REZENSION
Decoding the circuitry of consciousness: Aus
local microcircuits to brain-scale networks
Julien Modolo , Mahmoud Hassan , Fabrice Wendling
, and Pascal Benquet
University of Rennes, INSERM, LTSI–U1099, Rennes, Frankreich
Schlüsselwörter: Disorders of consciousness, Funktionale Konnektivität, Micro-circuitry, Communication
through coherence, Gating by inhibition, Elektroenzephalographie
ABSTRAKT
Identifying the physiological processes underlying the emergence and maintenance of
consciousness is one of the most fundamental problems of neuroscience, with implications
ranging from fundamental neuroscience to the treatment of patients with disorders of
Bewusstsein (DOCs). One major challenge is to understand how cortical circuits at
drastically different spatial scales, from local networks to brain-scale networks, operate in
concert to enable consciousness, and how those processes are impaired in DOC patients. In
this review, we attempt to relate available neurophysiological and clinical data with existing
theoretical models of consciousness, while linking the micro- and macrocircuit levels. Erste,
we address the relationships between awareness and wakefulness on the one hand, Und
cortico-cortical and thalamo-cortical connectivity on the other hand. Zweite, we discuss
the role of three main types of GABAergic interneurons in specific circuits responsible for
the dynamical reorganization of functional networks. Dritte, we explore advances in the
functional role of nested oscillations for neural synchronization and communication,
emphasizing the importance of the balance between local (high-frequency) and distant
(low-frequency) activity for efficient information processing. The clinical implications of
these theoretical considerations are presented. We propose that such cellular-scale
mechanisms could extend current theories of consciousness.
EINFÜHRUNG
Understanding how consciousness arises from communication among brain regions is a ques-
tion of the utmost importance in the field of neuroscience in general, and for the diagnosis and
treatment of patients suffering from disorders of consciousness (DOCs) insbesondere. The prob-
lem of consciousness can be seen as fundamental (z.B., “What is consciousness?” and “Why
do we have subjective, conscious experiences?”; such questions are referred to as the “hard”
problem of consciousness [Harnad, 1998]) or more empirical (z.B., “What are the processes
associated with the emergence and maintenance of consciousness?”; this forms the “soft” prob-
lem of consciousness [Harnad, 1998]). In this review, we aim at understanding (1) how brain
networks at different scales are involved in enabling and maintaining conscious processes of
information transmission and processing related to awareness and wakefulness, Und (2) Wie
these mechanisms are related to the disruptions of consciousness in DOC patients.
Many theories have been proposed to explain how consciousness originates, ranging from
abstract and informational concepts to neurophysiology-based theories. The most widespread
theories of consciousness have a fundamental assumption in common: information process-
ing in the human brain networks is inextricably linked with consciousness. A recent paper
Keine offenen Zugänge
Tagebuch
Zitat: Modolo, J., Hassan, M.,
Wendling, F., & Benquet, P. (2020).
Decoding the circuitry of
Bewusstsein: From local
microcircuits to brain-scale networks.
Netzwerkneurowissenschaften, 4(2), 315–337.
https://doi.org/10.1162/netn_a_00119
DOI:
https://doi.org/10.1162/netn_a_00119
Erhalten: 29 Juli 2019
Akzeptiert: 9 Dezember 2019
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Fabrice Wendling
fabrice.wendling@univ-rennes1.fr
Handling-Editor:
Olaf Sporns
Urheberrechte ©: © 2019
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Decoding the circuitry of consciousness
Disorders of consciousness:
Refers to pathological alterations of
the consciousness states, z.B., due to
brain trauma.
by Dehaene and colleagues summarizes this principle as follows (Dehaene, Lau, & Kouider,
2017): “What we call ‘consciousness’ results from specific types of information-processing
computations, physically realized by the hardware of the brain.”
In this review, we are mentioning three theories that appear to be the most prominent in the
field of consciousness research. Jedoch, the reader should be aware that numerous other the-
ories have been proposed, with different degrees of success. For the sake of this review focus-
ing on the micro- and macrocircuits involved in consciousness, we focus on those three main
theories, which can be linked with such considerations about brain network properties. Der
three main theories of consciousness are the Integrated Information Theory (IIT; Tononi, 2004),
the Dynamic Core Hypothesis (DCH; Tononi & Edelman, 1998), and the Global Workspace
Theory (GWT; Baars, 1988; Dehaene, Kerszberg, & Changeux, 1998; Dehaene, Sergent, &
Changeux, 2003). Historically, DCH theory has been the first to refer to the notion of informa-
tion processing involved in consciousness (Tononi & Edelman, 1998). This theory is based on
the central role of functional clusters in the thalamo-cortical system and reentrant interactions,
with high integration and differentiation of neuronal activity being crucial in the emergence
of conscious phenomena. IIT, which is an evolution and generalization of DCH, is based on
a set of axioms from which postulates are derived. IIT also provides a computable quantity,
Φ, also called integrated information, that quantifies the level of consciousness. In the case of
IIT, no specific physiological substrate has been suggested, since this theory is mainly focused
on understanding how consciousness arises from the integration of information among several
Systeme. In diesem Rahmen, if combining subelements increases information processing capa-
bility more than linearly adding these elements, then integrated information increases. GWT
is a theory of consciousness theory that is more directly connected with neurophysiology and
neuroanatomy. The main hypothesis of GWT is that information related to conscious processes
is globally available within the brain, and that two fundamentally different computational sys-
tems co-exist: (1) a network of distributed “local” processors operating in parallel in the brain
(“unconscious”), Und (2) a “global” workspace formed by a network of distributed intercon-
nected cortical areas involved in conscious perception (Baars, 1988). In the remainder of this
Rezension, the distinction should be made between the concepts of “consciousness” and “con-
scious perception” in terms of subjective experience. The concept of consciousness is much
broader than conscious perception, and involves among others the capability to remember,
perceive, and report, as well as a sense of selfhood (Seth, 2018). Conscious perception refers
more specifically to the reportability of the focus of our perception (Naccache, 2005). Der
key concept here is that the global workspace is composed of distant regions densely con-
nected through glutamatergic cortico-cortical connections as opposed to the network of local
processors operating in “isolation” (in parallel). It is worth noting that this distinction between
unconscious and conscious processes has been recently challenged, and might be an oversim-
plification (Melnikoff & Bargh, 2018). In GWT, conscious perception is associated with “igni-
tion,” a large-scale brain activation pattern induced by exposure to a stimulus (Dehaene et al.,
2003). If the stimulus does not trigger ignition, and if the induced brain response remains spa-
tially confined and is brief, then the perception will not reach consciousness. Mit anderen Worten, A
stimulus has to be sufficiently long and strong to reach consciousness, which suggests a form of
filtering mechanism that is consistent with the view that only a limited amount of information
effectively enters in the global workspace. Despite these successes in accounting for experi-
mental data in humans regarding subliminal (unconscious) and conscious perception (King,
Pescetelli, & Dehaene, 2016; Sergent & Dahaene, 2004), one drawback of GWT is that it does
not explicitly relate the large-scale recruitment of brain regions during conscious access with
cellular mechanisms. More precisely, what prevents ignition for short, irrelevant stimuli? And
Netzwerkneurowissenschaften
316
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Decoding the circuitry of consciousness
umgekehrt, what enables ignition for strong stimuli? The neuroanatomical, neurophysiologi-
cal and dynamical mechanisms behind ignition are of fundamental importance to understand-
ing how we become conscious of a stimulus, or how alterations of brain networks can result
in impaired consciousness in DOC patients. Since GWT and DCH are the two theories that
focus the most on the neurophysiological substrates of consciousness, and focus respectively
on cortico-cortical connectivity and thalamo-cortical connectivity, which are two key issues
detailed in this review, we are only discussing those two theories in the remainder of this
Rezension.
If one accepts that consciousness is associated with a sufficiently complex (in the algo-
rithmic sense of “less compressible”) information processing, then the emergence of con-
sciousness is critically dependent on three factors: (1) a physical network enabling interactions
between its components; (2) the flexibility to reorganize transiently subnetworks to achieve
greater computation capabilities by increasing the number of possible configurations and
input-output functions, through functional connectivity; Und (3) dynamic communications be-
tween its components. These three critical components have the potential to be altered, für
Beispiel, in lesions following traumatic brain injury. Although the physical large-scale network
linking brain regions is well defined and known as the connectome, there are still unresolved
questions regarding the transient organization of clusters performing specific computations
(functional networks), the associated means of communication (neural coding), and how large-
scale functional brain networks and information routing can reconfigure rapidly depending on
microcircuit regulation.
Whereas the main theories of consciousness introduced in this review have been focused
on the large-scale processes associated with consciousness, few efforts have been made to
understand how the microcircuit scale can help understand how large-scale coordinated ac-
tivity emerges. Bridging the micro- and macrocircuit levels appears indispensable to obtain
an integrated view of the physiological processes that underlie the maintenance of conscious-
ness (Changeux, 2017). daher, we formulate in this review the hypothesis that account-
ing for cellular-scale mechanisms could provide possible directions to extend current theories
of consciousness by encompassing several spatial scales of description. Such multiscale un-
derstanding is a prerequisite to understanding how brain networks become dysfunctional in
DOCs, and might contribute to reconciling GWT and DCH into a unified framework. More
specifically, we hypothesize that the co-occurrence of low- and high-frequency neuronal os-
cillations could provide distant coactivation (low-frequency, glutamatergic origin) that would
then enable binding (high-frequency, GABAergic origin). This would involve the formation
of a functional network of several brain regions through the low-frequency rhythm, prior to
information transfer and processing through gamma activity notably.
The review is organized as follows. Erste, we examine the relationship between the two di-
mensions of consciousness, namely wakefulness and awareness, with functional connectivity
between cortical regions and the thalamus. Zweite, we review the “means of communication”
enabling complex information processing linked with consciousness, which regulate cortico-
cortical communication, among which communication through coherence (CTC) and gating
by inhibition (GBI). The alteration of those mechanisms is presented through results from the
clinical literature. Dritte, we attempt to link these findings with concepts that have recently
emerged based on the communication between brain regions based on cross-frequency cou-
plings between oscillations with specific functional roles. Endlich, we suggest possible clinical
implications for this framework in terms of novel neuromodulation protocols in DOCs.
317
Functional networks:
Refers to a subset of brain regions for
which the temporal dynamics
exhibits some form of correlation
(z.B., phase synchronization) über
Zeit.
Communication through coherence:
An hypothesized mechanism of
communication between brain
regions through transient coherence
of gamma oscillations.
Gating by inhibition:
An hypothesized mechanism of
communication between brain
regions based on the inhibition of
task-irrelevant areas, enabling the
transient communication of
task-relevant areas.
Netzwerkneurowissenschaften
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Decoding the circuitry of consciousness
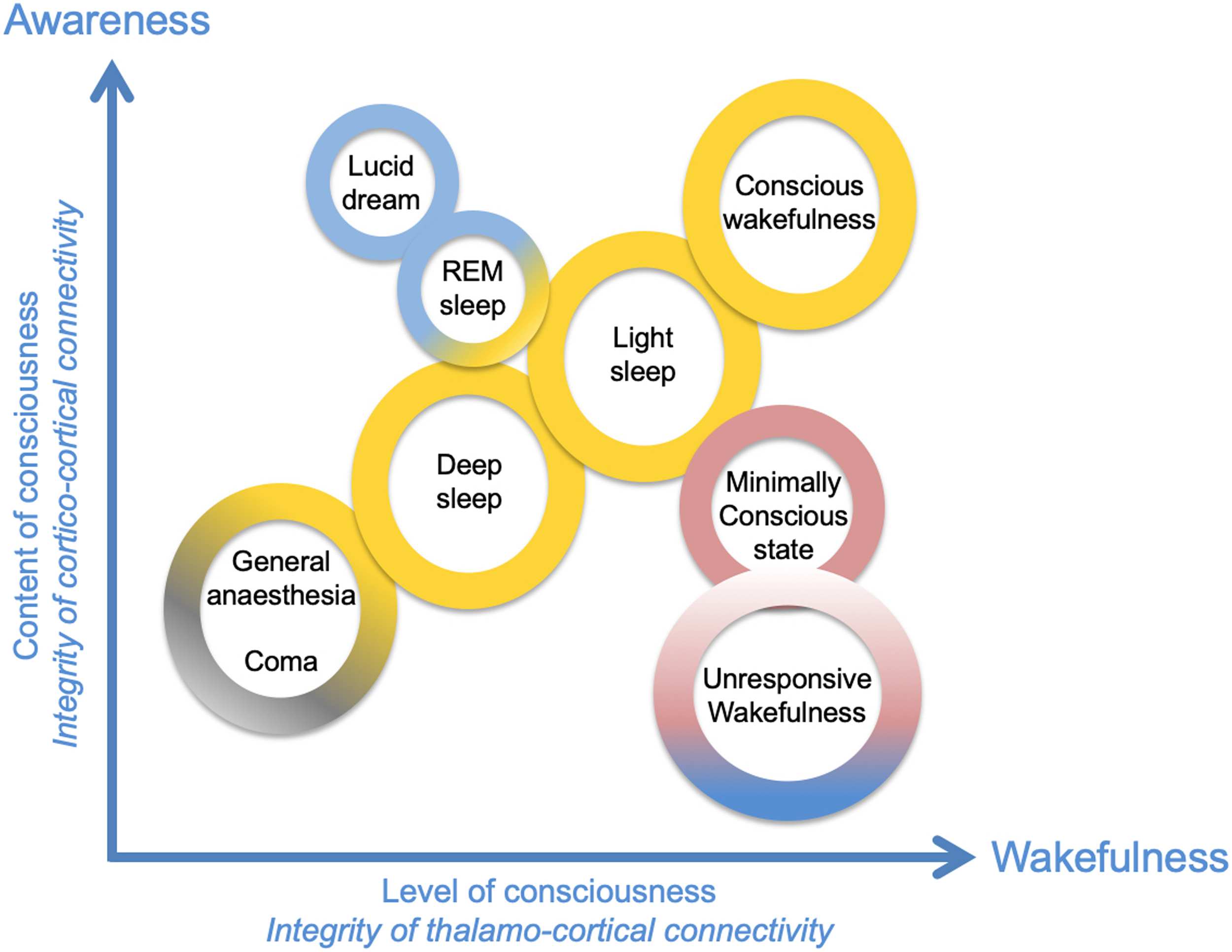
AWARENESS AND WAKEFULNESS: A SHORT REVIEW OF CONCEPTS
Conscious perception results from an interplay between two processes interacting with each
andere: awareness and wakefulness. Deep sleep switches awareness off, whereas being able of
conscious perception (awareness) of environmental stimuli usually implies a state of wakeful-
ness, as illustrated in Figure 1. Zum Beispiel, during general anesthesia, there is an absence of
conscious perception, das ist, awareness and wakefulness. In some peculiar cases, Jedoch,
these two components can be unrelated. Unresponsive wakefulness state (UWS) is an example
of a DOC in which wakefulness is present without any detectable signs of awareness (Laureys &
Boly, 2012). Auch, during lucid dreaming, there is a form of awareness in the absence of wake-
fulness (Während dem Schlafen) (Voss, Schermelleh-Engel, Windt, Frenzel, & Hobson, 2013). Ein anderer
example is spatial neglect syndrome, in which patients have no conscious awareness of visual
Reize, while being awake in the contralateral side of the cortical lesion (Le, Stojanoski, Khan,
Keough, & Niemeier, 2015). Let us mention that human consciousness is a multicomponent
concept extremely challenging to define, since it involves objective states and subjective ex-
perience, comprising different “contents” and “levels” of consciousness. Each author proposes
his or her own definition of consciousness, since these components are still under debate and
are not always precisely determined (Storm et al., 2017). Since the main goal of this article is
to discuss the neural correlates of consciousness (Crick & Koch, 1998), we restricted here the
notion of consciousness to the basic component of wakefulness (awake/sleep state, alertness,
vigilance) and awareness (content of conscious perception).
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 1. Wakefulness and awareness are two essential dimensions of consciousness. In diesem
diagram, several qualitatively different states of consciousness have been positioned on the two-
dimensional matrix as a function of the associated axes “content of consciousness” (awareness) Und
“level of consciousness” (Wachsamkeit). Adapted from Laureys (2005).
Netzwerkneurowissenschaften
318
Decoding the circuitry of consciousness
Awareness is supported by attentional, fronto-parietal networks that amplify synaptic con-
nections within specific cortical pathways (Tallon-Baudry, 2011). This amplification of relevant
stimuli enhances the activated network related to stimulus representation. In parallel, the con-
comitant inhibition of irrelevant surrounding networks (1) optimizes cortico-cortical routing
of information by constraining the possible propagation of neural activity throughout all pos-
sible cortical “routes,” which (2) restricts propagation to a limited number of stimulus-driven
Möglichkeiten, Und (3) increases the signal-to-noise ratio. Such mechanisms are related to the
concept of functional connectivity and are detailed further in this review. Wichtig, solch
mechanisms of active inhibition likely involve cortical inhibition with an active modulation
by thalamo-cortical inputs (Gabernet, Jadhav, Feldman, Carandini, & Scanziani, 2005), imply-
ing that the pattern of thalamo-cortical activity influences information processing in cortico-
cortical networks.
Wakefulness depends critically on thalamcortical connectivity and critical neuromodula-
tory brainstem inputs to the thalamus (including noradrenaline projections from the locus
coeruleus [Monti, 2011] and projections from the reticular formation system), the posterior
hypothalamus, and the thalamus itself (Lin, 2000). Zum Beispiel, during slow-wave sleep, A
low-frequency, synchronized activity between the cortex and thalamus (the so-called “up-and-
down” rhythm [Neske, 2015]) prevents transmission of subcortical inputs to the cortex during
schlafen. This provides an example in which thalamo-cortical activity drastically decreases infor-
mation processing by cortico-cortical networks, resulting in a loss of consciousness. Während
Wachsamkeit, thalamo-cortical activity is weakly synchronized (Gent, Bandarabadi, Herrera, &
Adamantidis, 2018), which is a necessary but not sufficient condition to enable consciousness.
Zum Beispiel, as aforementioned, wakefulness is present in UWS patients but cortico-cortical
communication is severely impaired (Noirhomme et al., 2010), interfering with the reportable
“awareness” component of consciousness. Another required condition for consciousness is
an efficient large-scale cortico-cortical communication that can support awareness through
the activation of attentional fronto-parietal networks (Luckmann, Jacobs, & Sack, 2014; Ptak,
Schnider, & Fellrath, 2017). daher, in terms of neuroanatomy, it is possible to link wake-
fulness with thalamo-cortical, “vertical” connectivity, while awareness depends on cortico-
cortical, “horizontal” connectivity. This is consistent with the recent view by Naccache (2018)
that minimally conscious state (MCS) Patienten, who are conscious to some degree, exhibit
“cortically mediated states,” whereas UWS patients do not exhibit such activity, presumably
because cortico-cortical connectivity (critical for awareness) is too severely impaired. More
specifically, a “critical mass” of information processing is required to occur for the emergence
and maintenance of consciousness, which is tightly regulated by thalamo-cortical and cortico-
cortical connectivity. Figur 1 presents, in a two-dimensional plane, the continuum of the states
of consciousness, as a function of awareness and wakefulness.
One popular paradigm in consciousness research that is used to investigate these two com-
ponents has been the use of reportability, das ist, instructing subjects when they perceive
consciously a stimulus, which enables the contrast of brain activity with/without a conscious
Bericht, supposedly pointing at key structures in conscious perception. This paradigm has led to
key insights on how information becomes conscious, as exemplified by works demonstrating
how “ignition” takes place in the brain, as formalized in the GWT. A stimulus presented very
briefly will lead to a short, localized activation, Zum Beispiel, in visual areas, and no perception
will be reported by the subject. If the stimulus is presented for a sufficiently long time, a large-
scale activation of brain networks will occur during an extended period of several hundreds
of milliseconds, leading to global information availability through ignition, and the subject
will be able to report the perception. Information that is processed by the brain but remains
Netzwerkneurowissenschaften
319
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Decoding the circuitry of consciousness
EEG (Elektroenzephalographie):
Neuroimaging technique consisting
in recording with scalp electrodes
the small currents induced by
neuronal activity at the cortex level.
confined and is not globally shared with other brain regions is then termed preconscious (Und
not unconscious, which would mean the complete absence of stimulus-related processing
by the brain). Endlich, let us mention that using reportability has been questioned (Tsuchiya,
Wilke, Frassle, & Lamme, 2015), since instructing subjects to report their perception induces
large prefrontal activation, Zum Beispiel, which is not directly linked with consciousness per se.
daher, using measures of consciousness not involving reportability should help reduce the
biases in estimating the minimal network of brain regions required to support consciousness
(the so-called NCC, or neural correlates of consciousness).
FUNCTIONAL NETWORKS: A FLEXIBLE ARCHITECTURE FOR CONSCIOUS PROCESSES
The main disadvantage of static network architectures is their limitation in terms of amount and
variety (complexity) of information processing that can take place. The notion of complexity
is central in the study of consciousness, therefore it has to be emphasized that numerous mea-
sures of complexity have been proposed and that no universal definition is satisfactory. Zum Beispiel-
reichlich, the Lampel-Ziv (LZ) complexity used to compute the Perturbational Complexity Index
consciousness metrics (Casali et al., 2013) is not fully appropriate: the Perturbational Complex-
ity Index is computed by applying the LZ algorithm on the TMS-evoked responses; Jedoch,
a fully random set of TMS-evoked responses would result in a maximal LZ complexity, welche
is obviously not realistic. Other measures of complexity have also been suggested to evaluate
the state of consciousness from EEG signals, such as multiscale entropy (Eagleman et al., 2018)
and Kolmogorov complexity (Ruffini, 2017), unter anderen (see Arsiwalla & Verschure, 2018,
for a detailed review on the topic). Despite this conceptual limitation of LZ complexity, völlig
random activity is likely unrealistic in the human brain, since there is always some degree of
temporal or spatial organization of neural activity due to the underlying micro-and macrocon-
nectivity. daher, in the limit of these theoretical limitations, LZ appears adapted to quantify
the complexity associated with the presence of conscious processes. An extensive study of
complexity metrics applied to the study of neural signals in various conditions of conscious-
ness would certainly move the field forward in this regard. Let us mention that all measures
of complexity do not suffer the limitation of being maximal for a random signal, which is an
issue discussed at length by Wang et al. (2017). Two different types of complexity measures
can be distinguished, so-called Type 1 measures of complexity linearly increase with the level
of randomness, whereas Type 2 measures increase with randomness, then reach a plateau, Sei-
fore decreasing and becoming null for a maximal randomness (bell-like curve). As suggested
by Wang et al. (2017), Typ 2 measures of complexity (such as fluctuation complexity) might
be better suited to quantify consciousness-related processes.
The brain takes advantage of different mechanisms that overcome this limitation by enabling
a dynamic, transient reconfiguration of brain networks increasing the repertoire of possible
responses to inputs (d.h., complexity of input-output functions) (Spurns, 2013). Such transient
networks involving only a few brain regions, coordinated to achieve a specific function limited
in duration, form what is termed functional connectivity. There is a growing interest regard-
ing the functional networks associated with specific cognitive tasks (Hassan et al., 2015), Und
novel frameworks have recently emerged (Avena-Koenigsberger, Misic, & Spurns, 2017) to ex-
plain how brain-scale anatomical connectivity relates to functional connectivity. Funktional
networks organize through the network “means of communication,” also termed communi-
cation dynamics, that govern the routing of information through specific networks, instead of
propagating information through the entire brain network (Avena-Koenigsberger et al., 2017).
If no such routing mechanisms were in place, any information generated by a single brain re-
gion would have the potential to induce phase synchronization in all anatomically connected
Netzwerkneurowissenschaften
320
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Decoding the circuitry of consciousness
Regionen (connectome), which in turn would result in brain-scale synchronized activity. Solch
synchronized activity at a large scale has a low informational content and complexity (z.B.,
in the case in seizure-induced loss of consciousness), and is therefore not compatible with
the maintenance of conscious processes. Zusammenfassend, the network means of communication
are critically linked with the neural states dynamical repertoire of the network, which is key
since conscious processes are associated with rich and diverse information processing among
large-scale brain networks. Let us mention that such key elements of brain networks function-
ing are likely not sufficient to enable the emergence of consciousness, but rather that they are
a necessary prerequisite. Zum Beispiel, considering peripheral body functions is likely impor-
tant to understand the emergence of consciousness, as highlighted in Azzalini, Rebollo, Und
Tallon-Baudry (2019).
More specifically, one fundamental question is, What are the mechanisms regulating com-
munication dynamics and enabling functional networks to emerge in brain-scale networks?
This question is central to understand how the brain optimizes its information processing ca-
pabilities, which are tightly linked with consciousness. We propose that the fundamental mech-
anisms underlying communication dynamics are actually cellular-scale mechanisms that (1)
prevent brain-scale neuronal synchronization following a stimulus, Und (2) enable the transient
coupling of specific distant brain regions. There has been a considerable amount of interest
for large-scale brain activity patterns linked with consciousness, since those can be measured
through various neuroimaging modalities (z.B., electroencephalography, functional magnetic
resonance imaging). Jedoch, mechanisms at the cellular scale have remained more chal-
lenging to address in humans for obvious reasons of invasiveness associated with the required
recording techniques in humans. In diesem Abschnitt, we review evidence for such mechanisms that
could bridge the microcircuit and brain-scale levels.
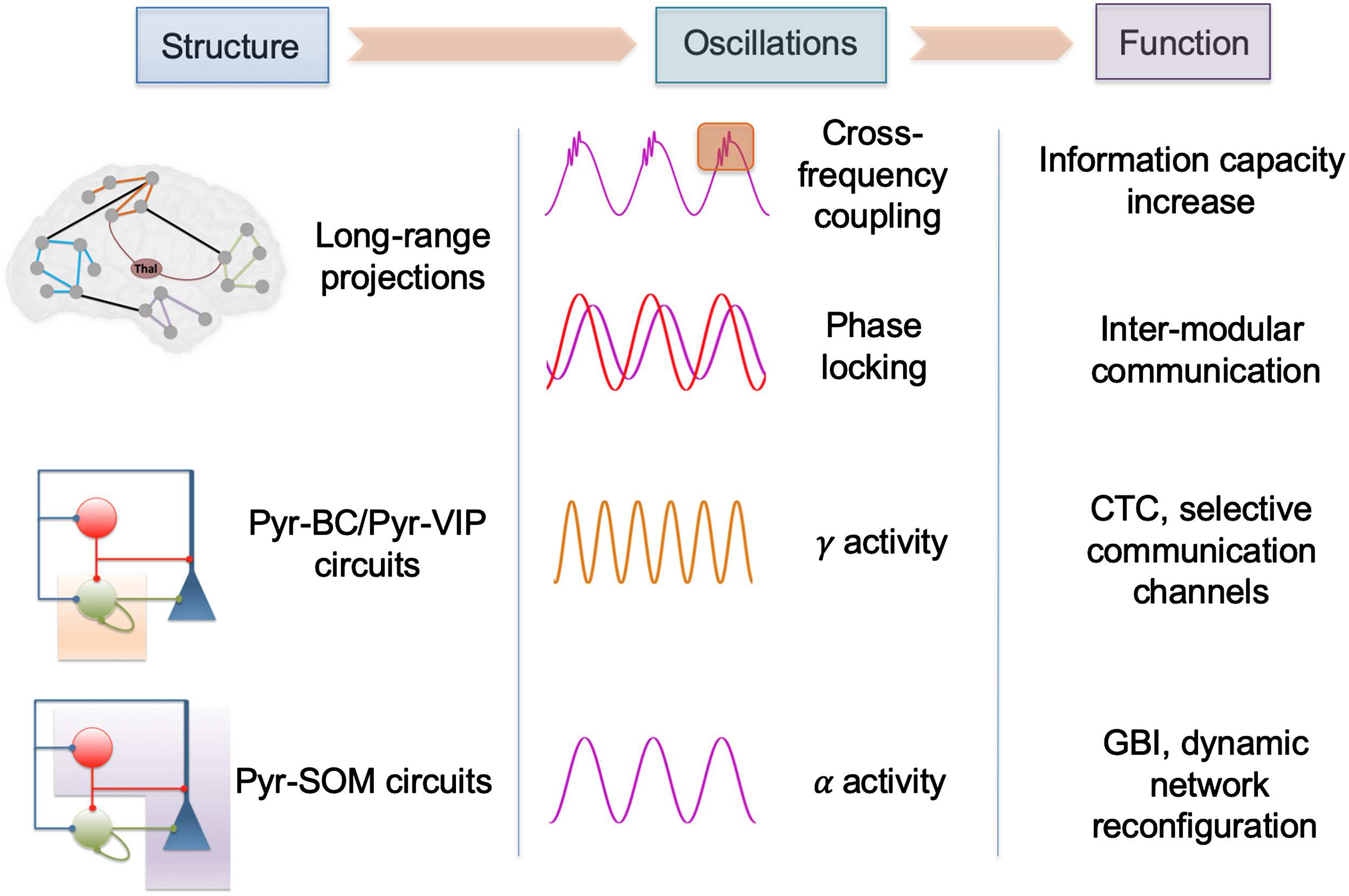
In terms of large-scale neuroanatomical pathways enabling consciousness, long-range glu-
tamatergic projections between pyramidal neurons through white matter fibers likely have a
critical role (Dehaene & Changeux, 2011) since they enable fast (because of myelin) com-
munication between distant regions. At the brain-scale level, these white matter fibers are
likely critical to enable conscious access, which involves the transient stabilization of neu-
ronal activity encoding a specific information pattern, in a network of high-level brain regions
interconnected by long-range connections, with the prefrontal cortex acting as a key node
(Berkovitch, Dehaene, & Gaillard, 2017; Dehaene, Changeux, Naccache, Sackur, & Sergent,
2006). On conscious trials, distributed gamma-band activity reflects a stabilization of local
information broadcasted to other areas. During conscious access to a specific information,
other surrounding global workspace neurons would be inhibited and unavailable for the pro-
cessing of other stimuli, therefore remaining preconscious (not reaching consciousness). In
addition to the characteristics of connectivity at the microcircuit scale, another key factor in
shaping neuronal oscillations is the presence of time delays, notably due to the finite con-
duction speed of action potentials along fibers. Time delays can result in the emergence and
stability of oscillations at specific frequencies (Petkoski & Jirsa, 2019; Petkoski, Verehrung, & Jirsa,
2018). daher, damage to cortico-cortical fibers not only impairs the transmission of activity
between regions, it has also the potential to alter the frequency content and stability of neural
oscillations, thereby impacting their function.
At the local scale, a microcircuit has also been identified as being involved in the com-
munication between distant brain regions: the projection from pyramidal neurons in a brain
region to VIP-positive (vasoactive intestinal peptide) neurons in another region. By activating
VIP-positive neurons in a distant region, this induces an inhibition of somatostatin-positive
Netzwerkneurowissenschaften
321
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Decoding the circuitry of consciousness
(SST) Neuronen, which target pyramidal cell dendrites, resulting in a disynaptic disinhibition
(Karnani et al., 2016). Through this disynaptic disinhibition, gamma activity generation can
occur through parvalbumin-parvalbumin (PV-PV) mutual inhibition, and binding between the
two involved regions can possibly take place (Munoz, Tremblay, Levenstein, & Rudy, 2017),
temporarily enabling information transfer and processing. These cellular-scale mechanisms are
summarized in Figure 2.
Few studies have addressed how traumatic brain injury (TBI) impacts specific cellular cell
types and how it could impact large-scale information processing in the brain. In animal mod-
els of TBI, a hyperexcitability induced in the long term (8–10 weeks posttreatment) (Allitt,
Iva, Yan, & Rajan, 2016; Alwis et al., 2016; Alwis, Yan, Morganti-Kossmann, & Rajan, 2012)
was linked to the selective reduction of dendrite-targeting inhibitory interneurons in that cortex
(Carron, Yan, Allitt, & Rajan, 2018). daher, one could suggest that TBI-induced loss of func-
tional dendritic-targeting interneurons will consequently affect both low-frequency oscilla-
tions generation and disinhibition through VIP interneurons. Darüber hinaus, in animal models, A
progressive loss of phasic inhibition with a corresponding loss of somatic-targeting PV+
GABAergic interneurons following TBI has also been shown (Cantu et al., 2015; Guerriero,
Giza, & Rotenberg, 2015), which might be at least partly responsible for impaired gamma os-
cillations and synchrony (C. Wang et al., 2017). The loss of PV+ interneurons has not only been
found in rodent models of TBI, but also in an immunochemistry study performed in postmortem
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
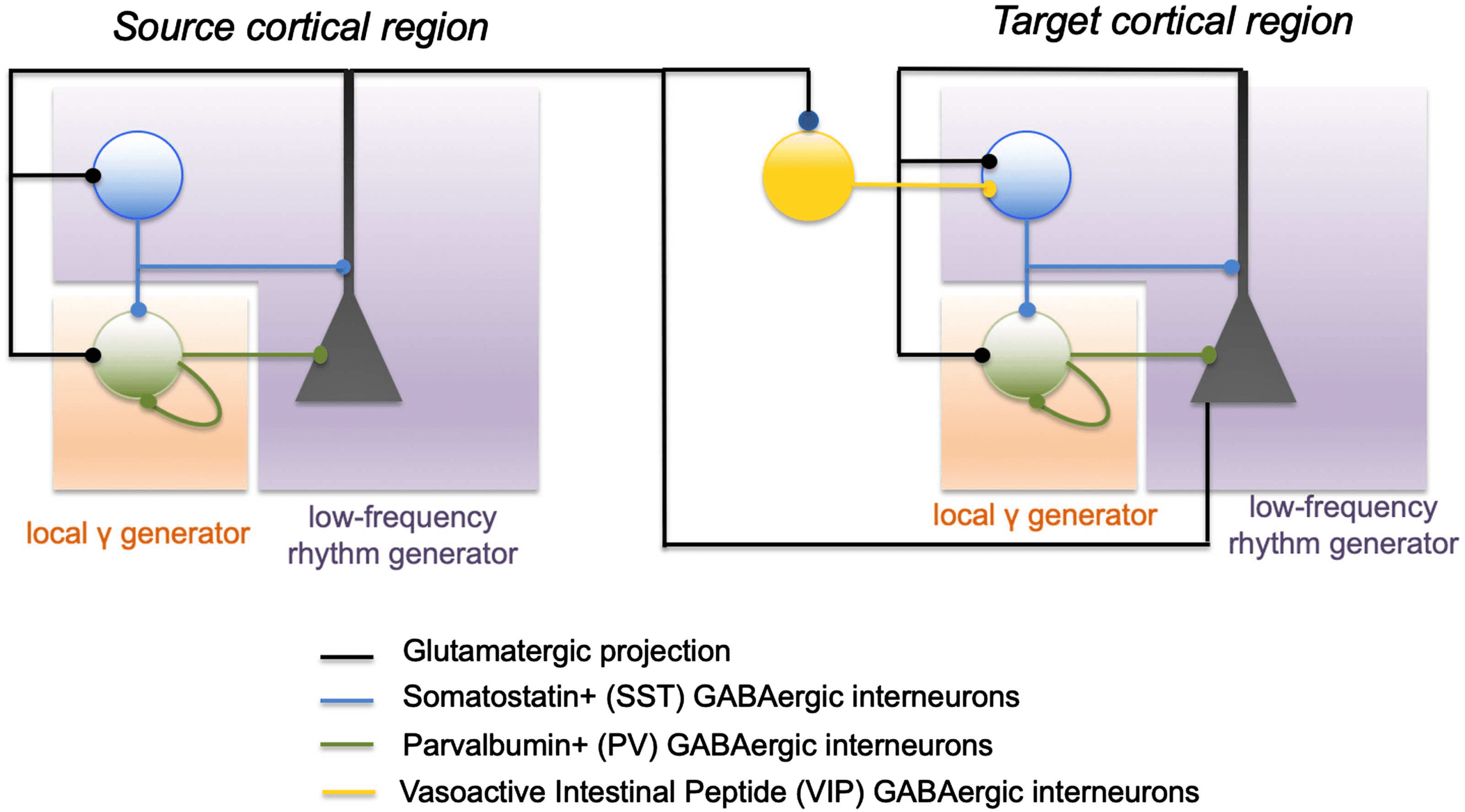
Figur 2. Basic (schematisch) circuits involved in the generation of local and distant oscillations.
In a local cortical network (cortical column), gap-junctional, mutual inhibition of soma-projecting
“fast” GABAergic interneurons are one of the basic mechanisms of generation for local gamma
Aktivität, along with the PING (pyramid-interneuron gamma). Umgekehrt, the feedback loop be-
tween dendrite-projecting “slow” GABAergic interneurons and pyramidal cells can generate low-
frequency activity. Wichtig, distant communication through the disynaptic pathway enables
transient generation of gamma oscillations in distant populations. Pyramidal cells in the source
Bevölkerung (left circuit) project on the pyramidal cells of the distant population (Rechts), but also on
VIP interneurons that project on dendrite-projecting SST neurons. Transient activation of VIP neu-
rons from the source population transiently inhibits SST neurons in the target population, enabling
the generation of gamma oscillations through the PV-PV and pyramidal-PV circuit. Once the input
on distant VIP neurons decreases, SST neurons resume their inhibitory input, which can terminate
gamma oscillations generation.
Netzwerkneurowissenschaften
322
Decoding the circuitry of consciousness
human brain samples (Schiavone, Neri, Trabace, & Turillazzi, 2017). Gesamt, in addition to the
TBI-induced damage of GABAergic interneurons involved in the generation of slow and fast
oscillations, the presence of abnormal excitatory synapses and axonal damage might also
participate to the alteration of neuronal connectivity (Harris et al., 2016).
Considering these cellular-scale mechanisms, one crucial question is to understand how
they are involved in the transient emergence and maintenance of functional networks such as
those supporting consciousness. It is established that the state of consciousness is critically
dependent on brain functional cortico-cortical connectivity (Marino, Bonanno, & Giorgio,
2016; Naro et al., 2018). As a reminder, functional connectivity refers to statistically signifi-
cant couplings between temporal courses of neuronal activity within different regions, wohingegen
anatomical connectivity denotes the physical connections between brain regions. Funktional
networks can therefore reflect indirect connections between brain regions, and are transient
depending on which tasks are performed, or which stimuli are perceived. Even in the absence
of any specific task or stimuli, it has been shown that resting-state networks (z.B., the default
mode network) are also transient (Kabbara, El Falou, Khalil, Wendling, & Hassan, 2017). Ex-
perimental evidence supports the idea that functional connectivity can shed light on the net-
works involved in various conscious states ( Jin & Chung, 2012). Zum Beispiel, during general
anesthesia-induced loss of consciousness, there is a breakdown in cortico-cortical functional
Konnektivität (Ferrarelli et al., 2010; Gomez et al., 2013; Hudetz, 2012), severely impairing the
capacity of cortical networks to integrate information and to make it available at a large scale,
as required for conscious perception in IIT or GWT. Ähnlich, in the transition from wakeful-
ness to slow-wave sleep, the firing rate in the cortex remains relatively unchanged during the
depolarizing phases of the slow sleep oscillation (Steriade, Timofeev, & Grenier, 2001), while
effective brain connectivity (effective connectivity is defined as the rapid, causal interaction
between brain regions; Rosanova et al., 2012) is dramatically altered (Esser, Hill, & Tononi,
2009; Tononi & Spurns, 2003). Upon falling into non-REM sleep, cortical activations also be-
come more local and stereotypical, impairing effective cortical connectivity (Massimini et al.,
2010), as shown using TMS-evoked EEG responses that remain very close to the stimulation
site, while these responses involve a network of distant brain regions undergoing complex dy-
namical patterns of successive activation during wakefulness (Casali et al., 2013; Casarotto
et al., 2016; Guillery & Sherman, 2002). These results also emphasize the crucial role of the
thalamo-cortical pathway in cortico-cortical functional connectivity. It is worth noting that the
essential role of the thalamo-cortical loop as well as so-called “reentrant interactions” were
previously considered as key in the DCH (Tononi & Edelman, 1998).
Consistently with these results obtained during sleep, this breakdown of cortico-cortical
connectivity has also been observed during general anesthesia and in DOC patients, Und
even explored through computational modeling (Casali et al., 2013; Esser et al., 2009). Im
GWT framework, this explains why consciousness is impaired in such states: large-scale
communication between distant brain areas is impaired because of thalamo-cortical modula-
tion, preventing ignition from occurring. In brain-damaged DOC patients, large-scale cortico-
cortical communication can be impaired through the partial destruction of long-range fibers,
physically impeding long-range brain synchrony. In terms of effects at the cellular scale, von-
struction of long-range fibers could prevent the synchronization of distant VIP interneurons,
which is critical to induce disynaptic disinhibition and associated gamma activity required for
CTC.
Pathological alterations of functional connectivity have been investigated using a variety
of modalities: (1) functional connectivity during “resting state” using fMRI or EEG, (2) pulsed
Netzwerkneurowissenschaften
323
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
N
e
N
_
A
_
0
0
1
1
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Decoding the circuitry of consciousness
stimulation using transcranial magnetic stimulation (TMS) during EEG recording, Und (3) andere
perturbation-based approaches investigating brain responses to sensory stimuli (Boly et al.,
2017). The advantage of functional connectivity is that it can be employed to improve the
evaluation and classification of DOCs (Holler et al., 2014; Naro et al., 2018; Rossi Sebastiano
et al., 2015; Sanders, Tononi, Laureys, & Sleigh, 2012). Zum Beispiel, in mild cognitive impair-
ment patients, it has been shown that impaired consciousness is associated with altered effec-
tive connectivity (Crone et al., 2015; Varotto et al., 2014). Failure of large-scale connectivity,
along with a hypersynchrony of local short-range delta and alpha activity were detected within
the DMN and were correlated with the level of awareness in patients with DOCs (Fingelkurts,
Fingelkurts, Bagnato, Boccagni, & Galardi, 2013; Kabbara et al., 2017; Maki-Marttunen, Diez,
Cortes, Chialvo, & Villarreal, 2013; Naro et al., 2018; Vanhaudenhuyse et al., 2010; Varotto
et al., 2014). Außerdem, the functional connectivity pattern of several brain regions, wie zum Beispiel
the posterior cingulate cortex and precuneus, may even predict UWS patients’ state of improve-
ment of consciousness with an accuracy superior to 80% (Wu et al., 2015). Let us mention
that fMRI has notably contributed to identifying changes in functional connectivity in vari-
ous resting-state networks (RSNs) in different conditions of consciousness (für eine Rezension, sehen
Heine et al., 2012. More specifically, in terms of resting-state fMRI, Demertzi et al. (2014)
used machine learning to automatically extract RSNs and determine their neuronal origin (als
opposed to artifactual). Results showed that, for most RSNs, the number of components of
neuronal origin decreased with the level of consciousness. In another study (Demertzi et al.,
2015), also based on the use of resting-state fMRI, the authors pointed at a decrease in func-
tional connectivity among the various RSNs (z.B., DMN, visuell, usw.), which correlated well
with the CRS-R score. Another recent fMRI study (Demertzi et al., 2019) has demonstrated that
the dynamics of fMRI functional networks could discriminate between different states of con-
sciousness (Kontrollen, MCS, UWS). More specifically, the probability of presence of different
modular states over time, along with their dwell time, was a reliable indicator of conscious-
ness. The complexity of the transition map between different modular states was also reduced
with the level of consciousness. The fact that both EEG and fMRI studies converge, albeit on
different timescales, on the reduction of complexity of dynamical responses, emphasize further
the fundamental relationship between the level of consciousness and the apparent complexity
of brain dynamics. The tight relationship between the BOLD signal, which is of low frequency
(<0.5 Hz, also called infraslow oscillations), and the low-frequency component of local field
potentials (LFPs) and electrocorticographic (ECOGs) signals (He, Snyder, Zempel, Smyth, &
Raichle, 2008), might provide a link between fMRI and EEG studies in the field of conscious-
ness research (for a review see He & Raichle, 2009). It has been indeed suggested that infraslow
oscillations in BOLD, which are linked to the low-frequency domain of LFP/ECOGs recordings
(for a thorough investigation of the correlation between the BOLD signal and electrophysio-
logical activity in different frequency bands see Scheeringa, Koopmans, van Mourik, Jensen,
& Norris, 2016) are a key contributor in the integration of information between distant corti-
cal regions. Interestingly, this proposal is in accordance with our proposal on the functional
role of low-frequency oscillations in the integration of information, while higher frequency
oscillations would be involved in binding.
Beside “passive” investigation of resting-state functional connectivity, the use of TMS-evoked
EEG responses enables the active “probing” of effective connectivity. For example, a drastic
breakdown of functional connectivity has been identified in UWS patients by using a spe-
cific TMS protocol, triggering, in these patients, a stereotyped local EEG response similar to
unconscious sleeping or anesthetized subjects (Rosanova et al., 2012). Restoring cortical large-
scale effective connectivity with transcranial brain stimulation, such as transcranial alternating
Network Neuroscience
324
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
n
e
n
_
a
_
0
0
1
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Decoding the circuitry of consciousness
tACS:
Transcranial current stimulation,
consisting in delivering electric
currents to the brain through
electrodes placed on the scalp, using
a constant (AC) current.
current stimulation (tACS), in DOCs could therefore be a useful approach to facilitate partial
recovery by enhancing oscillations and plasticity. One clinical result supporting this idea is
the recent demonstration that DLPFC (dorsolateral prefrontal cortex)-tACS was able to tran-
siently restore the connectivity breakdown in DOC individuals (Naro, Bramanti, Leo, Russo,
& Calabro, 2016).
Overall, we argue that a delicate balance of phase synchronization is required between
brain regions to enable optimal communication. Phase synchronization can occur through
the direct (or indirect through a common input) connections between two regions when some
form of activity in one changes activity in another. Such an increase in phase synchronization
underlies the so-called functional connectivity. Obviously, with low phase synchronization
(functional connectivity), no efficient transfer of information can take place between regions.
On the opposite end, excessive synchronization leads to a loss of complexity due to the activ-
ity becoming too similar in the involved regions. An extreme example is an epileptic complex
seizure, in which a large number of brain structures are strongly functionally connected and
have a similar seizure-like rhythm, and where there is a loss of consciousness (Blumenfeld
& Taylor, 2003). Therefore, we argue that there exists a “sweet spot” of synchronization at
specific frequencies that enables the proper maintenance of consciousness-related infor-
mation processing by a network of brain regions.
INFORMATION PROCESSING IN LARGE-SCALE FUNCTIONAL NETWORKS
THROUGH NESTED OSCILLATIONS
One fundamental microscopic-scale mechanism involved in information routing in the brain,
and contributing to form functional networks within the anatomical network, is GBI (Jensen
& Mazaheri, 2010). GBI involves inhibitory processes resulting in the selective activation of
subnetworks and inactivation of other subnetworks. By preventing brain-scale activation in re-
sponse to a stimulus, and restricting the number of brain regions engaged in performing tasks,
GBI also prevents states of low complexity (e.g., all brain regions displaying the exact same
activity) and therefore inefficient information processing. Thus, GBI processes suggest that the
role of inhibition is more complex than preventing excessive activation of brain networks,
contributing instead to shaping anatomical brain networks into functional networks (Avena-
Koenigsberger et al., 2017). Possible alterations of GBI were reported in studies showing that
EEG alpha power is lower in UWS than in MCS patients (Lehembre et al., 2012; Stefan et al.,
2018), hinting that the neurobiological mechanisms underlying alpha oscillations generation
and associated GBI are profoundly altered in unresponsive patients. Moreover, alpha activity
was highly synchronized and clustered in central and posterior cortical regions in UWS pa-
tients (Lehembre et al., 2012; Stefan et al., 2018), suggesting a possible failure of GBI in the
most severe DOCs. Indeed, during general anesthesia, propofol and ketamine modify frontal
alpha oscillations (Feshchenko, Veselis, & Reinsel, 2004; Purdon et al., 2013), suggesting that
reduced frontal-parietal connectivity in the alpha band might play a major role in the loss of
consciousness (Blain-Moraes, Lee, Ku, Noh, & Mashour, 2014). Coherent slow oscillations,
such as alpha (8–12 Hz) and delta (1–4 Hz) rhythms, appear across medial prefrontal cortex
and the thalamus during the loss of consciousness induced by propofol and is believed to
contribute to the functional disruption of these areas (Flores et al., 2017; Lewis et al., 2018).
More specifically, delta oscillations induced by propofol anesthesia in humans appear to im-
pair large-scale communication between brain regions, while preserving the activity of local
neural assemblies (Lewis et al., 2018). A recent study (Todorova & Zugaro, 2019) has reached
a similar conclusion regarding the functional role of delta oscillations during sleep, which
Network Neuroscience
325
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
n
e
n
_
a
_
0
0
1
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Decoding the circuitry of consciousness
isolate small networks within the cortex from the rest of the brain (thereby impairing integra-
tion), which seems to play a role in the reinforcement of memory during sleep.
Another key established processes by which distant brain regions engage together in an
activity pattern associated with the performance of a given task is CTC (Bonnefond, Kastner,
& Jensen, 2017; Deco & Kringelbach, 2016; Fries, 2005, 2009, 2015). CTC involves indeed
phase-coupled gamma activity between distant brain regions to enable information process-
ing. CTC has been suggested to be the substrate of “binding,” that is, the merging of different
features of a stimulus into a single, unified conscious perception (Singer, 2001). More pre-
cisely, the excitability fluctuation in a group of neurons provides a specific signature charac-
terized by a specific frequency band and pattern of discharge (Womelsdorf, Valiante, Sahin,
Miller, & Tiesinga, 2014), propagating through a large-scale network consisting of anatom-
ically interconnected brain areas and subsequently triggering activity in connected regions.
Information processing in the brain is strongly linked with phase-locked, coordinated-in-time
fluctuations of excitability (Fries, 2005) in networks of distributed neuronal populations. The
resulting oscillations generate a specific neuronal code, and coherence enables the associa-
tion of information and communication. Furthermore, CTC involves gamma activity, generated
mainly by GABAergic interneurons (PV-positive basket cells). Taken together, inhibitory pro-
cesses appear key for information routing and processing in brain-scale networks involved in
consciousness: GBI shapes brain networks spatially (which brain regions are involved, and
which ones are inhibited), while CTC controls them temporally (information flow). However,
this raises an intriguing question: If gamma rhythms are generated locally by interneuronal
GABAergic networks, how can distant brain regions, connected through glutamatergic long-
range fibers, communicate efficiently and achieve CTC? Our hypothesis could provide a pos-
sible explanation to this paradox by assuming that distant brain regions “lock” their activity
through low-frequency oscillations (which would be a prerequisite) and then enable binding
through high-frequency oscillation (gamma, through CTC).
Supporting this possibility, conscious perception is indeed characterized by an increase in
distributed gamma-band activity (Melloni et al., 2007; Wyart & Tallon-Baudry, 2009). Inter-
estingly, these fast oscillations are modulated by slow oscillations (Jensen, Gips, Bergmann, &
Bonnefond, 2014; Osipova, Hermes, & Jensen, 2008). It has recently been proposed that phase
synchronization of low-frequency oscillations, playing the role of a temporal reference frame
for information, carrying high-frequency activity, is a general mechanism of brain communica-
tion (Bonnefond et al., 2017). These nested oscillations might be a key mechanism, not only for
cortico-cortical communication and processing but also between subcortical structures. Emo-
tional memory is relevant to consider for our purpose, since emotional memory involves both
cortical and subcortical structures, and engages the synchronization of large networks through
theta-gamma oscillations (Bocchio, Nabavi, & Capogna, 2017). During in vivo experiments
performed in rodents, a perceived threat (a stimulus announcing a footshock) enhances theta
power and coherence in the amygdala, prefrontal cortex, and hippocampus (Lesting et al.,
2011; Likhtik, Stujenske, Topiwala, Harris, & Gordon, 2014), while fast gamma bursts are
phase-locked to theta oscillations (Stujenske, Likhtik, Topiwala, & Gordon, 2014). Overall,
these results support the idea that nested oscillations at theta and gamma frequencies are a
plausible substrate for information channel opening/routing (theta) and processing/transfer
(gamma) within the brain.
Attention is another key element for conscious processing, and it is involved in the syn-
chronization of distant brain regions (Niebur, Hsiao, & Johnson, 2002; Steinmetz et al., 2000).
The main underlying brain rhythms involved in attentional processes are alpha and gamma
Network Neuroscience
326
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
n
e
n
_
a
_
0
0
1
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Decoding the circuitry of consciousness
oscillations: brain regions synchronize gamma oscillations (Womelsdorf et al., 2014) and are
modulated by slow alpha oscillations. Slow oscillations enable inhibition of irrelevant networks
and influence local signal processing, widespread information exchange, and perception
(Sadaghiani & Kleinschmidt, 2016). Information flow is established by neuronal synchroniza-
tion at the lower frequency bands, namely in the theta (4–7 Hz), alpha (8–13 Hz), and beta
(14–25 Hz) bands (Bonnefond et al., 2017). One possible reason is that low-frequency ac-
tivity induces a transient change in excitability in target brain structures, which provides an
optimal window for binding neuronal signals from different regions through high-frequency
activity (i.e., gamma) (Canolty et al., 2006). This provides further support to the idea that low-
frequency neural oscillations are mainly involved in establishing transient long-range com-
munication through glutamatergic projections, whereas high-frequency neural oscillations are
involved in information processing/transfer. We propose that it is possible to relate the notion of
“integration” with this long-range glutamatergic coactivation, enabling brain-scale communi-
cation between brain regions; whereas “differentiation/segregation” would depend on locally
generated gamma activity (and in part on low-frequency activation level, which would result
in massively synchronized activity and reduced differentiation and complexity). An overview
of the aforementioned mechanisms is proposed in Figure 3.
Emerging evidence shows that the local versus global information processing balance can
be impaired in neurological disorders. Typically, a recent study investigating functional net-
works in Alzheimer’s disease patients identified a decrease in brain integration as quantified
by the participation coefficient (reflects communication between distant brain modules), while
segregation as quantified by the clustering coefficient (reflects local communication between
while brain regions) was increased (Kabbara et al., 2018). This is consistent with neurodegen-
erative processes, which likely impact the “locking” of specific brain regions or the inhibition
of irrelevant networks, thereby severely impairing large-scale integration of information. In the
context of DOC, recent clinical evidence (Chennu et al., 2017; Rizkallah et al., 2019) sup-
ports this view. In the study by Chennu et al., scalp-level networks were assessed from DOC
patients and pointed at decreased integration within the alpha band. More specifically, the
fronto-parietal network in the alpha band was discriminant between MCS and UWS patients.
In a recent study Rizkallah et al. (2019) quantified the level of local versus global information
processing in frequency-dependent functional networks (source level) in DOC patients and
controls. Integration in theta-band functional networks decreased with consciousness level,
and two anatomical regions were systematically involved between controls and any patient
group: a portion of the left orbitofrontal cortex and the left precuneus. One possibility is that
physical damage to long-range white matter fibers impairs large-scale integration and the local/
global information processing balance. One possible approach to study such anatomical dam-
age to white matter fibers is diffusion tensor imaging, as performed in DOC patients suffer-
ing severe brain injury (Fernandez-Espejo et al., 2011; Galanaud et al., 2012; Luyt et al.,
2012), which highlighted widespread disruptions of white matter. Lower fractional anisotropy
was indeed found in the subcortico-cortical and cortico-cortical fiber tracts of DOC patients as
compared with controls (Lant, Gonzalez-Lara, Owen, & Fernandez-Espejo, 2016; Weng et al.,
2017), suggesting that major consciousness deficits in DOC patients may be related to al-
tered white matter connections between the basal ganglia, thalamus, and frontal cortex. This
is also in line with the effect of lesion of myelinated fiber tracts, which can result in a failure
of communication between distant brain regions (Adams, Graham, & Jennett, 2000). There-
fore, it seems reasonable that white matter lesions can alter, modify, or prevent both CTC and
GBI between large-scale networks. Furthermore, we speculate that should the specific phase-
locking of gamma oscillations onto theta oscillations be perturbed, then clinical manifestations
Network Neuroscience
327
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
n
e
n
_
a
_
0
0
1
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Decoding the circuitry of consciousness
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
n
e
n
_
a
_
0
0
1
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
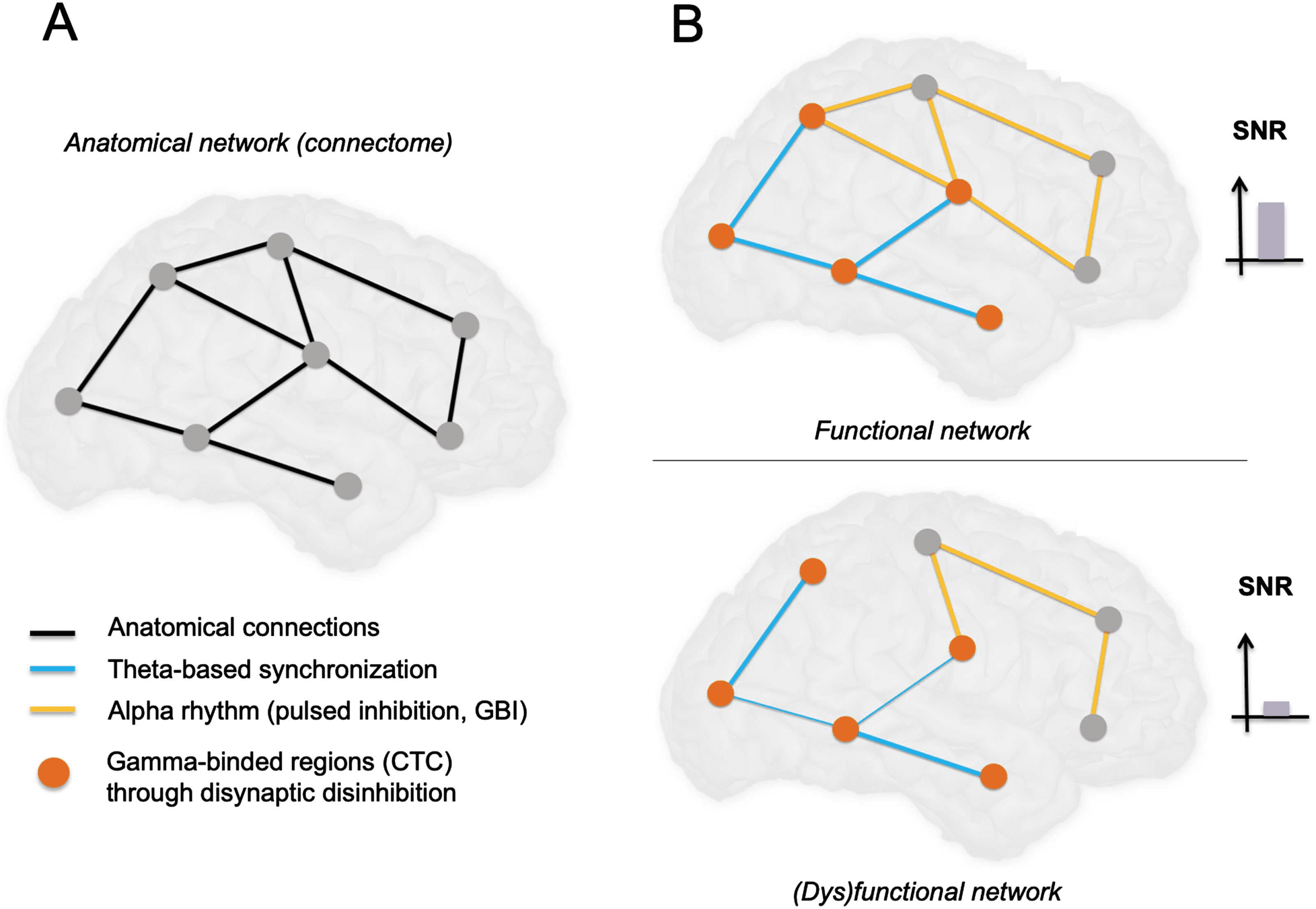
Figure 3. Proposed synthesis on how transient selective binding among cortical networks takes
place through cellular-scale mechanisms. (A) Schematic diagram of an anatomical network with
main projections between regions at the brain scale. (B) In top panel, selective binding (communi-
cation through coherence; Fries, 2015) in subset of cortical regions occurs through the generation
of gamma oscillations (mostly through microcircuits involving basket cells), while the distant disin-
hibition of specific brain regions occurs through disynaptic disinhibition (activation of distant VIP
neurons inhibits SST neurons, which in turn decrease their inhibitory projection on pyramidal neu-
rons). This contributes to shape the anatomical network into a functional network. The alpha rhythm
acts as a pulsed inhibition to inhibit “irrelevant” networks, increasing further the signal-to-noise ra-
tio (SNR). In bottom panel, decreased integration (e.g., following brain damage) leads to impaired
synchronization of distant brain regions (reflected by decreased low-frequency rhythm on the illus-
trative oscillation), and thereby a decrease of binding, which combined leads to a decrease in GBI
(Jensen & Mazaheri, 2010) efficiency and strongly decreases the overall SNR, leading to dysfunction
of the network in terms of integration and binding required for consciousness. Let us note that in
addition to decreased amplitude of the low-frequency rhythm, the phase relationship of the nested
gamma oscillation could be perturbed (i.e., more random) as compared with the physiological case.
Such potential relationship of nested theta/gamma oscillations remains to be explored in DOCs.
associated with DOCs might appear (loss of integration and decrease in the consciousness
level).
Let us also mention that, although not directly related, other pathologies can inform the
pathophysiology of DOCs since they share one crucial component: alterations in functional
networks. When those alterations impact key brain structures/networks, consciousness has the
potential to be altered. Therefore, understanding how deregulations of functional networks in
neurological disorders that do not involve any deterioration of the consciousness level (e.g.,
mild cognitive impairment patients) can still be informative in terms of fundamental mecha-
nisms underlying DOCs. In the same vein, since functional connectivity is dependent on the
level of phase synchronization between regions, it is of prime importance to highlight that in-
creasing the level of phase synchronization between the structures involved in a precise task
Network Neuroscience
328
Decoding the circuitry of consciousness
(e.g., the fronto-temporal network underlying working memory) also improves behavioral per-
formance of this task. Therefore, the tACS literature can provide novel directions on how to
regulate the functional networks most critically in conscious processes, for example, by in-
creasing the phase synchronization of key structures in specific frequency bands. This point is
discussed in the next section.
POSSIBLE CLINICAL IMPLICATIONS
In this review, we have attempted to reconcile the neuroanatomical and neurophysiological
knowledge at the level of micro- and macroscopic networks, regarding the processes that
underlie the emergence and maintenance of consciousness and its alterations in DOC pa-
tients. The multiplexing of neuronal rhythms through nested oscillations appears as a plau-
sible mechanism of coactivation in a network of specific distant brain regions (integration),
which is a prerequisite for a conscious perception. Furthermore, a key mechanism seems to
be the subtle balance between low-frequency activity (associated with “global” processing)
and high-frequency activity (associated with a more “local” processing), which could enable
the neuronal dynamics underlying optimal information routing and processing. Excessive low-
frequency activity (e.g., delta activity) results in massively synchronized activity, resulting in a
loss of complexity in terms of information processing, paralleled in such cases with a loss of
consciousness (e.g., sleep, seizures). Regarding the delta band, this rhythm does not appear
linked with the emergence or maintenance of conscious processes. This is supported by the
fact that delta activity is the most predominant in UWS (Sitt et al., 2014) and also during deep
sleep, where in both cases cortico-cortical communication is drastically reduced. UWS and
deep sleep are indeed examples of mental states where thalamo-cortical activity and synchro-
nization is high (“vertical” connectivity), while cortico-cortical (“horizontal”) synchronization
decreases. Since the delta rhythm has a thalamo-cortical origin, and since the thalamus is
so extensively connected to the entirety of the cortex, delta activity is associated with a high
level of synchronization throughout the cortex, thereby reducing integration and complexity,
and preventing conscious access. Furthermore, delta activity appears to modulate mainly the
“wakefulness” (thalamo-cortical) dimension of consciousness, whereas faster rhythms seem to
modulate the “awareness” (cortico-cortical) dimension of consciousness.
Similarly, a lack of fronto-parietal functional coupling (attentional network) has been
observed in a recent study, as quantified using high-resolution EEG in DOC patients (Chennu
et al., 2017), suggesting that a sufficient level of fronto-parietal coupling is required to achieve
sufficient neuronal integration and ignition for conscious perception. More generally, brain
dynamics in DOC patients are typically characterized by a loss of integration at a large-scale
(Chennu et al., 2017; Rizkallah et al., 2019), preventing efficient large-scale coordination of
distant brain regions to achieve conscious perception. This suggests that the low-frequency
rhythm required for long-range cortical communication is decreased, preventing binding in
the gamma range and therefore further processing information and ignition for consciousness
access. That being said, what are the possible implications of this slow/fast activity balance as
a candidate mechanism for the complex processing associated with consciousness?
An interesting perspective, which would also be a form of validation for this mechanism, is
the use of neuromodulation techniques in DOC patients to increase their level of conscious-
ness. The objective of such neuromodulation techniques could be to “rebalance” local versus
global processing, for example, through the use of transcranial direct or alternating stimulation
(tDCS/tACS), applied to both a frontal and a parietal site simultaneously in order to increase
low-frequency synchronization in the theta range. It is the current view that tACS can modulate
329
Multiplexing:
The combination of several
oscillations observed in the same
electrophysiological signal at the
level of the same brain region.
tDCS:
Transcranial current stimulation,
consisting in delivering electric
currents to the brain through
electrodes placed on the scalp,
using a constant (DC) current.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
1
5
1
8
6
6
5
9
8
n
e
n
_
a
_
0
0
1
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Decoding the circuitry of consciousness
endogenous brain rhythms by using relatively low-level electric fields (<1 V>