RESEARCH ARTICLE
Spatiotemporal Dynamics of Activation in Motor
and Language Areas Suggest a Compensatory
Role of the Motor Cortex in Second
Language Processing
Lili Tian1,2,3,4
, Hongjun Chen3
, Pyry Petteri Heikkinen5,6
,
Wenya Liu7
, and Tiina Parviainen1,2
1Abteilung für Psychologie, University of Jyväskylä, Jyväskylä, Finland
2Centre for Interdisciplinary Brain Research, University of Jyväskylä, Jyväskylä, Finland
3School of Foreign Languages, Dalian University of Technology, Dalian, China
4Language and Brain Research Centre, Sichuan International Studies University, Chongqing, China
5Department of Psychology and Logopedics, Faculty of Medicine, University of Helsinki, Helsinki, Finland
6Faculty of Educational Sciences, University of Helsinki, Helsinki, Finland
7Faculty of Information Technology, University of Jyväskylä, Jyväskylä, Finland
Schlüsselwörter: motor cortex involvement, magnetoencephalography, native language, zweite
Sprache, language proficiency, abstractness
ABSTRAKT
The involvement of the motor cortex in language understanding has been intensively
discussed in the framework of embodied cognition. Although some studies have provided
evidence for the involvement of the motor cortex in different receptive language tasks, the role
that it plays in language perception and understanding is still unclear. In der vorliegenden Studie, Wir
explored the degree of involvement of language and motor areas in a visually presented
sentence comprehension task, modulated by language proficiency (L1: native language,
L2: second language) and linguistic abstractness (literal, metaphorical, and abstract).
Magnetoencephalography data were recorded from 26 late Chinese learners of English. A
cluster-based permutation F test was performed on the amplitude of the source waveform
for each motor and language region of interest (ROI). Results showed a significant effect
of language proficiency in both language and motor ROIs, manifested as overall greater
involvement of language ROIs (short insular gyri and planum polare of the superior temporal
gyrus) in the L1 than the L2 during 300–500 ms, and overall greater involvement of motor ROI
(central sulcus) in the L2 than the L1 during 600–800 ms. We interpreted the over-recruitment
of the motor area in the L2 as a higher demand for cognitive resources to compensate for
the inadequate engagement of the language network. Allgemein, our results indicate a
compensatory role of the motor cortex in L2 understanding.
EINFÜHRUNG
The engagement of the motor cortex in language processing has been intensively discussed
within the framework of embodied cognition. Based on the embodied view, language process-
ing, specifically semantic processing (d.h., processing of meaning), involves not only classic
language-related regions but also the motor system to simulate the perceptual meaning con-
veyed by words (Barsalou et al., 2008; Fischer & Zwaan, 2008; Walisisch & Lakoff, 2005;
Keine offenen Zugänge
Tagebuch
Zitat: Tian, L., Chen, H., Heikkinen,
P. P., Liu, W., & Parviainen, T. (2023).
Spatiotemporal dynamics of activation
in motor and language areas suggest a
compensatory role of the motor cortex
in second language processing.
Neurobiology of Language, 4(1),
178–197. https://doi.org/10.1162
/nol_a_00093
DOI:
https://doi.org/10.1162/nol_a_00093
Erhalten: 15 Mai 2022
Akzeptiert: 14 November 2022
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Corresponding Authors:
Tiina Parviainen
tiina.m.parviainen@jyu.fi
Hongjun Chen
chenhj@dlut.edu.cn
Handling-Editor:
Kate Watkins
Urheberrechte ©: © 2023
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Compensatory role of the motor cortex in second language processing
Pulvermüller & Ermüdung, 2010; Zwaan, 2014). The embodied view of semantic processing has
been supported by neuroimaging and electrophysiological studies during the past decade,
showing neural activations and oscillations in the motor cortex during meaning understanding
(Fargier et al., 2012; Fernandino et al., 2013; Klepp et al., 2014, 2015; Mollo et al., 2016;
Moreno et al., 2013). Zusätzlich, the action-sentence compatibility effect (Glenberg &
Kaschak, 2002) has been taken as evidence for the involvement of the motor system in
action-related semantic processing. Faster response was found when the direction of move-
ment is congruent with the direction conveyed by the sentence (Glenberg et al., 2008;
Kaschak & Borreggine, 2008; Santana & de Vega, 2011; Zwaan & Taylor, 2006). Jedoch,
some recent studies failed to replicate any such motor compatibility effect (Greco, 2021;
Morey et al., 2022; Papesh, 2015).
Clinical studies have provided more direct evidence for the involvement of the motor cor-
tex in semantic processing by investigating patients with motor impairment (z.B., Parkinson’s
Krankheit, or PD; Buccino et al., 2018; Cardona et al. 2014; Desai et al., 2015; Fernandino et al.,
2013; Kargieman et al., 2014; Kemmerer et al., 2012; Monaco et al., 2019). These studies
showed that the motor-impaired participants had a selective difficulty in comprehending the
action-related words (z.B., kick), manifested as a lower accuracy rate, longer response time,
and an absence or attenuation of modulation of motor responses in patients with PD, com-
pared with the healthy control group. The revealed association of impaired motor skills and
deficits in understanding action-related meaning would support the embodied account of
semantic processing. Jedoch, some other lesion studies failed to find the causal effect of
motor cortex impairment on the processing of action-related meaning (Maieron et al., 2013;
Papeo et al., 2010). These studies showing the dissociation of motor impairment and motoric
semantic processing question the necessity of the motor cortex in language processing.
The emerging controversial findings have stirred up critiques and reflections on the
embodied assumptions of language processing. As has been pointed out, the rapidly growing
popularity of the embodied account is likely to result in the ignorance of other potential inter-
pretations (sehen, z.B., Chatterjee, 2010; Hauk & Tschentscher, 2013; Mahon, 2015; Mahon &
Hickok, 2016). Studies with the embodied hypothetical stance tended to interpret the data
within the theoretical framework of embodiment with a prior hypothetical bias. Zum Beispiel,
results showing motor activation in language tasks have been monotonically interpreted as the
result of mental simulation of motor-related meaning, and therefore taken as an additional
piece of evidence to confirm the embodied assumption. Jedoch, activation of the motor
cortex may not necessarily be due to the mental simulation of motoric meaning. It can be
ubiquitous in the language processing in general (Meteyard et al., 2012; Tian et al., 2020)
or related to other aspects beyond strict linguistic processing (Maieron et al., 2013).
Functional and Epiphenomenal Role
The emerging controversial findings impelled researchers to re-examine the role of the motor
cortex in language processing and test whether activations in the motor cortex reflect the
retrieval of lexical-semantic information (functional role) or arise as a byproduct of post-
semantic motor imagery (epiphenomenal role). Some studies attempted to disentangle the
functional and epiphenomenal role by scrutinizing the temporal information of motor activa-
tionen (García et al., 2019; van Elk et al., 2010). In van Elk et al.’s (2010) Studie, an early acti-
vation of the motor area indexed by the mu rhythm event-related desynchronization (ERD) War
found preceding semantic processing (around 400 ms after onset) and sustaining in parallel
with semantic processing (around 700 ms after onset). Based on the early latency of motor
Neurobiology of Language
179
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Compensatory role of the motor cortex in second language processing
Aktivierung, it was concluded that motor activation primarily reflected lexical-semantic retrieval
and integration rather than post-lexical motor imagery.
Compared with neurotypical studies, lesion (pathological and virtual transient dysfunctions
caused by repetitive transcranial magnetic stimulation [rTMS]) studies offered a more direct
pathway for scrutinizing the causal role of the motor cortex, since researchers were able to
detect the causality by manipulating stimulations over the motor cortex (Bocanegra et al.,
2017; Desai et al., 2015; Fernandino et al., 2013; Pulvermüller et al., 2005; Reilly et al.,
2019; Vukovic et al., 2017). In Vukovic et al.’s (2017) Studie, rTMS was employed over the left
motor cortex within 200 ms of word onset to examine whether the stimulation would affect the
processing of hand-related action words and abstract words in the lexical decision
task—which requires very shallow lexical-semantic processing—and semantic judgment
task—which requires explicit access to action-related meaning processing. The stimulation
impaired the comprehension of the action words but facilitated that of the abstract words,
compared with the performance in the lexical decision task. The interruptive effect of stimu-
lation on lexical-semantic processing suggested a functional role of the motor cortex in
semantic processing. Consistent results were also reported among studies concerning motor
disorders, where associations were found between the impairment in action performance
and the impairment in action-verb processing (Bocanegra et al., 2017; Desai et al., 2015;
Fernandino et al., 2013).
Umgekehrt, some studies reported dissociations between motor impairment and action
semantic deficits (Maieron et al., 2013; Papeo et al., 2010). In Maieron et al.’s (2013) Studie,
functional magnetic resonance imaging (fMRT) was employed to examine functional connec-
tivity between the language network and primary motor cortex (M1) in an action-verb naming
Aufgabe. Participants were patients whose lesions involved (or spared) the M1 and healthy con-
trols. It was found that lesions in the M1 did not degrade the performance of the action-verb
naming task compared with the healthy controls. Results of the functional connectivity further
revealed a lack of task-modulated connectivity between the M1 and language network in the
action-verb naming task for both lesion and healthy groups. These findings indicated an acces-
sory rather than functional role of the motor cortex in the processing of action words.
Gradations of Motor Cortex Involvement
Instead of confirming or refuting the embodied hypothesis, some studies turned to explore the
degree of motor cortex involvement, such as whether the motor cortex was differentially
involved in different language settings. As highlighted by Chatterjee (2010) and Meteyard
et al. (2012), the discussion of the graded nature of embodiment would shed light on the role
that the motor system plays in semantic processing.
The gradation of motor cortex involvement has been mostly explored from the perspective
of language proficiency (L1: native language; L2: second language) (Birba et al., 2020; Von
Grauwe et al., 2014; Monaco et al., 2021; Tian et al., 2020; Vukovic & Shtyrov, 2014; Zhang
et al., 2020). By employing a passive reading task involving action-related words, Vukovic and
Shtyrov (2014) found that the engagement of the motor cortex was greater for L1 than L2 for
German-English speakers, indexed by a stronger ERD for the L1 than the L2 at around 8–12 Hz
(mu rhythm). The stronger ERD for the L1 was interpreted as the result of a more integrated
perception-action circuit for the L1 lexical-semantic representation. Im Gegensatz, in our earlier
fMRT-Studie (Tian et al., 2020), stronger activation of the motor cortex was found for the L2 than
the L1, which was interpreted as the consequence of higher demand for cognitive resources as
compensation for a less proficient language. Ähnlich, Monaco et al. (2021) also reported
Neurobiology of Language
180
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Compensatory role of the motor cortex in second language processing
greater motor excitability for the L2 (English) than the L1 (French) in an action-related semantic
judgment task, indexed by a higher motor evoked potentials for the L2 when the TMS was given
275 ms after word onset. Jedoch, the authors only claimed a different degree of motor cortex
involvement between L1 and L2 semantic processing without further interpreting the implica-
tions underlying such differences. Andererseits, a similar degree of motor cortex activation
has been reported (De Grauwe et al., 2014) between the L1 (Dutch native speakers) and the L2
(German advanced learners of Dutch) groups in performing a lexical decision task involving
cognates and non-cognates with motor or non-motor-related meanings. The study therefore con-
cluded that the lexical-semantic representation of the L2 was adequate to induce a similar
degree of motor activation relative to the L1.
In addition to language proficiency, the gradation of motor cortex involvement has also been
explored by manipulating the level of linguistic abstractness (z.B., literal/metaphorical/abstract
Sprache; Desai et al., 2013; Schaller et al., 2017; Tian et al., 2020). In Desai et al.’s (2013)
Studie, four levels of linguistic abstractness were manipulated at sentence level, including literal
Aktion, metaphorical action, idiomatic action, and abstract verb. The blood oxygen level
dependent signals of fMRI showed attenuated activation in the motor regions with the increase
of linguistic abstractness (literal > metaphor > idiom > abstract). In our earlier study (Tian et al.,
2020), we reported a similar decremental trend of motor activation with a hierarchically
decreasing pattern of motor cortex activation from the literal to the abstract verb phrases.
The Present Study
Previous studies have advanced our understanding of the motor system in semantic processing
by exploring the gradations of motor cortex involvement in different linguistic circumstances.
Jedoch, the discussed studies using fMRI, electroencephalogram (EEG), or TMS lacked either
temporal or spatial accuracy in describing brain activation. Combining spatial and temporal
resolution is crucial for the comprehensive understanding of how (and when) the motor cortex
contributes to language understanding since timing and source dynamics of brain activation
needs to be extracted simultaneously from language and motor areas. Majority of previous
studies only focused on the motor regions of interest (ROIs), while ignoring the simultaneous
neural activities of the language regions. In der vorliegenden Studie, we employed magnetoenceph-
alography (MEG) with millisecond temporal resolution and sub-centimeter spatial resolution to
explore the temporal activation dynamics of motor and language areas in semantic processing.
Speziell, we aim to investigate whether the degree of the engagement of motor and
language areas is modulated by language proficiency (native language and second language)
and linguistic abstractness (literal, metaphorical, and abstract).
METHODEN
Teilnehmer
A total of 26 Teilnehmer (8 männlich, 18 weiblich) were recruited from the University of Jyväskylä,
Finland. Participants were Chinese-English speakers, who started to learn English at the mean
age of 9.77 (SD = 2.73) and had an average of 16.38 years’ (SD = 4.67) experience in learning
English. Participants had the Lextale vocabulary test (www.lextale.com; Lemhöfer & Broersma,
2012) to measure their L2 vocabulary knowledge (Mittelwert ± SD: 74.18 ± 8.35). All participants
were right-handed with normal or corrected-to-normal vision. None of the participants
reported having any history of neurological disorder. Participants gave informed consent prior
to participation. Participants were compensated for their participation in the experiment. Der
study was approved by the ethics committee of the University of Jyväskylä. Two participants
Neurobiology of Language
181
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Compensatory role of the motor cortex in second language processing
were excluded from data analysis due to the low accuracy rate in behavioral performance
(below 75%, mean = 93.04%, SD = 6%), ergebend 24 participants in the final analysis.
Experiment Design
To examine the effect of language proficiency and linguistic abstractness on the degree of
motor cortex involvement, L1 and L2 experiments were designed. Within each experiment,
the factor of linguistic abstractness was manipulated with a gradual increase of abstractness
from literal to metaphorical to abstract conditions. Each trial consisted of two verb phrases,
with the second verb phrase either semantically congruent or incongruent with the first one.
Participants were required to perform a semantic judgment task, where they needed to judge
whether the second verb phrase shared the same meaning as the first phrase by pressing the
response buttons.
Stimuli
A total of 180 verb phrases (60 in each condition) were used in both L1 and L2 experiment. Der
literal and metaphorical phrases contained an action-related verb, either hand or arm related.
The abstract phrase connoted the same meaning expressed by the metaphorical one (Tisch 1).
Phrases in the L1 experiment were semantically equivalent to those in the L2 experiment, mit
few exceptions in the metaphorical condition, due to the lack of Chinese equivalents of some
English metaphorical expressions. The verb phrases in both L1 and L2 experiments shared the
same syntactic structure: verb + Objekt. A frequency norming test and familiarity rating test were
conducted to ensure that stimuli across conditions did not differ significantly in the aspects of
word frequency and word familiarity (p > 0.01). Motor-relatedness of all stimuli was evaluated
on a 5-point scale (1: not related at all; 5: very related ): L1 experiment (literal: 4.60 ± 0.40;
metaphorical: 2.78 ± 1.24; abstract: 2.12 ± 1.23) and the L2 experiment (literal: 4.39 ±
0.58; metaphorical: 2.64 ± 0.96; abstract: 2.11 ± 1.04). Only the first verb phrase, welches ist
independent of task-related strategic manipulations, was used for further MEG analysis.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
.
/
l
Experimental Procedure
L1 and L2 experiments shared the same experimental procedure. As suggested by previous
Studien, L1 could have a stronger translation priming effect on L2 than the other way around
(d.h., asymmetrical cross-language priming effects; Chen et al., 2014; Keatley et al., 1994;
Smith et al., 2019). To avoid the translation priming effect, L1 experiment was presented after
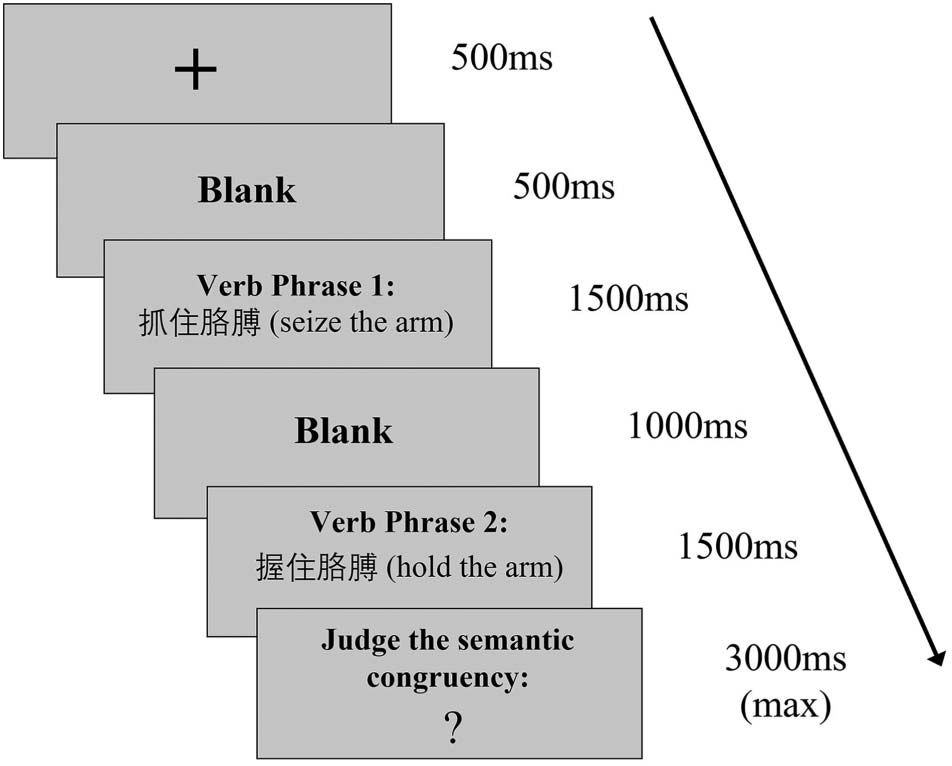
L2 experiment. Trials were shown in a pseudo-randomized order. As shown in Figure 1, jede
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Literal
Tisch 1.
An exemplar of stimuli in the L1 and L2 experiment
L1 (Chinese)
抓住胳膊–握住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
L2 (English)
seize the arm – hold the arm
抓住胳膊–摔伤胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
抓住胳膊
seize the arm – hurt the arm
Metaphorical
抓住机会–把握机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
seize the chance – grab the chance
抓住机会–错过机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
抓住机会
seize the chance – give up the chance
Abstrakt
珍惜机会–爱惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
cherish the chance – appreciate the chance
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会
珍惜机会–放弃机会
cherish the chance – abandon the chance
Notiz. The L1 and L2 stimuli are semantically equivalent.
Neurobiology of Language
182
Compensatory role of the motor cortex in second language processing
Figur 1.
Schematic view of the experimental procedure.
trial began with a 500 ms fixation at the center of the screen, followed by a 500 ms long blank
interval. Nachher, the first verb phrase was presented for a duration of 1,500 MS, followed by
A 1,000 ms long blank interval. The second verb phrase was then presented for 1,500 MS,
followed by “?” with a duration of maximal 3,000 MS. Participants were expected to give a
response after the “?” appeared. Visual stimuli were presented using Presentation software
(Neurobehavioral Systems, 2022). L1 stimuli were in KaiTi font and L2 stimuli in Times
New Roman font. The viewing distance from participants’ eyes to the stimuli on the projection
screen was one meter. The L1 stimuli subtended a horizontal visual angle of 3° 50, and the L2
stimuli subtended a horizontal visual angle of 4° 580.
MEG Data Recording
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
/
.
l
Continuous neuromagnetic signals were recorded using a 306-channel (102 magnetometers
Und 204 planar gradiometers) whole-head MEG system (MEGIN Oy, 2022) in a magnetically
shielded room at the Centre for Interdisciplinary Brain Research, University of Jyväskylä,
Finland. The head position of each subject was monitored by five head-position indicator
(HPI) coils attached over the forehead and behind each ear. Electrooculography signals were
recorded simultaneously by four electrodes attached around the eyes: above/below the right
eye, near the corner of the left/right eye. One ground electrode was attached to the collar
bone. The position of three fiducial landmarks (nasion, left/right preauricular points), sowie
as approximately 120 digitization points over the scalp, were acquired to establish the head
coordinate frame for the coregistration between MEG data and the MRI template. MEG signals
were online bandpass filtered at 0.1–330 Hz with a sampling rate of 1000 Hz.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
MEG Data Preprocessing and Source Estimation
Raw MEG data were processed in MaxFilter 2.2 (Elekta, 2010) with the time-domain extension
of the signal space separation method to minimize external magnetic disturbance and within-
sensor artifacts and to compensate for head movement (Taulu & Kajola, 2005). Head position
was estimated with a buffer length of 30 s and a correlation limit of 0.980. Head movement
correction was performed using a 200 ms window with a 10 ms step. The error limit of HPI coil
fit acceptance was 5 mm with a g-value of 0.98.
Neurobiology of Language
183
Compensatory role of the motor cortex in second language processing
The preprocessing was performed with Meggie, a graphic user interface built in-house based
on MNE-Python software (Gramfort et al., 2013). Erste, visual inspection was done to identify and
exclude the bad data segments in the continuous MEG data. Dann, MEG data were resampled to
250 Hz. A lowpass filter of 40 Hz (transition bandwidth 0.5 Hz, filter length 10 S) was applied.
Physiological artifacts related to heartbeat, blink, and saccade were removed using a semiauto-
matic independent component analysis method. Event-related epochs were extracted from
–200 ms to 1,000 ms relative to the onset of the first verb phrase. A 200 ms interval before
the onset was used as the baseline. MEG epochs with an amplitude exceeding 3,000 fT/cm for
gradiometers or 4,000 femtoteslas (fT) for magnetometers were rejected from further analysis.
In the calculation of evoked responses for the literal, metaphorical, and abstract conditions,
the first verb phrase was combined across the congruent and incongruent trials. Evoked
responses were obtained by averaging the signals of each condition (literal, metaphorical,
and abstract) in each experiment (L1 and L2 experiment).
Source estimation was performed in MNE-Python ( Version 0.17.0; Gramfort et al., 2013).
The CN200 template (https://www.nitrc.org/projects/us200_cn200; Yang et al., 2020), based
on T1-weighted magnetic resonance images of 250 healthy Chinese adults, was used for cor-
tical reconstruction and volumetric segmentation. Coregistration between the CN200 template
scalp and the digitized scalp was performed for each participant using a three-axis scaling
mode. Shrunk covariance with cross-validation was used to estimate the noise-covariance
Matrix (Engemann & Gramfort, 2015).
Dynamic statistical parametric mapping (dSPM; Dale et al., 2000), which is based on
minimum-norm estimate (Hämäläinen & Ilmoniemi, 1994), was used for source estimation
with a source space consisting of 4,098 vertices and 4,098 loose-constraint and depth-
weighted current dipoles (loose = 0.2, depth = 0.8) distributed on the cortical surface in each
hemisphere. Source estimation results were then noise normalized using the dSPM. The source
estimates across participants were morphed to the same cortical space (CN200 template).
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
ROI Selection
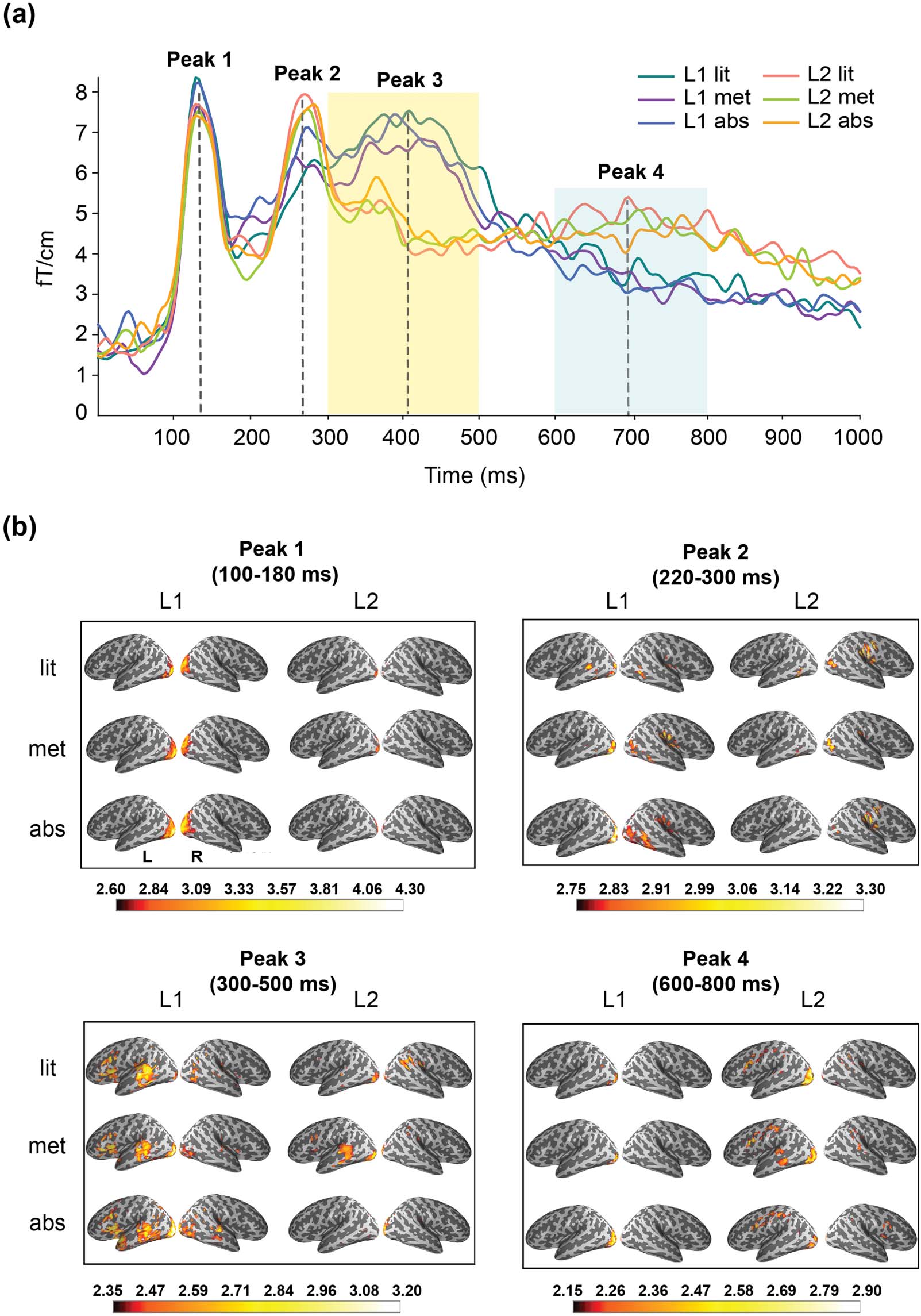
Regions of interest were selected in a hybrid way. Erste, based on the timing of peak activities in
the grand-averaged sensor waveform (Figure 2A), the spatial distribution of cortical sources
corresponding to each peak was identified (Figure 2B).
Nächste, the source distribution was compared against previous meta-analysis results of
neuroimaging studies pertaining to semantic processing and motor performance/imagery.
Brain regions appearing in both the data-derived cortical activation maps and previous
meta-analyses were selected as ROIs for the present study. The selection was done by using
MNE_analyze (https://mne.tools/0.17/manual/gui/analyze.html#the-labels-menu; Gramfort
et al., 2014). Erste, label names corresponding to the literature-derived brain regions were
selected from the parcellation list (Destrieux Atlas a2009s; Destrieux et al., 2010). Dann, Die
partition of the selected region was overlaid with the MEG data on the inflated cortical surface.
Only areas which showed prominent activation within the partitions were selected as ROIs.
Both language and motor ROIs were selected left-lateralized due to only minor activation in
the right hemisphere (Figure 2B). All ROIs were parcellated based on the Destrieux Atlas
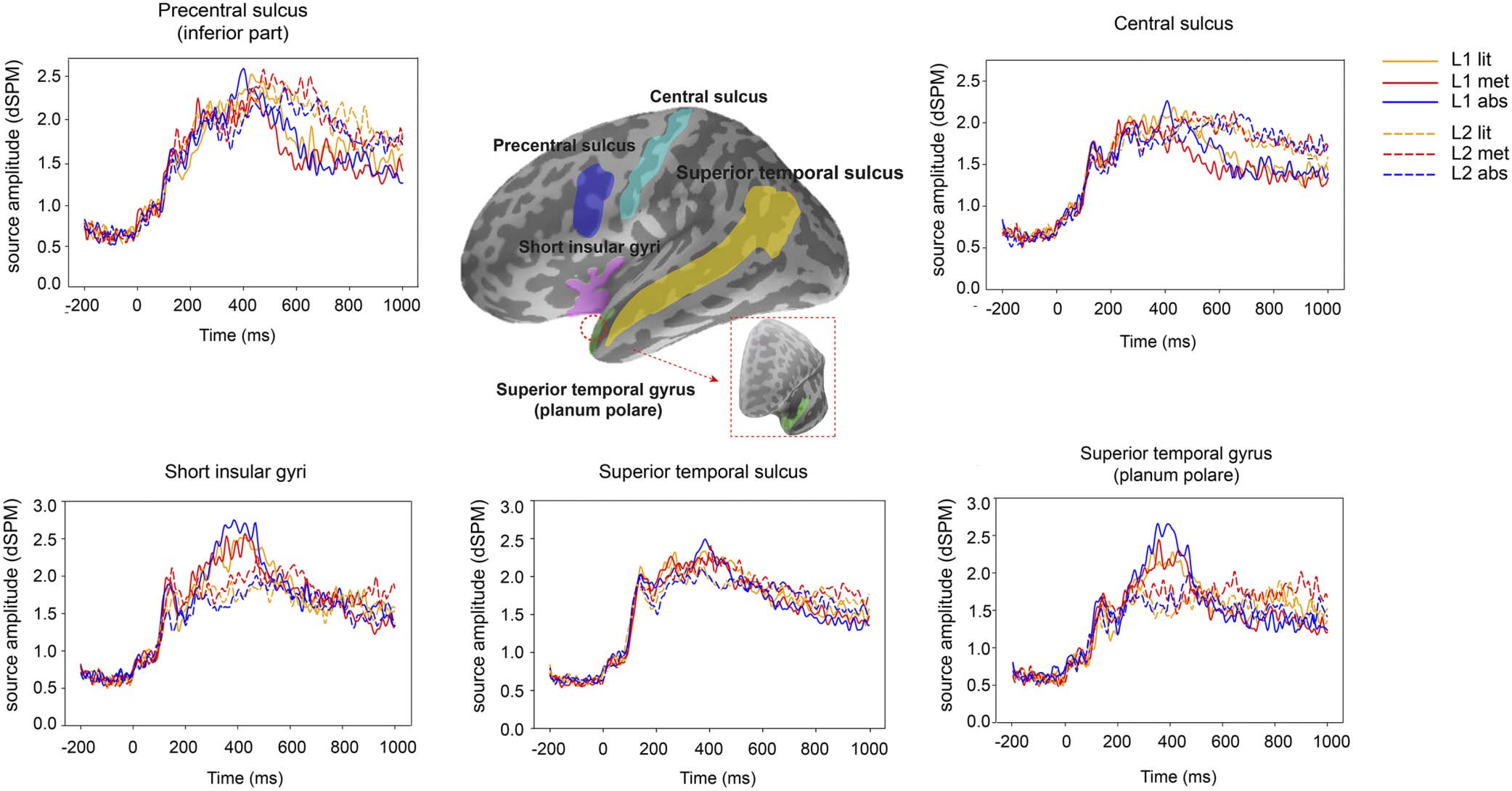
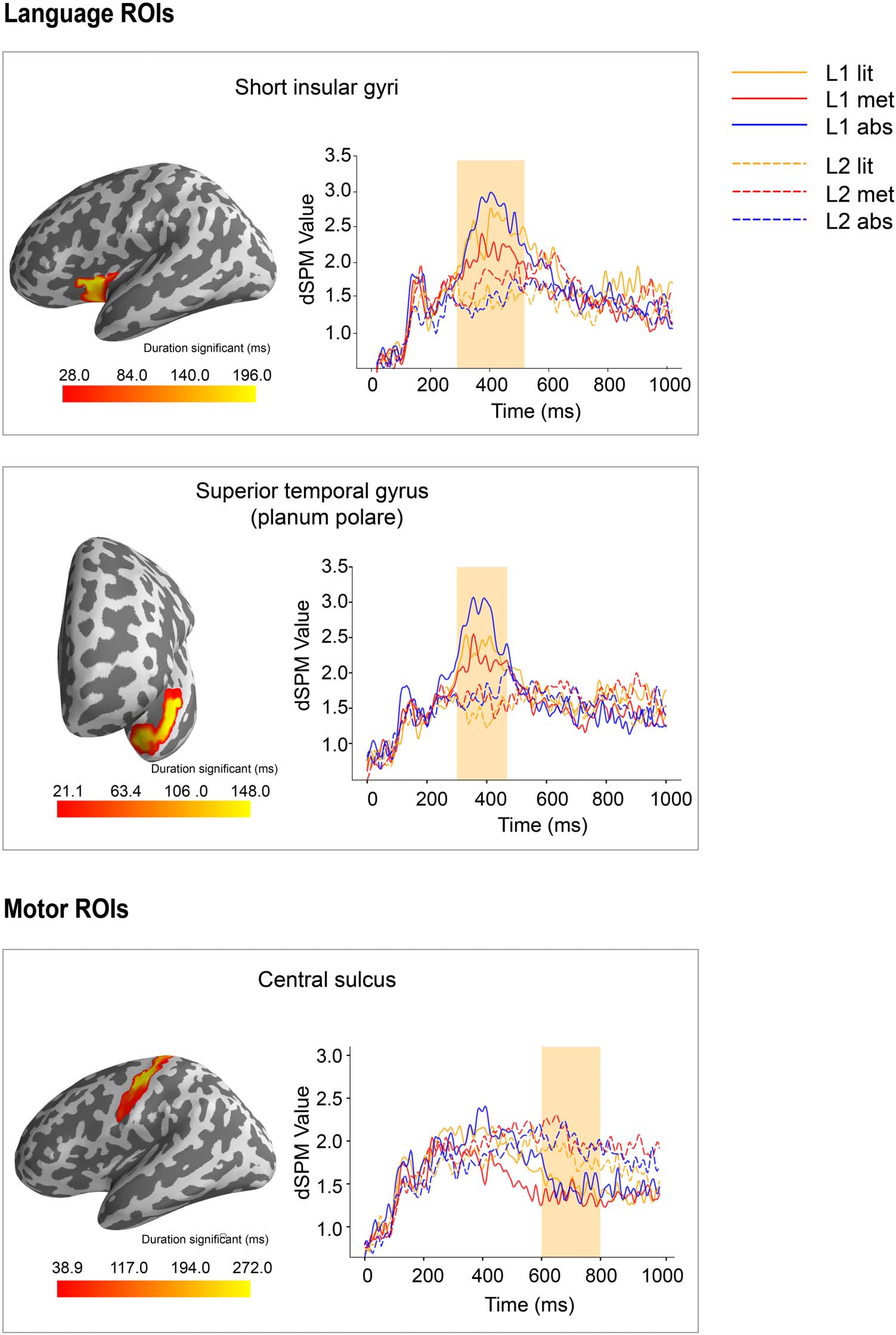
a2009s (Destrieux et al., 2010; see the schematic view of ROIs in Figure 3).
The above procedure resulted in the following language ROIs: short insular gyri (partially
overlapping with inferior frontal gyrus; Binder et al., 2009; Friederici et al., 2003; Rueckl et al.,
2015), planum polare of the superior temporal gyrus (part of anterior temporal cortex;
Neurobiology of Language
184
Compensatory role of the motor cortex in second language processing
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 2. Grand-averaged results at sensor and source level. (A) Grand-averaged sensor waveform (204 gradiometers) across conditions in
the L1 and L2 experiments; the light-shaded areas indicate the time window applied to the region-of-interest-based statistical analysis. (B)
Grand-averaged source activation quantified as mean dynamic statistical parametric mapping (dSPM) value over time points corresponding
to each peak: ±40 ms duration prior and after the relatively transient peaks (peak 1 and peak 2), and ±100 ms prior and after the relatively
sustainable peaks (peak 3 and peak 4). The intensity of the color in the cortical activation map indicates the degree of dSPM value. L1: einheimisch
Sprache (Chinese); L2: second language (English); zündete: literal; met: metaphorical; abs: abstract; L: left hemisphere; R: right hemisphere.
Neurobiology of Language
185
Compensatory role of the motor cortex in second language processing
Figur 3. Grand-averaged source time courses for the literal, metaphorical, and abstract conditions in the L1 and L2 experiments in the
indicated ROIs (language ROIs: short insular gyri, planum polare of the superior temporal gyrus, and superior temporal sulcus; motor ROIs:
inferior part of the precentral sulcus and central sulcus). The parcellation of each ROI is shown in the inflated brain surface with a lateral view.
For a better view, the planum polare of the superior temporal gyrus is also shown with a rostral view.
Carreiras et al., 2013; Lambon Ralph et al., 2017; Patterson et al., 2007), and superior
temporal sulcus (Citron et al., 2020; Rueckl et al., 2015). Motor ROIs were selected as
the inferior part of precentral sulcus and central sulcus (part of primary motor cortex; Hari
et al., 1998; Hétu et al., 2013; Michelon et al., 2006; Porro et al., 1996; Yousry et al., 1997).
The ROI-based source time courses are shown in Figure 3.
Time Window Selection
The time window was selected based on the latency of peak activities in the grand-averaged
sensor waveform (Figure 2A) and the corresponding time-resolved source activation maps
(Figure 2B). Based on the visual inspection, four peaks were identified in the sensor waveform:
peak 1 at around 140 MS, peak 2 bei 260 MS, peak 3 bei 400 MS, and peak 4 bei 700 MS.
Based on the source activation map, the first two peaks reflected activation in the visual cortex
(peak 1) and more distributed areas across occipital-temporal lobes (peak 2), which were not
included for statistical analysis. During peak 3 (300–500 ms, with peak activity at around 400 MS)
and peak 4 (600–800 ms, with peak activity at around 700 MS), activation was found within tem-
poral and frontal-central lobes, overlapping with our selected ROIs. daher, these two time
windows, TW1 (300–500 ms) and TW2 (600–800 ms), were used for further statistical analysis.
Statistical Analysis
Statistical analysis was performed on the amplitude of the source waveform (represented as
dSPM value) extracted from each ROI separately for TW1 and TW2. To examine the effect of
language proficiency and linguistic abstractness, the nonparametric two-way repeated measures
Neurobiology of Language
186
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
N
Ö
_
A
_
0
0
0
9
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Compensatory role of the motor cortex in second language processing
analysis of variance with spatiotemporal clustering was performed in MNE-Python. To solve the
multiple comparison problem, a cluster-based permutation test across time and space was
employed (Maris & Oostenveld, 2007). The randomization times of the permutation test were
1,000, with a threshold for cluster inclusion α = 0.05 and the permutation significance α = 0.05.
The p-values across language ROIs and motor ROIs were corrected for multiple comparison
using Benjamini-Hochberg false discovery rate (FDR; Benjamini & Hochberg, 1995).
ERGEBNISSE
Verhaltensergebnisse
We estimated the behavioral competence in L1 and L2 by analyzing the behavioral performance
in the semantic judgment task. There was no significant difference between the L1 and L2 in the
reaction time (p > 0.05, L1 (mean, SD): 587.12 ms ± 62.14, L2: 626.20 ms ± 71.59), aber die
accuracy rate was higher in L1 (96.25% ± 1.8%) than in L2 (91.94% ± 3.9%) (P < 0.001).
General Pattern and Time Course of Activation
The rough level activation timing (grand-averaged sensor waveform across the 204 gradiome-
ters) and spatial distribution (source activation within the major activation peaks) across con-
ditions in the L1 and L2 experiments are shown in Figure 2A and 2B. The source activation map
revealed robust activation in the occipital lobe at 130 ms for both the L1 and L2, with slightly
greater amplitude for the L1 than the L2. At around 260 ms, activation was found in the pos-
terior temporal area for the L1 and in the lateral occipital-temporal area for the L2. At the peak
around 400 ms, a notably greater amplitude was observed for the L1 than the L2. Activation in
L1 was broadly distributed to the insular area (partially overlapping with the inferior frontal
gyrus), posterior temporal area, anterior temporal area, inferior part of precentral area and cen-
tral area. For the L2 (mainly the metaphorical condition), robust activation was observed mainly
in the posterior temporal area. At around 700 ms, the pattern between L1 and L2 was reversed:
L2 showed greater amplitude than L1 in the central and precentral areas.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
.
/
l
Table 2.
Statistical results of region-of-interest (ROI) analyses on source data
Parcellation (Destrieux
Atlas a2009s)a
Language ROIs G_insular_short
G_temp_sup-Plan_polar
Terminologia
Anatomicab
Short insular gyri
Planum polare of
the superior
temporal gyrus
p-values (FDR-corrected)
#Vertice
732
A:B
0.095
300–500 ms
A
0.042*
B
0.258
A:B
0.243
600–800 ms
A
0.970
B
0.240
876
0.215
0.042*
0.258
0.406
0.613
0.290
S_temporal_sup
Superior temporal
5216
0.095
0.326
0.271
0.298
0.842
0.451
Motor ROIs
S_precentral-inf-part
sulcus
Inferior part of the
precentral sulcus
1587
0.056
0.466
0.859
0.465
0.056
0.246
S_central
Central sulcus
3139
0.073
0.515
0.512
0.465
0.020*
0.246
Note. A:B: interactions between language proficiency and linguistic abstractness; A: the main effect of language proficiency; B: the main effect of linguistic
abstractness. The anatomical parcellation was based on Destrieux Atlas a2009s. Statistical significance (p < 0.05) is marked in bold with an asterisk.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
a Destrieux et al. (2010).
b FIPAT (2019).
Neurobiology of Language
187
Compensatory role of the motor cortex in second language processing
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Results of permutation F test on the language and motor ROIs. (Left) Significant clusters
at source space (clusters exceeding the randomization distribution under H0 hypothesis). The inten-
sity of the color in the cortical map indicates the duration of the time window of clusters. (Right)
Source time courses (represented as dSPM value) extracted from significant spatiotemporal clusters;
light orange shading area shows time window of the significant cluster. dSPM: dynamic statistical
parametric mapping.
Neurobiology of Language
188
Compensatory role of the motor cortex in second language processing
Statistical Results
Cluster-based permutation F test on source data was performed for each language and motor
ROI in the TW1 (300–500 ms) and TW2 (600–800 ms) respectively. Statistical results are
shown in Table 2. Language and motor ROIs with significant spatiotemporal clusters are
shown in Figure 4.
For the language ROIs, in the TW1, the cluster-based permutation F test revealed a signif-
icant main effect of language proficiency in the short insular gyri (p = 0.042) and the planum
polare of the superior temporal gyrus (p = 0.042), manifested as greater activation within these
areas for the L1 than for the L2. Statistical analysis did not reveal any significant interaction
effect or the main effect of abstractness. In the TW2, no significant effect was found for
language ROIs.
For the motor ROIs, no significant effect was found in the TW1. In the TW2, results showed
a significant main effect of language proficiency in the central sulcus (p = 0.020), manifested
as greater activation in the L2 than in the L1. No significant interaction effect or the main effect
of abstractness was found.
DISCUSSION
In this MEG study, we investigated the degree of involvement of the language and motor areas
in a language comprehension task. We employed spatiotemporally sensitive MEG recordings,
which allowed us to examine the temporal trajectory of language and motor cortex activation.
Specifically, we investigated whether the degree of involvement of language and motor areas
in the stage of semantic processing was modulated by learner-specific factors (i.e., language
proficiency), and/or by stimulus-specific factors (i.e., level of abstractness of the language
stimuli).
Our source analysis evidenced a typical spatiotemporal trajectory of visual word process-
ing, which witnessed an early robust activation in the occipital area, followed by activation
flowing from the posterior to the anterior temporal and frontal areas (Brennan & Pylkkänen,
2012; Carreiras et al., 2013). In addition, the source estimation results showed neural activa-
tion of motor areas across all conditions (literal, metaphorical, and abstract) in both native
language (L1) and second language (L2). More importantly, our results showed an overall
greater involvement of language areas (short insular gyri and planum polare of the superior
temporal gyrus) in the L1 than in the L2 in the time window of 300–500 ms, which has been
broadly associated with semantic analysis (Kutas & Federmeier, 2011; Lau et al., 2008; Lau
et al., 2013). Although greater activation in the posterior superior temporal sulcus can be seen
for the L1 than the L2 in the grand-averaged source results (Figure 2B), it failed to show any
statistically significant difference. In addition, our results showed an overall greater involve-
ment of motor area (central sulcus) in the L2 than in the L1 in the late time window of 600–
800 ms, which might be associated with post-semantic analysis and integration.
Compensatory Role of the Motor Cortex in Late-Acquired L2 Processing
Our findings corroborate previous studies in showing that the motor cortex is involved in the
processing of not only the L1 but also the L2 (Birba et al., 2020; De Grauwe et al., 2014;
Monaco et al., 2021; Tian et al., 2020; Vukovic & Shtyrov, 2014; Zhang et al., 2020). In fact,
our results suggest a stronger role for motor areas in the L2 than the L1. Our findings are also in
line with earlier studies which suggested that the motor (or sensorimotor) area is involved in
the processing of not only action-related but also abstract meaning (Dreyer & Pulvermüller,
Neurobiology of Language
189
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
2018; Guan et al., 2013; Tian et al., 2020; Vukovic et al., 2017). These findings jointly indicate
that motor cortex involvement is ubiquitous in semantic processing, regardless of the linguistic
features of the stimuli.
The stronger involvement of the motor cortex in the L2 semantic processing, independent of
its linguistic abstractness, allows us to speculate on its role in language processing more gen-
erally. The finding is in line with some previous studies showing greater motor activation in the
L2 than the L1 (Monaco et al., 2021; Tian et al., 2020), though not exactly in the same time
window (275 ms after onset in Monaco et al.’s study, 600 ms in the present study). The some-
what earlier emergence of the effect in Monaco et al. may arise from the use of single verbs,
while in our study the stimuli were verb phrases, which are relatively more complex seman-
tically, and may evoke longer-lasting cortical engagement. In addition, the semantic task in
Monaco et al.’s study required explicit motor simulation, as participants needed to judge if
the verb represents a physical or mental action. In contrast, the task in our study only required
the evaluation of semantic congruency and did not require any action-related judgment.
Although the underlying process in L1 and L2 may be different between Monaco et al.’s study
and ours, both studies indicate stronger involvement of motor areas in L2.
However, there are also contradictory findings. The results of Vukovic and Shtyrov’s study
(2014) pointed to greater involvement of the motor cortex in the L1 than the L2, indicated by
stronger mu rhythm ERD. This apparently opposite pattern may at least partly be due to the
differences in the brain activation measures. ERD (and event-related synchronization) reflects
the temporal changes in the power of oscillations, and particularly the 10–20 Hz (hence mu
rhythm) is often associated with the level of top-down inhibitory control. Unlike ERD, evoked
responses, on the other hand, are time and phase locked to the onset of incoming sensory
input and are likely to reflect a different source of neuronal activation. Particularly for the later
stages of activation, evoked responses are likely to represent activation of a distributed net-
work, the center of which is represented by the spatial extent of the source model. In their
study, Vukovic and Shtyrov interpreted the stronger modulation for the L1 as the results of a
more integrated perception-action circuit for the L1 lexical-semantic representation and a
higher degree of embodiment for the L1. An alternative interpretation of their findings may,
however, be that even though the task did not require verbal output, L1 more readily and auto-
matically engages articulatory preparation, which may manifest as stronger predictive (i.e.,
top-down) allocation of resources in the motor areas. This interpretation would be in line with
the results of anticipatory alpha modulation in visual and language domains (Wang et al.,
2018) and challenges the embodied interpretation of the findings. The stronger and automatic
recruitment of motor representations in the L1 in early time windows would also be compat-
ible with increased engagement of motor areas in the L2 in later time windows (as shown in
our study). Indeed, given the strongly time-evolving nature of language processing in the brain,
it is conceivable that the role of the motor cortex may vary across time. As the source result
shows in our study, the activation in the L1 (but not the L2) extended to the precentral sulcus in
300–500 ms, although the difference between L1 and L2 did not show statistically significant
clusters.
The discussion of the role of the motor cortex in language comprehension may thus need to
be approached with increased resolution (both temporally and spatially), as different neuroim-
aging modalities, and even different neural measures derived by same modality suggest diver-
gent roles. It is also of crucial importance to acknowledge the time-varying nature of language
processing. In addition to the methodological concerns, the search for functional significance
of motor cortex also requires rigorous use of reasoning in interpreting the neuroscientific find-
ings. Indeed, it needs to be noted that the greater degree of motor cortex activation may not
Neurobiology of Language
190
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
necessarily imply a higher degree of embodiment. As has been pointed out, the involvement
of a certain cognitive process cannot be unequivocally inferred from the presence of brain
activation of a certain region (cf. reverse inference, e.g., Henson, 2006; Mahon & Hickok,
2016; Poldrack, 2006), as a particular brain region may carry multiple cognitive functions with
a primary or secondary role.
The difficulty in specifying the correspondence relationship between brain regions and
cognitive functions also applies to neuroimaging studies concerning action-related language
processing. Neural activation of the motor cortex has mostly been elucidated as the result of
utilizing the motor cortex for mentally simulating action-related meaning. The inference is
made based on the established fact that the motor cortex is engaged in motor execution, motor
planning, and motor imagery, as has been widely reported (Filimon et al., 2007; Hanakawa
et al., 2008; Leonardo et al., 1995). Consequently, motor activations in the studies of semantic
processing are believed to indicate the engagement of the motor cortex in the mental simula-
tion of action-related meanings. However, the motor cortex, in addition to its motor-related
cognitive functions, has also been shown to be functionally involved in other cognitive pro-
cesses in a sub-dominant way, including (procedural) memory retrieval, cognitive control,
inhibition, and integration (Francis, 2005; Miller, 2000; Mofrad et al., 2020; Lambon Ralph
et al., 2017; Ullman, 2004; Willems et al., 2010). In the context of language processing, as
mentioned in Maieron et al.’s (2013) study, the engagement of the primary motor cortex may
be related to other aspects of cognitive processing rather than specific linguistic processing,
which was inferred based on the lack of modulation of language-motor coupling during the
action-verb generation task for both lesion and healthy groups.
In the present study, greater activation of the motor cortex was found for the L2 than the L1
across conditions. Referring to the above reasoning, we are of the opinion that the greater acti-
vation of the motor cortex may not imply a greater degree of embodiment in the semantic
processing of the L2, but a higher demand for cognitive resources to compensate for its lower
proficiency and weaker semantic representation compared with the L1. The interpretation is
made based on the joint findings of the underactivation of the language areas (short insular gyri
and planum polare of the superior temporal gyrus) at the semantic processing stage and the
overactivation of the motor area (central sulcus) at the post-semantic processing stage in L2,
compared with L1. The planum polare of the superior temporal gyrus, as part of the anterior
temporal lobe, has been shown to be a semantic hub for integrating domain-specific concepts
and semantic integration in general (Lambon Ralph et al., 2017; see review by Visser et al.,
2010). Interpreted in the context of the present study, the L1 with richer semantic representa-
tion (compared with the L2) is likely to engage a greater degree of the anterior temporal lobe
for meaning processing. In contrast, the weaker semantic representation of L2 may cost longer
time for participants to access the meaning of L2, which might account for the early under-
activation in language areas. The motor cortex was over-recruited, presumably, to offset the
inadequate engagement of language areas, as a result of weaker semantic representation of L2,
compared with L1. However, it is important to collect more direct evidence on the causal role
of language and motor areas in linguistic tasks, as neuroimaging studies are necessarily cor-
relative in nature.
Similar interpretation about the compensatory mechanism has also been reported in a study
of individuals with dyslexia (Richlan et al., 2011), where underactivation in the left temporal
region and overactivation in the motor cortex was found in adults with reading difficulty. This
lends support to the idea that motor areas may represent general supportive functions in case
of lower proficiency. Indeed, it has been validated by converging empirical evidence, that the
retrieval of weakly encoded information relies more strongly on the control network (Lambon
Neurobiology of Language
191
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
Ralph et al., 2017). Based on the above discussion, we argue that the greater activation of the
motor cortex in the L2 may not signify a higher degree of embodiment, but a higher demand
for cognitive resources to compensate for the inadequate engagement of the language
network.
Functional Role of the Motor Cortex
By clarifying the role of learner-specific (i.e., language proficiency) and stimulus-specific (i.e.,
abstractness) factors, our findings shed light on the functional role of the motor cortex in
language processing. There has been a longstanding debate on the functional and epiphenom-
enal role of motor cortex involvement in the literature on embodied language processing
(Bocanegra et al., 2017; Desai et al., 2015; Fernandino et al., 2013; García et al., 2019; Reilly
et al., 2019; Repetto et al., 2013; van Elk et al., 2010; Vukovic et al., 2017). Similar to our
paradigm, some earlier studies attempted to disentangle these two roles by referring to the time
course of the motor cortex activation, compared with that of the language areas (García et al.,
2019; Papeo et al., 2009; Reilly et al., 2019; van Elk et al., 2010). The motor-related activa-
tions or modulations occurring at an early stage of semantic processing (130–190 ms post-
stimulus in García et al., 2019; 300 ms in Reilly et al., 2019; 400 ms in van Elk et al.,
2010) are considered as evidence supporting the assumption of the functional (i.e., necessary)
role, which claims that the motor cortex directly contributes to semantic processing, while
activations occurring at a later stage are considered to reflect post-semantic motor imagery
(500 ms in Papeo et al., 2009) and not necessarily contributing to language comprehension.
However, the onset of semantic processing is unlikely to be clearly defined by a fixed time
point, and it may vary considerably depending on learner-related factors (e.g., language pro-
ficiency and language experience) and language-related factors (e.g., language distance). For a
less proficient language, the latency of lexical-semantic retrieval and integration can be
delayed compared with the highly proficient native language. Considering the influence of
language proficiency, we assume that the greater activation of the motor cortex in the L2 in
our study is not the result of post-semantic motor imagery but reflects the general cognitive
processes that support semantic processing in an indirect way. It may thus be useful for the
discussion of the functional or epiphenomenal role of the motor network to focus not only on
latency of motor cortex activation, but also on language proficiency, which may lead to var-
iance in the latency of semantic access.
The Null Effect of Abstractness
Our study did not reveal any significant effect of abstractness, suggesting that neural responses
in the motor areas may not be modulated by the degree of abstractness of the linguistic input.
The finding is inconsistent with our prediction of decreased motor involvement with the
increase of abstractness. Our finding is also inconsistent with previous studies exploring the
effect of abstractness on a continuum (i.e., literal, metaphorical (idiomatic), and abstract; Desai
et al., 2013; Tian et al., 2020). In their studies, hierarchically attenuated motor activation was
found with the increase of linguistic abstractness.
So far, most studies concerning the effect of abstractness on motor cortex involvement
mainly focused on literal and figurative action-related language (mainly metaphorical and idi-
omatic). Some revealed greater involvement of motor cortex for literal than figurative language
(Cacciari et al., 2011), and some reported a similar degree of motor cortex involvement
between them (Boulenger et al., 2009; Boulenger et al., 2012). Inconsistently, some other
studies found motor cortex involvement only for the literal language, but not for the figurative
Neurobiology of Language
192
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
(Raposo et al., 2009). The discrepancy in findings may derive from methodological differences
across studies, including task demands (covert vs. overt motor association), stimulus properties
(word vs. phrase vs. sentence), and ways of presentation (word-by-word vs. whole item, visu-
ally vs. aurally). As has been highlighted, the recruitment of the motor cortex in action seman-
tic processing is task (Giacobbe et al., 2022; Tomasino et al., 2008) and context dependent
(Raposo et al., 2009).
Moreover, current findings call for a reflection on the relationship between artificial cate-
gorization of abstractness and its actual brain response. Although the stimuli do follow a
linguistically defined continuum of abstractness, the actual brain responses may not follow
such gradation. In future studies, it will be important to test the modulatory effect of abstract-
ness on the degree of motor cortex recruitment by using comparable approaches.
Limitations
Our study has some limitations. First, our study is a correlative study in nature, and interpre-
tations are mainly “bound” to earlier literature. Second, our study only included ROI-based
analysis motivated by its hypothesis-driven nature. The exclusion of whole-brain analysis
may cause the ignorance of important neural activity in other brain regions. Future studies
should further investigate the relationship between language and motor networks in bilingual
language processing by employing comparable approaches.
Conclusion
Our study explored the degree of involvement of language and motor areas modulated by
language proficiency and linguistic abstractness. We reported an overall greater activation in
the language areas for the L1 than the L2 at the semantic processing stage at 300–500 ms, and
an overall greater activation in the motor regions for the L2 than the L1 at the later post-semantic
processing stage at 600–800 ms. The over-recruitment of the motor areas in the L2 implied a
compensatory role of the motor area to offset the lower language proficiency of the L2 in
relative to the L1. Our study provides an alternative interpretation of motor cortex involvement
in language processing and invites further research to explore the factors that modulate this
relationship.
ACKNOWLEDGMENTS
The authors would like to thank Aino Sorsa for her assistance in data collection and Dr. Viki-
Veikko Elomaa for program setup. The authors would also like to thank Dr. Weiyong Xu,
Dr. Xiulin Wang, Dr. Simo Monto, and Erkka Heinilä for their assistance and suggestions in
data analysis.
FUNDING INFORMATION
Lili Tian, China Scholarship Council (https://dx.doi.org/10.13039/501100004543), Award ID:
201708500099.
AUTHOR CONTRIBUTIONS
Lili Tian: Conceptualization; Formal analysis; Investigation; Writing – original draft; Writing –
review & editing. Hongjun Chen: Conceptualization. Pyry Petteri Heikkinen: Formal analysis.
Wenya Liu: Formal analysis. Tiina Parviainen: Conceptualization; Resources; Writing – review
& editing.
Neurobiology of Language
193
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
DATA AND CODE AVAILABILITY STATEMENTS
The data are not publicly available due to the restrictions of research ethics stated in the Pri-
vacy Notice for Research Subjects in terms of the privacy of research participants.
The data that support the findings of this study are available upon reasonable request from
Tiina Parviainen (tiina.m.parviainen@jyu.fi) and Lili Tian (litian@jyu.fi).
In compliance with the General Data Protection Regulation, the following situation will be
approved when requesting the data: (1) actions aiming to confirm and verify the validity and
authenticity of the results of the current research; (2) actions related to scientific research or
other compatible purpose.
REFERENCES
Barsalou, L. W., Santos, A., Simmons, W. K., & Wilson, C. D.
(2008). Language and simulation in conceptual processing. In
M. de Vega, A. M. Glenberg, & A. C. Graesser (Eds.), Symbols,
embodiment, and meaning (pp. 245–283). Oxford University
Press. https://doi.org/10.1093/acprof:oso/9780199217274.003
.0013
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society Series B (Methodological),
57(1), 289–300. https://doi.org/10.1111/j.2517-6161.1995
.tb02031.x
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral Cortex,
19(12), 2767–2796. https://doi.org/10.1093/cercor/ bhp055,
PubMed: 19329570
Birba, A., Beltrán, D., Caro, M. M., Trevisan, P., Kogan, B., Sedeño,
L., Ibáñez, A., & García, A. M. (2020). Motor-system dynamics
during naturalistic reading of action narratives in first and second
language. NeuroImage, 216, Article 116820. https://doi.org/10
.1016/j.neuroimage.2020.116820, PubMed: 32278096
Bocanegra, Y., García, A. M., Lopera, F., Pineda, D., Baena, A.,
Ospina, P., Alzate, D., Buriticá, O., Moreno, L., Ibáñez, A., &
Cuetos, F. (2017). Unspeakable motion: Selective action-verb
impairments in Parkinson’s disease patients without mild cogni-
tive impairment. Brain and Language, 168, 37–46. https://doi.org
/10.1016/j.bandl.2017.01.005, PubMed: 28131052
Boulenger, V., Hauk, O., & Pulvermüller, F. (2009). Grasping ideas
with the motor system: Semantic somatotopy in idiom compre-
hension. Cerebral Cortex, 19(8), 1905–1914. https://doi.org/10
.1093/cercor/bhn217, PubMed: 19068489
Boulenger, V., Shtyrov, Y., & Pulvermüller, F. (2012). When do you
grasp the idea? MEG evidence for instantaneous idiom under-
standing. NeuroImage, 59(4), 3502–3513. https://doi.org/10
.1016/j.neuroimage.2011.11.011, PubMed: 22100772
Brennan, J., & Pylkkänen, L. (2012). The time-course and spatial
distribution of brain activity associated with sentence processing.
NeuroImage, 60(2), 1139–1148. https://doi.org/10.1016/j
.neuroimage.2012.01.030, PubMed: 22248581
Buccino, G., Volta, R. D., Arabia, G., Morelli, M., Chiriaco, C.,
Lupo, A., & Quattrone, A. (2018). Processing graspable object
images and their nouns is impaired in Parkinson’s disease
patients. Cortex, 100, 32–39. https://doi.org/10.1016/j.cortex
.2017.03.009, PubMed: 28413070
Cacciari, C., Bolognini, N., Senna, I., Pellicciari, M. C., Miniussi,
C., & Papagno, C. (2011). Literal, fictive and metaphorical
motion sentences preserve the motion component of the verb:
A TMS study. Brain and Language, 119(3), 149–157. https://doi
.org/10.1016/j.bandl.2011.05.004, PubMed: 21684590
Cardona, J. F., Kargieman, L., Sinay, V., Gershanik, O., Gelormini,
C., Amoruso, L., Roca, M., Peneda, D., Trujilla, N., Michon, M.,
García, A. M., Szenkman, D., Bekinschtein, T., Manes, F., &
Ibáñez, A. (2014). How embodied is action language? Neurolog-
ical evidence from motor diseases. Cognition, 131(2), 311–322.
https://doi.org/10.1016/j.cognition.2014.02.001, PubMed:
24594627
Carreiras, M., Armstrong, B. C., Perea, M., & Frost, R. (2013). The
what, when, where, and how of visual word recognition. Trends
in Cognitive Sciences, 18(2), 90–98. https://doi.org/10.1016/j.tics
.2013.11.005, PubMed: 24373885
Chatterjee, A. (2010). Disembodying cognition. Language and Cog-
nition, 2(1), 79–116. https://doi.org/10.1515/langcog.2010.004,
PubMed: 20802833
Chen, B., Zhou, H., Gao, Y., & Dunlap, S. (2014). Cross-language
translation priming asymmetry with Chinese-English bilinguals: A
test of the sense model. Journal of Psycholinguist Research, 43(3),
225–240. https://doi.org/10.1007/s10936-013-9249-3, PubMed:
23584838
Citron, F. M., Michaelis, N., & Goldberg, A. E. (2020). Metaphorical
language processing and amygdala activation in L1 and L2.
Neuropsychologia, 140, Article 107381. https://doi.org/10.1016
/j.neuropsychologia.2020.107381, PubMed: 32061649
Dale, A. M., Liu, A. K., Fischl, B. R., Buckner, R. L., Belliveau, J. W.,
Lewine, J. D., & Halgren, E. (2000). Dynamic statistical paramet-
ric mapping: Combining fMRI and MEG for high-resolution imag-
ing of cortical activity. Neuron, 26(1), 55–67. https://doi.org/10
.1016/S0896-6273(00)81138-1, PubMed: 10798392
De Grauwe, S., Willems, R. M., Rueschemeyer, S., Lemhöfer, K., &
Schriefers, H. (2014). Embodied language in first- and
second-language speakers: Neural correlates of processing motor
verbs. Neuropsychologia, 56, 334–349. https://doi.org/10.1016/j
.neuropsychologia.2014.02.003, PubMed: 24524912
Desai, R. H., Conant, L. L., Binder, J. R., Park, H., & Seidenberg,
M. S. (2013). A piece of the action: Modulation of sensory-motor
regions by action idioms and metaphors. NeuroImage, 83,
862–869. https://doi.org/10.1016/j.neuroimage.2013.07.044,
PubMed: 23891645
Desai, R. H., Herter, T., Riccardi, N., Rorden, C., & Fridriksson, J.
(2015). Concepts within reach: Action performance predicts
action language processing in stroke. Neuropsychologia, 71,
217–224. https://doi.org/10.1016/j.neuropsychologia.2015.04
.006, PubMed: 25858602
Destrieux, C., Fischl, B., Dale, A., & Halgren, E. (2010). Automatic
parcellation of human cortical gyri and sulci using standard
Neurobiology of Language
194
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
anatomical nomenclature. NeuroImage, 53(1), 1–15. https://doi
.org/10.1016/j.neuroimage.2010.06.010, PubMed: 20547229
Dreyer, F. R., & Pulvermüller, F. (2018). Abstract semantics in the
motor system?: An event-related fMRI study on passive reading of
semantic word categories carrying abstract emotional and mental
meaning. Cortex, 100, 52–70. https://doi.org/10.1016/j.cortex
.2017.10.021, PubMed: 29455946
Elekta. (2010). MaxFilter ( Version 2.2) [Software].
Engemann, D. A., & Gramfort, A. (2015) Automated model selec-
tion in covariance estimation and spatial whitening of MEG and
EEG signals. NeuroImage, 108, 328–342. https://doi.org/10.1016
/j.neuroimage.2014.12.040, PubMed: 25541187
Fargier, R., Paulignan, Y., Boulenger, V., Monaghan, P., Reboul, A.,
& Nazir, T. A. (2012). Learning to associate novel words with
motor actions: Language-induced motor activity following short
training. Cortex, 48(7), 888–990. https://doi.org/10.1016/j.cortex
.2011.07.003, PubMed: 21864836
Fernandino, L., Conant, L. L., Binder, J. R., Blindauer, K., Hiner, B.,
Spangler, K., Spangler, K., & Desai, R. H. (2013). Parkinson’s dis-
ease disrupts both automatic and controlled processing of action
verbs. Brain and Language, 127(1), 65–74. https://doi.org/10
.1016/j.bandl.2012.07.008, PubMed: 22910144
Filimon, F., Nelson, J. D., Hagler, D. J., & Sereno, M. I. (2007).
Human cortical representations for reaching: Mirror neurons
for execution, observation, and imagery. NeuroImage, 37(4),
1315–1328. https://doi.org/10.1016/j.neuroimage.2007.06.008,
PubMed: 17689268
FIPAT. (2019). Terminologia Anatomica (2nd ed.). Federative Inter-
national Programme for Anatomical Terminology. https://FIPAT
.library.dal.ca
Fischer, M. H., & Zwaan, R. A. (2008). Embodied language:
A review of the role of the motor system in language compre-
hension. Quarterly Journal of Experimental Psychology, 61(6),
825–850. https://doi.org/10.1080/17470210701623605,
PubMed: 18470815
Francis, W. S. (2005). Bilingual semantic and conceptual represen-
tation. In J. F. Kroll & A. M. De Groot (Eds.), Handbook of bilin-
gualism: Psycholinguistic approaches. Oxford University Press.
Friederici, A. D., Rüschemeyer, S. A., Hahne, A., & Fiebach, C. J.
(2003). The role of left inferior frontal and superior temporal
cortex in sentence comprehension: Localizing syntactic and
semantic processes. Cerebral Cortex, 13(2), 170–177. https://
doi.org/10.1093/cercor/13.2.170, PubMed: 12507948
Gallese, V., & Lakoff, G. (2005). The brain’s concepts: The role of
the sensory-motor system in conceptual knowledge. Cognitive
Neuropsychology, 22(3), 455–479. https://doi.org/10.1080
/02643290442000310, PubMed: 21038261
García, A. M., Moguilner, S., Torquati, K., García-Marco, E.,
Herrera, E., Muñoz, E., Castillo, E. M., Kleineschay, T., Sedeño,
L., & Ibáñez, A. (2019). How meaning unfolds in neural time:
Embodied reactivations can precede multimodal semantic effects
during language processing. NeuroImage, 197, 439–449. https://
doi.org/10.1016/j.neuroimage.2019.05.002, PubMed: 31059796
Giacobbe, C., Raimo, S., Cropano, M., & Santangelo, G. (2022).
Neural correlates of embodied action language processing: A
systematic review and meta-analytic study. Brain Imaging and
Behavior, 16(5), 2353–2374. https://doi.org/10.1007/s11682
-022-00680-3, PubMed: 35754077
Glenberg, A. M., & Kaschak, M. P. (2002). Grounding language in
action. Psychonomic Bulletin & Review, 9(3), 558–565. https://
doi.org/10.3758/BF03196313, PubMed: 12412897
Glenberg, A. M., Sato, M., Cattaneo, L., Riggio, L., Palumbo, D., &
Buccino, G. (2008). Processing abstract language modulates
motor system activity. Quarterly Journal of Experimental Psy-
c h o l o g y, 6 1 ( 6 ) , 9 0 5 – 9 1 9 . h t t p s : / / d o i . o r g / 1 0 . 1 0 8 0
/17470210701625550, PubMed: 18470821
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A., Strohmeier,
D., Brodbeck, C., Goj, R., Jas, M., Brooks, T., Parkkonen, L., &
Hämäläinen, M. (2013). MEG and EEG data analysis with
MNE-Python. Frontiers in Neuroscience, 7, Article 267. https://
doi.org/10.3389/fnins.2013.00267, PubMed: 24431986
Gramfort, A., Luessi, M., Larson, E., Engemann, D. Strohmeier, D.,
Brodbeck, C., Parkkonen, L., & Hämäläinen, M. (2014). MNE
software for processing MEG and EEG data. NeuroImage, 86,
446–460. https://doi.org/10.1016/j.neuroimage.2013.10.027,
PubMed: 24161808
Greco, A. (2021). Spatial and motor aspects in the “Action-
Sentence Compatibility Effect.” Frontiers in Psychology, 12,
Article 647899. https://doi.org/10.3389/fpsyg.2021.647899,
PubMed: 33897555
Guan, C. Q., Meng, W., Yao, R., & Glenberg, A. M. (2013). The
motor system contributes to comprehension of abstract language.
PLOS ONE, 8(9), Article e75183. https://doi.org/10.1371/journal
.pone.0075183, PubMed: 24086463
Hämäläinen, M. S., & Ilmoniemi, R. J. (1994). Interpreting magnetic
fields of the brain: Minimum norm estimates. Medical and Bio-
logical Engineering and Computing, 32(1), 35–42. https://doi.org
/10.1007/BF02512476, PubMed: 8182960
Hanakawa, T., Dimyan, M. A., & Hallett, M. (2008). Motor plan-
ning, imagery, and execution in the distributed motor network:
A time-course study with functional MRI. Cerebral Cortex, 18(12),
2775–2788. https://doi.org/10.1093/cercor/ bhn036, PubMed:
18359777
Hari, R., Forss, N., Avikainen, S., Kirveskari, E., Salenius, S., &
Rizzolatti, G. (1998). Activation of human primary motor cortex
during action observation: A neuromagnetic study. Proceedings
of the National Academy of Sciences, 95(25), 15061–15065.
https://doi.org/10.1073/pnas.95.25.15061, PubMed: 9844015
Hauk, O., & Tschentscher, N. (2013). The body of evidence: What
can neuroscience tell us about embodied semantics? Frontiers in
Psychology, 4, 50. https://doi.org/10.3389/fpsyg.2013.00050,
PubMed: 23407791
Henson, R. (2006). Forward inference using functional neuroimag-
ing: Dissociations versus associations. Trends in Cognitive Sci-
ences, 10(2), 64–69. https://doi.org/10.1016/j.tics.2005.12.005,
PubMed: 16406759
Hétu, S., Grégoire, M., Saimpont, A., Coll, M., Eugène, F., Michon,
P., & Jackson, P. L. (2013). The neural network of motor imagery:
An ALE meta-analysis. Neuroscience and Biobehavioral Reviews,
37(5), 930–949. https://doi.org/10.1016/j.neubiorev.2013.03
.017, PubMed: 23583615
Kargieman, L., Herrera, E., Baez, S., García, A. M., Dottori, M.,
Gelormini, C., Manes, F., Gershanik, O., & Ibáñez, A. (2014).
Motor-language coupling in Huntington’s disease families. Fron-
tiers in Aging Neuroscience, 6, Article 122. https://doi.org/10
.3389/fnagi.2014.00122, PubMed: 24971062
Kaschak, M. P., & Borreggine, K. L. (2008). Temporal dynamics of
the action-sentence compatibility effect. Quarterly Journal of
Experimental Psychology, 61(6), 883–895. https://doi.org/10
.1080/17470210701623852, PubMed: 18470819
Keatley, C. W., Spinks, J. A., & de Gelder, B. (1994). Asymmetrical
cross-language priming effects. Memory and Cognition, 22(1),
70–84. https://doi.org/10.3758/BF03202763, PubMed: 8035687
Kemmerer, D., Rudrauf, D., Manzel, K., & Tranel, D. (2012).
Behavioral patterns and lesion sites associated with impaired
processing of lexical and conceptual knowledge of actions.
Neurobiology of Language
195
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
Cortex, 48(7), 826–848. https://doi.org/10.1016/j.cortex.2010.11
.001, PubMed: 21159333
Klepp, A., Niccolai, V., Buccino, G., Schnitzler, A., & Biermann-
Ruben, K. (2015). Language-motor interference reflected in
MEG beta oscillations. NeuroImage, 109, 438–448. https://doi
.org/10.1016/j.neuroimage.2014.12.077, PubMed: 25576646
Klepp, A., Weissler, H., Niccolai, V., Terhalle, A., Geisler, H.,
Schnitzler, A., & Biermann-Ruben, K. (2014). Neuromagnetic
hand and foot motor sources recruited during action verb pro-
cessing. Brain and Language, 128(1), 41–52. https://doi.org/10
.1016/j.bandl.2013.12.001, PubMed: 24412808
Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting: Find-
ing meaning in the N400 component of the event-related brain
potential (ERP). Annual Review of Psychology, 62, 621–647.
https://doi.org/10.1146/annurev.psych.093008.131123, PubMed:
20809790
Lambon Ralph, M. A., Jefferies, E., Patterson, K., & Rogers, T. T.
(2017). The neural and computational bases of semantic cogni-
tion. Nature Reviews Neuroscience, 18(1), 42–55. https://doi.org
/10.1038/nrn.2016.150, PubMed: 27881854
Lau, E. F., Gramfort, A., Hamalainen, M. S., & Kuperberg, G. R.
(2013). Automatic semantic facilitation in anterior temporal
cortex revealed through multimodal neuroimaging. Journal of
Neuroscience, 33(43), 17174–17181. https://doi.org/10.1523
/JNEUROSCI.1018-13.2013, PubMed: 24155321
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network for
semantics: (De)constructing the N400. Nature Reviews Neurosci-
ence, 9(12), 920–933. https://doi.org/10.1038/nrn2532, PubMed:
19020511
Lemhöfer, K., & Broersma, M. (2012). Introducing LexTALE: A quick
and valid lexical test for advanced learners of English. Behavior
Research Methods, 44, 325–343. https://doi.org/10.3758/s13428
-011-0146-0, PubMed: 21898159
Leonardo, M., Fieldman, J., Sadato, N., Campbell, G., Ibañez, V.,
Cohen, L., Deiber, M.-P., Jezzard, P., Pons, R., Turner, R., Le
Bihan, D., & Hallett, M. (1995). A functional magnetic resonance
imaging study of cortical regions associated with motor task
execution and motor ideation in humans. Human Brain
Mapping, 3(2), 83–92. https://doi.org/10.1002/hbm.460030205
Mahon, B. Z. (2015). What is embodied about cognition?
Language, Cognition and Neuroscience, 30(4), 420–429.
https://doi.org/10.1080/23273798.2014.987791, PubMed:
25914889
Mahon, B. Z., & Hickok, G. (2016). Arguments about the nature of
concepts: Symbols, embodiment, and beyond. Psychonomic
Bulletin & Review, 23(4), 941–958. https://doi.org/10.3758
/s13423-016-1045-2, PubMed: 27282991
Maieron, M., Marin, D., Fabbro, F., & Skrap, M. (2013). Seeking a
bridge between language and motor cortices: A PPI study. Fron-
tiers in Human Neuroscience, 7, Article 249. https://doi.org/10
.3389/fnhum.2013.00249, PubMed: 23761753
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical testing
of EEG- and MEG-data. Journal of Neuroscience Methods, 164(1),
177–190. https://doi.org/10.1016/j.jneumeth.2007.03.024,
PubMed: 17517438
MEGIN. (2022). TRIUX MEG system [Apparatus]. https://megin.fi
/triux-neo/
Meteyard, L., Cuadrado, S. R., Bahrami, B., & Vigliocco, G. (2012).
Coming of age: A review of embodiment and the neuroscience of
semantics. Cortex, 48(7), 788–804. https://doi.org/10.1016/j
.cortex.2010.11.002, PubMed: 21163473
Michelon, P., Vettel, J. M., & Zacks, J. M. (2006). Lateral somatoto-
pic organization during imagined and prepared movements.
Journal of Neurophysiology, 95(2), 811–822. https://doi.org/10
.1152/jn.00488.2005, PubMed: 16207787
Miller, E. K. (2000). The prefrontal cortex and cognitive control.
Nature Reviews Neuroscience, 1(1), 59–65. https://doi.org/10
.1038/35036228, PubMed: 11252769
Mofrad, F. T., Jahn, A., & Schiller, N. O. (2020). Dual function of
primary somatosensory cortex in cognitive control of language:
Evidence from resting state fMRI. Neuroscience, 446(15), 59–68.
https://doi.org/10.1016/j.neuroscience.2020.08.032, PubMed:
32866600
Mollo, G., Pulvermüller, F., & Hauk, O. (2016). Movement priming
of EEG/MEG brain responses for action-words characterizes the
link between language and action. Cortex, 74, 262–276. https://
doi.org/10.1016/j.cortex.2015.10.021, PubMed: 26706997
Monaco, E., Jost, L. B., Gygax, P. M., & Annoni, J. M. (2019).
Embodied semantics in a second language: Critical review
and clinical implications. Frontiers in Human Neuroscience,
13, 110. https://doi.org/10.3389/fnhum.2019.00110, PubMed:
30983983
Monaco, E., Jost, L. B., Lancheros, M., Harquel, S., Schmidlin, E., &
Annoni, J. M. (2021). First and second language at hand: A
chronometric transcranial-magnetic stimulation study on seman-
tic and motor resonance. Journal of Cognitive Neuroscience,
33(8), 1563–1580. https://doi.org/10.1162/jocn_a_01736,
PubMed: 34496375
Moreno, I., de Vega, M., & León, I. (2013). Understanding action
language modulates oscillatory mu and beta rhythms in the same
way as observing actions. Brain and Cognition, 82(3), 236–242.
https://doi.org/10.1016/j.bandc.2013.04.010, PubMed:
23711935
Morey, R. D., Kaschak, M. P., Díez-Álamo, A. M., Glenberg, A. M.,
Zwaan, R. A., Lakens, D., Ibáñez, A., García, A., Gianelli, C.,
Jones, J. L., Madden, J., Alifano, F., Bergen, B., Bloxsom, N. G.,
Bub, D. N., Cai, Z. G., Chartier, C. R., Chatterjee, A., Conwell, E.,
… Ziv-Crispel, N.
(2022). A pre-registered, multi-lab
non-replication of the action-sentence compatibility effect
(ACE). Psychonomic Bulletin & Review, 29(2), 613–626. https://
doi.org/10.3758/s13423-021-01927-8, PubMed: 34755319
Neurobehavioral Systems. (2022). Presentation [Software]. https://
www.neurobs.com/
Papeo, L., Negri, G., Zadini, A., & Rumiati, R. I. (2010). Action per-
formance and action-word understanding: Evidence of double
dissociations in left-damaged patients. Cognitive Neuropsychol-
ogy, 27(5), 428–461. https://doi.org/10.1080/02643294.2011
.570326, PubMed: 21718215
Papeo, L., Vallesi, A., Isaja, A., & Rumiati, R. I. (2009). Effects of TMS
on different stages of motor and non-motor verb processing in the
primary motor cortex. PLOS ONE, 4(2), Article e4508. https://doi
.org/10.1371/journal.pone.0004508, PubMed: 19240793
Papesh, M. H. (2015). Just out of reach: On the reliability of the
action-sentence compatibility effect. Journal of Experimental Psy-
chology: General, 144(6), e116–e141. https://doi.org/10.1037
/xge0000125, PubMed: 26595844
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you
know what you know? The representation of semantic knowl-
edge in the human brain. Nature Reviews Neuroscience, 8(12),
976–987. https://doi.org/10.1038/nrn2277, PubMed: 18026167
Poldrack, R. A. (2006). Can cognitive processes be inferred from
neuroimaging data? Trends in Cognitive Sciences, 10(2), 59–63.
https://doi.org/10.1016/j.tics.2005.12.004, PubMed: 16406760
Porro, C. A., Francescato, M. P., Cettolo, V., Diamond, M. E.,
Baraldi, P., Zuiani, C., Bazzocchi, M., & di Prampero, P. E.
(1996). Primary motor and sensory cortex activation during
Neurobiology of Language
196
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Compensatory role of the motor cortex in second language processing
motor performance and motor imagery: A functional magnetic
resonance study. Journal of Neuroscience, 16(23), 7688–7698.
https://doi.org/10.1523/ JNEUROSCI.16-23-07688.1996,
PubMed: 8922425
Pulvermüller, F., & Fadiga, L. (2010). Active perception: Sensorimo-
tor circuits as a cortical basis for language. Nature Reviews
Neuroscience, 11(5), 351–360. https://doi.org/10.1038/nrn2811,
PubMed: 20383203
Pulvermüller, F., Hauk, O., Nikulin, V. V., & Ilmoniemi, R. J. (2005).
Functional links between motor and language systems. European
Journal of Neuroscience, 21(3), 793–797. https://doi.org/10.1111
/j.1460-9568.2005.03900.x, PubMed: 15733097
Raposo, A., Moss, H. E., Stamatakis, E. A., & Tyler, L. K. (2009).
Modulation of motor and premotor cortices by actions, action
words and action sentences. Neuropsychologia, 47(2), 388–396.
https://doi.org/10.1016/j.neuropsychologia.2008.09.017,
PubMed: 18930749
Reilly, M., Howerton, O., & Desai, R. H. (2019). Time-course of
motor involvement in literal and metaphoric action sentence pro-
cessing: A TMS study. Frontiers in Psychology, 10, 371. https://doi
.org/10.3389/fpsyg.2019.00371, PubMed: 30863346
Repetto, C., Colombo, B., Cipresso, P., & Riva, G. (2013). The
effects of rTMS over the primary motor cortex: The link between
action and language. Neuropsychologia, 51(1), 8–13. https://doi
.org/10.1016/j.neuropsychologia.2012.11.001, PubMed:
23142706
Richlan, F., Kronbichler, M., & Wimmer, H. (2011). Meta-analyzing
brain dysfunctions in dyslexic children and adults. NeuroImage,
56(3), 1735–1742. https://doi.org/10.1016/j.neuroimage.2011
.02.040, PubMed: 21338695
Rueckl, J. G., Paz-Alonso, P. M., Molfese, P. J., Kuo, W.-J., Bick, A.,
Frost, S. J., Hancock, R., Wu, D. H., Mencl, W. E., Duñabeitia,
J. A., Lee, J.-R., Oliver, M., Zevin, J. D., Hoeft, F., Carreiras, M.,
Tzeng, O. J. L, Pugh, K. R., & Frost, R. (2015). Universal brain
signature of proficient reading: Evidence from four contrasting
languages. Proceedings of the National Academy of Sciences,
11 2 ( 5 0 ) , 1 5 5 1 0– 1 5 5 1 5. h t t p s : / / d o i . o rg / 1 0 . 1 0 7 3/ p na s
.1509321112, PubMed: 26621710
Santana, E., & de Vega, M. (2011). Metaphors are embodied, and
so are their literal counterparts. Frontiers in Psychology, 2, 90.
h t t p s : / / d o i . o r g / 1 0 . 3 3 8 9 / f p s y g . 2 0 11 . 0 0 0 9 0 , P u b M e d :
21687459
Schaller, F., Weiss, S., & Müller, H. M. (2017). EEG beta-power
changes reflect motor involvement in abstract action language
processing. Brain and Language, 168, 95–105. https://doi.org
/10.1016/j.bandl.2017.01.010, PubMed: 28189047
Smith, Y., Walters, J., & Prior, A. (2019). Target accessibility contrib-
utes to asymmetric priming in translation and cross-language
semantic priming in unbalanced bilinguals. Bilingualism: Lan-
guage and Cognition, 22(1), 157–176. https://doi.org/10.1017
/S1366728917000645
Taulu, S., & Kajola, M. (2005). Presentation of electromagnetic mul-
tichannel data: The signal space separation method. Journal of
Applied Physics, 97, Article 124905. https://doi.org/10.1063/1
.1935742
Tian, L., Chen, H., Zhao, W., Wu, J., Zhang, Q., De, A., Leppänen,
P., Cong, F., & Parviainen, T. (2020). The role of motor system in
action-related language comprehension in L1 and L2: An fMRI
study. Brain and Language, 201, Article 104714. https://doi.org
/10.1016/j.bandl.2019.104714, PubMed: 31790907
Tomasino, B., Fink, G. R., Sparing, R., Dafotakis, M., & Weiss, P. H.
(2008). Action verbs and the primary motor cortex: A compara-
tive tms study of silent reading, frequency judgments, and motor
imagery. Neuropsychologia, 46(7), 1915–1926. https://doi.org/10
.1016/j.neuropsychologia.2008.01.015, PubMed: 18328510
Ullman, M. T. (2004). Contributions of memory circuits to lan-
guage: The declarative/procedural model. Cognition, 92(1–2),
231–270. https://doi.org/10.1016/j.cognition.2003.10.008,
PubMed: 15037131
van Elk, M., van Schie, H. T., Zwaan, R. A., & Bekkering, H. (2010).
The functional role of motor activation in language processing:
Motor cortical oscillations support lexical-semantic retrieval.
NeuroImage, 50(2), 665–677. https://doi.org/10.1016/j
.neuroimage.2009.12.123, PubMed: 20060478
Visser, M., Jefferies, E., & Lambon Ralph, M. A. (2010). Semantic
processing in the anterior temporal lobes: A meta-analysis of
the functional neuroimaging literature. Journal of Cognitive Neu-
roscience, 22(6), 1083–1094. https://doi.org/10.1162/jocn.2009
.21309, PubMed: 19583477
Vukovic, N., Feurra, M., Shpektor, A., Myachykov, A., & Shtyrov, Y.
(2017). Primary motor cortex functionally contributes to lan-
guage comprehension: An online rTMS study. Neuropsychologia,
96, 222–229. https://doi.org/10.1016/j.neuropsychologia.2017
.01.025, PubMed: 28122198
Vukovic, N., & Shtyrov, Y. (2014). Cortical motor systems are
involved in second-language comprehension: Evidence from
rapid mu-rhythm desynchronization. NeuroImage, 102(2),
695–703. https://doi.org/10.1016/j.neuroimage.2014.08.039,
PubMed: 25175538
Wang, L., Hagoort, P., & Jensen, O. (2018). Language prediction is
reflected by coupling between frontal gamma and posterior
alpha oscillations. Journal of Cognitive Neuroscience, 30(3),
432–447. https://doi.org/10.1162/jocn_a_01190, PubMed:
28949823
Willems, R. M., Toni, I., Hagoort, P., & Casasanto, D. (2010). Neural
dissociations between action verb understanding and motor
imagery. Journal of Cognitive Neuroscience, 22(10), 2387–2400.
https://doi.org/10.1162/jocn.2009.21386, PubMed: 19925195
Yang, G., Zhou, S., Bozek, J., Dong, H.-M., Han, M., Zuo, X.-N.,
Liu, H., & Gao, J.-H. (2020). Sample sizes and population differ-
ences in brain template construction. NeuroImage, 206, Article
116318. https://doi.org/10.1016/j.neuroimage.2019.116318,
PubMed: 31689538
Yousry, T. A., Schmid, U. D., Alkadhi, H., Schmidt, D., Peraud, A.,
Buettner, A., & Winkler, P. (1997). Localization of the motor hand
area to a knob on the precentral gyrus: A new landmark. Brain,
120(1), 141–157. https://doi.org/10.1093/ brain/120.1.141,
PubMed: 9055804
Zhang, X., Yang, J., Wang, R., & Li, P. (2020). A neuroimaging study
of semantic representation in first and second languages.
Language, Cognition and Neuroscience, 35(10), 1223–1238.
https://doi.org/10.1080/23273798.2020.1738509
Zwaan, R. A. (2014). Embodiment and language comprehension:
Reframing the discussion. Trends in Cognitive Sciences, 18(5),
229–234. https://doi.org/10.1016/j.tics.2014.02.008, PubMed:
24630873
Zwaan, R. A., & Taylor, L. J. (2006). Seeing, acting, understanding:
Motor resonance in language comprehension. Journal of Experi-
mental Psychology: General, 135(1), 1–11. https://doi.org/10
.1037/0096-3445.135.1.1, PubMed: 16478313
Neurobiology of Language
197
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
1
1
7
8
2
0
7
4
4
6
7
n
o
_
a
_
0
0
0
9
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3