RESEARCH ARTICLE
Cerebral White Matter Mediation of Age-Related
Differences in Picture Naming Across Adulthood
Sara B. W. Troutman1
, David J. Madden2,3
, and Michele T. Diaz1,4
1Abteilung für Psychologie, Pennsylvania State University, University Park, PA, USA
2Brain Imaging and Analysis Center & Department of Psychiatry and Behavioral Sciences,
Duke University Medical Center, Durham, NC, USA
3Center for Cognitive Neuroscience, Duke University, Durham, NC, USA
4Sozial, Life, & Engineering Sciences Imaging Center, Pennsylvania State University, University Park, PA, USA
Schlüsselwörter: diffusion tensor imaging, picture naming, Altern, language production
ABSTRAKT
As people age, one of the most common complaints is difficulty with word retrieval. A wealth of
behavioral research confirms such age-related language production deficits, yet the structural
neural differences that relate to age-related language production deficits remains an open area
of exploration. daher, the present study used a large sample of healthy adults across
adulthood to investigate how age-related white matter differences in three key left-hemisphere
language tracts may contribute to age-related differences in language ability. Speziell, Wir
used diffusion tensor imaging to measure fractional anisotropy (FA) and radial diffusivity (RD)
which are indicators of white matter structure. We then used a series of path models to test
whether white matter from the superior longitudinal fasciculus (SLF), the inferior longitudinal
fasciculus, and the frontal aslant tract (FAT) mediated age-related differences in one form of
language production, picture naming. We found that FA, as well as RD from the SLF and FAT
mediated the relation between age and picture naming performance, whereas a control tract
(corticospinal) was not a mediator. Darüber hinaus, differences between mediation of picture naming
and a control naming condition suggest that left SLF has a greater role in higher-order aspects of
Wir, such as semantic and lexical selection whereas left FAT is more sensitive to
sensorimotor aspects of fluency or speech motor planning. These results suggest that dorsal
white matter contributes to age-related differences in generating speech and may be
particularly important in supporting word retrieval across adulthood.
EINFÜHRUNG
One of the most common age-related complaints is word retrieval difficulty (Ossher et al.,
2013). Such age-related challenges in language production are well documented (Burke &
Shafto, 2008): Compared to younger cohorts, older adults produce speech more slowly
(Gollan et al., 2008), make more errors (Feyereisen, 1997), use more filler words and pauses
(Kemper et al., 1992), produce more off-topic speech (Arbuckle & Gold, 1993), and experience
more tip-of-the-tongue events and word finding difficulties (Burke et al., 1991). Behavioral
research suggests that age-related deficits in language production may be related to deficits in
accessing the sounds of words. In the context of the transmission deficit hypothesis (Burke et al.,
1991), age-related weakening among phonological connections may underlie language pro-
duction deficits. Although recent functional magnetic resonance imaging (MRT) research found
similar neural and behavioral sensitivity to phonological characteristics for younger and older
Keine offenen Zugänge
Tagebuch
Zitat: Troutman, S. B. W., Madden,
D. J., & Diaz, M. T. (2022). Zerebral
white matter mediation of age-related
differences in picture naming across
Erwachsensein. Neurobiology of Language,
3(2), 272–286. https://doi.org/10.1162
/nol_a_00065
DOI:
https://doi.org/10.1162/nol_a_00065
zusätzliche Informationen:
https://doi.org/10.1162/nol_a_00065
Erhalten: 4 Mai 2021
Akzeptiert: 2 Januar 2022
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Michele T. Diaz
mtd143@psu.edu
Handling-Editor:
Kate Watkins
Urheberrechte ©: © 2022
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
Magnetic resonance imaging (MRT):
A technique that uses strong
magnetic fields and radio frequencies
to provide images of the brain and
other parts of the body.
Diffusion tensor imaging (DTI):
An MRI sequence that provides
information regarding the rate and
directionality of water diffusion. Das
is often used as an assessment of
white matter integrity.
Fractional anisotropy (FA):
Reflects the degree to which water
diffusion is directional (anisotropic)
rather than random or equal in all
directions (isotropic). This is most
often used as an indicator of fiber
tract integrity.
Radial diffusivity (RD):
Reflects the rate of diffusivity
perpendicular to the principal
Richtung. This is often used as an
indicator of myelination.
Erwachsene, age-related differences in picture naming across adulthood were associated with
increases in functional activation (Diaz et al., 2021). Jedoch, it remains unclear how brain
structure may relate to these age-related language production deficits.
One possible neural mechanism for age-related increases in language production difficulty
is the structural degradation of cerebral white matter, which comprises the myelin-coated axo-
nal fibers conveying signals across the brain. Information regarding cerebral white matter can
be obtained by MRI through susceptibility-weighted data. Diffusion tensor imaging (DTI) Profi-
vides information regarding the rate and directionality of molecular water motion at the voxel
Ebene (Basser, 1995; Jones et al., 2013; Pierpaoli et al., 1996; A. W. Song et al., 1996; S.-K.
Song et al., 2003, 2005). Within a tensor model, Zum Beispiel, fractional anisotropy (FA) reflects
the degree to which diffusivity is directional rather than random (isotropic), and radial diffu-
sivity (RD) reflects the rate of diffusivity perpendicular to the principal direction (eigenvalue) von
the tensor. Daher, assuming that many white matter fibers contribute to the estimates obtained
from each voxel, higher spatial coherence of fibers would lead to increasing FA (d.h., higher
directionality of diffusion), and a greater hindrance to diffusion across the fibers, provided by
more complete myelination, would lead to lower RD (S.-K. Song et al., 2003, 2005). Daher,
these DTI measures are informative regarding the underlying structural integrity of white mat-
ter, despite the fact that these measures are indirect and influenced by other variables (Jones
et al., 2013; Wheeler-Kingshott & Cercignani, 2009).
Structural disconnection, specifically changes in the white matter tracts that emerge through
Altern, have been linked to age-related differences in several areas of cognition, especially in mea-
sures of executive function and processing speed (Bennett & Madden, 2014; Gazes et al., 2016;
Hedden et al., 2016; Johnson et al., 2015; Salami et al., 2012), though the degree to which white
matter has an independent or interactive effect with age is unclear (Madden et al., 2017, 2020;
Salami et al., 2012). Darüber hinaus, a number of studies have pointed to the importance of white matter
connections in language production ability (De Zubicaray et al., 2011; Dick et al., 2014; Houston
et al., 2019; Madhavan et al., 2014; Stamatakis et al., 2011; Troutman & Diaz, 2019). Studies using
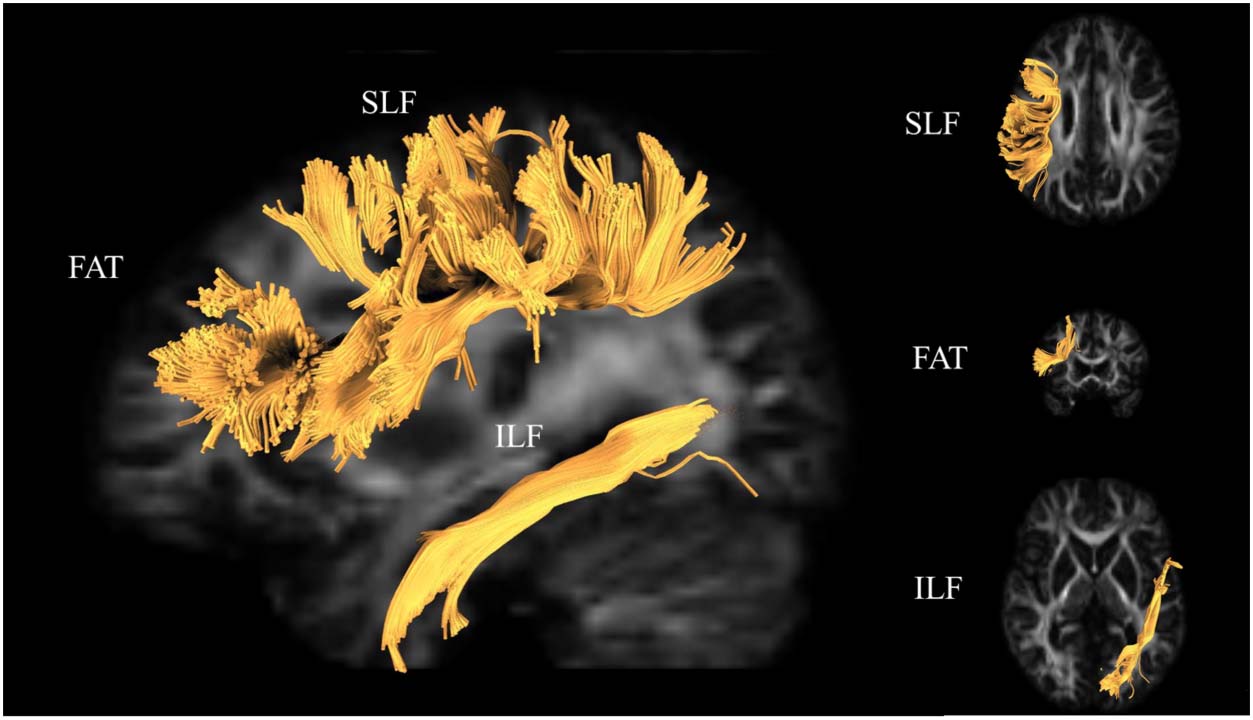
DTI-derived metrics of white matter structure have shown that white matter along several key tracts
within the language network are sensitive to age-related differences. These tracts include the supe-
rior longitudinal fasciculus-III (SLF-III; Houston et al., 2019; Madhavan et al., 2014; Stamatakis
et al., 2011; Troutman & Diaz, 2019), the inferior longitudinal fasciculus (ILF; Kantarci et al.,
2011; Stamatakis et al., 2011; Troutman & Diaz, 2019), and the frontal aslant tract (FAT; Catani
et al., 2013; Rizio & Diaz, 2016; Troutman & Diaz, 2019). It is important to note that there are
several models of these tracts (for reviews, see Dick et al., 2014; Dick & Tremblay, 2012; Friederici,
2009, 2012; Varriano et al., 2020). Zum Beispiel, the SLF-III is in close proximity to the arcuate fas-
ciculus, which also supports language (z.B., Perron et al., 2021; Tremblay et al., 2019), and there is
debate about where each terminates posteriorly (z.B., posterior parietal, posterior superior temporal
gyrus). With respect to the FAT, which connects inferior frontal gyrus regions with supplemental
motor regions (SMA & vor SMA) in superior frontal gyrus, it has recently been suggested that there
is an additional anterior portion that may play a role in working memory (Varriano et al., 2020).
Several groups have reported that, among both younger and older adults, higher FA along
the left SLF correlated with better naming performance (Houston et al., 2019; Madhavan et al.,
2014; Stamatakis et al., 2011; Troutman & Diaz, 2019; for a review see Dick et al., 2014; Dick
& Tremblay, 2012). Darüber hinaus, Stamatakis et al. (2011) found that age-related word-finding
failures were linked to lower FA, particularly in the posterior portion of the left SLF. This sug-
gests that white matter deficits, particularly along the left posterior SLF, may play a role in driv-
ing age-related language production deficits. Madhavan et al. (2014) and Houston et al. (2019)
found that lower FA across the SLF was linked to poorer performance on clinical tests of language
Neurobiology of Language
273
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
production (d.h., the Controlled Oral Word Association Test (Benton et al., 1983) and the Boston
Naming Test (Kaplan et al., 1983), jeweils). More recently, Troutman and Diaz (2019) stud-
ied white matter using both FA and RD. They found that higher FA within dorsal tracts, einschließlich
the SLF and FAT, was linked to better picture naming during a phonological picture-word inter-
ference task. While the effect of FA in their study was localized to dorsal tracts, specifically the
SLF and FAT, the effect of RD was more widespread. Language production performance was
linked to RD along dorsal language tracts (the SLF and FAT), as well as ventral language tracts
(the ILF and middle longitudinal fasciculus) and the frontostriatal tract. Das ist, participants with
higher FA along dorsal language tracts and lower RD along all tracts named pictures the most
accurately. Wichtig, effects of age and white matter shared variance in explaining behavioral
differences in picture naming, suggesting that age-related declines in white matter, insbesondere
from dorsal pathways, have a substantive role in age-related language production deficits.
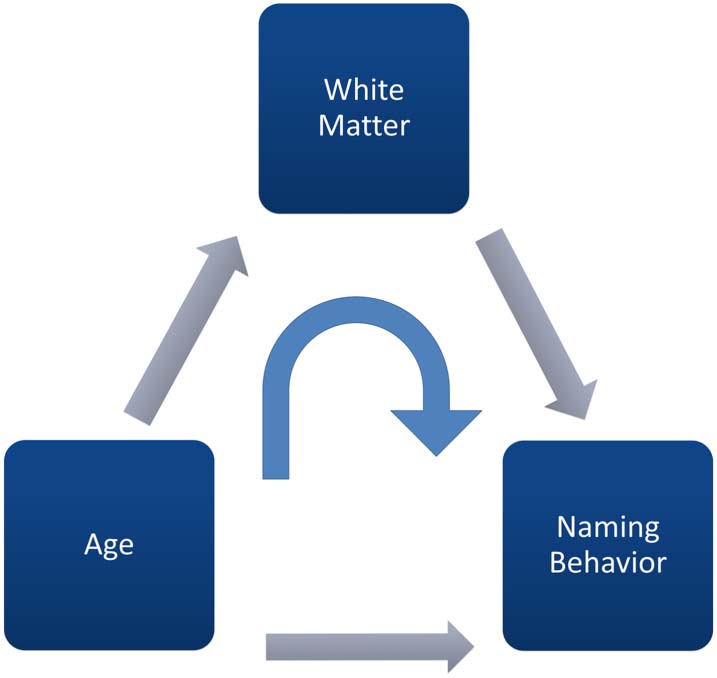
The Current Study
We sought to extend prior studies examining the relations between aging, language production,
and white matter in several ways. Erste, we took a more mechanistic approach by investigating the
influence of white matter on the relation between age and language production with mediation
analyses by building a series of path models to test the role of the SLF, FAT, ILF, and a control tract,
the corticospinal (CS) tract on naming performance. Zweite, we examined the role of white matter
and age by comparing naming under two conditions (object naming and repeating a word to an
abstract image). This allowed us to distinguish between sensorimotor aspects of naming (z.B., motor
Kontrolle, articulation) from lexical and semantic aspects of naming. Dritte, we included a broad sam-
ple of adults of different ages, who completed a picture-naming task and MRI scanning, einschließlich
diffusion-weighted imaging (DWI), to allow us to examine these relations across adulthood.
Given the prior literature, we hypothesized that dorsal language tracts (SLF, FAT) would be
most important for selection and retrieval aspects of picture naming, and that FA would dis-
criminate naming performance between picture naming and our control condition. Weil
previous research has found effects of RD on picture naming within both dorsal and ventral
language tracts, we hypothesized that RD might be sensitive to naming in general (d.h., across
Bedingungen). We expected higher FA (d.h., more cohesive white matter) and lower RD (d.h.,
more myelinated tracts) would predict faster and more accurate picture naming. We also
expected that age would be negatively correlated with FA and positively correlated with
RD, reflecting well-documented age-related deficits in white matter. Critically, we hypothe-
sized a mediating path, whereby the effect of dorsal white matter would mediate the effect
of age on picture naming, such that increases in FA and decreases in RD would be associated
with better picture naming performance.
MATERIALS AND METHODS
Teilnehmer
Ninety-three healthy, right-handed, monolingual, native American-English speaking adults
participated in this study. Participants were community-dwelling adults, recruited from cam-
pus and the local area via radio ads and flyers. Data from one participant were removed
because of poor picture naming performance (>3 SDs from the mean) and a second partici-
pant’s data were removed due to a high score on our depression screening (Geriatric Depres-
sion Scale; Guerin et al., 2018; Sheikh & Yesavage, 1986). The final sample included data from
91 participants aged 20–75 (Durchschnittsalter = 47.40 Jahre, SD = 17.45 Jahre), 54 weiblich, 37 männlich.
All participants had normal or corrected-to-normal vision as indicated by the Freiburg Visual
Neurobiology of Language
274
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
Acuity Test (Bach, 1996). Participants reported no history of neurological, psychologisch, oder
major medical conditions nor did they report any contraindications to MRI scanning (z.B., hav-
ing a pacemaker; Christensen et al., 1992). Participants also completed a battery of psycho-
metric and neuropsychological tests to assess cognitive functions such as speed, working
Erinnerung, executive function, and language. Participant demographics and cognitive scores
are reported in Table 1. A detailed description of the battery is available in Diaz et al.
(2021), where results from the fMRI analysis are also reported. All procedures were approved
by the Pennsylvania State University Institutional Review Board, and all participants provided
written, informed consent.
Tisch 1.
Participant demographic and neuropsychological testing scores
Demographic information
N
Alter (Jahre)
Gender (M/F)
Education (Jahre)
Cognitive assessments–age correlation
Education
MMSE
Depression (GDS)
Speed RT (Auswahl)
WAIS-III digit symbol RT
WAIS-III digit span forward
WAIS-III digit span backward
Stroop effect
Verbal working memory
CVLT immediate recall
CVLT delayed recall
Category fluency (Tiere)
Phonemic fluency (F, A, S)
WAIS-III vocabulary
Author recognition task
91
47.40 (20–75, 17.45)
37/54
16.9 (12–25, 2.5)
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
0.24*
−0.19
−0.13
0.56***
0.69***
−0.20
−0.26*
0.33***
−0.40***
−0.27*
−0.26*
−0.30**
−0.11
0.07
0.47***
Notiz. The color Stroop task was used; the Stroop effect was the difference in reaction times (RT) between word-
font incongruent minus word-font congruent trials. The author recognition task (Acheson et al., 2008) Verwendet
author name recognition to assess reading habits, which may be less biased compared to self-report. Demo-
graphic information for age and education is the mean, with range and standard deviation in parentheses.
MMSE = Mini-Mental State Exam (Folstein et al., 1975); GDS = Geriatric Depression Scale (Guerin et al.,
2018; Sheikh & Yesavage, 1986); WAIS-III = Wechsler Adult Intelligence Scale (Wechsler, 1997); CVLT =
California Verbal Learning Test (Delis et al., 1987). *P < 0.05, **p < 0.01, ***p < 0.001.
Neurobiology of Language
275
White matter mediation in picture naming
Stimuli and Procedure
During their functional MRI scan, participants named a series of pictures and said “picture” to
abstract images (191 color photographs of objects and 50 diffeomorphically scrambled control
images (Stojanoski & Cusack, 2014), 396 pixels × 396 pixels, duration = 1,500 ms). Photo-
graphs were of everyday objects (e.g., animals, fruit, vehicles, household objects, etc.), and
were largely selected from normed databases (Brodeur et al., 2010, 2014; Moreno-Martinez
& Montoro, 2012). All photographs underwent a separate norming procedure with a different
group of younger participants (N = 28; mean age = 19.48, SD = 1.37; 16 females) to ensure
high name agreement (mean H-index = 0.25, SD = 0.40, range = 0–1.55; H-index is a measure
of name agreement that accounts for the variability in acceptable names given by participants
for a particular image by considering both the number of acceptable names and their propor-
tions [Snodgrass & Vanderwart, 1980]; lower H-index values correspond to higher name
agreement). Photographs were presented in a random order with a variable inter-stimulus
interval (range = 1.5–15 s, mean = 3.40 s) to optimize the hemodynamic response (Dale,
1999). Responses were recorded and filtered using an MR-compatible, dual-channel, fiber-
optic microphone system (Optoacoustics Ltd., Or-Yehuda, Israel). For additional details on
the stimuli and task procedures see Diaz et al. (2021).
Behavioral Data Analyses
Responses to the photographs of objects were considered to be correct if the participant pro-
vided the anticipated picture name (e.g., airplane for airplane), the plural form of an otherwise
correct name, or an acceptable alternative (e.g., jet for airplane). Across all participants, this
method yielded a total of 17,381 observations before data cleaning, including 12,698 modal
answers, 1,629 acceptable alternatives, and 3,054 incorrect answers. In cases where partici-
pants provided an acceptable alternative, word characteristics such as length and frequency
were calculated for the provided response. Alternate names were, on average, more frequent,
shorter, and had more phonological neighbors.
For naming latencies, only correct trials as defined above were considered. To calculate
naming latencies we used custom scripts in PRAAT (Boersma, 2001). These scripts used pitch
and intensity cues to identify potential word onsets, which were manually verified by trained
research assistants (additional details can be found in the Supporting Information at https://doi
.org/10.1162/nol_a_00065). Then, we calculated naming latencies by taking the difference
between the stimulus onset and the onset of the participant’s response. We excluded trials
where the naming latency was implausibly low or not recorded (i.e., >200 MS; n = 219) oder
if the naming latency constituted an outlier (d.h., greater than or less than 2.5 SDs from that
participant’s mean naming latency or mean accuracy; n = 636). Daher, there were a total of 855
excluded outliers (∼4.92% of total responses), leaving a final data set of 13,472 total picture
naming observations.
In order to minimize the influence of speed-accuracy trade-offs in the analysis, we calcu-
lated an inverse efficiency score (IES) at the participant level (Horowitz & Wolfe, 2003). Wir
constructed the IESs by dividing each participant’s average naming latency by their average
accuracy. Similar to naming latencies, a higher IES indicates slower naming while a lower IES
indicates faster naming, but an IES adjusts latency by the associated accuracy. Responses to
control images were considered correct if participants replied “picture” to the scrambled
image as per the instructions. In Summe, participants accurately identified 4,437 scrambled
Bilder. Trials with implausible or no response times (d.h., <200 ms, n = 138) and subject-level
Neurobiology of Language
276
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
>0.9; Tucker–Lewis
index [TLI] >0.90; root mean square error of approximation [RMSEA] <0.08; see Supplemental
Table 2 for details; Hooper et al., 2008). To minimize the influence of false positives, we used a
Bonferroni-corrected significance threshold of p = 0.006 (i.e., 0.05 / 8 = 0.006). To compare
the strength of relationships between paths, we used Fisher r to z transformations.
RESULTS
Behavioral Results
Consistent with age-related slowing, IESs were positively correlated with age (t = 36.74, p <
0.0001). There was no significant main effect of age on naming time. However, age was asso-
ciated with lower accuracies (t = −2.92, p < 0.005, eta squared = −0.08), and there was a
marginally significant quadratic effect of age on accuracy (p = 0.08), with the strongest effect
of age on accuracy among the oldest adults. Thus, the age-related differences in IESs were
most likely driven by age-related differences in accuracy.
Fractional Anisotropy
Consistent with age-related slowing, IESs were positively correlated with age (t = 36.74, p <
0.0001); for additional details see Supporting Information. With respect to the DTI measures,
simple bivariate correlations confirmed that increases in FA were related to faster naming. In
the path models, age was negatively related to FA from all tracts: the SLF (β = −0.092, SE =
0.009, p < 0.001), ILF (β = −0.131, SE = 0.009, p < 0.001), FAT (β = −0.073, SE = 0.01, p <
0.001), and CS (β = −0.030, SE = 0.01, p = 0.002). Picture naming latency increased with age
in most models: SLF (β = 62.89, SE = 1.56, p < 0.001), ILF (β = 31.95, SE = 1.73, p < 0.001),
FAT (β = −0.073, SE = 0.009, p < 0.001), and CS (β = 56.33, SE = 1.65, p < 0.001). See Table 2
for model details.
Table 2.
Path model estimates for the relationship between age, FA, and naming
Naming pictures of everyday objects
Age–FA
Age–IES
FA–IES
FA mediation of Age–IES
Naming abstract control items
FA–IES
FA mediation of Age–IES
SLF
−0.092 (0.009)*
62.89 (1.56)*
13.55 (1.51)*
−1.26 (0.19)*
SLF
−2.76 (1.66)
0.25 (0.16)
ILF
−0.131 (0.009)*
31.95 (1.73)*
−0.004 (1.53)
FAT
−0.073 (0.01)*
−0.073 (0.01)*
−9.64 (1.63)*
0.099 (0.20)
0.71 (0.15)*
ILF
0.29 (1.76)
−0.04 (0.25)
FAT
24.78 (1.30)*
−2.02 (0.44)*
CS
−0.030 (0.01)*
56.33 (1.65)*
−7.16 (1.62)*
−0.93 (0.46)
CS
−7.78 (1.80)*
0.23 (0.15)
Note. Values provided are beta estimates with standard errors in parentheses. *p < 0.006, for statistically significant relationships, the significance threshold
accounts for multiple comparisons.
Neurobiology of Language
279
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
n
o
_
a
_
0
0
0
6
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
White matter mediation in picture naming
FA from the SLF, FAT, and CS were related to picture naming latency (β = 13.55, SE = 1.51,
p < 0.001; β = −9.64, SE = 1.63, p < 0.001; β = −7.16, SE = 1.62, p < 0.001). Moreover, FA
from the SLF and the FAT mediated the relation between age and naming latency (β = −1.26,
SE = 0.19, p < 0.001; β = 0.71, SE = 0.15, p < 0.001). FA from the CS did not (p = 0.01).
Importantly, the relationships between picture naming and SLF FA and picture naming and
FAT FA were significantly stronger than the corresponding effect of FA from the CS control tract
(z = 7.092, p < 0.001; z = 2.810, p = 0.005). There were no significant relationships between
picture naming and FA from the ILF (β = −0.004, SE = 1.53, p = 0.613), nor did this mediate
the relationship between age and picture naming.
The role of FA on the control condition (i.e., saying “picture” in response to abstract images)
revealed that FAT FA was positively related to naming the control trials (β = 24.78, SE = 1.30,
p < 0.001) and FAT FA mediated the relationship between age and naming control trials (β =
−2.02, SE = 0.44, p < 0.001). Though, CS FA was related to naming latency (β = −7.78, SE =
1.80, p < 0.001), the relationship between age and naming control trials was not mediated
by FA from the CS (p = 0.124). Moreover, the mediating effect of FAT FA was significantly
greater than the corresponding relationship between CS FA and naming latency (z = 4.875,
p < 0.001). FA from the SLF and ILF did not show significant relationships with the control
condition naming latency (ps > 0.10), nor were these significant mediators (ps > 0.09).
Radial Diffusivity
Simple bivariate correlations indicated that increases in RD were associated with slower
naming times. In the path models, age was positively related to RD from all models: Die
SLF (β = 0.37, SE = 0.009, P < 0.001), ILF (β = 0.08, SE = 0.009, p < 0.001), FAT (β =
0.42, SE = 0.009, p < 0.001), and CS (β = 0.28, SE = 0.01, p < 0.001). See Table 3 for
model details. Picture naming latency increased with age in all models: SLF (β = 66.96,
SE = 1.70, p < 0.001), ILF (β = 52.42, SE = 1.52, p < 0.001), FAT (β = 60.09, SE = 1.70,
p < 0.001), and CS (β = 52.53, SE = 1.73, p < 0.001).
RD from the SLF (β = −14.09, SE = 1.62, p < 0.001) and FAT (β = −18.37, SE = 1.66, p <
0.001) were negatively related to picture naming latency. RD of both the SLF (β = −5.16, SE =
Table 3.
Path model estimates for the relationship among age, RD, and naming
Naming pictures of everyday objects
Age–RD
Age–IES
RD–IES
RD mediation of Age–IES
Naming abstract control items
RD–IES
RD mediation of Age–IES
SLF
0.37 (0.009)*
ILF
0.08 (0.009)*
FAT
0.42 (0.009)*
CS
0.28 (0.01)*
66.96 (1.70)*
52.42 (1.52)*
60.09 (1.70)*
52.53 (1.73)*
−14.09 (1.62)*
−5.16 (0.61)*
2.84 (1.54)
0.22 (0.12)
−18.37 (1.66)*
−7.68 (0.71)*
−3.37 (1.67)
−0.93 (0.46)
SLF
9.39 (1.78)*
3.43 (0.67)*
ILF
−4.97 (1.77)*
−0.50 (0.20)
FAT
−13.88 (1.96)*
−5.83 (0.81)*
CS
2.04 (1.87)
0.57 (0.52)
Note. Values provided are beta estimates with standard errors in parentheses. *p < 0.006, for statistically significant relationships, the significance threshold
accounts for multiple comparisons.
Neurobiology of Language
280
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
n
o
_
a
_
0
0
0
6
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
White matter mediation in picture naming
0.61, p < 0.001) and FAT (β = −7.68, SE = 0.71, p < 0.001) also mediated the relationship
between age and picture naming latency. Moreover, for the SLF and FAT, the mediating rela-
tionship between naming latencies and RD was significantly stronger than the corresponding
path in the CS model (z = 5.568, p < 0.001; z = 7.961, p < 0.001, respectively). RD from the
ILF and CS were not significantly related to naming latency and did not mediate the age-
naming latency relationship (ps > 0.01).
Tests of the role of RD on naming control trials revealed that RD from the SLF, ILF, and FAT
were related to naming latency (β = 9.39, SE = 1.78, P < 0.001; β = −4.97, SE = 1.77 p = 0.005; β =
−13.88, SE = 1.96, p < 0.001, respectively) and that SLF and FAT RD mediated the relationship
between age and naming control trials (β = 3.43, SE = 0.67, p < 0.001; β = −5.83, SE = 0.81,
p < 0.001, respectively), though ILF RD did not (p = 0.01). Again, the mediating paths
between SLF and FAT RD and naming control trials were significantly stronger than the cor-
responding path in the CS model (z = 3.527, p < 0.001, z = 6.849, p < 0.001, respectively).
The CS RD was not significantly related to naming control trials, nor did it significantly medi-
ate the relationship between age and naming (ps > 0.1).
DISKUSSION
A wealth of behavioral research has shown that as we age, word retrieval difficulties frustrat-
ingly increase; Jedoch, the neural bases of these behaviors are less well understood. Eins
potential neural factor underlying age-related differences in language production is white
matter. Hier, we tested how white matter relates to age-related language production differ-
ences in picture naming by examining DTI-derived metrics that are sensitive to white matter
Struktur. Based on the prior literature, we hypothesized that behaviorally, older adults would
have slower and less accurate naming ability. Measuring picture naming with a combined
estimate of speed and accuracy (IES), we found that, across all the path models, age was pos-
itively correlated with IES values (d.h., increased age was associated with slower accuracy-
adjusted naming times). With respect to neural measures and in line with prior literature,
we hypothesized that older adults would have lower white matter integrity. Consistent with
our hypothesis, we found that age was negatively correlated with FA and positively correlated
with RD in the left hemisphere language tracts that we measured (SLF, ILF, and FAT), reflecting
age-related white matter deficits in the spatial coherence of estimated fibers (FA) and diffusion
across the fibers (RD).
Most importantly, we hypothesized that left-hemisphere dorsal language tracts (SLF, FAT)
would have a stronger role in picture naming compared to our control naming condition,
and that these white matter tracts would mediate age-related differences in picture naming.
Consistent with this, our analysis showed that FA and RD from dorsal language tracts (SLF
and FAT) mediated age-related language production deficits in picture naming. Darüber hinaus,
these significant mediation effects were stronger than the corresponding relations within the
CS control tract, which were nonsignificant. The significant mediation effects suggest that
white matter connections within dorsal portions of the language network contribute to age-
related differences in language production over and above chronological age.
Wichtig, FA within the FAT and RD in the SLF, ILF, and FAT also mediated the relation-
ship between chronological age and naming latency of a control condition that placed less
demand on word retrieval systems (d.h., saying “picture” to an abstract image). Gesamt, diese
findings are consistent with the prior literature (Houston et al., 2019; Madhavan et al., 2014;
Stamatakis et al., 2011; Troutman & Diaz, 2019) suggesting that dorsal white matter tracts play
a particularly important role in age-related language production deficits. Darüber hinaus, Die
Neurobiology of Language
281
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
differentiation between picture naming and control trial naming suggests that the SLF has a
greater role in higher-order aspects of naming, such as semantic and lexical selection. Das
is consistent with previous work that has linked posterior regions of the SLF to word retrieval
failures (Stamatakis et al., 2011).
Interessant, the FAT’s sensitivity to both picture naming and control trial naming highlights
its importance for speech in general, but it is perhaps more closely related to sensorimotor
aspects of speech motor planning or fluency. This is consistent with Catani et al.’s (2013) clin-
ical work that showed greater FAT atrophy among individuals with the nonfluent variety of
primary progressive aphasia compared to individuals with the semantic variant of primary pro-
gressive aphasia, which is more closely linked to semantic deficits as opposed to naming dif-
ficulty per se. Darüber hinaus, verbal fluency ability across their entire sample (including healthy
Kontrollen) was significantly correlated with fiber structure of the FAT. Ähnlich, Dick et al.
(2019) have suggested that the left FAT may support speech motor control and may be
involved in motor plan selection during speech. Likewise, in their study of picture-word inter-
ference in healthy adults, Troutman and Diaz (2019) found that higher FA and lower RD from
dorsal tracts, including the FAT, were linked to better naming performance in younger and
older adults. Results from the present study build upon previous literature by providing a direct
test of the role of dorsal white matter as a mechanism of age-related deficits in speech produc-
tion and suggest that the left SLF may be more directly related to lexical or semantic selection,
while the left FAT has a role in fluency and speech motor planning.
Critically, the observed mediation effects were specific to language-relevant tracts, Und
white matter from the CS, as measured by either FA or RD, did not mediate the relationship
between age and picture naming behavior. Post hoc comparisons confirmed that the signifi-
cant mediating paths (z.B., between picture naming and SLF and FAT FA and RD) were sig-
nificantly different from the nonsignificant effects in the control CS path. Daher, age-related
deficits in language white matter tracts have specific cognitive consequences for language pro-
duktion. Some prior studies of cognitive aging have suggested that age-related changes in the
properties of white matter have relatively global relations to cognition, particularly executive
function and processing speed (Gazes et al., 2016; Hedden et al., 2016; Johnson et al., 2015;
Madden et al., 2017, 2020; Salami et al., 2012). Although our results do not speak to cognitive
effects more broadly, we were able to differentiate white matter contributions to naming pic-
tures vs. abstract items, and also to differentiate our results within left-hemisphere language
tracts vs. a CS control tract. If these effects were due to general effects of cognition, we might
have expected less differentiation across conditions and regions in our results.
Related to the discussion of global vs. local effects, we also observed greater specificity in
mediating effects of picture naming in our FA measures, as compared to the RD measures. Das
suggests that RD may be more closely related to sensorimotor aspects of speech production, als
opposed to picture naming specifically.
Natürlich, this study only focused on one aspect of language production (Wir), and it is
possible that the age-related differences in white matter tracts that we observed may also influ-
ence other cognitive abilities. Zum Beispiel, Rizio and Diaz (2016) found that higher FA along
the FAT and SLF was linked to better working memory in both older and younger adults, Und
working memory has been linked both to language comprehension (Caplan & Waters, 2005)
and language production (MacDonald, 2013). Future work should test whether white matter
from the FAT and SLF contributes independently to age-related declines in both working mem-
ory and language production, or whether declines in one of these domains precipitates
declines in the other. Darüber hinaus, we focused on a subset of language-relevant tracts, and did
Neurobiology of Language
282
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
not include the uncinate fasciculus, which connects orbital frontal regions with anterior tem-
poral regions. This tract may also have a role in picture naming, particularly in the naming of
proper nouns (z.B., Papagno, 2011; Papagno et al., 2016), and future work should evaluate its
role in production more thoroughly. Zusätzlich, the present results were obtained from edu-
cated, high-functioning, community dwelling adults. In many ways, the age-related decline
observed here may be smaller than what might be expected in a more socioeconomically
diverse sample, as increased education and higher socioeconomic status have been found
to have protective effects on neural decline and cognition (z.B., Chan et al., 2018; Farah,
2017; Hurst et al., 2013; Stern et al., 2019).
One unexpected result of this study was the direction of some of the relationships between
white matter metrics and naming behaviors. Lower FA and higher RD are typically interpreted as
markers of poorer white matter integrity and consequently associated with poorer behavioral
outcomes. Jedoch, in our mediation models higher SLF FA and lower SLF and FAT RD predicted
higher IES values (d.h., slower naming). Bivariate correlations largely confirmed the expected
direction of results (d.h., increases in RD = slower naming, increases in FA = faster naming),
and path model fit metrics confirmed good model fits for the path models. One consideration
is that our dependent variable, efficiency scores, incorporates both response time and accuracy.
Gesamt, the behavioral effects of age were strongest in our accuracy measure and it could be that
the efficiency scores were more sensitive to this aspect of the age-related behavioral differences.
A final important consideration when interpreting these and all DTI results is the indirect
nature of the white matter measures. Previous studies have confirmed that FA and RD are highly
correlated with white matter structure and that they are also influenced by other microstructural
properties not considered here (Jones et al., 2013; Wheeler-Kingshott & Cercignani, 2009).
Trotz dieser Einschränkungen, these results highlight the importance of white matter in support-
ing language production, and white matter as one mechanism underlying the commonly
observed age-related differences in language production. Consistent with prior reports, Wir
found that increases in age were associated with slower and less accurate picture naming.
We also observed well-established, age-related differences in FA and RD measures of white
matter integrity. Wichtig, these white matter measures from the SLF and FAT mediated the
relationship between age and behavioral performance and suggest that left SLF may be impor-
tant for lexical and semantic selection, while left FAT may have a role for speech fluency or
speech motor planning. Data from this broad sample of adults suggest that age-related deficits
in dorsal white matter contribute to older adults’ deficits in language production above and
beyond effects of age.
ACKNOWLEDGMENTS
We thank the staff and scientists at the Social, Life, & Engineering Sciences Imaging Center and
the Center for Language Science, where the data were collected. This publication was sup-
ported by funding from the National Institute on Aging NIH NIA R01 AG034138 (Michele
T. Diaz). Sara B. W. Troutman was supported by funding from a National Institute on Aging
T32 fellowship while she worked on these analyses (NIH NIA T32 AG049676 to David
Almeida and Lynn Martire).
FUNDING INFORMATION
Michele T. Diaz, National Institute on Aging (https://dx.doi.org/10.13039/100000049), Award
ID: R01 AG034138. Sara B. W. Troutman, National Institute on Aging (https://dx.doi.org/10
.13039/100000049), Award ID: T32 AG049676.
Neurobiology of Language
283
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
BEITRÄGE DES AUTORS
Sara B. W. Troutman: Formale Analyse; Untersuchung; Visualisierung; Writing – original draft.
David J. Madden: Akquise von Fördermitteln; Writing – review & Bearbeitung. Michele T. Diaz: Concep-
tualization; Formale Analyse; Akquise von Fördermitteln; Projektverwaltung; Aufsicht; Writ-
ing – original draft.
VERWEISE
Acheson, D. J., Wells, J. B., & MacDonald, M. C. (2008). New and
updated tests of print exposure and reading abilities in college
students. Behavior Research Methods, 40(1), 278–289. https://
doi.org/10.3758/BRM.40.1.278, PubMed: 18411551
Andersson, J. L. R., Skare, S., & Aschenbrenner, J. (2003). How to correct
susceptibility distortions in spin-echo echo-planar images: Appli-
cation to diffusion tensor imaging. NeuroImage, 20(2), 870–888.
https://doi.org/10.1016/S1053-8119(03)00336-7, PubMed:
14568458
Arbuckle, T. Y., & Gold, D. P. (1993). Altern, inhibition, and verbos-
ität. Journal of Gerontology, 48(5), 225–232. https://doi.org/10
.1093/geronj/48.5.P225, PubMed: 8366267
Bach, M. (1996). The Freiburg visual acuity test—Automatic mea-
surement of visual acuity. Optometry and Vision Science, 73(1),
49–53. https://doi.org/10.1097/00006324-199601000-00008,
PubMed: 8867682
Basser, P. J. (1995). Inferring microstructural features and the phys-
iological state of tissues from diffusion-weighted images. NMR in
Biomedicine, 8(7), 333–344. https://doi.org/10.1002/nbm
.1940080707, PubMed: 8739270
Bennett, ICH. J., & Madden, D. J. (2014). Disconnected aging: Zerebral
white matter integrity and age-related differences in cognition.
Neurowissenschaften, 276, 187–205. https://doi.org/10.1016/j
.neuroscience.2013.11.026, PubMed: 24280637
Benton, A. L., Hamsher, K. D., & Sivan, A. B. (1983). Multilingual
aphasia examination (2nd ed.). Iowa City: AJA Associates.
Boersma, P. (2001). PRAAT, a system for doing phonetics by com-
puter. Glot International, 5(9/10), 341–345.
Brodeur, M. B., Dionne-Dostie, E., Montreuil, T., & Lepage, M.
(2010). The bank of standardized stimuli (BOSS), a new set of
480 normative photos of objects to be used as visual stimuli in
cognitive research. PLOS ONE, 5(5), Article e10773. https://doi
.org/10.1371/journal.pone.0010773, PubMed: 20532245
Brodeur, M. B., Guerard, K., & Bouras, M. (2014). Bank of standard-
ized stimuli (BOSS) phase II: 930 new normative photos. PLOS
ONE, 9(9), Article e106953. https://doi.org/10.1371/journal
.pone.0106953, PubMed: 25211489
Burke, D. M., Mackay, D. G., Worthley, J. S., & Wade, E. (1991).
On the tip of the tongue: What causes word finding failures in
young and older adults? Journal of Memory & Language, 30(5),
542–579. https://doi.org/10.1016/0749-596X(91)90026-G
Burke, D. M., & Shafto, M. A. (2008). Language and aging. In
F. ICH. M. Craik & T. A. Salthouse (Hrsg.), The Handbook of Aging
and Cognition (3rd ed., S. 373–443). Psychology Press.
Caplan, D., & Waters, G. (2005). The relationship between age,
processing speed, working memory capacity, and language com-
prehension. Memory, 13(3–4), 403–413. https://doi.org/10.1080
/09658210344000459, PubMed: 15952262
Catani, M., & die Schotten, M. T. (2012). Atlas of human brain con-
nections. Oxford University Press. https://doi.org/10.1093/med
/9780199541164.001.0001
Catani, M., Mesulam, M. M., Jakobsen, E., Malik, F., Martersteck,
A., Wieneke, C., Thompson, C. K., Thiebaut de Schotten, M.,
Dell’Acqua, F., & Weintraub, S. (2013). A novel frontal pathway
underlies verbal fluency in primary progressive aphasia. Gehirn,
136(8), 2619–2628. https://doi.org/10.1093/ brain/awt163,
PubMed: 23820597
Chan, M. Y., Bereits, J., Agres, P. F., Savalia, N. K., Park, D. C., & Wig,
G. S. (2018). Socioeconomic status moderates age-related differ-
ences in the brain’s functional network organization and anatomy
across the adult lifespan. Proceedings of the National Academies
of Sciences, 115(22), E5144–E5153. https://doi.org/10.1073/pnas
.1714021115, PubMed: 29760066
Christensen, K. J., Moye, J., Armson, R. R., & Kern, T. M. (1992).
Health screening and random recruitment for cognitive aging
Forschung. Psychologie & Altern, 7(2), 204–208. https://doi.org/10
.1037/0882-7974.7.2.204, PubMed: 1610509
Dale, A. M. (1999). Optimal experimental design for event-related
fMRT. Kartierung des menschlichen Gehirns, 8(2-3), 109–114. https://doi.org/10
.1002/(WISSENSCHAFT)1097-0193(1999)8:2/3<109::AID-HBM7>3.0.CO;2-W
Delis, D. C., Kramer, J. H., Kaplan, E., & Ober, B. A. (1987). CVLT,
California verbal learning test. Psychological Corporation.
De Zubicaray, G. ICH., Rose, S. E., & McMahon, K. L. (2011). Der
structure and connectivity of semantic memory in the healthy
older adult brain. NeuroImage, 54(2), 1488–1494. https://doi
.org/10.1016/j.neuroimage.2010.08.058, PubMed: 20807579
Diaz, M. T., Karimi, H., Troutman, S. B. W., Gertel, V. H., Cosgrove,
A. L., & Zhang, H. (2021). Neural sensitivity to phonological
characteristics is stable across the lifespan. NeuroImage, 225,
Article 117511. https://doi.org/10.1016/j.neuroimage.2020
.117511, PubMed: 33129928
Dick, A. S., Bernal, B., & Tremblay, P. (2014). The language con-
nectome: New pathways, new concepts. Neuroscientist, 20(5),
453–467. https://doi.org/10.1177/1073858413513502,
PubMed: 24342910
Dick, A. S., Garic, D., Graziano, P., & Tremblay, P. (2019). Der
frontal aslant tract (FAT) and its role in speech, language and
executive function. Kortex, 111, 148–163. https://doi.org/10
.1016/j.cortex.2018.10.015, PubMed: 30481666
Dick, A. S., & Tremblay, P. (2012). Beyond the arcuate fasciculus:
Consensus and controversy in the connectional anatomy of lan-
Spur. Gehirn, 135(12), 3529–3550. https://doi.org/10.1093/brain
/aws222, PubMed: 23107648
Farah, M. J. (2017). The neuroscience of socioeconomic status:
Correlates, causes, and consequences. Neuron, 96(1), 56–71.
https://doi.org/10.1016/j.neuron.2017.08.034, PubMed:
28957676
Feyereisen, P. (1997). A meta-analytic procedure shows an
age-related decline in picture naming: Comments on Goulet,
Ska, and Kahn (1994). Journal of Speech, Language, and Hearing
Forschung, 40(6), 1328–1333. https://doi.org/10.1044/jslhr.4006
.1328, PubMed: 9430752
Neurobiology of Language
284
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). “Mini-
mental state”: A practical method for grading the cognitive state
of patients for the clinician. Journal of Psychiatric Research, 12(3),
189–198. https://doi.org/10.1016/0022-3956(75)90026-6
Friederici, A. D. (2009). Pathways to language: Fiber tracts in the
human brain. Trends in den Kognitionswissenschaften, 13(4), 175–181.
https://doi.org/10.1016/j.tics.2009.01.001, PubMed: 19223226
Friederici, A. D. (2012). The cortical language circuit: From audi-
tory perception to sentence comprehension. Trends im kognitiven Bereich
Wissenschaften, 16(5), 262–268. https://doi.org/10.1016/j.tics.2012.04
.001, PubMed: 22516238
Gazes, Y., Bowman, F. D., Razlighi, Q. R., O’Shea, D., Stern, Y., &
Habeck, C. (2016). White matter tract covariance patterns pre-
dict age-declining cognitive abilities. NeuroImage, 125, 53–60.
https://doi.org/10.1016/j.neuroimage.2015.10.016, PubMed:
26477658
Gollan, T. H., Montoya, R. ICH., Cera, C., & Sandoval, T. C. (2008).
More use almost always a means a smaller frequency effect:
Altern, bilingualism, and the weaker links hypothesis. Zeitschrift
of Memory & Language, 58(3), 787–814. https://doi.org/10
.1016/j.jml.2007.07.001, PubMed: 19343088
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain image
alignment using boundary-based registration. NeuroImage, 48(1),
63–72. https://doi.org/10.1016/j.neuroimage.2009.06.060,
PubMed: 19573611
Guerin, J. M., Copersino, M. L., & Schretlen, D. J. (2018). Klinisch
utility of the 15-item geriatric depression scale (GDS-15) for use
with young and middle-aged adults. Journal of Affective Disor-
ders, 241, 59–62. https://doi.org/10.1016/j.jad.2018.07.038,
PubMed: 30096593
Hedden, T., Schultz, A. P., Rieckmann, A., Mormino, E. C., Johnson,
K. A., Sperling, R. A., & Buckner, R. L. (2016). Multiple brain
markers are linked to age-related variation in cognition. Zerebral
Kortex, 26(4), 1388–1400. https://doi.org/10.1093/cercor
/bhu238, PubMed: 25316342
Hooper, D., Coughlan, J., & Mullen, M. R. (2008). Equation model-
ling: Guidelines for determining model fit. Electronic Journal of
Business Research Methods, 6(1), 53–60.
Horowitz, T. S., & Wolfe, J. M. (2003). Memory for rejected distrac-
tors in visual search? Visuelle Wahrnehmung, 10(3), 257–298. https://
doi.org/10.1080/13506280143000005
Houston,
J., Allendorfer,
J., Nenert, R., Guter Mann, A. M., &
Szaflarski, J. P. (2019). White matter language pathways and
language performance in healthy adults across ages. Grenzen in
Neurowissenschaften, 13, Article 1185. https://doi.org/10.3389/fnins
.2019.01185, PubMed: 31736704
Hurst, L., Stafford, M., Cooper, R., Hardy, R., Richards, M., & Kuh,
D. (2013). Lifetime socioeconomic inequalities in physical and
kognitives Altern. American Journal of Public Health, 103(9),
1641–1648. https://doi.org/10.2105/AJPH.2013.301240,
PubMed: 23865666
Jenkinson, M., Bannister, P. R., Brady, J. M., & Schmied, S. M. (2002).
Improved optimisation for the robust and accurate linear registra-
tion and motion correction of brain images. NeuroImage, 17(2),
825–841. https://doi.org/10.1006/nimg.2002.1132, PubMed:
12377157
Johnson, M. A., Diaz, M. T., & Madden, D. J. (2015). Global versus
tract-specific components of cerebral white matter integrity:
Relation to adult age and perceptual-motor speed. Brain Struc-
tur & Function, 220(5), 2705–2720. https://doi.org/10.1007
/s00429-014-0822-9, PubMed: 24972959
Jones, D. K., Knösche, T. R., & Turner, R. (2013). Weiße Substanz
integrity, fiber count, and other fallacies: The do’s and don’ts
of diffusion MRI. NeuroImage, 73, 239–254. https://doi.org/10
.1016/j.neuroimage.2012.06.081, PubMed: 22846632
Kantarci, K., Senjem, M., Avula, R., Zhang, B., Samikoglu, A., Weigand,
S., Przybelski, S., Edmonson, H., Vemuri, P., Knopman, D., Boeve,
B. F., Ivnik, R. J., Schmied, G. E., Petersen, R. C., & Jack, C. R. (2011).
Diffusion tensor imaging and cognitive function in older adults
with no dementia. Neurologie, 77(1), 26–34. https://doi.org/10
.1212/ WNL.0b013e31822313dc, PubMed: 21593440
Kaplan, E., Goodglass, H., & Weintrab, S. (1983). The Boston nam-
ing test. Philadelphia: Lea & Febiger.
Kemper, S., Kynette, D., & Norman, S. (1992). Age differences in
spoken language. In R. L. Westen & J. D. Sinnott (Hrsg.), Everyday
memory and aging (S. 138–152). Springer. https://doi.org/10
.1007/978-1-4613-9151-7_9
MacDonald, M. C. (2013). How language production shapes
language form and comprehension. Grenzen in der Psychologie, 4,
Article 226. https://doi.org/10.3389/fpsyg.2013.00226,
PubMed: 23637689
Madden, D. J., Jain, S., Monge, Z. A., Cook, A. D., Lee, A., Huang,
H., Howard, C. M., & Cohen, J. R. (2020). Influence of structural
and functional brain connectivity on age-related differences in
fluid cognition. Neurobiologie des Alterns, 96, 205–222. https://
doi.org/10.1016/j.neurobiolaging.2020.09.010, PubMed:
33038808
Madden, D. J., Parks, E. L., Tallman, C. W., Boylan, M. A., Hoagey,
D. A., Cocjin, S. B., Packard, L. E., Johnson, M. A., Chou, Y., Töpfer,
G. G., Chen, N. K., Siciliano, R. E., Monge, Z. A., Honin, J. A., &
Diaz, M. T. (2017). Sources of disconnection in neurocognitive
Altern: Cerebral white matter integrity, resting-state functional con-
nectivity, and white matter hyperintensity volume. Neurobiology
of Aging, 54, 199–213. https://doi.org/10.1016/j.neurobiolaging
.2017.01.027, PubMed: 28389085
Madhavan, K. M., McQueeny, T., Howe, S. R., Shear, P., &
Szaflarski, J. (2014). Superior longitudinal fasciculus and lan-
guage functioning in healthy aging. Gehirnforschung, 1562,
11–22. https://doi.org/10.1016/j.brainres.2014.03.012, PubMed:
24680744
Moreno-Martinez, F. J., & Montoro, P. R. (2012). An ecological
alternative to Snodgrass & Vanderwart: 360 high quality colour
images with norms for seven psycholinguistic variables. PLOS
ONE, 7(5), Article e37527. https://doi.org/10.1371/journal.pone
.0037527, PubMed: 22662166
Ossher, L., Flegal, K. E., & Lustig, C. (2013). Everyday memory
errors in older adults. Altern, Neuropsychology, and Cognition,
20(2), 220–242. https://doi.org/10.1080/13825585.2012
.690365, PubMed: 22694275
Papagno, C. (2011). Naming and the role of the uncinate fasciculus
in language function. Current Neurology and Neuroscience
Reports, 11(6), Article 553. https://doi.org/10.1007/s11910-011
-0219-6, PubMed: 21853238
Papagno, C., Casarotti, A., Comi, A., Pisoni, A., Lucchelli, F., Bizzi, A.,
Riva, M., & Bello, L. (2016). Long-term proper name anomia after
removal of the uncinate fasciculus. Brain Structure & Function,
221(1), 687–694. https://doi.org/10.1007/s00429-014-0920-8,
PubMed: 25348267
Perron, M., Theaud, G., Descoteaux, M., & Tremblay, P. (2021).
The frontotemporal organization of the arcuate fasciculus and
its relationship with speech perception in young and older ama-
teur singers and non-singers. Kartierung des menschlichen Gehirns, 42(10),
3058–3076. https://doi.org/10.1002/ hbm.25416, PubMed:
33835629
Pierpaoli, C., Jezzard, P., Basser, P. J., Barnett, A., & Di Chiro, G.
(1996). Diffusion tensor MR imaging of the human brain.
Neurobiology of Language
285
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
White matter mediation in picture naming
Radiology, 201(3), 637–648. https://doi.org/10.1148/radiology
.201.3.8939209, PubMed: 8939209
R Core Team. (2014). R: A language and environment for statistical
computing. https://www.R-project.org
Rizio, A. A., & Diaz, M. T. (2016). Language, Altern, and cognition:
Frontal aslant tract and superior longitudinal fasciculus contribute
toward working memory performance in older adults. Neuroreport,
27(9), 689–693. https://doi.org/10.1097/wnr.0000000000000597,
PubMed: 27138951
Rosseel, Y. (2012). lavaan: An R package for structural equation
modeling. Journal of Statistical Software, 48(2), 1–36. https://doi
.org/10.18637/jss.v048.i02
RStudio Team. (2020). RStudio: Integrated development for R.
RStudio PBC. https://www.rstudio.com/
Salami, A., Eriksson, J., Nilsson, L.-G., & Nyberg, L. (2012). Alter-
related white matter microstructural differences partly mediate
age-related decline in processing speed but not cognition. Bio-
chimica Et Biophysica Acta (BBA) – Molecular Basis of Disease,
1822(3), 408–415. https://doi.org/10.1016/j.bbadis.2011.09.001,
PubMed: 21930202
Sheikh, J. ICH., & Yesavage, J. A. (1986). Geriatric Depression Scale
(GDS): Recent evidence and development of a shorter version.
Clinical Gerontologist: The Journal of Aging and Mental Health,
5(1–2), 165–173. https://doi.org/10.1300/J018v05n01_09
Schmied, S. M. (2002). Fast robust automated brain extraction. Human
Brain Mapping, 17(3), 143–155. https://doi.org/10.1002/ hbm
.10062, PubMed: 12391568
Schmied, S. M., Jenkinson, M., Woolrich, M. W., Beckman, C. F.,
Behrens, T. E. J., Johansen-Berg, H., Bannister, P. R., De Luca,
M., Drobnjak, ICH., Flitney, D. E., Niazy, R., Saunders, J., Vickers,
J., Zhang, Y., De Stefano, N., Brady, J. M., & Matthews, P. M.
(2004). Advances in functional and structural MR image analysis
and implementation as FSL. NeuroImage, 23(S1), 208–219.
https://doi.org/10.1016/j.neuroimage.2004.07.051, PubMed:
15501092
Snodgrass, J. G., & Vanderwart, M. (1980). A standardized set of
260 pictures: Norms for name agreement, image agreement,
Vertrautheit, and visual complexity. Journal of Experimental Psy-
chology: Human Learning and Memory, 6(2), 174–215. https://
doi.org/10.1037/0278-7393.6.2.174, PubMed: 7373248
Song, A. W., Wong, E. C., Bräunen, S. G., & Hyde, J. S. (1996). Diffusion
weighted fMRI at 1.5 T. Magnetic Resonance in Medicine, 35(2),
155–158. https://doi.org/10.1002/mrm.1910350204, PubMed:
8622577
Song, S.-K., Sun, S.-W., Ju, W.-K., Lin, S.-J., Cross, A. H., & Neufeld,
A. H. (2003). Diffusion tensor imaging detects and differentiates
axon and myelin degeneration in mouse optic nerve after retinal
ischemia. NeuroImage, 20(3), 1714–1722. https://doi.org/10
.1016/j.neuroimage.2003.07.005, PubMed: 14642481
Song, S.-K., Yoshino, J., Le, T. Q., Lin, S.-J., Sun, S.-W., Cross, A. H.,
& Armstrong, R. C. (2005). Demyelination increases radial
diffusivity in corpus callosum of mouse brain. NeuroImage, 26(1),
132–140. https://doi.org/10.1016/j.neuroimage.2005.01.028,
PubMed: 15862213
Stamatakis, E. A., Shafto, M. A., Williams, G., Tam, P., & Tyler, L. K.
(2011). White matter changes and word finding failures with
increasing age. PLOS ONE, 6(1), Article e14496. https://doi.org
/10.1371/zeitschrift.pone.0014496, PubMed: 21249127
Stern, Y., Barnes, C. A., Grady, C., Jones, R. N., & Raz, N. (2019).
Brain reserve, cognitive reserve, compensation, and maintenance:
Operationalization, validity, and mechanisms of cognitive
Widerstandsfähigkeit. Neurobiologie des Alterns, 83, 124–129. https://doi.org/10
.1016/j.neurobiolaging.2019.03.022, PubMed: 31732015
Stojanoski, B., & Cusack, R. (2014). Time to wave good-bye to phase
scrambling: Creating controlled scrambled images using diffeo-
morphic transformations. Journal of Vision, 14(12), Article 6.
https://doi.org/10.1167/14.12.6, PubMed: 25301014
Tournier, J.-D., Schmied, R., Raffelt, D., Tabbara, R., Dhollander, T.,
Pietsch, M., Christiaens, D., Jeurissen, B., Yeh, C.-H., & Connelly,
A. (2019). MRtrix3: A fast, flexible and open software framework
for medical image processing and visualisation. NeuroImage,
202, Article 116137. https://doi.org/10.1016/j.neuroimage.2019
.116137, PubMed: 31473352
Tremblay, P., Perron, M., Deschamps, ICH., Kennedy-Higgins, D.,
Houde, J.-C., Dick, A. S., & Descoteaux, M. (2019). Die Rolle
of the arcuate and middle longitudinal fasciculi in speech per-
ception in noise in adulthood. Kartierung des menschlichen Gehirns, 40(1),
226–241. https://doi.org/10.1002/ hbm.24367, PubMed:
30277622
Troutman, S. B. W., & Diaz, M. T. (2019). White matter disconnec-
tion is related to age-related phonological deficits. Brain Imaging
and Behavior, 14(5), 1555–1565. https://doi.org/10.1007/s11682
-019-00086-8, PubMed: 30937829
Varriano, F., Pascual-Diaz, S., & Prats-Galino, A. (2020). Unterscheidbar
components in the right extended frontal aslant tract mediate
language and working memory performance: A tractography-
informed VBM Study. Frontiers in Neuroanatomy, 14, Article 21.
https://doi.org/10.3389/fnana.2020.00021, PubMed: 32372922
Veraart, J., Novikov, D. S., Christiaens, D., Ades-Aron, B., Sijbers, J.,
& Fieremans, E. (2016). Denoising of diffusion MRI using random
matrix theory. NeuroImage, 142, 394–406. https://doi.org/10
.1016/j.neuroimage.2016.08.016, PubMed: 27523449
Wechsler, D. (1997). WAIS-III administration and scoring manual.
The Psychological Corporation.
Wheeler-Kingshott, C. A., & Cercignani, M. (2009). About “axial”
and “radial” diffusivities. Magnetic Resonance in Medicine, 61(5),
1255–1260. https://doi.org/10.1002/mrm.21965, PubMed:
19253405
Neurobiology of Language
286
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
3
2
2
7
2
2
0
0
4
9
5
9
N
Ö
_
A
_
0
0
0
6
5
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3