Saccade Reorienting Is Facilitated by Pausing the
Oculomotor Program
Antimo Buonocore1, Simran Purokayastha2, and Robert D. McIntosh2
Abstrakt
■ As we look around the world, selecting our targets, compet-
ing events may occur at other locations. Depending on current
Ziele, the viewer must decide whether to look at new events or
to ignore them. Two experimental paradigms formalize these
response options: double-step saccades and saccadic inhibition.
In the first, the viewer must reorient to a newly appearing tar-
get; in the second, they must ignore it. Until now, the relation-
ship between reorienting and inhibition has been unexplored.
In three experiments, we found saccadic inhibition ∼100 msec
after a new target onset, regardless of the task instruction.
Darüber hinaus, if this automatic inhibition is boosted by an irrele-
vant flash, reorienting is facilitated, suggesting that saccadic in-
hibition plays a crucial role in visual behavior, as a bottom–up
brake that buys the time needed for decisional processes to act.
Saccadic inhibition may be a ubiquitous pause signal that pro-
vides the flexibility for voluntary behavior to emerge. ■
EINFÜHRUNG
As we look around the world, selecting the targets of our
eye movements, the scene may change. Our next planned
target may move or an event of greater urgency may occur
at another location. Depending on current goals, Die
viewer must decide whether to look at new events or to
ignore them. These response options have been formal-
ized within two powerful experimental paradigms: double-
step saccades and saccadic inhibition. In the first, Die
viewer must reorient to a target appearing shortly before
a planned saccade; in the second, they must ignore it.
Each paradigm has been studied extensively, but the rela-
tionship between reorienting and inhibition is hitherto
unexplored. This study seeks to bridge that gap, testing
the presence and possible functional role of oculomotor
distraction in reorienting.
Irrelevant distractors robustly delay saccade execution
(z.B., Walker, Kentridge, & Findlay, 1995; Weber & Fischer,
1994). Analysis of latency distributions reveals that distrac-
tors cause a specific dip in saccadic activity ∼60–120 msec
after distractor onset, with the suppressed saccades launch-
ing later (Bompas & Sumner, 2011; Edelman & Xu, 2009;
Buonocore & McIntosh, 2008). This pause in visual be-
havior, known as “saccadic inhibition,” is a fast, reflexive
response of the oculomotor system to any salient visual
ändern, and it occurs regardless of the task instruction
(Reingold & Stampe, 1999, 2002, 2003, 2004). We should
thus expect that saccadic inhibition will accompany the
change in target position in double-step tasks.
1Tübingen University, 2University of Edinburgh
The literature on double-step saccadic performance
contains hints that this may be the case. Allgemein, sac-
cades launching immediately after a target step do not
have time to reflect the step and are directed toward
the initial target, whereas saccades launching at longer
delays are progressively more likely to go toward the sec-
ond target (Becker & Jürgens, 1979; Lisberger, Fuchs,
King, & Evinger, 1975). At least for large target steps, Das
“transition function” is discontinuous, with saccades
launched less than ∼60 msec after the target step going to
the initial target, saccades launched more than ∼120 msec
after the step going to the new target, and few if any
saccades in the intervening time window (Aslin & Shea,
1987; Findlay & Harris, 1984). There is thus a pause in
behavior in double-step tasks that corresponds roughly
with the time course of saccadic inhibition. Speculation
that the target step in double-step tasks has a distract-
ing effect can be found quite far back in the literature
(Sheliga, Braun, & Miles, 2002; Findlay & Walker, 1999),
yet this important possibility has never been formally
examined.
The first aim of this study is to assess whether the sig-
nature of saccadic inhibition is present in a double-step
Kontext, comparing between two tasks, matched for stim-
ulus events, in which a new visual event is to be responded
to as a target or ignored as irrelevant. We demonstrate
that saccadic inhibition does indeed occur in a double-
step context, just as in a distractor context. Experiment 3
is then designed to study the consequences of this reflex-
ive inhibition for reorienting behavior. This is done by
boosting the inhibitory signal during double-step reorient-
ing, with a salient generalized flash presented at the
same time as the target jump. One might expect that this
© 2017 Massachusetts Institute of Technology. Published under a
Creative Commons Atrribution 3.0 Unportiert (CC BY 3.0) Lizenz.
Zeitschrift für kognitive Neurowissenschaften 29:12, S. 2068–2080
doi:10.1162/jocn_a_01179
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
T
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
irrelevant event would interfere with the ability to respond
to the new target; but a more interesting, counterintuitive
possibility is that the flash will interfere mainly with the
planned response to the initial target, creating a pause
in behavior that would increase the subsequent likelihood
of successful reorienting. Wenn ja, we would predict that
reorienting would be enhanced under conditions of
boosted inhibition, indexed by an increased proportion
of saccades being successfully redirected. This would pro-
vide strong support for a long-standing but little-discussed
idea that saccadic inhibition is a feature, not a bug of the
oculomotor system, which buys time for the system to
evaluate any salient change and to alter the next saccade
if necessary (Reingold & Stampe, 2002).
METHODEN
Teilnehmer
Eighteen volunteers, aged between 18 Und 30 Jahre, Par-
ticipated in the three experiments (six in each). Das
study is concerned with basic design features of the
oculomotor system, so we are targeting only very consis-
tent effects that should be near-universal. The number of
participants per experiment is therefore small (n = 6),
with effort directed to maximizing the number of obser-
vations per participant to ensure robust individual param-
eter estimates. Our focus is on a high consistency of
behavior across participants, and individual parameters
are reported for each participant in each experiment.
All were free from neurological and visual impairments.
This experiment was conducted in accordance with the
1964 Declaration of Helsinki, with the approval of the
University of Edinburgh Psychology research ethics
committee.
Apparatus, Stimuli, and Procedure
Stimuli were black on a mid-gray background, vorgeführt
on a 19-in. CRT monitor (1024 × 768 pixels) bei 100 Hz.
All the experiments were implemented in Experiment
Builder (SR Research, Ottawa, ON, Kanada). Teilnehmer
were seated with their head on a chinrest, their eyes
aligned with the center of the screen at a distance of
80 cm. Eye movements were recorded with the EyeLink
1000 System (SR Research; detection algorithm: pupil
and corneal reflex; 1,000 Hz sampling; saccade detection
based on 30°/s velocity and 8,000°/s2 acceleration thresh-
olds). Each trial began with drift correction and a tone
followed by a 0.5° central fixation cross. In Experiments
1 Und 2 (but not Experiment 3), 1° black outline circles
were also presented at 6° to left and right of fixation as
placeholders for potential targets. In all experiments, A
5-point calibration on the horizontal and vertical axes
was run at the beginning of each session and after three
consecutive blocks. Additional calibrations were run if
the participant’s head moved from the chinrest.
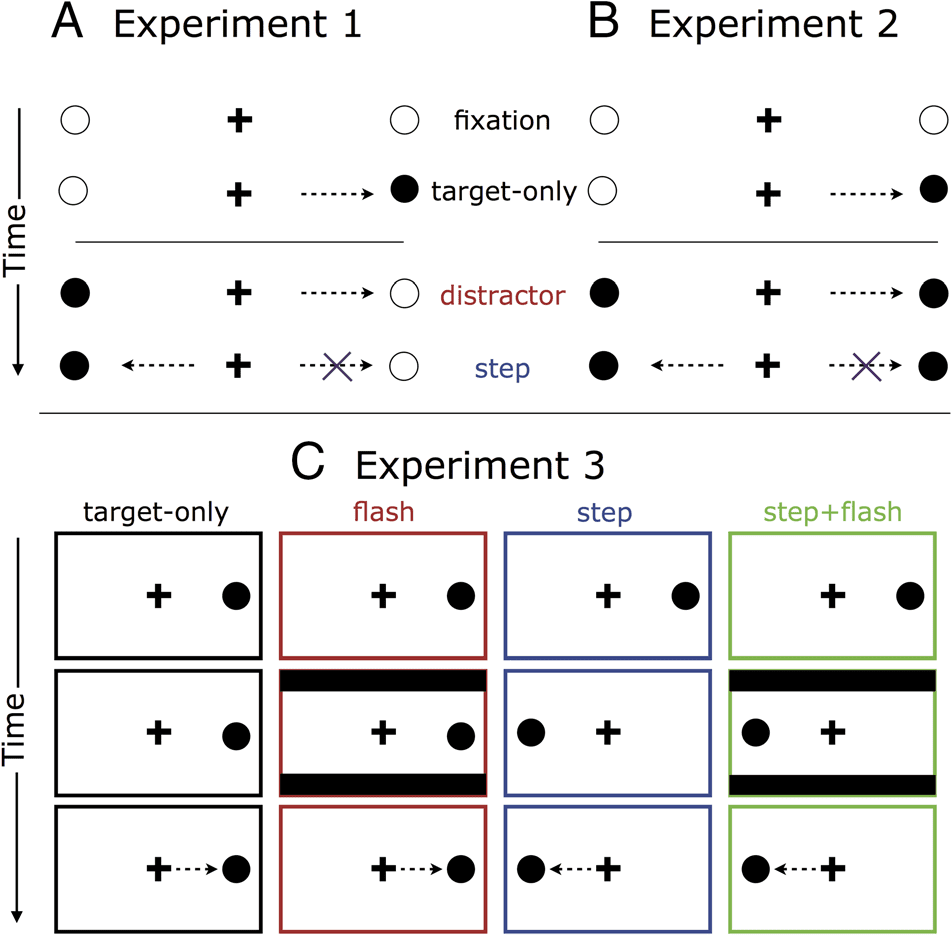
Experiment 1 (Figure 1A)
Each trial began with fixation followed by the onset of
Target 1, created by filling in one of the two lateralized
outline circles. In half of the trials, Target 1 remained
filled until the screen went gray 700 msec later (target-
only trials). In the other half of the trials, Target 1 unfilled
and the outline on the opposite side filled simultaneously
(Target 2), after a delay determined individually for each
participant (see Preliminary Block section). In the “dis-
tractor” condition, participants were required to ignore
Target 2 and to move their eyes to the Target 1 location.
In the “step” condition, participants were required to
move their eyes to the second target if it appeared.
Each participant completed two sessions for the dis-
tractor and step conditions,1 following an ABBA se-
quence, counterbalanced across participants. Within
each session, there were 10 blocks of 24 Versuche; Target 1
appeared equally often on each side and, for each side,
was followed by Target 2 in half of the trials. Teilnehmer
thus completed 240 target-only and 240 distractor trials
in the distractor condition and 240 target-only and 240 step
trials in the step condition.
Experiment 2 (Figure 1B)
This was identical to Experiment 1, except that Target 1
did not unfill when Target 2 was presented. Stattdessen, für
the trials in which Target 2 appeared, both targets re-
mained filled until the end of the trial. Daher, wohingegen
Experiment 1 used stimulus conditions typical of double-
step tasks (Target 1 replaced by Target 2), Experiment 2
used conditions more typical of distractor tasks (Target 1
joined by Target 2).
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Figur 1. Schematic diagram of trial events. See Methods for details.
Buonocore, Purokayastha, and McIntosh
2069
Experiment 3 (Abbildung 1C)
Saccadic Inhibition Analysis
This was similar to Experiments 1 Und 2, with some im-
portant changes. After the initial fixation period, a 1°
black dot appeared on the left or the right (Target 1).
After the delay determined individually for each par-
ticipant (see Preliminary Block section), one or two
changes could occur: In flash trials (25% of trials), Dort
was a distracting flash of maximum contrast, dauerhaft
30 msec and covering the top and bottom thirds of
the screen; in step trials (25% of trials), Target 1 dis-
appeared and was replaced by an identical target on
the opposite side (Target 2); in step-flash trials (25% von
Versuche), both changes happened simultaneously; in target-
only trials (25% of trials), neither change happened.
In all trials, the participant was required to move their
eyes as rapidly as possible to the target, thus to change
plan if the target stepped. Note that the flash condition
of Experiment 3 is similar to the distractor conditions of
Experimente 1 Und 2, except that the distracting event is
a large generalized flash.
Each participant completed two sessions of 10 blocks
jede. Within each block, there were 48 Versuche; Target 1
appeared equally often on the left and the right, Und
for each target side, there were six trials in each of the
stimulus conditions (target-only, flash, step, step-flash).
Participants thus completed 240 trials per condition.
Preliminary Block
Before each experiment, participants performed a block
von 70 trials of the target-only condition for that experi-
ment. The median saccadic RT of the last 50 trials was
berechnet, Und 100 msec was subtracted from this value
to set the timing of transient events for that participant in
the subsequent experiment. These participant-specific
timings ensure that saccadic inhibition, which is maximal
around 90–100 msec after a transient event, will impact
upon a dense portion of the expected saccadic distribution.
The individual transient onset times relative to initial target
onset are listed per participant: Experiment 1: 150, 150,
120, 120, 90, 170; Experiment 2: 120, 90, 140, 130, 150,
140; Experiment 3: 20, 120, 120, 170, 130, 80.
Data Processing
All the data processing and analysis were performed in
MATLAB (The MathWorks, Inc., Natick, MA). Only the
first eye movement following target onset was analyzed.
A total of 9% (Experiment 1), 2.7% (Experiment 2), Und
4.3% (Experiment 3) of trials were excluded from the
analysis due to blinks, latencies shorter than 70 ms
or longer than 500 ms, and saccades smaller than
2° amplitude or with a duration exceeding 100 ms. Für
the target-only condition, we also excluded saccades made
in the wrong direction (Experiment 1: 0.67%, Experiment 2:
1.31%, Experiment 3: 0.21%).
To chart the time course of saccadic activity, we applied a
distribution analysis to the saccadic RTs (vgl. McIntosh &
Buonocore, 2014; Bompas & Sumner, 2011; Reingold &
Stampe, 2002). In each experiment, für jeden Teilnehmer
and condition, we computed the probability density esti-
mate of saccades at each millisecond, with time zero de-
fined by the transient change. The estimate was smoothed
using a kernel-smoothing window of 8 ms. For each par-
ticipant, for each experiment, the target-only condition was
taken as the baseline distribution of saccadic activity. Wir
berechnet, at each millisecond, the proportional change
relative to this baseline in each of the transient change
Bedingungen (distractor or step in Experiments 1 Und 2;
flash, step, or step-flash in Experiment 3) using Equation 1,
which expresses reduced saccadic activity as positive:
ptarget tð Þ − pchange tð Þ
ptarget tð Þ
(1)
where p(T) is the probability for the target-only condition
und p(C) is the probability for the transient change con-
dition. To keep the estimate of proportional change reli-
able, considering the low frequency of saccades in the
initial and final part of the distribution, we extracted
data only from 45 Zu 250 msec after the change. Aus
each of these profiles, we extracted inhibition magnitude
(maximum inhibition), inhibition latency (time to maxi-
mum inhibition), and inhibition start and end times,
defined operationally as the times at which inhibition
crossed 50% of its maximum level, before and after the
maximum, jeweils (Reingold & Stampe, 2002).
Double-step Analysis
To characterize reorienting behavior for each condition
that had a “step” (d.h., in which the participant was re-
quired to reorient to Target 2), we derived a direction
transition function. Erste, we classified each saccade as
directed at Target 1 or Target 2, according to the hemi-
field in which it landed (the distribution of landing posi-
tions was bimodal, as is clear in Figures 2, 4, Und 6). Wir
then computed the probability of responding to Target 1
as function of time since the step and fitted a logistic
function using Equation 2:
1
D
1 þ e
−a * t−b
D
Þ
Þ
(2)
where e is the natural logarithm base, a represents the
steepness of the curve, b is the inflection point, Und
the numerator represents the maximum probability.
Probability of Responses to Target 1
In each experiment, for each condition, we estimated
the proportion of responses to Target 1, with Target 1
responses coded as 1 and Target 2 responses coded as 0.
2070
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
T
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
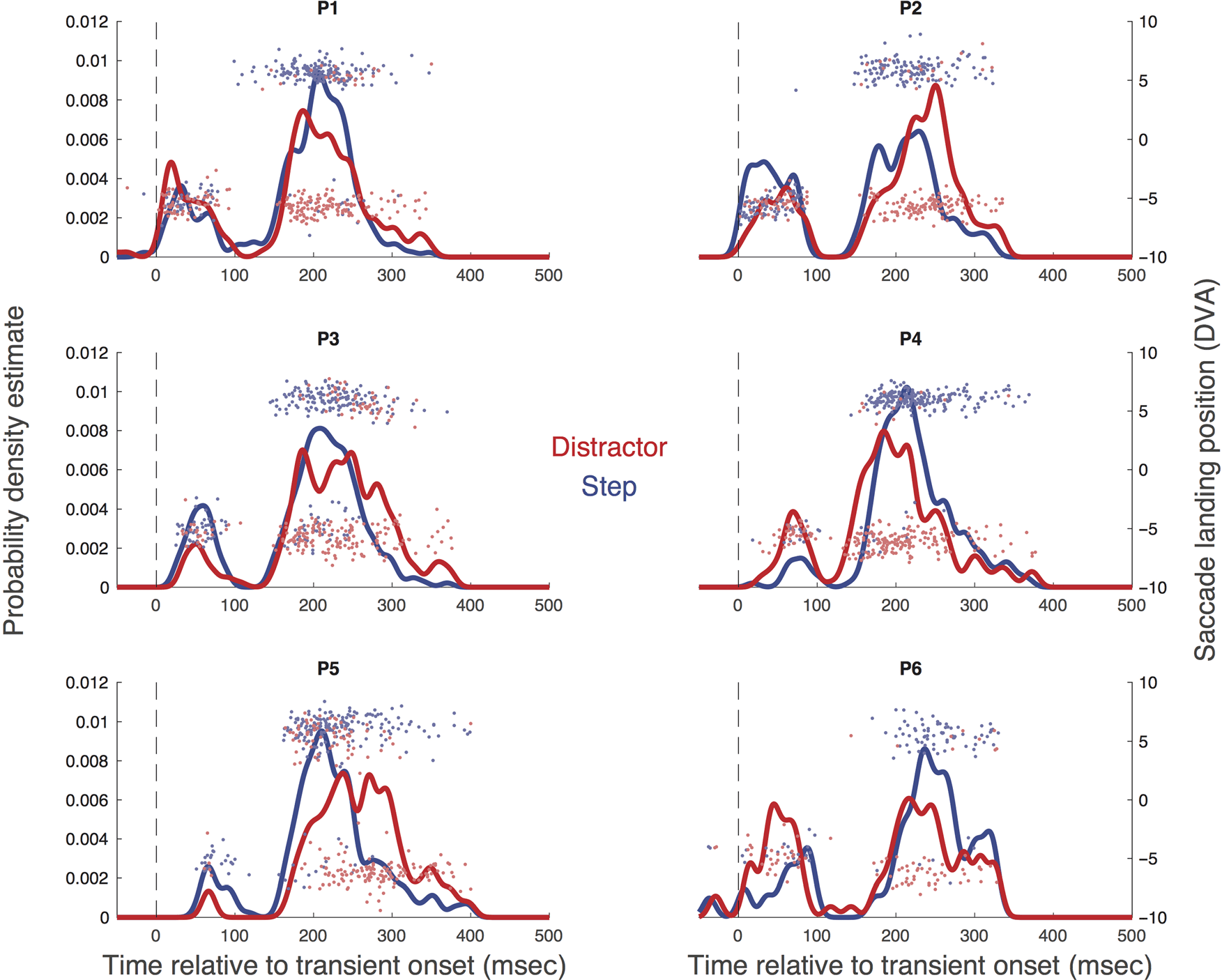
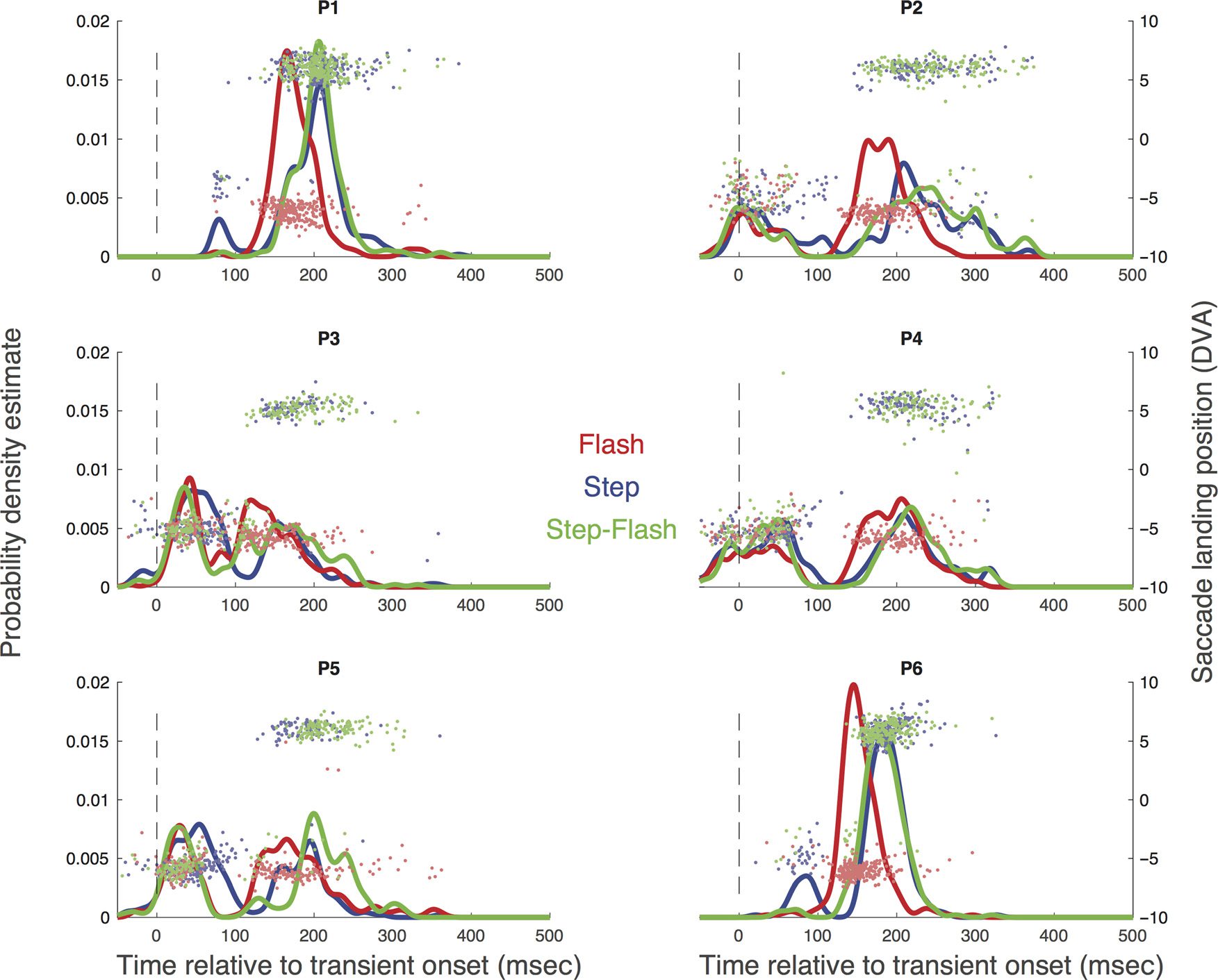
Figur 2. Each panel shows the probability density estimate derived from the raw data of each participant (P1–P6) in the distractor (Rot) and step
(Blau) conditions of Experiment 1. The horizontal components of the saccade landing positions are overlaid (negative values indicate Target 1
location, and positive values indicate Target 2 location). Time on the x axis is coded relative to transient onset (d.h., positive numbers indicate
saccades launched after distractor onset), rather than initial target onset to reveal the time course of responses to the transient change.
In distractor and flash conditions, Teilnehmer waren
required to respond to Target 1, so the expected pro-
bability is very high. Umgekehrt, in step and step-flash
Bedingungen, participants had to reorient to Target 2, mak-
ing the expected probability to Target 1 lower, to the
extent that the participant was able to reorient. We were
most interested in these proportions for step and step-
flash conditions in Experiment 3, in which lower num-
bers indicate more reliable reorienting (d.h., lower error
rate).
Reorienting Latency
For conditions involving a step, we also estimated the
average latency of successful reorienting responses, als
the median saccadic RT for saccades to Target 2, coded
relative to the target step.
Statistical Analysis
Experimente 1 Und 2 were designed as exploratory stud-
ies to assess whether the signature of saccadic inhibition
was present in a double-step context. There were no
critical hypotheses for these two experiments, so formal
statistical tests were not used. Experiment 3 was then
designed to test whether saccadic inhibition facilitated
reorienting. For this experiment, we ran a series of pair-
wise t-test comparisons to test differences among param-
eters of interest. Given the low number of participants
(N = 6), which makes parametric assumptions hard to
verify, we also ran nonparametric statistics. We report
the parametric analysis in the Results section, but all non-
parametric comparisons had similar outcomes and do
not modify our findings.
ERGEBNISSE
Experimente 1 Und 2 were designed to provide a descrip-
tive comparison of saccadic activity in distraction and
double-step tasks. We expected to observe the classic re-
duction in saccade frequency around 100 msec after the
transient event. Experiment 3 was designed to test the
influence of saccadic inhibition upon measures of re-
orienting and the influence of the reorienting instruction
Buonocore, Purokayastha, and McIntosh
2071
upon measures of inhibition. Formal statistical tests are
reported for Experiment 3 nur.
probability of Target 1 = 0.26, SD = 0.1), selbst nach
the maximum of inhibition.
Experiment 1
Figur 2 shows the saccadic RT profiles for each partici-
keuchen, showing the probability of saccade launching by
time after transient onset. Saccadic inhibition is indicated
by the bimodality of these distributions; its consistency
across participants is striking, with a depression of sac-
cadic activity centered around 100 msec after the tran-
sient change (Buonocore, McIntosh, & Melcher, 2016;
Buonocore & McIntosh 2008, 2012, 2013; Bompas &
Sumner, 2011; Edelman & Xu 2009; Reingold & Stampe,
1999, 2000, 2002, 2004). Even more striking is that this
bimodal pattern emerged regardless of whether the par-
ticipant was required to ignore or to reorient to a target
step. The data show, for the first time, that the disconti-
nuity of saccadic behavior in double-step reorienting
(Ludwig, Mildinhall, & Gilchrist, 2007; Findlay & Harris,
1984; Becker & Jürgens, 1979) closely matches the char-
acteristic time course of saccadic inhibition.
The horizontal components of the saccade landing
positions are overlaid in each panel of Figure 2. For the
step condition, these clouds of points cluster to Target 1
or Target 2 along the time dimension, reflecting the
double-step transition. Virtually no eye movements to-
ward Target 2 were launched before the start of the inhi-
bition. In all participants bar one (P02), reorienting was
permeated by erroneous saccades to Target 1 (mean
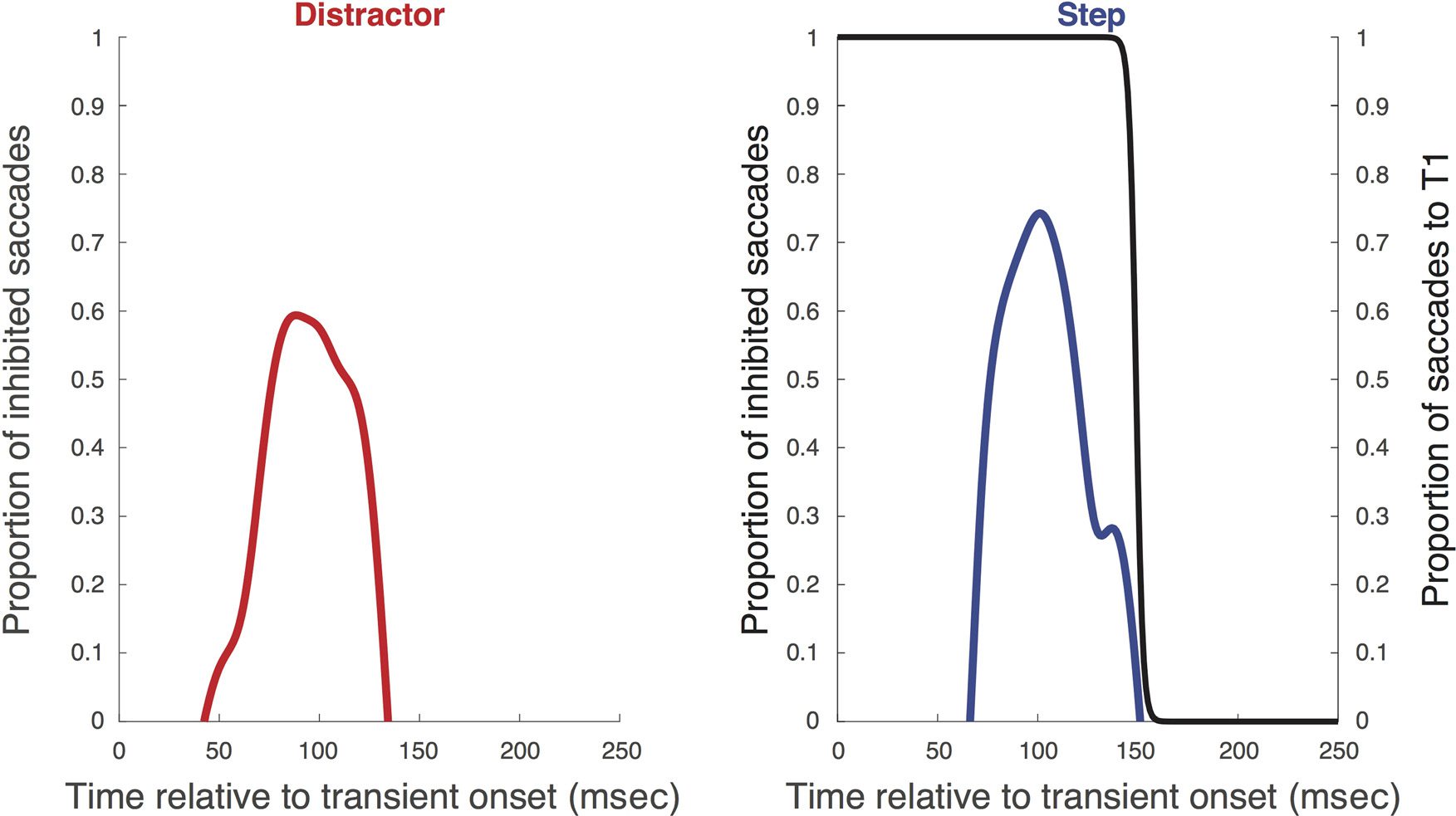
The group average time course of inhibition derived
from the saccadic inhibition analysis for each condition
is shown in Figure 3, and the individual parameters are
shown in Table 1. Inhibition starts around 81 ms, mit
saccadic activity driven almost to zero by 119 ms, Re-
turning to normal levels by 165 ms. The overlaid group
average direction transition function for the step condi-
tion implies that the transition is parallel to the time
course of saccadic inhibition, with its inflection point
on average ∼16 msec after the peak of inhibition (sehen
Tisch 2 for individual parameters).
Experiment 1 thus demonstrates saccadic inhibition in
a double-step context and further shows a temporal over-
lap of saccadic inhibition and reorienting. Experiment 2
asks whether the pattern holds under stimulus condi-
tions more similar to a classic distractor paradigm, In
which Target 1 does not disappear with the appearance
of Target 2 but persists (Buonocore & McIntosh, 2008;
Walker et al., 1995).
Experiment 2
Figur 4 shows the distributions for each participant,
with horizontal landing positions overlaid. As in Experi-
ment 1, there is clear impression of bimodality consistent
with saccadic inhibition in distractor and step conditions;
this is, if anything, more pronounced in the step condi-
tion. The landing positions show that erroneous saccades
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
/
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
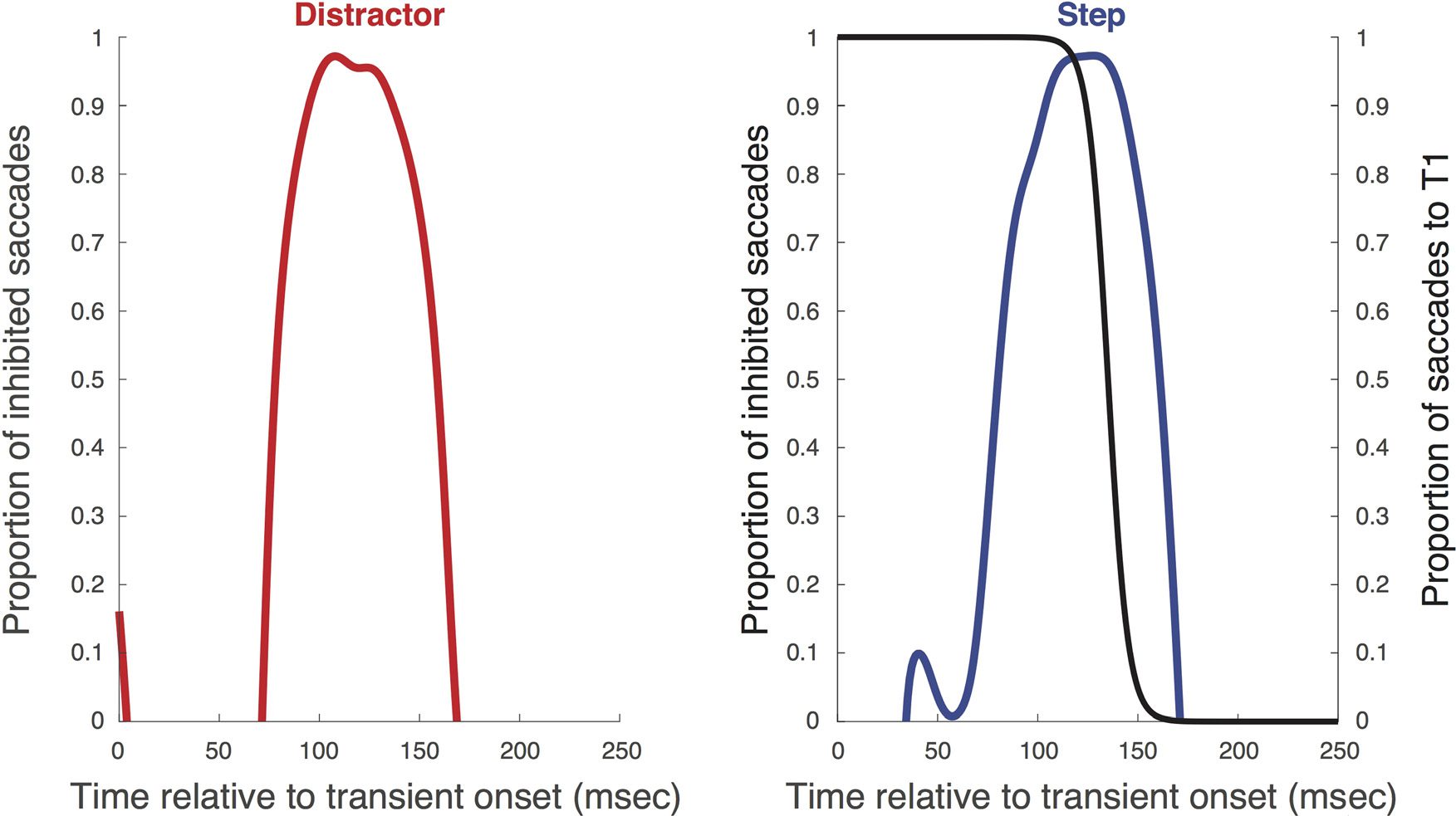
Figur 3. Group average time course of proportional inhibition derived from the saccadic inhibition analysis for distractor (Rot) and step (Blau)
conditions of Experiment 1. For the step condition, the black line represents the group average direction transition function coded as the proportion
of saccades directed to Target 1 (right y axis). Time on the x axis is coded relative to transient onset (d.h., positive numbers indicate saccades
launched after distractor onset), rather than initial target onset to reveal the time course of responses to the transient change.
2072
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 12
D
N
E
N
Ö
ich
T
ich
B
ich
H
N
ICH
T
R
A
T
S
N
Ö
ich
T
ich
B
ich
H
N
ICH
j
C
N
e
T
A
L
N
Ö
ich
T
ich
B
ich
H
N
ICH
e
D
u
T
ich
N
G
A
M
N
Ö
ich
T
ich
B
ich
H
N
ICH
H
S
A
l
F
–
P
e
T
S
P
e
T
S
R
Ö
T
C
A
R
T
S
ich
D
H
S
A
l
F
–
P
e
T
S
P
e
T
S
R
Ö
T
C
A
R
T
S
ich
D
H
S
A
l
F
–
P
e
T
S
P
e
T
S
R
Ö
T
C
A
R
T
S
ich
D
H
S
A
l
F
–
P
e
T
S
P
e
T
S
R
Ö
T
C
A
R
T
S
ich
D
T
N
A
P
ich
C
ich
T
R
A
P
T
N
e
M
ich
R
e
P
X
E
N
Ö
ich
T
ich
D
N
Ö
C
D
N
A
T
N
e
M
ich
R
e
P
X
E
H
C
A
E
R
Ö
F
S
e
l
ich
F
Ö
R
P
N
Ö
ich
T
ich
B
H
N
ICH
ich
C
ich
D
A
C
C
A
S
e
H
T
M
Ö
R
F
D
e
T
C
A
R
T
X
E
S
N
Ö
ich
T
A
ich
v
e
D
D
R
A
D
N
A
T
S
D
N
A
,
S
N
A
e
M
,
S
R
e
T
e
M
A
R
A
P
l
A
u
D
ich
v
ich
D
N
ICH
.
1
e
l
B
A
T
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
5
5
1
8
6
1
8
0
1
5
6
1
4
2
1
7
4
1
5
4
1
7
5
1
8
5
1
2
5
1
4
6
1
6
6
1
4
9
1
5
6
1
7
5
1
8
6
1
7
5
1
5
4
1
4
7
1
3
6
1
1
6
1
0
0
.
5
1
9
0
.
0
1
3
5
1
4
4
1
6
0
1
0
5
1
4
3
1
8
6
1
3
4
1
8
4
1
3
3
1
6
2
1
3
0
1
0
3
1
3
2
1
7
2
1
9
0
.
1
2
9
6
.
4
1
3
5
1
2
8
1
4
3
1
3
5
1
0
4
1
3
5
1
3
5
1
6
3
1
5
4
1
6
0
1
7
3
1
8
1
1
4
2
1
8
2
1
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
A
0
5
9
6
2
5
2
6
6
5
3
5
7
5
7
7
4
8
8
7
6
7
5
7
8
9
1
8
6
7
0
8
3
6
9
7
8
7
0
8
6
7
6
7
.
8
4
5
.
6
1
7
1
8
3
7
9
5
9
8
4
7
5
7
7
0
.
0
1
7
7
3
7
5
8
8
6
9
6
9
8
7
7
1
1
1
6
7
6
6
6
5
1
7
4
7
6
7
3
7
.
8
1
A
0
5
9
6
4
5
3
5
6
5
A
0
5
5
5
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
9
0
1
9
9
2
7
4
1
1
8
8
4
0
1
8
9
2
9
5
1
1
7
1
1
4
2
1
6
3
1
0
3
1
9
1
1
5
1
1
0
2
1
7
2
1
2
1
1
3
0
1
1
0
1
3
1
1
9
3
.
5
1
4
9
.
9
4
3
1
1
1
1
2
9
2
3
1
3
1
1
6
0
1
5
1
1
5
3
1
5
0
1
5
8
8
7
5
1
1
6
0
1
4
0
1
0
0
.
6
1
1
6
.
0
2
6
2
1
3
3
1
3
1
1
7
1
1
9
1
1
7
2
1
3
2
1
8
9
9
9
7
6
6
0
1
6
8
2
8
0
9
4
8
.
3
2
5
5
.
6
1
8
3
.
4
1
1
2
.
7
9
5
.
8
9
0
.
7
2
4
.
5
1
3
4
.
7
1
2
.
4
1
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
0
0
.
1
0
0
.
1
6
8
.
0
0
0
.
1
0
0
.
1
0
0
.
1
8
9
.
0
6
0
.
0
2
9
.
0
0
0
.
1
0
0
.
1
0
0
.
1
0
0
.
1
0
0
.
1
8
9
.
0
3
0
.
0
5
8
.
0
5
8
.
0
4
9
.
0
1
8
.
0
0
9
.
0
0
0
.
1
9
8
.
0
7
0
.
0
7
9
.
0
7
9
.
0
7
8
.
0
7
9
.
0
7
8
.
0
0
0
.
1
4
9
.
0
6
0
.
0
0
0
.
1
0
0
.
1
0
0
.
1
7
9
.
0
0
0
.
1
7
9
.
0
9
9
.
0
1
0
.
0
7
4
.
0
1
8
.
0
9
6
.
0
9
5
.
0
9
6
.
0
5
8
.
0
9
6
.
0
4
1
.
0
0
0
.
1
0
0
.
1
8
7
.
0
0
0
.
1
0
0
.
1
6
9
.
0
6
9
.
0
9
0
.
0
1
P
2
P
3
P
4
P
5
P
6
P
N
A
e
M
D
S
1
P
2
P
3
P
4
P
5
P
6
P
N
A
e
M
D
S
1
P
2
P
3
P
4
P
5
P
6
P
N
A
e
M
D
S
1
2
3
C
e
S
M
0
5
w
Ö
e
B
l
e
M
ich
T
T
R
A
T
S
N
Ö
ich
T
ich
B
H
N
ich
ich
N
A
D
e
T
u
P
M
Ö
C
M
H
T
ich
R
Ö
G
l
A
e
H
T
D
N
A
,
H
S
A
l
F
A
H
T
ich
w
S
N
Ö
ich
T
ich
D
N
Ö
C
e
H
T
N
ich
N
Ö
ich
T
u
B
ich
R
T
S
ich
D
e
D
A
C
C
A
S
e
H
T
F
Ö
T
R
A
P
T
S
e
ich
l
R
A
e
e
H
T
M
Ö
R
F
G
N
Ö
R
T
S
S
A
w
N
Ö
ich
T
ich
B
H
N
ich
ich
C
ich
D
A
C
C
A
S
,
3
T
N
e
M
ich
R
e
P
X
E
N
ich
6
P
R
Ö
F
A
.
N
Ö
ich
T
ich
D
N
Ö
C
H
S
A
l
F
e
H
T
Ö
T
S
D
N
Ö
P
S
e
R
R
Ö
C
3
T
N
e
M
ich
R
e
P
X
E
F
Ö
N
Ö
ich
T
ich
D
N
Ö
C
R
Ö
T
C
A
R
T
S
ich
D
e
H
T
T
B
u
Ö
D
Ö
N
S
ich
e
R
e
H
T
,
S
N
Ö
ich
T
ich
D
N
Ö
C
e
S
e
H
T
N
ich
1
P
R
Ö
F
N
ich
A
T
R
e
C
N
u
S
ich
e
M
ich
T
T
e
S
N
Ö
F
Ö
e
T
A
M
ich
T
S
e
e
S
ich
C
e
R
P
e
H
T
H
G
u
Ö
H
T
l
A
.
e
T
A
M
ich
T
S
e
l
l
A
C
ich
G
Ö
Ö
ich
S
j
H
P
Ö
R
u
e
N
e
B
ich
S
u
A
l
P
e
R
Ö
M
A
l
R
Ö
F
C
e
S
M
0
5
F
Ö
e
u
l
A
v
A
Ö
T
S
ich
H
T
D
e
C
R
e
Ö
C
e
w
;
)
C
e
S
M
1
4
,
.
e
.
ich
(
.
N
Ö
ich
T
ich
D
N
Ö
C
P
e
T
S
e
H
T
N
ich
N
Ö
ich
T
ich
B
H
N
ich
ich
F
Ö
T
e
S
N
Ö
e
H
T
N
A
H
T
R
e
ich
l
R
A
e
S
ich
T
ich
T
A
H
T
Buonocore, Purokayastha, and McIntosh
2073
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Tisch 2. Individual Parameters, Means, and Standard Deviations for Direction Transition Function (DTF) and Other Measures of
Reorienting Behavior for Each Experiment and Condition
Experiment
Participant
Schritt
Step-flash
Schritt
Step-flash Distractor
Schritt
Step-flash
Schritt
Step-flash
DTF Slope
DTF Inflection
Probability T1
Reorienting Latency
1
2
3
P1
P2
P3
P4
P5
P6
Mean
SD
P1
P2
P3
P4
P5
P6
Mean
SD
P1
P2
P3
P4
P5
P6
Mean
SD
−0.90
−0.09
−0.02
−0.06
−0.06
−0.06
−0.20
0.35
−0.05
−0.05
−3.30
−0.07
−0.05
−0.06
−0.60
1.32
−0.11
−0.86
−0.26
−1.74
−1.51
−1.49
−1.00
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
−1.34
−3.10
−0.14
−0.34
−1.01
−0.29
−1.04
88
108
152
149
152
162
135
29.69
165
152
109
145
138
185
149
25.71
116
135
126
136
121
127
127
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
108
154
124
115
152
146
133
0.69
1.11
7.80
19.91
0.89
0.91
0.80
0.94
0.69
0.90
0.86
0.10
0.94
0.92
0.92
0.99
0.98
0.99
0.96
0.03
1.00
1.00
1.00
1.00
0.99
1.00
1.00
0.01
0.26
0.36
0.37
0.12
0.18
0.28
0.26
0.10
0.38
0.33
0.23
0.37
0.37
0.29
0.33
0.06
0.10
0.44
0.65
0.51
0.61
0.14
0.41
0.24
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
0.01
0.34
0.51
0.44
0.47
0.08
0.31
0.21
208
217
222
220
223
251
223
14.34
205
199
162
188
199
243
199
26.30
220
256
188
239
203
202
218
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
219
278
211
250
242
198
233
25.62
29.31
to Target 1 (failures of reorienting) were relatively common
in the step condition (mean probability of Target 1 = 0.33,
SD = 0.06). Noch einmal, virtually no reorienting saccades
to Target 2 were launched before the start of inhibition.
In Abbildung 5, the group average transition function for
the step condition is overlaid on the average profile from
the saccadic inhibition analysis and alongside the inhi-
bition profile for the distractor condition. Da ist ein
broad superimposition of the temporal profiles of inhi-
bition and reorienting, and the point of inflection in
the transition function lags the peak of inhibition (von
∼34 msec). Jedoch, visual inspection suggests that
the transition function now extends further beyond the
end of the inhibition profile than in Experiment 1. Das
is not due to a change in the direction transition function,
which is closely similar between Experiments 1 Und 2, Aber
instead to alterations in the inhibition profile. Descriptively,
the magnitude of the inhibition is reduced in Experiment 2
(due to the persistence of Target 1), and this is associated
with a weaker and contracted inhibitory profile, a lower
maximum inhibition being reached relatively sooner, Und
an inhibition ending earlier (the numerical differences
between these parameters can be appreciated in Table 1).
Comparing inhibition profiles between the step and
distractor condition of Experiment 2 (Figur 5 Und
Tisch 1), the onset times are similar, but the maximum
inhibition is stronger (in every participant) and often
ends later (in five of six participants) in the step than in
the distractor condition of Experiment 2. This may sug-
gest that the bottom–up inhibition associated with the
target step accounts for only a proportion of its inhibitory
effect in the step condition, having its main influence in
2074
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
/
.
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
Ö
6
C
8
N
5
_
7
A
/
_
J
0
Ö
1
C
1
N
7
9
_
A
P
_
D
0
1
B
1
j
7
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
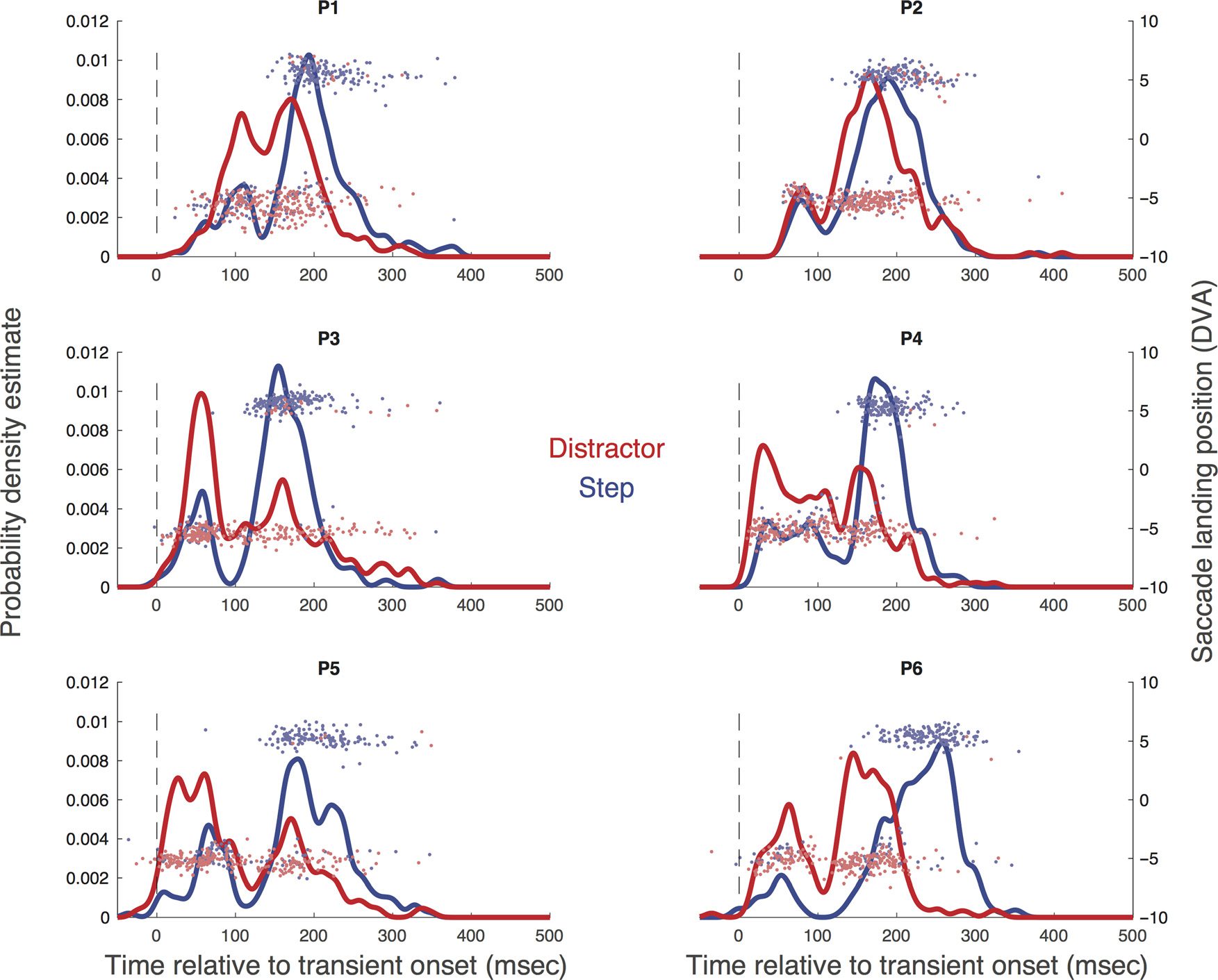
Figur 4. Each panel shows the probability density estimate derived from the raw data of each participant (P1–P6) in the distractor (Rot) Und
step (Blau) conditions of Experiment 2. The horizontal components of the saccade landing positions are overlaid. Time on the x axis is coded relative
to transient onset, rather than initial target onset, to reveal the time course of responses to the transient change.
the early part of the profile, and that the latter part of
the profile is additionally shaped by reorienting behavior.
Experiment 2 thus replicates the finding of saccadic in-
hibition in a double-step context, but additionally sug-
gests some possible dissociability from the time course
of reorienting.
Having demonstrated that saccadic inhibition occurs in
double-step tasks, Experiment 3 is designed to ask
whether inhibition plays a functional role in reorienting
behavior by testing the counterintuitive idea that re-
orienting behavior might actually benefit from additional
bottom–up interference, in the form a salient irrelevant
flash. Darüber hinaus, because Experiment 3 manipulates re-
flexive inhibition (the occurrence of flash) and voluntary
reorienting (the occurrence of a target step) without any
differences in task instruction, it provides a firmer basis
for assessing possible dissociations between the time
courses of inhibition and reorienting.
Experiment 3
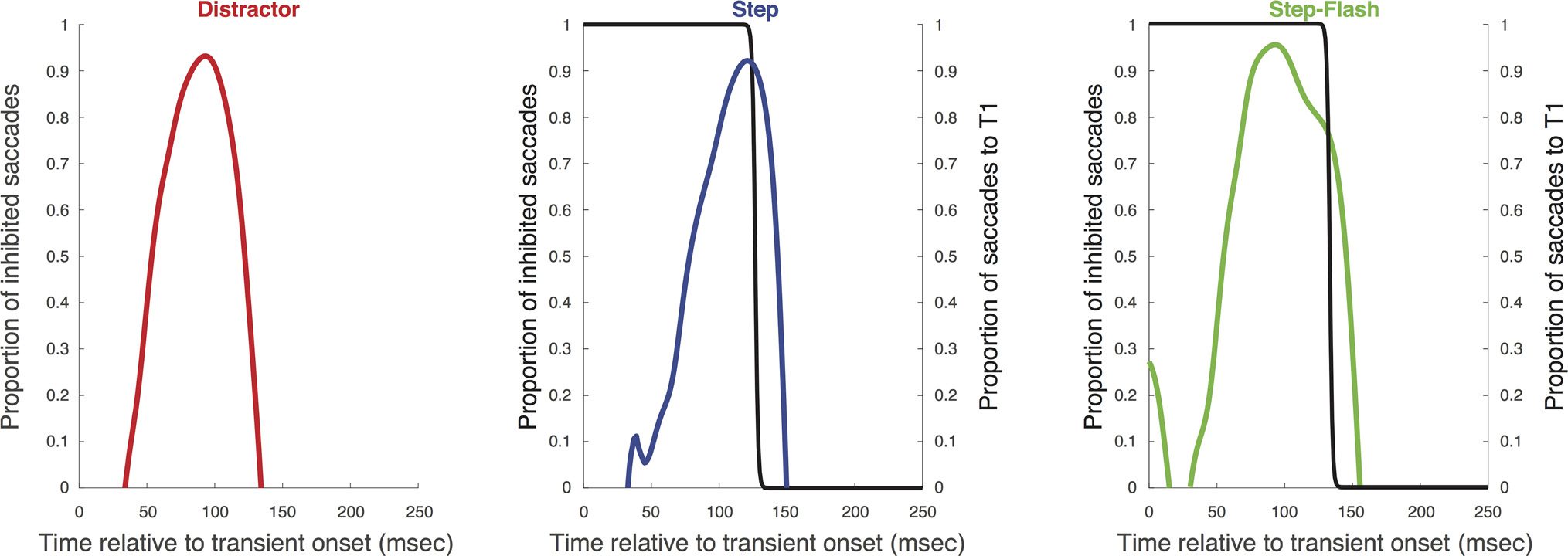
Figur 6 shows the distributions for each participant,
with horizontal landing positions overlaid. Da ist ein
clear reduction in saccadic activity for all the conditions,
within a common temporal window. Yet we can also ob-
serve consistent differences between conditions, welche
are clear from the inhibition profiles in Figure 7 Und
the individual parameters in Table 1. The latency of inhi-
bition was lower in the flash and step-flash conditions
than in the step condition, T(5) = 6.27, p = .002; T(5) =
4.47, p = .007, and inhibition began earlier in these con-
ditions, T(5) = 4.05, P < .01; t(5) = 3.48, p = .02. By con-
trast, inhibition ended later in the step and step-flash
conditions compared with the flash condition, t(5) =
7.57, p = .001; t(5) = 3.96, p = .01. These patterns strongly
support the idea that the total inhibitory period during
reorienting is a combination of an early bottom–up effect
(driven by the salient visual change) and a sustained
effect (driven by goal-related reorienting). The addition
of the flash to the step manipulation boosts the early
component, and the intention to reorient influences the
later component.
Because the addition of the flash shifts the start of
inhibition to an earlier moment, we can examine how
this impacts upon reorienting behavior. The first thing
to note is that there is no corresponding shift in the
Buonocore, Purokayastha, and McIntosh
2075
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
o
6
c
8
n
5
_
7
a
/
_
j
0
o
1
c
1
n
7
9
_
a
p
_
d
0
1
b
1
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Group average time course of proportional inhibition derived from the saccadic inhibition analysis for distractor (red) and step (blue)
conditions of Experiment 2. For the step condition, the black line represents the group average direction transition function (right y axis). Time on
the x axis is coded relative to transient onset (i.e., positive numbers indicate saccades launched after distractor onset), rather than initial target onset,
to reveal the time course of responses to the transient change.
direction transition function (average function in Figure 7;
individual parameters in Table 2): The time course of
reorienting is invariant across the shift in inhibition
onset. Table 2 similarly shows that the median latency
of reorienting to Target 2 was not reduced by the flash
(if anything, there was a tendency toward an in-
crease). This implies that saccadic inhibition does not
facilitate faster reorienting to Target 2, but this does not
mean that it has no functional benefit for reorienting
behavior.
The most crucial data are the probabilities of Target 1
responses in step and step-flash conditions (i.e., the error
rate), reflecting the overall proportion of saccades in
which reorienting was unsuccessful. The probability of
Target 1 responses is relatively high in the step condition
(mean = 0.41, SD = 0.24), but significantly reduced in
the step-flash condition (mean probability of Target 1 =
0.31, SD = 0.21), a difference that was seen in every par-
ticipant, t(5) = 6.78, p = .001. Reorienting thus becomes
more reliable when inhibition is boosted by the flash. We
infer that reflexive inhibition rapidly interrupts ongoing
activity, allowing a higher proportion of saccades to be re-
programmed for the new target, yet without speeding
their redirection.
DISCUSSION
Our experiments show that saccadic inhibition occurs in
typical double-step tasks and that it promotes successful
reorienting. In our critical third experiment, double-step
reorienting was improved by a distracting flash, coinci-
dent with the change in target position, even though
the flash carried no spatial information about the target.
However, although reoriented saccades were more likely
after a flash, we did not find that they took any less time
to emerge. Thus, oculomotor inhibition improves re-
orienting, but the benefit is quite specific: Inhibition helps
to countermand the planned response at short latency,
making an alternative response possible, but it does not
speed the generation of that alternative response.
The short-latency inhibitory effect of a visual change is
apparent across all conditions of our three experiments.
The onset of inhibition is insensitive to the task relevance
of a visual change but is modulated by its salience. In
Experiment 3, the onset of inhibition was thus earlier
in conditions with a salient flash (flash and step-flash con-
ditions) than in the simple target step condition, consis-
tent with a low-level reflexive response to the flash
impacting on a time scale close to the minimum neural
delays for the visual information to reach the superior
colliculus (Rizzolatti, Buchtel, Camarda, & Scandolara,
1980). By contrast, the late, recovery portion of the inhi-
bition profile was affected by the task context, with the
offset of inhibition relatively later in conditions requiring
reorienting, presumably because it takes longer to pro-
gram a new saccade than to restore a prior plan. So, re-
flexive inhibition facilitates reorienting by automatically
suppressing a planned saccade, but the generation of a
replacement response depends on participant intentions
and unfolds over a longer time scale.
2076
Journal of Cognitive Neuroscience
Volume 29, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
o
6
c
8
n
5
_
7
a
/
_
j
0
o
1
c
1
n
7
9
_
a
p
_
d
0
1
b
1
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 6. Each panel shows the probability density estimate derived from the raw data of each participant (P1–P6) in the flash (red), step (blue),
and step-flash (green) conditions of Experiment 3. The horizontal components of the saccade landing positions are overlaid. Time on the x axis is
coded relative to transient onset, rather than initial target onset, to reveal the time course of responses to the transient change.
These findings are consistent with a contemporary
model of double-step behavior, which suggests that re-
orienting cannot succeed simply by activation of a re-
placement response but also requires a STOP process
to countermand the initial planned response (Camalier
et al., 2007; see also Bissett & Logan, 2013). This “inde-
pendent horse race” model involves three processes: a
GO process (GO1) accumulating activation to respond
to the first target; a STOP process, triggered by the target
step, accumulating activation to countermand the GO1
response; and a second GO process (GO2), also trig-
gered by the target step, accumulating activation to re-
spond to the new target. Within this model, each
process develops independently of the others, except
that the STOP process inhibits the GO1 process if it
reaches threshold first. Note that this mutual indepen-
dence implies that a GO2 response will not be speeded
by a successful STOP process; it will just be more likely to
occur because GO1 has been withdrawn from the race.
Applying this model to the present context, saccadic
inhibition would be a rapidly rising STOP process, trig-
gered by the target step, which countermands the initially
planned (GO1) response whenever it reaches threshold
first; on these occasions, the GO2 response will sub-
sequently complete, and a reorienting saccade will follow.
In our Experiment 3, the addition of a large flash, simul-
taneous with the target step, would selectively boost the
STOP signal. This would raise the likelihood that the
STOP process achieves threshold before GO1, reducing
the frequency of Target 1 saccades and thereby increas-
ing the frequency of Target 2 saccades, yet without
speeding them ( just as we observed). We can similarly
apply the model to the differences between Experiments 1
and 2, in which all stimulus events were matched, except
that the initial target persisted after the step in Experiment 2.
This selective boosting of the GO1 process in Experiment 2
would make it less likely to be countermanded by STOP
and thereby less likely to be superseded by GO2, so that
inhibition would be reduced and reorienting less success-
ful (again as we observed). Saccadic inhibition seems like
a phenomenon ready-made for the role of STOP process
in this independent race model.
But, although functional independence of STOP and
GO processes may work within computational models of
double-step behavior, mutual independence seems less
plausible at the neurophysiological level. Reingold and
Buonocore, Purokayastha, and McIntosh
2077
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
o
6
c
8
n
5
_
7
a
/
_
j
0
o
1
c
1
n
7
9
_
a
p
_
d
0
1
b
1
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 7. Group average time course of proportional inhibition derived from the saccadic inhibition analysis for flash (red), step (blue), and step-
flash (green) conditions of Experiment 3. In the step and step-flash conditions, the black lines represent the group average direction transition
function (right y axis). Time on the x axis is coded relative to transient onset, rather than initial target onset, to reveal the time course of responses to
the transient change.
Stampe (2002) originally speculated that saccadic inhibi-
tion might arise from competitive interactions between
populations of neurons, within the motor maps of the
intermediate superior colliculus, building up activity for
saccades to target and distractor locations (Olivier, Dorris,
& Munoz, 1999). Dorris, Olivier, and Munoz (2007) sub-
sequently confirmed that preparatory activity for an ex-
pected saccade target is decreased transiently by the
onset of a distractor elsewhere in the visual field (nearby
distractors can conversely facilitate build-up activity).
Recently, a physiologically inspired model, incorporating
long-range inhibition and local facilitation, has been found
to simulate empirical patterns of saccadic inhibition with
impressive accuracy (Bompas & Sumner, 2011, 2015).
Superior colliculus neurons thus show competitive inter-
actions sufficient for causal involvement in saccade gener-
ation and countermanding (see also Paré & Hanes, 2003),
though extrinsic connections may also be involved, partic-
ularly with FEF (Brown, Hanes, Schall, & Stuphorn, 2008;
Hanes et al., 1998) and BG (Schmidt, Leventhal, Mallet,
Chen, & Berke, 2013; Hikosaka, Takikawa, & Kawagoe,
2000). In addition, a more direct inhibitory effect may in-
volve omnipause neurons in the brainstem, which gate
the activity of saccade burst neurons and show spikes of
activity time-locked to sudden visual changes (Missal &
Keller, 2002; Everling, Paré, Dorris, & Munoz, 1998). This
signal might delay the execution of a planned saccade,
creating a pause in behavior, during which competitive
interactions between target and distractor activity would
have time to play out. The saccadic inhibition phenome-
non might then result from a combination of competitive
integration within the superior colliculus and saccade
gating in the brainstem followed by top–down signals
from cortical areas such as the FEF (Peel, Hafed, Dash,
Lomber, & Corneil, 2016), promoting either reorienting
or a reinstatement of the original plan.
How can we reconcile evidence for neural interactions
among oculomotor STOP and GO signals, with an inde-
pendent race model of double-step behavior? This “neural
paradox” has been noted already within the response in-
hibition literature, and to address it, an interactive race
model was proposed, which allows interactions between
STOP and GO processes (Boucher, Palmeri, Logan, &
Schall, 2007; Boucher, Stuphorn, Logan, Schall, & Palmeri,
2007). This interactive model produces behavior equiva-
lent to that of the independent version, provided that the
influence of the STOP process is brief and potent (Boucher,
Palmeri, et al., 2007; see also Salinas & Stanford, 2013). This
would be consistent with the character of saccadic inhi-
bition, most of the latency of which is accounted for by
afferent delays, implying a brief and potent inhibitory inter-
action. Frameworks thus exist for modeling response inhi-
bition and response selection that allow underlying neural
interactions while giving the appearance of independent
processes at the behavioral level.
Conclusions
Saccadic inhibition is a reflexive phenomenon that fol-
lows whenever a visual change occurs while a saccade
is planned, even if the change is known to be irrelevant
(e.g., Buonocore & McIntosh, 2008; Reingold & Stampe,
2002). We suggest that it plays a crucial role in behavior,
acting as a bottom–up brake that buys the time needed
for voluntary processes to act. Reflexive inhibition follows
a visual change at latencies too brief for voluntary control;
the voluntary part is whether to subsequently recover the
original plan (distractor context) or to switch to an alter-
native (double-step context). In a rapid behavioral system
like the ocuolomotor one, saccadic inhibition may be a
ubiquitous pause signal that provides the flexibility for
voluntary behavior to emerge.
2078
Journal of Cognitive Neuroscience
Volume 29, Number 12
Acknowledgments
The authors are grateful to Eyal Reingold for discussion of the
ideas behind these experiments and to Ziad Hafed and Antje
Nuthmann for comments on an earlier draft of this paper. A. B.
and R. D. M. designed the experiments. A. B. and S. P. carried
out data collection. A. B. and R. D. M. analyzed and interpreted
the data, and drafted the paper. All authors approved the final
version of the paper for submission.
Reprint requests should be sent to Dr. Antimo Buonocore,
Werner Reichardt Centre for Integrative Neuroscience, Tübingen
University, Otfried-Müller-Str. 25, 72076 Tübingen, Germany,
or via e-mail: antimo.buonocore@cin.uni-tuebingen.de.
Note
1. Participant 6 in Experiment 1 completed only one session
per condition.
REFERENCES
Aslin, R. N., & Shea, S. L. (1987). The amplitude and angle of
saccades to double-step target displacements. Vision Research,
27, 1925–1942.
Becker, W., & Jürgens, R. (1979). An analysis of the saccadic
system by means of double step stimuli. Vision Research, 19,
967–983.
Bissett, P. G., & Logan, G. D. (2013). Stop before you leap:
Changing eye and hand movements requires stopping.
Journal of Experimental Psychology. Human Perception
and Performance, 39, 941–946.
Bompas, A., & Sumner, P. (2011). Saccadic inhibition reveals
the timing of automatic and voluntary signals in the human
brain. Journal of Neuroscience, 31, 12501–12512.
Bompas, A., & Sumner, P. (2015). Saccadic inhibition and the
remote distractor effect: One mechanism or two? Journal of
Vision, 15, 15.
Boucher, L., Palmeri, T. J., Logan, G. D., & Schall, J. D. (2007).
Inhibitory control in mind and brain: An interactive race
model of countermanding saccades. Psychological Review,
114, 376–397.
Boucher, L., Stuphorn, V., Logan, G. D., Schall, J. D., & Palmeri,
T. J. (2007). Stopping eye and hand movements: Are the
processes independent? Perception and Psychophysics, 69,
785–801.
Brown, J. W., Hanes, D. P., Schall, J. D., & Stuphorn, V. (2008).
Relation of frontal eye field activity to saccade initiation
during a countermanding task. Experimental Brain Research,
190, 135–151.
Buonocore, A., & McIntosh, R. D. (2008). Saccadic inhibition
underlies the remote distractor effect. Experimental Brain
Research, 191, 117–122.
Buonocore, A., & McIntosh, R. D. (2012). Modulation of
saccadic inhibition by distractor size and location. Vision
Research, 69, 32–41.
Buonocore, A., & McIntosh, R. D. (2013). Attention modulates
saccadic inhibition magnitude. Quarterly Journal of
Experimental Psychology, 66, 1051–1059.
Buonocore, A., McIntosh, R. D., & Melcher, D. (2016). Beyond
the point of no return: Effects of visual distractors on saccade
amplitude and velocity. Journal of Neurophysiology, 115,
752–762.
Camalier, C. R., Gotler, A., Murthy, A., Thompson, K. G., Logan,
G. D., Palmeri, T. J., et al. (2007). Dynamics of saccade
target selection: Race model analysis of double step and search
step saccade production in human and macaque. Vision
Research, 47, 2187–2211.
Dorris, M. C., Olivier, E., & Munoz, D. P. (2007). Competitive
integration of visual and preparatory signals in the superior
colliculus during saccadic programming. Journal of
Neuroscience, 27, 5053–5062.
Edelman, J. A., & Xu, K. Z. (2009). Inhibition of voluntary
saccadic eye movement commands by abrupt visual onsets.
Journal of Neurophysiology, 101, 1222–1234.
Everling, S., Paré, M., Dorris, M. C., & Munoz, D. P. (1998).
Comparison of the discharge characteristics of brain stem
omnipause neurons and superior colliculus fixation
neurons in monkey: Implications for control of fixation
and saccade behavior. Journal of Neurophysiology, 79,
511–528.
Findlay, J. M., & Harris, L. R. (1984). Small saccades to double-
stepped targets moving in two dimensions. Advances in
Psychology, 22, 71–78.
Findlay, J. M., & Walker, R. (1999). A model of saccade
generation based on parallel processing and competitive
inhibition. Behavioral and Brain Sciences, 22, 661–721.
Hanes, D. P., Ii, W. F. P., Schall, J. D., Doug, P., Patterson, W. F.,
& Schall, J. D. (1998). Role of frontal eye fields in
countermanding saccades: Visual, movement, and fixation
activity. Journal of Neurophysiology, 79, 817–834.
Hikosaka, O., Takikawa, Y., & Kawagoe, R. (2000). Role of
the basal ganglia in the control of purposive saccadic eye
movements. Physiological Reviews, 80, 953–978.
Lisberger, S. G., Fuchs, A. F., King, W. M., & Evinger, L. C.
(1975). Effect of mean reaction time on saccadic responses to
two-step stimuli with horizontal and vertical components.
Vision Research, 15, 1021–1025.
Ludwig, C. J. H., Mildinhall, J. W., & Gilchrist, I. D. (2007). A
population coding account for systematic variation in
saccadic dead time. Journal of Neurophysiology, 97,
795–805.
McIntosh, R. D., & Buonocore, A. (2014). Saccadic inhibition
can cause the remote distractor effect, but the remote
distractor effect may not be a useful concept. Journal of
Vision, 14, 15.
Missal, M., & Keller, E. L. (2002). Common inhibitory
mechanism for saccades and smooth-pursuit eye movements.
Journal of Neurophysiology, 88, 1880–1892.
Olivier, E., Dorris, M. C., & Munoz, D. P. (1999). Lateral
interactions in the superior colliculus, not an extended
fixation zone, can account for the remote distractor effect.
Behavioral and Brain Sciences, 22, 694–695.
Paré, M., & Hanes, D. P. (2003). Controlled movement
processing: Superior colliculus activity associated with
countermanded saccades. Journal of Neuroscience, 23,
6480–6489.
Peel, T. R., Hafed, Z. M., Dash, S., Lomber, S. G., & Corneil,
B. D. (2016). A causal role for the cortical frontal eye
fields in microsaccade deployment. PLoS Biology, 14,
e1002531.
Reingold, E. M., & Stampe, D. M. (1999). Saccadic inhibition
in complex visual tasks. In W. Becker, H. Deubel, &
T. Mergner (Eds.), Current oculomotor research (pp. 249–255).
Boston: Springer US.
Reingold, E. M., & Stampe, D. M. (2000). Saccadic inhibition
and Gaze Contingent Research Paradigms. In A. Kennedy,
R. Radach, D. Heller, & J. Pynte (Eds.), Reading as a
Perceptual Process (pp. 119–145). Oxford: Elsevier.
Reingold, E. M., & Stampe, D. M. (2002). Saccadic inhibition
in voluntary and reflexive saccades. Journal of Cognitive
Neuroscience, 14, 371–388.
Reingold, E. M., & Stampe, D. M. (2003). Using the saccadic
inhibition paradigm to investigate saccadic control in reading.
Buonocore, Purokayastha, and McIntosh
2079
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
o
6
c
8
n
5
_
7
a
/
_
j
0
o
1
c
1
n
7
9
_
a
p
_
d
0
1
b
1
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
In J. Hyönä, R. Radach & H. Duebel (Eds.), The Mind’s Eye
(pp. 347–360). Amsterdam: Elsevier.
Reingold, E. M., & Stampe, D. M. (2004). Saccadic inhibition
in reading. Journal of Experimental Psychology: Human
Perception and Performance, 30, 194–211.
Rizzolatti, G., Buchtel, H. A., Camarda, R., & Scandolara, C.
(1980). Neurons with complex visual properties in the
superior colliculus of the macaque monkey. Experimental
Brain Research, 38, 37–42.
Salinas, E., & Stanford, T. R. (2013). The countermanding task
revisited: Fast stimulus detection is a key determinant of
psychophysical performance. Journal of Neuroscience, 33,
5668–5685.
Schmidt, R., Leventhal, D. K., Mallet, N., Chen, F., & Berke, J. D.
(2013). Canceling actions involves a race between basal
ganglia pathways. Nature Neuroscience, 16, 1118–1124.
Sheliga, B. M., Brown, V. J., & Miles, F. A. (2002). Voluntary
saccadic eye movements in humans studied with a double-cue
paradigm. Vision Research, 42, 1897–1915.
Walker, R., Kentridge, R. W., & Findlay, J. M. (1995). Independent
contributions of the orienting of attention, fixation offset
and bilateral stimulation on human saccadic latencies.
Experimental Brain Research, 103, 294–310.
Weber, H., & Fischer, B. (1994). Differential effects of non-
target stimuli on the occurrence of express saccades in man.
Vision Research, 34, 1883–1891.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
2
1
0
2
6
/
8
2
1
0
9
6
5
8
3
/
2
1
9
8
7
8
o
6
c
8
n
5
_
7
a
/
_
j
0
o
1
c
1
n
7
9
_
a
p
_
d
0
1
b
1
y
7
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
2080
Journal of Cognitive Neuroscience
Volume 29, Number 12