RESEARCH ARTICLE
Role of Family Risk and of Pre-Reading Auditory
and Neurostructural Measures in Predicting
Reading Outcome
Keine offenen Zugänge
Tagebuch
Lauren Blockmans1
Jan Wouters1
, Narly Golestani2,3,4
, Pol Ghesquière6
, Josué Luiz Dalboni da Rocha3,5
,
, and Maaike Vandermosten1
1Research Group ExpORL, Department of Neuroscience, KU Leuven, Leuven, Belgien
2Brain and Language Lab, Cognitive Science Hub, University of Vienna, Vienna, Österreich
3Abteilung für Psychologie, Faculty of Psychology and Educational Sciences, University of Geneva, Genf, Schweiz
4Department of Behavioral and Cognitive Biology, Faculty of Life Sciences, University of Vienna, Vienna, Österreich
5St. Jude Children’s Research Hospital, Memphis, TN, USA
6Parenting and Special Education Research Unit, Faculty of Psychology and Educational Sciences, KU Leuven, Leuven, Belgien
Schlüsselwörter: pre-reading, T1-weighted MRI, family risk, speech-in-noise perception, Anstiegszeit
discrimination, auditory cortex
ABSTRAKT
Some children who develop dyslexia show pre-reading auditory and speech processing
difficulties. Außerdem, left auditory cortex structure might be related to family risk for
dyslexia rather than to reading outcome. Jedoch, it remains unclear to what extent auditory
and speech processing and auditory cortex structure mediate the relationship between family
risk and reading. In the current longitudinal study, we investigated the role of family risk
(measured using parental reading questionnaires) and of pre-reading auditory measures in
predicting third grade word reading. We measured auditory and speech processing in 162 pre-
readers varying in family risk. In 129 of them, we also acquired structural magnetic resonance
Bildgebung (MRT). We quantified surface area and duplication patterns of the bilateral transverse
temporal gyri (TTG(S)), and surface area of the bilateral planum temporale (PT). We found
effects of pre-reading auditory and speech processing, surface area of the left first TTG and of
bilateral PT and of left TTG duplication pattern on later reading. Higher pre-reading values
on these measures were predictive of better word reading. Although we also found some
evidence for an effect of family risk on auditory and speech processing, these latter measures
did not mediate the strong relationship between family risk and later reading. Our study shows
the importance of pre-reading auditory and speech processing and of auditory cortex anatomy
for later reading. A better understanding of such interrelations during reading development will
facilitate early diagnosis and intervention, which can be especially important given the
continuity of family risk in the general population.
EINFÜHRUNG
Reading achievement can be predicted by phonological processing skills (Quinn et al., 2015)
and features of the neural reading network (Chyl et al., 2021) measured as early as in kinder-
garten (d.h., pre-reading). These cognitive and neural processes associated with reading are
influenced by genetic and environmental factors, as described in the theoretical models of
van Bergen et al. (2014) and van Atteveldt et al. (2021). Genauer, genetic variations
Zitat: Blockmans, L., Golestani, N.,
Dalboni da Rocha, J. L., Wouters, J.,
Ghesquière, P., & Vandermosten, M.
(2023). Role of family risk and of pre-
reading auditory and neurostructural
measures in predicting reading
outcome. Neurobiology of Language,
4(3), 474–500. https://doi.org/10.1162
/nol_a_00111
DOI:
https://doi.org/10.1162/nol_a_00111
zusätzliche Informationen:
https://doi.org/10.1162/nol_a_00111
Erhalten: 16 Januar 2023
Akzeptiert: 22 Mai 2023
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Lauren Blockmans
lauren.blockmans@kuleuven.be
Handling-Editor:
Kate Watkins
Urheberrechte ©: © 2023
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Phonological processing:
Refers to cognitive processes of
recognizing, manipulating and
accessing speech sounds, welche sind
crucial in developing reading
fluency.
are partly responsible for characteristics at the neural level, which in turn influence the cog-
nitive processes that underlie reading development (Galaburda et al., 2006; Giraud & Ramus,
2013). In line with these models, previous studies have found that phonological processing
skills (van Bergen et al., 2015) and white matter connectivity in the left temporal lobe
(Vandermosten et al., 2017) partially mediate the relationship between parental reading abil-
ities and the reading outcomes of their children. These findings support the idea that factors at
multiple levels can interact with and mediate the intergenerational transfer of reading skills
between parents and children. Zusätzlich, previous longitudinal studies related pre-reading audi-
tory and speech processing skills to later reading, while a few brain imaging studies have indirectly
suggested that left auditory cortex anatomy is related to family risk for reading difficulties, often defined
as having at least one first-degree relative with a formal dyslexia diagnosis, rather than to reading
outcome (Beelen et al., 2019; Łuniewska et al., 2019; Vanderauwera et al., 2018; van der Leij
et al., 2013; Vandermosten, Correia, et al., 2020). Taken together, these auditory (Kortex) factors
may have a similar mediating role in early reading development, as previously observed for
phonological processing and white matter connectivity. In this study, we examined the extent to
which children’s pre-reading auditory skill and auditory cortex structure mediated the effect of
their parent’s reading abilities (indexed indirectly using a continuous measure of family risk) An
third grade word reading outcome.
Pre-reading Auditory and Speech Processing
It is well known that phonological processing plays a crucial role during typical (Clayton et al.,
2020; Quinn et al., 2015) and atypical (Ozernov-Palchik & Gaab, 2016; Snowling & Melby-
Lervåg, 2016) reading development, although recent accounts have been questioning its
causal role (sehen, z.B., Blockmans et al., 2023; Landerl et al., 2019; Peterson et al., 2018). Inter-
estingly, there is also some evidence linking lower level, auditory processing of speech and
non-speech signals to reading development. Several lower level features of auditory signals
such as frequency and duration have been shown to be important in typical (Goswami,
2022) and atypical (Hämäläinen et al., 2013; Serrallach et al., 2016) reading development.
Genauer, Goswami (2011, 2022) has proposed that especially the ability to discrim-
inate temporal information embedded in the amplitude envelope, reflected, Zum Beispiel, In
stimuli differing in the time taken to reach the maximum amplitude (also known as rise time),
can be related to the development of phonological processes important for reading. In line
with this theory, rise time discrimination (RTD) has been indirectly linked to reading skills.
Zum Beispiel, RTD was found to predict phonological awareness and letter knowledge longi-
tudinally in the year before formal reading instruction, das ist, at the pre-reading stage
(Corriveau et al., 2010), and at the start of the first year of reading instruction (Vanvooren
et al., 2017). Also at later stages, during primary school, RTD is associated with phonological
awareness, both concurrently (Poelmans et al., 2011; Richardson et al., 2004) and longitudi-
schließlich (Goswami et al., 2021). Interessant, some studies reported a direct link between pre-
reading rise time processing and reading outcome after at least one year of reading instruction
(Law et al., 2017; Plakas et al., 2013), and even later, between rise time processing at age eight
and reading outcome at age nine (Kuppen et al., 2014). Darüber hinaus, several studies identified
RTD deficits in children with severe reading difficulties, otherwise known as dyslexia (Law
et al., 2017; Poelmans et al., 2011; Richardson et al., 2004; Serrallach et al., 2016), und in
adults with dyslexia (Law et al., 2014; Leong et al., 2011; Van Hirtum et al., 2019), with aver-
age effect sizes of 0.8 (Hämäläinen et al., 2013).
Efficient auditory processing, in particular RTD, contributes to the temporal segmentation of
more complex auditory signals such as speech (Goswami, 2011). The relationship between
Neurobiology of Language
475
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Duplication pattern:
A quantitative measure reflecting the
shape of the transverse temporal gyri.
speech processing and reading is typically assessed using speech presented in a challenging
Umfeld, Zum Beispiel, with background noise. Better pre-reading speech-in-noise percep-
tion has been linked to better reading outcome during primary school years (Boets et al., 2008,
2011; Vandewalle et al., 2012; Vanvooren et al., 2017; White-Schwoch et al., 2015). Weiter-
mehr, speech-in-noise perception is more often anomalous in individuals with dyslexia com-
pared to typical reading controls (Boets et al., 2011; Boets, Wouters, et al., 2007; Calcus et al.,
2018; Rosen, 2003; Ziegler et al., 2009). Wichtig, deficits in auditory and speech process-
ing are only present in a subset of persons with dyslexia, with studies reporting incidence rates
ranging from 30% (Boets et al., 2006; Boets, Ghesquière, et al., 2007; Calcus et al., 2018;
Rosen, 2003) Zu 60% (Law et al., 2014; Ramus et al., 2003). Zusammenfassend, several lines of
research suggest that auditory and speech processing are important for successful reading
Entwicklung.
Pre-reading Auditory Cortex Structure
Studies have tried to identify brain structural predictors or correlates of reading outcome,
including traditional structural measures such as volume, surface area, and thickness and mea-
sures of shape such as gyrification indices or gyral duplication patterns. The earliest work in
this field was related to the anatomy of the PT, a secondary auditory cortex region known to be
functionally involved in the spectrotemporal analysis of speech sounds (Griffiths & Warren,
2002). The PT surface area is usually left-lateralized in the general population (Tzourio-
Mazoyer & Mazoyer, 2017), with the left PT surface area being larger than the right one.
Reversed or absent asymmetry has been related to atypical reading outcome (Altarelli et al.,
2014; Sanchez Bloom et al., 2013), but others could not replicate this finding (Eckert &
Leonard, 2000; Vanderauwera et al., 2018). Methodological differences, such as the applied
delineation criteria and sample size restrictions, might lie at the base of these discrepancies
across studies (Ramus et al., 2018).
In addition to the PT, early stages of auditory and speech processing are subserved by other
auditory regions, including Heschl’s gyrus (HG), other secondary regions such as the planum
polare, and associative cortices. The primary auditory cortex is most often located within the
Erste, most anterior TTG (Von Economo & Horn, 1930), which corresponds to HG in cases
where there is a single gyrus. The first TTG can also be partially separated by an intermediate
Sulkus, in the case of common stem duplications (CSDs; Marie et al., 2015). Different defini-
tions exist for determining the posterior border of HG, which is also the anterior border of the
PT, in particular in the presence of CSDs. Older definitions assign the posterior branch of the
CSD to the PT in cases where the length of the intermediate sulcus is more than half the length
of HG (Golestani et al., 2007; Leonard et al., 2001; Penhune et al., 1996). Other researchers
have defined the posterior branch of CSDs as belonging to HG, regardless of the length of the
intermediate sulcus (Schneider et al., 2005; Seither-Preisler et al., 2014; Wengenroth et al.,
2014; Zoellner et al., 2019), which is the definition used by the present work. Additional, völlig
duplicated TTG, when present, also belong to the PT. The first TTG and any additional TTG(S))
are located on the superior surface of the superior temporal gyrus, within the Sylvian fissure.
Structural characteristics and measures of shape of the TTG have been related to language
and musical skills, both of which rely on adequate auditory processing (Turker & Reiterer,
2021). Positive associations have been reported between gray matter volume of HG/TTG
and musical abilities (Palomar-García et al., 2020; Schneider et al., 2002; Seither-Preisler
et al., 2014), phonetic expertise (Golestani et al., 2011), language aptitude (Turker et al.,
2019), non-native speech sound learning skill (Golestani et al., 2007), linguistic pitch learning
Neurobiology of Language
476
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
skill (Wong et al., 2008), and bilingualism (Ressel et al., 2012). These associations have been
reported in the right HG/TTG (Palomar-García et al., 2020; Seither-Preisler et al., 2014; Turker
et al., 2019), the left HG/TTG (Golestani et al., 2007, 2011; Palomar-García et al., 2020; Wong
et al., 2008), and the bilateral HG/TTG (Ressel et al., 2012; Schneider et al., 2002). Likewise,
the presence of multiple TTGs in both the left and right hemispheres has been related to adults’
musicality (Benner et al., 2017; Turker et al., 2017), phonetic expertise (Golestani et al., 2011),
and language aptitude (Turker et al., 2021). Paradoxerweise, several studies have shown that
people with dyslexia are more likely to have complete posterior duplications (d.h., fully sepa-
rated additional TTG(S)). Speziell, additional TTG(S) occur more often in the right hemi-
sphere in children (Altarelli et al., 2014; Serrallach et al., 2016) and in the left hemisphere in adults
with dyslexia (Leonard et al., 2001) compared to people with typical reading skills. The aspects
of TTG duplication that are beneficial versus detrimental in language/musical skill versus in dys-
lexia, jeweils, remain to be understood. Außerdem, it is unlikely that the mixed findings
on left versus right hemisphere involvement reflect developmental changes, in light of the known
relative stability of auditory cortex anatomy. While gray matter volume across the whole brain
develops rapidly in the first two years of life, it is much more stable in later childhood and ado-
lescence (Gennatas et al., 2017; Gilmore et al., 2018). Gray matter volume of the auditory cortex
specifically has been shown to be very stable in nine-year-olds tested longitudinally over
approximately one year (Seither-Preisler et al., 2014). The mixed findings are more likely due
to the behavioral measures under investigation and/or to sample characteristics. Zum Beispiel,
sex differences could also explain these contradictory results, as the studies finding right hemi-
spheric duplications in children oversampled males (Serrallach et al., 2016) or found the effect
only in males (Altarelli et al., 2014). Vor allem, none of these studies accounted for family history
of dyslexia, a factor which we elaborate on below (see Influences of Genetics, Heritability, Und
Family Risk, below).
Other features of auditory cortex shape also appear to be different in dyslexia, such as the
gyrification index, which assesses the local ratio of curvature perpendicular to the cortical
surface as opposed to the shape of HG parallel to the cortical surface (Schaer et al.,
2008). A recent study looking at HG specifically reported a higher gyrification index of the
left HG in children with dyslexia compared to peers with typical reading skills (Kuhl et al.,
2020). This was found to be the case in the same children assessed once before (Alter 5) Und
once during (Alter 8) reading instruction, although the effect of time itself was not investigated
in this study. Clark et al. (2014) found thinner cortex of the left HG in pre-reading children
(Alter 6) who go on to develop dyslexia. Considering the small sample sizes in the studies of
Kuhl et al. (2020; n = 32 of which 16 developed dyslexia) and Clark et al. (2014; n = 17 von
welche 7 developed dyslexia), it remains unknown if these pre-reading differences in HG
anatomy would replicate in larger samples. In the study by Clark et al. (2014), the thickness
difference in the left HG was still present after a few years of reading instruction (Alter 12), Aber
other studies relating dyslexia to cortical thickness of HG after reading acquisition reported
contradictory results (Ramus et al., 2018). Ma et al. (2015) reported thicker right HG in
school-aged children with dyslexia, whereas neither Altarelli et al. (2014) nor Skeide et al.
(2018) found any differences in cortical thickness of HG between individuals with and
without dyslexia. Diverging findings might be related to experience-dependent plasticity of
cortical thickness, since this structural feature has been shown to change with experience
and with language-related training (Hervais-Adelman et al., 2017; Lövdén et al., 2013;
Mårtensson et al., 2012).
Other studies have also looked at gray matter volume of the auditory cortex in children after
the pre-reading stage. Children with dyslexia exhibited lower gray matter volume of the left
Neurobiology of Language
477
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Heritability:
The proportion of individual
differences in a characteristic that
can be attributed to genetic factors.
Epigenetics:
The study of changes in gene
expression by environmental factors,
without changes to the underlying
DNA sequence.
HG and PT in comparison to peers without dyslexia (Xia et al., 2016). Jedoch, while
Serrallach et al. (2016) also found lower gray matter volume in bilateral HG, they found higher
gray matter volume in bilateral PT. Last, Sutherland et al. (2012) found a positive relationship
between gray matter density in bilateral HG and auditory processing of non-speech stimuli in a
sample of 10-year-olds. The latter was correlated positively with reading, but the investigation
of a direct link between HG and reading was lacking. Zusamenfassend, there is ample evidence that not
only traditional structural features of the TTG and of the PT such as volume, surface area, Und
Dicke, but also shape, relate to language and music skills. Jedoch, evidence concerning
the predictive role of pre-reading TTG and PT anatomy in the context of reading skills is limited
and contradictory (Ramus et al., 2018), warranting further research.
Influences of Genetics, Heritability, and Family Risk
It is known that both surface area and cortical thickness are influenced by genetics and
heritability, with large regional cortical variation in the heritability of these different brain
structural features (Grasby et al., 2020; Strike et al., 2019). A recent twin study in infants found
strong, significant effects of heritability on surface area as opposed to weak, nonsignificant
effects on cortical thickness (Jha et al., 2018). Consistent with this, a recent large-scale
meta-analysis looking both at molecular genetics and at twin heritability studies has shown
a stronger influence of molecular genetics on surface area than on cortical thickness. Specif-
isch, this study showed that surface area is more likely to be influenced by early develop-
mental factors and that cortical thickness is more likely to be influenced by learning and
experience during adulthood (Grasby et al., 2020). In this study, genetics and heritability
were the focus, but this does not exclude the role of other early factors on brain development
(Hoeft & Hancock, 2017), ones including prenatal hormones (Miranda & Sousa, 2018) Und
epigenetic effects (Schmied, 2011). Grasby et al. (2020) showed regional variation in the effects
of genetics in different cortical regions, in line with other studies reporting high heritability for
HG specifically compared to other cortical regions (Hulshoff Pol et al., 2006; Winkler et al.,
2010). Daher, in agreement with what is proposed by theories of reading development (Transporter
Atteveldt et al., 2021; van Bergen et al., 2014), genetic and environmental factors can have
a distinct impact on different neurostructural properties, possibly with a greater influence of
genetics on surface area than on thickness.
Zusätzlich, behavioral studies have identified pre-reading auditory and speech perception
difficulties in children with, compared to children without, family risk for dyslexia (Boets,
Ghesquière, et al., 2007; Gerrits & de Bree, 2009; Kalashnikova et al., 2018; Noordenbos
et al., 2012; Richardson et al., 2003), although it should be noted that some could not detect
such group differences (Boets et al., 2006; Vanvooren et al., 2017). Emerging evidence from
several neuroimaging modalities further indirectly suggests that family risk for dyslexia (eher
than reading skill itself ) could explain some of the brain differences in the temporal cortex,
including the auditory cortex, during reading development (van der Leij et al., 2013). Erste,
studies using event-related potentials showed poorer auditory processing in children with a
family risk for dyslexia compared to peers without such risk, and this difference was not
dependent on later reading outcome itself. Genauer, this was found for the auditory
processing of frequency differences between tone stimuli (Hakvoort et al., 2015; Leppänen
et al., 2010). Family risk driven deficits were also shown in the processing of temporal pattern
changes (van Zuijen et al., 2012) and rise time differences (Plakas et al., 2013) between tone
stimuli in young children, but note that the latter could not be replicated in older children
(Hakvoort et al., 2015). Zweite, functional MRI studies suggested that brain activation can
depend on the family risk status. Zum Beispiel, activation elicited by two phonemes in the
Neurobiology of Language
478
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
bilateral superior temporal gyri was distinct in children without a family risk but not in children
with such risk, and these activation differences were unrelated to the actual reading outcome
(Vandermosten, Correia, et al., 2020). Children with family risk further exhibited hypoactiva-
tion in the bilateral temporal gyri during a phonological task, irrespective of their reading out-
kommen (Łuniewska et al., 2019). Dritte, concerning structural MRI measures of these regions,
Beelen et al. (2019) found a smaller surface area in bilateral temporal gyri in pre-reading chil-
dren with a family risk, again unrelated to later reading outcome. Zusätzlich, the typical PT
surface area asymmetry was reported to be reversed in children with a family risk, with right-
ward rather than leftward structural lateralization (Vanderauwera et al., 2018).
Zusammenfassend, it seems likely that some auditory and speech processing deficits and neural
properties of related brain regions reflect family risk status but do not relate to actual reading
outcome. Jedoch, these previous findings used group comparisons, whereas family risk is
known to be continuous rather than categorical (Erbeli et al., 2022; Snowling et al., 2003;
van Bergen et al., 2014), and whereas it is known that arbitrary diagnostic dichotomies do
not accurately reflect reading skills (Astle et al., 2022). A few studies assessed reading abilities
of the parents as a continuous measure for family risk. They observed that the relationship
between family risk and reading outcome was mediated by phonological processing skills (Transporter
Bergen et al., 2015) and by white matter connectivity in the temporal cortex (Vandermosten
et al., 2017). Vor allem, these factors only partially mediated the family risk–reading relationship,
and as such do not exclude the well-established presence of a robust direct link (Snowling &
Melby-Lervåg, 2016). Currently, it remains unknown whether a similar mediating role pertains
to auditory and speech processing and to auditory cortex anatomy. Daher, a direct investigation
of the possible mediating role of auditory (Kortex) factors on the relationship between family
risk and reading is lacking.
The Current Study
In the current study, we investigated whether pre-reading behavioral measures of auditory and
speech processing and auditory cortex anatomy mediated the relationship between family risk
for dyslexia and third grade word reading outcome. We recruited a relatively large sample of
Teilnehmer (n = 162), that varied in terms of their family risk. Pre-reading auditory processing
was assessed with non-speech (RTD) and speech (speech-in-noise perception) tasks. For the
pre-reading auditory cortex measures, we examined surface area of bilateral first TTG, addi-
tional TTG(S) (when present) and PT and surface area asymmetry of PT because of previous
work showing a greater influence of heritability and genetics on surface area rather than on
Dicke. Due to previous findings of TTG shape differences in dyslexia, we also examined
bilateral TTG duplication patterns by assessing the total number of gyri along the Sylvian
fissure. Surface area of the first and of additional TTG(S) were extracted using an automatic
auditory cortex segmentation toolbox (Toolbox for the Automated Segmentation of Heschl’s
Gyrus, or TASH; Dalboni da Rocha et al., 2020), which was adapted to suit the pediatric data
at hand. Since family risk is known to be continuous, we assessed it using a continuous
messen, namely parental reading questionnaires. We expected an effect of family risk
(Snowling & Melby-Lervåg, 2016) and of pre-reading auditory and speech processing and
auditory cortex structure (Boets et al., 2011; Clark et al., 2014; Kuhl et al., 2020; Law et al.,
2017) on third grade reading outcome. Zusätzlich, we hypothesized that auditory and
speech processing and surface area of bilateral first TTG, additional TTG(S) and PT would
partially mediate the effect of family risk on reading outcome, since these measures are
often influenced by familial factors (Beelen et al., 2019; Jha et al., 2018; Plakas et al.,
2013; Vanderauwera et al., 2018).
Neurobiology of Language
479
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
MATERIALS AND METHODS
Teilnehmer
This study is part of a longitudinal research project of the Dyslexia Collaboration Leuven
(DYSCO) in which children are followed up throughout their early reading development.
The study was approved by the University Hospital Leuven Ethical Committee (S55139 and
S54415). Written informed consent by the parents and oral assent by the children was
erhalten. Participant recruitment and the auditory and speech processing data collection took
place at the start of the third and final year of kindergarten (age 4–5). Zusätzlich, strukturell
brain images were acquired at the end of kindergarten, using MRI. Reading skills were not
formally assessed in kindergarten, but since this study was conducted in Flanders (Belgien),
where formal reading instruction is not yet initiated in kindergarten (https://onderwijsdoelen
.be/), the first stage of our study is considered the pre-reading stage. Reading data were col-
lected at the start of second (age 6–7) and third grade (age 7–8). Participants received a small
present after each test session, and parents were sent a report about their child’s individual
behavioral results.
The study sample consisted of two cohorts with a total of 163 Teilnehmer. Children in both
cohorts were followed up at the same time points during reading development using identical
Maßnahmen, allowing compilation of the two cohorts into one study sample. The first cohort (n =
87) was recruited in 2011. Half of these children had at least one first-degree relative with a
formal dyslexia diagnosis, reflecting an increased familial risk for dyslexia. For the second
cohort, recruited in 2018, we selected 76 children without a familial risk but with an increased
cognitive risk for dyslexia, out of a large-scale screening effort (n = 1,225; Verwimp et al.,
2020). Cognitive risk corresponded to a score below the 30th percentile compared to the
entire screening data set, on at least two out of three pre-reading tasks (phonological aware-
ness, rapid automatized naming, and letter knowledge). Cognitive risk was also assessed ret-
rospectively in the 2011 cohort. These recruitment efforts resulted in the total sample (n =
163), consisting of 27 children with only a family risk, 64 children with only a cognitive risk,
36 children with a combined family and cognitive risk, Und 36 children without any risk. Für
Die 2011 cohort, all measures were collected prior to the COVID-19 pandemic. For the 2018
cohort, all the pre-reading measures and brain imaging were also assessed before the COVID-
19 pandemic and thus before nationwide school closures. Jedoch, children in this cohort
experienced school closures during first grade (March–May 2020). daher, we chose to
include third grade reading data (collected in September 2021), since by then, these partici-
pants had received an entire school year of uninterrupted in-person classroom reading
instruction.
Across both cohorts, all children were monolingual Dutch speakers, without previous his-
tory of hearing loss, vision deficits, brain damage, or ADHD. The data of one participant were
excluded from all analyses due to an incidental brain finding, resulting in a study sample of
162 participants for all analyses including only behavioral data. For analyses including brain
imaging data, 56 additional subjects were excluded due to various reasons (24 did not agree to
participate in the MRI session, 7 did not complete the MRI session, 20 showed excessive head
Bewegung, 3 had unexpected brain segmentation errors, Und 2 dropped out during the course of
the study), resulting in a sample of 106 Kinder. We report general participant characteristics,
nämlich, Sex, handedness, Alter, intelligence, and parental education, in Table 1. Handedness
was evaluated using the parents’ report of the Edinburgh Handedness Inventory (EHI; Oldfield,
1971). Three participants were ambidextrous according to the EHI manual, which classifies
scores below −40 as left-handed and scores above +40 as right-handed. Jedoch, the scores
Neurobiology of Language
480
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Variable
Sex (female/male)
Handedness (left/right)
Alter (in months) – kindergarten behavioral tasks
Alter (in months) – kindergarten MRI
Alter (in months) – third grade behavioral tasks
Verbal intelligence
Nonverbal intelligence
Parental education
Notiz. MRI = magnetic resonance imaging.
a Mean (range) or occurrence (N).
Tisch 1.
Participant characteristics.
Gesamt (n = 162)A
80/82
2011 (n = 87)A
36/51
16/113
63 (56–72)
73 (66–81)
9/65
62 (56–68)
74 (68–81)
Cohort
2018 (n = 75)A
44/31
7/48
5 (59–72)
72 (66–78)
98 (92–105)
98 (93–105)
98 (92–104)
9.8 (3.0–17.0)
9.3 (4.0–15.0)
10.4 (3.0–17.0)
9.7 (3.0–19.0)
10.0 (3.0–18.0)
9.2 (3.0–19.0)
4.0 (2.0–6.0)
3.8 (2.0–6.0)
4.2 (2.0–6.0)
p valueb
0.028
0.923
< 0.001
0.001
0.451
0.012
0.038
0.039
b Group differences were assessed using a Pearson’s Chi-squared test for sex, a Fisher’s exact test for handedness, and a Wilcoxon rank sum test for age,
intelligence, and parental education.
of these three participants were very close to the cut-off values (i.e., −35, −36 and +38), and
for ease of interpretation their handedness was categorized as left-, left-, and right-handed,
respectively. As a proxy for intelligence, we administered the Vocabulary (verbal) and Block
Design (nonverbal) subtests of the Wechsler Intelligence Scale for Children ( WISC-III-NL; Kort
et al., 2005) at the start of second grade. Scores of these subtests were normed using the WISC-
III-NL manual with M = 10 and SD = 3. Parental education, as a proxy for socioeconomic
status (Duncan & Magnuson, 2012), was quantified by the parents’ highest level of education,
ranging from one (high school degree), to two (bachelor degree), and three (master or doctoral
degree). The maternal and paternal educational levels were summated into one continuous
measure with values ranging between two and six (Robitzsch, 2020). For each of these partic-
ipant characteristics, descriptive statistics of the total study sample as well as a comparison
between the two cohorts is shown in Table 1.
Materials and Procedure
Behavioral measures of auditory and speech processing at the pre-reading stage
Two auditory and speech processing tasks were administered at the start of the final year of
kindergarten. Both tasks were performed using APEX software (Francart et al., 2008), and
stimuli were presented using a Fireface UC soundcard (RME Audio, Germany) with calibrated
HDA 200 headphones (Sennheiser, Germany).
An RTD task was used to measure auditory processing of nonspeech sounds. In this task,
participants had to discriminate noise stimuli that differed in rise time, that is, the duration it
takes from the onset of the stimulus to reach maximum amplitude. They were presented with
speech-weighted noise stimuli of 800 ms with a linear fall time of 75 ms at an intensity level of
70 dB SPL (sound pressure level). The rise times were adapted logarithmically, between 15 ms
and 699 ms, during the task. On each trial, the participants heard three stimuli (interstimulus
interval = 350 ms); two reference stimuli with the shortest rise time of 15 ms and one target
stimulus with a longer rise time. The order of target stimulus presentation relative to the
Neurobiology of Language
481
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Oddity paradigm:
A paradigm in which participants
have to identify the stimulus that is
perceptually distinct from other
stimuli (odd one out).
references was randomized across trials and across subjects. Using a three-alternative forced
choice oddity paradigm, they were instructed to identify the target stimulus. The rise time of
the next trial’s target stimulus was adapted using a two-down, one-up staircase procedure. The
discrimination threshold was estimated after eight reversals and calculated as the average of
the last four reversals; the threshold corresponded to the rise time of the target stimulus for
which the participant could still discriminate the target from the reference. The children first
practiced the task, and then they completed a test and a retest measurement (test–retest reli-
ability = 0.74). To minimize both learning and attentional effects, we used the best threshold of
the test and retest measurement. Lower thresholds indicated better performance. The task was
embedded in an interactive computer game (Laneau et al., 2005).
Higher-order auditory processing was assessed using a speech-in-noise perception (SPIN)
task designed specifically for young children, the Leuven Intelligibility Peutertest (van
Wieringen & Wouters, 2023). Consonant–vowel–consonant words were presented to the right
ear in stationary speech-weighted noise. The noise had a fixed level of 65 dB SPL, whereas the
level of the speech was adapted during the course of the task. First, during the training phase,
one list of 11 words was used to ensure understanding of the task procedure. The level of the
speech during the training list was equal to the level of the noise, resulting in a signal-to-noise
ratio (SNR) of 0 dB SNR. Then, during the testing phase, lists of 11 words each were adminis-
tered at SNRs of −2, −5, and −8 dB SNR, meaning the level of the speech was 2, 5, and 8 dB SPL
more quiet than the level of the noise, respectively. The first word of each list served solely to
engage the attention of the participant, whereas the next 10 words were used to assess perfor-
mance. Participants had to repeat the words, or, in cases they could not understand the entire
word, to repeat each phoneme they could understand. Each correctly repeated phoneme was
counted (i.e., phoneme-level scoring), allowing calculation of the percentage of correctly
repeated phonemes per list. A test and a retest list were administered for each SNR. By dividing
the standard deviation of differences between the raw scores of these test and retest measure-
ffiffiffi
, we measured test–retest reliability while taking into account the possibility that
2
ments by
performance improved during testing (i.e., learning effect; Smits & Houtgast, 2005). Measure-
ment error was less than 1.5 dB, in line with previous findings of SPIN tasks (Van den Borre
et al., 2021). For further analyses, we therefore averaged the test and retest values for each
SNR. Using a generalized linear model with cumulative Gaussian distribution, we fitted a psy-
chometric performance-intensity curve for each participant and estimated the SNR at which
50% of the phonemes were still accurately repeated (i.e., speech reception threshold). Lower
speech reception thresholds indicated better performance. The above-mentioned psychometric
analyses were performed in R ( Version 4.1.2; R Core Team, 2021), including the MASS
(Venables & Ripley, 2002) and modelfree (Marin-Franch et al., 2012) packages.
p
Image acquisition and processing of auditory cortex at the pre-reading stage
To observe structural characteristics of the auditory cortex, we collected MRI data at the end of
the final year of kindergarten at the University Hospital of Leuven. The MRI scanner was
replaced with a new scanner of the same type between data collection of the two cohorts.
Thus, for both cohorts, we acquired T1-weighted structural sequences on a 3T Philips Achieva
scanner (Philips, Best, The Netherlands) equipped with a 32-channel head coil. The T1-
sequences had the following parameters: 182 contiguous coronal slices, 9.7 ms repetition
time, 4.61 ms echo time, 8° flip angle, 0.98 × 0.98 × 1.20 mm voxel size and 6 m 22 s acqui-
sition time.
We used a child-friendly protocol, in line with the recommendations of Theys et al. (2014),
to limit excessive head motion that can occur in young children and thus confound structural
Neurobiology of Language
482
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
brain measures (Pardoe et al., 2016; White et al., 2018). Prior to the MRI appointment, the
children were acquainted with the MRI procedure through an informative video. On the
day of the scan, the researcher explained different aspects of undergoing an MRI scan to
the participant (e.g., limiting head movement). Lastly, the actual scanner was decorated to look
less intimidating, and the children could choose a movie to watch while being scanned.
During data processing, we performed a visual quality check for head motion, rated the scans
using Blumenthal’s scale and excluded scans that were contaminated with severe motion arti-
facts (Blumenthal et al., 2002). In total, 20 out of 129 subjects that had MRI data (= 15.5%)
were excluded due to severe motion artifacts (based on the Blumenthal rating). Two indepen-
dent researchers completed the Blumenthal rating for all participants. Interrater reliability, cal-
culated using Cohen’s weighted kappa, was 0.78, indicating substantial agreement.
Pre-processing of the raw T1 images was performed using the recon-all function of Free-
Surfer ( Version 6; Fischl, 2012). The resulting output was then used for an automated segmen-
tation of the TTG. Most previous studies on the anatomy of Heschl’s gyrus relied on the
visual/manual segmentation of this region (Marie et al., 2015; Schneider et al., 2009), but
we used the recently developed TASH (Dalboni da Rocha et al., 2020) to segment the TTG
in a time-efficient, reproducible, and standardized way. TASH was originally developed on
adult brain images. Due to the smaller size of pediatric brains, as used in this study, the TASH
parameter for the automatic check for existing TTG clusters was decreased from 100 to 50
vertices, thus eliminating only clusters smaller than 50 vertices. The bilateral TTG were then
segmented and the surface area (in mm2) of the first TTG and of additional TTG(s), when pres-
ent, were extracted. Note that TASH segments the first TTG as a whole, regardless of the length
of the sulcus intermedius (SI) in the case of a common stem duplication. In three subjects, the
first TTG was not correctly segmented by TASH, and their scans were excluded from further
analyses. TASH segments the gyri specifically, but does not include segmentation of the supe-
rior temporal plane, that is, the superior surface of the superior temporal gyrus (Shapleske
et al., 1999). Therefore, we also extracted surface area of the bilateral PT segmented by Free-
Surfer, using the Destrieux et al. (2010). This automatic PT segmentation, in line with some
studies that used manual segmentations (Hugdahl et al., 2003; Sanchez Bloom et al., 2013;
Shapleske et al., 1999; Steinmetz et al., 1990), was more liberal in defining the posterior bor-
der of the PT compared to other studies, which did not include posterior rami (Altarelli et al.,
2014; Vanderauwera et al., 2018). To quantify surface area asymmetry of the PT, we calculated
the asymmetry index (AI) using the formula AI = [(R − L)/(R + L)/2] as described in Marie and
Golestani (2017). Since surface area was calculated in native space, and to ensure that poten-
tial effects were specific to the TTG, we included the corresponding total hemispheric surface
area as a covariate in the statistical analyses.
To examine TTG duplication patterns, the number of gyri along the bilateral TTG was
assessed visually, blind to participant characteristics. Specifically, the rater (NG) quantified
the number of gyri by giving a value of 1 when a single gyrus was present, a value of 2 when
two fully separated gyri were present, and so on. Further, the ratings of the first TTG (i.e., of
HG) only was not binary but continuous, since it is known that HG can be either a single gyrus
or split in the form of a common stem duplication (Marie et al., 2015). Thus, for HG only, the
rater provided a number ranging between 1 and 1.99 reflecting the degree of gyrification or, in
other words, reflecting the depth of the SI, when present. For example, a single HG got a rating
of 1; a common stem duplication having an SI spanning half the length of HG got a rating of
1.5; and a common stem duplication having a SI spanning 8/10ths of the length of HG got a
rating of 1.8, and so on. The HG value was then simply incremented by 1 every time an addi-
tional, fully separated TTG was present. Since duplication patterns are region specific (Ronan
Neurobiology of Language
483
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
et al., 2014), there was no need to control for any global or corresponding hemispheric value
in the statistical analyses. In summary, auditory cortex morphology was quantified using five
measures: surface area of bilateral first TTG, surface area of bilateral additional TTG(s), surface
area of bilateral PT, surface area asymmetry of PT, and duplication patterns of bilateral TTG(s).
Measure of family risk for dyslexia
Family risk for dyslexia was quantified using the parental self-report Adult Reading History
Questionnaire (ARHQ; Lefly & Pennington, 2000), translated to Dutch from the original
English version (Verwimp et al., 2020). This questionnaire included 23 items on a 5-point
Likert scale and inquired about the parents’ history with learning to read in primary school,
as well as their current experience with and attitude toward reading. To quantify family risk,
we divided the total score by the maximum score of 92, resulting in a continuous variable
ranging between zero and one. Scores closer to one indicated more self-reported reading
difficulties in the parents. Both the biological mother and father were requested to fill in the
questionnaire. We only included the scores if both maternal and paternal scores were avail-
able, and both scores were averaged into one familial risk score for each participant.
Measure of reading ability
As our outcome of interest, we assessed word reading ability at the start of third grade. We
administered the standardized Eén-minuut-toets (Brus & Voeten, 1999), where children had
to read aloud as many words as possible during one minute. The outcome measure was the
number of correctly read words in one minute.
Statistical Analyses
All statistical analyses were performed in R ( Version 4.1.2; R Core Team, 2021), including the
lavaan package (Rosseel, 2012). The analyses script and anonymized data sheet are available
at the Open Science Framework (osf.io/vk8tx). First, an exploratory correlation analysis was
performed to identify which of the pre-reading measures of auditory and speech processing
and auditory cortex were related to third grade word reading. Then, we fitted mediation
models to investigate whether these pre-reading measures mediated the effect of family risk
for dyslexia on third grade word reading outcome (see Figure 1). We fitted the models to
our data using robust maximum likelihood (MLR) estimation and full information maximum
likelihood (FIML) to account for missing data and to ensure usage of all available data. The
model syntax included the direct effect of family risk on reading outcome, the indirect effect
via pre-reading measures, and the total effect combining these direct and indirect paths. We
reported standardized parameter estimates and interpreted effect sizes following the guidelines
of Gignac and Szodorai (2016), who found effects of 0.10, 0.20, and 0.30 to be small, typical,
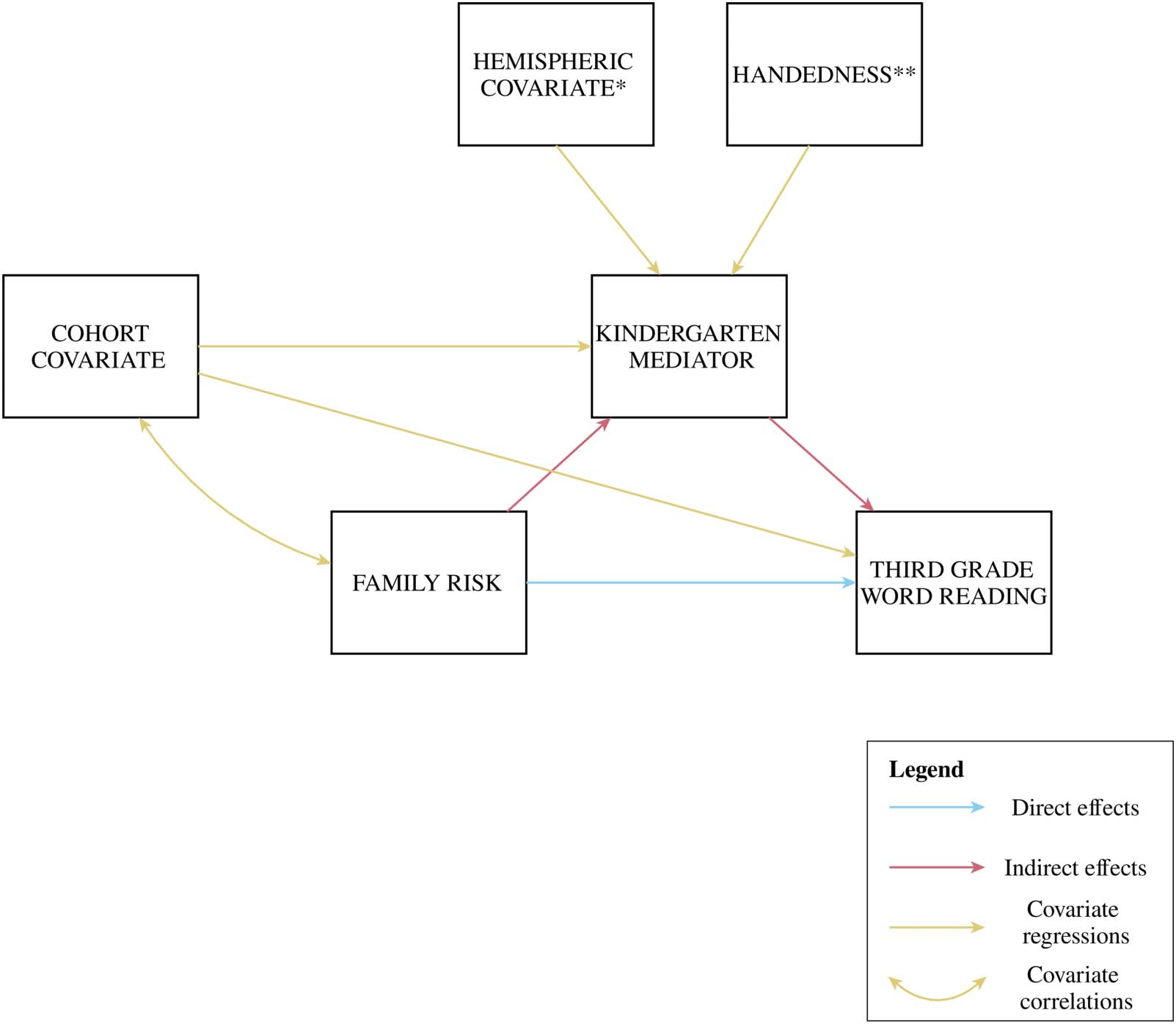
and large, respectively. Additionally, three covariates were taken into account (see Figure 1),
using regressions for endogenous variables and covariances for exogenous variables (Little,
2013). First, we included the cohort variable as a covariate to each mediation model, to
account for variance explained by differences between the cohorts (i.e., COVID-19 pandemic
and associated nationwide school closures [see Participants section]; participant characteris-
tics [see Table 1]; and scanner [see Image Acquisition and Processing of Auditory Cortex at the
Pre-reading Stage]). Second, in the mediation models where surface area was included, we
allowed the local surface area to covary with the corresponding hemispheric surface area
(Genon et al., 2022). Third, given the presence of both left- and right-handed participants in
our study sample, we checked for group differences related to handedness. For each mediation
model including a neuroanatomical measure, we performed the Wilcoxon rank sum test to
Mediation model:
A model that examines the
relationship between an independent
and dependent variable through a
proposed mediating variable.
Endogenous variables:
Variable that are influenced by other
variables in the statistical model,
whereas exogenous variables are not.
Neurobiology of Language
484
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Mediation model. * The hemispheric covariate was only included in the models including surface area, but not in models including
duplication patterns, rise time discrimination, or speech-in-noise perception. ** The handedness covariate was only included in case of a
group difference between left- and right-handed participants.
compare left- and right-handed participants. For the neuroanatomical measures that were sig-
nificantly different between left- and right-handed participants, we added handedness to the
mediation model as a covariate.
Note that before running the mediation analyses, we linearly rescaled some of the variables
as described in Kline (2011). More specifically, RTD thresholds and surface area measures
were divided by 100, word reading scores were divided by 10, and familial risk scores were
multiplied by 10. This procedure resulted in more homogeneous variances across the variables
without affecting the underlying interrelations, which facilitated model estimation.
RESULTS
Descriptive statistics are presented in Table 2. First, we used the Shapiro–Wilk normality test to
assess the extent to which the data followed a normal distribution. This test showed deviations
from a normal distribution for all kindergarten measures except SPIN, surface area of the left and
right first TTG and PT asymmetry (see Supplementary Table 1 in the Supporting Information,
available at https://doi.org/10.1162/nol_a_00111). Family risk and third grade word reading
did not show deviations from a normal distribution. Next, we explored zero-order correlations
to see how the kindergarten measures correlated with third grade word reading (see Table 3).
Given the normality test results, we used Spearman coefficients for bivariate correlations that
Neurobiology of Language
485
Linking pre-reading auditory cortex with reading and family risk
Table 2. Descriptive statistics.
Variable
Family risk for dyslexia
Measure
Parental ARHQ
Kindergarten auditory and speech
RTD threshold
processing
(in ms)
SPIN threshold
(in dB SNR)
Kindergarten surface area first TTG
(in mm2)
Kindergarten surface area additional
TTG(s) (in mm2)
Left
Right
Left
Right
Kindergarten surface area PT (in mm2)
Left
Right
PT surface area asymmetry
Kindergarten TTG duplication patterns
Left
Third grade word reading
Right
Correctly read
words in 1 min.
N
144
158
158
106
106
106
106
106
106
106
106
106
156
Missing
18
Centera
0.32
Spreadb
0.10
Minimum
0.09
Maximum
0.57
4
4
56
56
56
56
56
56
56
56
56
6
289
366
35
648
−5.4
1.3
−9.2
−1.8
296.16
267.68
84.25
85.47
260.50
134.25
151.50
149.00
76.00
81.00
0.00
0.00
484.00
493.00
752.00
437.00
650.00
232.25
299.00
1,269.00
529.00
118.75
221.00
948.00
−0.18
3.00
2.75
0.20
0.70
1.00
38
15
−0.69
1.00
1.00
8
0.34
5.00
4.30
77
Note. TTG = transverse temporal gyrus; PT = planum temporale; ARHQ = Adult Reading History Questionnaire; RTD = rise time discrimination; SPIN = speech-
in-noise perception; SNR = signal-to-noise ratio.
a Center was quantified by the mean for family risk, SPIN, surface area of bilateral first TTG, planum temporale asymmetry, and word reading and by the median
for all other variables.
b Spread was quantified by the standard deviation for family risk, SPIN, surface area of bilateral first TTG, planum temporale asymmetry, and word reading and
by the interquartile range for all other variables.
included at least one non-normally distributed variable (i.e., RTD, surface area of left or right
additional TTG(s), surface area of left or right PT, or duplication patterns of left or right TTG(s)),
and Pearson coefficients for the other bivariate correlations. RTD and SPIN had a significant
negative correlation with word reading. In other words, higher RTD or SPIN thresholds, corre-
sponding to worse performance, were associated with worse word reading scores. Surface area
of left first TTG, surface area of left and right PT, and duplication patterns of left TTG(s) had a
significant positive correlation with word reading. Each of these correlations had an absolute
value of 0.20 or more (typical effect size; Gignac & Szodorai, 2016). The full zero-order corre-
lation table can be found in Supplementary Table 2.
Then, we conducted mediation analyses to test whether the kindergarten measures that cor-
related significantly with reading also acted as mediators of the relationship between family
risk and reading. The standardized estimates and p values for the total, direct, and indirect
effects of these mediation analyses are shown in Table 4.
The total effect provides an estimate of the relationship between family risk and third grade
reading outcome, while taking into account putative mediation through the kindergarten audi-
tory (cortex) measures. Standardized estimates of this relationship indicated a large effect size,
with βs between −0.45 and −0.49 across the mediation models. The total effect was further
Neurobiology of Language
486
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Table 3.
Correlation coefficients between kindergarten measures and third grade word reading.
Pre-reading measure
Rise time discrimination
Speech-in-noise perception
Surface area left first TTG
Surface area right first TTG
Surface area left additional TTG(s)
Surface area right additional TTG(s)
Surface area left PT
Surface area right PT
PT surface area asymmetry
Duplication patterns left TTG(s)
Duplication patterns right TTG(s)
Correlation
r = −0.29, p < 0.001*
r = −0.28, p < 0.001*
r = 0.21, p = 0.032*
r = 0.14, p = 0.15
r = −0.08, p = 0.426
r = 0.16, p = 0.104
r = 0.26, p = 0.007*
r = 0.22, p = 0.021*
r = −0.09, p = 0.336
r = 0.20, p = 0.038*
r = 0.02, p = 0.823
Note. Coefficients denoted with asterisks are significant at α = 0.05. TTG = transverse temporal gyrus; PT =
planum temporale.
divided into an estimation of the direct effect of family risk on reading outcome, and of the
indirect effect running through the kindergarten auditory (cortex) measures. The direct effect
was significant in each of the mediation models, with βs between −0.41 and −0.48 (large
effect sizes). In other words, every increase by one standard deviation in family risk was
associated with a decrease in third grade word reading outcome of between 0.41 and 0.48
standard deviations.
Concerning the indirect paths, family risk significantly predicted kindergarten RTD and
SPIN (typical effect sizes of β = 0.23 and β = 0.22, respectively), with higher family risk pre-
dicting higher/worse RTD or SPIN thresholds. In turn, kindergarten RTD and SPIN predicted
third grade word reading (small effect sizes of β = −0.18 and β = −0.19, respectively), with
Table 4.
Standardized estimates and associated p values of the direct, indirect, and total effects of all mediation analyses.
Mediator
Rise time discrimination
Speech-in-noise perception
Surface area left first TTG
Surface area left PT
Surface area right PT
Duplication patterns left TTG(s)
Direct + Indirect
β
−0.46
−0.45
−0.47
−0.46
−0.45
−0.49
p
< 0.001*
< 0.001*
< 0.001*
< 0.001*
< 0.001*
< 0.001*
−0.41
−0.47
−0.44
−0.46
−0.48
Total effect
Direct effect
FR → Reading
β
−0.42
p
< 0.001*
FR → Mediator
β
0.23
p
0.001*
Indirect effect
Mediator →
Reading
p
0.033*
β
−0.18
FR → Mediator →
Reading
β
−0.04
−0.04
p
0.057
0.050
< 0.001*
0.22
0.002*
−0.19
0.006*
< 0.001*
0.02
0.864
0.18
0.026*
0.00
0.864
< 0.001*
−0.10
0.159
0.18
0.026*
−0.02
0.226
< 0.001*
0.06
0.339
0.20
0.024*
0.01
0.375
< 0.001*
−0.06
0.588
0.15
0.056
−0.01
0.587
Note. Parameters denoted with asterisks are significant at α = 0.05.
Neurobiology of Language
487
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
higher/worse RTD or SPIN thresholds predicting lower word reading skills. However, the
indirect effect combining these two paths (β = −0.04 for both mediation models) as well
as the proportion mediated (the ratio of the indirect effect to the total effect: 0.09 or 9%
for both mediation models) were very small. As a result, these indirect paths were not statis-
tically significant (p = 0.057 and p = 0.050, respectively). Further, we found no evidence
that any of the kindergarten neuroanatomical measures mediated the family risk—reading
relationship. Even though family risk showed a significant negative correlation with surface
area of the left PT (rs = −0.23; see Supplementary Table 2), the standardized estimate of the
path between family risk and this neuroanatomical measure in the mediation model was very
small and not significant (β = −0.10, p = 0.159). Family risk also did not significantly predict
any of the other pre-reading neuroanatomical measures. Results did show a significant
positive effect of surface area of the left first TTG, surface area of the left PT, and surface
area of the right PT on third grade reading, in line with the exploratory correlations (see
Table 3), with small effects sizes (βs between 0.18 and 0.20). Duplication patterns of the left
TTG(s) also correlated positively with reading (see Table 3) but this relationship did not
survive in the mediation model (β = 0.15, p = 0.056).
All of the results shown in Table 4 were present above and beyond covariate effects of cohort
and hemispheric surface area. Separate estimates for each path in the models (including the
covariate effects) can be found in Supplementary Table 3a through Supplementary Table 3f in
the Supporting Information. With regard to handedness, there were no group differences between
left- and right-handed participants for surface area of the left first TTG (W = 644, p = 0.653),
surface area of the left PT (W = 662, p = 0.534), and left TTG duplication patterns (W = 716,
p = 0.214). Surface area of the right PT did differ significantly between left- and right-handed
participants (W = 795, p = 0.047). Handedness was therefore added as a covariate to the medi-
ation model of the right PT, and this did not alter the pattern of results. Since we did not find
evidence for mediation of the family risk–reading relationship by any of the pre-reading auditory
(cortex) measures, we investigated post hoc whether pre-reading phonological awareness had a
mediating role. Descriptive statistics and mediation results are reported in Supplementary Table 4a
and Supplementary Table 4b, respectively. In short, we did not find evidence that pre-reading
phonological awareness mediated the family risk–reading relationship.
DISCUSSION
In the current longitudinal study, we investigated whether or not word reading in third grade
can be predicted by nonspeech (RTD) and by speech (SPIN) processing, and by anatomy of the
bilateral TTG(s) and PT in kindergarten. We found relationships between pre-reading RTD,
SPIN, surface area of the left first TTG, left PT, and right PT and duplication patterns of the left
TTG(s) with later word reading. Moreover, we looked at the possible mediating role of these
measures on the relationship between family risk and reading. Although family risk did
influence RTD and SPIN, we found no evidence for mediation of the family risk–reading rela-
tionship by any of the kindergarten measures under study.
Predictors of Reading Outcome
First and foremost, we found a very large effect of family risk for dyslexia on third grade word
reading outcome. This effect has been reported in many previous studies examining typical
(Andreola et al., 2021) and atypical (Snowling & Melby-Lervåg, 2016) reading development,
and as a result, family risk is generally considered a robust predictor of reading outcome. We
also found a longitudinal effect of pre-reading auditory and speech processing on later word
Neurobiology of Language
488
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
reading skills. Poor thresholds of both RTD and SPIN measured in kindergarten were predictive
of poor word reading outcome in third grade. By analogy, better auditory thresholds predicted
better word reading outcome. Although some previous studies could not find direct links
between RTD and reading (Johnson et al., 2009; Papadopoulos et al., 2012) or SPIN and read-
ing (Miller et al., 2018; Vanvooren et al., 2017), the results of the current study are in line with
other research reporting involvement of auditory factors in reading development. For example,
cross-sectional studies in school-aged children reported an association between RTD and
reading (Corriveau et al., 2007) and between SPIN and reading (Ziegler et al., 2009). The
cross-sectional design of these studies limited their analyses to concurrent associations, but
the findings have been extended in longitudinal studies starting at the pre-reading stage and
following up children throughout their reading development. Consistent with our longitudinal
results, these latter studies found direct effects of pre-reading RTD (Law et al., 2017; Plakas
et al., 2013) and pre-reading SPIN (Boets et al., 2008, 2011; Vandewalle et al., 2012;
White-Schwoch et al., 2015) on later reading outcome.
Compared to the extensive literature reporting auditory and speech processing links to
reading, structural properties of pre-reading auditory cortex in relation to reading outcome
are less documented. To the best of our knowledge, only two studies with small samples sizes
looked at properties of HG specifically at the pre-reading stage in relation to the development
of dyslexia. Children who later developed dyslexia demonstrated pre-reading thinner cortex
(Clark et al., 2014) and a higher local curvature perpendicular to the cortex (also known as
gyrification index; Kuhl et al., 2020) in left HG. Regarding the former study (Clark et al., 2014),
methodological concerns have been described that challenge their conclusions (Kraft et al.,
2015; Ramus et al., 2018). Other studies investigating properties of HG in school-aged chil-
dren and adults with dyslexia reported lower left volume (Xia et al., 2016), lower left and right
volume (Serrallach et al., 2016), higher right thickness (Ma et al., 2015), or no differences in
cortical thickness (Altarelli et al., 2014; Skeide et al., 2018). In sum, findings still vary a lot
across studies, both with regard to the specific properties of HG and the direction of findings.
Our results confirm a predictive role of auditory cortex anatomy in reading outcome, beyond a
binary classification of typical versus poor reading skills, by showing predictive effects on later
reading outcome measured continuously. More specifically, we found that pre-reading surface
area of the left first TTG (including the posterior branch of common stem duplications, regard-
less of the length of the intermediate sulcus, when present) positively predicted later reading
outcome.
Additional gyri of TTG (posterior to the first TTG), which are part of the PT, have also been a
subject of investigation in dyslexia research, as previous studies have shown a higher likeli-
hood of full posterior duplications in this reading disorder. Notably, the automatic toolbox for
segmenting the TTG that we applied in the current study (TASH; Dalboni da Rocha et al.,
2020), included quantification of the surface area of the gyri, but not of the non-gyrus portions
of the superior temporal plane. Therefore, we looked at both the surface area of the additional
TTG(s) posterior to the first TTG segmented by TASH and at the surface area of the PT
segmented by FreeSurfer. Our analyses showed that the surface area of left and right PT was
positively predictive of later reading outcome. Interestingly, surface area of additional TTG(s)
did not predict word reading, even though there is some anatomical overlap between the
surface area of additional TTG(s) and PT. This suggests that the relationship with reading is
driven by the variation in the PT itself, as opposed to merely the surface area of the gyri.
However, it could also be that a relationship for the PT was easier to detect since surface area
of the PT is larger compared to the surface area of only the gyri. Together, our results showing
predictive strength of variations in TTG and PT anatomy suggest that there may also be other
Neurobiology of Language
489
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
brain functional as well as functional and/or structural connectivity differences within net-
works including these regions that are crucial to the development of a healthy reading net-
work. Functional brain imaging studies suggest that HG is primarily involved in the processing
of low-level auditory information and that the PT is involved in speech sound processing
(Griffiths & Warren, 2002; Hickok & Poeppel, 2007). However, the role of PT is still debated
and could also involve top-down processes from higher-order regions such as the inferior
frontal gyrus, inferior parietal cortex, superior temporal sulcus, and middle temporal gyrus
(Davis et al., 2011; Friederici, 2012; Golestani et al., 2013; Rutten et al., 2019).
As described above, we found a positive relationship with reading of both surface area of
the left first TTG and of the left PT. This is in line with results of Xia et al. (2016), who found
lower gray matter volume of both HG and PT in children with dyslexia. However, it is inter-
esting that Serrallach et al. (2016) reported a different pattern in children with dyslexia, with
gray matter volume of PT having an inverse effect compared to HG. While both cross-sectional
studies included participants in the same age range (age 10 and older), there were large dif-
ferences in their analyses approach, which could have contributed to this inconsistency. On
the one hand, Xia et al. (2016) first performed a whole brain, voxel-based analysis and then
investigated significant clusters. This resulted in several clusters, of which one spanned HG
and PT, but which did not specifically distinguish between the two regions. On the other hand,
Serrallach et al. (2016) applied a semiautomated method to segment HG and PT specifically,
which is a more reliable method for differentiating the effects arising from HG from those aris-
ing from the PT. In the current longitudinal study, we do not find the inverse effect, indicating
that greater pre-reading surface area of both the left first TTG and of the left PT can be bene-
ficial for later word reading performance.
With regard to the duplication patterns of the TTG(s), we found a significant positive cor-
relation between pre-reading left TTG(s) duplication patterns and third grade word reading.
Greater left TTG(s) duplications in kindergarten were associated with better third grade word
reading. This relationship is in line with other research reporting that the presence of two or
more gyri is more common in language aptitude and in musical expertise (Benner et al., 2017;
Dalboni da Rocha et al., 2023; Golestani et al., 2011; Schneider et al., 2009; Turker et al.,
2019). However, it does not explain why, on average, studies have reported more duplications
in children (Altarelli et al., 2014; Serrallach et al., 2016) and in adults with dyslexia (Leonard
et al., 2001). Notably, this relationship did not survive in the mediation model where family
risk was also included. We speculate that including family risk caused a decrease in this effect.
However, given the p value ( p = 0.056) in our analyses, it would be of interest to examine the
effect of TTG(s) duplication patterns in future research by applying statistical inference such as
Bayesian statistics that would allow quantification of evidence in favor of or against the null
hypothesis (Dienes, 2014).
Nevertheless, the effects of the auditory cortex anatomy (both surface area and duplication
patterns) further corroborate our results showing that pre-reading auditory and speech process-
ing play a role in the prediction of later word reading outcome, consistent with recent theo-
retical accounts of reading development, such as the intergenerational multiple deficit model
(IMDM; van Bergen et al., 2014). In this model, it is posited that the interplay between multiple
factors at multiple levels, including the processing of auditory information and neural proper-
ties, underlie the development of severe reading difficulties (dyslexia). Such dynamic interre-
lations are not only involved in atypical reading development, but also assumed to also exist
for the entire distribution of reading abilities in the general population (van Atteveldt et al.,
2021). According to these theoretical models, and supported by recent molecular genetic stud-
ies (Doust et al., 2022), genetics are also important to consider.
Neurobiology of Language
490
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Mediation of the Family Risk–Reading Relationship
We investigated whether the large effect of family risk on later reading described above was
mediated by any of the kindergarten auditory (cortex) measures by applying mediation anal-
yses in a relatively large sample that had varying degrees of family risk. Family risk had a sig-
nificant effect on pre-reading RTD and SPIN, in agreement with findings of neurophysiological
studies suggesting a contribution of family risk to auditory processing (van der Leij et al.,
2013). Although RTD and SPIN in turn had a significant effect on later reading (see Predictors
of Reading Outcome, above), neither RTD nor SPIN significantly mediated the effect of family
risk on reading. The proportion mediated by RTD or SPIN was very small (9%) and not signif-
icant in light of the very large direct effect of family risk on reading. Thus, while predisposition
for reading difficulties seems to be related to how well people can process auditory input and
degraded speech input, this relationship does not seem to contribute more generally to reading
outcome. This could be because additional processes come into play to compensate for audi-
tory and degraded speech processing, for example, processes involving harnessing top-down
resources to compensate for degraded bottom-up input (Giraud & Ramus, 2013; Mattys et al.,
2012). Arguably, processing skills that are related more closely to reading outcomes, such as
phonological processing skills, are more likely to mediate the family risk–reading relationship.
Although it was not within the scope of the current study, we did check post hoc whether pre-
reading phonological processing (measured using syllable- and phoneme-level tasks) had a
mediating role (see Supplementary Table 4b). Results of this supplementary analysis did not
reveal mediation by these measures, in contrast to the findings of van Bergen et al. (2015).
Further, we found no evidence that family risk predicted any of the kindergarten neuroan-
atomical measures. By extension, these measures also did not play a mediating role in the
family risk–reading relationship. This was unexpected in light of previous studies suggesting
a relationship between family risk and brain structural differences in auditory and temporal
cortices beyond HG (Beelen et al., 2019; Vanderauwera et al., 2018; note that our participants
partly overlapped with these previous studies). Surface area of the left first TTG, left PT, and
right PT in kindergarten did predict third grade word reading, as discussed in Predictors of
Reading Outcome (above), indicating that these measures were related to reading indepen-
dently of family risk.
It is noteworthy that family risk and surface area of the left PT correlated negatively and
significantly, and that the parameter estimate within the mediation model was larger in abso-
lute value compared to the estimates of the family risk paths to the other neuroanatomical
measures. However, this relationship was not significant in the mediation model. Arguably,
the measure we used to estimate the putative predisposition-driven contributions in our medi-
ation analyses, though valid, is only an indirect proxy of genetic risk. We assessed family risk
for dyslexia using the continuous outcome of a parental self-report questionnaire. We chose an
indirect measure for its time- and cost-efficiency, and because it has been shown to be an
adequate proxy for the heritability of reading skills (Swagerman et al., 2017), as well as a valid
alternative for parents’ actual reading skills (Khanolainen et al., 2023). We recognize the need
for replication using direct genetic data compiled in polygenic risk scores (Belsky & Harden,
2019) or by applying the familial control method as proposed by Hart et al. (2021), since our
measure of family risk confounds influences of environment and genetics (Hart et al., 2021).
Furthermore, our family risk measure captures information about the parents’ current and past
reading behavior, which might be more closely related to their offspring’s behavioral outcomes
than to properties of the auditory cortex. Thus, it remains unknown whether a similar pattern of
results would be present when indexing family risk using a parental brain measure, as
Polygenic risk scores:
An estimate of genetic risk based on
the combined effects of multiple
genetic variants.
Neurobiology of Language
491
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
suggested by previous studies looking at the structure of other regions in the reading network
(Fehlbaum et al., 2022), connectivity within the reading network (Vandermosten, Schevenels,
et al., 2020), and sulcal patterns across the whole brain (Ahtam et al., 2021).
Overall, the absence of mediation effects in the current study is consistent with an additive
risk model, indicating that the auditory processing and auditory cortex measures contribute to
reading independently from family risk (Bishop, 2006, 2013). Other unmeasured behavioral
and neural mediators, especially in interaction with each other (Galaburda et al., 2006; Giraud
& Ramus, 2013), might still contribute to the family risk–reading relationship. Phonological
processing would be a reasonable alternative (van Bergen et al., 2015), although a supplemen-
tary analysis could not confirm this in the current study sample. Perhaps, given the indirect
evidence for family risk effects in brain regions of the temporal cortex (Beelen et al., 2019;
Łuniewska et al., 2019), other possible mediators include white matter connections in the neu-
ral reading network that pass through temporal regions such as the inferior fronto-occipital
fasciculus (see, e.g., Vanderauwera et al., 2017; Vandermosten et al., 2017).
Methodological Considerations and Recommendations
Apart from our indirect family risk measure discussed above, some other methodological con-
siderations of the current study are also important to address. First, we looked to elucidate
which regions of the auditory cortex specifically related to later reading, using different audi-
tory cortex measures (surface area and duplication patterns) of both HG and the PT. While
exploratory, we found small significant effects in a study sample that was relatively large
compared to previous studies looking at pre-reading auditory cortex in relation to reading.
Nevertheless, we acknowledge the need for replication in independent samples in order to
demonstrate the robustness of these findings, especially given the challenges that come with
pediatric brain imaging (Turesky et al., 2021). The development of large publicly available
data sets such as the ABCD Data Repository (Casey et al., 2018) is very promising for this
purpose, but currently, large-scale longitudinal data sets that include both behavioral and
brain measures starting in the pre-reading stage are still lacking.
Second, since auditory processing relies on efficient processing of temporal information, we
do not rule out putative mediation through other structural features, such as the microstructural
property of myelination, which is known to facilitate efficient processing (Long et al., 2018). It
has been suggested that myelination and white matter more generally are in part responsible for
predicting individual differences in non-speech processing (Warrier et al., 2009) and reading
skill (Marie et al., 2018), and myelination differences have been found in the auditory cortex of
adults with and without dyslexia (Skeide et al., 2018). In addition, the myelination differences
found in two of the studies described above (Marie et al., 2018; Skeide et al., 2018) were local-
ized to the PT. Finally, although PT surface area asymmetry did not correlate with third grade
reading outcome in the current study, this neural measure did correlate positively with family
risk (higher family risk associated with a more rightward lateralization, see Supplementary
Table 2). Taken together with findings of altered PT asymmetries in relation to family risk
(Vanderauwera et al., 2018) and dyslexia (Altarelli et al., 2014; Galaburda et al., 1985; Sanchez
Bloom et al., 2013), and findings of altered HG to PT volume ratios in dyslexia (Serrallach et al.,
2016), mediation of the family risk–reading relationship cannot be ruled out entirely. Extension of
our approach to regions beyond those examined in the current study and to neural measures such
as myelination and functional activation could benefit the understanding of the complex inter-
play between the relative influences of predisposition and of the environment on different levels,
as described in the IMDM, involved during reading development.
Myelination:
Refers to the myelin sheaths around
neurons that allow for faster and
more efficient neural transmission.
Neurobiology of Language
492
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
CONCLUSION
In conclusion, we found predictive effects of pre-reading auditory and speech processing and
of auditory cortex anatomy on later word reading outcome. Even though we also found some
evidence for family risk effects on these pre-reading auditory measures, they did not mediate
the strong relationship between family risk and later reading. This study contributes to under-
standing to what extent auditory measures are involved in reading acquisition. As such, it may
help to guide future research, to ultimately help clinicians in achieving early identification of
and intervention for children at risk for severe reading difficulties.
ACKNOWLEDGMENTS
We thank all children and parents for their cooperation in our study. We would like to thank
the researchers involved in longitudinal data collection of the first cohort: Sophie Dandache,
Astrid De Vos, Jolijn Vanderauwera, and Sophie Vanvooren. We are also grateful to all students
who assisted in data collection.
FUNDING INFORMATION
Maaike Vandermosten, Fonds Wetenschappelijk Onderzoek (https://dx.doi.org/10.13039
/501100003130), Award ID: G077018N. Narly Golestani, Swiss National Science Foundation,
Award ID: 100014_182381. Narly Golestani, National Centre of Competence in Research
E v o l v i n g L a n g u a g e ( h t t p s : / / d x . d o i . o rg / 1 0 . 1 3 0 3 9 / 5 0 11 0 0 0 2 3 5 5 5 ) , Aw a r d I D :
#51NF40_180888.
AUTHOR CONTRIBUTIONS
Lauren Blockmans: Conceptualization; Data curation; Formal analysis; Investigation; Method-
ology; Software; Visualization; Writing – original draft; Writing – review & editing. Narly
Golestani: Conceptualization; Formal analysis; Methodology; Software; Supervision; Writing –
review & editing. Josué Luiz Dalboni da Rocha: Formal analysis; Software; Writing – review &
editing. Jan Wouters: Conceptualization; Funding acquisition; Resources; Supervision; Writing –
review & editing. Pol Ghesquière: Conceptualization; Funding acquisition; Resources; Supervi-
sion; Writing – review & editing. Maaike Vandermosten: Conceptualization; Funding acquisition;
Methodology; Resources; Supervision; Writing – review & editing.
DATA AND CODE AVAILABILITY STATEMENT
The analyses script and anonymized data sheet are available at the Open Science Framework
(osf.io/vk8tx).
REFERENCES
Ahtam, B., Turesky, T. K., Zöllei, L., Standish, J., Grant, P. E., Gaab,
N., & Im, K. (2021). Intergenerational transmission of cortical
sulcal patterns from mothers to their children. Cerebral Cortex,
31(4), 1888–1897. https://doi.org/10.1093/cercor/ bhaa328,
PubMed: 33230560
Altarelli, I., Leroy, F., Monzalvo, K., Fluss, J., Billard, C., Dehaene-
Lambertz, G., Galaburda, A. M., & Ramus, F. (2014). Planum
temporale asymmetry in developmental dyslexia: Revisiting an
old question. Human Brain Mapping, 35(12), 5717–5735.
https://doi.org/10.1002/hbm.22579, PubMed: 25044828
Andreola, C., Mascheretti, S., Belotti, R., Ogliari, A., Marino, C.,
Battaglia, M., & Scaini, S. (2021). The heritability of reading and
reading-related neurocognitive components: A multi-level meta-
analysis. Neuroscience and Biobehavioral Reviews, 121, 175–200.
https://doi.org/10.1016/j.neubiorev.2020.11.016, PubMed: 33246020
Astle, D. E., Holmes, J., Kievit, R., & Gathercole, S. E. (2022).
Annual Research Review: The transdiagnostic revolution in neu-
rodevelopmental disorders. Journal of Child Psychology and Psy-
chiatry, 63(4), 397–417. https://doi.org/10.1111/jcpp.13481,
PubMed: 34296774
Neurobiology of Language
493
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Beelen, C., Vanderauwera, J., Wouters, J., Vandermosten, M., &
Ghesquière, P. (2019). Atypical gray matter in children with
dyslexia before the onset of reading instruction. Cortex, 121,
399–413. https://doi.org/10.1016/j.cortex.2019.09.010,
PubMed: 31704534
Belsky, D. W., & Harden, K. P. (2019). Phenotypic annotation:
Using polygenic scores to translate discoveries from genome-
wide association studies from the top down. Current Directions
in Psychological Science, 28(1), 82–90. https://doi.org/10.1177
/0963721418807729
Benner, J., Wengenroth, M., Reinhardt, J., Stippich, C., Schneider,
P., & Blatow, M. (2017). Prevalence and function of Heschl’s
gyrus morphotypes in musicians. Brain Structure and Function,
222(8), 3587–3603. https://doi.org/10.1007/s00429-017-1419-x,
PubMed: 28397108
Bishop, D. V. M. (2006). Developmental cognitive genetics: How
psychology can inform genetics and vice versa. Quarterly Journal
of Experimental Psychology, 59(7), 1153–1168. https://doi.org/10
.1080/17470210500489372, PubMed: 16769616
Bishop, D. V. M. (2013). Cerebral asymmetry and language devel-
opment: Cause, correlate, or consequence? Science, 340(6138),
Article 1230531. https://doi.org/10.1126/science.1230531,
PubMed: 23766329
Blockmans, L., Kievit, R., Wouters, J., Ghesquière, P., & Vandermosten,
M. (2023). Dynamics of cognitive predictors during reading
acquisition in a sample of children overrepresented for dyslexia
risk. Developmental Science. Advance online publication. https://
doi.org/10.1111/desc.13412, PubMed: 37219071
Blumenthal, J. D., Zijdenbos, A., Molloy, E., & Giedd, J. N. (2002).
Motion artifact in magnetic resonance imaging: Implications for
automated analysis. NeuroImage, 16(1), 89–92. https://doi.org/10
.1006/nimg.2002.1076, PubMed: 11969320
Boets, B., Ghesquière, P., van Wieringen, A., & Wouters, J. (2007).
Speech perception in preschoolers at family risk for dyslexia:
Relations with low-level auditory processing and phonological
ability. Brain and Language, 101(1), 19–30. https://doi.org/10
.1016/j.bandl.2006.06.009, PubMed: 16887179
Boets, B., Vandermosten, M., Poelmans, H., Luts, H., Wouters, J., &
Ghesquière, P. (2011). Preschool impairments in auditory pro-
cessing and speech perception uniquely predict future reading
problems. Research in Developmental Disabilities, 32(2),
560–570. https://doi.org/10.1016/j.ridd.2010.12.020, PubMed:
21236633
Boets, B., Wouters, J., van Wieringen, A., & Ghesquière, P. (2006).
Auditory temporal information processing in preschool children at
family risk for dyslexia: Relations with phonological abilities and
developing literacy skills. Brain and Language, 97(1), 64–79.
https://doi.org/10.1016/j.bandl.2005.07.026, PubMed: 16112723
Boets, B., Wouters, J., van Wieringen, A., de Smedt, B., &
Ghesquière, P. (2008). Modelling relations between sensory
processing, speech perception, orthographic and phonological
ability, and literacy achievement. Brain and Language, 106(1),
29–40. https://doi.org/10.1016/j.bandl.2007.12.004, PubMed:
18207564
Boets, B., Wouters, J., van Wieringen, A., & Ghesquière, P. (2007).
Auditory processing, speech perception and phonological ability
in pre-school children at high-risk for dyslexia: A longitudinal
study of the auditory temporal processing theory. Neuropsycho-
l o g i a , 4 5 ( 8 ) , 1 6 0 8 – 1 6 2 0 . h t t p s : / / d o i . o r g / 1 0 . 1 0 1 6 / j
.neuropsychologia.2007.01.009, PubMed: 17303197
Brus, T., & Voeten, M. (1999). Eén-minuut-test (EMT). Harcourt.
Calcus, A., Hoonhorst, I., Colin, C., Deltenre, P., & Kolinsky, R.
(2018). The “rowdy classroom problem” in children with
dyslexia: A review. In T. Lachmann & T. Weis (Eds.), Reading
and dyslexia (pp. 183–211). Springer. https://doi.org/10.1007
/978-3-319-90805-2_10
Casey, B. J., Cannonier, T., Conley, M. I., Cohen, A. O., Barch,
D. M., Heitzeg, M. M., Soules, M. E., Teslovich, T., Dellarco,
D. V., Garavan, H., Orr, C. A., Wager, T. D., Banich, M. T., Speer,
N. K., Sutherland, M. T., Riedel, M. C., Dick, A. S., Bjork, J. M.,
Thomas, K. M., … Dale, A. M. (2018). The Adolescent Brain Cog-
nitive Development (ABCD) study: Imaging acquisition across 21
sites. Developmental Cognitive Neuroscience, 32, 43–54. https://
doi.org/10.1016/j.dcn.2018.03.001, PubMed: 29567376
Chyl, K., Fraga-González, G., Brem, S., & Jednoróg, K. (2021). Brain
dynamics of (a)typical reading development—A review of longi-
tudinal studies. NPJ Science of Learning, 6(1), Article 4. https://
doi.org/10.1038/s41539-020-00081-5, PubMed: 33526791
Clark, K. A., Helland, T., Specht, K., Narr, K. L., Manis, F. R., Toga,
A. W., & Hugdahl, K. (2014). Neuroanatomical precursors of
dyslexia identified from pre-reading through to age 11. Brain,
137(12), 3136–3141. https://doi.org/10.1093/ brain/awu229,
PubMed: 25125610
Clayton, F. J., West, G., Sears, C., Hulme, C., & Lervåg, A. (2020). A
longitudinal study of early reading development: Letter-sound
knowledge, phoneme awareness and RAN, but not letter-sound
integration, predict variations in reading development. Scientific
Studies of Reading, 24(2), 91–107. https://doi.org/10.1080
/10888438.2019.1622546
Corriveau, K. H., Goswami, U., & Thomson, J. M. (2010). Auditory
processing and early literacy skills in a preschool and kindergar-
ten population. Journal of Learning Disabilities, 43(4), 369–382.
https://doi.org/10.1177/0022219410369071, PubMed:
20457882
Corriveau, K. [H.], Pasquini, E., & Goswami, U. (2007). Basic audi-
tory processing skills and specific language impairment: A new
look at an old hypothesis. Journal of Speech, Language, and
Hearing Research, 50(3), 647–666. https://doi.org/10.1044/1092
-4388(2007/046), PubMed: 17538107
Dalboni da Rocha, J. L., Kepinska, O., Schneider, P., Benner, J.,
Degano, G., Schneider, L., & Golestani, N. (2023). Multivariate
Concavity Amplitude Index (MCAI) for characterizing Heschl’s
gyrus shape. NeuroImage, 272, Article 120052. https://doi.org
/10.1016/j.neuroimage.2023.120052, PubMed: 36965861
Dalboni da Rocha, J. L., Schneider, P., Benner, J., Santoro, R.,
Atanasova, T., Van De Ville, D., & Golestani, N. (2020). TASH:
Toolbox for the Automated Segmentation of Heschl’s Gyrus. Sci-
entific Reports, 10, Article 3887. https://doi.org/10.1038/s41598
-020-60609-y, PubMed: 32127593
Davis, M. H., Ford, M. A., Kherif, F., & Johnsrude, I. S. (2011). Does
semantic context benefit speech understanding through “top–
down” processes? evidence from time-resolved sparse fMRI.
Journal of Cognitive Neuroscience, 23(12), 3914–3932. https://
doi.org/10.1162/jocn_a_00084, PubMed: 21745006
Dienes, Z.
Destrieux, C., Fischl, B., Dale, A., & Halgren, E. (2010). Automatic
parcellation of human cortical gyri and sulci using standard ana-
tomical nomenclature. NeuroImage, 53(1), 1–15. https://doi.org
/10.1016/j.neuroimage.2010.06.010, PubMed: 20547229
(2 014). Usin g Bayes to get
th e m ost out of
non-significant results. Frontiers in Psychology, 5, Article 781.
https://doi.org/10.3389/fpsyg.2014.00781, PubMed: 25120503
Doust, C., Fontanillas, P., Eising, E., Gordon, S. D., Wang, Z.,
Alagöz, G., Molz, B., 23andMe Research Team, Quantitative
Trait Working Group of the GenLang Consortium, St Pourcain,
B., Francks, C., Marioni, R. E., Zhao, J., Paracchini, S., Talcott,
J. B., Monaco, A. P., Stein, J. F., Gruen, J. R., Olson, R. K., …
Neurobiology of Language
494
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
n
o
_
a
_
0
0
1
1
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Luciano, M. (2022). Discovery of 42 genome-wide significant
loci associated with dyslexia. Nature Genetics, 54(11),
1621–1629. https://doi.org/10.1038/s41588-022-01192-y,
PubMed: 36266505
Duncan, G. J., & Magnuson, K. (2012). Socioeconomic status and
cognitive functioning: Moving from correlation to causation.
Wiley Interdisciplinary Reviews: Cognitive Science, 3(3), 377–386.
https://doi.org/10.1002/wcs.1176, PubMed: 26301469
Eckert, M. A., & Leonard, C. M. (2000). Structural imaging in dys-
lexia: The planum temporale. Mental Retardation and Develop-
mental Disabilities Research Reviews, 6(3), 198–206. https://doi
.org/10.1002/1098-2779(2000)6:3<198::AID-MRDD7>3.0.CO
;2-1, PubMed: 10982497
Erbeli, F., Rice, M., & Paracchini, S. (2022). Insights into dyslexia
genetics research from the last two decades. Gehirnwissenschaften,
12(1), Article 27. https://doi.org/10.3390/ brainsci12010027,
PubMed: 35053771
Fehlbaum, L. V., Peters, L., Dimanova, P., Roell, M., Borbás, R.,
Ansari, D., & Raschle, N. M. (2022). Mother-child similarity in
brain morphology: A comparison of structural characteristics of
the brain’s reading network. Developmental Cognitive Neurosci-
enz, 53, Article 101058. https://doi.org/10.1016/j.dcn.2022
.101058, PubMed: 34999505
Fischl, B. (2012). FreeSurfer. NeuroImage, 62(2), 774–781. https://
doi.org/10.1016/j.neuroimage.2012.01.021, PubMed: 22248573
Francart, T., van Wieringen, A., & Wouters, J. (2008). APEX 3: A
multi-purpose test platform for auditory psychophysical experi-
gen. Journal of Neuroscience Methods, 172(2), 283–293.
https://doi.org/10.1016/j.jneumeth.2008.04.020, PubMed:
18538414
Friederici, A. D. (2012). The cortical language circuit: From audi-
tory perception to sentence comprehension. Trends im kognitiven Bereich
Wissenschaften, 16(5), 262–268. https://doi.org/10.1016/j.tics.2012.04
.001, PubMed: 22516238
Galaburda, A. M., LoTurco, J., Ramus, F., Fitch, R. H., & Rosen,
G. D. (2006). From genes to behavior in developmental dyslexia.
Naturneurowissenschaften, 9(10), 1213–1217. https://doi.org/10.1038
/nn1772, PubMed: 17001339
Galaburda, A. M., Sherman, G. F., Rosen, G. D., Aboitiz, F., &
Geschwind, N. (1985). Developmental dyslexia: Four consecu-
tive patients with cortical anomalies. Annals of Neurology,
18(2), 222–233. https://doi.org/10.1002/ana.410180210,
PubMed: 4037763
Gennatas, E. D., Avants, B. B., Wolf, D. H., Satterthwaite, T. D.,
Ruparel, K., Ciric, R., Hakonarson, H., Gur, R. E., & Gur, R. C.
(2017). Age-related effects and sex differences in gray matter
density, Volumen, mass, and cortical thickness from childhood
to young adulthood. Zeitschrift für Neurowissenschaften, 37(20),
5065–5073. https://doi.org/10.1523/JNEUROSCI.3550-16.2017,
PubMed: 28432144
Genon, S., Eickhoff, S. B., & Kharabian, S. (2022). Linking interin-
dividual variability in brain structure to behaviour. Natur
Reviews Neuroscience, 23(5), 307–318. https://doi.org/10.1038
/s41583-022-00584-7, PubMed: 35365814
Gerrits, E., & de Bree, E. (2009). Early language development of
children at familial risk of dyslexia: Speech perception and pro-
duktion. Journal of Communication Disorders, 42(3), 180–194.
https://doi.org/10.1016/j.jcomdis.2008.10.004, PubMed:
19100994
Gignac, G. E., & Szodorai, E. T. (2016). Effect size guidelines for
individual differences researchers. Personality and Individual
Unterschiede, 102, 74–78. https://doi.org/10.1016/j.paid.2016.06
.069
Gilmore, J. H., Knickmeyer, R. C., & Gao, W. (2018). Imaging struc-
tural and functional brain development in early childhood.
Nature Reviews Neurowissenschaften, 19(3), 123–137. https://doi.org
/10.1038/nrn.2018.1, PubMed: 29449712
Giraud, A.-L., & Ramus, F. (2013). Neurogenetics and auditory
processing in developmental dyslexia. Current Opinion in Neu-
robiology, 23(1), 37–42. https://doi.org/10.1016/j.conb.2012.09
.003, PubMed: 23040541
Golestani, N., Hervais-Adelman, A., Obleser, J., & Scott, S. K.
(2013). Semantic versus perceptual interactions in neural pro-
cessing of speech-in-noise. NeuroImage, 79, 52–61. https://doi
.org/10.1016/j.neuroimage.2013.04.049, PubMed: 23624171
Golestani, N., Molko, N., Dehaene, S., LeBihan, D., & Pallier, C.
(2007). Brain structure predicts the learning of foreign speech
Geräusche. Hirnrinde, 17(3), 575–582. https://doi.org/10
.1093/cercor/bhk001, PubMed: 16603709
Golestani, N., Price, C. J., & Scott, S. K. (2011). Born with an ear for
dialects? Structural plasticity in the expert phonetician brain.
Zeitschrift für Neurowissenschaften, 31(11), 4213–4220. https://doi.org/10
.1523/JNEUROSCI.3891-10.2011, PubMed: 21411662
Goswami, U.
(2011). A temporal sampling framework for
developmental dyslexia. Trends in den Kognitionswissenschaften, 15(1),
3–10. https://doi.org/10.1016/j.tics.2010.10.001, PubMed:
21093350
Goswami, U. (2022). Language acquisition and speech rhythm pat-
Seeschwalben: An auditory neuroscience perspective. Royal Society Open
Wissenschaft, 9(7), Article 211855. https://doi.org/10.1098/rsos
.211855, PubMed: 35911192
Goswami, U., Huss, M., Mead, N., & Fosker, T. (2021). Auditiv
sensory processing and phonological development in high IQ
and exceptional readers, typically developing readers, and chil-
dren with dyslexia: A longitudinal study. Kinderentwicklung,
92(3), 1083–1098. https://doi.org/10.1111/cdev.13459,
PubMed: 32851656
Grasby, K. L., Jahanshad, N., Painter, J. N., Colodro-Conde, L.,
Bralten, J., Hibar, D. P., Lind, P. A., Pizzagalli, F., Ching,
C. R. K., McMahon, M. A. B., Shatokhina, N., Zsembik, L. C. P.,
Thomopoulos, S. ICH., Zhu, A. H., Strike, L. T., Agartz, ICH., Alhusaini,
S., Almeida, M. A. A., Alnæs, D., … Medland, S. E. (2020). Der
genetic architecture of the human cerebral cortex. Wissenschaft,
367(6484), Article eaay6690. https://doi.org/10.1126/science
.aay6690, PubMed: 32193296
Griffiths, T. D., & Warren, J. D. (2002). The planum temporale as a
computational hub. Trends in den Neurowissenschaften, 25(7), 348–353.
https://doi.org/10.1016/S0166-2236(02)02191-4, PubMed:
12079762
Hakvoort, B., van der Leij, A., Maurits, N., Maassen, B., & Transporter
Zuijen, T. L. (2015). Basic auditory processing is related to
familial risk, not to reading fluency: An ERP study. Kortex, 63,
90–103. https://doi.org/10.1016/j.cortex.2014.08.013, PubMed:
25243992
Hämäläinen, J. A., Salminen, H. K., & Leppänen, P. H. T. (2013).
Basic auditory processing deficits in dyslexia: Systematic reviews
of the behavioral and event-related potential/field evidence. Jour-
nal of Learning Disabilities, 46(5), 413–427. https://doi.org/10
.1177/0022219411436213, PubMed: 22323280
Hart, S. A., Little, C., & van Bergen, E. (2021). Nurture might be
nature: Cautionary tales and proposed solutions. NPJ Science
of Learning, 6(1), Article 2. https://doi.org/10.1038/s41539-020
-00079-z, PubMed: 33420086
Hervais-Adelman, A., Moser-Mercer, B., Murray, M. M., &
Golestani, N. (2017). Cortical thickness increases after simulta-
neous interpretation training. Neuropsychologie, 98, 212–219.
Neurobiology of Language
495
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
https://doi.org/10.1016/j.neuropsychologia.2017.01.008,
PubMed: 28077311
Hickok, G., & Kacke, D. (2007). The cortical organization of
speech processing. Nature Reviews Neurowissenschaften, 8(5),
393–402. https://doi.org/10.1038/nrn2113, PubMed: 17431404
Hoeft, F., & Hancock, R. (2017). Intergenerational transmission of
reading and reading brain networks. In A. Galaburda, N. Gaab,
F. Hoeft, & P. McCardle (Hrsg.), Dyslexia and neuroscience:
The Geschwind-Galaburda hypothesis, 30 years later (1st ed.,
S. 167–176). Paul H. Brookes.
Hugdahl, K., Heiervang, E., Ersland, L., Lundervold, A., Steinmetz, H.,
& Smievoll, A. ICH. (2003). Significant relation between MR measures
of planum temporale area and dichotic processing of syllables in
dyslexic children. Neuropsychologie, 41(6), 666–675. https://doi
.org/10.1016/S0028-3932(02)00224-5, PubMed: 12591024
Hulshoff Pol, H. E., Schnack, H. G., Posthuma, D., Mandl, R. C. W.,
Baaré, W. F., van Oel, C., van Haren, N. E., Collins, D. L., Evans,
A. C., Amunts, K., Bürgel, U., Zilles, K., de Geus, E., Boomsma,
D. ICH., & Kahn, R. S. (2006). Genetic contributions to human brain
morphology and intelligence. Zeitschrift für Neurowissenschaften, 26(40),
10235–10242. https://doi.org/10.1523/ JNEUROSCI.1312-06
.2006, PubMed: 17021179
Jha, S. C., Xia, K., Schmitt, J. E., Ahn, M., Girault, J. B., Murphy,
V. A., Li, G., Wang, L., Shen, D., Zou, F., Zhu, H., Styner, M.,
Knickmeyer, R. C., & Gilmore, J. H. (2018). Genetic influences
on neonatal cortical thickness and surface area. Menschliches Gehirn
Mapping, 39(12), 4998–5013. https://doi.org/10.1002/ hbm
.24340, PubMed: 30144223
Johnson, E. P., Pennington, B. F., Lee, N. R., & Boada, R. (2009).
Directional effects between rapid auditory processing and
phonological awareness in children. Journal of Child Psychology
and Psychiatry, 50(8), 902–910. https://doi.org/10.1111/j.1469
-7610.2009.02064.X, PubMed: 19298469
Kalashnikova, M., Goswami, U., & Burnham, D. (2018). Mothers
speak differently to infants at-risk for dyslexia. Developmental
Wissenschaft, 21(1), Article e12487. https://doi.org/10.1111/desc
.12487, PubMed: 27785865
Khanolainen, D., Salminen, J., Eklund, K., Lerkkanen, M.-K., &
Torppa, M. (2023). Intergenerational transmission of dyslexia:
How do different identification methods of parental difficulties
influence the conclusions regarding children’s risk for dyslexia?
Reading Research Quarterly, 58(2), 220–239. https://doi.org/10
.1002/rrq.482
Kline, R. B. (2011). Relative variances. In Principles and practice of
structural equation modeling (3rd ed., S. 67–68). Guilford.
Kort, W., Schittekatte, M., Dekker, P. H., Verhaeghe, P., Compaan,
E. L., Bosmans, M., & Vermeir, G. (2005). WISC-III NL Wechsler
Intelligence Scale for Children. Handleiding en verantwoording.
Harcourt.
Kraft, ICH., Cafiero, R., Schaadt, G., Brauer, J., Neef, N. E., Müller, B.,
Kirsten, H., Wilcke, A., Boltze, J., Friederici, A. D., & Skeide,
M. A. (2015). Cortical differences in preliterate children at
familiar risk of dyslexia are similar to those observed in dyslexic
readers. Gehirn, 138(9), Article e378. https://doi.org/10.1093
/brain/awv036, PubMed: 25701065
Kuhl, U., Neef, N. E., Kraft, ICH., Schaadt, G., Dörr, L., Brauer, J.,
Czepezauer, ICH., Müller, B., Wilcke, A., Kirsten, H., Emmrich, F.,
Boltze, J., Friederici, A. D., & Skeide, M. A. (2020). The emer-
gence of dyslexia in the developing brain. NeuroImage, 211,
Article 116633. https://doi.org/10.1016/j.neuroimage.2020
.116633, PubMed: 32061802
Kuppen, S., Huss, M., & Goswami, U. (2014). A longitudinal study
of basic auditory processing and phonological skills in children
with low IQ. Applied Psycholinguistics, 35(6), 1109–1141.
https://doi.org/10.1017/S0142716412000719
Landerl, K., Freudenthaler, H. H., Heene, M., de Jong, P. F.,
Desrochers, A., Manolitsis, G., Parrila, R., & Georgiou, G. K.
(2019). Phonological awareness and rapid automatized naming
as longitudinal predictors of reading in five alphabetic orthogra-
phies with varying degrees of consistency. Scientific Studies of
Reading, 23(3), 220–234. https://doi.org/10.1080/10888438
.2018.1510936
Laneau, J., Boets, B., Moonen, M., van Wieringen, A., & Wouters, J.
(2005). A flexible auditory research platform using acoustic or
electric stimuli for adults and young children. Journal of Neuro-
science Methods, 142(1), 131–136. https://doi.org/10.1016/j
.jneumeth.2004.08.015, PubMed: 15652626
Law, J. M., Vandermosten, M., Ghesquière, P., & Wouters, J. (2014).
The relationship of phonological ability, speech perception, Und
auditory perception in adults with dyslexia. Frontiers in Human
Neurowissenschaften, 8, Article 482. https://doi.org/10.3389/fnhum
.2014.00482, PubMed: 25071512
Law, J. M., Vandermosten, M., Ghesquière, P., & Wouters, J. (2017).
Predicting future reading problems based on pre-reading audi-
tory measures: A longitudinal study of children with a familial
risk of dyslexia. Grenzen in der Psychologie, 8, Article 124. https://
doi.org/10.3389/fpsyg.2017.00124, PubMed: 28223953
Lefly, D. L., & Pennington, B. F. (2000). Reliability and validity of
the adult reading history questionnaire. Journal of Learning
D i s a b i l i t i e s , 3 3 ( 3 ) , 2 8 6–2 9 6. h t t p s : / / d o i . o rg/ 1 0 . 11 7 7
/002221940003300306, PubMed: 15505966
Leonard, C. M., Eckert, M. A., Lombardino, L. J., Oakland, T.,
Kranzler, J., Mohr, C. M., King, W. M., & Freeman, A. (2001).
Anatomical risk factors for phonological dyslexia. Zerebral
Kortex, 11(2), 148–157. https://doi.org/10.1093/cercor/11.2
.148, PubMed: 11208669
Leong, V., Hämäläinen, J., Soltész, F., & Goswami, U. (2011). Rise
time perception and detection of syllable stress in adults with
developmental dyslexia. Journal of Memory and Language,
64(1), 59–73. https://doi.org/10.1016/j.jml.2010.09.003
Leppänen, P. H. T., Hämäläinen, J. A., Salminen, H. K., Eklund,
K. M., Guttorm, T. K., Lohvansuu, K., Puolakanaho, A., &
Lyytinen, H. (2010). Newborn brain event-related potentials
revealing atypical processing of sound frequency and the
subsequent association with later literacy skills in children with
familial dyslexia. Kortex, 46(10), 1362–1376. https://doi.org/10
.1016/j.cortex.2010.06.003, PubMed: 20656284
Little, T. D. (2013). Covariate/control variables. In Longitudinal
structural equation modeling (S. 190–198). Guilford.
Long, P., Wan, G., Roberts, M. T., & Corfas, G. (2018). Myelin
Entwicklung, plasticity, and pathology in the auditory system.
Developmental Neurobiology, 78(2), 80–92. https://doi.org/10
.1002/dneu.22538, PubMed: 28925106
Lövdén, M., Wenger, E., Mårtensson, J., Lindenberger, U., &
Backmann, L. (2013). Structural brain plasticity in adult learning
und Entwicklung. Neuroscience and Biobehavioral Reviews,
37(9B), 2296–2310. https://doi.org/10.1016/j.neubiorev.2013
.02.014, PubMed: 23458777
Łuniewska, M., Chyl, K., De¸bska, A., Banaszkiewicz, A.,
Żelechowska, A., Marchewka, A., Grabowska, A., & Jednoróg,
K. (2019). Children with dyslexia and familial risk for dyslexia
present atypical development of the neuronal phonological
Netzwerk. Frontiers in Neuroscience, 13, Article 1287. https://doi
.org/10.3389/fnins.2019.01287, PubMed: 31849595
Ma, Y., Koyama, M. S., Milham, M. P., Castellanos, F. X., Quinn,
B. T., Pardoe, H., Wang, X., Kuzniecky, R., Devinsky, O., Thesen,
Neurobiology of Language
496
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
T., & Blackmon, K. (2015). Cortical thickness abnormalities
associated with dyslexia, independent of remediation status.
NeuroImage: Klinisch, 7, 177–186. https://doi.org/10.1016/j.nicl
.2014.11.005, PubMed: 25610779
Marie, D., & Golestani, N. (2017). Brain structural imaging of
receptive speech and beyond: A review of current methods. Lan-
Spur, Cognition and Neuroscience, 32(7), 870–890. https://doi
.org/10.1080/23273798.2016.1250926
Marie, D., Jobard, G., Crivello, F., Perchey, G., Petit, L., Mellet, E.,
Joliot, M., Zago, L., Mazoyer, B., & Tzourio-Mazoyer, N. (2015).
Descriptive anatomy of Heschl’s gyri in 430 gesunde Freiwillige,
einschließlich 198 left-handers. Brain Structure and Function, 220(2),
729–743. https://doi.org/10.1007/s00429-013-0680-x, PubMed:
24310352
Marie, D., Rutten, S., Marques, J., & Golestani, N. (2018). Auditiv
cortex myelination abnormalities in dyslexia and relationships
with reading performance [Poster presentation]. Alpine Brain
Meeting, Champéry, Schweiz.
Marin-Franch, ICH., Zychaluk, K., & Foster, D. H. (2012). modelfree:
Model-free estimation of a psychometric function. R package
( Version 1.1-1) [Software]. https://cran.r-project.org/package
=modelfree
Mårtensson, J., Eriksson, J., Bodammer, N. C., Lindgren, M.,
Johansson, M., Nyberg, L., & Lövdén, M. (2012). Growth of
language-related brain areas after foreign language learning.
NeuroImage, 63(1), 240–244. https://doi.org/10.1016/j
.neuroimage.2012.06.043, PubMed: 22750568
Mattys, S. L., Davis, M. H., Bradlow, A. R., & Scott, S. K. (2012).
Speech recognition in adverse conditions: Eine Rezension. Language
and Cognitive Processes, 27(7–8), 953–978. https://doi.org/10
.1080/01690965.2012.705006
Müller, G., Lewis, B., Benchek, P., Buss, E., & Calandruccio, L.
(2018). Masked speech recognition and reading ability in
school-age children: Is there a relationship? Journal of Speech,
Language, and Hearing Research, 61(3), 776–788. https://doi
.org/10.1044/2017_JSLHR-H-17-0279, PubMed: 29507949
Miranda, A., & Sousa, N. (2018). Maternal hormonal milieu
influence on fetal brain development. Brain and Behavior, 8(2),
Article e00920. https://doi.org/10.1002/ brb3.920, PubMed:
29484271
Noordenbos, M. W., Segers, E., Serniclaes, W., Mitterer, H., &
Verhoeven, L. (2012). Allophonic mode of speech perception
in Dutch children at risk for dyslexia: A longitudinal study.
Research in Developmental Disabilities, 33(5), 1469–1483.
https://doi.org/10.1016/j.ridd.2012.03.021, PubMed: 22522205
Oldfield, R. C. (1971). Assessment and analysis of handedness: Der
Edinburgh inventory. Neuropsychologie, 9(1), 97–113. https://doi
.org/10.1016/0028-3932(71)90067-4, PubMed: 5146491
Ozernov-Palchik, O., & Gaab, N. (2016). Tackling the “dyslexia
paradox”: Reading brain and behavior for early markers of devel-
opmental dyslexia. Wiley Interdisciplinary Reviews: Kognitiv
Wissenschaft, 7(2), 156–176. https://doi.org/10.1002/wcs.1383,
PubMed: 26836227
Palomar-García, M. Á., Hernández, M., Olcina, G., Adrián-
Ventura, J., Costumero, V., Miró-Padilla, A., Villar-Rodríguez,
E., & Ávila, C. (2020). Auditory and frontal anatomic correlates
of pitch discrimination in musicians, non-musicians, and chil-
dren without musical training. Brain Structure and Function,
225(9), 2735–2744. https://doi.org/10.1007/s00429-020-02151-1,
PubMed: 33029708
Papadopoulos, T. C., Georgiou, G. K., & Parrila, R. K. (2012).
Low-level deficits in beat perception: Neither necessary nor suf-
ficient for explaining developmental dyslexia in a consistent
orthography. Research in Developmental Disabilities, 33(6),
1841–1856. https://doi.org/10.1016/j.ridd.2012.04.009,
PubMed: 22695074
Pardoe, H. R., Kucharsky Hiess, R., & Kuzniecky, R. (2016). Motion
and morphometry in clinical and nonclinical populations. Neu-
roImage, 135, 177–185. https://doi.org/10.1016/j.neuroimage
.2016.05.005, PubMed: 27153982
Penhune, V. B., Zatorre, R. J., MacDonald, J. D., & Evans, A. C.
(1996). Interhemispheric anatomical differences in human primary
auditory cortex: Probabilistic mapping and volume measurement
from magnetic resonance scans. Hirnrinde, 6(5), 661–672.
https://doi.org/10.1093/cercor/6.5.661, PubMed: 8921202
Peterson, R., Arnett, A. B., Pennington, B. F., Byrne, B., Samuelsson,
S., & Olson, R. K. (2018). Literacy acquisition influences chil-
dren’s rapid automatized naming. Developmental Science,
21(3), Article e12589. https://doi.org/10.1111/desc.12589,
PubMed: 28812316
Plakas, A., van Zuijen, T. L., van Leeuwen, T., Thomson, J. M., &
van der Leij, A. (2013). Impaired non-speech auditory processing
at a pre-reading age is a risk-factor for dyslexia but not a predic-
tor: An ERP study. Kortex, 49(4), 1034–1045. https://doi.org/10
.1016/j.cortex.2012.02.013, PubMed: 22542727
Poelmans, H., Luts, H., Vandermosten, M., Boets, B., Ghesquière,
P., & Wouters, J. (2011). Reduced sensitivity to slow-rate dynamic
auditory information in children with dyslexia. Research in
Developmental Disabilities, 32(6), 2810–2819. https://doi.org
/10.1016/j.ridd.2011.05.025, PubMed: 21645986
Quinn, J. M., Spencer, M., & Wagner, R. K. (2015). Individual dif-
ferences in phonological awareness and their role in learning to
read. In P. Afflerbach (Ed.), Handbook of individual differences in
reading (S. 80–92). Routledge.
R Core Team. (2021). R: A language and environment for statistical
computing. R Foundation for Statistical Computing. https://www
.r-project.org/
Ramus, F., Altarelli, ICH., Jednoróg, K., Zhao, J., & Scotto di Covella, L.
(2018). Neuroanatomy of developmental dyslexia: Pitfalls and
promise. Neuroscience and Biobehavioral Reviews, 84,
434–452. https://doi.org/10.1016/j.neubiorev.2017.08.001,
PubMed: 28797557
Ramus, F., Rosen, S., Dakin, S. C., Day, B. L., Castellote, J. M.,
White, S., & Frith, U. (2003). Theories of developmental dys-
lexia: Insights from a multiple case study of dyslexic adults.
Gehirn, 126(4), 841–865. https://doi.org/10.1093/brain/awg076,
PubMed: 12615643
Ressel, V., Pallier, C., Ventura-Campos, N., Díaz, B., Roessler, A.,
Ávila, C., & Sebastián-Gallés, N.
(2012). An effect of bilin-
gualism on the auditory cortex. Zeitschrift für Neurowissenschaften, 32(47),
16597–16601. https://doi.org/10.1523/ JNEUROSCI.1996-12
.2012, PubMed: 23175815
Richardson, U., Leppänen, P. H. T., Leiwo, M., & Lyytinen, H.
(2003). Speech perception of infants with high familial risk for
dyslexia differ at the age of 6 months. Developmental Neuro-
psy cho lo gy, 23( 3 ) , 3 8 5–397 . https://doi.org/10.1207
/S15326942DN2303_5, PubMed: 12740192
Richardson, U., Thomson, J. M., Scott, S. K., & Goswami, U.
(2004). Auditory processing skills and phonological representa-
tion in dyslexic children. Dyslexia, 10(3) 215–233. https://doi
.org/10.1002/dys.276, PubMed: 15341199
Robitzsch, A. (2020). Why ordinal variables can (almost) always be
treated as continuous variables: Clarifying assumptions of robust
continuous and ordinal factor analysis estimation methods. Fron-
tiers in Education, 5, Article 589965. https://doi.org/10.3389
/feduc.2020.589965
Neurobiology of Language
497
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
Ronan, L., Voets, N., Rua, C., Alexander-Bloch, A., Hough, M.,
Mackay, C., Crow, T. J., James, A., Giedd, J. N., & Fletcher,
P. C. (2014). Differential tangential expansion as a mechanism
for cortical gyrification. Hirnrinde, 24(8), 2219–2228.
https://doi.org/10.1093/cercor/bht082, PubMed: 23542881
Rosen, S. (2003). Auditory processing in dyslexia and specific lan-
guage impairment: Is there a deficit? What is its nature? Does it
explain anything? Journal of Phonetics, 31(3–4), 509–527.
https://doi.org/10.1016/S0095-4470(03)00046-9
Rosseel, Y. (2012). lavaan: An R package for structural equation
modeling. Journal of Statistical Software, 48(2), 1–36. https://doi
.org/10.18637/jss.v048.i02
Rutten, S., Santoro, R., Hervais-Adelman, A., Formisano, E., &
Golestani, N. (2019). Cortical encoding of speech enhances
task-relevant acoustic information. Natur menschliches Verhalten,
3(9), 974–987. https://doi.org/10.1038/s41562-019-0648-9,
PubMed: 31285622
Sanchez Bloom, J., Garcia-Barrera, M. A., Müller, C. J., Müller, S. R.,
& Hynd, G. W. (2013). Planum temporale morphology in chil-
dren with developmental dyslexia. Neuropsychologie, 51(9),
1684–1692. https://doi.org/10.1016/j.neuropsychologia.2013.05
.012, PubMed: 23707683
Schaer, M., Cuadra, M. B., Tamarit, L., Lazeyras, F., Eliez, S., &
Thiran, J.-P. (2008). A surface-based approach to quantify local
cortical gyrification. IEEE Transactions on Medical Imaging,
27(2), 161–170. https://doi.org/10.1109/ TMI.2007.903576,
PubMed: 18334438
Schneider, P., Andermann, M., Wengenroth, M., Goebel, R., Flor,
H., Rupp, A., & Diesch, E. (2009). Reduced volume of Heschl’s
gyrus in tinnitus. NeuroImage, 45(3), 927–939. https://doi.org/10
.1016/j.neuroimage.2008.12.045, PubMed: 19168138
Schneider, P., Scherg, M., Dosch, H. G., Specht, H. J., Gutschalk,
A., & Rupp, A. (2002). Morphology of Heschl’s gyrus reflects
enhanced activation in the auditory cortex of musicians. Natur
Neurowissenschaften, 5(7), 688–694. https://doi.org/10.1038/nn871,
PubMed: 12068300
Schneider, P., Sluming, V., Roberts, N., Scherg, M., Goebel, R., Specht,
H. J., Dosch, H. G., Bleeck, S., Stippich, C., & Rupp, A. (2005).
Structural and functional asymmetry of lateral Heschl’s gyrus
reflects pitch perception preference. Naturneurowissenschaften, 8(9),
1241–1247. https://doi.org/10.1038/nn1530, PubMed: 16116442
Seither-Preisler, A., Parncutt, R., & Schneider, P. (2014). Size and
synchronization of auditory cortex promotes musical, literacy,
and attentional skills in children. Zeitschrift für Neurowissenschaften,
34(33), 10937–10949. https://doi.org/10.1523/ JNEUROSCI
.5315-13.2014, PubMed: 25122894
Serrallach, B., Groß, C., Bernhofs, V., Engelmann, D., Benner, J.,
Gündert, N., Blatow, M., Wengenroth, M., Seitz, A., Brunner,
M., Seither, S., Parncutt, R., Schneider, P., & Seither-Preisler, A.
(2016). Neural biomarkers for dyslexia, ADHD, and ADD in the
auditory cortex of children. Frontiers in Neuroscience, 10, Article
324. https://doi.org/10.3389/fnins.2016.00324, PubMed:
27471442
Shapleske, J., Rossell, S. L., Woodruff, P. W. R., & David, A. S.
(1999). The planum temporale: A systematic, quantitative review
of its structural, functional and clinical significance. Gehirn
Research Reviews, 29(1), 26–49. https://doi.org/10.1016/S0165
-0173(98)00047-2, PubMed: 9974150
Skeide, M. A., Bazin, P. L., Trampel, R., Schäfer, A., Männel, C., von
Kriegstein, K., & Friederici, A. D. (2018). Hypermyelination of the
left auditory cortex in developmental dyslexia. Neurologie, 90(6),
e492–e497. https://doi.org/10.1212/ WNL.0000000000004931,
PubMed: 29321232
Schmied, S. D. (2011). Approach to epigenetic analysis in language
disorders. Journal of Neurodevelopmental Disorders, 3(4),
356–364. https://doi.org/10.1007/s11689-011-9099-y, PubMed:
22113455
Smits, C., & Houtgast, T. (2005). Results from the Dutch
speech-in-noise screening test by telephone. Ear and Hearing,
26(1), 89–95. https://doi.org/10.1097/00003446-200502000
-00008, PubMed: 15692307
Snowling, M., Gallagher, A., & Frith, U. (2003). Family risk of dys-
lexia is continuous: Individual differences in the precursors of
reading skill. Kinderentwicklung, 74(2), 358–373. https://doi
.org/10.1111/1467-8624.7402003, PubMed: 12705560
Snowling, M., & Melby-Lervåg, M. (2016). Oral language deficits in
familial dyslexia: A meta-analysis and review. Psychological
Bulletin, 142(5), 498–545. https://doi.org/10.1037/bul0000037,
PubMed: 26727308
Steinmetz, H., Rademacher, J., Jäncke, L., Huang, Y., Thron, A., &
Zilles, K. (1990). Total surface of temporoparietal intrasylvian
Kortex: Diverging left-right asymmetries. Brain and Language,
39(3), 357–372. https://doi.org/10.1016/0093-934X(90)90145
-7, PubMed: 2285859
Strike, L. T., Hansell, N. K., Couvy-Duchesne, B., Thompson, P. M.,
de Zubicaray, G. ICH., McMahon, K. L., & Wright, M. J. (2019).
Genetic complexity of cortical structure: Differences in genetic
and environmental factors influencing cortical surface area and
Dicke. Hirnrinde, 29(3), 952–962. https://doi.org/10
.1093/cercor/bhy002, PubMed: 29377989
Sutherland, M. E., Zatorre, R. J., Watkins, K. E., Hervé, P.-Y.,
Leonard, G., Pike, B. G., Witton, C., & Paus, T. (2012). Anatomical
correlates of dynamic auditory processing: Relationship to literacy
during early adolescence. NeuroImage, 60(2), 1287–1295.
https://doi.org/10.1016/j.neuroimage.2012.01.051, PubMed:
22266413
Swagerman, S. C., van Bergen, E., Dolan, C., de Geus, E. J. C.,
Koenis, M. M. G., Hulshoff Pol, H. E., & Boomsma, D. ICH.
(2017). Genetic transmission of reading ability. Brain and
Language, 172, 3–8. https://doi.org/10.1016/j.bandl.2015.07
.008, PubMed: 26300341
Theys, C., Wouters, J., & Ghesquière, P. (2014). Diffusion tensor
imaging and resting-state functional MRI-scanning in 5- Und
6-year-old children: Training protocol and motion assessment.
PLOS ONE, 9(4), Article e94019. https://doi.org/10.1371
/zeitschrift.pone.0094019, PubMed: 24718364
Turesky, T. K., Vanderauwera, J., & Gaab, N. (2021). Imaging the
rapidly developing brain: Current challenges for MRI studies in
the first five years of life. Developmental Cognitive Neuroscience,
47, Article 100893. https://doi.org/10.1016/j.dcn.2020.100893,
PubMed: 33341534
Turker, S., & Reiterer, S. M. (2021). Gehirn, musicality, and language
aptitude: A complex interplay. Annual Review of Applied Linguis-
Tics, 41, 95–107. https://doi.org/10.1017/S0267190520000148
Turker, S., Reiterer, S. M., Schneider, P., & Seither-Preisler, A.
(2019). Auditory cortex morphology predicts language learning
potential in children and teenagers. Frontiers in Neuroscience,
13, Article 824. https://doi.org/10.3389/fnins.2019.00824,
PubMed: 31447639
Turker, S., Reiterer, S. M., Seither-Preisler, A., & Schneider, P.
(2017). “When music speaks”: Auditory cortex morphology as
a neuroanatomical marker of language aptitude and musicality.
Grenzen in der Psychologie, 8, Article 2096. https://doi.org/10.3389
/fpsyg.2017.02096, PubMed: 29250017
Turker, S., Seither-Preisler, A., & Reiterer, S. M. (2021). Examining
individual differences in language learning: A neurocognitive
Neurobiology of Language
498
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
model of language aptitude. Neurobiology of Language, 2(3),
389–415. https://doi.org/10.1162/nol_a_00042, PubMed:
37213255
Tzourio-Mazoyer, N., & Mazoyer, B. (2017). Variations of planum
temporale asymmetries with Heschl’s gyri duplications and asso-
ciation with cognitive abilities: MRI investigation of 428 healthy
volunteers. Brain Structure and Function, 222(6), 2711–2726.
https://doi.org/10.1007/s00429-017-1367-5 , PubMed:
28164245
van Atteveldt, N., Vandermosten, M., Weeda, W., & Bonte, M.
(2021). How to capture developmental brain dynamics: Gaps
and solutions. NPJ Science of Learning, 6(1), Article 10. https://
doi.org/10.1038/s41539-021-00088-6, PubMed: 33941785
van Bergen, E., Bishop, D., van Zuijen, T. L., & de Jong, P. F. (2015).
How does parental reading influence children’s reading? A study
of cognitive mediation. Scientific Studies of Reading, 19(5),
325–339. https://doi.org/10.1080/10888438.2015.1050103
van Bergen, E., van der Leij, A., & de Jong, P. F. (2014). The inter-
generational multiple deficit model and the case of dyslexia.
Grenzen der menschlichen Neurowissenschaften, 8, Article 346. https://doi.org
/10.3389/fnhum.2014.00346, PubMed: 24920944
Van den Borre, E., Denys, S., van Wieringen, A., & Wouters, J.
(2021). The digit triplet test: A scoping review. International Jour-
nal of Audiology, 60(12), 946–963. https://doi.org/10.1080
/14992027.2021.1902579, PubMed: 33840339
Vanderauwera, J., Wouters, J., Vandermosten, M., & Ghesquière, P.
(2017). Early dynamics of white matter deficits in children
developing dyslexia. Developmental Cognitive Neuroscience,
27, 69–77. https://doi.org/10.1016/j.dcn.2017.08.003, PubMed:
28823983
Vanderauwera, J., Altarelli, ICH., Vandermosten, M., De Vos, A.,
Wouters, J., & Ghesquière, P. (2018). Atypical structural asymme-
try of the planum temporale is related to family history of dys-
lexia. Hirnrinde, 28(1), 63–72. https://doi.org/10.1093
/cercor/bhw348, PubMed: 29253247
van der Leij, A., van Bergen, E., van Zuijen, T. L., de Jong, P.,
Maurits, N., & Maassen, B. (2013). Precursors of developmental
dyslexia: An overview of the longitudinal Dutch Dyslexia Pro-
gramme study. Dyslexia, 19(4), 191–213. https://doi.org/10
.1002/dys.1463, PubMed: 24133035
Vandermosten, M., Correia, J., Vanderauwera, J., Wouters, J.,
Ghesquière, P., & Bonte, M. (2020). Brain activity patterns
of phonemic representations are atypical in beginning readers
with family risk for dyslexia. Developmental Science, 23(1),
Article e12857. https://doi.org/10.1111/desc.12857, PubMed:
31090993
Vandermosten, M., Cuynen, L., Vanderauwera, J., Wouters, J., &
Ghesquière, P. (2017). White matter pathways mediate parental
effects on children’s reading precursors. Brain and Language,
173, 10–19. https://doi.org/10.1016/j.bandl.2017.05.002,
PubMed: 28558269
Vandermosten, M., Schevenels, K., Economou, M., & Hoeft, F.
(2020). The influence of intergenerational transfer of white matter
tracts on early reading development. bioRxiv. https://doi.org/10
.1101/2020.10.09.333096
Vandewalle, E., Boets, B., Ghesquière, P., & Zink, ICH. (2012).
Auditory processing and speech perception in children with
specific language impairment: Relations with oral language and
literacy skills. Research in Developmental Disabilities, 33(2),
635–644. https://doi.org/10.1016/j.ridd.2011.11.005, PubMed:
22155538
Van Hirtum, T., Ghesquière, P., & Wouters, J. (2019). Atypical neu-
ral processing of rise time by adults with dyslexia. Kortex, 113,
128–140. https://doi.org/10.1016/j.cortex.2018.12.006,
PubMed: 30640141
Vanvooren, S., Poelmans, H., De Vos, A., Ghesquière, P., &
Wouters, J. (2017). Do prereaders’ auditory processing and
speech perception predict later literacy? Research in Develop-
mental Disabilities, 70, 138–151. https://doi.org/10.1016/j.ridd
.2017.09.005, PubMed: 28938227
van Wieringen, A., & Wouters, J. (2023). Lilliput: Speech percep-
tion in speech-weighted noise and in quiet in young children.
International Journal of Audiology, 62(8), 747–755. https://doi
.org/10.1080/14992027.2022.2086491, PubMed: 35732012
van Zuijen, T. L., Plakas, A., Maassen, B. A. M, Been, P., Maurits,
N. M., Krikhaar, E., van Driel, J., & van der Leij, A. (2012).
Temporal auditory processing at 17 months of age is associated
with preliterate language comprehension and later word reading
fluency: An ERP study. Neurowissenschaftliche Briefe, 528(1), 31–35.
https://doi.org/10.1016/j.neulet.2012.08.058, PubMed:
22981882
Venables, W. N., & Ripley, B. D. (2002). Modern applied statistics
with S (4th ed.). Springer. https://doi.org/10.1007/978-0-387
-21706-2
Verwimp, C., Vanden Bempt, F., Kellens, S., Economou, M.,
Vandermosten, M., Wouters, J., Ghesquière, P., & Vanderauwera,
J. (2020). Pre-literacy heterogeneity in Dutch-speaking kinder-
gartners: Latent profile analysis. Annals of Dyslexia, 70(3),
275–294. https://doi.org/10.1007/s11881-020-00207-9,
PubMed: 33074483
Von Economo, C., & Horn, L. (1930). Uber windungsrelief mabe
und Rindenarchitektonic der supratemparalflache, ihre indivi-
duellen und seitenunterschiede. Zeitschrift für die gesamte Neu-
rologie und Psychiatrie, 130, 678–757. https://doi.org/10.1007
/BF02865945
Warrier, C., Wong, P., Penhune, V., Zatorre, R., Parrish, T., Abrams,
D., & Kraus, N. (2009). Relating structure to function: Heschl’s
gyrus and acoustic processing. Zeitschrift für Neurowissenschaften, 29(1),
61–69. https://doi.org/10.1523/ JNEUROSCI.3489-08.2009,
PubMed: 19129385
Wengenroth, M., Blatow, M., Heinecke, A., Reinhardt, J., Stippich,
C., Hofmann, E., & Schneider, P. (2014). Increased volume and
function of right auditory cortex as a marker for absolute pitch.
Hirnrinde, 24(5), 1127–1137. https://doi.org/10.1093
/cercor/bhs391, PubMed: 23302811
White, T., Jansen, P. R., Muetzel, R. L., Sudre, G., El Marroun, H.,
Tiemeier, H., Qiu, A., Shaw, P., Michael, A. M., & Verhulst, F. C.
(2018). Automated quality assessment of structural magnetic res-
onance images in children: Comparison with visual inspection
and surface-based reconstruction. Kartierung des menschlichen Gehirns,
39(3), 1218–1231. https://doi.org/10.1002/ hbm.23911,
PubMed: 29206318
White-Schwoch, T., Woodruff Carr, K., Thompson, E. C., Anderson,
S., Nicol, T., Bradlow, A. R., Zecker, S. G., & Kraus, N. (2015).
Auditory processing in noise: A preschool biomarker for literacy.
PLOS Biology, 13(7), Article e1002196. https://doi.org/10.1371
/journal.pbio.1002196, PubMed: 26172057
Winkler, A. M., Kochunov, P., Blangero, J., Almasy, L., Zilles, K.,
Fuchs, P. T., Duggirala, R., & Glahn, D. C. (2010). Cortical
thickness or grey matter volume? The importance of selecting
the phenotype for imaging genetics studies. NeuroImage, 53(3),
1135–1146. https://doi.org/10.1016/j.neuroimage.2009.12.028,
PubMed: 20006715
Wong, P. C. M., Warrier, C. M., Penhune, V. B., Roy, A. K., Sadehh,
A., Parrish, T. B., & Zatorre, R. J. (2008). Volume of left Heschl’s
gyrus and linguistic pitch learning. Hirnrinde, 18(4),
Neurobiology of Language
499
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Linking pre-reading auditory cortex with reading and family risk
828–836. https://doi.org/10.1093/cercor/ bhm115, PubMed:
17652466
Xia, Z., Hoeft, F., Zhang, L., & Shu, H. (2016). Neuroanatomical anom-
alies of dyslexia: Disambiguating the effects of disorder, perfor-
Mance, and maturation. Neuropsychologie, 81, 68–78. https://doi
.org/10.1016/j.neuropsychologia.2015.12.003, PubMed: 26679527
Ziegler, J. C., Pech-Georgel, C., George, F., & Lorenzi, C. (2009).
Speech-perception-in-noise deficits in dyslexia. Developmental
Wissenschaft, 12(5), 732–745. https://doi.org/10.1111/j.1467-7687
.2009.00817.X, PubMed: 19702766
Zoellner, S., Benner, J., Zeidler, B., Seither-Preisler, A., Christiner,
M., Seitz, A., Goebel, R., Heinecke, A., Wengenroth, M., Blatow,
M., & Schneider, P. (2019). Reduced cortical thickness in
Heschl’s gyrus as an in vivo marker for human primary auditory
Kortex. Kartierung des menschlichen Gehirns, 40(4), 1139–1154. https://doi.org
/10.1002/hbm.24434, PubMed: 30367737
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
3
4
7
4
2
1
5
6
6
0
3
N
Ö
_
A
_
0
0
1
1
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
9
S
e
P
e
M
B
e
R
2
0
2
3
Neurobiology of Language
500