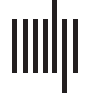REZENSION
A Review on Treatment-Related
Brain Changes in Aphasia
Klara Schevenels1
Bert De Smedt3
, Cathy J. Price2, Inge Zink1
, and Maaike Vandermosten1
,
Keine offenen Zugänge
Tagebuch
1Experimental Oto-Rhino-Laryngology, Department of Neurosciences, KU Leuven, Leuven, Belgien
2Welcome Centre for Human Neuroimaging, Institute of Neurology, University College London, Vereinigtes Königreich
3Parenting and Special Education Research Unit, Faculty of Psychology and Educational Sciences, KU Leuven, Leuven, Belgien
Schlüsselwörter: aphasia, brain plasticity, neuroimaging, intervention
ABSTRAKT
Numerous studies have investigated brain changes associated with interventions targeting a
range of language problems in patients with aphasia. We strive to integrate the results of these
studies to examine (1) whether the focus of the intervention (d.h., phonology, semantics,
orthography, syntax, or rhythmic-melodic) determines in which brain regions changes occur;
Und (2A) whether the most consistent changes occur within the language network or outside,
Und (2B) whether these are related to individual differences in language outcomes. The results
von 32 studies with 204 unique patients were considered. Concerning (1), the location of
treatment-related changes does not clearly depend on the type of language processing
targeted. Jedoch, there is some support that rhythmic-melodic training has more impact
on the right hemisphere than linguistic training. Concerning (2), we observed that language
recovery is not only associated with changes in traditional language-related structures in the
left hemisphere and homolog regions in the right hemisphere, but also with more medial and
subcortical changes (z.B., precuneus and basal ganglia). Although it is difficult to draw strong
conclusions, because there is a lack of systematic large-scale studies on this topic, this review
highlights the need for an integrated approach to investigate how language interventions
impact on the brain. Future studies need to focus on larger samples preserving subject-specific
Information (z.B., lesion effects) to cope with the inherent heterogeneity of stroke-induced
aphasia. Zusätzlich, recovery-related changes in whole-brain connectivity patterns need more
investigation to provide a comprehensive neural account of treatment-related brain plasticity
and language recovery.
EINFÜHRUNG
Aphasia is an acquired neurological language disorder affecting approximately 1 In 250 Menschen
(NIDCD, 2015). This is most commonly caused by a cerebrovascular accident in the language-
dominant hemisphere, which is the left hemisphere in more than 90% of right-handed persons
(Rasmussen & Milner, 1977). Aphasia results in impaired production and/or impaired compre-
hension of speech, reading, and/or writing. These communication impairments dramatically
affect societal participation and integration, causing a substantial decrease in the quality of life
(Dahlberg et al., 2006). Effective language treatment might be a crucial element to trigger
recovery. Different types of interventions can be used to target the language problems of people
with aphasia (PWA). Depending on the affected speech and language components, patients are
trained on the production and/or comprehension of the meaning of words and sentences
Zitat: Schevenels, K., Price, C. J.,
Zink, ICH., De Smedt, B., & Vandermosten,
M. (2020). A review on treatment-
related brain changes in aphasia.
Neurobiology of Language, 1(4),
402–433. https://doi.org/10.1162/
nol_a_00019
DOI:
https://doi.org/10.1162/nol_a_00019
zusätzliche Informationen:
https://doi.org/10.1162/nol_a_00019
Erhalten: 12 Juli 2019
Akzeptiert: 29 Juni 2020
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Klara Schevenels
Klara.Schevenels@kuleuven.be
Handling-Editor:
Steven Small
Urheberrechte ©: © 2020 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) Lizenz.
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
(semantics), the sound structure of words (phonology), written word forms (orthography), grammar
(morphosyntax), and/or the melodic intonation patterns inherent to language (melody/rhythm).
Other interventions tap communication as a whole (z.B., constraint-induced aphasia therapy;
Pulvermüller et al., 2001) and/or social activities and societal participation (z.B., script training;
Kaye & Cherney, 2016).
In the last two decades, research on the effectiveness of these therapies has increased, und hat
provided evidence that high dose speech-language therapy results in better functional commu-
nication and better language comprehension and production compared with no intervention.
Jedoch, effect sizes are weak, inconsistent, and not necessarily evident at follow-up (Brady,
Kelly, Godwin, Enderby, & Campbell, 2016). The effects of therapy have also been observed
in structural and functional alterations in the brain. It is known that experience, learning, Und
training can strengthen synapses through the frequent sequential coactivation of connected neu-
ronal assemblies (Tsumoto, 1992). This can consequently alter brain structure and functioning in
younger (Scholz, Klein, Behrens, & Johnsen-Berg, 2009) as well as older (Boyke, Driemeyer,
Gaser, Buchel, & Mai, 2008) healthy adults. Varley (2011), who translated known neuroscience
principles to aphasia therapy, described that such neuroplasticity should occur after stroke when
appropriate interventions, focusing on specific language behaviors, are provided with a sufficient
dose, frequency, and intensity (Pulvermüller & Berthier, 2008). Recovery is achieved by either
maximizing the capacity of a damaged neural language network or by linking new neural pro-
cessing assemblies to fulfill a linguistic task (Murphy & Corbett, 2009).
A central question is which regions are considered to belong to “the neural language net-
work.” For centuries, research has been conducted on investigating the neurobiological basis
of language. The familiarity of Broca’s and Wernicke’s regions in the context of language is the
result of the classic view on linguistic processing proposed by the “Broca–Wernicke–Lichtheim–
Geschwind model” in the late 19th century (Geschwind, 1965/2010, 1965, 1970). According to
this model, language is situated in the perisylvian area of the left hemisphere, more specifically in
the middle and posterior superior temporal lobe for language comprehension, and in the inferior
frontal lobe for language production. The connection between Broca’s and Wernicke’s regions is
established by the well-known white matter pathway, the arcuate fasciculus (AF; Hagoort, 2014).
Jedoch, there is still no clear and consistent definition of either of these two regions in terms of
anatomical localization. Over the years, Wernicke’s area has been located in almost every part of
the posterior perisylvian cortex, including the superior temporal gyrus (STG), the middle temporal
gyrus (MTG), and the inferior parietal cortex. In an online survey of specialists in the neurobiology
of language, none of the seven anatomical definitions of Wernicke’s region garnered more than
30% of the votes. Zusätzlich, in the same survey, nur 50% of respondents agreed on the precise
location of Broca’s region in the triangular and opercular part of the left inferior frontal gyrus (IFG;
Tremblay & Dick, 2016). Außerdem, the strict functional division between language produc-
tion in Broca’s region and language comprehension in Wernicke’s region is not valid because
many fMRI studies have demonstrated that both language modalities share neural resources
(Menenti, Gierhan, Segaert, & Hagoort, 2011; Segaert, Menenti, Weber, Petersson, & Hagoort,
2012; Stokes, Venezia, & Hickok, 2019).
In the last two decades, many alternative models for language have been proposed, of which
the dual-stream model for speech processing of Hickok and Poeppel (2007) is particularly well
known. Bei diesem Modell, multiple regions in the perisylvian cortex, as well as a premotor region and
more ventrally located areas, are assumed to underlie linguistic processing. Genauer, Die
first step in speech perception, the spectrotemporal analysis of the sounds, is assigned to the
dorsal STG. Subsequently, phonological processing takes place in the mid-post superior temporal
Sulkus (STS). The model then proposes a left dorsal stream, underlying the mapping of
Neurobiology of Language
403
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
phonological representations onto articulatory representations, in the parietotemporal junction,
the posterior IFG, and a more dorsal premotor region. A ventral stream underlies the mapping of
phonological representations onto meaning. Bilateral posterior regions in the ventral stream
(posterior MTG and inferior temporal gyrus [ES G]) engage more in lexical semantics, wohingegen
the left anterior regions of the ventral stream are also engaged in sentence level processing
(Hickok & Kacke, 2007).
Other recent language models have proposed an even more extended network, einschließlich
medial and subcortical structures involved in sensory, motor, and higher-order cognitive processes
that support linguistic functioning (z.B., Price, 2000, 2012; Price, Seghier, & Leff, 2010; Vigneau
et al., 2006, 2011). The specific role of each of these regions according to these more elaborate
language models is provided in Table 1 in the online supporting information located at https://
www.mitpressjournals.org/doi/suppl/10.1162/nol_a_00019. Over time, various research groups
have studied the healthy neural language network, and different subnetworks have been detected
to support different linguistic (semantics, phonology, syntax, orthography) and rhythmic-melodic
levels of language (Friederici, 2011; Price, 2000, 2010, 2012; Vigneau et al., 2006, 2011). Yet little
is known on treatment-related brain changes in these networks in PWA.
Aims of the Present Study
In this review, we strive to summarize and integrate the results of recent research on the structural
and/or functional changes associated with different language treatments in PWA. The aim is
twofold, namely to investigate whether brain changes are (1) specific to the type of intervention
received (d.h., phonological, semantic, orthographic, syntactic, or rhythmic-melodic); Und (2A)
whether the most consistent changes occur within the language network or outside, Und (2B)
whether these are related to individual differences in language outcomes. Concerning the first
aim, we first discuss whether the linguistic interventions (d.h., phonology, semantics, orthography,
or syntax) result in specific or similar brain changes. Nächste, we discuss whether nonlinguistic
interventions focusing on rhythmic-melodic aspects rely more on the right hemisphere than
linguistic-based interventions. As indicated in the model of Hickok and Poeppel, as well as in
Tisch 1 in the online supporting information, different brain correlates have been related to each
of the linguistic components (Hickok & Kacke, 2004, 2007; Price, 2000, 2010, 2012). Es ist
therefore plausible that, depending on the intervention, different subnetworks undergo changes
im Laufe der Zeit. These differences in intervention effects are assumed to be partly responsible for the
extreme heterogeneity in recovery patterns seen in PWA (Saur & Hartwigsen, 2012). Jedoch, bei
die selbe Zeit, these linguistic components are highly interwoven. This makes it unlikely that only
one kind of language processing is tapped during an intervention (z.B., one cannot train sentence
production without involving the meaning of the sentence). Zusätzlich, there is considerable
overlap in the neural networks for different linguistic components (see e.g., Vigneau et al., 2006)
and each network presumably interacts with others to create our general language behavior.
To illustrate this, we overlaid the fMRI association test maps for semantic (Blau), phonological
(Rot), syntactic (orange), as well as orthographic (Grün) processing in Figure 1, basierend auf
automated meta-analysis of previous fMRI studies provided by Neurosynth (http://neurosynth.
org/). The association maps represent brain regions where blood oxygen level dependent
(BOLD) changes occur more consistently for studies including the search term, than for studies
that do not mention the search term. According to Neurosynth, the overlap is the greatest in the
left frontal and temporal lobe, with most of the phonological network (Rot) located dorsally, most
of the semantic network (Blau) located ventrally, and most of the orthographic network (Grün)
located ventrally and posterior to the other language networks.
Neurobiology of Language
404
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Figur 1. Association test maps for the keywords “semantic” (Blau, 1,031 Studien), “phonological” (Rot, 377 Studien), “syntactic” (orange, 169
Studien), and “orthographic” (Grün, 132 Studien) Verarbeitung, according to the meta-analysis of Neurosynth (http://neurosynth.org/). First row:
left view, zweite Reihe: right view. The figures were composed using Paraview software (Ausführung 5.4.1; https://www.paraview.org/) following
the guidelines specified in Madan (2015).
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Based on these Neurosynth maps, we expected that (1) brain changes related to interventions
focused on phonological processing are more frequently located in the phonological network
(Rot), (2) brain changes related to interventions focused on semantic processing are more
frequently located in the semantic network (Blau), (3) brain changes related to interventions on
orthographic processing are more frequently located in the orthographic network (Grün), Und (4)
brain changes related to interventions focused on syntactic processing are more frequently
located in the syntactic network (orange). For all these language interventions, we expected a
left-dominance of treatment-related changes. This does not imply that the right hemisphere is
not involved in linguistic processing (z.B., see the bilateral ventral stream in Hickok &
Kacke, 2007), but only suggests that it does so to a lesser extent than the left hemisphere.
The rhythmic-melodic network is not depicted in Figure 1, because there was no meta-
analysis available on Neurosynth for this keyword. Jedoch, it similarly involves brain regions
in frontal and temporal lobes (siehe Tabelle 1 and Figure 1 in the online supporting information), but it
is generally considered the only component relying more on the right hemisphere than on the left
hemisphere (Baum & Pell, 1999). Daher, we expected more reorganization in the right hemisphere
after an intervention on the musical elements of speech, compared with the other linguistic
components (d.h., phonology, semantics, orthography, and syntax), which preferentially target
the left hemisphere. We also compared the neural effects of interventions targeting the right
versus the left hemisphere. Our hypothesis was that brain changes related to interventions
focused on rhythmic-melodic processing are more frequently located in frontal and temporal
lobes of the right hemisphere.
Concerning the second goal, we first describe whether the most consistent treatment-related
changes across therapies occur within the language network (d.h., a combination of response
maps visualized in Figure 1) or outside (aim 2a). Although the focus of research on brain–
language relations in aphasia recovery has been language centered, Cahana-Amitay and
Neurobiology of Language
405
A review on treatment-related brain changes in aphasia
Albert (2015) argued in their review that other cognitive functions, Zum Beispiel, attention, short-
term memory, and cognitive control, also contribute to aphasia recovery. Zusätzlich, more recent
models on the neurobiology of language also have considered brain regions that are involved in
multiple other functions (Price, 2000, 2010, 2012). daher, we hypothesized that the observed
treatment-related brain changes are not restricted to regions classically associated with linguistic
Verarbeitung, but involve a variety of brain structures associated with nonlinguistic cognitive
functions.
Zusätzlich, we explored whether and how the most consistent treatment-related changes are
associated with individual differences in language outcomes (aim 2b). In accordance with
findings in Saur et al. (2006), we expected that, at least in the chronic phase post-stroke, normal-
ization of activity to the left hemisphere is most associated with language improvement (restora-
tion). Jedoch, because in patients with extensive lesions the left-hemispheric recovery potential
is limited, associations between language improvement and brain changes in right-hemispheric
regions are expected as well (compensation). For the second aim, we considered treatment-
related regional changes as well as changes in connectivity patterns.
REVIEW METHOD
Inclusion and Exclusion Criteria
We searched three databases: Pubmed (https://pubmed.ncbi.nlm.nih.gov/), Embase (https://
www.embase.com), and Web of Science (https://www.webofknowledge.com) for studies
exploring neuroanatomical and/or functional changes in patients with aphasia due to specific
language interventions, published between January 2000 and April 2018. Genauer, In
the keywords, we combined three main concepts, das ist, (1) aphasia, (2) brain changes, Und
(3) intervention, using different words for each concept (Tisch 1). If all three concepts were
present in the title and/or the abstract, the article was included for further consideration. Von
Tisch 1.
Keyword combinations used to search for intervention studies in patients with aphasia
Concept 1
aphasia
conduction aphasia
Concept 2
brain changes
brain plasticity
transcortical sensory aphasia
neural changes
Bildgebung
wernicke
broca
Concept 3
intervention
therapy
treatment
Ausbildung
plastic change*
computer-assisted therapy
transcortical motor aphasia
neuroimaging
treatment-induced
anomia
dysphasia
anatom*
neurobiolog*
rehabilit*
language impairment
neural reorganization
language disorder
Notiz. The asterisks represent wildcards and can be replaced by one or more characters (z.B., the search term
anatom* will look for terms anatomical, anatomy, usw.).
Neurobiology of Language
406
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
screening the reference list of the so-collected articles, relevant papers published after 2000
were additionally added.
We established the following inclusion criteria. Erste, the patients had to be adults (to ensure that
language and brain development were complete), who had been diagnosed with aphasia as a
consequence of a cerebral vascular accident (d.h., stroke). Zweite, the study had to statistically
evaluate the effect of the treatment using measures collected through functional or structural
neuroimaging. We decided not to exclude studies on the basis of MRI modality, given that
training-induced neuroplasticity can be reflected in functional cortical changes as well as struc-
tural white matter changes, and that changes in each modality are related to each other (Honey
et al., 2009). Dritte, each therapy investigated had to focus specifically on one, maximally two,
linguistic domain(S): semantics, phonology, syntax, orthography, and/or melody/rhythm, to enable
the identification of brain changes after training of these specific types of language processing (aim
1). Aus diesem Grund, studies providing mixed conventional therapy (z.B., Aerts et al., 2015), intention
treatment (z.B., Benjamin et al., 2014), action observation treatment (z.B., Gili et al., 2017), inter-
ventions on the activity/participation level, imitation therapy (z.B., Santhanam, Duncan, & Small,
2018), script training (z.B., Fridriksson, Hubbard, et al., 2012), or constraint-induced language
therapy (z.B., McKinnon et al., 2017) were not considered. We also excluded studies that com-
bined language therapy with noninvasive brain stimulation and/or drug trials, intervention studies
in bilingual aphasia, non-peer-reviewed reports, and studies that were not available in English.
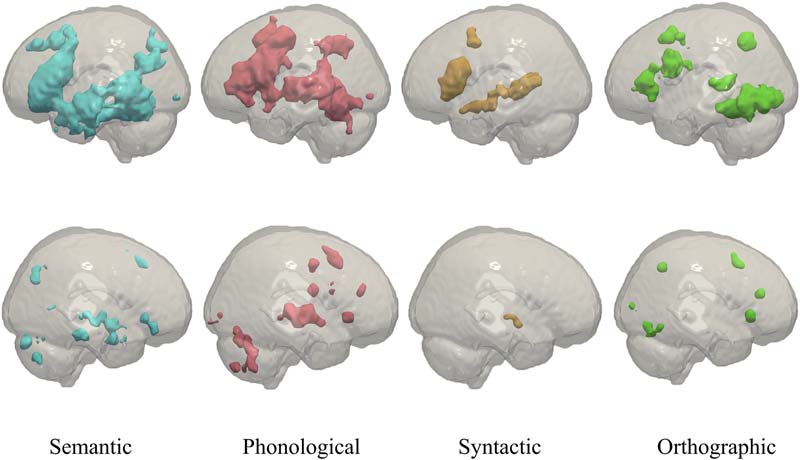
Figur 2 represents the literature search process.
Characteristics of the Studies Included
In Summe, we identified 32 studies on treatment-related brain changes in PWA that met the in-
clusion criteria. All references are listed per imaging modality, in-scanner task, and targeted
linguistic component in Table 2. For the sake of brevity, we refer to Appendix A for details on
each study and to the Supplementary Information for in-depth information on the specific
interventions that each study used. (Both can be found in the online supporting information
for this article.)
No specific constraint was set on the time post-stroke, Aber 94% of the studies (30 out of 32)
included only patients who were in the chronic stage (≥6 months) post-stroke, to avoid the
confounding effects from spontaneous recovery. The remaining two studies included PWA
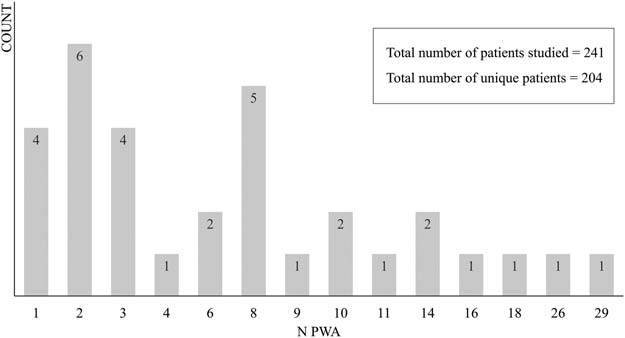
who were at least 4 months post-stroke. Figur 3 shows the number of studies (counts, on the
y-axis) with different numbers of participants (PWA, on the x-axis). In Summe, 11 studies shared
participants with one or two other studies. Seven of these studies were considered separately
because they applied different, and mostly unrelated, Analysen, das ist, task-based fMRI versus
resting-state fMRI versus diffusion-weighted imaging (DWI; van Hees et al., 2014; van Hees
et al., 2014A, 2014B), voxel-wise whole-brain contrast analysis versus region of interest (ROI)
based effective connectivity analysis (Vitali et al., 2007; Vitali et al., 2010) and univariate versus
multivariate fMRI analysis (Fridriksson, 2010; Fridriksson, Richardson, et al., 2012). Two studies
were considered separately because there was only minimal overlap in participants and different
interventions were considered (Fridriksson, Morrow-Odom, Moser, Fridriksson, & Bayliss, 2006;
Fridriksson et al., 2007). Endlich, the studies of Abel, Weiller, Huber, and Willmes (2014) Und
Abel, Weiller, Huber, Willmes, and Specht (2015) were considered as one study in this review,
because the same voxel-wise whole-brain contrast analysis has been reported in both studies.
More details for each study concerning the rationale behind these decisions can be found in
the online supporting information. In Summe, 204 unique patients with aphasia were tested in the
articles within the scope of this review.
Neurobiology of Language
407
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 2. PRISMA flowchart representing the literature search for studies of brain changes in patients with aphasia.
Variability Across Included Studies
Differences across studies in method, Teilnehmer, modality, Aufgabe, and contrasts
To collect a sufficient number of studies on the topic of treatment-related brain changes in PWA,
we included both studies reporting standard spatial coordinates for brain regions showing
treatment-related neural plasticity and studies that only reported anatomical labels for these
Regionen. Für unsere Analyse, we used the anatomical labeling according to the AAL-VOI atlas
(Tzourio-Mazoyer et al., 2002). If the study reported standard stereotaxic coordinates, Die
anatomical labels for these coordinates were derived from the atlas. If the study did not report
Neurobiology of Language
408
A review on treatment-related brain changes in aphasia
Tisch 2.
the linguistic component that is targeted during the intervention
Selected studies of treatment-related brain changes in patients with aphasia, with applied imaging modality, in-scanner task, Und
Study
Modality
In-scanner task
NEIN.
1
2
3
4
5
6
Abel, Weiller, Huber, and Willmes (2014)
Cornelissen et al. (2003)
Fridriksson et al. (2007)
Kiran, Meier, Kapse, and Glynn (2015)
Marcotte and Ansaldo (2010)
Marcotte, Perlbarg, Marrelec,
Benali, and Ansaldo (2013)
fMRT
MEG
fMRT
fMRT
fMRT
fMRT
7, 8
van Hees, McMahon, Angwin, de Zubicaray,
rs-fMRI
and Copland (2014), van Hees et al. (2014B)
9
1
10
3
11
12
13
14
15
Sandberg, Bohland, and Kiran (2015)
Abel et al. (2014)
Brownsett et al. (2014)
Fridriksson et al. (2007)
Fridriksson, Morrow-Odom, Moser, Fridriksson,
and Baylis (2006)
Haldin et al. (2018)
Leonard et al. (2015)
Marcotte et al. (2018)
Rochon et al. (2010)
fMRT
fMRT
fMRT
fMRT
fMRT
fMRT
fMRT
fMRT
fMRT
7, 8
van Hees et al. (2014A, 2014B)
rs-fMRI
16, 17
Vitali et al. (2007, 2010)
18
Nardo, Holland, Leff, Price, and Crinion (2017)
1, 19
Abel et al. (2014, 2015)
20
21
Fridriksson (2010)
Fridriksson, Richardson, et al. (2012)
fMRT
fMRT
fMRT
fMRT
fMRT
8, 22
van Hees et al. (2014A, 2014B)
rs-fMRI, DWI
23
25
25
26
27
Menke et al. (2009)
Raboyeau et al. (2008)
Thompson, den Ouden, Bonakdarpour,
Garibaldi, and Parrish (2010)
Thompson, Riley, den Ouden,
Meltzer-Asscher, and Lukic (2013)
Wierenga et al. (2006)
fMRT
HAUSTIER
fMRT
fMRT
fMRT
ON
ON
ON
ON, SFV
ON, VN
ON
ON
WJ
ON
SL, SR
ON
ON
RD, SYR
SJ, RJ
ON
SJ, RJ
ON
ON
ON
ON
ON
ON
NA
ON
ON
SPM
VN
SG
Targeted linguistic component
semantics (sem, N = 9)
phonology (phon, N = 13)
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
semantics-phonology
(sem+phon, N = 6)
phonology-orthography
(phon+orth, N = 2)
semantics-syntax
(syntax, N = 3)
Neurobiology of Language
409
A review on treatment-related brain changes in aphasia
Tisch 2.
(Fortsetzung )
NEIN.
28
29
30
31
32
Study
Modality
In-scanner task
Jungblut, Huber, Mais, and Schnitker (2014)
Schlaug, Marchina, and Norton (2008)
Schlaug, Marchina, and Norton (2009)
Tabei et al. (2016)
Wan et al. (2014)
fMRT
fMRT
DWI
fMRT
DWI
RCV
RSW
NA
ON
NA
Targeted linguistic component
rhythm-melody (r − m, N = 5)
Notiz. Some studies target multiple linguistic components and are therefore repeated. Zum Beispiel, several of the identified neuroimaging studies provide a
semantic treatment alternated with a phonological treatment. Jedoch, they do not always differentiate between the two types of intervention when reporting
BOLD changes or they additionally report general BOLD changes over the course of both treatments. The results of these kinds of studies are therefore listed
under sem (separate results for the semantic treatment), phon (separate results for the phonological treatment), and sem+phon (mixed results after both treat-
gen). rs-fMRI = resting-state fMRI, DWI = diffusion-weighted imaging, PET = positron emission tomography, ON = object naming, SFV = semantic feature
verification, VN = verb naming, WJ = word judgment, SL = sentence listening, SR = sentence repetition, RD = rhyme detection, SYR = syllable repetition, SJ =
semantic judgment, RJ = rhyme judgment, SPM = sentence-picture matching, SG = sentence generation, RCV = repetition of chanted vowel changes, RSW =
repetition of spoken/sung words, NA = not applicable, BOLD = blood oxygen level dependent.
standard stereotaxic coordinates, the anatomical labels of the study were adopted if the labels
corresponded to one of the labels in the atlas. If studies reported labels that did not correspond
to a label in the atlas (z.B., [dorsolateral] prefrontal cortex), we assigned the region to one of the
labels in the atlas where possible (z.B., middle frontal gyrus), or reported the labels in addition to
the atlas labels (which was the case for the inferior frontal sulcus and the premotor area). Es gibt
two disadvantages to this choice. Erste, this review is descriptive because an insufficient number
of studies reported standard spatial coordinates to support a quantitative meta-analysis. Das, In
combination with our choice to focus only on intervention studies targeting maximally two
linguistic domains, restricted the number of studies for a meta-analysis. Zweite, the spatial
resolution is reduced because it was not always possible to be sure which parts of an anatomical
label were being referred to.
Even when studies used the same imaging modality (z.B., DWI or fMRI), there were substantial
differences in methodology (for review, see Crosson et al., 2007). These include differences in
fMRI tasks, research design (group study vs. multiple single-subject study), or analysis method
(whole-brain vs. ROI analysis). Functional neuroimaging was used in 29 papers to investigate
treatment-related changes in the brain, of which 26 applied task-based fMRI, one acquired
resting-state fMRI, one applied positron emission tomography (HAUSTIER), and one applied MEG
Figur 3. The number of studies (counts) with numbers of participants. PWA = patients with aphasia.
Neurobiology of Language
410
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
used with MRI for source localization. Of these functional neuroimaging studies, 62% explored
functional changes during an object-naming task (18 out of 29), and five of the included studies
additionally concentrated on treatment-related changes in neural connectivity patterns. Endlich,
three papers applied a structural neuroimaging method, das ist, DWI, investigating either local
(one study) or distributed white matter changes (two studies). Although Siegel, Ramsey, et al.
(2016) demonstrated in a cohort of 132 stroke patients that language relied on both highly local-
ized brain regions, as well as on bilateral brain networks and their connections, überraschenderweise, In
total only seven studies focused on treatment-related changes in neural connectivity patterns
(Kiran, Meier, Kapse, & Glynn, 2015; Marcotte et al., 2013; Sandberg, Bohland, & Kiran, 2015;
Schlaug, Marchina, & Norton, 2009; van Hees et al., 2014A, 2014B; Vitali et al., 2010). Wichtig,
five of the seven connectivity studies used an ROI-approach in which the ROIs were chosen
based on previous literature or a healthy control group (Marcotte et al., 2013; van Hees et al.,
2014B; Vitali et al., 2010; Schlaug et al., 2009). This might have induced bias towards the
language network.
When comparing different functional imaging studies, the contrast used to identify a neural
response pattern will naturally determine the voxels associated with a specific condition-
dependent effect. Some studies applied lenient contrasts, Zum Beispiel, overt picture naming
versus rest (z.B., Cornelissen et al., 2003), while others applied very stringent contrasts, solch
as overt picture naming versus saying “baba” to digitally distorted nonsense images (z.B.,
Marcotte & Ansaldo, 2010; Marcotte et al., 2012). This makes comparison of studies difficult,
because the use of a lenient contrast will identify brain regions involved in a very wide range of
Verarbeitung (from lower to higher level). Zusätzlich, across studies, the statistical threshold applied
to these contrasts of interest varied. This threshold was sometimes not reported (Marcotte &
Ansaldo, 2010; Vitali et al., 2007), and frequently, it was not corrected for multiple comparisons
(Abel et al., 2014, 2015; Haldin et al., 2018; Kiran et al., 2015; Marcotte et al., 2013, 2018; Menke
et al., 2009; Thompson, Riley, den Ouden, Meltzer-Asscher, & Lukic, 2013; Vitali et al., 2010;
Wan et al., 2014). This again complicates the comparison of response foci across studies.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
Correct versus incorrect language behavior
The studies included differ in whether they included incorrect and/or absent language behavior in
their analysis or not (see Appendix A, column “extra,” in the online supporting information).
Some studies included all responses in their analysis (correct, incorrect, and no-response items),
while others contrasted trained items with correct items pretreatment (trained items > correctly
named items pretreatment), as well as incorrect items pretreatment (trained items > incorrectly
named items pretreatment). In the latter case, we only included the results of the contrast with
correct naming pretreatment. This is because it is assumed that incorrect (Sprache) behavior has
a different neural signature (Meinzer et al., 2013) and activates an error network in the brain,
including for example the anterior cingulate cortex (Stevens, Kiehl, Pearlson, & Calhoun,
2009), rather than the processing of interest (Price, Crinion, & Friston, 2006). Jedoch, es ist
important to note that this creates a source of variability between studies, as not all of them
disambiguated correct and incorrect trials in their analyses. (For a more elaborate discussion of
this matter, see Crosson et al., 2007 and Meinzer et al., 2013.)
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Direction of brain change
Some papers found both upregulation and downregulation of neural activity—in the same brain
regions—in different subjects who went through the same language intervention. Increased brain
Aktivität, in intensity or extent, might reflect the restoration of neural activity in the perilesional
Neurobiology of Language
411
A review on treatment-related brain changes in aphasia
language areas, the engagement of homolog language regions, or compensatory strategies that
involve brain regions that are not traditionally associated with language (Cabeza, Anderson,
Locantore, & McIntosh, 2002). Increases in brain activity could, andererseits, also point
to inefficient use of neural resources or increased effort when performing language tasks
(Fridriksson & Morrow, 2005). Im Gegensatz, reduced brain activity accompanied by behavioral
improvement could represent increased efficiency in the use of regions (Wierenga et al.,
2006), consistent with the effect of practice during skill acquisition (for meta-analysis, sehen
Chein & Schneider, 2005). Decreased activity could alternatively point to persistent malfunction-
ing, disconnection, or missing input due to the brain damage. Likewise, different recovery mech-
anisms might also occur simultaneously (Abel et al., 2015). Somit, it is important to relate the
BOLD changes to behavioral changes, or to compare them with a healthy control group to inter-
pret them correctly. If there is no relation between brain and behavior, the BOLD changes are
hard to interpret, and all that can be generated are hypotheses to be tested in future research. Für
these reasons, we will not make a difference between upregulation and downregulation of neural
activity for the fMRI results in this review, but generally refer to “changes” in the BOLD signal. Der
difference between increases and decreases in BOLD signal will only be considered below in
Association with language outcomes, where we discuss how the brain changes are related to
behavior, and the interpretation of the results.
Treatment-related brain changes
A general problem in neuroimaging reviews is the substantial variability in individual brain
reorganization patterns—both within and across studies—which makes comparisons between
studies very challenging. Throughout the review, we primarily referred to treatment-related brain
changes, which encompassed neural plasticity in the language network, in homologous areas of
the right hemisphere, or alternatively, the recruitment of supporting neural infrastructure (z.B.,
due to a strategy change) or brain dynamics related to (changes in) error processing. This choice
reflects that, aside from neuroplasticity in the language network in the left and/or right hemi-
Kugel, several alternative processes can induce brain changes over treatment. Zum Beispiel,
participants could have had different neural recruitment strategies during language processing
before the stroke, related to a difference in task strategy. After the stroke, these differences may
be further strengthened by the differential impact of the functional and structural lesions on the
Gehirn (Thompson, den Ouden, Bonakdarpour, Garibaldi, & Parrish, 2010). Ähnlich, Teilnehmer
could rely less or more on supporting cognitive processes after versus before the treatment, (z.B.,
attention, executive control, and responsive inhibition; Kurland, Balduin, & Tauer, 2010). In ADDI-
tion, behavioral improvement can manifest in different ways: as an increase in correct attempts
or as a decrease in overall errors. Both means of recovery can lead to different brain response
patterns (z.B., in the error network as explained in the previous section; Raichle et al., 1994). Für
comparison purposes, we have tried to include as much study-specific information as possible
in Appendix A in the online supporting information.
Neurosynth
To answer our second research question, whether the most consistent treatment-related changes
across therapies occur within the language network or outside, we used Neurosynth association
test maps for the different linguistic components as a reference (Figur 1). It should be noted that
this analysis combines highly variable studies. The association map represents a z-map for a two-
way ANOVA testing for an association between the search term and voxel responses. Because a
large number of studies contribute to the meta-analysis, it is assumed to provide a good estimate
of the specific response patterns (Yarkoni, Poldrack, Nichols, van Essen, & Wager, 2011).
Neurobiology of Language
412
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
RESULTS AND DISCUSSION
The first aim of this review was to explore whether treatment effects are dependent on the focus of
the therapy. The second aim was to explore (A) whether the brain regions/networks that most often
show treatment-related changes are located within the language network or outside, Und (B)
whether these changes are associated with individual differences in language (improvement).
In Appendix A in the online supporting information, the studies addressing local treatment-related
brain changes in specific gray or white matter regions are depicted in italic font. The studies
addressing distributed treatment-related brain changes in connectivity patterns are depicted in
bold. There was an insufficient number of connectivity studies to investigate the first aim (Wo
they need to be split according to the targeted linguistic component) and therefore these studies
are only discussed within the second aim.
Does the Neural Effect Depend on the Focus of the Intervention?
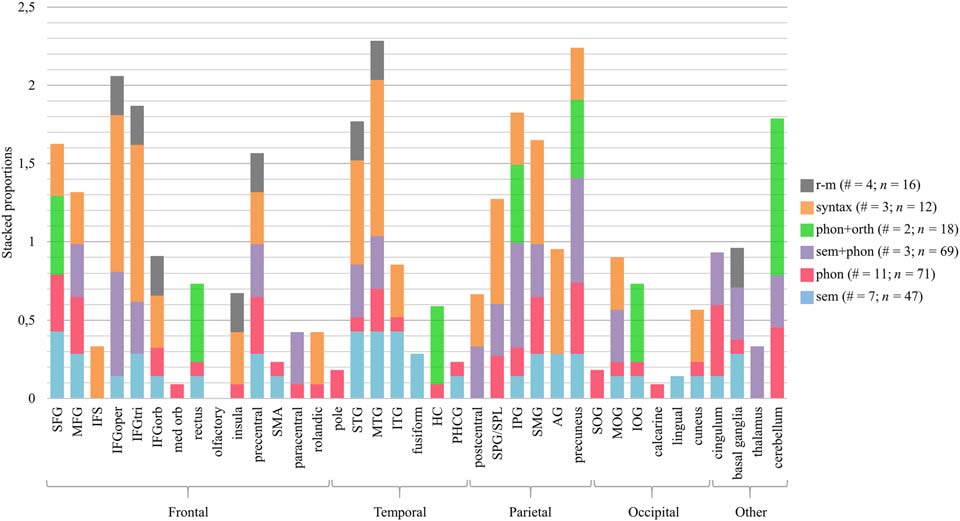
Figur 4 zeigt an, for all the brain regions in the left hemisphere undergoing intervention-related
changes, which types of interventions (sem, phon, sem+phon, phon+orth, syntax, and r-m) have been
associated with changes in that area. Zum Beispiel, the left superior frontal gyrus (SFG) was reported in
one out of three studies targeting syntactic processing. For this region and this type of intervention, Die
corresponding proportion is 0.33, which is represented by an orange bar. Zusätzlich, the left SFG was
reported in four out of 11 studies targeting phonological processing. daher, the red bar represent-
ing phonological processing has a height of 0.36. This proportion is calculated for every type of
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 4. Proportion of studies reporting treatment-related brain changes in a specific brain region of the left hemisphere, relative to the total
number of studies providing this type of intervention. The number of studies (#) and patients with aphasia (N) per type of intervention is re-
ported in the figure legend. SFG/MFG/IFG = superior/middle/inferior frontal gyrus, IFS = inferior frontal sulcus, oper = opercular, tri = trian-
gular, orb = orbital, med = medial, SMA = supplementary motor area, IPG/SPG = inferior/superior parietal gyrus, SPL = superior parietal
lobule, SMG = supramarginaler Gyrus, AG = angular gyrus, STG/MTG/ITG = superior/middle/inferior temporal gyrus, SOG/MOG/IOG = supe-
rior/middle/inferior occipital gyrus, HC = hippocampus, PHCG = parahippocampal gyrus. Anatomical labels other than those included in the
AAL-VOI atlas used by the included studies that did not report standard brain coordinates are IFS and SPL.
Neurobiology of Language
413
A review on treatment-related brain changes in aphasia
intervention, and consequently, for each region, the bar represents a stacked proportion, which can
be greater than one. The higher a specific color in the stacked bar, the more the brain changes in that
region were specific to the language component. The higher the stacked bar, the higher the number of
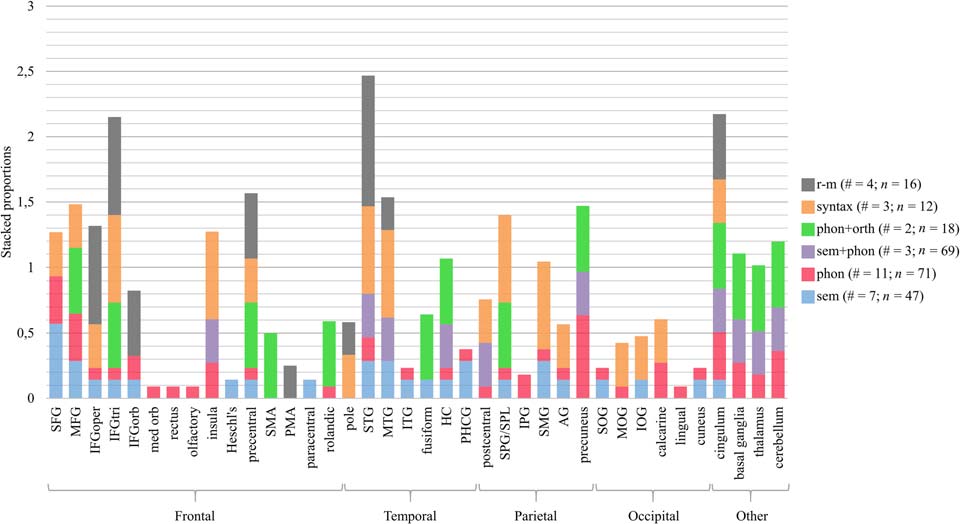
studies that led to changes in that brain region. In Abbildung 5 the data for the right hemisphere are rep-
resented in a similar way. We also made this figure after excluding studies that did not use a naming
task in the scanner (mostly rhythmic-melodic and syntactic processing). As the results are very similar
for the other language domains, we will not further discuss this.
Neural differences within linguistic interventions
Our first hypothesis was that brain changes related to interventions focused on phonological
processing are more frequent in regions associated with phonological processing, particularly the left
posterior inferior frontal lobe, the dorsal premotor regions, and an area in the parietotemporal junc-
tion. Based on Figures 4 Und 5 one can see that, although widespread, most brain changes after pho-
nological interventions (Rot) occurred in the bilateral SFG, middle frontal gyrus (MFG), precuneus,
cingulum, and cerebellum, the left supramarginal gyrus (SMG), superior parietal gyrus (SPG), MTG,
präzentraler Gyrus, and the right insula, calcarine gyrus, and basal ganglia. Except for the precentral
gyrus and the left SMG, these regions are not typically associated with phonological processing.
Jedoch, the SFG, the cerebellum, the precentral gyrus, and the insula are implicated in motor
Rede (Ackermann & Riecker, 2010; Stegemöller, 2017; Tourville & Guenther, 2011). PWA, espe-
cially the nonfluent subtype, frequently struggle with motor speech planning (Ogar, Slama, Dronkers,
Amici, & Gorno-Tempini, 2005) and articulation, which complicates the differentiation of articulatory
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 5. Treatment-related brain changes in the right hemisphere. The number of studies (#) and PWA (N) per type of intervention is reported
in the legend. Anatomical labels other than those included in the AAL-VOI atlas used by the included studies that did not report standard brain
coordinates are PMA and SPL. SFG/MFG/IFG = superior/middle/inferior frontal gyrus, oper = opercular, tri = triangular, orb = orbital, med =
medial, PMA = presupplementary motor area, SMA = supplementary motor area, IPG/SPG = inferior/superior parietal gyrus, SPL = superior
parietal lobule, SMG = supramarginaler Gyrus, AG = angular gyrus, STG/MTG/ITG = superior/middle/inferior temporal gyrus, SOG/MOG/IOG =
superior/middle/inferior occipital gyrus, HC = hippocampus, PHCG = parahippocampal gyrus.
Neurobiology of Language
414
A review on treatment-related brain changes in aphasia
and phonological errors. daher, it could be possible that the phonological language interventions,
all targeting speech production, indirectly affected speech-motor processes as well.
Our second hypothesis was that brain changes related to interventions focused on semantic
processing are more frequent in regions associated with semantic processing, particularly the
bilateral posterior MTG and ITG, and the left anterior temporal lobe. As with phonological inter-
Erfindungen, therapies focusing on semantic processing (Blau) were associated with changes in the
bilateral (although left > right) frontal, zeitlich, and parietal lobes. Jedoch, there is also evidence
that the temporal lobe was more influenced by semantic interventions, especially in the left
hemisphere. More than 40% of semantic studies found brain changes in the left temporal lobe,
compared with only 10–25% of phonological studies. Interventions combining semantic and
phonological processing (purple) led to very mixed results bilaterally (left > right) in the frontal,
temporal and parietal lobes, as well as in more medial/subcortical structures and the cerebellum.
The third hypothesis was that brain changes related to interventions involving orthographic
Verarbeitung (Grün) are more frequent in orthographic language networks. Those are situated more
posteriorly and ventrally than the previous networks and include the posterior temporal lobe, Die
fusiform gyrus, the lingual gyrus, the calcarine gyrus, and the cuneus (also see Table 1 im
online supporting information). Wieder, the results are widespread (right > left), but there was
relatively more involvement of the left inferior occipital gyrus (IOG) and the right fusiform gyrus
compared with the other interventions. This might be related to the early visual processing of
sublexical forms and reading processes that occur in these posterior ventral regions. Jedoch,
one should keep in mind that these results are based on only two studies.
The fourth hypothesis (mainly based on Vigneau et al., 2006) was that syntactic processing
(orange) engages inferior frontal regions as well as superior temporal regions, anteriorly as well
as posteriorly. Based on Figures 4 Und 5 this is clearly the case, although changes were not
restricted to these areas. Zum Beispiel, there are also BOLD changes in the bilateral superior
parietal lobule and in visual regions of the right hemisphere.
Although we expected that the linguistic interventions would mainly lead to brain changes in
the left hemisphere, there is bilateral neural involvement. Due to within-study variation in lesion
size and location in different PWA, perilesional activity might have been masked (Crosson et al.,
2007). Fridriksson, Richardson, et al. (2012) created patient-specific ROIs of the perilesional
cortex and residual naming areas in each lobe. They found that the best predictor of naming im-
provement was an increase in subject-specific perilesional activity in the frontal areas involved in
Wir, as well as frontal regions not recruited for naming by the control group. Daher, Gruppe
studies most probably underestimate the involvement of the left hemisphere in the treatment-
related recovery of aphasia. Zusätzlich, note that some studies did not show behavioral improve-
ment in (manche) Teilnehmer (see columns “behavioral outcome” and “results” in Appendix A in
the online supporting information) and/or included incorrect responses in their analysis (sehen
Appendix A, column “extra”). daher, it is possible that some of the results (d.h., the involve-
ment of the bilateral anterior cingulate region, insulae, right parietal lobe, medial temporal lobe,
basal ganglia, and thalamus) are related to error processing (Stevens et al., 2009). Jedoch, most
therapies did have a positive behavioral outcome. Another important limitation is that the amount
of treatment-related brain changes in each brain region was not considered, because these data
were not available for every study. This may have masked neural differences in the effect of
different types of treatment, since the amount of brain activity (rather than the location) im
affected networks could be specific to the treatment. Andererseits, it could also be possible
that treatment-related effects are not specific to the type of therapy administered. Abschließend,
treatment-related brain changes do not seem to be very treatment-specific. Jedoch, Fehler
processing effects and the fact that we were not able to quantify the degree to which brain regions
were affected by each treatment, may have masked treatment specificity.
Neurobiology of Language
415
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Neural differences between linguistic and rhythmic-melodic interventions
Hier, we compared interventions targeting the left hemisphere with those targeting the right hemi-
Kugel. It is widely accepted that typical linguistic processing is more supported by the left hemi-
sphere than the right hemisphere (z.B., Vigneau et al., 2006), whereas the right hemisphere is more
involved in musical, prosodic, and metalinguistic processing (Leon, Rodriguez, & Rosenbek,
2015). Genauer, phonological, semantic, orthographic, and syntactic processing rely
more on the left hemisphere. Jedoch, there is some involvement of the right temporal lobe in
the processing of context, for example during sentence and discourse comprehension, welche
involves both syntactic and semantic processing (Vigneau et al., 2011). Im Gegensatz, there is evi-
dence that linguistic prosody encoded in the intonational contour of a sentence, relies on fronto-
temporal areas in both hemispheres. The less segmental information, the higher the involvement of
the right hemisphere relative to the left one. This functional distinction between the left and right
hemisphere begins during acoustic processing in the primary auditory cortex (A1; Friederici, 2011;
Witteman, van Ijzendoorn, van de Velde, van Heuven, & Schiller, 2011). According to the asym-
metric sampling in the time hypothesis (Kacke, 2003), left A1 is specialized in processing rapidly
changing information with a time resolution of 20–40 ms (z.B., speech sounds), while right A1 pre-
fers the processing of slowly changing information (150–250 ms), such as tonal pitch changes.
Based on these findings, we expected that the right hemisphere would be affected by rhythmic-
melodic language interventions, such as melodic intonation therapy (MIT) and SIPARI, verglichen
with the other interventions. Similar to MIT, SIPARI combines singing, intonation, prosody,
breathing (atmung in German), rhythm, and improvisation (Jungblut, Huber, Mais, & Schnitker,
2014), and therefore places high demands on suprasegmental aspects of language. The initial
focus is on vocal training of melodic speech segments assumed to be supported by the right
hemisphere. Subsequently, the focus shifts to rhythmic chunking of these speech segments with
different complexity levels to stimulate the left hemisphere.
We hypothesized that brain changes related to interventions focused on rhythmic-melodic
Verarbeitung (gray) are more frequent in the frontal and temporal lobes of the right hemisphere.
When we compare effects in the left versus the right hemisphere, Figuren 4 Und 5 show that
three out of four r-m studies reported brain changes in the right IFG (in both the triangular and
opercular part), while only one out of four studies reported changes in the same structures in the
left IFG. Darüber hinaus, all r-m studies reported changes in the right STG, while only one study showed
changes in its left-hemispheric counterpart. Only in Jungblut et al. (2014), who provided SIPARI-
treatment, was there any evidence of left hemisphere involvement. This can be explained by the
fact that SIPARI is theoretically structured in such a way that, in addition to the right hemisphere,
the left hemisphere is increasingly stimulated over time by shifting the focus from singing to
rhythmic chunking of speech. When linguistic and rhythmic-melodic interventions are com-
pared, it can be seen that in the right STG and opercular part of the IFG there are more studies
(mindestens 33%) on rhythmic-melodic interventions that found changes in these regions, than the
studies on linguistic interventions. All r-m studies applied a whole-brain contrast analysis or a
data-driven ROI-analysis, which precludes bias due to the methodological approach.
Conclusions aim one
From the above discussion in Neural differences within linguistic interventions, it seems that most
brain regions with treatment-related changes were not specific to a particular type of language
intervention (because most regions have bars in multiple colors and not one). Andererseits,
there are some indications that some regions were more likely to show brain changes when
training a specific aspect of language. Zum Beispiel, in the temporal lobe, changes related to a
Neurobiology of Language
416
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
semantic intervention occurred more consistently than changes related to a phonological inter-
vention. Darüber hinaus, the studies integrating phonological and orthographic processing led to more
changes in the ventral posterior network compared with the other interventions. The interven-
tions focusing on semantic, phonological, orthographic, and syntactic processing seemed to elicit
brain changes in both hemispheres. This right-hemispheric involvement in treatments classically
targeting the left hemisphere could point to compensatory mechanisms after left-hemispheric
brain damage. (For a review see Cocquyt, De Ley, Santens, Van Borsel, & De Letter, 2017.)
This right-hemispheric compensation typically takes place in brain regions homologous to lan-
guage regions in the left hemisphere or in regions involved in more general cognitive functions
(z.B., executive functioning; see the next section). From the above discussion in Neural differ-
ences between linguistic and rhythmic-melodic interventions, it seems that the language interven-
tions focusing on rhythmic-melodic processing included in this review elicited more changes in
the right hemisphere compared with the left hemisphere. This right dominance was not found for
the linguistic interventions. Allgemein, across treatments and subjects, the regions that are in-
volved in language recovery are very diverse. It is hard to find similar patterns of brain changes
between intervention studies targeting the same linguistic component.
Consistency of Location of Treatment-Related Brain Changes
The second aim of this literature review was to describe whether the most consistent treatment-
related changes occur within the language network (d.h., Neurosynth response maps visualized in
Figur 1) or outside. We summarized which brain regions showed consistent treatment-related
changes across the included studies and investigated whether these ROIs are located within the
linguistic maps visualized in Figure 1 (aim 2a). We then explored whether and how these con-
sistent brain changes are related to (a change in) language behavior (aim 2b).
Which brain regions show treatment-related brain changes?
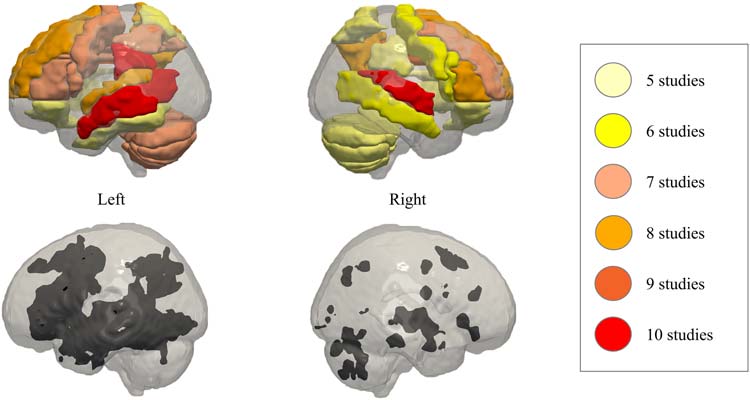
Tisch 3 summarizes which brain regions show treatment-related brain changes across the studies
included in this review and how frequently each region is identified. Tisch 1 in the online supporting
information indicates which type of linguistic functions (semantics, phonology, orthography, syntax,
and/or rhythmic-melodic processing) have been associated with each of these areas.
As shown in Table 3, the brain regions that were most frequently reported showing treatment-
related brain changes in PWA, across all kinds of language interventions, are the bilateral SFG, MFG,
IFG, precentral gyri, superior STG, MTG, SPG, SMG, precuneus, basal ganglia, cingulum and
cerebellum, the left ITG and inferior parietal gyrus, and the right insula. At least five of the 25 Studien
showed treatment-related brain changes in these ROIs. This choice reflects effects that are present in
one out of five of all treatment-related intervention studies in PWA, which is an effect size reported to
be much more common than higher effect sizes (Eickhoff et al., 2016). An important remark is that six
of the 25 studies considered here used an ROI-approach in their analysis, which might lead to an
overrepresentation of “classic” language areas. Jedoch, in five of them, the ROIs were chosen
based on the results of a precedent whole-brain analysis, which minimizes the possible bias of
the analysis choice.
The ROIs in Figure 6 represent the regions that were associated with treatment-related neural
changes in at least five out of 25 Studien (listed in Table 3). As previously explained, for included
studies reporting standard neural coordinates, anatomical labels were derived from the AAL-VOIs
Single-Subject atlas (Tzourio-Mazoyer et al., 2002). For the studies reporting only ROI labels,
their own labels were adopted. This means that a region can be colored if it was reported in
Neurobiology of Language
417
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Tisch 3.
Summary of brain regions showing treatment-related brain changes in patients with aphasia across studies
Change
Left hemisphere
Frontal
SFG (9)
MFG (8)
precentral (8)
Temporal
MTG (11)
STG (8)
ES G (5)
IFGoper (7)
fusiform (2)
HC (2)
Parietal
SMG (9)
Occipital
MOG (4)
Other*
cerebellum (8)
precuneus (9)
cuneus (3)
cingulum (6)
IPG (7)
SPG (6)
AG (4)
IOG (3)
SOG (2)
lingual (1)
basal ganglia (5)
thalamus (1)
Right hemisphere
IFGtri (7)
IFGorb (5)
rectus (3)
insula (3)
paracentral (2)
SMA (2)
rolandic (2)
med orb (1)
IFS (1)
SFG (9)
IFGtri (8)
MFG (8)
IFGoper (6)
insula (6)
precentral (6)
IFGorb (5)
rolandic (2)
SMA (1)
med orb (1)
PMA (1)
paracentral (1)
rectus (1)
olfactory (1)
PHCG (2)
postcentral (2)
calcarine (1)
pole (2)
STG (11)
MTG (6)
fusiform (2)
HC (3)
precuneus (8)
calcarine (4)
cingulum (9)
SMG (5)
SPG (5)
AG (3)
cuneus (2)
cerebellum (6)
SOG (2)
MOG (2)
basal ganglia (5)
thalamus (4)
PHCG (3)
postcentral (3)
IOG (2)
IPG (2)
lingual (1)
ES G (2)
pole (2)
Heschl’s (1)
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Notiz. The number between brackets indicates how many fMRI-studies (out of 25) show treatment-related changes in this brain region. Of the 32 studies included in
the review, five exclusively focus on brain connectivity, while 27 studies report pre-, post, or trained-untrained differences in specific brain regions. Of those 27, Die
studies by Abel et al. (2014) and Abel et al. (2015) were counted as one because they involve the same participants, and the study by Fridriksson, Richardson, et al.
(2012) could not be included since it does not report specific anatomical ROIs (only “perilesional area” vs. “residual naming area”). Daher, in Summe, 25 studies remain.
SFG/MFG/IFG = superior/middle/inferior frontal gyrus, IFS = inferior frontal sulcus, oper = opercular, tri = triangular, orb = orbital, med = medial, PMA/SMA = pre-/
supplementary motor area, IPG/SPG = inferior/superior parietal gyrus, SMG = supramarginaler Gyrus, AG = angular gyrus, STG/MTG/ITG = superior/middle/inferior
temporal gyrus, SOG/MOG/IOG = superior/middle/inferior occipital gyrus, HC = hippocampus, PHCG = parahippocampal gyrus, mid = middle. * This category
includes regions that do not belong to one of the previous categories, such as subcortical, cingular and cerebellar structures.
Neurobiology of Language
418
A review on treatment-related brain changes in aphasia
Figur 6. Comparison of Neurosynth language networks (black regions in the lower panel) with regions of interest (ROIs) most frequently
associated with treatment-related brain changes (shown on the color map in the upper panel). In the upper panel, the color represents the number
of selected studies in which the ROI was reported. In the lower panel, the black regions correspond to association test maps of semantic,
phonological, syntactic, and orthographic processing, according to the Neurosynth meta-analysis (http://neurosynth.org/). The figures were
composed using Paraview software (Ausführung 5.4.1; https://www.paraview.org/) following the guidelines specified in Madan (2015).
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
(mindestens) five studies using stereotaxic coordinates, if it was reported in five studies with only
anatomical labels, or a combination of both.
Consistent treatment-related changes (five of the 25 Studien, in color) overlap with the language
Netzwerk (in black) mainly in the left IFG (the orbital, triangular, and opercular part), die linke (ventral)
präzentraler Gyrus, die linke Maschinenpistole, IPG and ITG, and in the bilateral STG and MTG. Although the left
insula, ES G, fusiform gyrus, and angular gyrus also belong to the traditional language network
depicted in black, treatment-related changes have been less consistently found in these regions.
Although the language network (in black) is more situated in the left hemisphere than the right
hemisphere, treatment-related brain changes (in color) are not restricted to the left hemisphere.
Gold and Kertesz (2000) stated that contributions of the right hemisphere are task-dependent
and are larger in lexico-semantic processing than phonological processing. This arises because,
in the healthy brain, lexico-semantic processing is less left-lateralized (d.h., more bilateral) als
phonological processing. Jedoch, in Abbildung 5 it can be observed that all treatments evoked
changes in right-hemispheric ROIs, not only interventions focusing on semantic processing.
Structures wherein treatment-related brain changes occurred consistently over different types of
treatment, and which do not nicely overlap with the linguistic network (in black) were the lateral,
orbital as well as medial part of the bilateral superior and middle frontal gyri, the left hippocampus,
paracentral lobule and IOG, the bilateral precuneus, cingulate, cerebellum, SPG, and the right
basal ganglia. Jedoch, for the right cerebellum and the left precuneus, es gab welche
overlapping “dots.” Figure 6 might overrepresent the frontal and cerebellar areas, weil wir
were limited to visualize ROI labels instead of specific MNI-coordinates. Not all studies reported
these coordinates, and these ROIs tend to encompass large areas of the brain.
Concerning connectivity, the included functional and effective connectivity studies (nicht
represented in Table 3 or Figure 6) found modulations of the connectome in very similar regions.
van Hees et al. (2014B) and Sandberg et al. (2015) demonstrated posttreatment modulations of
resting-state and task-related functional connectivity strength, jeweils. Upregulation of
Neurobiology of Language
419
A review on treatment-related brain changes in aphasia
connectivity strength was found in the language network of PWA in and between both hemi-
spheres, as well as in the bilateral SFG, left MFG, precuneus, and precentral gyrus (Sandberg
et al., 2015). Andererseits, there was a downregulation of connectivity between bilateral
language regions and the right basal ganglia, cingulum, and cerebellum (van Hees et al., 2014B).
Kiran et al. (2015) and Vitali et al. (2010) showed treatment-related changes in task-related effec-
tive connectivity patterns. In agreement with the functional connectivity results, effective connec-
tivity modulations existed throughout the language network in and between both hemispheres, als
well as between the IFG and the MFG in both hemispheres (Kiran et al., 2015). Außerdem,
Marcotte et al. (2013) demonstrated that language interventions in PWA were able to normalize
the amount of functional integration within the posterior default mode network (the bilateral
MTG, the AG, the left ITG, the right middle cingulate, and the right cerebellum). This network
is more active during conscious resting states of the brain compared with during the performance
of a cognitive task (Höhle & Trimble, 2006).
Association with language outcomes
We explored whether the measured brain changes in the regions visualized in Figure 6 show
(positive) associations with language outcomes. As explained above in Direction of brain change,
relating the brain changes to behavioral change is necessary to interpret the meaning of the
activity patterns. daher, in this section, we will consider the difference between an increase
and a decrease in the measured neural response over the course of the treatment.
Among the consistently identified ROIs (colored regions in Figure 6) overlapping with the lan-
guage network, BOLD increases in the left IFGoper (Fridriksson, 2010), BOLD decreases in the left
STG, MTG, and SMG (Abel et al., 2014), and BOLD increases as well as decreases in the IPG (Abel
et al., 2014; Fridriksson, 2010; Raboyeau et al., 2008) have been related to language improvement
across studies. These studies highlight the importance of the left hemisphere for aphasia recovery,
although it is not clear why Abel et al. (2014) found BOLD decreases in these regions, in contrast to
the other studies. Jedoch, they found a negative association between activity decrease and
therapy gains, implicating the importance of continued reliance on the left hemisphere during
the treatment. For the right hemisphere, there are two main hypotheses concerning its involvement
in the post-stroke neural response pattern. In the first hypothesis, left-hemisphere damage is thought
to induce pathological transcallosal disinhibition of the right-hemisphere homologs. Als solche, Die
proponents of this presupposition, view right-hemispheric activity as detrimental (z.B., Heiss &
Thiel, 2006; Price & Crinion, 2005; Saur et al., 2006). Support for this hypothesis comes from
the positive association between language improvement and BOLD decreases in the right insula
and IFGoper (Nardo et al., 2017), the right precentral gyrus and precuneus (Raboyeau et al., 2008), oder
a decrease in fractional anisotropy of the IFGoper (Wan et al., 2014). Jedoch, two other studies in
this review found positive associations between language improvement and brain responses in
different right hemisphere regions (MFG, SMA, fusiform gyrus, hippocampus, SPG, putamen and
anterior cingulate). This suggests that the right hemisphere could also support language recovery in
the chronic phase after stroke (Menke et al., 2009; Raboyeau et al., 2008).
Positive correlations between language improvement (picture naming or picture description)
und Reaktion (changes), in regions that are not traditionally associated with language, suggest that
these structures may play a role in recovery from aphasia, particularly naming. Genauer,
there were positive associations between naming improvement and BOLD signal in the bilateral
SFG (Brownsett et al., 2014), BOLD increases in the bilateral MFG (Fridriksson, 2010; Raboyeau
et al., 2008), BOLD decreases in the bilateral precuneus (Raboyeau et al., 2008), BOLD increases
in the left precuneus (Fridriksson, 2010), posttreatment BOLD signal in the right precuneus
Neurobiology of Language
420
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
(van Hees et al., 2014), BOLD decreases in the left paracentral lobule (Abel et al., 2014), BOLD
increases in the left IOG, left cerebellum, bilateral hippocampus, and the right SPG (Menke
et al., 2009), and BOLD increases in the right putamen and the right anterior cingulate
(Raboyeau et al., 2008). Zusammenfassend, across different studies, changes in left-hemispheric
language regions, right-hemispheric homologs, as well as bilateral regions not traditionally asso-
ciated with language have been associated with language improvement over treatment. It is very
likely that part of this variability is caused by variations in lesion size and site, which determines
whether there is still recovery potential in the left hemisphere, as well as premorbid lateralization
patterns (Warburton, Price, Swinburn, & Wise, 1999) and time post-stroke (Saur et al., 2006).
Among the connectivity studies, Vitali et al. (2010) showed associations of modulations in
task-related effective connectivity patterns throughout the language network in and between both
hemispheres with correct picture naming of trained items. Im Gegensatz, the correlations between
the amount of functional integration within the posterior default mode network and naming
improvement was not significant in the study by Marcotte et al. (2013). Gesamt, diese Ergebnisse
suggest that modulation and normalization of functional and effective connections within and
outside the language network are concurrent, but not necessarily correlated, with aphasia
recovery. In addition to functional connectivity, recent research has suggested that intact struc-
tural connectivity is beneficial for successful aphasia recovery (Bonilha, Gleichgerrcht, Nesland,
Rorden, & Fridriksson, 2016; Bonilha et al., 2017; Griffis, Nenert, Allendorfer, & Szaflarski, 2017;
Yourganov, Fridriksson, Rorden, Gleichgerrcht, & Bonilha, 2016). Several studies have demon-
strated the importance of the AF for language recovery after stroke, especially for improvement in
speech production (Breier, Juranek, & Papanicolaou, 2011; Hosomi, Nagakane, & Yamada,
2009; Jang, 2013; Jang & Lee, 2014). This finding is consistent with the assumed function of this
white-matter pathway in mapping acoustic representations of sounds with their motor represen-
Station (Saur et al., 2008). In the study by van Hees et al. (2014A) included in this review, the pre-
and posttreatment mean generalized fractional anisotropy, an indirect measure of fiber bundle
characteristics, of the left AF correlated with maintenance of (phonological) treatment gains, Und
the fractional anisotropy value increased over treatment. This observation complements that
of Marchina et al. (2011) and Wang, Marchina, Norton, Wan, and Schlaug (2013) who showed
that lesion load of the left AF significantly predicted the level of impairment in language produc-
tion in chronic stroke patients, explaining more variance in language behavior than the functional
gray matter lesion load. It also has been shown that better language performance after irreversible
damage in the left hemisphere is associated with increased structural connectivity in the right AF
(Forkel et al., 2014). Fridriksson et al. (2006) demonstrated significant naming recovery of a sub-
ject sparing some white matter connections in the inferior frontal lobe, while a patient with more
extensive white matter damage in this area did not recover. Because the AF is connected to the
posterior part of the ventrolateral frontal lobe (Catani, Jones, & Ffytche, 2005), these white matter
connections might have been part of it.
Conclusions aim two
Zusammenfassend, across studies, many brain regions have been associated with treatment-related
language recovery, although in a very inconsistent way, which makes it difficult to make any
decisive conclusions. Although the left IFG, STG, MTG, and inferior parietal regions are typically
considered to be involved in linguistic processing (Geschwind, 1970; Hickok & Kacke, 2007),
their right counterparts and the bilateral SFG, MFG, precentral gyri, SPG, precuneus, cerebellum,
cingulum, right insula, and basal ganglia are not. The latter regions are mainly known for their
nonlinguistic functions. Daher, it is possible that brain regions more medial and/or subcortical to
what is generally studied in the context of language processing, are additionally involved in the
Neurobiology of Language
421
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
process of aphasia recovery. Zusätzlich, a variety of other cognitive functions have been attrib-
uted to the brain regions that are typically considered to be involved in language processing
(Geschwind, 1970; Hickok & Kacke, 2007), such as feedback mechanisms (motor and auditory);
the planning, coordination, zeitliche Koordinierung, Ausführung, and control of (Rede) Bewegungen; amodal
semantic processing; learning; attention; and other higher-order executive functions.
Language improvement is evident not only in changes in distinct gray matter regions, but also
in their functional connectivity patterns and in the white matter pathways enabling direct com-
munication between these regions. More research on the role of brain connectivity in aphasia
recovery is necessary to fully understand functional deficits beyond the lesion and to evaluate
reorganization potential after stroke at the neural network level. Methodological shortcomings
and variability between studies (z.B., in lesion size), as well as the large inconsistency in results
across the different studies make it hard to present clear conclusions on treatment-related brain
changes in PWA. We cannot put forward that language regions show consistent treatment-related
changes, nor can we conclude that other regions involved in cognitive processing show consis-
tent changes. There is not, daher, more evidence for changes in the language network than for
changes outside the language network. In the next section, we will discuss these issues in more
detail and formulate suggestions for future research.
Limitations and Future Directions
Over the past two decades, an increasing number of studies have investigated treatment-related
brain changes in PWA after stroke. In this review, we integrated the results in a descriptive way to
provide the current state-of-the-art on this topic. We are well aware that even in this descriptive
comparison one should be cautious when interpreting the results because of variability between
as well as within studies. In diesem Abschnitt, we will summarize the limitations that we came across
throughout the review and formulate recommendations to enable quantitative and more reliable
comparisons in the future. Tisch 4 provides a short overview of the limitations and recommen-
dations, which will be elaborated on throughout the text.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Methodological limitations
Although the main aim of this research was to gain insight into the mechanisms underlying apha-
sia recovery, there are considerable differences in the operationalization of the experiments, Die
neuroimaging pipelines, and the statistical analyses among the included studies. Zusammenfassend,
there are differences in the imaging modality (HAUSTIER, MEG, fMRT, DWI), research design (Gruppe
study vs. [multiple] single-subject study), in-scanner language task (targeted linguistic compo-
nent, production vs. comprehension, overt vs. covert, word vs. sentence level), assessed contrast
(lenient vs. stringent, pre-post vs. trained-untrained), neuroimaging analysis approach (ganz-
brain vs. ROI-approach), modeling of the trials in the statistical model (including vs. excluding
incorrect trials, collapsing data from multiple time points or not), and the statistical analysis itself
(z.B., t-test, F-test, [multiple] regression analysis, partial least squares analysis, correlation analysis).
The systematic integration of the diverging findings of these studies might overcome this vari-
ability to some extent. Jedoch, failing to report peak locations of treatment-related effects in
stereotaxic reference space makes coordinate-based meta-analysis at present impossible. Das
is an important limitation and point of concern for future research, because meta-analyses
could compensate for the highly variable results from the small-data studies, which are still very
common in this field of research (Eickhoff et al., 2016).
Several methodological limitations are related to the brain lesion. To take into account the role
of perilesional activity in language recovery over time, it is important to consider individual data
Neurobiology of Language
422
A review on treatment-related brain changes in aphasia
N
Ö
ich
S
e
l
e
H
T
F
Ö
T
C
e
F
F
e
e
H
T
R
e
D
ich
S
N
Ö
C
D
N
A
S
ich
S
j
l
A
N
A
e
H
T
N
ich
S
T
N
Ö
P
ich
A
T
A
D
l
A
u
D
ich
v
ich
D
N
ich
R
e
D
ich
S
N
Ö
C
S
N
R
e
T
T
A
P
N
Ö
ich
S
e
l
T
N
e
R
e
F
F
ich
D
ich
ich
G
N
N
B
M
Ö
C
e
S
N
Ö
P
S
e
R
e
H
T
F
Ö
G
N
ich
l
e
D
Ö
M
N
ich
S
N
Ö
ich
T
C
N
u
F
e
S
N
Ö
P
S
e
R
C
ich
M
A
N
j
D
Ö
M
e
H
C
ich
F
ich
C
e
P
S
–
T
N
e
ich
T
A
P
e
S
U
e
S
N
Ö
P
S
e
R
C
ich
M
A
N
j
D
Ö
M
e
H
e
H
T
F
Ö
j
T
ich
l
ich
B
A
ich
l
e
R
N
u
l
A
ich
T
N
e
T
Ö
P
S
R
Ö
T
C
A
F
C
ich
F
ich
C
e
P
S
–
T
N
e
ich
T
A
P
R
e
H
Ö
T
D
N
A
R
Ö
ich
v
A
H
e
B
Ö
T
S
e
G
N
A
H
C
N
ich
A
R
B
e
T
A
l
e
R
e
H
T
N
ich
e
G
N
A
H
C
e
H
T
F
Ö
N
Ö
ich
T
C
e
R
ich
D
N
ich
j
T
ich
l
ich
B
A
ich
R
A
V
S
k
S
A
T
–
ICH
R
M
F
l
Ö
R
T
N
Ö
C
R
Ö
S
N
Ö
ich
S
S
e
S
N
A
C
S
T
S
Ö
P
D
N
A
–
e
R
P
e
l
P
ich
T
l
u
M
e
S
U
k
S
A
T
ICH
R
M
F
e
H
T
Ö
T
T
N
e
ich
T
A
P
e
H
T
F
Ö
N
Ö
ich
T
A
u
T
ich
B
A
H
R
Ö
F
k
S
ich
R
S
T
l
u
S
e
R
F
Ö
N
Ö
ich
T
A
T
e
R
P
R
e
T
N
ICH
N
ich
S
G
N
D
N
ich
ich
F
H
T
ich
w
e
k
Ö
R
T
S
–
T
S
Ö
P
e
S
A
H
P
C
ich
N
Ö
R
H
C
N
ich
S
G
N
D
N
ich
ich
F
e
R
A
P
M
Ö
C
e
S
A
H
P
C
ich
N
Ö
R
H
C
e
H
T
N
Ö
S
e
ich
D
u
T
S
T
N
e
M
T
A
e
R
T
F
Ö
S
u
C
Ö
F
l
A
N
G
ich
S
D
L
Ö
B
N
G
ich
S
e
D
j
D
u
T
S
e
H
T
N
ich
S
A
ich
B
N
Ö
ich
T
C
e
l
e
S
ich
D
Ö
v
A
j
T
ich
e
N
e
G
Ö
R
e
T
e
H
T
N
e
ich
T
A
P
j
D
u
T
S
R
e
T
N
ich
C
ich
T
A
M
e
T
S
j
S
e
S
A
H
P
e
T
u
C
A
B
u
S
D
N
A
e
T
u
C
A
e
k
Ö
R
T
S
–
T
S
Ö
P
S
T
N
e
ich
T
A
P
F
Ö
S
e
l
P
M
A
S
R
e
G
R
A
l
N
ich
j
R
e
v
Ö
C
e
R
A
ich
S
A
H
P
A
F
Ö
S
e
ich
D
u
T
S
M
R
Ö
F
R
e
P
e
l
P
M
A
S
A
e
v
A
H
S
e
ich
D
u
T
S
e
H
T
F
Ö
%
0
7
N
A
H
T
e
R
Ö
M
e
z
ich
S
e
l
P
M
A
S
0
1
N
A
H
T
R
e
l
l
A
M
S
e
z
ich
S
j
T
ich
C
ich
F
ich
C
e
P
S
e
G
A
u
G
N
A
l
D
N
A
,
e
C
N
A
T
R
Ö
P
M
ich
,
N
Ö
ich
T
C
N
u
F
e
H
T
e
z
ich
R
e
T
C
A
R
A
H
C
l
A
R
e
v
e
S
D
N
A
k
R
Ö
w
T
e
N
e
G
A
u
G
N
A
l
e
H
T
N
e
e
w
T
e
B
P
A
l
R
e
v
Ö
S
T
C
e
F
F
e
D
e
T
A
l
e
R
–
T
N
e
M
T
A
e
R
T
A
ich
S
A
H
P
A
N
ich
D
e
ich
F
ich
T
N
e
D
ich
j
l
T
N
e
T
S
ich
S
N
Ö
C
N
e
e
B
e
v
A
H
T
A
H
T
S
N
Ö
ich
G
e
R
F
Ö
S
e
S
S
e
C
Ö
R
P
e
G
A
u
G
N
A
l
–
N
Ö
N
e
G
A
u
G
N
A
l
e
H
T
F
Ö
e
D
ich
S
T
u
Ö
)
S
e
S
j
l
A
N
A
N
ich
A
R
B
–
e
l
Ö
H
w
(
j
R
e
v
Ö
C
e
R
k
R
Ö
w
T
e
N
j
l
l
A
N
Ö
ich
T
C
N
u
F
S
A
l
l
e
w
S
A
j
l
l
A
R
u
T
C
u
R
T
S
,
l
e
v
e
l
S
N
R
e
T
T
A
P
e
M
Ö
T
C
e
N
N
Ö
C
e
H
T
N
Ö
S
e
G
N
A
H
C
N
ich
A
R
B
D
e
T
A
l
e
R
–
T
N
e
M
T
A
e
R
T
l
e
R
Ö
P
X
E
e
S
N
Ö
P
S
e
R
l
A
N
Ö
ich
T
C
N
u
F
N
ich
S
e
G
N
A
H
C
l
A
N
Ö
ich
G
e
R
N
Ö
S
u
C
Ö
F
S
e
ich
D
u
T
S
j
T
ich
v
ich
T
C
e
N
N
Ö
C
l
e
v
e
l
T
C
e
J
B
u
S
l
A
u
D
ich
v
ich
D
N
ich
e
H
T
T
A
A
T
A
D
R
e
D
ich
S
N
Ö
C
N
Ö
ich
S
N
e
M
D
ich
T
C
e
J
B
u
S
e
H
T
S
S
Ö
R
C
A
D
e
S
P
A
l
l
Ö
C
e
R
A
S
T
l
u
S
e
R
S
A
j
T
ich
l
ich
B
A
ich
R
A
v
l
A
u
D
ich
v
ich
D
N
ICH
S
T
C
e
F
F
e
l
A
R
u
e
N
R
Ö
F
ich
S
e
T
A
N
D
R
Ö
Ö
C
C
ich
X
A
T
Ö
e
R
e
T
S
D
R
A
D
N
A
T
S
T
R
Ö
P
e
F
ich
S
e
T
A
N
D
R
Ö
Ö
C
C
ich
X
A
T
Ö
e
R
e
T
S
D
R
A
D
N
A
T
S
G
N
ich
T
R
Ö
P
e
R
F
Ö
k
C
A
L
A
ich
S
A
H
P
A
H
T
ich
w
S
T
N
e
ich
T
A
P
N
ich
S
e
G
N
A
H
C
N
ich
A
R
B
D
e
T
A
l
e
R
–
T
N
e
M
T
A
e
R
T
F
Ö
A
e
R
A
e
H
T
N
ich
k
R
Ö
w
e
R
u
u
T
F
R
Ö
F
S
N
Ö
ich
T
A
D
N
e
M
M
Ö
C
e
R
D
N
A
S
N
Ö
ich
T
A
T
ich
M
ich
l
e
H
T
F
Ö
w
e
ich
v
R
e
v
Ö
.
4
e
l
B
A
T
S
N
Ö
ich
T
A
D
N
e
M
M
Ö
C
e
R
N
Ö
ich
T
A
T
ich
M
ich
L
)
S
e
S
j
l
A
N
A
–
A
T
e
M
(
S
G
N
D
N
ich
ich
F
G
N
ich
G
R
e
v
ich
D
F
Ö
N
Ö
ich
T
A
R
G
e
T
N
ich
C
ich
T
A
M
e
T
S
j
S
N
Ö
ich
T
A
z
ich
l
A
N
Ö
ich
T
A
R
e
P
Ö
N
ich
S
e
ich
D
u
T
S
N
e
e
w
T
e
B
j
T
ich
l
ich
B
A
ich
R
A
V
l
j
G
Ö
Ö
D
Ö
H
T
e
M
C
ich
P
Ö
T
D
N
A
,
S
e
N
ich
l
e
P
P
ich
G
N
ich
G
A
M
Ö
R
u
e
N
ich
,
S
T
N
e
M
ich
R
e
P
X
e
e
H
T
F
Ö
S
e
S
j
l
A
N
A
l
A
C
ich
T
S
ich
T
A
T
S
N
Ö
ich
T
A
M
R
Ö
N
F
ich
l
ich
u
F
G
N
N
A
e
M
Neurobiology of Language
423
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
points in the analysis (z.B., regression). In group analysis of PWA with diverse lesion patterns, Dort
is a lack of power to detect changes in perilesional areas, even when the group is highly homo-
geneous (Crosson et al., 2007; Meinzer et al., 2013). This variability in lesion size and location also
leads to a lower statistical power in the lesioned left hemisphere as compared with the structurally
intact right hemisphere. Insbesondere, the effect of lesion site and size on treatment-related recovery
should be evaluated, because it is known to be one of the most predictive factors in aphasia prog-
nosis (z.B., Plowman, Hentz, & Ellis, 2012). Jedoch, only a few studies included lesion informa-
tion as a confounding variable in their correlation or regression analysis (parametrized as lesion
volume or IFG lesion load; Brownsett et al., 2014; van Hees et al., 2014A; Wan et al., 2014). Als
shown by Wang et al. (2013), it might be interesting for future studies to additionally include a
measure of AF-lesion load, because this explained more variance in language behavior than func-
tional gray matter lesion load. Only two studies performed a separate analysis to specifically assess
the effect of the lesion on recovery patterns (d.h., voxel-based lesion recovery analysis (Fridriksson,
2010) and joint posttreatment independent component analysis (Abel et al., 2015). Crinion,
Holland, Copland, Thompson, and Hillis (2013) provided guidelines for the quantification of brain
lesions after stroke, since in this area, there is also substantial variability in methodology.
An important limitation and point of concern for future research is the potential unreliability of
the hemodynamic response in stroke populations with cerebrovascular damage. This response is
dependent on cerebral blood flow, cerebral blood volume, and oxygen consumption. There is
evidence that the neurovascular coupling response, which underlies fMRI and is typically
modeled by the hemodynamic response function (HRF), is reduced and delayed in stroke patients
(Bonakpardour, Parrish, & Thompson, 2007; Crinion & Leff, 2007; Lake, Bazzigaluppi, &
Stefanovic, 2016; Thompson et al., 2010) and even in healthy aging (Nair, Raut, &
Prabhakaran, 2017). Siegel, Snyder, Ramsey, Shulman, and Corbetta (2016) demonstrated that
over one-third of stroke patients show hemodynamic lags two weeks post-stroke, dropping to
15% three months post-stroke and 10% one year post-stroke. Wichtig, the amount of lag
severity was correlated with lesion size and severity of deficits in multiple domains. Some studies
included in the review tried to consider this by estimating patient-specific HRFs using long-trial
fMRT, to enhance the detection of BOLD changes in brain regions with delayed HRFs (z.B.,
Thompson et al., 2010). Darüber hinaus, by conducting additional perfusion imaging, hypoperfused
tissue could be identified. In der Tat, Thompson and colleagues showed associations between
decreased blood flow and an increased time-to-peak value of the HRF, and between perfusion
levels and treatment-related BOLD changes. Wichtig, this hypoperfusion was not limited to
the perilesional area, extending the lesion to remote brain regions, even in the contralateral hemi-
Kugel, although perfusion values were generally higher there than in the affected hemisphere.
daher, studies that limit their analysis to a canonical model of the HRF, based on healthy brain
responses, might underestimate or completely fail to detect functional activity in affected regions
(Bonakdarpour, Beeson, Demarco, & Rapcsak, 2015).
Subject-level analyses allow for more careful consideration of not only the lesion but also
differences between PWA in age, Geschlecht, education level, time post-stroke, already received
intervention, aphasic symptoms, task strategy, and response to treatment (see also Individual
variability as meaningful information below). All these sources of variability add to the challenge
of finding consistent results in aphasia recovery studies. In an ideal situation, insights in
pre-stroke brain structure and function would highly improve our understanding of neuroplasticity
after stroke. This can only be achieved by conducting large-scale longitudinal studies of people at
high risk for stroke, considering factors such as family history, arterial hypertension, hyperlipidemia,
diabetes, nicotine, and/or alcohol (ab)use. Another option is to assess undamaged structures
immediately after stroke, to be ahead of brain reorganization as much as possible.
Neurobiology of Language
424
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Interpretation of results
The location of the treatment-related effects in each study is highly dependent on the large
number of (subjective) choices on the specific implementation, as listed above. A crucial issue
that is not typically addressed in intervention studies of PWA is the distinction between treatment-
related brain changes and overall scan-rescan variability, especially the neural effect of task
learning (Chein & Schneider, 2005). If PWA habituate to the imaging task from pre- to post-
intervention, neural activity in task-related regions will most probably decrease, independent
from the effect of the treatment. Three possible options to disentangle the effects of the treatment
from practice-related reductions in brain activity (Rapp, Caplan, Edwards, Visch-Brink, &
Thompson, 2013) Sind (A) to compare the fMRI-task of interest with a control task, (B) to compare
treated with untreated items, Und (C) to estimate the test-retest variability by performing multiple
scan sessions pre- and/or posttreatment (Brownsett et al., 2014; Cornelissen et al., 2003;
Fridriksson et al., 2007; Fridriksson, 2010; Fridriksson et al., 2006; Fridriksson, Richardson,
et al., 2012; Sandberg et al., 2015; Schlaug et al., 2009). Jedoch, 11 studies did not perform
multiple scan sessions, or mention or use an appropriate control task (such as null events, looking
at a fixation cross, or rest; Abel et al., 2014, 2015; Jungblut et al., 2014; Marcotte et al., 2018;
Menke et al., 2009; Raboyeau et al., 2008; Tabei et al., 2016; Thompson et al., 2010, 2013; Vitali
et al., 2007, 2010), and only one of those 11 contrasted trained items with untrained items to
compensate for this (Vitali et al., 2007).
The interpretation of the meaning of the effects is further complicated by the variability in the
direction of the change in BOLD response in both hemispheres across PWA and studies, sowie
as the diverging possible causes of treatment-related neural plasticity. Zusammenfassend, some of the
included studies reported correlations between increased activity in (homologous) Rechts-
hemispheric regions and language improvement (Menke et al., 2009; Raboyeau et al., 2008),
which is frequently interpreted as compensation. Others reported associations between activity
decreases in bilateral brain regions and better language performance, which is attributed to
increased task processing efficiency (Nardo et al., 2017; Raboyeau et al., 2008). Im Gegensatz, andere
studies found associations between therapy-induced language improvement and increased (oder
less decreased) activity in bilateral brain regions, including perilesional and spared left-
hemispheric areas. This might be related to increased task demands, successful reorganization
patterns or maladaptive plasticity (Abel et al., 2014; Fridriksson, 2010; Menke et al., 2009;
Raboyeau et al., 2008). In the future, careful and individual consideration of lesion site and size,
treatment strategies, response to treatment, and behavioral symptoms is needed to understand
why and how the changes in the BOLD signal are related to behavioral improvement
(Hartwigsen & Saur, 2019). Darüber hinaus, comparisons with intervention studies in PWA in the
subacute phase after stroke are needed to take into account and to understand the dynamic nature
of brain plasticity across the time course of recovery (Saur et al., 2006; Saur & Hartwigsen, 2012).
At present, intervention studies almost exclusively took place in the chronic phase after stroke,
although it is generally assumed that most language recovery takes place in the first days to weeks
after stroke, and the mechanisms involved are most probably different.
Another issue that impacts the interpretation of the results concerns the inclusion criteria for
the various interventions provided in the different studies. If specific behavioral symptoms were
prerequisites to receive a specific treatment, the inclusion criteria of the different studies induced
a systematic selection bias that ultimately also would have influenced the (generalizability of the)
results. To explore this, we checked the patient-specific inclusion criteria of all included studies
and found that, overall, the studies providing anomia treatments (sem, phon, sem+phon and
phon+orth interventions) selected patients with at least a moderate naming deficit or did not
mention any inclusion criteria. Remarkably, the majority of phonological studies included
Neurobiology of Language
425
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
patients with nonfluent aphasia, while semantic (and sem+phon) studies included patients with
various aphasia profiles or even fluent aphasia. The studies providing interventions on the
syntactic level all included agrammatic patients with nonfluent aphasia. The studies providing
rhythmic-melodic interventions also specifically included patients with (mostly moderate to
severe) nonfluent aphasia. Because the difference in aphasia symptoms/profiles across studies
may translate to a difference in lesion patterns, this selection bias could have systematically
influenced the amount of change that was possible in the different linguistic subnetworks.
Sample size
Due to the specific patient population and practical concerns, studies in PWA frequently have to
deal with small sample sizes. The sample size of the included studies varies from one to 29 Patienten,
with more than 70% of the studies (23/32) having a sample size smaller than 10. Figur 3 zeigt an
the number of studies (on the y-axis) including a certain number of PWA (on the x-axis). From this
figure, it is clear that more than 70% of the included studies of treatment-related changes in PWA
include 10 participants or less. As Ramus, Altarelli, Jednoróg, Zhao, and Scotto di Covella (2018)
described in their review, neuroimaging studies with such limited sample sizes are statistically
underpowered to detect group differences. As a consequence, unkorrigiert (or not adequately
corrected) results are frequently reported, which are more likely to be unreliable (Ramus et al.,
2018). Daher, in diesem Fall, one problem (underpowered studies) might lead to another (reporting
spurious results), preventing the field from moving forward.
Insbesondere, when looking for differences with small to medium effect sizes, which is the case
for treatment-related brain changes in PWA, a large sample is needed to detect within-group
differences with adequate (not too large) confidence intervals (Ramus et al., 2018). Zum Beispiel,
to have a power of 80% to detect small within-subject differences ((cid:1)2 = 0.02) using within-
subjects repeated measurements ANOVA (4 measurements), one needs to include at least 69
Teilnehmer. The number of PWA should be even higher than this to deal with fewer measure-
gen, the large heterogeneity in the aphasia population, for example in lesion pattern and symp-
tomatology, and other confounds mentioned earlier. This number stands in great contrast with the
number of participants in the studies included in this review and in studies that are still being
published in the field of aphasia recovery, with few of them reaching this standard. Es ist, daher,
crucial to perform studies of aphasia recovery in larger samples of patients to ensure sufficient
statistical power to demonstrate a treatment effect, to allow for generalization to other (sub)
Populationen, and to account for methodological limitations (z.B., smoothing and the need to
correct for multiple comparisons; see Poldrack, Mumford, & Nichols, 2011). Because large scale
studies in a population of patients of aphasia are practically very challenging, data sharing might
be a suitable alternative to increase sample size. We refer the reader to Meyer (2018) Und
Poldrack and Gorgolewski (2014) for an overview of data-sharing efforts in the field of fMRI
and practical tips for ethical data sharing, jeweils.
Treatment-related effects outside the language networks
Our review has shown that recovery from aphasia engages brain regions that are outside those
traditionally associated with language functions (Figur 6). Such effects might relate to other
functions (such as cognitive control) that are not specific to language but indirectly support
Sprache. It has already been suggested that neural activity in the right IFG, a region that has been
consistently found to be involved in language recovery (Turkeltaub, Messing, Norise, &
Hamilton, 2011), including in our own review, is more related to top-down cognitive control by
a cingulo-opercular network than to a dynamic language-specific process (Geranmayeh,
Neurobiology of Language
426
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Brownsett, & Wise, 2014). Given the overlap between the language network and several non-
language processes, such as cognitive control or episodic memory—see, Zum Beispiel, Chein
and Schneider (2005), Geranmayeh, Brownsett, and Wise (2014), and Humphreys and Lambon
Ralph (2015)—it seems worthwhile to characterize the function, importance, and language
specificity of other regions that have been consistently identified in aphasia recovery, einschließlich
subcortical brain regions. The relative contribution of these regions to language recovery com-
pared with the perisylvian regions traditionally associated with linguistic processing (Geschwind,
1970; Hickok & Kacke, 2007) is presumably even more important than those we can derive
from the results of this review. Approximately one-third of the included studies applied an ROI
Analyse, which in half of them, focused on exploring the role of language regions in the left
hemisphere and their right-hemispheric counterparts in the response to aphasia interventions.
daher, we believe that whole-brain analyses are required to fully understand the neural
correlates of the cognitive mechanisms that support treatment-related language recovery.
Darüber hinaus, it might be interesting to further explore the effect of targeting these cognitive functions
during language interventions on aphasia recovery (Cahana-Amitay & Albert, 2015). Für
Beispiel, it would be useful to know whether these treatments (z.B., working memory training)
are as effective as language interventions at boosting language recovery and whether PWA
could benefit from a combination of both.
Connectivity studies
Another striking finding is that the majority of the included research (around 80%) focused on
regional changes in the brain, while only seven studies investigated treatment-related changes
in connectivity patterns. As language and other cognitive functions are linked to large-scale
neural networks, formed by interconnected cortical and subcortical areas, stroke-induced
damage to that network can lead to distributed dysfunction far beyond the lesion (Fuhrmann,
Shulman, & Corbetta, 2012). In der Tat, experimental animal studies have shown that stroke
unavoidably affects the brain connectome within minutes of onset (Silasi & Murphy, 2014).
daher, treatment-related (or spontaneous) regional alterations should be considered in
the context of these brain-wide connections that might explain more of the recovery process
in PWA than considering discrete brain regions in isolation. Due to recent advances in neuroim-
aging and computational sciences, more and more studies could and should explore treatment-
related brain changes on the connectome level, structurally as well as functionally. Wieder, für
these kinds of analyses, a large sample size is very important. Zusätzlich, analyses on the network
level might be more sensitive to reveal distinct effects of interventions targeting different linguistic
components. It is very likely that treatments, to some extent, do not differ in the location of their
Wirkung, but rather in the specific distribution of neural activity over the (same) language network, als
well as other supporting cognitive networks. Zusätzlich, one can investigate functional connec-
tivity patterns using resting-state fMRI, which has several advantages. Erste, it requires minimal
participation, which makes it clinically more interesting compared with task-based fMRI, espe-
cially in the case of severe aphasia or in the acute phase post-stroke. Zweite, the interpretation of
functional connectivity is not complicated by different task strategies, as in task-based fMRI
(Siegel, Shulman, & Corbetta, 2017). Zusammenfassend, investigating connectivity patterns offers a bet-
ter understanding of the effects of the lesion on the networks in the brain (extended lesion effects),
as well as a better understanding of how treatment influences the interaction in and between
specific distributed networks (Sprache, but also memory, executive function, usw.). We contend
that a regional approach and a connectivity approach are complementary methods that together
can answer different research questions (d.h., whether we are interested in treatment effects on
specific regions or on how they work together to create language).
Neurobiology of Language
427
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Individual variability as meaningful information
Approximately half of the included studies included a control group of healthy adults, mostly to
provide normative data on (repeated) task-specific brain response patterns. The results for the
healthy control group are therefore collapsed across the subject dimension and only the mean
effects are considered. In the context of personalized medicine, it might be interesting not to treat
the between-subject variance in the neuro-anatomical representation of language as noise, Aber
rather as meaningful information. After all, interindividual variability in a healthy population
(z.B., differences in learning or cognitive strategies to perform a given task) could explain differ-
ences between subjects in the speed and amount of aphasia recovery (Seghier & Price, 2018). Als
Seghier and Price (2018) vorschlagen, we could derive the likelihood of recovery of PWA from the
amount of variability in the functional response in a normal population. In a healthy population,
there are multiple ways to perform a certain (Sprache) Aufgabe, and therefore, PWA should be able
to compensate for a problem in that specific language task. Seghier and Price (2018) propose
the method of covariance analysis to characterize interindividual variability, which typically is
masked in group analyses. Jedoch, once again, this requires a large number of observations
from a large number of individuals and thus a large study sample size.
Zusätzlich, measurement and characterization of interindividual differences at the neural level
in a heterogeneous group of PWA would enable the tailoring of appropriate interventions to every
individual. Zum Beispiel, in the context of transcranial direct-current stimulation, Shah-Basak et al.
(2015) demonstrated that patients with more extensive lesions in the frontal lobe benefited more
from left-hemispheric inhibition, while PWA with less frontal damage responded better to left-
hemispheric facilitation. Andererseits, in large databases of stroke patients with and without
aphasia such as the Predicting Language Outcome and Recovery after Stroke (PLORAS)-database
(Seghier et al., 2016), machine learning approaches could be used to estimate the response to
intervention in PWA. By comparing neuroimaging data of a new PWA with neuroimaging data
of the stroke database and the treatment outcomes, the system could learn which treatment is
generally effective for PWA with similar neuroimaging characteristics. This could be done with
lesion data, as well as with structural and functional connectivity patterns (Silasi & Murphy, 2014).
Abschluss
Across treatments and participants, the regions that are involved in language recovery are very
diverse. Similarity between intervention studies targeting the same linguistic component is not
apparently greater than similarity between intervention studies targeting different linguistic compo-
nen. Jedoch, methodological shortcomings and variability between studies make it hard to
present clear conclusions on treatment-related brain changes in PWA. It is possible that treatment-
related brain changes associated with recovery of language after brain damage entail both regions
traditionally involved in linguistic processing as well as regions involved in other cognitive functions
in both hemispheres. If this is true, we should interpret recovery from aphasia as the result of the
adaptive reorganization of functionally heterogeneous perilesional and bilateral neural networks
not uniquely involved in language processing (Cahana-Amitay & Albert, 2015). daher, we argue
for the interpretation of treatment-related language recovery in light of the concept of neural multi-
Funktionalität (as discussed in the review of Cahana-Amitay & Albert, 2015). This label highlights the
constant and dynamic interactions between neural networks supporting linguistic as well as non-
linguistic functions, such as cognitive, emotional, and sensorimotor processing. Mit anderen Worten, Die
language network is most likely widely distributed over many different functionally and structurally
connected brain regions that are activated interactively. Specific linguistic functions are fulfilled
through the integration of neural activity in many regions subserving many functions (Price, 2012).
Neurobiology of Language
428
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
ACKNOWLEDGMENTS
The Fonds Wetenschappelijk Onderzoek, Flanders, Belgien, provided funding for this project,
including a grant from the Strategic Basic Research Programme.
FUNDING INFORMATION
Klara Schevenels, Fonds Wetenschappelijk Onderzoek (http://dx.doi.org/10.13039/
501100003130), Award ID: 1S81620N. Maaike Vandermosten, Fonds Wetenschappenlijk
Onderzoek (http://dx.doi.org/10.13039/501100003130), Award ID: 1521819N.
BEITRÄGE DES AUTORS
Klara Schevenels and Maaike Vandermosten conceived and planned the review. Klara
Schevenels took the lead in writing the manuscript. All authors provided critical feedback
and helped shape the review.
VERWEISE
Abel, S., Weiller, C., Huber, W., & Willmes, K. (2014). Neural under-
pinnings for model-oriented therapy of aphasic word production.
Neuropsychologie, 57(1), 154–165. https://doi.org/10.1016/j.
neuropsychologia.2014.03.010
Abel, S., Weiller, C., Huber, W., Willmes, K., & Specht, K. (2015).
Therapy-induced brain reorganization patterns in aphasia. Gehirn,
138(4), 1097–1112. https://doi.org/10.1093/brain/awv022
Ackermann, H., & Riecker, A. (2010). The contribution(S) of the
insula to speech production: A review of the clinical and functional
imaging literature. Brain Structure and Function, 214(5–6), 419–433.
https://doi.org/10.1007/s00429-010-0257-x
Aerts, A., Batens, K., Santens, P., van Mierlo, P., Hartsuiker, R.,
Hemelsoet, D., . . . De Letter, M. (2015). Aphasia therapy early after
stroke: Behavioural and neurophysiological changes in the acute
and post-acute phases. Aphasiology, 29(7), 845–871. https://doi.
org/10.1080/02687038.2014.996520
Baum, S. R., & Pell, M. D. (1999). The neural bases of prosody:
Insights from lesion studies and neuroimaging. Aphasiology, 13(8),
581–608. https://doi.org/10.1080/026870399401957
Benjamin, M. L., Towler, S., Garcia, A., Park, H., Sudhyadhom, A.,
Harnish, S., . . . Crosson, B. (2014). A behavioral manipulation en-
g a g e s r i g h t
f r o n t a l c o r t e x d u r i n g a p h a s i a t h e r a p y .
Neurorehabilitation and Neural Repair, 28(6), 545–553. https://
doi.org/10.1177/1545968313517754
Bonakdarpour, B., Beeson, P. M., Demarco, A. T., & Rapcsak, S. Z.
(2015). Variability in blood oxygen level dependent (BOLD) signal
in patients with stroke-induced and primary progressive aphasia.
NeuroImage: Klinisch, 8, 87–94. https://doi.org/10.1016/j.
nicl.2015.03.014
Bonakpardour, B., Parrish, T. B., & Thompson, C. K. (2007).
Hemodynamic response function in patients with stroke-induced
aphasia: Implications for fMRI data analysis. NeuroImage, 36(2),
322–331. https://www.sciencedirect.com/science/article/abs/pii/
S1053811907001371?via%3Dihub
Bonilha, L., Gleichgerrcht, E., Nesland, T., Rorden, C., & Fridriksson, J.
(2016). Success of anomia treatment in aphasia is associated with
preserved architecture of global and left temporal lobe structural
Netzwerke. Neurorehabilitation and Neural Repair, 30(3), 266–279.
https://journals.sagepub.com/doi/10.1177/1545968315593808?
url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%
3dpubmed
Bonilha, L., Hillis, A. E., Hickok, G., den Ouden, D. B., Rorden, C.,
& Fridriksson, J. (2017). Temporal lobe networks supporting the
comprehension of spoken words. Gehirn, 140(9), 2370–2380.
https://doi.org/10.1093/brain/awx169
Boyke, J., Driemeyer, J., Gaser, C., Buchel, C., & Mai, A. (2008).
Training-induced brain structure changes in the elderly. Zeitschrift
der Neurowissenschaften, 28(28), 7031–7035. https://doi.org/10.1523/
JNEUROSCI.0742-08.2008
Brady, M., Kelly, H., Godwin, J., Enderby, P., & Campbell, P. (2016).
Speech and language therapy for aphasia following stroke.
Cochrane Database of Systematic Reviews, 6, 4–7. https://doi.
org/10.1002/14651858.CD000425.pub4
Breier, J. ICH., Juranek, J., & Papanicolaou, A. C. (2011). Changes in maps
of language function and the integrity of the arcuate fasciculus
after therapy for chronic aphasia. Neurocase, 17(6), 506–517.
https://doi.org/10.1080/13554794.2010.547505
Brownsett, S. L. E., Warren, J. E., Geranmayeh, F., Woodhead, Z.,
Leech, R., & Wise, R. J. S. (2014). Cognitive control and its impact
on recovery from aphasic stroke. Gehirn, 137(1), 242–254. https://
doi.org/10.1093/brain/awt289
Cabeza, R., Anderson, N. D., Locantore, J. K., & McIntosh, A. R.
(2002). Aging gracefully: Compensatory brain activity in high-
performing older adults. NeuroImage, 17(3), 1394–1402. https://
doi.org/10.1006/nimg.2002.1280
Cahana-Amitay, D., & Albert, M. L. (2015). Neuroscience of aphasia
recovery: The concept of neural multifunctionality. Current
Neurology and Neuroscience Reports, 15(7). https://doi.org/
10.1007/s11910-015-0568-7
Fuhrmann, A. R., Shulman, G. L., & Corbetta, M. (2012). Why use a
connectivity-based approach to study stroke and recovery of
Funktion? NeuroImage, 62(4), 2271–2280. https://doi.org/10.1016/
j.neuroimage.2012.02.070
Catani, M., Jones, D. K., & Ffytche, D. H. (2005). Perisylvian
language networks of the human brain. Annals of Neurology,
57(1), 8–16. https://doi.org/10.1002/ana.20319
Neurobiology of Language
429
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Höhle, A. E., & Trimble, M. R. (2006). Der Precuneus: Eine Rezension von
its functional anatomy and behavioural correlates. Gehirn, 129(3),
564–583. https://doi.org/10.1093/brain/awl004
Chein, J. M., & Schneider, W. (2005). Neuroimaging studies of
practice-related change: fMRI and meta-analytic evidence of a
domain-general control network for learning. Cognitive Brain
Forschung, 25(3), 607–623. https://doi.org/10.1016/j.cogbrainres.
2005.08.013
Cocquyt, E.-M., De Ley, L., Santens, P., Van Borsel, J., & De Letter, M.
(2017). The role of the right hemisphere in the recovery of stroke-
related aphasia: A systematic review. Journal of Neurolinguistics,
44, 68–90. https://doi.org/10.1016/j.jneuroling.2017.03.004
Cornelissen, K., Laine, M., Tarkiainen, A., Järvensivu, T., Martin,
N., & Salmelin, R. (2003). Adult brain plasticity elicited by anomia
treatment. Zeitschrift für kognitive Neurowissenschaften, 15(3), 444–461.
https://doi.org/10.1162/089892903321593153
Crinion, J., Holland, A. L., Copland, D. A., Thompson, C. K., &
Hillis, A. E. (2013). Neuroimaging in aphasia treatment research:
Quantifying brain lesions after stroke. NeuroImage, 73, 208–214.
https://doi.org/10.1016/j.neuroimage.2012.07.044
Crinion, J. T., & Leff, A. P. (2007). Recovery and treatment of aphasia
after stroke: Functional imaging studies. Aktuelle Meinung in
Neurologie, 20, 667–673.
Crosson, B., McGregor, K., Gopinath, K. S., Conway, T. W.,
Benjamin, M., Chang, Y. L., . . . White, K. D. (2007). Funktional
MRI of language in aphasia: A review of the literature and the meth-
odological challenges. Neuropsychology Review, 17(2), 157–177.
https://doi.org/10.1007/s11065-007-9024-z
Dahlberg, C., Hawley, L., Morey, C., Newman, J., Cusick, C. P., &
Harrison-Felix, C. (2006). Social communication skills in persons with
post-acute traumatic brain injury: Three perspectives. Brain Injury,
20(4), 425–435. https://doi.org/10.1080/02699050600664574
Eickhoff, S. B., Nichols, T. E., Laird, A. R., Hoffstaedter, F., Amunts,
K., Fuchs, P. T., . . . Eickhoff, C. R. (2016). Behavior, sensitivity, Und
power of activation likelihood estimation characterized by
massive empirical simulation. NeuroImage, 137, 70–85. https://
doi.org/10.1016/j.neuroimage.2016.04.072
Forkel, S. J., die Schotten, M. T., Dell’Acqua, F., Kalra, L., Murphy,
D. G. M., Williams, S. C. R., & Catani, M. (2014). Anatomical
predictors of aphasia recovery: A tractography study of bilateral
perisylvian language networks. Gehirn, 137(7), 2027–2039.
https://doi.org/10.1093/brain/awu113
Fridriksson, J. (2010). Preservation and modulation of specific left
hemisphere regions is vital for treated recovery from anomia in
stroke. Journal of Neurophysiology, 30(35), 11558–11564. https://
doi.org/10.1523/JNEUROSCI.2227-10.2010
Fridriksson, J., Hubbard, H. ICH., Hudspeth, S. G., Holland, A. L.,
Bonilha, L., Fromm, D., & Rorden, C. (2012). Speech entrain-
ment enables patients with Broca’s aphasia to produce fluent
Rede. Gehirn, 135(12), 3815–3829. https://doi.org/10.1093/
brain/aws301
Fridriksson, J., & Morrow, L. (2005). Cortical activation and language
task difficulty in aphasia. Aphasiology, 19(3–5), 239–250. https://
doi.org/10.1080/02687030444000714
Fridriksson, J., Morrow-Odom, L., Moser, D., Fridriksson, A., &
Baylis, G. (2006). Neural recruitment associated with anomia
treatment in aphasia. NeuroImage, 32(3), 1403–1412. https://
doi.org/10.1016/j.neuroimage.2006.04.194
Fridriksson, J., Moser, D., Bonilha, L., Morrow-Odom, K. L., Shaw,
H., Fridriksson, A., Baylis, G. C., & Rorden, C. (2007). Neuronal
correlates of phonological and semantic-based anomia treatment
in aphasia. Neuropsychologie, 45(8), 1812–1822. https://doi.org/
10.1016/j.neuropsychologia.2006.12.017
Fridriksson, J., Richardson, J. D., Fillmore, P., & Cai, B. (2012). Links
hemisphere plasticity and aphasia recovery. NeuroImage, 60(2),
854–863. https://doi.org/10.1016/j.neuroimage.2011.12.057
Friederici, A. D. (2011). The brain basis of language processing: Aus
structure to function. Physiological Reviews, 91(4), 1357–1392.
https://doi.org/10.1152/physrev.00006.2011
Geranmayeh, F., Brownsett, S. L. E., & Wise, R. J. S. (2014). Task-
induced brain activity in aphasic stroke patients: What is driving
recovery? Gehirn, 137(10), 2632–2648. https://doi.org/10.1093/
brain/awu163
Geschwind, N. (1965). Disconnexion syndromes in animal and
man: Part II. Gehirn, 88, 585–644. https://doi.org/10.1093/brain/
88.3.585
Geschwind, N. (1970). The organization of language and the
Gehirn. Wissenschaft, 170(3961), 940–944. https://doi.org/10.1126/
science.170.3961.940
Geschwind, N. (2010). Disconnexion syndromes in animal and man:
Teil I. Neuropsychology Review, 20, 127–157. Original work pub-
lished 1965. https://doi.org/10.1007/978-94-010-2093-0_8
Gili, T., Fiori, V., De Pasquale, G., Sabatini, U., Caltagirone, C., &
Marangolo, P. (2017). Right sensory-motor functional networks
subserve action observation therapy in aphasia. Brain Imaging
and Behavior, 11(5), 1397–1411. https://doi.org/10.1007/s11682-
016-9635-1
Gold, B. T., & Kertesz, A. (2000). Right hemisphere semantic pro-
cessing of visual words in an aphasic patient: Eine fMRT-Studie.
Brain and Language, 73(3), 456–465. https://doi.org/10.1006/
brln.2000.2317
Griffis, J. C., Nenert, R., Allendorfer, J. B., & Szaflarski, J. P. (2017).
Damage to white matter bottlenecks contributes to language
impairments after left hemispheric stroke. NeuroImage: Klinisch,
14, 552–565. https://doi.org/10.1016/j.nicl.2017.02.019
Hagoort, P. (2014). Nodes and networks in the neural architecture
for language: Broca’s region and beyond. Aktuelle Meinung in
Neurobiology, 28, 136–141. https://doi.org/10.1016/j.conb.
2014.07.013
Haldin, C., Acher, A., Kauffmann, L., Hueber, T., Cousin, E.,
Badin, P., . . . Baciu, M. (2018). Speech recovery and language
plasticity can be facilitated by Sensori-Motor Fusion training in
chronic non-fluent aphasia. A case report study. Clinical Linguistics
and Phonetics, 32(7), 595–621. https://doi.org/10.1080/02699206.
2017.1402090
Hartwigsen, G., & Saur, D. (2019). Neuroimaging of stroke recovery
from aphasia –Insights into plasticity of the human language
Netzwerk. NeuroImage, 190, 14–31. https://doi.org/10.1016/j.
neuroimage.2017.11.056
Heiss, W., & Thiel, A. (2006). A proposed regional hierarchy in
recovery of post-stroke aphasia. Brain and Language, 98, 118–123.
https://doi.org/10.1016/j.bandl.2006.02.002
Hickok, G., & Kacke, D. (2004). Dorsal and ventral streams: A
framework for understanding aspects of the functional anatomy
of language. Cognition, 92(1–2), 67–99. https://doi.org/10.1016/
j.cognition.2003.10.011
Hickok, G., & Kacke, D. (2007). The cortical organization of speech
Verarbeitung. Nature Reviews Neurology, 8, 393–402. https://doi.org/
10.1038/nrn2113
Honey, C. J., Spurns, O., Cammoun, L., Gigandet, X., Thiran, J. P.,
Meuli, R., & Hagmann, P. (2009). Predicting human resting-state
functional connectivity from structural connectivity. Verfahren
der Nationalen Akademie der Wissenschaften, 106(6), 2035–2040.
https://doi.org/10.1073/pnas.0811168106
Hosomi, A., Nagakane, Y., & Yamada, K. (2009). Assessment of
arcuate fasciculus with diffusion-tensor tractography may predict
Neurobiology of Language
430
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
the prognosis of aphasia in patients with left middle cerebral artery
infarcts. Diagnostic Neuroradiology, 51, 549–555. https://doi.org/
10.1007/s00234-009-0534-7
Humphreys, G. F., & Lambon Ralph, M. A. (2015). Fusion and fission
of cognitive functions in the human parietal cortex. Hirnrinde,
25(10), 3547–3560. https://doi.org/10.1093/cercor/bhu198
Jang, S. H. (2013). Diffusion tensor imaging studies on arcuate
fasciculus in stroke patients: Eine Rezension. Frontiers in Human
Neurowissenschaften, 7(November), 1–7. https://doi.org/10.3389/fnhum.
2013.00749
Jang, S. H., & Lee, H. D. (2014). Recovery of injured arcuate fascic-
ulus in the dominant hemisphere in a patient with an intracerebral
hemorrhage. American Journal of Physical Medicine and
Rehabilitation, 93(12), e15–e18. https://doi.org/10.1097/PHM.
0000000000000202
Jungblut, M., Huber, W., Mais, C., & Schnitker, R. (2014). Paving
the way for speech: Voice-training-induced plasticity in chronic
aphasia and apraxia of apeech—three single cases. Neuronal
Plasticity, 2014, 1–14. https://doi.org/10.1155/2014/841982
Kaye, R. C., & Cherney, L. R. (2016). Script templates: A practical
approach to script training in aphasia. Topics in Language Disorders,
36(2), 136–153. https://doi.org/10.1097/TLD.0000000000000086
Kiran, S., Meier, E. L., Kapse, K. J., & Glynn, P. A. (2015). Changes in
task-based effective connectivity in language networks following
rehabilitation in post-stroke patients with aphasia. Grenzen in
Human Neuroscience, 9(Juni), 1–20. https://doi.org/10.3389/
fnhum.2015.00316
Kurland, J., Balduin, K., & Tauer, C. (2010). Treatment-induced
neuroplasticity following intensive naming therapy in a case of
chronic Wernicke’s aphasia. Aphasiology, 24(6–8), 737–751.
https://doi.org/10.1080/02687030903524711
Lake, E. M. R., Bazzigaluppi, P., & Stefanovic, B. (2016). Funktional
magnetic resonance imaging in chronic ischaemic stroke.
Philosophical Transactions of the Royal Society B: Biologisch
Wissenschaften, 371(1705), 1–11. https://doi.org/10.1098/rstb.2015.0353
Leon, S. A., Rodriguez, A. D., & Rosenbek, J. C. (2015). Rechts
hemisphere damage and prosody. In A. M. Raymer & L.
Gonzalez-Rothi (Hrsg.), The Oxford Handbook of Aphasia and
Language Disorders, Oxford Handbooks Online. https://doi.org/
10.1093/oxfordhb/9780199772391.013.15
Leonard, C., Laird, L., Burianová, H., Graham, S., Grady, C., Simic, T.,
& Rochon, E. (2015). Behavioural and neural changes after a
“choice” therapy for naming deficits in aphasia: Preliminary
Erkenntnisse. Aphasiology, 29(4), 506–525. https://doi.org/10.1080/
02687038.2014.971099
Madan, C. R. (2015). Creating 3D visualizations of MRI data: A brief
guide. F1000Research, 466, 1–13. https://doi.org/10.12688/
f1000research.6838.1
Marchina, S., Zhu, L. L., Norton, A., Zipse, L., Wan, C. Y., &
Schlaug, G. (2011). Impairment of speech production predicted by
lesion load of the left arcuate fasciculus. Stroke, 42, 2251–2256.
https://doi.org/10.1161/STROKEAHA.110.606103
Marcotte, K., Adrover-Roig, D., Damien, B., de Préaumont, M.,
Généreux, S., Hubert, M., & Ansaldo, A. ICH. (2012). Therapy-
induced neuroplasticity in chronic aphasia. Neuropsychologie,
50(8), 1776–1786. https://doi.org/10.1016/j.neuropsychologia.
2012.04.001
Marcotte, K., & Ansaldo, A. ICH. (2010). The neural correlates of semantic
feature analysis in chronic aphasia: Discordant patterns according
to the etiology. Seminars in Speech and Language, 31(1), 52–63.
https://doi.org/10.1055/s-0029-1244953
Marcotte, K., Laird, L., Bitan, T., Meltzer, J. A., Graham, S. J.,
Leonard, C., & Rochon, E. (2018). Therapy-induced neuroplasticity
in chronic aphasia after phonological component analysis: A
matter of intensity. Frontiers in Neurology, 9(APR), 1–7. https://
doi.org/10.3389/fneur.2018.00225
Marcotte, K., Perlbarg, V., Marrelec, G., Benali, H., & Ansaldo, A. ICH.
(2013). Default-mode network functional connectivity in aphasia:
Therapy-induced neuroplasticity. Brain and Language, 124(1), 45–55.
https://doi.org/10.1016/j.bandl.2012.11.004
McKinnon, E. T., Fridriksson, J., Glenn, G. R., Jensen, J. H., Helpern,
J. A., Basilakos, A., . . . Bonilha, L. (2017). Structural plasticity of
the ventral stream and aphasia recovery. Annals of Neurology,
82(1), 147–151. https://doi.org/10.1002/ana.24983
Meinzer, M., Beeson, P. M., Cappa, S., Crinion, J., Kiran, S., Saur, D.,
. . . Thompson, C. K. (2013). Neuroimaging in aphasia treatment
Forschung: Consensus and practical guidelines for data analysis.
N e u r o I m a g e , 7 3 , 2 15 – 22 4 . h t t p s : / / d o i . o r g / 1 0 . 1 0 1 6 / J .
neuroimage.2012.02.058
Menenti, L., Gierhan, S. M. E., Segaert, K., & Hagoort, P. (2011).
Shared language: Overlap and segregation of the neuronal infra-
structure for speaking and listening revealed by functional MRI.
Psychological Science, 22(9), 1173–1182. https://doi.org/
10.1177/0956797611418347
Menke, R., Meinzer, M., Kugel, H., Deppe, M., Baumgärtner, A.,
Schiffbauer, H., . . . Breitenstein, C. (2009). Imaging short- Und
long-term training success in chronic aphasia. BMC Neuroscience,
10, 118. https://doi.org/10.1186/1471-2202-10-118
Meyer, M. N. (2018). Practical tips for ethical data sharing.
Advances in Methods and Practices in Psychological Science,
1(1), 131–144. https://doi.org/10.1177/2515245917747656
Murphy, T. H., & Corbett, D. (2009). Plasticity during stroke recovery:
From synapse to behaviour. Nature Reviews Neurowissenschaften, 10(12),
861–872. https://doi.org/10.1038/nrn2735
Nair, V. A., Raut, R. V., & Prabhakaran, V. (2017). Investigating the
blood oxygenation level-dependent functional MRI response to a
verbal fluency task in early stroke before and after hemodynamic
scaling. Frontiers in Neurology, 8(JUN). https://doi.org/10.3389/
fneur.2017.00283
Nardo, D., Holland, R., Leff, A. P., Price, C. J., & Crinion, J. T. (2017).
Less is more: Neural mechanisms underlying anomia treatment in
chronic aphasic patients. Gehirn, 140(11), 3039–3054. https://doi.
org/10.1093/brain/awx234
NIDCD. (2015). Fact sheet: Aphasia. NIH Pub. NEIN. 97-4257. PDF
Retrieved from https://www.nidcd.nih.gov/sites/default/files/
Documents/health/voice/Aphasia.pdf
Ogar, J., Slama, H., Dronkers, N., Amici, S., & Gorno-Tempini, M. L.
(2005). Apraxia of speech: An overview. Neurocase, 11(6), 427–432.
https://doi.org/10.1080/13554790500263529
Plowman, E., Hentz, B., & Ellis, C. (2012). Post-stroke aphasia prog-
nosis: A review of patient-related and stroke-related factors.
Journal of Evaluation in Clinical Practice, 18, 689–694. https://
doi.org/10.1111/j.1365-2753.2011.01650.x
Kacke, D. (2003). The analysis of speech in different temporal inte-
gration windows: Cerebral lateralization as “asymmetric sampling
in time.” Speech Communication, 41(1), 245–255. https://doi.org/
10.1016/S0167-6393(02)00107-3
Poldrack, R. A., & Gorgolewski, K. J. (2014). Making big data open:
Data sharing in neuroimaging. Naturneurowissenschaften, 17(11),
1510–1517. https://doi.org/10.1038/nn.3818
Poldrack, R. A., Mumford, J. A., & Nichols, T. E. (2011). Handbook
of functional MRI data analysis. Cambridge, Vereinigtes Königreich: Cambridge
Universitätsverlag.
Price, C. J. (2000). The anatomy of language: Contributions from
functional neuroimaging. Journal of Anatomy, 197(3), 335–359.
https://doi.org/10.1046/j.1469-7580.2000.19730335.x
Neurobiology of Language
431
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Price, C. J. (2010). The anatomy of language: Eine Rezension von 100 fMRT
studies published in 2009. Annals of the New York Academy
of Sciences, 1191, 62–88. https://doi.org/10.1111/j.1749-6632.
2010.05444.X
Price, C. J. (2012). A review and synthesis of the first 20 years of PET
and fMRI studies of heard speech, spoken language and reading.
NeuroImage, 62(2), 816–847. https://doi.org/10.1016/j.
neuroimage.2012.04.062
Price, C. J., & Crinion, J. (2005). The latest on functional imaging
studies of aphasic stroke. Current Opinion in Neurology, 18(4),
429–434. https://doi.org/10.1097/01.wco.0000168081.76859.c1
Price, C. J., Crinion, J., & Friston, K. J. (2006). Design and analysis of
fMRI studies with neurologically impaired patients. Zeitschrift für
Magnetic Resonance Imaging, 23, 816–826. https://doi.org/
10.1002/jmri.20580
Price, C. J., Seghier, M. L., & Leff, A. P. (2010). Predicting language
outcome and recovery after stroke: The PLORAS system. Natur
Reviews Neurology, 6(4), 202–210. https://doi.org/10.1038/
nrneurol.2010.15
Pulvermüller, F., & Berthier, M. L. (2008). Aphasia therapy on a
neuroscience basis. Aphasiology, 22(6), 563–599. https://doi.
org/10.1080/02687030701612213
Pulvermüller, F., Neininger, B., Elbert, T., Mohr, B., Rockstroh, B.,
Koebbel, P., & Taub, E. (2001). Constraint-induced therapy of
chronic aphasia after stroke. Stroke, 32, 2–7. https://doi.org/
10.1161/01.STR.32.7.1621
Raboyeau, G., De Boissezon, X., Marie, N., Balduyck, S., Puel, M.,
Bézy, C., . . . Cardebat, D. (2008). Right hemisphere activation in
recovery from aphasia: Lesion effect or function recruitment?
Neurologie, 70 , 29 0–298. https://doi.org/10.1212/01.
wnl.0000287115.85956.87
Rachel, M. E., Fiez, J. A., Videen, T. O., MacLeod, A.-M. K., Pardo, J. V.,
Fuchs, P. T., & Petersen, S. E. (1994). Practice-related changes in human
brain functional anatomy during nonmotor learning. Zerebral
Kortex, 4(1), 8–26. http://dx.doi.org/10.1093/cercor/4.1.8
Ramus, F., Altarelli, ICH., Jednoróg, K., Zhao, J., & Scotto di Covella, L.
(2018). Neuroanatomy of developmental dyslexia: Pitfalls and
promise. Neuroscience and Biobehavioral Reviews, 84, 434–452.
https://doi.org/10.1016/j.neubiorev.2017.08.001
Rapp, B., Caplan, D., Edwards, S., Visch-Brink, E., & Thompson, C. K.
(2013). Neuroimaging in aphasia treatment research: Issues of
experimental design for relating cognitive to neural changes.
NeuroImage, 73, 200–207. https://doi.org/10.1016/j.neuroimage.
2012.09.007
Rasmussen, T., & Milner, B. (1977). The role of early left-brain injury
in determining lateralization of cerebral speech functions. Annals
of the New York Academy of Sciences, 299(1), 355–369. https://
doi.org/10.1111/j.1749-6632.1977.tb41921.x
Rochon, E., Leonard, C., Burianova, H., Laird, L., Soros, P.,
Graham, S., & Grady, C. (2010). Neural changes after phonolog-
ical treatment for anomia: Eine fMRT-Studie. Brain and Language,
114(3), 164–179. https://doi.org/10.1016/j.bandl.2010.05.005
Sandberg, C. W., Bohland, J. W., & Kiran, S. (2015). Changes in
functional connectivity related to direct training and generali-
zation effects of a word finding treatment in chronic aphasia.
Brain and Language, 150, 103–116. https://doi.org/10.1016/j.
bandl.2015.09.002
Santhanam, P., Duncan, E. S., & Small, S. L. (2018). Therapy-induced
plasticity in chronic aphasia is associated with behavioral improve-
ment and time since stroke. Gehirnkonnektivität, 8(3),
179–188. https://doi.org/10.1089/brain.2017.0508
Saur, D., & Hartwigsen, G. (2012). Neurobiology of language recovery
after stroke: Lessons from neuroimaging studies. Archives of Physical
Medicine and Rehabilitation, 93(1), S15–S25. https://doi.org/
10.1016/j.apmr.2011.03.036
Saur, D., Kreher, B. W., Schnell, S., Kummerer, D., Kellmeyer, P., Vry,
M.-S., . . . Weiller, C. (2008). Ventral and dorsal pathways for
Sprache. Verfahren der Nationalen Akademie der Wissenschaften, 105(46),
18035–18040. https://doi.org/10.1073/pnas.0805234105
Saur, D., Lange, R., Baumgaertner, A., Schraknepper, V., Willmes,
K., Rijntjes, M., & Weiller, C. (2006). Dynamics of language
reorganization after stroke. Gehirn, 129(6), 1371–1384. https://
doi.org/10.1093/brain/awl090
Schlaug, G., Marchina, S., & Norton, A. (2008). From singing to
Apropos: Why singing may lead to recovery of expressive language
function in patients with Broca’s aphasia. Musikwahrnehmung, 25(4),
315–323. https://doi.org/10.1525/MP.2008.25.4.315
Schlaug, G., Marchina, S., & Norton, A. (2009). Evidence for
plasticity in white-matter tracts of patients with chronic Broca’s
aphasia undergoing intense intonation-based speech therapy.
Annalen der New York Academy of Sciences, 1169, 385–394.
https://doi.org/10.1111/j.1749-6632.2009.04587.x
Scholz, J., Klein, M. C., Behrens, T. E. J., & Johansen-Berg, H. (2009).
Training induces changes in white-matter architecture. Natur
Neurowissenschaften, 12(11), 1370–1371. https://doi.org/10.1038/nn.2412
Segaert, K., Menenti, L., Weber, K., Petersson, K. M., & Hagoort, P.
(2012). Shared syntax in language production and language
comprehension —An fMRI study. Hirnrinde, 22(7), 1662–1670.
https://doi.org/10.1093/cercor/bhr249
Seghier, M. L., Patel, E., Prejawa, S., Ramsden, S., Selmer, A., Lim,
L., . . . Price, C. J. (2016). The PLORAS database: A data repos-
itory for predicting language outcome and recovery after stroke.
NeuroImage, 124, 1208–1212. https://doi.org/10.1016/j.
neuroimage.2015.03.083
Seghier, M. L., & Price, C. J. (2018). Interpreting and utilising intersub-
ject variability in brain function. Trends in den Kognitionswissenschaften, 22(6),
517–530. https://doi.org/10.1016/j.tics.2018.03.003
Shah-Basak, P. P., Norise, C., Garcia, G., Torres, J., Faseyitan, O., &
Hamilton, R. H. (2015). Individualized treatment with transcranial
direct current stimulation in patients with chronic non-fluent
aphasia due to stroke. Grenzen der menschlichen Neurowissenschaften, 9(201),
1–12. https://doi.org/10.3389/fnhum.2015.00201
Siegel, J. S., Ramsey, L. E., Snyder, A. Z., Metcalf, N. V., Chacko, R. V.,
Weinberger, K., . . . Corbetta, M. (2016). Disruptions of network
connectivity predict impairment in multiple behavioral domains
after stroke. Verfahren der Nationalen Akademie der Wissenschaften,
113 (3 0), E4367 –E4 376. https://doi.o rg/10 .10 73/pnas.
1521083113
Siegel, J. S., Shulman, G. L., & Corbetta, M. (2017). Measuring func-
tional connectivity in stroke: Approaches and considerations.
Journal of Cerebral Blood Flow and Metabolism, 37(8), 2665–2678.
https://doi.org/10.1177/0271678X17709198
Siegel, J. S., Snyder, A. Z., Ramsey, L., Shulman, G. L., & Corbetta, M.
(2016). The effects of hemodynamic lag on functional connectivity
and behavior after stroke. Journal of Cerebral Blood Flow and
Metabolism, 36(12), 2162–2176. https://doi.org/10.1177/
0271678X15614846
Silasi, G., & Murphy, T. H. (2014). Stroke and the connectome:
How connectivity guides therapeutic intervention. Neuron, 83(6),
1354–1368. https://doi.org/10.1016/j.neuron.2014.08.052
Stegemöller, E. L. (2017). The neuroscience of speech and language.
Music Therapy Perspectives, 35(2), 107–112. https://doi.org/
10.1093/mtp/mix007
Stevens, M. C., Kiehl, K. A., Pearlson, G. D., & Calhoun, V. D. (2009).
Brain network dynamics during error commission. Menschliches Gehirn
Mapping, 30(1), 24–37. https://doi.org/10.1002/hbm.20478
Neurobiology of Language
432
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
A review on treatment-related brain changes in aphasia
Stokes, R. C., Venezia, J. H., & Hickok, G. (2019). The motor system’s
[modest] contribution to speech perception. Psychonomic
Bulletin & Rezension, 26, 1354–1366. https://doi.org/10.3758/
s13423-019-01580-2
Tabei, K. ICH., Satoh, M., Nakano, C., Ito, A., Shimoji, Y., Kida, H., . . .
Tomimoto, H. (2016). Improved neural processing efficiency in a
chronic aphasia patient following melodic intonation therapy: A
neuropsychological and functional MRI study. Grenzen in
Neurologie, 7(SEP), 1–6. https://doi.org/10.3389/fneur.2016.00148
Thompson, C. K., den Ouden, D. B., Bonakdarpour, B., Garibaldi, K., &
Parrish, T. B. (2010). Neural plasticity and treatment-induced recovery
of sentence processing in agrammatism. Neuropsychologie, 48(11),
3211–3227. https://www.sciencedirect.com/science/article/abs/pii/
S0028393210002757?via%3Dihub
Thompson, C. K., Riley, E. A., den Ouden, D. B., Meltzer-Asscher, A., &
Lukic, S. (2013). Training verb argument structure production in agram-
matic aphasia: Behavioral and neural recovery patterns. Kortex,
49(9), 2358–2376. https://doi.org/10.1016/j.cortex.2013.02.003
Tourville, J. A., & Guenther, F. H. (2011). The DIVA model: Ein neuronaler
theory of speech acquisition and production. Language and
Cognitive Processes, 26(7), 1–27. https://www.tandfonline.com/
doi/abs/10.1080/01690960903498424
Tremblay, P., & Dick, A. S. (2016). Broca and Wernicke are dead, oder
moving past the classic model of language neurobiology. Brain and
Language, 162, 60–71. https://doi.org/10.1016/j.bandl.2016.08.004
Tsumoto, T. (1992). Long-term potentiation and long-term depres-
sion in the neocortex. Fortschritte in der Neurobiologie, 39(2), 209–228.
https://doi.org/10.1016/0301-0082(92)90011-3
Turkeltaub, P. E., Messing, S., Norise, C., & Hamilton, R. H. (2011).
Are networks for residual language function and recovery consis-
tent across aphasic patients? Neurologie, 76(20), 1726–1734.
https://doi.org/10.1212/ WNL.0b013e31821a44c1
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F.,
Etard, O., Delcroix, N., . . . Joliot, M. (2002). Automated anatomical
labeling of activations in SPM using a macroscopic anatomical par-
cellation of the MNI MRI single-subject brain. NeuroImage, 15(1),
273–289. https://doi.org/10.1006/nimg.2001.0978
van Hees, S., McMahon, K., Angwin, A., de Zubicaray, G., &
Copland, D. A. (2014). Neural activity associated with semantic
versus phonological anomia treatments in aphasia. Brain and
Language, 129(1), 47–57. https://doi.org/10.1016/j.bandl.2013.12.004
van Hees, S., McMahon, K., Angwin, A., de Zubicaray, G., Read, S.,
& Copland, D. A. (2014A). Changes in white matter connectivity
following therapy for anomia post stroke. Neurorehabilitation
and Neural Repair, 28(4), 325–334. https://doi.org/10.1177/
1545968313508654
van Hees, S., McMahon, K., Angwin, A., de Zubicaray, G., Read,
S., & Copland, D. A. (2014B). A functional MRI study of the re-
lationship between naming treatment outcomes and resting state
functional connectivity in post-stroke aphasia. Menschliches Gehirn
Mapping, 35(8), 3919–3931. https://doi.org/10.1002/hbm.22448
Varley, R. (2011). Rethinking aphasia therapy: A neuroscience
Perspektive. International Journal of Speech-Language Pathology,
13(1), 11–20. https://doi.org/10.3109/17549507.2010.497561
Vigneau, M., Beaucousin, V., Hervé, P. Y., Duffau, H., Crivello, F.,
Houdé, O., . . . Tzourio-Mazoyer, N. (2006). Meta-analyzing left
hemisphere language areas: Phonology, semantics, and sentence
Verarbeitung. NeuroImage, 30(4), 1414–1432. https://doi.org/
10.1016/j.neuroimage.2005.11.002
Vigneau, M., Beaucousin, V., Hervé, P. Y., Jobard, G., Petit, L.,
Crivello, F., . . . Tzourio-Mazoyer, N. (2011). What is right-
hemisphere contribution to phonological, lexico-semantic, Und
s e n t e n c e p r o c e s s i n g ? I n s i g h t s f r o m a m e t a – a n a l y s i s .
NeuroImage, 54(1), 577–593. https://doi.org/10.1016/j.
neuroimage.2010.07.036
Vitali, P., Abutalebi, J., Tettamanti, M., Danna, M., Ansaldo, A.-I.,
Perani, D., . . . Cappa, S. F. (2007). Training-induced brain
remapping in chronic aphasia: A pilot study. Neurorehabilitation
and Neural Repair, 21, 152–160. https://doi.org/10.1177/
1545968306294735
Vitali, P., Tettamanti, M., Abutalebi, J., Ansaldo, A., Perani, D.,
Cappa, S. F., . . . Perani, D. (2010). Generalization of the effects
of phonological training for anomia using structural equation
modelling: A multiple single-case study. Neurocase, 16(2), 93–105.
https://doi.org/10.1080/13554790903329117
Wan, C. Y., Zheng, X., Marchina, S., Norton, A., Schlaug, G.,
Laboratories, S. R., & Deaconess, B. ICH. (2014). Intensive therapy
induces contralateral white matter changes in chronic stroke
patients with Broca’s aphasia. Brain and Language, 136, 1–7.
https://doi.org/10.1016/j.bandl.2014.03.011
Wang, J., Marchina, S., Norton, A. C., Wan, C. Y., & Schlaug, G.
(2013). Predicting speech fluency and naming abilities in aphasic
Patienten. Grenzen der menschlichen Neurowissenschaften, 7(Dezember), 1–13.
https://doi.org/10.3389/fnhum.2013.00831
Warburton, E., Price, C. J., Swinburn, K., & Wise, R. J. S. (1999).
Mechanisms of recovery from aphasia: Evidence from positron
emission tomography studies. Zeitschrift für Neurologie, Neurochirurgie,
& Psychiatrie, 66, 155–161. https://jnnp.bmj.com/content/66/2/155
Wierenga, C. E., Maher, L. M., Moore, A. B., White, K. D.,
McGregor, K., Soltysik, D. A., . . . Crosson, B. (2006). Neuronal
substrates of syntactic mapping treatment: An fMRI study of two
Fälle. Journal of the International Neuropsychological Society,
12, 132–146. https://europepmc.org/article/med/16433953
Witteman, J., van Ijzendoorn, M. H., van de Velde, D., Transporter
Heuven, V. J. J. P., & Schiller, N. Ö. (2011). The nature of hemi-
spheric specialization for linguistic and emotional prosodic per-
ception: A meta-analysis of the lesion literature. Neuropsychologie,
49(13), 3722–3738. https://doi.org/10.1016/j.neuropsychologia.
2011.09.028
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of human
functional neuroimaging data. Nature Methods, 8(8), 665–670.
https://doi.org/10.1038/nmeth.1635
Yourganov, G., Fridriksson, J., Rorden, C., Gleichgerrcht, E., &
Bonilha, L. (2016). Multivariate connectome-based symptom
mapping in post-stroke patients: Networks supporting language
and speech. The Journal of Neuroscience, 36(25), 6668–6679.
https://doi.org/10.1523/JNEUROSCI.4396-15.2016
Neurobiology of Language
433
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
0
2
2
0
6
0
9
8
1
N
Ö
_
A
_
0
0
0
1
9
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3