RESEARCH ARTICLE
Thematic Reanalysis in the Left Posterior
Parietal Sulcus: A TMS Study
Chiara Finocchiaro1, Luigi Cattaneo2, Carlotta Lega3, and Gabriele Miceli1,2,4
1Department of Psychology and Cognitive Sciences, University of Trento, Trient, Italien
2Center for Mind/ Brain Sciences (CIMeC), University of Trento, Trient, Italien
3Abteilung für Psychologie, University of Milan Bicocca, Milan, Italien
4Beniamino Segre Interdisciplinary Center, National Academy of the Lincei, Rome, Italien
Schlüsselwörter: transcranial magnetic stimulation, thematic role assignment, reanalysis, posterior parietal
Sulkus
ABSTRAKT
Understanding who does what to whom is at the core of sentence comprehension. The actors
that contribute to the verb meaning are labeled thematic roles. We used transcranial magnetic
stimulation (TMS) to verify the possible impact of verb semantics on the thematic role
encoding process that has been shown to involve the posterior portion of the left posterior
parietal sulcus (PPS; Finocchiaro et al., 2015). Sixteen participants underwent TMS and sham
stimulation sessions while performing an agent-decision task, in which they had to decide
by key press which of the two arguments was the agent of visually presented sentences or
pseudo-sentences. Der (pseudo)sentences were all reversible and were presented in the
active or passive diathesis. Double pulse TMS was delivered to the posterior part of the
intraparietal sulcus in an event-related fashion, at two different time windows: 200–400 ms (T1)
or 600–800 ms (T2) time-locked to the presentation of the (pseudo)Satz. Results showed
that TMS increased accuracy on passive sentences and pseudo-sentences as compared to
active sentences and to the baseline, sham condition. In der Tat, the presence of a verb with a full
semantic representation was not a necessary precondition for the TMS-induced facilitation of
passive (pseudo)Sätze. Stimulation timing had no effect on accuracy for sentences vs.
pseudo-sentences. These observations support the idea that the posterior parietal site is
recruited when the correct interpretation of a sentence requires reanalysis of temporarily
encoded thematic roles (as in reversible passive sentences) even when the verb is not an entry
in the lexicon and hence does not have a semantic representation. Results are consistent with
previous evidence and deserve further investigation in larger experimental samples. Increasing
the number and variety of stimulus sentences, and administering TMS to additional control
sites will be key to further articulate the conclusions allowed by these initial findings.
EINFÜHRUNG
The ability to understand who does what to whom is a core function of sentence comprehen-
sion. In der Tat, understanding a verb means knowing the arguments it takes and the thematic
roles it assigns to them (Chomsky, 1965, 1981). The who does what to whom structure con-
tains all the main thematic roles: the agent is who does the action, the theme is who/what is
involved in that action, the goal is the point to which that action is directed (to whom). Across
Keine offenen Zugänge
Tagebuch
Zitat: Finocchiaro, C., Cattaneo, L.,
Lega, C., & Miceli, G. (2021). Thematic
reanalysis in the left posterior parietal
Sulkus: A TMS study. Neurobiology of
Language, 2(3), 416–432. https://doi.org
/10.1162/nol_a_00043
DOI:
https://doi.org/10.1162/nol_a_00043
zusätzliche Informationen:
https://doi.org/10.1162/nol_a_00043
Erhalten: 11 Oktober 2019
Akzeptiert: 14 Mai 2021
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Chiara Finocchiaro
chiara.finocchiaro@unitn.it
Handling-Editor:
Kate Watkins
Urheberrechte ©: © 2021
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
Thematic role:
Semantic role that a given noun
phrase may play in relation to the
action/event/state denoted by a
governing verb.
languages, the most commonly used verbs are those whose thematic grid—in which the the-
matic roles associated to a given verb are specified—requires the roles of agent and theme. In
Indo-European languages like English or Italian these verbs tend to correspond to transitive
verbs (d.h., those that usually refer to an action performed by someone and involving
someone/something else). When a transitive verb is used in the active diathesis, the thematic
role of its arguments (agent and theme, jeweils) matches their syntactic function (Thema
and object, jeweils). Zum Beispiel, in the sentence Judith bites the apple, Judith is both the
subject of the sentence and the agent of the bite-action, and the apple is both the object of the
sentence and the theme of the bite-action. Except for a few idiomatic expressions that are not
admissible in the passive voice, transitive verbs normally allow the passive transformation.
Das ist, the same event as Judith bites the apple can be described by the passive sentence
The apple is bitten by Judith. The apple is now the subject of the sentence, but its thematic
role is the same as in the active sentence (d.h., theme) as the apple does not start the biting-
action but is still the element that undergoes biting. Judith is the agent because she gives origin
to the biting, even though she is no longer the subject of the sentence and is now introduced
by a by-phrase. Similar considerations apply to a sentence like Judith bites Claire. In the active
diathesis, Claire is both the object and the theme but, when the sentence is transformed in the
passive diathesis (Claire is bitten by Judith), Claire becomes the grammatical subject while
remaining the logical theme.
There is, Jedoch, a major difference between the two sentences Judith bites the apple /
The apple is bitten by Judith, and the two sentences Judith bites Claire / Claire is bitten by
Judith: Whereas pragmatic cues are available for the first sentence pair, thus allowing one
to assign the agent role to Judith irrespective of the sentence diathesis, the same cues are
not available for the second sentence pair. This is because we know that apples do not bite
Menschen, whereas we do not have any expectations about the biting habits of women whose
names are Judith or Claire. daher, sentences like Judith bites Claire and its passive version
are called reversible (both arguments could in principle be agents); by contrast, sentences like
Judith bites the apple and its passive version are called irreversible (the two arguments have a
very different likelihood of being the agent).
Note that bare thematic violations do not violate argument structure, and as such are dif-
ferent from subcategorization violations in which the argument realization is not allowed by
das Verb (Garnsey et al., 1997; McElree & Griffith, 1995). Daher, in The apple bites Judith,
there is no syntactic violation, but the apple is not marked for animacy as would be required
for being the agent of a biting action. There is also a critical difference between active and
passive sentences. In the passive diathesis there is a systematic mismatch between the ex-
pected order of the syntactic constituents and their thematic roles. In sentence comprehen-
sion, speakers show the tendency to interpret the first argument of a sentence as the subject
and the agent of the denoted event (Ferreira, 2003; Grodzinsky, 2000; Meyer et al., 2012).
This linear order cue may help the comprehension of active sentences, but leads to incorrect
predictions for passive sentences, thus complicating the comprehension of passives and forc-
ing the listener to reanalyze temporarily encoded thematic roles. Darüber hinaus, the passive di-
athesis is particularly difficult when, in addition to linear order cues being deceptive, Die
application of pragmatic cues also fails (d.h., in reversible passive sentences). Turner and
Rommetveit (1967) proposed a hierarchy of sentence complexity, starting from the least com-
plex nonreversible active, to reversible active, nonreversible passive, and ending with the
most complex reversible passive.
Because of the absence of pragmatic or word order cues, reversible passives have been used
to isolate the syntactic component of sentence comprehension in both neuropsychological and
Neurobiology of Language
417
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
Reversible sentences:
Sentences for which both arguments
could in principle be agents (there is
no semantic cue).
neuroimaging studies. In their seminal paper, Brookshire and Nicholas (1980) showed that
aphasic and nonaphasic participants were slower and less accurate on passive than on active
reversible sentences. The first report of a highly selective difficulty in the production and com-
prehension of thematic roles was provided by Caramazza and Miceli (1991). Patient EB, von-
spite being able to flawlessly process the morphological structure of a sentence, was impaired
in both comprehension and production of reversible sentences, especially (Aber, not exclusively)
when sentences were passive. Other studies focused on reversible sentences as a means to
manipulate the order (canonical vs. noncanonical) of the structural elements subject-verb-
Objekt (S-V-O). In Ferreira (2003), healthy participants were slower and less accurate in iden-
tifying the agent of reversible than nonreversible sentences, especially when the canonical
S-V-O order was violated (z.B., reversible passives). Using the eye-tracking-while-listening
paradigm, Meyer et al. (2012) found that upon hearing the first argument (N1) of active and
passive sentences both aphasic and control participants immediately looked at the picture in
which N1 was the agent.
A growing number of imaging studies have focused on the neural correlates of thematic role
Abtretung. Studies reporting on comprehension disorders in aphasic patients, with emphasis
on reversible sentences in the active and passive diathesis (Magnusdottír et al., 2013; Rogalsky
et al., 2017; Thothathiri et al., 2012), documented substantial lesion overlap in posterior tem-
poral and parietal regions (superior temporal, angular, and supramarginal gyri). In a study by
Thompson et al. (2013) agrammatic patients were trained to exploit the argument properties of
verbs through tasks that required verb naming and sentence generation. After training, Die
authors observed an improvement in verb production that generalized to untrained verbs
and recruited posterior perisylvian regions as well as parietal and sensory-motor cortices
bilaterally.
Studies also show that the complexity of a verb’s thematic grid can influence the neural
mechanisms involved in the comprehension of reversible sentences. Thematic role complexity
has been defined in different ways, als (1) the number of arguments a given verb may take
(Thompson et al., 2010); (2) the transitive vs. intransitive opposition as a particular instance
of the contrast between one-argument vs. two-argument verbs; (3) the number of thematic
grids that may be associated with a single verb (Meltzer-Asscher et al., 2013). In der Tat,
alternating-transitivity verbs may be associated to more than one thematic grid, as they allow
for transitive as well as intransitive use (z.B., to eat in Judith has already eaten, intransitive, Und
in Judith has already eaten her sandwich, transitive).
As to the neural correlates of verb comprehension processes, most fMRI studies show in-
creasing activity in a (mostly left-sided) posterior perisylvian network and in the parietal lobe,
that parallels increasing thematic role complexity. These phenomena involve the same regions
found to be associated with verb production after training in Thompson et al. (2013).
Other neuroimaging studies used sentences instead of words. In Richardson et al. (2010),
participants were asked to listen to or read silently reversible and nonreversible sentences. Der
left temporo-parietal boundary was found to be significantly more activated for reversible as
opposed to nonreversible sentences. Mack et al. (2013) contrasted active and passive sen-
tences and found greater activation for passives in the inferior frontal gyrus bilaterally and
in left temporal regions.
The picture that emerges from the fMRI literature, despite differences across studies, con-
verges with the neuropsychological literature in showing the involvement of left temporo-
parietal regions in the assignment of thematic roles. In a recent study, Finocchiaro et al.
(2015) used focal transcranial magnetic stimulation (TMS) to investigate the role of the parietal
Neurobiology of Language
418
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
cortex in thematic role assignment. Focal TMS has an immediate effect on the neural activity
of the stimulated portion of cortex, so that any TMS-related change in behavioral performance
is likely to reflect the physiological function of the stimulated region. In their study, a reversible
active or passive sentence was presented together with a picture. Participants had to judge, von
key press, whether or not the sentence matched the picture (sentence-picture matching task).
Three sites covering the whole length of the intraparietal sulcus (IPS) were stimulated with a
short train of event-related double pulse TMS. Accuracy for reversible passive sentences
increased following stimulation of the most posterior site along the IPS (probably corresponding
to area V6A in the monkey) commonly thought to process visual information in a spatial frame
of reference and possibly specialized in establishing relations between different components,
as in depth judgments with objects or textures (Grefkes & Fink, 2005). Finocchiaro et al.
interpreted their results as showing that the neural machinery in the posterior IPS is involved
in linguistic processes and specifically in the assignment of thematic roles, especially when the
reanalysis of transiently encoded thematic roles is required, as is the case in passive reversible
Sätze.
There is no consensus, Jedoch, on the theoretical status of the notion of thematic role.
Fillmore’s original view (Fillmore, 1968) of thematic roles as primitive, verb-independent en-
tities has been replaced by a number of accounts that vary greatly as to the participation of
verb-specific semantics in the qualification of the agent and the theme. One of the most influ-
ential models was proposed by Dowty (1989, 1991). According to that model, thematic roles
may be represented as groups of prototypical entailments imposed by a group of verbs on their
arguments. Taken individually, the features associated to “proto-agents” and “proto-patients”
are not necessary and sufficient to define an argument as agent or patient/theme, but together
they contribute to the likelihood that a given argument is interpreted as an agent or a
patient/theme. Das ist, whereas the “volitional involvement in the event or state” (Dowty,
1991, P. 572) is an entailment associated to protypical agents, there could be less prototypical
agents that do not volitionally start the event/state. Along the same line, the model interprets
other features more or less prototypically associated with agents or patients/themes. Other
accounts, though not incompatible with Dowty’s view, emphasize the importance of verb-
specific semantics for the characterization of thematic roles (McRae et al., 1997). These ap-
proaches focus on the characteristics normally associated with the thematic roles of agent and
patient/theme of a specific verb. On this view, the interest shifts from verb-independent fea-
tures to the verb-specific features associated with the agent of, Zum Beispiel, to kill as opposed
to to hide, aiming at a richer characterization of the features that agents and patients may re-
ceive depending on the verb. (Other studies have stressed the importance of typological con-
cepts such as prominence scales [animacy, definiteness/specificity, case marking, and linear
Befehl] on the comprehension of simple transitive sentences, [z.B., Bornkessel et al., 2005;
Bornkessel-Schlesewsky & Schlesewsky, 2009]. These concepts are not directly addressed
in our study.)
Be this as it may, all these accounts share the view that thematic roles cannot be assigned
independently from verb semantics (see also Van Valin’s, 1999, notion of “generalized seman-
tic roles”; für eine Rezension, see Lebani et al., 2015, and Levin & Rappaport Hovav, 2005). The goal
of the present study is to better qualify the nature of the thematic role encoding processes
shown to involve the posterior portion of the left parietal sulcus (PPS) in Finocchiaro et al.
(2015). The crucial question is whether the semantics of a verb is crucial for the facilitatory
effect on passives that follows PPS stimulation.
To answer this question, we presented participants with pseudo-sentences in addition to
real sentences. If verb-specific semantic features are considered in the reanalysis process for
Neurobiology of Language
419
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
which the PPS is recruited, the facilitation effect should be limited to real sentences. Wenn, on the
andererseits, the process relies on primitive semantic features related to a proto-meaning of the
type someone did or suffered something, facilitation should extend to pseudo-sentences, für
which that proto-meaning may be recovered.
It is important to point out that “reanalysis” is used here as a theoretically-free label. In diesem
Studie, we deliberately do not take a position in the linguistic debate on the involvement of
syntactic movement in passive sentences. Our point is that both movement-based accounts
(z.B., Chomsky, 1981) and lexical-thematic accounts (Bresnan, 2000; Pollard & Sag, 1994)
share the view that passive sentences—as well as other types of noncanonical sentences—
require a process of thematic reanalysis in order to revise the initial mapping of thematic roles.
This process is not required in the case of active sentences (and canonical sentences in
allgemein).
Introducing pseudo-sentences led to a major difference in the experimental design. In
Finocchiaro et al. (2015), TMS was delivered during a sentence-picture verification task, In
which participants were asked to decide whether a written sentence was the appropriate
description of the action represented in a picture. In the current study, pictures could not
be used as stimuli, as pseudo-sentences cannot be depicted. Folglich, the sentence-
picture matching task was substituted with an agent-decision task. In this task, the stimulus
(pseudo)sentence is presented at the top center of the screen. The stimulus, being reversible,
has two arguments. The two arguments are presented simultaneously below the (pseudo)
Satz, one on top of the other. The participant is asked to decide by key-press which of
the two arguments is the agent.
Based on the results reported in Finocchiaro et al. (2015), we focused on the stimulation site
shown to be involved in the processing of passive sentences, das ist, the posterior portion of the
left IPS. The simplification of the overall TMS design allowed introducing a sham TMS condi-
tion as a within-participant variable, which permitted a quantitative as well as a qualitative
comparison between the sham TMS and the TMS conditions.
We also introduced two different timings of TMS. In der vorliegenden Studie, double pulse TMS
was delivered at 200–400 ms or at 600–800 ms, in order to explore whether the effects of TMS
varied as a function of the stimulation window and/or of the sentence/pseudo-sentence status
of the target. Since sentence comprehension is an incremental process, showing that the ef-
fects of TMS differ as a function of stimulation timing may shed light on the time window in
which reanalysis takes place.
To sum up, we expect to replicate previous findings by showing the involvement of the PPS
in the processing of passive sentences relative to active sentences. Zusätzlich, manipulating
the amount of semantic information available for verbs (complete for real sentences, very em-
bryonal for pseudo-sentences) should allow inferences about the nature of the information that
is relevant for the PPS. Since the PPS is not known to have a specific semantic role, we also
expect the effect of TMS on passive processing to involve both sentences and pseudo-
Sätze. Andererseits, the manipulation of the TMS timing is exploratory, as we are
not aware of relevant findings that could help shape our predictions.
MATERIALS AND METHODS
Teilnehmer
Sixteen native Italian speakers took part in the experiment for course credit. They were all
right-handed (Oldfield, 1971), aged between 19 Und 35 Jahre (mean: 24.7), and had an
Neurobiology of Language
420
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
Pseudo-sentences:
Sentences made up of pseudo-words.
Word-endings and function words
are real elements in a given
Sprache.
Pseudo-words:
In a given language, möglich
but not existent strings of
graphemes/phonemes.
educational level between 14 Und 23 Jahre (mean: 18). All participants had normal or
corrected-to-normal vision, normal hearing, and no history of neurological or psychiatric
disorders. They were preliminarily screened for any relative or absolute contraindication to
TMS, received exhaustive information on the TMS procedure, and signed an informed consent
bilden. Information about the purpose of the study was provided at the end of the test session.
The study was approved by the ethics committee of the University of Trento.
Materials
Two sets of stimuli were created. Stimuli in the first set (N = 60) were sentences containing real
Wörter; stimuli in the second set (N = 60) were pseudo-sentences containing pseudo-words.
Pseudo-sentences contained real free-standing (determiners, prepositions, auxiliaries) Und
bound grammatical morphemes (gender and number morphemes for nouns; Zeitform, mood,
and person morphemes for verbs) but pseudo-word roots. Sentences and pseudo-sentences
were presented half in the active and half in the passive diathesis, and they were all reversible
(as the verb arguments used in pseudo-sentences are meaningless, there is no pragmatic con-
straint on their thematic roles).
Pseudo-words used in the pseudo-sentence set had an N-count (d.h., number of real words
that can be obtained by substituting one letter at a time in each pseudo-word) equal to 0.
Forty-three transitive verbs and 92 nouns were used in the sentence set. They appeared once
or twice (in this latter case, in different blocks) during the experiment. Sentences and pseudo-
sentences were of the same length in terms of number of (pseudo)Wörter (5 for active (pseudo)
Sätze, 6 for passive (pseudo)Sätze). We paid particular attention to excluding verbs
that denoted spatial relations (z.B., folgen, precede), as there is evidence that the processing of
spatial relations may recruit parietal regions (Amorapanth et al., 2010; Baumann & Mattingley,
2014).
Stimuli were divided into three blocks of 40 items each. The four experimental conditions
(Satz, active; Satz, passive; pseudo-sentence, active; pseudo-sentence, passive) war
equally represented in each block. Sixteen additional stimuli were used as practice trials. Sie
were presented to participants before the experiment proper began. They had the same char-
acteristics as the experimental stimuli and were equally distributed across the experimental
Bedingungen.
Verfahren
The study was completed in one session that lasted about one hour per participant. The ex-
periment was preceded by a practice block with the same characteristics as the experimental
blocks, except that TMS was not delivered. The practice session could be repeated for a max-
imum of three times until the participant produced at least 80% correct responses. The exper-
iment proper was repeated twice, once with TMS and once with a sham TMS. The order of
presentation of the two conditions was counterbalanced across participants. For the TMS con-
dition, double pulse TMS was delivered at T1 (zwischen 200 Und 400 MS) or T2 (zwischen 600
Und 800 MS). The same number of pulses was delivered at T1 and T2 for sentences and
pseudo-sentences. The rationale for exploring these specific time periods with TMS is three-
fold. Erste, the average response time (RT) in this and similar tasks is around 1,500 MS. Während
this time window, the participant reads the sentence and the two alternative targets (sehen
Figur 1) and produces a manual response. Given our experimental question, we aimed at
exploring the initial and middle part of the RT, when the participant is actually reading the
complete sentence. Zweite, studies on syntactic violations with event-related potentials hint
Neurobiology of Language
421
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
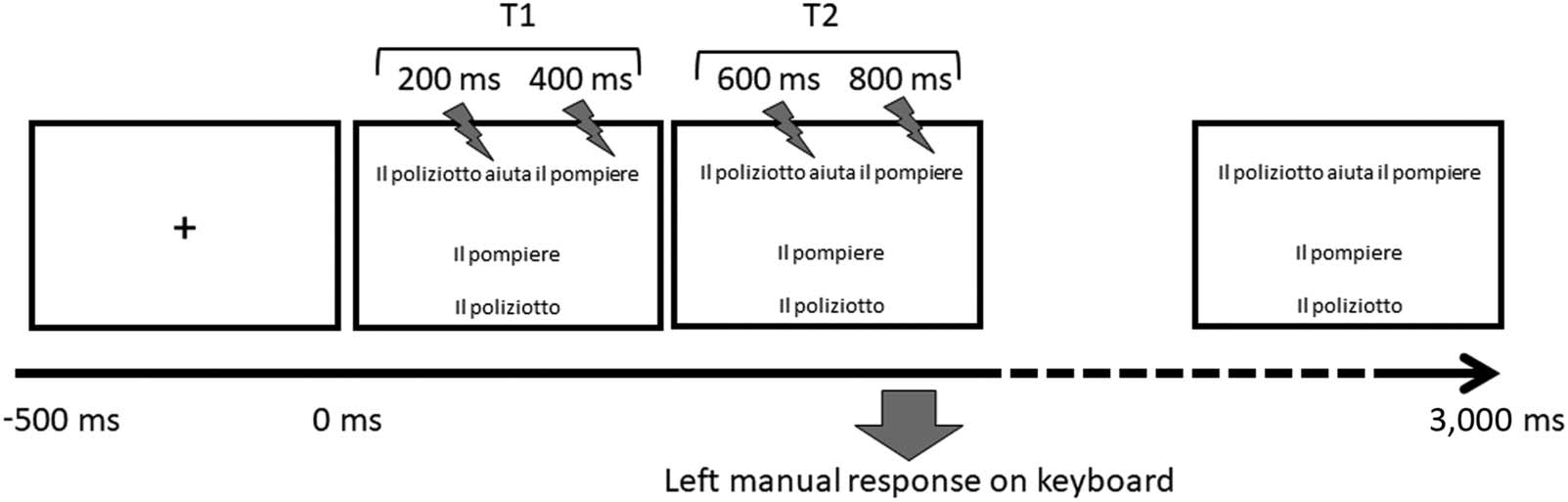
Figur 1.
Schematization of trial timeline and structure.
that syntactic processing occurs in three specific time windows, starting at 150 ms up to 800 MS
from the syntactic feature (Friederici, 1995, 2002; Friederici & Kotz, 2003). Dritte, active and
passive sentences have syntactic features in different positions and are likely to be processed at
slightly different timings, hence the need to explore at least two separate time windows.
At the beginning of each trial a fixation cross appeared on-screen for 500 MS, immediately
followed by the stimulus. Each stimulus consisted of a full (pseudo)sentence center aligned in
the upper half of the screen (z.B., Il poliziotto aiuta il pompiere ‘The policeman helps the fire-
man’ or Il pompiere è aiutato dal poliziotto ‘The fireman is helped by the policeman’), welche
appeared at the same time as the two arguments (Il pompiere ‘The fireman’ and Il poliziotto
‘The policeman’), shown one above the other, center aligned in the lower half of the screen.
The position of the two arguments was counterbalanced across conditions, so that the agent
was above or below the theme an equal number of times. The stimulus remained on-screen
until participants responded or 3,000 ms had elapsed, whichever came first. Participants were
asked to read the sentence and to decide, by pressing the “B” or the “N” key with their left
Hand, whether the agent was the argument appearing in the top or in the bottom line. Der
agent was previously defined as the person who does the action denoted by the verb. Wir
chose the left hand as effector to avoid any interference of left hemispheric TMS on the motor
processes involved in producing a hand movement. A single trial is schematically represented
in Abbildung 1. Gesamt, the study was designed for repeated measures with trials fully distributed
according to four within-subject factors: TMS (effective or sham), TIME of stimulation (zwei
levels), SEMANTICS (word or pseudo-word), DIATHESIS (active or passive). The presentation
of the stimuli within each block was completely randomized. The order of block presentation
was counterbalanced across participants according to a Latin-square design. Stimulus presen-
tation was controlled by E-Prime software (https://pstnet.com/products/e-prime/).
TMS
Magnetic stimuli were delivered with a double 70 mm figure-eight coil connected to a
Magstim Rapid (The Magstim Company, Vereinigtes Königreich) biphasic stimulator that was externally triggered
by TTL pulses at appropriate timing by the E-Prime software through the parallel port of the
computer. Following up on the results obtained by Finocchiaro et al. (2015), we delivered the
double pulse TMS to the posterior third of the left IPS (d.h., to the site where TMS affected
the processing of passive sentences). In der vorliegenden Studie, the TMS target was identified by means
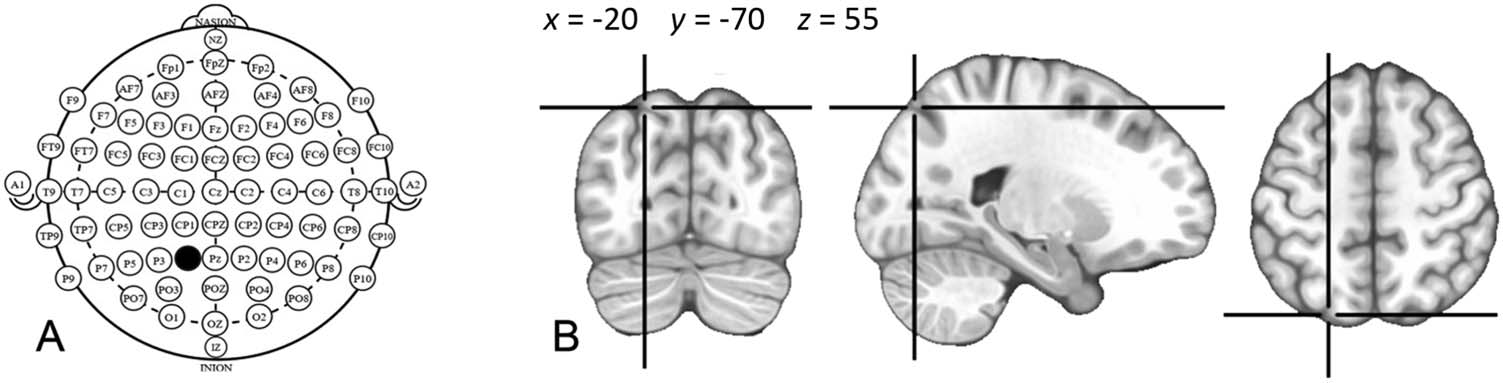
of the international 10-20 coordinates (siehe Abbildung 2). Several atlases provide probabilistic maps
of the cortical structures that correspond to specific 10-20 system coordinates on the scalp
(Herwig et al., 2003; Kabdebon et al., 2014; Koessler et al., 2009; Okamoto et al., 2004;
Neurobiology of Language
422
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
Figur 2.
(A) Reference coordinates of the international 10-20 system for EEG electrode placement.
Coordinate P1, used in the present study as stimulation target is indicated in black. (B) According to
probabilistic scalp-brain atlases (Herwig et al., 2003; Kabdebon et al., 2014; Koessler et al., 2009;
Okamoto et al., 2004; Sparing et al., 2010; Yoo et al., 1997), the cortical projection of the P1 scalp
coordinate is probabilistically distributed between-subjects over a region that has a maximum of
probability (d.h., is localized in most subjects) at the −20, −70, 55 coordinates of the standardized
MNI space, indicated in the figure with the crosshair on a standard MNI brain.
Sparing et al., 2010). Based on these atlases we identified the P1 coordinate (located halfway
between the Pz and P3 coordinates in the extended 10-20 System) as the one most probably
overlying the posterior part of the IPS. The coil was oriented so as to be roughly perpendicular
to the IPS, at 90° to the midline, the handle pointing medially. Stimulation intensity was indi-
vidually set to 100% of the visually-assessed motor threshold (d.h., to the intensity required to
produce a visible twitch in hand muscles in exactly 50% of eight trials). The sham TMS con-
dition was achieved by tilting the coil by 90°, so that only the edge of the TMS coil touched the
scalp and therefore the subject experienced the mechanical and acoustical experience of
effective TMS, without actual cortical stimulation.
Data Pre-processing
Analyses were carried out on RTs and on accuracy (Acc). The experimental design was based
on repeated measures with four within-subject factors: TMS (2 levels: sham or effective),
DIATHESIS (2 levels: active or passive), SEMANTICS (2 levels: sentence or pseudo-sentence)
and TIME (2 levels: TMS delivered at 200–400 ms [T1] and TMS delivered at 600–800 ms
[T2]). Es gab 15 trials in each cell of this 2 × 2 × 2 × 2 Design. Trials were trimmed
according to RTs. Any trials with RTs longer or shorter than the mean +/−2 standard deviations
(SDs) were excluded from further analysis. As expected from a normal distribution, RTs fell >2
SD below the mean in 1% of the trials. Zusätzlich, we excluded from further analysis trials in
which no response was given after 3 S. After data trimming, the numerosity of trials/cell went
aus 15 to an average of 14.1, ranging from 10 Zu 15. Trials with incorrect responses were
excluded from the RT analysis, but were obviously included in the Acc analysis.
Statistical Analysis
Linear mixed-effect models were used (Baayen et al., 2008). The model was estimated using R
(R Development Core Team, 2016) and the lme4 package (Bates et al., 2015). Post hoc tests
were conducted using the R-package phia (Ausführung 0.2-0; De Rosario-Martinez et al., 2015)
and applying the Bonferroni-Holm correction for multiple comparisons. A mixed logistic
model was estimated using accuracy as the dependent variable. TMS, DIATHESIS,
SEMANTICS, and TIME and their interaction were entered as fixed-effect factors in a mixed
logistic model that predicted accuracy. Intercepts were also included in the model as
random-effect factors across participants and across session. An identical analysis was per-
formed on RTs as the dependent variable.
Neurobiology of Language
423
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
Effect Sizes
Calculating effect size for mixed models is difficult, and there is little consensus among stat-
isticians on how to do it (see Nakagawa & Schielzeth, 2013). Reporting the effect size for anal-
yses such as mixed-effects regression modeling and hierarchical linear modeling can be
difficult. For these reasons, most reports so far have avoided reporting effect size when using
mixed model analysis. One of the best approximations of effect size for general linear model is
Cohen’s f 2 (Selya et al., 2012), “which allows an evaluation of local effect size, d.h., one var-
iable’s effect size within the context of a multivariate regression model” (Selya et al., 2012,
P. 1). Cohen’s f 2 is calculated starting from the R2, which is the proportion of variance ac-
counted for by the model. Mixed models have 2 different types of R2: the marginal R2, welche
represents the variance explained by the fixed effects, and the conditional R2, which repre-
sents the variance explained by the entire model, including both fixed and random effects.
2 (marginal)
daher, we calculated two different f 2 for each significant effect, namely the fm
2 (conditional). Vor allem, for multi-level models, effect sizes calculated using residual
and the fc
variance and proportion of variance explained should be interpreted with caution, Weil
adding variables to the model may increase residual variance resulting in negative estimates
of explained variance and even of effect size (Snijders & Bosker, 1994). In the present work we
proceeded as follows. The model was estimated using R (R Development Core Team, 2016)
and the lme4 package (Bates et al., 2015). Post hoc tests were conducted using the R-package
phia (Ausführung 0.2-0; De Rosario-Martinez et al., 2015) and applying the Bonferroni-Holm
correction for multiple comparisons. A mixed logistic model was estimated using accuracy
as the dependent variable. TMS, DIATHESIS, SEMANTICS, and TIME and their interaction
were entered as fixed-effect factors in a mixed logistic model that predicted accuracy.
Intercepts were also included in the model as random-effect factors across participants and
across session.
ERGEBNISSE
Planned Analysis
None of the participants reported side effects, immediate or delayed. Accuracy values and
latencies are presented in Tables 1 Und 2.
The analysis on RTs did not yield any effect of interest (d.h., any effect including the TMS factor).
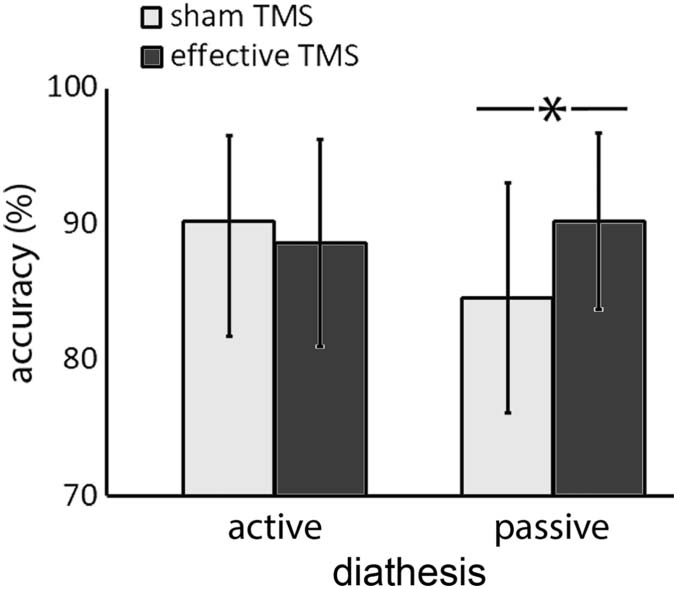
Umgekehrt, the analysis on accuracies revealed a significant main effect of DIATHESIS, (Chi2 (1) =
2 = 0.002), and a main effect of TIME (Chi2 (1) = 7.28, p = 0.006,
4.18, p = 0.040, fm
2 = 0.003). The interaction between TIME and DIATHESIS was also significant (Chi2
2 = 0.002, fc
fm
2 = 0.003) (siehe Abbildung 3). Post hoc analyses (Bonferroni-Holm
(1) = 11.33, P < 0.001, fm
2 = 0.001, fc
2 = 0.003, fc
Table 1.
Accuracy values (%) in all experimental conditions
TMS type
Effective TMS
TMS time
t1
Word
89.2 (3.2)
Pseudo-word
91.2 (2.3)
Word
90.9 (3.0)
Pseudo-word
93.3 (2.5)
Active diathesis
Passive diathesis
Sham TMS
t2
t1
t2
86.4 (5.0)
87.1 (3.8)
88.8 (4.8)
88.0 (2.8)
89.7 (3.3)
93.4 (2.5)
84.6 (4.2)
84.5 (4.0)
88.8 (3.7)
88.8 (2.4)
81.1 (4.9)
87.3 (3.3)
Note. The standard error of the mean is indicated in parentheses.
Neurobiology of Language
424
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
n
o
_
a
_
0
0
0
4
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Thematic reanalysis in the left PPS
Table 2.
Reaction times (ms) in all experimental conditions
TMS type
Effective TMS
TMS time
t1
Word
1,670 (74)
Pseudo-word
1,696 (73)
Word
1,764 (79)
Pseudo-word
1,836 (78)
Active diathesis
Passive diathesis
Sham TMS
t2
t1
t2
1,678 (79)
1,797 (76)
1,756 (64)
1,740 (72)
1,677 (67)
1,748 (80)
1,756 (71)
1,773 (71)
1,724 (71)
1,798 (58)
1,751 (73)
1,757 (78)
Note. The standard error of the mean is indicated in parentheses.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
n
o
_
a
_
0
0
0
4
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. TMS and DIATHESIS interaction. Asterisk denotes significant difference. Error bars rep-
resent standard deviation of the mean.
correction) indicated a significant difference between sham and effective TMS for passive
DIATHESIS (Chi2 (1) = 12.58, p < 0.001), but not for active DIATHESIS (Chi2 (1) = 2.07, p =
0.14). None of the other main effects or interactions reached significance (all ps > 0.08).
DISKUSSION
In a recent TMS study, Finocchiaro et al. (2015) showed increased comprehension accuracy
for passive sentences relative to active sentences following stimulation of the left posterior IPS.
The present study replicates and extends those findings, as the same facilitation for passives
was obtained when directly contrasting within-participant performance under sham and TMS
conditions in an agent-decision task. In line with Finocchiaro et al. (2015), we interpret this
result as showing that the posterior IPS is involved in thematic role assignment, speziell
when transiently encoded thematic roles need to be reanalyzed.
The main goal of the present paper was to check whether the semantics of the verb is crucial
for the facilitation of passive diathesis interpretation. Auf der einen Seite, the most credited linguistic
accounts posit the involvement of verb semantics in thematic role encoding (Dowty, 1989,
1991; Lebani et al., 2015; Levin & Rappaport Hovav, 2005; McRae et al., 1997; Van Valin,
1999). Andererseits, the PPS is not known to be devoted to semantic processing, and thus
the type of information relevant to the PPS may not concern verb-specific semantics. To inves-
tigate this issue, both sentences and pseudo-sentences were presented. Simple reversible
sentences in active and passive diathesis were randomly alternated with active and passive
pseudo-sentences, which contained real free-standing and bound grammatical markers but
pseudo-word roots. Critically, both passive sentences and passive pseudo-sentences were
Neurobiology of Language
425
Thematic reanalysis in the left PPS
facilitated, and the magnitude of the effect was comparable for the two stimulus types. It should be
noted that the main contrast on which our results are based is the comparison between the effects
of effective TMS applied to one single spot with the effects of sham TMS. In the present work we
focus on the posterior portion of the IPS, which was shown to be involved in diathesis processing in
Finocchiaro et al. (2015). The results reported here clearly show that the meaningfulness of the
verb is not a necessary precondition for the repetitive TMS induced facilitation of passive
Sätze. Perhaps the nature of the pseudo-sentences (pseudo-word roots, real grammatical
morphemes) allowed participants to recover a proto-meaning of the stimulus, according to
which someone did something to somebody else or someone suffered something because of
jemand anderes. In the absence of a full-verb’s semantics, participants may have treated the
meaningless arguments as “proto-agent” and “proto-patient” respectively, thus attributing to
them the features that are prototypically assigned to agents and patients/themes (Dowty 1989,
1991). In diesem Rahmen, one can tentatively conclude that the process of reanalysis, at least to
the extent to which it recruits the PPS, is relatively insensitive to verb-specific semantics (sehen
McRae et al., 1997). This does not imply that the semantics of the stimuli has no influence what-
soever on reanalysis; eher, it suggests that the left posterior IPS is insensitive to the semantic
characteristics of the stimuli that are relevant to the reanalysis process.
Remarkably, the TMS-induced facilitation for passives was not affected by stimulation tim-
ing. In der Tat, performance did not significantly change depending on whether double pulse
TMS was delivered at T1 (200–400 ms) or T2 (600–800 ms). It is commonly assumed that
the integration of the lexical, semantic, and syntactic elements of a simple sentence is com-
pleted between 500 Und 1,000 MS (Friederici, 2002). In the present experiment, stimulation
time was manipulated in order to explore the incremental processing of active and passive
(pseudo)sentences as it relates to the involvement of the left posterior IPS. Our results do
not shed light on this issue, probably for reasons related to the complexity of the experimental
Aufgabe. In der Tat, to complete each trial the participant had to read the sentence, analyze it, decide
which argument was the agent, and finalize their choice. The duration of each step may vary
across participants and even from trial to trial, thus leading to the huge variation in response
latencies observed across and within participants. Such variability may have hidden differen-
tial effects (wenn überhaupt) of the stimulation timing.
The polarity of the effect deserves a separate comment. As in Finocchiaro et al. (2015), In
the present experiment TMS improved accuracy on passive sentences. Finocchiaro et al.
(2015) accounted for the TMS-induced facilitation on passives by assuming that “the pos-
terior intraparietal site processes the relevant information after the TMS train is delivered”
(page 230). In light of the lack of effects of different TMS timings on performance, this expla-
nation is unlikely. It has been assumed that the neural and behavioral effects of TMS depend
on the state of the cortex being stimulated (Pasley et al., 2009; Perini et al., 2012; Silvanto
et al., 2008). The TMS of neurons with lower-than-average firing patterns is more likely to
facilitate behavior, whereas the TMS of neurons with hyper-active firing patterns is more likely
to disrupt it. This model has been repeatedly supported in the literature in experimental designs
in which the excitability of a given neural population is manipulated by adaptation or priming
Verfahren (Barchiesi et al., 2012; Cattaneo, 2010; Cattaneo et al., 2010, 2011; Mazzoni et al.,
2017; Silvanto et al., 2007). On this account, the polarity of TMS effects is unpredictable
unless the initial cortex excitability is known. Since the present experiment was not designed
to highlight state-dependent effects, no clear conclusion can be drawn on this issue.
Our data show that TMS-induced facilitation for passive sentences and pseudo-sentences is
not tied to full-verb semantics, and that it is probably linked to the fact that interpreting passive
sentences requires the reanalysis of thematic roles. This effect could be interpreted purely in
Neurobiology of Language
426
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
terms of working memory. In der Tat, many studies have shown that the posterior parietal region
is involved in the encoding of thematic roles as well as in working memory processes in sen-
tences presented both auditorily and visually (Papagno et al., 2007; Romero Lauro et al., 2006;
Romero Lauro et al., 2010). The data also show that area BA40 is strictly tied to the phono-
logical loop (Romero Lauro et al., 2010). The task used in the experiment discussed here re-
quires participants to keep phonological information active by relying on their phonological
store (Baldo & Dronkers, 2006). Jedoch, an account based entirely on working memory can-
not accommodate our results. On the reasonable assumption that working memory demands
are greater when processing pseudo-sentences, which are more difficult to keep in memory
than real sentences, such an account would predict a difference between passive sentences
and passive pseudo-sentences, with pseudo-sentences showing a greater facilitation effect.
Another potential account of the observed effects could stem from consideration of the key
role of the posterior portion of the parietal lobe in encoding spatial relations (Amorapanth
et al., 2010; Baumann & Mattingley, 2014). It has been proposed that thematic relations
can be translated into non-linguistic spatial representations (Chatterjee et al., 1995; Coslett,
1999). On this view, the effect putatively attributed to thematic reanalysis could be reduced
to a spatial effect. At face value, this possibility is unlikely, as we expressly avoided using verbs
or prepositions that denote relative positions or trajectory (z.B., precede, folgen, in front of,
behind, über, below). Darüber hinaus, the argument hardly applies to pseudo-sentences, welche
do not convey any obvious spatial content.
Gesamt, an account of the role of left PPS in sentence processing in terms of thematic role
encoding fits better with evidence from neuropsychological (Thothathiri et al., 2012) and fMRI
Studien (Mack et al., 2013; Meltzer-Asscher et al., 2013; Richardson et al., 2010; Thompson
et al., 2010, 2013) showing the involvement of temporo-parietal regions in the processing of
thematic roles, particularly for reversible and passive sentences. Zum Beispiel, a correlation
between impaired performance on reversible sentences and temporo-parietal damage was ob-
served in aphasic participants by Magnusdottír et al. (2013), Rogalsky et al. (2017) Und
Thothathiri et al. (2012). Darüber hinaus, Thothathiri et al. (2012) also showed that difficulty with
thematic roles persists after controlling for canonical/noncanonical word order and covarying
for working memory. In combination with these data, the present results add information on
the dynamics of the sentence comprehension process, via a technique that allows a fine-
grained temporal resolution during an agent-decision task.
We applied TMS to the posterior third of the left IPS. This region is referred to as superior
parietal-occipital cortex in humans and is considered homologous to areas V6A and PEc in
nonhuman primates. The posterior portion of IPS has been dubbed as the “parietal reach area,”
based on its key role in behavioral tasks (Battaglia-Mayer et al., 2014; Galletti & Fattori, 2018;
Grefkes & Fink, 2005). It is embedded in the “dorsal” visual stream (Ungerleider & Mishkin,
1982), which encodes visual stimuli in distinct but mostly body-centered frames of reference.
Like all dorsal stream regions, it is assumed to encode visual information for goal-directed ac-
tion and transform it into targets of reaching movements. Owing to the integration of visual
and somatosensory information, it also presumably provides online feedback on reaching
Bewegungen (Gallivan & Culham, 2015; Grefkes & Fink, 2005; Fattori et al., 2012; Piserchia
et al., 2017). Consistent with this view, classical neuropsychology reports show that damage to
the posterior portion of the IPS and the superior parietal lobule results in optic ataxia (z.B.,
Andersen et al., 2014), das ist, the inability to direct reaching movements to a half of a visual
Raum. Given the anatomic-functional aspects of posterior IPS, an intriguing question arises:
Why should an area unanimously deemed critical for visuospatial behavior be relevant for
the processing of passive sentences?
Neurobiology of Language
427
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
The anatomical localization of the TMS target is not entirely precise. We refer to it as the
posterior part of the IPS, but obviously TMS cannot reach the deeper part of its banks; daher,
the current data suffer from an anatomical bias toward the surface of the IPS region. This said,
the posterior-superior portion of IPS has been shown to be involved in nonmotor tasks, solch
as the processing of series of digits or letters (Bueti and Walsh, 2009; Eger et al., 2003; Jacob
& Nieder, 2008; Marshuetz et al., 2001; Schubotz et al., 2000; Schwenzer & Mathiak, 2011). A
core feature of all these tasks is the need to encode the serial order of the elements com-
prising the stimulus. Crucially, encoding the order of single elements is also a critical feature
of sentence processing. Daher, the effect reported here may be linked to the processing of
argument order, or more likely reflect the mapping from argument order to thematic roles,
systematically reversed in passive sentences.
In diesem Rahmen, the hypothesis that a cortical region that implements spatial representa-
tions also plays a critical role in syntactic processing is worth considering. Syntax relies strongly
on order. Scrambling word order relations in a sentence affects syntax, but not other
language functions. The human brain is well known to use its machinery flexibly, for appar-
ently heterogeneous functions. Based on the view that the brain can successfully accomplish
the “cultural recycling of cortical maps” (Dehaene & Cohen, 2007, P. 384), behavioral require-
ments that are relatively recent could be supported by neural hardware originally evolved for
different functions. Other cognitive domains in the human behavioral repertoire have been
linked to networks dedicated to other functions in nonhuman beings, as is the case of reading
(Dehaene & Cohen, 2011), time perception, or action understanding (Cattaneo, 2010;
Schubotz et al., 2000). The same may have happened for mechanisms involved in aspects of
syntactic processing that pose similar computational demands in nonlanguage contexts.
Limitations
Even though the findings are promising and in line with a previous TMS study, some method-
ological shortcomings suggest caution in the interpretation of the results and must be overcome
in future work. A first issue is related to the lack of a control site. Given that the main contrast is
between TMS and sham TMS when applied to one single spot, it cannot be completely ruled
out that the observed TMS effects are due to general arousal or alerting effects. Tatsächlich, sham
TMS, even though conducted according to conventional experimental practice, does not pro-
vide participants with the full sensory experience of effective TMS. In der Tat, applying TMS to just
one cortical spot without any active TMS control sites may lead to circular reasoning in which
the premises of the experiment turn out to be the conclusions (Cattaneo, 2018). This possibility
is partially mitigated by the consideration that the present experiment capitalizes on the results
of previous work in which the entire linear extension of the IPS had been mapped with 3 active
TMS spots, showing that passive sentences were affected only by stimulation of the spot selected
for the present experiment (Finocchiaro et al., 2015). Trotzdem, the presence of a control site would
strengthen the reliability of the data reported here. Other concerns stem from the limited sample
Größe (16 Teilnehmer) and the small number of trials in each experimental condition. In der Tat, für
each stimulation type (effective vs. sham) three variables were considered: semantics, diathesis,
and time (d.h., stimulation timing T1 vs. T2), each with two levels, thus leading to only 15 Versuche
per participant in each experimental condition. The low number of participants may undermine
the possibility of generalizing the observed effect to different participants and items and therefore
suggests caution. Tatsächlich, percentage scores in the region of 80–90% correspond to changes in
accuracy of only 1 oder 2 items per condition. Further studies with more participants, an increased
number of trials in each experimental condition, and an additional control site should assess the
reliability of the present findings.
Neurobiology of Language
428
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
ACKNOWLEDGMENTS
The authors wish to thank Martina Costamagna for her precious support in data collection.
FUNDING INFORMATION
The study was funded in part by a grant from PAT (Provincia Autonoma di Trento) to GM.
BEITRÄGE DES AUTORS
Chiara Finocchiaro: Konzeptualisierung: Equal; Ressourcen: Equal; Writing − original draft:
Lead; Writing − review and editing: Lead; Aufsicht: Lead. Luigi Cattaneo:
Konzeptualisierung: Equal; Formale Analyse: Lead; Ressourcen: Lead; Writing − original draft:
Equal; Writing − review and editing: Lead. Carlotta Lega: Formale Analyse: Lead; Ressourcen:
Equal; Writing − review and editing: Supporting. Gabriele Miceli: Konzeptualisierung: Equal;
Funding Acquisition: Lead; Writing − original draft: Equal; Writing − review and editing:
Supporting; Aufsicht: Equal. The first two authors (Chiara Finocchiaro and Luigi
Cattaneo) contributed to this project to an equal extent and thus share the role of first author.
VERWEISE
Amorapanth, P. X., Widjck, P., & Chatterjee, A. (2010). The neural
basis for spatial relations. Zeitschrift für kognitive Neurowissenschaften,
22(8), 1739–1753. https://doi.org/10.1162/jocn.2009.21322,
PubMed: 19642889
Andersen, R. A., Andersen, K. N., Hwang, E. J., & Hauschild, M.
(2014). Optic ataxia: From Balint’s syndrome to the parietal
reach region. Neuron, 81(5), 967–983. https://doi.org/10.1016/j
.neuron.2014.02.025, PubMed: 24607223
Baayen, R. H., Davidson, D. J., & Bates, D. M. (2008). Mixed-
effects modeling with crossed random effects for subjects and
Artikel. Journal of Memory and Language, 59(4), 390–412.
https://doi.org/10.1016/j.jml.2007.12.005
Baldo, J. V., & Dronkers, N. F. (2006). The role of inferior parietal
and inferior frontal cortex in working memory. Neuropsychology,
20(5), 529–538. https://doi.org/10.1037/0894-4105.20.5.529,
PubMed: 16938015
Barchiesi, G., Wache, S., & Cattaneo, L. (2012). The frames of
reference of the motor-visual aftereffect. PLOS ONE, 7(7),
e40892. https://doi.org/10.1371/journal.pone.0040892,
PubMed: 22848406
Bates, D., Maechler, M., Bolker, B., & Walker, S. (2015). Fitting linear
mixed-effects models using lme4. Journal of Statistical Software,
67(1), 1–48.
Battaglia-Mayer, A., Buiatti, T., Caminiti, R., Ferraina, S.,
Lacquaniti, F., & Shallice, T. (2014). Correction and suppression
of reaching movements in the cerebral cortex: Physiological and
neuropsychological aspects. Neurowissenschaften & Biobehavioral
Rezensionen, 42, 232–251. https://doi.org/10.1016/j.neubiorev
.2014.03.002, PubMed: 24631852
Baumann, O., & Mattingley, J. B. (2014). Dissociable roles of the
hippocampus and parietal cortex in processing of coordinate
and categorical spatial information. Frontiers in Human
Neurowissenschaften, 17(8), 73. https://doi.org/10.3389/fnhum.2014
.00073, PubMed: 24596551
Bornkessel, ICH., Zysset, S., Friederici, A. D., von Cramon, D. Y., &
Schlesewsky, M. (2005). Who did what to whom? The neural
basis of argument hierarchies during language comprehension.
NeuroImage, 26(1), 221–233. https://doi.org/10.1016/j
.neuroimage.2005.01.032, PubMed: 15862222
Bornkessel-Schlesewsky, ICH., & Schlesewsky, M. (2009). Die Rolle von
prominence information in the real-time comprehension of tran-
sitive constructions: A cross-linguistic approach. Language and
Linguistics Compass, 3(1), 19–58. https://doi.org/10.1111/j.1749
-818X.2008.00099.x
Bresnan, J. (2000). Lexical-functional syntax. Blackwell.
Brookshire, R. H., & Nicholas, L. E. (1980). Verification of active and
passive sentences by aphasic and nonaphasic subjects. Zeitschrift für
Speech, Language, and Hearing Research, 23(4), 878–893. https://
doi.org/10.1044/jshr.2304.878, PubMed: 7442218
Bueti, D., & Walsh, V. (2009). The parietal cortex and the represen-
tation of
Zeit, Raum, number and other magnitudes.
Philosophical transactions of the Royal Society of London.
Serie B, Biological Sciences, 364(1525), 1831–1840. https://doi
.org/10.1098/rstb.2009.0028, PubMed: 19487186
Caramazza, A., & Miceli, G. (1991). Selective impairment of thematic
role assignment in sentence processing. Brain and Language, 41(3),
402–436. https://doi.org/10.1016/0093-934X(91)90164-V,
PubMed: 1933265
Cattaneo, L. (2010). Tuning of ventral premotor cortex neurons to
distinct observed gras p t ypes: A TMS-primin g stu dy.
Experimentelle Hirnforschung, 207(3–4), 165–172. https://doi.org
/10.1007/s00221-010-2454-5, PubMed: 20963579
Cattaneo, L. (2018). Fancies and fallacies of spatial sampling with
transcranial magnetic stimulation (TMS). Grenzen in der Psychologie,
9, 3371. https://doi.org/10.3389/fpsyg.2018.01171, PubMed:
30026721
Cattaneo, L., Barchiesi, G., Tabarelli, D., Arfeller, C., Sato, M., &
Glenberg, A. M. (2011). One’s motor performance predictably
modulates the understanding of others’ actions through adapta-
tion of premotor visuo-motor neurons. Social Cognitive and
Affective Neuroscience, 6(3), 301–310. https://doi.org/10.1093
/scan/nsq099, PubMed: 21186167
Cattaneo, L., Sandrini, M., & Schwarzbach, J. (2010). Zustand-
dependent TMS reveals a hierarchical representation of observed
Neurobiology of Language
429
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
acts in the temporal, parietal, and premotor cortices. Zerebral
Kortex, 20(9), 2252–2258. https://doi.org/10.1093/cercor
/bhp291, PubMed: 20051360
Chatterjee, A., Maher, L. M., Rothi, L. J. G., & Heilman, K. M.
(1995). Asyntactic thematic role assignment: The use of a
temporal-spatial strategy. Brain and Language, 49(2), 125–139.
https://doi.org/10.1006/brln.1995.1024, PubMed: 7648248
Chomsky, N. (1965). Aspects of the theory of syntax. MIT Press.
Chomsky, N. (1981). Lectures on government and binding. Foris.
Coslett, H. B. (1999). Spatial influences on motor and language
Funktion. Neuropsychologie, 37(6), 695–706. https://doi.org/10
.1016/S0028-3932(98)00116-X, PubMed: 10390031
Dehaene, S., & Cohen, L. (2007). Cultural recycling of cortical
maps. Neuron, 56(2), 384–398. https://doi.org/10.1016/j.neuron
.2007.10.004, PubMed: 17964253
Dehaene, S., & Cohen, L. (2011). The unique role of the visual
word form area in reading. Trends in den Kognitionswissenschaften, 15(6),
254–262. https://doi.org/10.1016/j.tics.2011.04.003, PubMed:
21592844
De Rosario-Martinez, H., Fuchs, J., & R Core Team. (2015). phia:
Post-Hoc Interaction Analysis. (Version 0.2-0) R Foundation for
Statistical Computing, Vienna, Österreich. https://cran.r-project.org
/web/packages/phia/index.html
Dowty, D. (1989). On the semantic content of the notion “thematic
role.” In G. Chierchia, B. H. Partee, & R. Turner (Hrsg.), Properties,
types and meanings (Bd. II, S. 69–130). Kluwer. https://doi.org
/10.1007/978-94-009-2723-0_3
Dowty, D. (1991). Thematic proto-roles and argument selection.
Language, 67(3), 547–619. https://doi.org/10.1353/ lan.1991
.0021, https://doi.org/10.2307/415037
Eger, E., Sterzer, P., Russ, M. O., Giraud, A. L., & Kleinschmidt, A.
(2003). A supramodal number representation in human intrapar-
ietal cortex. Neuron, 37(4), 719–725. https://doi.org/10.1016
/S0896-6273(03)00036-9, PubMed: 12597867
Fattori, P., Breveglieri, R., Raos, V., Bosco, A., & Galletti, C.
(2012). Vision for action in the macaque medial posterior
parietal cortex. Zeitschrift für Neurowissenschaften, 32(9), 3221–3234.
https://doi.org/10.1523/ JNEUROSCI.5358-11.2012, PubMed:
22378893
Ferreira, F. (2003). The misinterpretation of noncanonical sentences.
Cognitive Psychology, 47, 164–203. https://doi.org/10.1016/S0010
-0285(03)00005-7, PubMed: 12948517
Fillmore, C. J. (1968). The case for case. In E. Bach & R. T. Harms
(Hrsg.), Universals in linguistics theory (S. 1–88). Holt, Rinehart,
and Winston.
Finocchiaro, C., Capasso, R., Cattaneo, L., Zuanazzi, A., & Miceli, G.
(2015). Thematic role assignment in the posterior parietal cortex:
A TMS study. Neuropsychologie, 77, 223–232. https://doi.org/10
.1016/ J.NEUROPSYCHOLOGIA.2015.08.025, PubMed:
26318240
Friederici, A. D. (1995). The time course of syntactic activation
during language processing: A model based on neuropsycho-
logical and neurophysiological data. Brain and Language, 50(3),
259–281. https://doi.org/10.1006/ brln.1995.1048, PubMed:
7583190
Friederici, A. D. (2002). Towards a neural basis of auditory sen-
tence processing. Trends in Cognitive Science, 6(2), 78–84.
https://doi.org/10.1016/S1364-6613(00)01839-8, PubMed:
15866191
Friederici, A. D., & Kotz, S. A. (2003). The brain basis of syntactic
processes: Functional imaging and lesion studies. NeuroImage,
20(S1), 8–17. https://doi.org/10.1016/j.neuroimage.2003.09
.003, PubMed: 14597292
Galletti, C., & Fattori, P. (2018). The dorsal visual stream revisited:
Stable circuits or dynamic pathways? Kortex, 98, 203–217.
https://doi.org/10.1016/j.cortex.2017.01.009, PubMed:
28196647
Gallivan, J. P., & Culham, J. C. (2015). Neural coding within human
brain areas involved in actions. Aktuelle Meinung in der Neurobiologie,
33, 141–149. https://doi.org/10.1016/j.conb.2015.03.012,
PubMed: 25876179
Garnsey, S. M., Pearlmutter, N. J., Myers, E., & Lotocky, M. A.
(1997). The contributions of verb bias and plausibility to the
comprehension of temporarily ambiguous sentences. Zeitschrift für
Memory and Language, 37, 58–93. https://doi.org/10.1006/jmla
.1997.2512
Grefkes, C., & Fink, G. R. (2005). The functional organization of the
intraparietal sulcus in humans and monkeys. Journal of Anatomy,
207(1), 3–17. https://doi.org/10.1111/j.1469-7580.2005.00426.x,
PubMed: 16011542
Grodzinsky, Y. (2000). The neurology of syntax: Language use
without Broca’s area. Behavioral and Brain Sciences, 23(1),
1–21. https://doi.org/10.1017/S0140525X00002399, PubMed:
11303337
Herwig, U., Satrapi, P., & Schönfeldt-Lecuona, C. (2003). Using the
international 10–20 EEG system for positioning of transcranial
magnetic stimulation. Brain Topography, 16(2), 95–99. https://
doi.org/10.1023/ B:BRAT.0000006333.93597.9D, PubMed:
14977202
Jacob, S. N., & Nieder, A. (2008). The ABC of cardinal and ordinal
number representations. Trends in den Kognitionswissenschaften, 12(2),
41–43. https://doi.org/10.1016/j.tics.2007.11.006, PubMed:
18178515
Kabdebon, C., Leroy, F., Simmonet, H., Perrot, M., Dubois, J., &
Dehaene-Lambertz, G. (2014). Anatomical correlations of the inter-
National 10-20 sensor placement system in infants. NeuroImage,
99, 342–356. https://doi.org/10.1016/j.neuroimage.2014.05.046,
PubMed: 24862070
Koessler, L., Maillard, L., Benhadid, A., Vignal, J. P., Felblinger, J.,
Vespignani, H., & Braun, M. (2009). Automated cortical projec-
tion of EEG sensors: Anatomical correlation via the international
10-10 System. NeuroImage, 46(1), 64–72. https://doi.org/10
.1016/j.neuroimage.2009.02.006, PubMed: 19233295
Lebani, G. E., Bondielli, A., & Lenci, A. (2015). You are what you do.
An empirical characterization of the semantic content of the
thematic roles for a group of Italian verbs. Zeitschrift für Kognition
Wissenschaft, 16(4), 401–430. https://doi.org/10.17791/jcs.2015.16
.4.401
Levin, B., & Rappaport Hovav, M. (2005). Argument realization.
C a m b r i d g e U n i v e r s i t y P r e s s . h t t p s : / / d o i . o r g / 1 0 . 1 0 1 7
/CBO9780511610479
Mack, J. E., Meltzer-Asscher, A., Barbieri, E., & Thompson, C. K.
(2013). Neural correlates of processing passive sentences. Gehirn
S c i e n c e s , 3 ( 3 ) , 1 1 9 8 – 1 2 1 4 . h t t p s : / / d o i . o r g / 1 0 . 3 3 9 0
/brainsci3031198, PubMed: 24961525
Magnusdottír, S., Fillmore, P., den Ouden, D. B., Hjaltason, H.,
Rorden, C., Kjartansson, O., Bonilha, L., & Fridriksson, J.
(2013). Damage to left anterior temporal cortex predicts impair-
ment of complex syntactic processing: A lesion-symptom map-
ping study. Kartierung des menschlichen Gehirns, 34(10), 2715–2723. https://
doi.org/10.1002/hbm.22096, PubMed: 22522937
Marshuetz, C., Schmied, E. E., Jonides, J., DeGutis, J., & Chenevert,
T. L. (2001). Order information in working memory: fMRT
evidence for parietal and prefrontal mechanisms. Zeitschrift für
Cognitive Neuroscience, 12(S2), 130–144. https://doi.org/10
.1162/08989290051137459, PubMed: 11506653
Neurobiology of Language
430
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
Mazzoni, N., Jacobs, C., Venuti, P., Silvanto, J., & Cattaneo, L.
(2017). State-dependent TMS reveals representation of affective
body movements in the anterior intraparietal cortex. Zeitschrift für
Neurowissenschaften, 37(30), 7231–7239. https://doi.org/10.1523
/JNEUROSCI.0913-17.2017, PubMed: 28642285
McElree, B., & Griffith, T. (1995). Syntactic and thematic processes in
sentence comprehension: Evidence for a temporal dissociation.
Journal of Experimental Psychology: Learning, Memory, Und
Cognition, 21, 134–157. https://doi.org/10.1037/0278-7393.21
.1.134
McRae, K., Ferretti, T. R., & Amyote, L. (1997). Thematic roles
as verb-specific concepts. Language and Cognitive Processes,
12(2-3), 137–176. https://doi.org/10.1080/016909697386835
Meltzer-Asscher, A., Schuchard,
J., den Ouden, D.-B., &
Thompson, C. K. (2013). The neural substrates of complex
argument structure representations: Processing “alternating
transitivity” verbs. Language and Cognitive Processes, 28(8),
1154–1168. https://doi.org/10.1080/01690965.2012.672754,
PubMed: 26139954
Meyer, A. M., Mack, J. E., & Thompson, C. K. (2012). Tracking pas-
sive sentence comprehension in agrammatic aphasia. Zeitschrift für
Neurolinguistics, 25(1), 31–43. https://doi.org/10.1016/j
.jneuroling.2011.08.001, PubMed: 22043134
Nakagawa, S., & Schielzeth, H. (2013). A general and simple
method for obtaining R2 from generalized linear mixed-effects
Modelle. Methods in Ecology and Evolution, 4, 133–142. https://
doi.org/10.1111/j.2041-210x.2012.00261.x
Okamoto, M., Und, H., Sakamoto, K., Takeo, K., Shimizu, K.,
Kohno, S., Oda, ICH., Isobe, S., Suzuki, T., Kohyama, K., & Und,
ICH. (2004). Three-dimensional probabilistic anatomical cranio-
cerebral correlation via the international 10-20 system oriented
for transcranial functional brain mapping. NeuroImage, 21(1),
99–111. https://doi.org/10.1016/j.neuroimage.2003.08.026,
PubMed: 14741647
Oldfield, R. C. (1971). The assessment and analysis of handed-
ness: The Edinburg inventory. Neuropsychologie, 9(1), 97–113.
https://doi.org/10.1016/0028-3932(71)90067-4, PubMed:
5146491
Papagno, C., Cecchetto, C., Reati, F., & Bello, L. (2007). Processing
of syntactically complex sentences relies on verbal short-term
Erinnerung: Evidence from a short-term memory patient. Kognitiv
Neuropsychology, 24(3), 292–311. https://doi.org/10.1080
/02643290701211928, PubMed: 18416493
Pasley, B. N., Allen, E. A., & Freeman, R. D.
(2009). Zustand-
dependent variability of neuronal responses to transcranial mag-
netic stimulation of the visual cortex. Neuron, 62(2), 291–303.
https://doi.org/10.1016/j.neuron.2009.03.012, PubMed:
19409273
Perini, F., Cattaneo, L., Carrasco, M., & Schwarzbach, J. V. (2012).
Occipital transcranial magnetic stimulation has an activity-
dependent suppressive effect. Zeitschrift für Neurowissenschaften, 32(36),
12361–12365. https://doi.org/10.1523/ JNEUROSCI.5864-11
.2012, PubMed: 22956826
Piserchia, V., Breveglieri, R., Hadjidimitrakis, K., Bertozzi, F.,
Galletti, C., & Fattori, P. (2017). Mixed body/ hand reference
frame for reaching in 3D space in macaque parietal area PEc.
Hirnrinde, 27(3), 1976–1990. https://doi.org/10.1093
/cercor/bhw039, PubMed: 26941385
Pollard, C., & Sag, ICH. A. (1994). Head-driven phrase structure grammar.
University of Chicago Press.
R Development Core Team. (2016). R: A language and environment
for statistical computing. R Foundation for Statistical Computing,
Vienna, Österreich. https://www.R-project.org/
Richardson, F. M., Thomas, M. S. C., & Price, C. J. (2010). Neuronal
activation for semantically reversible sentences. Zeitschrift für
Cognitive Neuroscience, 22(6), 1283–1298. https://doi.org/10
.1162/jocn.2009.21277, PubMed: 19445603
Rogalsky, C., LaCroix, A. N., Chen, K. H., Anderson, S. W.,
Damasio, H., Liebe, T., & Hickok, G. (2017). The neurobiology
of agrammatic sentence comprehension: A lesion study. Zeitschrift
of Cognitive Neuroscience, 30(2), 234–255. https://doi.org/10
.1162/jocn_a_01200, PubMed: 29064339
Romero Lauro, L. J., Reis, J., Cohen, L. G., Cecchetto, C., &
Papagno, C. (2010). A case for the involvement of phonological
loop in sentence comprehension. Neuropsychologie, 48(14),
4003–4011. https://doi.org/10.1016/j.neuropsychologia.2010.10
.019, PubMed: 20969883
Romero Lauro, L. J., Walsh, V., & Papagno, C. (2006). The neural
correlates of phonological short term memory: A repetitive trans-
cranial magnetic stimulation study. Zeitschrift für Kognition
Neurowissenschaften, 18(7), 1147–1155. https://doi.org/10.1162/jocn
.2006.18.7.1147, PubMed: 16839288
Schubotz, R. ICH., Friederici, A. D., & von Cramon, D. Y. (2000). Time
perception and motor timing: A common cortical and subcortical
basis revealed by fMRI. NeuroImage, 11(1), 1–12. https://doi.org
/10.1006/nimg.1999.0514, PubMed: 10686112
Schwenzer, M., & Mathiak, K. (2011). Numeric aspects in pitch
identification: Eine fMRT-Studie. BMC Neuroscience, 12, Article
26. https://doi.org/10.1186/1471-2202-12-26, PubMed:
21392373
Selya, A. S., Rose, J. S., Dierker, L. C., Hedeker, D., & Mermelstein,
R. J. (2012). A practical guide to calculating Cohen’s f 2, A
measure of local effect size, from PROC MIXED. Grenzen in
Psychologie, 3, 111. https://doi.org/10.3389/fpsyg.2012.00111,
PubMed: 22529829
Silvanto, J., Muggleton, N. G., Cowey, A., & Walsh, V. (2007).
Neural adaptation reveals state-dependent effects of transcranial
magnetic stimulation. Europäisches Journal für Neurowissenschaften, 25(6),
1874–1881. https://doi.org/10.1111/j.1460-9568.2007.05440.x,
PubMed: 17408427
Silvanto, J., Muggleton, N., & Walsh, V. (2008). State-dependency
in brain stimulation studies of perception and cognition. Trends
in Cognitive Sciences, 12(12), 447–454. https://doi.org/10.1016/j
.tics.2008.09.004, PubMed: 18951833
Snijders, T. A., & Bosker, R. J. (1994). Modeled variance in two-level
Modelle. Sociological Methods & Forschung, 22(3), 342–363.
https://doi.org/10.1177/0049124194022003004
Sparing, R., Hesse, M. D., & Fink, G. R. (2010). Neuronavigation for
transcranial magnetic stimulation (TMS): Where we are and
where we are going. Kortex, 46(1), 118–120. https://doi.org/10
.1016/j.cortex.2009.02.018, PubMed: 19371865
Thompson, C. K., Bonakdarpour, B., & Fix, S. C. (2010). Neuronal
mechanisms of verb argument structure processing in agram-
matic aphasic and healthy age-matched listeners. Zeitschrift für
Cognitive Neuroscience, 22(9), 1993–2011. https://doi.org/10
.1162/jocn.2009.21334, PubMed: 19702460
Thompson, C. K., Riley, E. A., den Ouden, D.-B., Meltzer-
Asscher, A., & Lukic, S. (2013). Training verb argument struc-
ture production in agrammatic aphasia: Behavioral and neural
recovery patterns. Kortex, 49(9), 2358–2376. https://doi.org/10
.1016/j.cortex.2013.02.003, PubMed: 23514929
Thothathiri, M., Kimberg, D. Y., & Schwartz, M. F. (2012). The neural
basis of reversible sentence comprehension: Evidence from voxel-
based lesion-symptom mapping in aphasia. Zeitschrift für Kognition
Neurowissenschaften, 24(1), 212–222. https://doi.org/10.1162/jocn_a
_00118, PubMed: 21861679
Neurobiology of Language
431
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Thematic reanalysis in the left PPS
Turner, E. A., & Rommetveit, R. (1967). The acquisition of sentence
voice and reversibility. Kinderentwicklung, 38(3), 649–660.
https://doi.org/10.2307/1127243, PubMed: 6049629
Ungerleider, L. G., & Mishkin, M. (1982). Two cortical visual sys-
Systeme. In D. J. Ingle, M. A. Goodale, & R. J. W. Mansfield (Hrsg.),
Analysis of visual behavior (S. 549–586). MIT Press.
Van Valin, R. D. (1999). Generalized semantic roles and the
syntax-semantics interface. In F. Corbin, C. Dobrovie-Sorin, &
J. M. Marandin (Hrsg.), Empirical issues in formal syntax and seman-
Tics (Bd. 2, S. 373–389). Theus.
Yoo, S. S., Guttmann, C. R. G., Ives, J. R., Panych, L. P., Kikinis, R.,
Schomer, D. L., & Jolesz, F. A. (1997). 3D localization of surface
10-20 EEG electrodes on high resolution anatomical MR images.
Electroencephalography and Clinical Neurophysiology, 102(4),
335–339. https://doi.org/10.1016/S0013-4694(96)95088-9,
PubMed: 9146495
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
4
1
6
1
9
5
8
6
7
3
N
Ö
_
A
_
0
0
0
4
3
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurobiology of Language
432