.66, p = .32).
Although stronger responses to an attended visual
stimulus were not associated with enhanced target detec-
tion, we found that this measure of performance across
attention-to-wedge runs was significantly negatively cor-
related with the amplitude of nonstimulus frequencies
in all studied topographic areas ( V1: t = −4.10, p =
5.9 × 10−3; V2: t = −5.26, p = 5.9 × 10−3; V3: t =
−4.22, p = 5.9 × 10−3; V3A/B: t = −2.97, p = .017;
V4: t = −3.69, p = 7.5 × 10−3; IPS0: t = −4.04, p =
5.9 × 10−3; IPS1: t = −2.99, p = .017; IPS2: t =
−3.90, p = 6.2 × 10−3; LO1: t = −4.30, p = 5.9 × 10−3;
LO2: t = −4.77, p = 5.9 × 10−3; VO1: t = −3.33,
p = .012; two-tailed t tests, FDR-corrected, df = 9).
daher, suppression of slow endogenous fluctua-
tions that were unrelated to the stimulus significantly
predicted target detection, as measured behaviorally
(Figure 7B).
Correlations of target detection with amplitude at stim-
ulus and nonstimulus frequencies were also computed
separately for participants in the 3T and 4T scanners.
There were no differences between these two participant
groups for either correlation in any of the topographic
cortical areas (Figur 7; all ps >.05, two-tailed t tests,
FDR-corrected, df = 8).
We repeated these brain–behavior correlation analyses
but used RTs instead of percentage of targets correctly
detected. RTs were not significantly correlated with ei-
ther the amplitude at the stimulus frequency or at non-
stimulus frequencies across 5-min runs in any of the
topographic cortical areas we studied (all ps >. 05, zwei-
tailed t tests, FDR-corrected; df = 9). Jedoch, partici-
pants were not under time pressure to respond in the
target detection task in our study, and it is possible that
correlations between RTs and the fMRI measures we de-
scribe in this study would have been evident if the task
had emphasized speed of response.
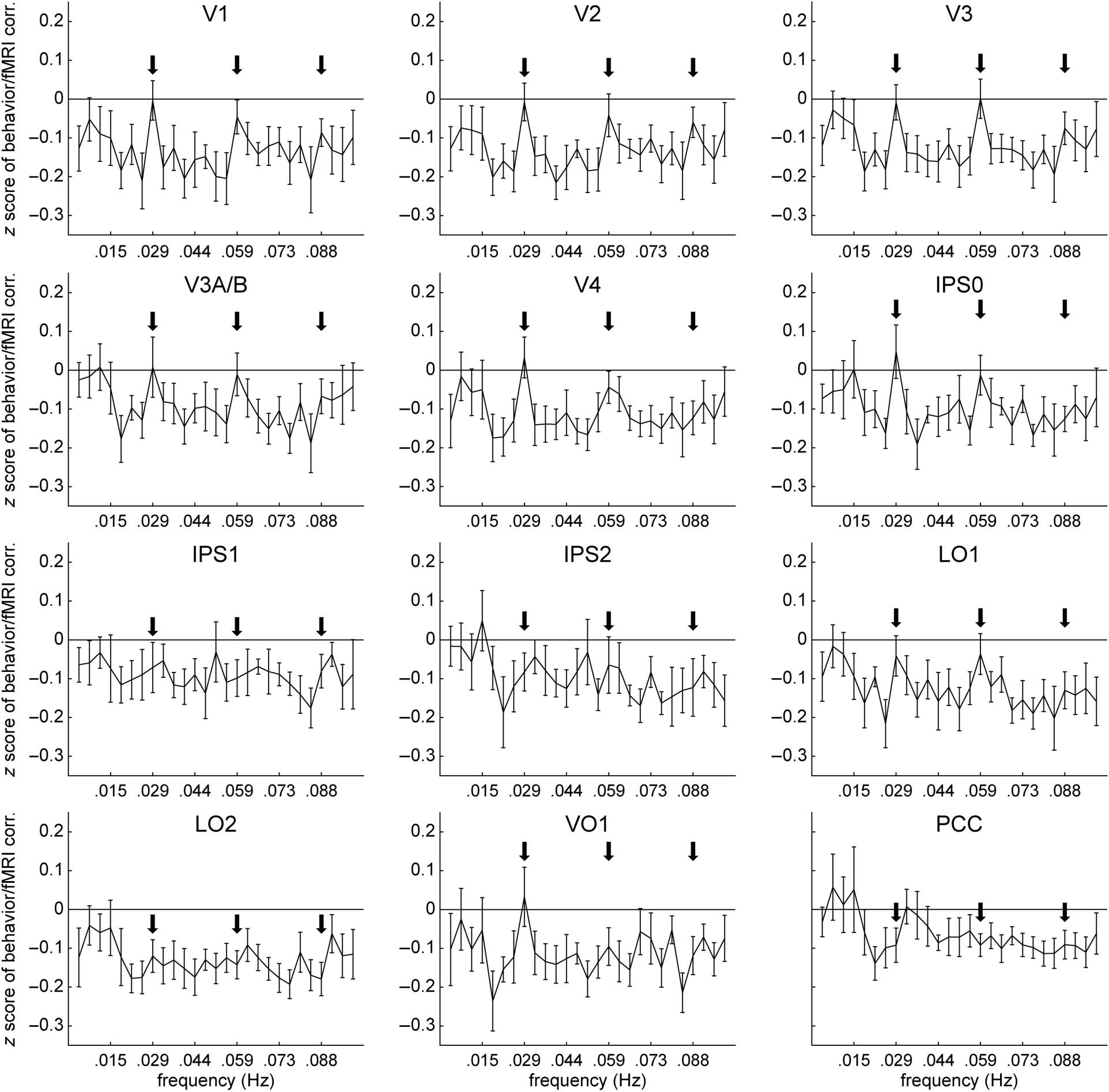
The frequency distribution of the correlation between
percentage of targets correctly detected and suppression
of slow endogenous fluctuations differed from the fre-
quency distribution of attentional suppression itself.
Although suppression of endogenous frequencies was
most pronounced at very low frequencies (zwischen
0.0037 Hz, the lowest frequency we could measure, Zu
etwa 0.03 Hz in most areas), the inverse corre-
lation between target detection performance and ampli-
tude of endogenous fluctuations was generally strongest
at frequencies above 0.02 Hz and often extended to
0.1 Hz, the highest frequency we analyzed (Figur 8).
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
1
e
8
–
5
P
D
1
9
F
3
/
1
3
1
2
4
/
4
1
/
Ö
8
C
5
N
/
_
A
1
_
8
0
6
1
1
4
4
7
0
0
0
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
4
N
7
0
0
8
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
T
/
.
F
Ö
N
0
5
M
A
j
2
0
2
1
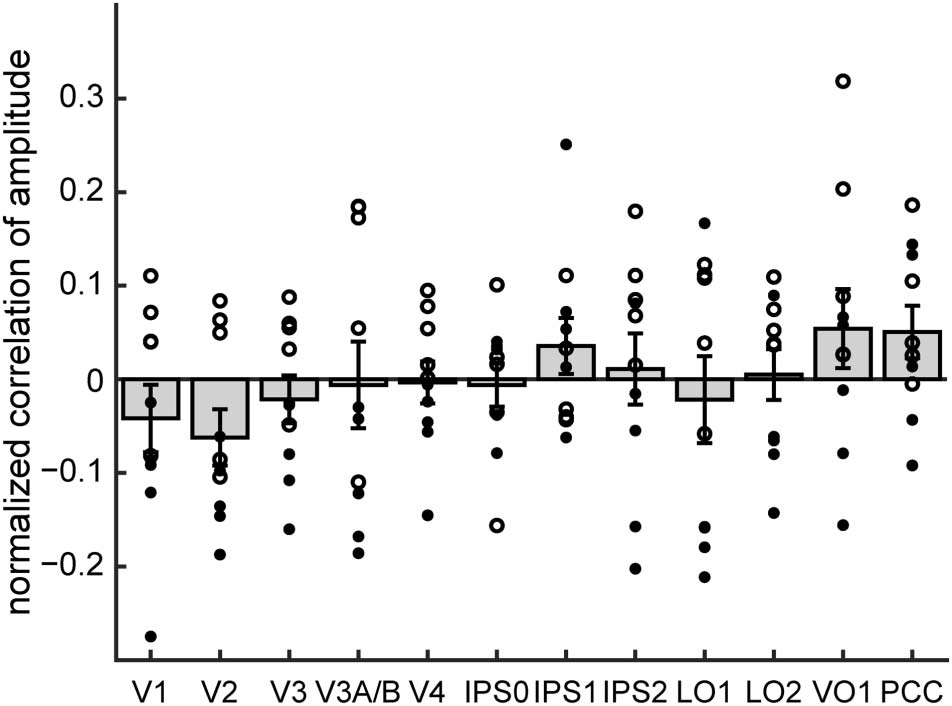
Figur 7. Correlation between performance and fMRI amplitude at stimulus and nonstimulus frequencies. (A) Across 5-min runs, no topographically
organized area showed a significant correlation between amplitude at the stimulus frequency and the percentage of wedge targets that were correctly
detected. (B) Behavioral performance was significantly negatively correlated with amplitude of fMRI fluctuations at nonstimulus frequencies. Open
circles: individual participant data from 3T scanner; filled circles: individual participant data from 4T scanner. Error bars represent SEMs across
Teilnehmer.
Bressler, Rokem, and Silver
93
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
1
e
8
–
5
P
D
1
9
F
3
/
1
3
1
2
4
/
4
1
/
Ö
8
C
5
N
/
_
A
1
_
8
0
6
1
1
4
4
7
0
0
0
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
4
N
7
0
0
8
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
F
T
/
Ö
N
0
5
M
A
j
2
0
2
1
Figur 8. Correlation between behavioral performance and amplitude of fluctuations across the frequency spectrum. In all topographically organized
Bereiche, negative correlations between performance and endogenous fMRI signal fluctuations tended to be weaker at the slowest nonstimulus
frequencies (<0.015 Hz) compared with higher nonstimulus frequencies. Stimulus frequency (0.029 Hz) and associated harmonics (0.059 and
0.088 Hz) are indicated with arrows. Error bars represent SEMs across participants.
Lack of Attentional Effects in a Control
Nontopographic PCC Region
The consistent attentional suppression of fMRI activity
fluctuations at nonstimulus frequencies across many to-
pographically organized cortical areas raises the possibil-
ity of a global effect of allocation of spatial attention to
the wedge, relative to attending to the central fixation
point. Fluctuations in arterial carbon dioxide concentra-
tion ( Wise, Ide, Poulin, & Tracey, 2004), cardiac rate
(Shmueli et al., 2007), and respiratory rate (Birn,
Diamond, Smith, & Bandettini, 2006) occur within the
0.0037–0.1 Hz range that we used to measure fMRI
fluctuations in nonstimulus frequencies, and these non-
neural fluctuations likely influenced our measured
BOLD signals. Additionally, although we matched task
difficulty in the attention-to-fixation and attention-to-
wedge conditions for every participant in our study, it
is still possible that allocating covert spatial attention to
the wedge resulted in a global reduction in the amplitude
of endogenous BOLD fluctuations in nontopographic
brain regions.
To control for this possibility, we anatomically de-
fined an ROI in the PCC/precuneus (Greicius et al.,
2003) of each participant (see the Methods section). The
PCC/precuneus is part of a network of regions that
94
Journal of Cognitive Neuroscience
Volume 32, Number 1
exhibits reduced activity during attention-demanding
cognitive processing (Greicius et al., 2003) and spontane-
ous activity that is negatively correlated with activity in
visual cortical regions (Fransson, 2006; Fox et al., 2005).
Relative to the attention-to-fixation condition, attending
to the wedge did not have a significant effect on either
the amplitude of the stimulus-evoked response (t =
−0.70, p = .53; Figure 3A) or endogenous fluctuations at
nonstimulus frequencies (t = −0.63, p = .53; Figure 3B)
in the PCC/precuneus (two-tailed t tests, FDR-corrected,
df = 59; see also Figures 4 and 5). Moreover, attentional
enhancement of the stimulus-evoked response was signif-
icantly greater in all topographic regions (V1: t = 5.93, p =
3.7 × 10−7; V2: t = 5.98, p = 3.7 × 10−7; V3: t = 6.06, p =
3.7 × 10−7; V3A/B: t = 4.17, p = 1.9 × 10−4; V4: t = 8.22,
p = 2.6 × 10−10; IPS0: t = 3.16, p = 3.9 × 10−3; IPS1: t =
2.25, p = .034; IPS2: t = 2.28, p = .034; VO1: t = 7.83, p =
5.9 × 10−10) except LO1 and LO2 (LO1: t = 1.12, p = .27;
LO2: t = 1.88, p = .071) than in PCC/precuneus (two-tailed
paired t tests, FDR-corrected for 11 statistical tests, df =
59), and attentional suppression at nonstimulus frequen-
cies was significantly greater in all topographic regions
( V1: t = −2.52, p = .020; V2: t = −2.70, p = .014; V3:
t = −3.05, p = 6.2 × 10−3; V3A/B: t = −4.07, p = 1.5 ×
10−3; IPS0: t = −3.81, p = 1.8 × 10−3; IPS1: t = −3.60,
p =1.8 × 10−3; LO1: t = −3.36, p = 3.0 × 10−3; LO2:
t = −3.62, p = 1.8 ×10−3) except V4, IPS2, and VO1
(V4: t = −1.60, p = .13; IPS2: t =−1.88, p = .079; VO1:
t = −1.27, p = .21) than in PCC/precuneus (two-tailed
paired t tests, FDR-corrected, df = 59).
In addition, there was not a significant correlation be-
tween behavioral performance in attend-to-wedge runs
and either response amplitude (t = −2.08, p = .40;
Figure 7A) or amplitude at nonstimulus frequencies
(t = −2.05, p = .070; Figure 7B) in the PCC/precuneus
(two-tailed t tests, FDR-corrected, df = 9; see also
Figure 8). However, direct statistical comparisons of
PCC/precuneus correlation values with those of individ-
ual topographically organized areas revealed no signifi-
cant differences for raw stimulus frequency ( V1: t =
1.17, p = .50; V2: t = 1.18, p = .50; V3: t = 1.20, p =
.50; V3A/B: t = 1.05, p = .51; V4: t = 1.89, p = .50;
IPS0: t = 1.50, p = .50; IPS1: t = 0.23, p = .90; IPS2:
t = 0.13, p = .90; LO1: t = 0.65, p = .73; LO2: t =
−0.40, p = .85; VO1: t = 1.41, p = .50), normalized
stimulus frequency ( V1: t = 2.07, p = .21; V2: t = 1.90,
p = .21; V3: t = 1.65, p = .24; V3A/B: t = 1.03, p = .45;
V4: t = 2.06, p = .21; IPS0: t = 1.94, p = .21; IPS1: t =
0.29, p = .78; IPS2: t = 0.55, p = .67; LO1: t = 1.17, p =
.43; LO2: t = 0.53, p = .67; VO1: t = 1.87, p = .21), or
nonstimulus frequencies ( V1: t = −1.94, p = .28; V2: t =
−2.16, p = .28; V3: t = −1.62, p = .28; V3A/B: t = −0.73,
p = .48; V4: t = −1.85, p = .28; IPS0: t = −1.08, p = .39;
IPS1: t = −0.78, p = .48; IPS2: t = −1.05, p = .39; LO1:
t = −1.47, p = .28; LO2: t = −1.71, p = .28; VO1: t =
−1.45, p = .28; paired two-tailed t tests, FDR-corrected,
df = 9).
DISCUSSION
Allocation of spatial attention to a visual stimulus im-
proves the reliability of the BOLD response to that stim-
ulus (Bressler & Silver, 2010). In this study, we show that
this increase in reliability is partly due to a reduction in
the strength of slow endogenous fluctuations that are un-
related to the visual stimulus. This attentional suppres-
sion of endogenous activity was observed in every
topographic cortical area we studied except IPS2, V4
and VO1. An increase in the amplitude of the stimulus-
evoked response when attending the wedge also contrib-
uted to increased response reliability in early and ventral
visual cortical areas. However, although the magnitude of
the stimulus-evoked response was not predictive of par-
ticipants’ ability to detect targets within the wedge across
5-min runs, suppression of slow endogenous fluctuations
was highly correlated with successful target detection in
all studied topographic areas.
Attentional Enhancement of Stimulus-Evoked
Responses
We found that allocating spatial attention to the rotating
wedge stimulus enhanced the BOLD response evoked by
the wedge. This enhancement was strongest in ventral
areas V4 and VO1; moderate in early visual areas V1,
V2, and V3; small but significant in dorsal areas V3A/B
and IPS0; and not significant in posterior parietal cortical
areas IPS1/2 and lateral occipital areas LO1/2. The lack of
attentional modulation of the stimulus-evoked response
in IPS areas is consistent with that reported in Corbetta,
Kincade, Ollinger, McAvoy, and Shulman (2000) and does
not necessarily contradict previous studies showing
robust attentional modulation of BOLD activity in this re-
gion (Silver et al., 2005; Yantis et al., 2002), as these stud-
ies did not directly compare IPS responses to attended
and unattended visual stimuli.
Additionally, we found no attentional enhancement of
the stimulus-evoked response in LO1/2, consistent with
a previous study showing a similar lack of attentional en-
hancement of LO visual responses for stimuli with
greater than 30% contrast (Murray & He, 2006). However,
we previously reported robust attentional enhancement
of fMRI responses in LO1/2 (Bressler, Fortenbaugh,
Robertson, & Silver, 2013). There are multiple differ-
ences in experimental design and attentional demands
between that study and this study, including the spatial
extent of the attended region (much larger in this study),
continuous attentional tracking of a moving stimulus
versus discrete shifts of attention to an unpredictable
location in Bressler et al. (2013), and the presence of
distractors in Bressler et al. (2013). Further research is
needed to determine the relative contribution of these
various factors to modulation of LO1/2 responses by
spatial attention.
Bressler, Rokem, and Silver
95
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
f
/
t
o
n
0
5
M
a
y
2
0
2
1
Attentional Suppression of Slow
Endogenous Fluctuations
We found attentional suppression of slow (<0.1 Hz) en-
dogenous fluctuations in every topographic cortical area
we studied except IPS2, V4, and VO1. We matched the
visual stimulus and task difficulty in the two attention
conditions, allowing us to attribute changes in the mag-
nitude of slow fluctuations to changes in spatial attention.
Moreover, our task and stimulus design are uniquely
suited to reveal effects of attention at very slow frequen-
cies of fluctuations in brain activity that are unrelated to
stimulus-evoked responses. On alternate 5-min runs,
attention was directed either to a central fixation point
or to a peripheral visual stimulus that was rotating at
0.03 Hz. Fourier analysis of the fMRI response therefore
distinguishes between effects of attention on amplitude
at the stimulus frequency from effects on amplitude of
endogenous fluctuations in fMRI signals at other frequen-
cies. Most fMRI studies of spatial attention employ either
event-related or block designs in trial-based paradigms to
assess the effects of discrete shifts of spatial attention. In
these designs, the experimental effects are typically not
constrained to specific frequency bands, and they are
usually not compatible with measuring effects of atten-
tion on slow fluctuations in brain activity (Huk, Bonnen,
& He, 2018). Therefore, our design enables us to assess
effects that would be very difficult to discover using more
standard visual attention paradigms.
Relationships among Slow fMRI Fluctuations,
Neural Activity, and Behavior
Fluctuations in fMRI signal in the frequency range that we
have studied (<0.1 Hz) are likely to be influenced by
both neural and nonneural factors. For example, fluctua-
tions in arterial carbon dioxide concentration ( Wise et al.,
2004), cardiac rate (Shmueli et al., 2007), and respiratory
rate (Birn et al., 2006) all occur at these frequencies. For
this reason, many studies apply a high-pass filter during
preprocessing of the fMRI BOLD response to reduce the
contributions of these sources of “noise.” However,
neural activity also contributes to slow endogenous
fluctuations in fMRI signals. The amplitude of BOLD re-
sponses to a given stimulus is correlated with levels of
single-unit spiking, multiunit activity, and local field po-
tential gamma power (Mukamel et al., 2005; Logothetis,
Pauls, Augath, Trinath, & Oeltermann, 2001), and similar
relationships between fMRI signals and direct measures
of neuronal activity have also been observed for slow en-
dogenous fluctuations (Keller et al., 2013; Schölvinck,
Maier, Ye, Duyn, & Leopold, 2010; Shmuel & Leopold,
2008). Therefore, the BOLD signals in the very slow
range that we have studied (<0.1 Hz) likely include com-
ponents that reflect neural activity.
Moreover, fluctuations in brain activity on this time-
scale are cognitively and behaviorally relevant. The global
component of resting state fMRI measurements has been
associated with general levels of arousal and alertness
(Turchi et al., 2018; Wong, Olafsson, Tal, & Liu, 2013).
Within the frequency range we have studied (<0.1 Hz),
stimulus detection is predicted by spontaneous slow neg-
ative cortical shifts in EEG signals (Devrim, Demiralp,
Kurt, & Yücesir, 1999) and by the phase of ongoing
EEG oscillations in the infraslow range (Monto et al.,
2008). Additionally, previous studies have demonstrated
relationships between performance on visual detection
tasks and prestimulus fMRI activity recorded a few
seconds before stimulus onset (Coste & Kleinschmidt,
2016; Wohlschläger et al., 2016; Schölvinck, Friston, &
Rees, 2012; Hesselmann et al., 2008), and fMRI connec-
tivity between brain regions at very slow frequencies is
significantly correlated with performance across partici-
pants in an executive control task (Xu et al., 2014) and
a visual attention task (Griffis, Elkhetali, Burge, Chen, &
Visscher, 2015). Our results extend these findings by
showing that spatial attention suppresses slow endoge-
nous fluctuations in occipital and parietal cortex and that
reduced endogenous activity predicts performance on a
visual detection task.
Various theories have been proposed regarding the
roles of slow endogenous activity fluctuations in cortical
function, including self-regulation of cortical excitability
to avoid overactivation and insensitivity (Birbaumer,
Elbert, Canavan, & Rockstroh, 1990), generation of
predictive internal models of the environment (Berkes,
Orbán, Lengyel, & Fiser, 2011), providing a substrate
for top–down expectation signals (Ringach, 2009), and in-
tegrating information across wide regions of cortex to
support consciousness (He & Raichle, 2009). One addi-
tional possibility is that slow endogenous fluctuations
are important for coordinating neural activity across dis-
tant regions of cortex in the service of nonperceptual
functions but that they can also divert resources from
performance of specific perceptual tasks and/or interfere
with representations of sensory information.
Differential Amounts of Attentional Enhancement
of Stimulus-Evoked Responses and Attentional
Suppression of Endogenous Fluctuations across
Cortical Areas
We found that the relative amounts of attentional en-
hancement of the visually evoked response and suppres-
sion of endogenous fluctuations varied across areas. In
early and ventral visual cortical areas, an increase in the
strength of the evoked response contributed to en-
hanced response reliability, but this effect was not ob-
served in posterior parietal cortical areas IPS1 and IPS2
or in lateral occipital cortical areas. In contrast, every to-
pographic cortical area we studied, except IPS2, V4 and
VO1, exhibited a significant decrease in endogenous fluc-
tuations at nonstimulus frequencies that also contributed
to enhanced response reliability.
96
Journal of Cognitive Neuroscience
Volume 32, Number 1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
1
e
8
-
5
p
d
1
9
f
3
/
1
3
1
2
4
/
4
1
/
o
8
c
5
n
/
_
a
1
_
8
0
6
1
1
4
4
7
0
0
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
7
0
0
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
f
/
o
n
0
5
M
a
y
2
0
2
1
Ventral cortical areas V4 and VO1 exhibited the stron-
gest increases in stimulus-evoked response and the
weakest reductions in the strength of ongoing fluctua-
tions. In contrast, dorsal areas V3A/B, IPS0, and IPS1, as
well as lateral occipital areas LO1 and LO2, showed little
or no increase in evoked response and the strongest re-
ductions in endogenous fluctuations. Early visual cortical
areas V1–V3 displayed intermediate levels of both re-
sponse enhancement and suppression at nonstimulus
frequencies by attention.
Effects of Attention on Enhancement, Suppression,
and Response Reliability
Although our results reveal an inverse relationship be-
tween the amount of attentional enhancement of the re-
sponse to the stimulus and attentional suppression of
endogenous fluctuations across cortical areas, these two
effects are unrelated across fMRI runs within a cortical
area. That is, the amplitude of the stimulus-evoked re-
sponse and the amplitude of slow endogenous fluctua-
tions of nonstimulus frequencies were not significantly
correlated, suggesting that they may reflect dissociable
mechanisms, at least at the timescale studied here
(5-min runs). Previous studies have also shown dissocia-
tions between effects of attention on evoked responses
and on correlated activity fluctuations in pairs of neurons
in macaque visual cortex (Ruff & Cohen, 2014; Herrero
et al., 2013).
Studies employing microelectrode recordings have
revealed that spatial attention enhances response reliabil-
ity, both by increasing the evoked response amplitude
(McAdams & Maunsell, 1999) and by decreasing intertrial
response variability (Cohen & Maunsell, 2009; Mitchell
et al., 2007, 2009). Attention can also enhance response
reliability by decorrelating fluctuations in spiking within
neuronal populations (Ruff & Cohen, 2014; Herrero
et al., 2013; Cohen & Maunsell, 2009; Mitchell et al.,
2009). These studies describe attentional suppression
of correlated activity on much shorter timescales than
those for which we have demonstrated attentional sup-
pression of endogenous fluctuations of fMRI signals.
However, the strength of local interneuronal correlations
is itself correlated with BOLD activity (Nir et al., 2007).
Attentional enhancement of the stimulus-evoked re-
sponse was significantly greater in all topographically de-
fined regions (except LO1 and LO2) than in a control
nontopographic cortical region (PCC/precuneus). In ad-
dition, attentional suppression of endogenous fluctua-
tions was significantly greater in all topographically
defined regions (except V4, IPS2, and VO1) than in
PCC/precuneus. However, the distinction between topo-
graphic areas and this control area was less clear for cor-
relations between behavior and strength of endogenous
fluctuations: Although all topographic regions exhibited a
significant correlation and the control PCC/precuneus re-
gion did not, there were no significant differences in
correlation coefficients between individual topographic
regions and PCC/precuneus.
These results suggest that the negative correlation be-
tween behavioral performance on the attention-to-wedge
task and the strength of slow endogenous fluctuations
(Figure 7B) may be more widespread across cortical
areas than the effects of attending to the wedge (as com-
pared with attending to fixation) on the amplitude of
slow endogenous fluctuations (Figure 3B). In addition,
attentional suppression of endogenous fluctuations was
strongest at very low temporal frequencies (<0.03 Hz;
Figure 4), whereas the negative correlation between tar-
get detection performance and the amplitude of endog-
enous fluctuations was more evident at higher temporal
frequencies (>0.02 Hz; Figur 8).
Conclusions
In addition to increasing the strength of visually evoked
BOLD responses, sustained covert visual spatial attention
to a periodic stimulus decreased the strength of endoge-
nous BOLD fluctuations. Reduced endogenous activity
substantially contributed to improved response reliability
in every topographically organized cortical area we stud-
ied except IPS2, V4, and VO1 and was inversely correlated
with behavioral performance in all topographic areas. In
Kontrast, attention’s effects on visually evoked BOLD re-
sponses contributed to improved response reliability
only in early and ventral cortical areas, and the amplitude
of stimulus-evoked responses did not predict behavioral
performance in any area. daher, our results provide
evidence that attentional suppression of endogenous
fluctuations in brain activity enhances cortical representa-
tions of visual stimuli and substantially benefits visual
perception.
Danksagungen
This work was supported by the National Eye Institute at the
Nationale Gesundheitsinstitute (R01 EY025278 to M. A. S.,
Training Grant in Vision Science T32 EY07043, and Core Grant
for Vision Research P30 EY003176), the National Science
Foundation (Graduate Research Fellowship to D. W. B. Und
Major Instrumentation Program grant BCS-0821855), und das
National Institute on Aging at the National Institutes of Health
(NRSA F31 AG032209 to A. R.).
Reprint requests should be sent to Michael Silver, School of
Optometry, 360 Minor Hall, Universität von Kalifornien, Berkeley,
Berkeley, CA 94720-2020, oder per E-Mail: masilver@berkeley.edu.
VERWEISE
Berkes, P., Orbán, G., Lengyel, M., & Fiser, J. (2011).
Spontaneous cortical activity reveals hallmarks of an optimal
internal model of the environment. Wissenschaft, 331, 83–87.
Birbaumer, N., Elbert, T., Canavan, A. G., & Rockstroh, B.
(1990). Slow potentials of the cerebral cortex and behavior.
Physiological Review, 70, 1–41.
Bressler, Rokem, and Silver
97
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
1
e
8
–
5
P
D
1
9
F
3
/
1
3
1
2
4
/
4
1
/
Ö
8
C
5
N
/
_
A
1
_
8
0
6
1
1
4
4
7
0
0
0
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
4
N
7
0
0
8
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
T
/
F
Ö
N
0
5
M
A
j
2
0
2
1
Birn, R. M., Diamond, J. B., Schmied, M. A., & Bandettini, P. A.
(2006). Separating respiratory-variation-related fluctuations
from neuronal-activity-related fluctuations in fMRI.
Neurobild, 31, 1536–1548.
Bressler, D. W., Fortenbaugh, F. C., Robertson, L. C., & Silver, M. A.
(2013). Visual spatial attention enhances the amplitude of
positive and negative fMRI responses to visual stimulation in an
eccentricity-dependent manner. Sehforschung, 85, 104–112.
Bressler, D. W., & Silver, M. A. (2010). Spatial attention
improves reliability of fMRI retinotopic mapping signals in
occipital and parietal cortex. Neurobild, 53, 526–533.
Brewer, A. A., Liu, J., Wade, A. R., & Wandell, B. A. (2005). Visual
field maps and stimulus selectivity in human ventral occipital
Kortex. Naturneurowissenschaften, 8, 1102–1109.
Busch, N. A., Dubois, J., & VanRullen, R. (2009). The phase of
ongoing EEG oscillations predicts visual perception. Zeitschrift
der Neurowissenschaften, 29, 7869–7876.
Carrasco, M. (2011). Visual attention: The past 25 Jahre. Vision
Forschung, 51, 1484–1525.
Cohen, M. R., & Maunsell, J. H. R. (2009). Attention improves
performance primarily by reducing interneuronal
correlations. Naturneurowissenschaften, 12, 1594–1600.
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., &
Shulman, G. L. (2000). Voluntary orienting is dissociated
from target detection in human posterior parietal cortex.
Naturneurowissenschaften, 3, 292–297.
Coste, C. P., & Kleinschmidt, A. (2016). Cingulo-opercular network
activity maintains alertness. Neurobild, 128, 264–272.
Devrim, M., Demiralp, T., Kurt, A., & Yücesir, ICH. (1999). Slow
cortical potential shifts modulate the sensory threshold in
human visual system. Neurowissenschaftliche Briefe, 270, 17–20.
Engel, S. A., Rumelhart, D. E., Wandell, B. A., Lee, A. T., Glover,
G. H., Chichilnisky, E. J., et al. (1994). fMRI of human visual
Kortex. Natur, 369, 525.
Fuchs, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Rachel, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
102, 9673–9678.
Fransson, P. (2006). How default is the default mode of brain
Funktion? Further evidence from intrinsic BOLD signal
fluctuations. Neuropsychologie, 44, 2836–2845.
Gandhi, S. P., Heeger, D. J., & Boynton, G. M. (1999). Spatial
attention affects brain activity in human primary visual cortex.
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
96, 3314–3319.
Genovese, C. R., Lazar, N. A., & Nichols, T. (2002). Thresholding
of statistical maps in functional neuroimaging using the false
discovery rate. Neurobild, 15, 870–878.
Greicius, M. D., Krasnow, B., Reiss, A. L., & Menon, V. (2003).
Functional connectivity in the resting brain: A network
analysis of the default mode hypothesis. Verfahren der
Nationale Akademie der Wissenschaften, USA., 100, 253–258.
Griffis, J. C., Elkhetali, A. S., Burge, W. K., Chen, R. H., &
Visscher, K. M. (2015). Retinotopic patterns of background
connectivity between V1 and fronto-parietal cortex are
modulated by task demands. Frontiers in Human
Neurowissenschaften, 9, 338.
Er, B. J., & Rachel, M. E. (2009). The fMRI signal, slow cortical
potential and consciousness. Trends in den Kognitionswissenschaften,
13, 302–309.
Herrero, J. L., Gieselmann, M. A., Sanayei, M., & Thiele, A.
(2013). Attention-induced variance and noise correlation
reduction in macaque V1 is mediated by NMDA receptors.
Neuron, 78, 729–739.
Hesselmann, G., Kell, C. A., & Kleinschmidt, A. (2008). Ongoing
activity fluctuations in hMT+ bias the perception of coherent
visual motion. Zeitschrift für Neurowissenschaften, 28, 14481–14485.
Huk, A., Bonnen, K., & Er, B. J. (2018). Beyond trial-based
paradigms: Continuous behavior, ongoing neural activity, Und
natural stimuli. Zeitschrift für Neurowissenschaften, 38, 7551–7558.
Imamoglu, F., Heinzle, J., Imfeld, A., & Haynes, J. D. (2014).
Activity in high-level brain regions reflects visibility of low-
level stimuli. Neurobild, 102, 688–694.
Jenkinson, M., Bannister, P., Brady, M., & Schmied, S. (2002).
Improved optimization for the robust and accurate linear
registration and motion correction of brain images.
Neurobild, 17, 825–841.
Keller, C. J., Bickel, S., Honey, C. J., Groppe, D. M., Entz, L.,
Craddock, R. C., et al. (2013). Neurophysiological
investigation of spontaneous correlated and anticorrelated
fluctuations of the BOLD signal. Zeitschrift für Neurowissenschaften,
33, 6333–6342.
Larsson, J., & Heeger, D. J. (2006). Two retinotopic visual areas
in human lateral occipital cortex. Journal of Neurosciencce,
26, 13128–13142.
Logothetis, N. K., Pauls, J., Augath, M., Trinath, T., &
Oeltermann, A. (2001). Neurophysiological investigation of
the basis of the fMRI signal. Natur, 412, 150–157.
Mathewson, K. E., Gratton, G., Fabiani, M., Beck, D. M., & Ro, T.
(2009). To see or not to see: Prestimulus alpha phase
predicts visual awareness. Zeitschrift für Neurowissenschaften, 29,
2725–2732.
Maunsell, J. H. R. (2015). Neuronal mechanisms of visual
attention. Annual Review of Vision Science, 1, 373–391.
McAdams, C. J., & Maunsell, J. H. R. (1999). Effects of attention
on the reliability of individual neurons in monkey visual
Kortex. Neuron, 23, 765–773.
Mitchell, J. F., Sundberg, K. A., & Reynolds, J. H. (2007). Differential
attention-dependent response modulation across cell
classes in macaque visual area V4. Neuron, 55, 131–141.
Mitchell, J. F., Sundberg, K. A., & Reynolds, J. H. (2009). Spatial
attention decorrelates intrinsic activity fluctuations in
macaque area V4. Neuron, 63, 879–888.
Monto, S., Verehrung, S., Voipio, J., & Verehrung, J. M. (2008). Very slow
EEG fluctuations predict the dynamics of stimulus detection
and oscillation amplitudes in humans. Zeitschrift für
Neurowissenschaften, 28, 8268–8272.
Mukamel, R., Gelbard, H., Arieli, A., Hasson, U., Fried, ICH., &
Malach, R. (2005). Coupling between neuronal firing, field
potentials, and fMRI in human auditory cortex. Wissenschaft,
309, 951–954.
Müller, N. G., & Kleinschmidt, A. (2004). The attentional
‘spotlight’s’ penumbra: Center-surround modulation in
striate cortex. NeuroReport, 15, 977–980.
Murray, S. O., & Er, S. (2006). Contrast invariance in the human
lateral occipital complex depends on attention. Current
Biology, 16, 606–611.
Nir, Y., Fisch, L., Mukamel, R., Gelbard-Sagiv, H., Arieli, A.,
Fried, ICH., et al. (2007). Coupling between neuronal firing rate,
gamma LFP, and BOLD fMRI is related to interneuronal
correlations. Aktuelle Biologie, 17, 1275–1285.
Ress, D., Backus, B. T., & Heeger, D. J. (2000). Activity in
primary visual cortex predicts performance in a visual
detection task. Naturneurowissenschaften, 3, 940–945.
Ringach, D. L. (2009). Spontaneous and driven cortical activity:
Implications for computation. Aktuelle Meinung in
Neurobiology, 19, 439–444.
Ruff, D. A., & Cohen, M. R. (2014). Attention can either increase
or decrease spike count correlations in visual cortex. Natur
Neurowissenschaften, 17, 1591–1597.
Schölvinck, M. L., Friston, K. J., & Rees, G. (2012). The influence
of spontaneous activity on stimulus processing in primary
visual cortex. Neurobild, 59, 2700–2708.
Schölvinck, M. L., Maier, A., Ye, F. Q., Duyn, J. H., & Leopold,
D. A. (2010). Neural basis of global resting-state fMRI activity.
98
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 1
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
1
e
8
–
5
P
D
1
9
F
3
/
1
3
1
2
4
/
4
1
/
Ö
8
C
5
N
/
_
A
1
_
8
0
6
1
1
4
4
7
0
0
0
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
4
N
7
0
0
8
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
/
.
T
F
Ö
N
0
5
M
A
j
2
0
2
1
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
107, 10238–10243.
Sereno, M. ICH., Dale, A. M., Reppas, J. B., Kwong, K. K., Belliveau,
J. W., Brady, T. J., et al. (1995). Borders of multiple visual
areas in humans revealed by functional magnetic resonance
Bildgebung. Wissenschaft, 268, 889–893.
Shmuel, A., & Leopold, D. A. (2008). Neuronal correlates of
spontaneous fluctuations in fMRI signals in monkey visual
Kortex: Implications for functional connectivity at rest.
Kartierung des menschlichen Gehirns, 29, 751–761.
Shmueli, K., van Gelderen, P., de Zwart, J. A., Horovitz, S. G.,
Fukunaga, M., Jansma, J. M., et al. (2007). Low-frequency
fluctuations in the cardiac rate as a source of variance in the
resting-state fMRI BOLD signal. Neurobild, 38, 306–320.
Silver, M. A., Ress, D., & Heeger, D. J. (2005). Topographic
maps of visual spatial attention in human parietal cortex.
Journal of Neurophysiology, 94, 1358–1371.
Silver, M. A., Ress, D., & Heeger, D. J. (2007). Neural correlates
of sustained spatial attention in human early visual cortex.
Journal of Neurophysiology, 97, 229–237.
Supèr, H., van der Togt, C., Spekreijse, H., & Lamme, V. A. F.
(2003). Internal state of monkey primary visual cortex ( V1)
predicts figure-ground perception. Zeitschrift für Neurowissenschaften,
23, 3407–3414.
Turchi, J., Chang, C., Ye, F. Q., Russ, B. E., Yu, D. K.,
Cortes, C. R., et al. (2018). The basal forebrain regulates
global resting-state fMRI fluctuations. Neuron, 97,
940–952.
Wandell, B. A., Chial, S., & Backus, B. T. (2000). Visualisierung
and measurement of the cortical surface. Zeitschrift für
Cognitive Neuroscience, 12, 739–752.
Weissman, D. H., Roberts, K. C., Visscher, K. M., & Woldorff,
M. G. (2006). The neural bases of momentary lapses in
attention. Naturneurowissenschaften, 9, 971–978.
Wise, R. G., Ide, K., Poulin, M. J., & Tracey, ICH. (2004). Resting
fluctuations in arterial carbon dioxide induce significant low
frequency variations in BOLD signal. Neurobild, 21,
1652–1664.
Wohlschläger, A. M., Glim, S., Shao, J., Draheim, J., Köhler, L.,
Lourenço, S., et al. (2016). Ongoing slow fluctuations in V1
impact on visual perception. Frontiers in Human
Neurowissenschaften, 10, 411.
Wong, C. W., Olafsson, V., Tal, O., & Liu, T. T. (2013). Der
amplitude of the resting-state fMRI global signal is related to
EEG vigilance measures. Neurobild, 83, 983–990.
Xu, J., Rees, G., Yin, X., Song, C., Han, Y., Ge, H., et al. (2014).
Spontaneous neuronal activity predicts intersubject variations in
executive control of attention. Neurowissenschaften, 263, 181–192.
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L.,
Steinmetz, M. A., Pekar, J. J., et al. (2002). Transient neural
activity in human parietal cortex during spatial attention
shifts. Naturneurowissenschaften, 5, 995–1002.
Zarahn, E., Aguirre, G. K., & D'Esposito, M. (1997). Empirical
analyses of BOLD fMRI statistics. ICH. Spatially unsmoothed data
collected under null-hypothesis conditions. Neurobild,
5, 179–197.
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
1
e
8
–
5
P
D
1
9
F
3
/
1
3
1
2
4
/
4
1
/
Ö
8
C
5
N
/
_
A
1
_
8
0
6
1
1
4
4
7
0
0
0
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
4
N
7
0
0
8
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
T
/
.
F
Ö
N
0
5
M
A
j
2
0
2
1
Bressler, Rokem, and Silver
99