RESEARCH ARTICLE
Effect of Bilateral Opercular Syndrome
on Speech Perception
Keine offenen Zugänge
Tagebuch
Grant M. Walker1, Patrick Sarahan Rollo2
, Nitin Tandon2, and Gregory Hickok1,3
1Department of Cognitive Sciences, Universität von Kalifornien, Irvine
2Department of Neurosurgery, University of Texas Medical School at Houston
3Department of Language Science, Universität von Kalifornien, Irvine
Schlüsselwörter: speech perception, sensorimotor integration, audiovisual integration, bilateral surgical
Resektion, opercular syndrome
ABSTRAKT
Speech perception ability and structural neuroimaging were investigated in two cases of
bilateral opercular syndrome. Due to bilateral ablation of the motor control center for
the lower face and surrounds, these rare cases provide an opportunity to evaluate the
necessity of cortical motor representations for speech perception, a cornerstone of some
neurocomputational theories of language processing. Speech perception, einschließlich
audiovisual integration (d.h., the McGurk effect), was mostly unaffected in these cases,
although verbal short-term memory impairment hindered performance on several tasks that
are traditionally used to evaluate speech perception. The results suggest that the role of the
cortical motor system in speech perception is context-dependent and supplementary, nicht
inherent or necessary.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
EINFÜHRUNG
Despite centuries of discussion and debate, the role of the motor system in speech perception
continues to inspire curiosity and empirical investigations. More recently, the discovery of
mirror neurons has renewed interest in motor theories of speech perception. These theories posit
that motoric gesture representations play either a primary or supportive role in perceiving speech
sounds via articulatory recoding (motoric simulation), thereby assisting the resolution of the
indeterminacy problem in mapping from acoustic to phonological perception. Consistent with
this theory, functional imaging research has shown conclusively that frontal motor speech-
related areas activate during speech listening (Hickok et al., 2003; Watkins & Paus, 2004;
Wilson et al., 2004) and transcranial magnetic stimulation (TMS) of premotor and primary motor
areas can modulate performance on some perception tasks in an effector-specific manner
(D’Ausilio et al., 2009; Möttönen & Watkins, 2012; Pulvermüller & Ermüdung, 2010). For in depth
Bewertungen, Diskussion, and debate see Galantucci et al. (2006), Gallese et al. (2011), Hickok
(2014), Hickok et al. (2011), Liebenthal and Möttönen (2018), Massaro and Chen (2008),
Skipper et al. (2017).
Nowadays, researchers generally agree that a strong version of the motor theory—that
motor speech systems are necessary for speech perception—is untenable. This conclusion is
based on several facts. Erste, functional imaging evidence, where there is robust evidence for
Zitat: Walker, G. M., Rollo, P. S.,
Tandon, N., & Hickok, G. (2021). Effect
of bilateral opercular syndrome on
speech perception. Neurobiology of
Language, 2(3), 335–353. https://doi.org
/10.1162/nol_a_00037
DOI:
https://doi.org/10.1162/nol_a_00037
zusätzliche Informationen:
https://doi.org/10.1162/nol_a_00037
Erhalten: 28 April 2020
Akzeptiert: 23 Marsch 2021
Konkurrierende Interessen:
The authors have declared that
no competing interests exist.
Korrespondierender Autor:
Grant M. Walker
grantw@uci.edu
Handling-Editor:
Kate Watkins
Urheberrechte ©: © 2021
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
Speech perception in opercular syndrome
Sublexical:
Refers to language representations that
are smaller than a word, wie zum Beispiel
syllables or individual speech sounds.
Anarthria:
The complete inability to control the
muscles for articulating speech.
Wada procedures:
Involve injecting anesthetic into one
hemisphere of the brain to deactivate it.
Operculum:
A region on the lateral surface of
the brain that includes portions of
primary motor and sensory areas.
motor activation during perception, Ist, nonetheless, only correlational and cannot speak to the
causal involvement of motor areas in perception. Zweite, while TMS has provided evidence
of causation, these effects are typically small and only evident under noisy listening conditions
or during tasks requiring explicit attention to sublexical phonological features (D’Ausilio et al.,
2012; Sato et al., 2009), which are known to doubly dissociate from phonological ability
under more ecologically valid conditions (Hickok & Kacke, 2000, 2004, 2007). Dritte, evi-
dence from the speech perception abilities of pre-lingual infants (Eimas et al., 1971), Menschen
with anarthria due to cerebral palsy (Bishop et al., 1990), and non-human animals (Kuhl &
Müller, 1975) shows that a functioning motor speech system is not necessary for speech per-
ception. Vierte, unilateral damage to motor speech areas, which can substantially disrupt
speech fluency, does not produce a substantial (or in some cases any) concomitant deficit
in speech perception (Rogalsky et al., 2011). Endlich, acute unilateral deactivation of the left
hemisphere causing complete expressive mutism during Wada procedures leaves word
comprehension relatively spared (Hickok et al., 2008).
In light of such observations, most researchers promote a more nuanced view of the role of
the motor system in receptive speech, arguing for a modulatory role that may engage, für
Beispiel, under noisy listening conditions (Hickok et al., 2011; Skipper et al., 2017; Wilson,
2009). Entsprechend, the research questions are becoming more nuanced as well, with an aim
toward quantifying the magnitude of motor influence and specifying the condition(S) unter
which it holds (Skipper et al., 2017). Zum Beispiel, Stokes et al. (2019) used a behavioral psy-
chometric approach to estimate the effect size of motor interference (articulatory suppression)
on minimal pair perception in noise. They reported that motor interference reduced speech
perception under noisy conditions by an average of approximately 1 dB, which is to say that
increasing the stimulus volume by just 1 dB was enough to overcome the perceptual decrement
of motor suppression. In two studies, another group compared the effects of TMS stimulation
under two different task conditions, a sublexical task (4-alternative forced-choice syllable iden-
tification) (D’Ausilio et al., 2009) and a word comprehension task (Schomers et al., 2015).
(Recall that these tasks are known to dissociate, as mentioned above.) Motor stimulation signif-
icantly increased errors only for the sublexical task, resulting in between 5–10% change in
accuracy under noisy conditions, which approximates a 1 dB size effect (Stokes et al., 2019).
The effects of motor stimulation during word comprehension affected only reaction times but
not accuracy. Daher, existing evidence suggests that motor modulation during speech percep-
tion has an effect size of approximately 1 dB, holding only under noisy listening conditions, Und
may be less during normal comprehension compared to artificial laboratory tasks.
The present study contributes to this body of evidence via a detailed study of two cases of
opercular syndrome. Clinical symptoms of opercular syndrome include severe disruption of
voluntary control of the orofacial, lingual, pharyngeal, and masticatory muscles. Speech artic-
ulation is severely impaired, often to the point of mutism, despite automatic control of this mus-
culature being relatively spared (Desai et al., 2013; Groswasser et al., 1988; Milanlioglu et al.,
2013; Silveri et al., 2017). The syndrome is typically associated with bilateral lesions of the
frontal operculum, including speech-related zones, although unilateral and subcortical lesions
have also been documented (Bakar et al., 1998; Starkstein et al., 1988). Various etiologies, solch
as epileptic disorders, cerebrovascular events, degenerative diseases, or CNS infections, can
cause opercular syndrome, which can be acquired or congenital, and persistent or intermittent.
The syndrome is also known as Foix-Chavany-Marie syndrome, and the developmental form in
children is known as Worcester-Drought syndrome (Christen et al., 2007). Bilateral perisylvian
polymicrogyria is a developmental condition that also can lead to some of the symptoms
observed in opercular syndrome, often severely affecting speech production and speech
Neurobiology of Language
336
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
perception together, but without severely compromising other cognitive abilities (d.h., IQ); Wie-
immer, polymicrogyria is generally not specific to motor systems and often affects sensory cortices
in the perisylvian region as well (Boscariol et al., 2010; Jansen et al., 2005; Saletti et al., 2007).
Opercular syndrome due to surgical resection, as we report here, offers a unique opportunity to
study the effects of severe and specific motor speech disruption due to bilateral cortical/
subcortical lesions on receptive speech ability.
Two patients who had been clinically diagnosed with opercular syndrome based on clinical
examination, medical reports, and neuroimaging were referred to us for an in-depth assessment
by the neurosurgeon managing their current care. During multiple visits over the course of
several months, the patients completed informal interviews and a comprehensive examination
of their receptive speech abilities, as well as undergoing new, structural neuroimaging with MRI
to provide a precise delineation of the surgical lesions’ anatomical extent. In our discussion of
these cases, we consider how the specific patterns of performance and anatomical disruption
bear on the limits of the motor system’s contributions to speech perception at the phonemic,
word, and sentence levels, as well as to audio-visual integration of speech signals.
MATERIALS AND METHODS
Teilnehmer
Two patients with opercular syndrome due to bilateral surgical resection were tested on speech
perception and language comprehension tasks. A control sample, selected for convenience,
included four neurologically healthy lab members (3 männlich, 1 weiblich), ranging 24–26 years in
Alter, who performed a subset of the same tasks under identical testing conditions as the patients.
Not all tasks were performed, as some of them were only administered to one of the patients during
post-hoc, follow-up investigation of observed behavioral effects. All study procedures were
performed after approval from the UT Health Committee for the Protection of Human Subjects.
All subjects provided written informed consent. For details about the patients’ motor and commu-
nicative abilities at time of testing, see Additional Observations in the Results section.
Case history 1
Patient is a 27-year-old female with a history of seizures that began at age 15. The patient’s intrac-
table epilepsy was managed by a pediatric neurosurgeon (not the author, N. T.) at the age of 16.
Prior to surgery, the Weschler Abbreviated Scale of Intelligence (WASI) indicated average Full
Scale IQ (100), average Verbal IQ (93), and average Performance IQ (106), with noted weaknesses
in verbal fluency to semantic cueing, naming to confrontation, Arbeitsgedächtnis, sustained
attention, and speeded sequential processing. She underwent invasive seizure monitoring with
subdural grid electrodes and then several resective operations. Erste, a resection of right frontal
opercular epileptogenic focus and, 6 months later, an anterior 2/3 corpus callosotomy. The patient
then had an additional subdural grid implantation on the left side, followed by a left frontal
topectomy and left parietal sub-pial transections. Two months after this last resective operation,
the patient had a ventriculoperitoneal shunt placed and, at age 18, had a vagus nerve stimulation
device implanted. The patient continues to suffer from seizure disorder. No postoperative neuro-
psychological testing was done. At the time of enrolling in the current study, she had not spoken
In 9 Jahre, since her last surgery.
Case history 2
Patient is a 54-year-old female with a history of focal seizures that began when she was 12. Bei
the age of 18 she suffered a stroke secondary to the bleeding from a cavernous malformation of
Neurobiology of Language
337
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
the left side of the brain. Multiple other cavernous hemangiomas were identified in the brain at
Alter 22. The patient had a left frontal opercular cavernoma resected about 20 years ago. This led
to a marked impairment of her language function and then a recovery over many months. Sie
then had a hemorrhagic stroke in the right frontal lobe. A second resection was performed on the
left side at age 42. She has had a total of nine spontaneous episodes of hemorrhagic strokes. Der
patient continues to suffer from seizure disorder, with fewer seizures and less intense seizures
than when she was younger. She has a spastic hemiparesis on her left side. Preoperative neuro-
psychological exam results: On the WASI, she obtained a Full Scale IQ of 90. Her Performance
IQ (99) was much higher than her Verbal IQ (86). Achievement screening with the Wide Range
Achievement Test–3rd edition (Snelbaker et al., 2001) indicated average performance on a read-
ing subtest (Standard Score: 107, post-HS grade equivalent) and a spelling subtest (Standard
Score: 92, HS grade equivalent), and low average performance on a math subtest (Standard
Score: 83, sixth grade equivalent). Neuropsychological testing indicated relative reductions in ver-
bal fluency, fine motor speed and coordination, recall of rote verbal and nonverbal information fol-
lowing a delay, mental flexibility, and problem solving. She also reported moderate anxiety and
moderate-to-severe depression. No postoperative neuropsychological testing was done. At time of
enrolling in the current study, she had not spoken in 12 Jahre, since her last surgery.
Materials
Tests of receptive processing of phonemes included:
(1) Word Discrimination (Rogalsky et al., 2011)—The patient is auditorily presented with two
single-syllable words and instructed to judge whether they are the same or different. Stimuli
differed by only a single phonetic feature in the initial phoneme (d.h., minimal pairs, wie zum Beispiel
Might-Night or Face-Vase) and were matched for phonotactic frequency and phonological
neighborhood density. Es gab 20 “same” trials and 20 “different” trials.
(2) Nonword Discrimination (Rogalsky et al., 2011)—The patient is auditorily presented
with two single-syllable pseudowords and instructed to judge whether they are the same or
anders. As with the word discrimination task, stimuli differed by only a single phonetic fea-
ture in the initial phoneme, involved all of the same contrasts as in the word discrimination
Aufgabe, and were matched for phonotactic frequency and phonological neighborhood density.
Es gab 20 same trials and 20 different trials.
(3) Word-to-Picture Matching Phonological Foils (Rogalsky et al., 2011)—The patient is
auditorily presented with a probe word and instructed to choose which picture matches the
word from among four alternatives. The alternatives are all phonological distractors that rhyme
with the target. Es gab 20 total trials.
(4) Adaptive Word-to-Picture Matching (Stokes et al., 2019)—The patient was auditorily
presented with a synthetic one-syllable word embedded in white noise followed by visual pre-
sentation of two black-and-white drawings on the left and right side of a computer screen, Und
the patient was instructed to press a key on the keyboard to indicate the matching picture. Der
volume of the auditorily presented word was adaptive, decreasing after correct trials and
increasing after incorrect trials, while the volume of the white noise was constant. Es gab
200 trials per condition. There were four stimulus pair conditions ( pie-buy, tie-die, buy-die, pie-
tie). The order of auditory tokens from a given pair and the orientation of the pictures on the
screen were randomized. A logistic psychometric curve was fit to the data, and the signal-
to-noise ratio (dB) that yielded 75% performance accuracy is reported. Because dB is on a log
scale, a positive value indicates that the signal was louder than the noise, a negative value in-
dicates that the noise was louder than the signal, and zero indicates equal amplitudes for signal
Neurobiology of Language
338
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
and noise. Lower dB values indicate better performance (greater noise tolerance); the maximum
dB value was capped at 30 dB, indicating total failure to discriminate the target word, even with
relatively negligible background noise.
(5) Audiovisual Integration (Hickok et al., 2018)—In the auditory-only condition, the patient
was auditorily presented with different instances of a naturally spoken syllable by a male speaker
from the set {/pa/, /ka/} and instructed to point to the written syllable that matched among three
alternatives on a computer screen {/pa/, /ka/, /ta/}. Es gab 30 trials in this condition. In
the audiovisual condition, the patient was auditorily presented with a syllable from the set
{/pa/, /ka/} synchronously with a video of the same male speaker that was congruent or incon-
gruent (auditory /pa/ with visual /ka/), and the patient was instructed to point to the written
syllable that matched the auditory percept among three alternatives on a computer screen
{/pa/, /ka/, /ta/}. Incongruent stimuli were pilot tested to include instances that were most likely
to evoke a fused percept. Es gab 10 congruent /pa/ trials, 10 congruent /ka/ trials, Und 10
incongruent trials.
Tests of receptive processing of words included:
(1) Word-to-Picture Matching Mixed Foils (Rogalsky et al., 2011)—The patient is auditorily
presented with a probe word and instructed to choose which picture matches the word from
among four alternatives. The alternatives include a semantic distractor from the same taxo-
nomic category as the target, a phonological distractor that rhymes with the target, und ein
unrelated distractor. Es gab 20 total trials.
(2) Word-to-Picture Matching Mixed Foils in Noise (Rogalsky et al., 2011)—The patient is
auditorily presented with a probe word embedded in 14 dB Gaussian white noise and instructed
to choose which picture matches the word from among four alternatives. The alternatives
include a semantic distractor from the same taxonomic category as the target, a phonological
distractor that rhymes with the target, and an unrelated distractor. Es gab 20 total trials.
(3) Word-to-Picture Matching Semantic Foils (Rogalsky et al., 2011)—The patient is auditorily
presented with a probe word and instructed to choose which picture matches the word from
among four alternatives. The alternatives are all semantic distractors from the same taxonomic
category as the target. Es gab 20 total trials.
(4) The Western Aphasia Battery ( WAB; Kertesz, 1982), Auditory Word Recognition—The
patient is instructed to identify items from a given category by pointing. Categories include real
Objekte, drawn objects, shapes, letters, Zahlen, colors, furniture, body parts, fingers, right-left
body parts. Es gab 60 total items.
Tests of receptive processing of sentences included:
(1) WAB, Yes/No Questions—Questions are presented verbally to the patient with the answer
being either yes or no. Es gab 20 total questions.
(2) WAB, Sequential Commands—The patient is verbally presented with a command that
requires a sequence of two to four actions. Es gab 8 total commands involving 19 Aktionen.
(3) Sentence-to-Picture Matching (subject-relative, object-relative, active, and passive
[SOAP]; Liebe & Oster, 2002)—A sentence is verbally presented to the patient, and the patient
is instructed to choose which picture matches the sentence among three alternatives. The “se-
mantic distractor” alternative includes a different subject, Objekt, and action from the target,
while the “syntactic distractor” alternative reverses the roles of the subject and object. The sen-
tences had different syntactic constructions, including active voice, passive voice, Thema-
relative clauses, and object-relative clauses. Es gab 10 sentences of each type.
Neurobiology of Language
339
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
(4) Sentence-Picture Verification (SOAP)—Each of the subject-relative and object-relative
sentences from SOAP were presented verbally with either the correct picture or the syntactic
distractor picture, and the patient was instructed to judge whether the sentence matched the
picture.
(5) Sentence Plausibility (Rogalsky et al., 2018)—The patient is auditorily presented with a
sentence and instructed to judge whether it is semantically plausible. The sentences had
different syntactic constructions, including active voice, passive voice, subject-relative clauses,
and object-relative clauses. Es gab 20 sentences of each type.
Tests of short-term memory included:
(1) Digit Span (Forward)—The patient was auditorily presented with a sequence of non-
repeating digits and instructed to point to the same sequence on a number line. The list length,
beginning with just a single digit, increased until errors occurred on all trials. The number line
was not visible during presentation of the sequence. Es gab 4 trials per list length.
(2) Word Span—The patient was auditorily presented with a sequence of non-repeating, eins-
syllable words, selected from the set {bags, cage, dump, gash, king, mock, peach, rake, shoe,
tent}, and instructed to point to the same sequence on an alphabetically arranged response sheet.
The response sheet was not visible during presentation of the sequence. The list length, begin-
ning with just a single word, increased until errors occurred on all trials of a list length condition.
Es gab 4 trials per list length condition.
(3) Nonword Span—The patient was auditorily presented with a sequence of non-repeating,
one-syllable pseudowords, selected from the set {bav, coaf, dook, fave, giz, kag, mide, nabe,
perb, roash, tast}, and instructed to point to the same sequence on an alphabetically arranged
response sheet. The response sheet was not visible during presentation of the sequence. The list
Länge, beginning with just a single pseudoword, increased until errors occurred on all trials of a
list length condition. Es gab 4 trials per list length condition.
(4) 4-Digit Recall—The patient was presented with a sequence of four digits and instructed
to point to the sequence on a number line after a 15 second delay. The number line was not
visible during presentation of the sequence or during the maintenance interval. Es gab 4
total trials.
Auxiliary tests included:
(1) WAB, Picture Description—The patient is presented with a picture of a picnic scene and
instructed to describe what they see. Responses were written.
(2) WAB, Object Naming—The patient is presented with a set of real objects and instructed
to name them. Responses were written due to mutism. Es gab 15 total objects. The safety
pin, eraser, padlock, pipe, and matches were not presented. The gun was replaced by a mirror,
and the pencil was replaced by a pen.
(3) WAB, Reading Comprehension of Sentences—The patient is presented with a written
Satz (or pair of sentences) with the final word elided, and the patient is instructed to
choose the final word from a set of four alternatives. Es gab 8 total sentences.
(4) WAB, Writing to Dictation—The patient is verbally presented with the sentence, “Pack
my box with five dozen jugs of liquid veneer.” The patient is instructed to write the sentence.
(5) WAB, Apraxia—The patient is instructed to pantomime actions with the upper limb,
Gesicht, imagined instruments, and imagined complex situations such as driving a car. Dort
war 20 total actions.
Neurobiology of Language
340
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
Apraxie:
A disorder of motor planning for
Aktionen.
Central sulcus:
An indentation or “valley” on the
brain’s surface, between primary
motor and sensory areas.
MRIcroGL:
Free software for visualizing and
editing neuroimaging data.
Testing Schedule
Participant 1 completed three testing sessions with six weeks intervening between each visit.
The first testing session lasted approximately 3 H. During the first testing session, she completed
the SOAP, Word-to-Picture Matching with Mixed Foils, Word Discrimination, Nonword
Discrimination, Digit Span, Word Span, Nonword Span, WAB (written picture description,
yes/no questions, auditory word recognition, sequential commands, object naming, reading
comprehension of sentences, writing to dictation, and apraxia subtests), and three conditions
of the Adaptive Word-to-Picture Matching task. Structural MRI (1.5 T) was also acquired during
the first visit; 3 T and functional imaging were contraindicated due to an implanted medical
device. The second testing session lasted approximately one and half hours. During the second
testing session, she completed the object- and subject-relative conditions of Sentence-to-Picture
Matching, Sentence Verification, Digit Span, 4-Digit Recall, Word Span, Nonword Span, Word-
to-Picture Matching with Semantic Foils, Sentence Plausibility, and three conditions of the
Adaptive Word-to-Picture Matching task. The three sentence processing tasks were added to
the testing schedule to confirm and investigate the object-relative sentence processing deficit
observed during the first testing session. The third testing session lasted approximately one hour.
During the third testing session, she completed Word-to-Picture Matching with Phonological
Foils, Word-to-Picture Matching with Mixed Foils in Noise, Audiovisual Integration, and four
conditions of the Adaptive Word-to-Picture Matching task.
Participant 2 completed two testing sessions with eight weeks intervening. The first testing
session lasted approximately 2 H. During the first testing session, she completed the SOAP,
Word-to-Picture Matching with Mixed Foils, Word Discrimination, Nonword Discrimination,
Digit Span, 4-Digit Recall, WAB (auditory verbal comprehension, word recognition, sequential
commands, and apraxia subsections), and one condition of the Adaptive Word-to-Picture
Matching task. Structural and functional MRI (3 T) was also acquired during the first visit.
Functional MRI tasks included covert object naming, covert action naming, and covert famous
face naming. During the second testing session, she completed Word Span, Nonword Span,
Word-to-Picture Matching with Phonological Foils, Word-to-Picture Matching with Mixed
Foils in Noise, Audiovisual Integration, and four conditions of the Adaptive Word-to-Picture
Matching task.
Neuroimaging
Anatomical MRI scans were obtained for Case 2 using a 3 T whole-body magnetic resonance
scanner (Philips Medical Systems) fitted with a 16-channel SENSE head coil. Images were
collected using a magnetization-prepared 180 radio-frequency pulse and rapid gradient-echo
sequence with 1 mm sagittal slices and an in-plane resolution of 0.938 X 0.938 mm. The same
specifications were used for Case 1, with the exception of a 1.5 T field strength and a transmit/
receive head coil. Images and renderings were generated with MRIcroGL (https://www.nitrc.org
/projects/mricrogl/). Lesion segmentations were drawn on axial slices by a trained neuropsychol-
ogist (author G.W.) using MRIcroGL and checked for accuracy by a neurologist (author N.T.).
Cortical atrophy and ventricle dilation were not identified as part of the lesion segmentation. Der
central sulcus on each patient’s scan was traced by hand to provide an anatomical reference
(with some approximation required inside the lesions).
Axial:
The plane passing through the head
that gives a top-down or bottom-up
view; both hemispheres are visible.
ERGEBNISSE
Results for all behavioral tests are presented in Table 1. Lesion maps are presented in Figure 1
and Figure 2.
Neurobiology of Language
341
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
Test
Maximum score
Case 1
Case 2
Controls
Tisch 1.
Behavioral test results for bilateral patients and neurotypical controls
Phonemes
Word discrimination (same)
Word discrimination (anders)
0
Word discrimination A
Nonword discrimination (same)
Nonword discrimination (anders)
0
Nonword discrimination A
Word-to-picture matching (phon foil)
Adaptive word-to-picture matching (buy-pie 75% threshold)
Adaptive word-to-picture matching (die-tie 75% threshold)
Adaptive word-to-picture matching (buy-die 75% threshold)
Adaptive word-to-picture matching ( pie-tie 75% threshold)
Audiovisual integration (A only)
Audiovisual integration (A/ V congruent)
Audiovisual integration (A/ V incongruent fuse/audio/visual)
Word-to-picture matching (sem foil)
Word-to-picture matching (mix foil)
Word-to-picture matching (mix foil+noise)
WAB word recognition
WAB yes/no questions
WAB sequential commands
Sentence-to-picture matching (active)
Sentence-to-picture matching (passive)
Sentence-to-picture matching (subject relative)
Sentence-to-picture matching (object relative)
Sentence-picture verification, matching (subject relative)
Sentence-picture verification, non-matching (subject relative)
0
Sentence-picture verification, A
(subject relative)
Sentence-picture verification, matching (object relative)
20
20
1
20
20
1
20
–
–
–
–
30
20
10
20
20
20
60
20
19
10
10
10
10
5
5
1
5
Words
Sentences
18
19
0.96
16
9
0.72
19
20
20
1
20
20
1
20
[20, 20]
21, >30 dB
>30 dB
[3.5, 8.5] dB
>30, 28, 19 dB
5.5, 1 dB
[−5.5, 2] dB
>30, 1.5, 19 dB
−14 dB
[−16, −14] dB
>30, >30 dB
−2.5 dB
[−15, −8.5] dB
29
19
30
20
3/5/2
10/0/0
[20, 20]
[20, 20]
[16, 19]
20
20
16
59
20
15
9
9
9, 8
4, 3
5
4
0.95
4
20
20
17
60
20
19
10
10
10
9
–
–
–
–
Neurobiology of Language
342
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
Tisch 1.
(Fortsetzung )
Test
Sentence-picture verification, non-matching (object relative)
Maximum score
5
Case 1
0
Case 2
–
Controls
0
Sentence-picture verification, A
(object relative)
Sentence plausibility, plausible (active)
Sentence plausibility, implausible (active)
0
Sentence plausibility, A
(active)
Sentence plausibility, plausible, (passive)
Sentence plausibility, implausible (passive)
0
Sentence plausibility, A
(passive)
Sentence plausibility, plausible, (subject relative)
Sentence plausibility, implausible (subject relative)
0
Sentence plausibility, A
(subject relative)
Sentence plausibility, plausible, (object relative)
Sentence plausibility, implausible (object relative)
0
Sentence plausibility, A
(object relative)
Digit span
Word span
Nonword span
4-digit recall (15 S)
1
10
10
1
10
10
1
10
10
1
10
10
1
Short-term memory
4
0
6
10
0.90
7
9
0.88
7
9
0.88
7
5
0.67
4
3
2
1
–
–
–
–
–
–
–
–
–
–
–
–
–
5
4
3
2
6A
Notiz. The range of control performance is reported in square brackets, [Mindest., max.]. Multiple, non-bracketed, comma-separated entries within a cell reflect
multiple attempts on different testing occasions. For the Adaptive Word-to-Picture Matching task, the signal-to-noise ratio (dB) that yielded 75% Leistung
accuracy is reported. Because dB is on a log ratio scale, a positive value indicates that the signal was louder than the noise, a negative value indicates that the
noise was louder than the signal, and zero indicates equal amplitudes for signal and noise. Lower dB values indicate better performance (greater noise toler-
0
ance); the maximum dB value was capped at 30 dB, indicating total failure to discriminate the target word, even with relatively negligible background noise. A
represents “an estimate of proportion correct corrected for response bias” (see Rogalsky, Pitz, et al., 2008, P. 168). A: Audio-; A/ V: audio-visual; phon: phono-
logical distractor; sem: semantic distractor; mix: mixed distractor. A only: audio signal only; A/ V congruent: audio-visual signals from the same production.
a Selnes et al., 1991 – 5th percentile.
Word and Nonword Discrimination
0
0
=
Both participants performed well on the word discrimination task: Case 1 scored 92.5% (A
represents “an estimate of proportion correct corrected for
0.96) and Case 2 scored 100%. (A
response bias”; see Rogalsky, Pitz, et al., 2008, P. 168). The two cases diverged in their ability to
perform the nonword discrimination task. Case 1 had significant difficulty, scoring only 62.5%
0
(A
= 0.76), whereas Case 2 scored 100%. This suggests that Case 1 has a verbal short-term mem-
ory deficit that limits her ability to maintain and compare two phonological forms without
lexical-semantic support. In a follow-up analysis, we assessed whether we could induce a dis-
sociation between word and nonword discrimination by testing two of the authors (neurotypical
listeners) on a version of these tasks where the stimuli were presented in noise to bring perfor-
mance down from ceiling levels. Performance was indeed worse on the noise version of the
Neurobiology of Language
343
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
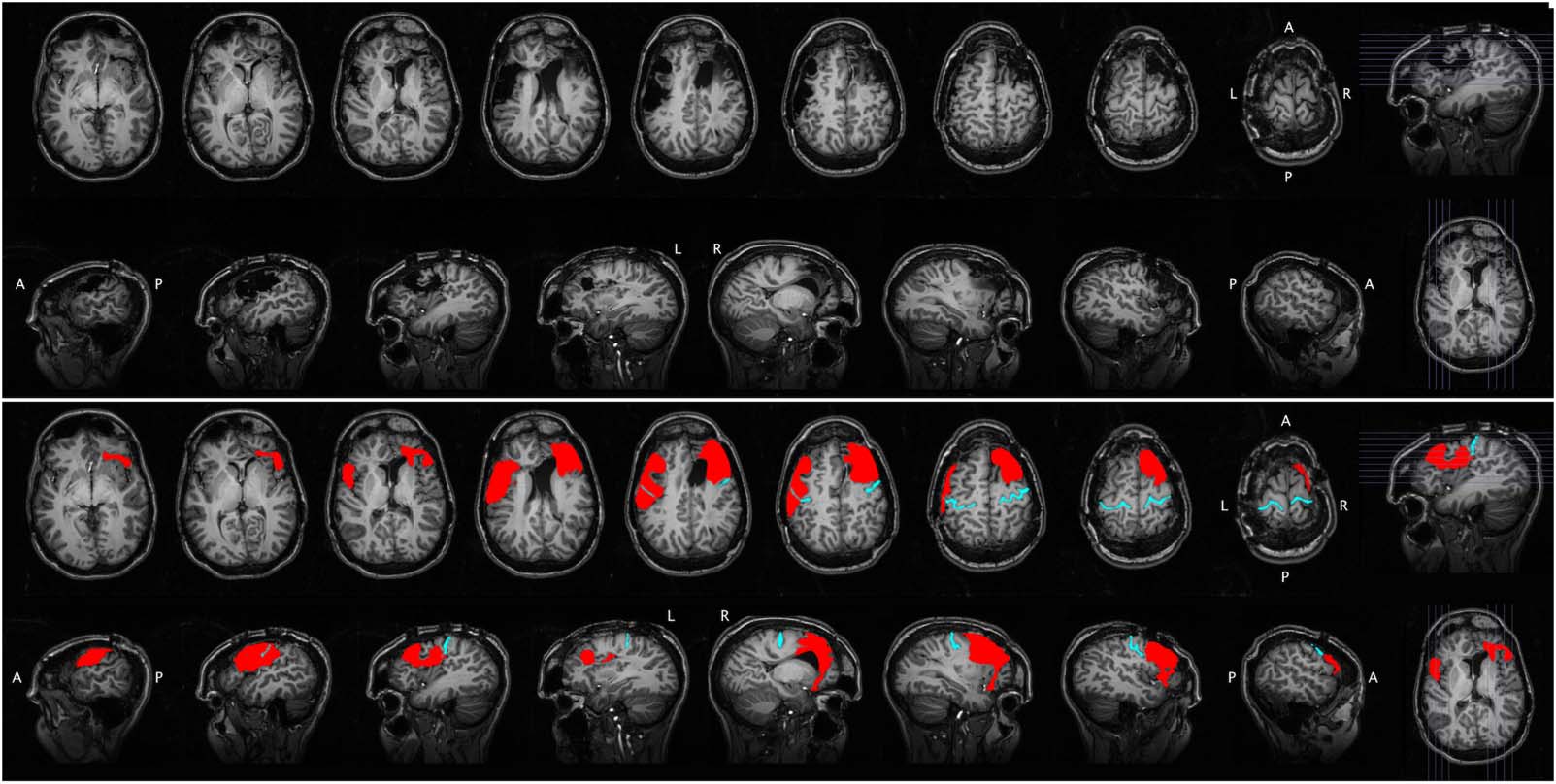
Figur 1.
Structural MRI for Case 1. The raw scan is shown in the top portion of the figure, with axial slices (first row) and sagittal slices
(zweite Reihe). The same scan is shown in the bottom portion of the figure, with the lesion segmentation highlighted in red and the central
sulcus highlighted in cyan for reference. A = anterior, P = posterior, L = left, R = right.
tasks but was similar for words (listener 1: 80% correct, listener 2: 85% correct) versus non-
Wörter (listener 1: 80% correct, listener 2: 82.5% correct). Daher, the dissociation observed in
Case 1 is likely not attributable to general differences in the difficulty of the two versions.
Performance on the adaptive word-comprehension in noise task was relatively poor for
both patients. Case 1 was essentially at floor levels, converging on an identifiable threshold
on only 2 out of 40 staircase runs (both in the easiest, buy/die, condition). Case 2 exhibited
mixed performance on this task, performing poorly on two of the phonemic contrasts ( pie/tie
and buy/pie) and within normal limits on the other two contrasts (die/tie and buy/die).
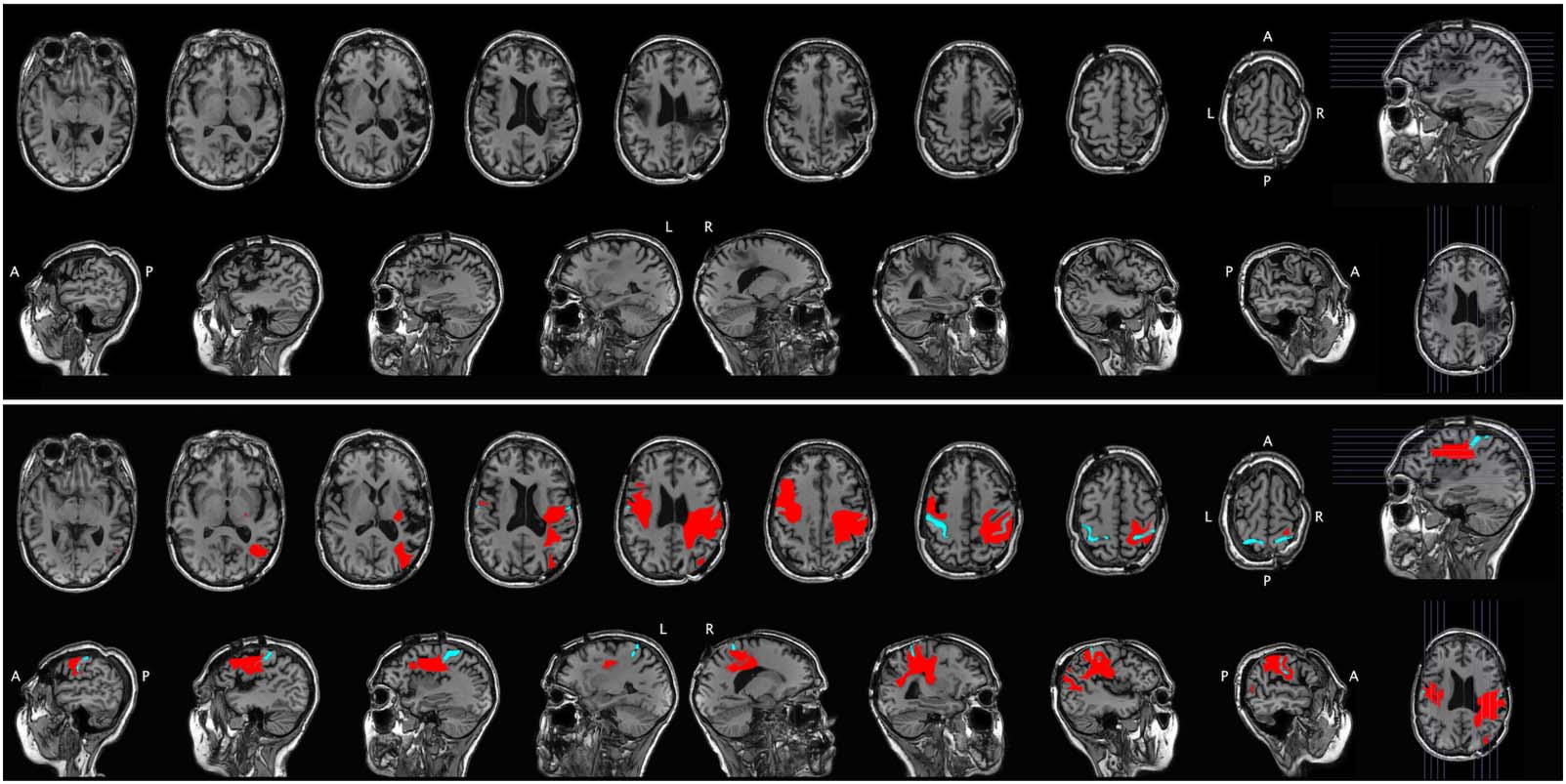
Figur 2.
Structural MRI for Case 2. The raw scan is shown in the top portion of the figure, with axial slices (first row) and sagittal slices
(zweite Reihe). The same scan is shown in the bottom portion of the figure, with the lesion segmentation highlighted in red and the central
sulcus highlighted in cyan for reference. A = anterior, P = posterior, L = left, R = right.
Neurobiology of Language
344
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
The McGurk effect:
When incongruent auditory and
visual speech signals are perceived
as a completely different speech
Klang.
Audiovisual Tasks
Both participants performed well on the auditory only and congruent audiovisual (AV) Aufgabe
Bedingungen, scoring 95% correct or better. For the incongruent AV task condition, Case 2 showed
a robust McGurk effect, reporting fused percepts on every trial. Case 1’s phonemic perception
was also influenced by incongruent AV stimuli, reporting a percept corresponding to the audio
signal only 50% of the time (vgl. 95% in the auditory only condition); the remaining 50% of her
responses were fused (3/5) and visual (2/5).
Word Comprehension
Both patients performed nearly flawlessly on all tests of non-noisy word comprehension (word-
picture matching with phonological, semantic, and mixed foils, WAB Word Recognition). Case 1
scored 98.3% correct (118/120) and Case 2 scored 100% on these tests. Performance was within
the normal range on the mixed foil word comprehension in noise task for both patients, although
lower, Natürlich, than performance on the clear speech version of the task. Daher, word compre-
hension appears well-preserved in both patients.
Sentence Comprehension
Comprehension of simple sentences was largely preserved for both of our participants, WHO
scored perfectly on the WAB yes/no questions and 90% or better on the sentence-picture
matching task for active, passive, and subject-relative sentences. When the sentence structure
increased working memory load ( WAB sequential commands and object-relative sentences),
Case 1’s performance dropped to 66% correct whereas Case 2 performed well (97% correct).
Case 1 exhibited a strong primacy effect on the sequential commands task; all of the initial
actions were correct. Given the marked deficit that Case 1 exhibited for object-relative pro-
cessing on the original sentence comprehension task, follow-up tests were administered that
reduced the working memory demands associated with choosing among response alterna-
tives. Trotzdem, a similar working memory load pattern was observed for Case 1 on the sentence
verification and plausibility tasks, with notable deficits for object-relative structure only, indi-
cating that the deficit observed in sentence-picture matching did not depend on selection
among multiple response alternatives. Case 1 exhibited a strong bias toward “yes” answers
on the object-relative sentence verification task, suggesting a reliance on identification of lexical
items instead of their syntactic roles.
Span Tasks
Both participants showed some evidence of reduced short-term memory. Case 1 durchgeführt
consistently worse than Case 2 across the tasks, which is consistent with her poorer perfor-
mance on language tasks that involve higher short-term memory load.
Additional Observations
Case 1 exhibited mild aphasic deficits on written production tasks. She scored 13/15 on WAB
written naming (rubber band ! “rubber bamb”, screwdriver ! “flatliner”). Written picture
description initially consisted only of single words (picnic, sailboat, beach, fishing, house,
lunch). After being prompted to use complete sentences, she produced a few short phrases
(“Man has his shoes off”; “Dog chasing a kid”; “It’s windy”; “It’s sunny also windy”; “Car has
no color”). On writing to dictation of Pack my box with five dozen jugs of liquid veneer,
she wrote, “Pack my box with 5 dozens of jugs of liquid vanery.” Although there was not
Neurobiology of Language
345
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
enough time to formally evaluate written production for Case 2, her use of her smartphone
for text-to-speech communication was rapid, fluent, and errorless. During a break from testing,
she communicated about her memories of being in the hospital for surgery, her family medical
Geschichte, and suggestions for sightseeing while visiting her hometown. Both participants used
smartphones for text or text-to-speech communication in their daily lives.
Case 1 was completely unable to phonate voluntarily, although her mother reported that she
could (very rarely) produce spontaneous vocalizations in response to pain or fear, such as when
waking from a nightmare. Case 2 was able to voluntarily produce guttural sounds using the
larynx with rising or falling pitch, which she used to communicate, Zum Beispiel, indicating
confusion/understanding or affirmative/negative, often paired with appropriate head move-
ments and facial expressions involving the muscles around the eyes and the brow. The sponta-
neous Duchenne smile was observed in both cases during testing. Both patients reported having
the experience of inner speech.
Both patients were able to pantomime upper limb gestures (z.B., making a fist), instrumental
Gesten (z.B., using a hammer), and complex gestures (z.B., pretending to knock at the door and
open it). Both patients were also able to close their eyes on command, and were able to attempt
gestures with the lower face. Case 1 was able to pantomime sniffing a flower and blowing out a
match, but was unable to stick out her tongue or whistle. Case 2 was unable to perform any of
these lower face gestures, and was unable to attempt whistling. Case 1 was able to take food
orally, including sucking on a breath mint and drinking from a cup or straw. Case 2 had dyspha-
gia and so was unable to take food orally, relying on a feeding tube; she used a hand towel to
manage involuntary production of saliva that could not be swallowed.
Structural Neuroimaging
Case 1 has a large left fronto-parietal lesion centered on lower sensorimotor and premotor (BA6)
Kortex. The lesion extends anteriorly to include the pars opercularis and most if not all of the pars
triangularis; inferior portions of the posterior middle frontal gyrus are also involved. Posteriorly,
the lesion extends into the inferior parietal lobule but spares the posterior supramarginal gyrus. In
the right hemisphere, the lesion is restricted to the frontal lobe, much of which is involved,
including lateral premotor cortex, Broca’s area homolog, and inferior motor cortex. Spared
regions include the frontal pole, ventromedial structures, posterior dorsomedial structures,
mid-superior portions of the precentral gyrus, and portions of the frontal operculum.
Case 2 turned out to have more asymmetric damage. In the left hemisphere, damage is centered
on the pars opercularis/premotor cortex with posterior extension into inferior primary sensorimotor
Kortex. The pars triangularis of Broca’s area is largely spared. In the right hemisphere the lesion is
predominantly in the parietal lobe involving virtually all of it from the postcentral gyrus (einschließlich
the portions of the lower precentral gyrus) to posterior parietal cortex (even including portions of
the angular gyrus, but with some islands of sparing around the supramarginal gyrus) and from the
parietal operculum to dorsal and medial parietal areas, sparing only the posterior cingulate and
posterior precuneus region. The lesion extends in depth to the lateral ventricle, thus interrupting
most of the dorsal stream white matter pathways.
DISKUSSION
We evaluated the receptive speech and language processing abilities in two cases of opercular
syndrome. Both have damage to left hemisphere motor speech areas including lower primary
sensorimotor cortex, lower premotor cortex (BA6), and the pars opercularis of Broca’s area. Case
1 also has involvement of right hemisphere motor speech areas (M1, premotor cortex, Broca’s
Neurobiology of Language
346
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
Bereich), whereas Case 2 has an extensive parietal lobe lesion interrupting dorsal stream
sensorimotor processing. Behaviorally, both were incapable of voluntary control of their vocal
tract and lower facial muscles (although Case 2 was able to produce guttural sounds for limited
communication), consistent with opercular syndrome. Despite these severe motor speech
deficits, both participants performed remarkably well on the clear speech word-to-picture
matching comprehension task and word-pair discrimination tasks, even when subtle phonemic
cues had to be resolved for correct performance. Case 1 had some difficulty with nonword
discrimination and with the identification of synthesized speech in noise, which we argue stems
from a phonological short-term/working memory deficit (siehe unten). Both participants showed
sensitivity to mismatched AV speech signals and were able to comprehend simple sentences.
Case 2 was also able to comprehend syntactically complex sentences showing largely preserved
receptive language abilities despite her motor speech impairment. Case 1 had difficulty with
complex syntax and limited immediate recall span, consistent with a phonological short-term
Erinnerung (pSTM) deficit. A previous report in the literature identified a case of opercular syn-
drome with similar difficulty comprehending center embedded sentences, which they also
linked to a pSTM deficit (Silveri et al., 2017).
Implications for Motor-Based Theories of Speech Perception
The present study adds to the large body of evidence arguing against a strong version of the
motor theory of speech perception, that an intact motor speech system is required to perceive
Rede. Both of the present cases performed well on the word-to-picture matching tests and
the word discrimination test, even though the ability to distinguish between highly similar
phones was required for success. A strong version of the motor theory predicts impaired
performance on all tasks requiring this degree of phonemic discrimination.
Weaker, modulatory theories of the role of the motor speech system in perception can
claim some tentative, qualified support, Jedoch. Speziell, Case 1, who had more exten-
sive bilateral motor involvement, was impaired on nonword discrimination and was at floor on
our adaptive speech-in-noise task. If one assumes that Case 2, whose right precentral motor
system was largely spared, performed well on these same tasks because of the sparing of her
right motor system, then one could argue that the motor system contributes substantially to the
performance on these tasks. This is a tentative conclusion because Case 2 became anarthric
only after the right hemisphere event, indicating that the right hemisphere lesion, in combina-
tion with the left hemisphere lesion, indeed permanently interrupted motor speech function.
dennoch, the discussion below will assume, for the sake of argument in favor of motor
theories, that the difference in performance between Case 1 and Case 2 is due to the difference
in motor cortex involvement.
The support for a weak version of the motor theory is qualified because our data suggest that
it is task-dependent, holding only for tasks that involve nonwords or degraded stimuli presented
in noise. The contrast in performance between the word and nonword discrimination tasks is
particularly telling. For both tasks, listeners have to make fine phonemic discriminations in order
to detect the difference between the items in each pair. Case 1 could perform this task success-
fully when the items were real words, but largely failed when the items were nonwords. Success
on the word stimuli indicate that fine phonemic discrimination is well preserved and enables
access to lexical-semantic representations. Given this, failure on the nonword discrimination
task cannot be a result of phonemic perception impairment generally—otherwise word discrim-
ination would be impossible—but rather, it depends on the ability to make a discrimination on
the basis of phonemic information alone, since lexical-semantic access is not possible for
Neurobiology of Language
347
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
nonwords. This suggests a pSTM explanation of Case 1’s difficulty with the nonword task, als
we detail later.
Case 1 also had much more trouble on the adaptive word-comprehension-in-noise task than
Case 2 hatte, performing essentially at floor. Taken at face value, this could be interpreted as
support for the claim that the motor system is important for speech perception under noisy
listening conditions. Jedoch, Case 1’s performance was much better (80% correct) and well
above chance (25%) on the speech-in-noise word-picture matching task with natural speech
Reize. In der Tat, this level of performance is just a single error below the performance range of
controls tested as part of another study and in an acoustically controlled setting (91% correct,
SD = 4.9%, range 85–100%; Rogalsky et al., 2011, unpublished data). It is also within the range
of scores, albeit on the low end, from the four control participants tested in the same room under
the same conditions and similar to Case 2’s performance (85% correct). If the dramatic failure on
the adaptive task was a result of the lack of motor system input to perceiving speech in noise, Wir
would expect a more substantial deficit on our other speech-in-noise task. Das deutet darauf hin
the adaptive task is tapping into something different. We will argue that it is again attributable
to a short-term phonological memory problem. We turn to this issue next.
Short-Term Memory, Speech Perception, and the Motor System
It is well-established that the motor speech system plays a critical role in pSTM via the articula-
tory rehearsal component of the phonological loop (Baddeley, 1992; Buchsbaum et al., 2011;
Buchsbaum & D'Esposito, 2008; Hickok et al., 2003). Severe damage to the motor speech sys-
tem, as in our present cases, should, daher, cause pSTM deficits, which we observed, partic-
ularly for Case 1. The question we address here is whether a pSTM deficit can confound
performance on speech perception tasks. Das ist, do some speech perception tasks rely in part
on pSTM?
Case 1 performed poorly on our nonword discrimination task. This clearly involves some
kind of pSTM, because the first item must be maintained until after the second item is pre-
gesendet, and then the two representations are compared. Because the items are nonwords, Dort
is no opportunity to recode the stimuli semantically. This places the burden on some form of
either auditory or phonological STM. Daher, there is no question that discrimination tasks require
some degree of STM that is not required during word comprehension, which involves process-
ing a single word-form and activating its associated lexical representation incrementally even
before the whole word is perceived (W. D. Marslen-Wilson, 1987; W. Marslen-Wilson & Tyler,
1980). Aber, one might argue, listening to only two items would seem to impose a minimal
pSTM burden, and even Case 1 had a nonword span of two items. Jedoch, the items in the
nonword span task were phonologically dissimilar, which benefits pSTM (Baddeley, 1992), In
contrast to the minimal pair comparisons in the discrimination task. Daher, the nonword span
task may overestimate the STM resources available for performance on the discrimination task.
Case 1 also performed very poorly on our adaptive word-to-picture matching task, welche
involved degraded (synthesized) speech stimuli presented in noise. On this task, Teilnehmer
were not instructed to perform discrimination directly (d.h., they were instructed to match a
word to a picture, rather than compare two word-forms), and so it should not tax pSTM in
the same way as the discrimination task. Als solche, Case 1’s poor performance might be viewed
as support for the motor system’s involvement in augmenting speech perception under noisy
listening conditions (Moineau et al., 2005; Wilson, 2009), perhaps using some form of predic-
tive coding (Hickok et al., 2011; Londei et al., 2009; Schwartz et al., 2012). On closer inspec-
tion, Jedoch, we noticed that some of the synthesized tokens were difficult to identify when
Neurobiology of Language
348
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
presented in isolation (informal testing confirmed this; stimuli publicly available at https://osf
.io/pw35n/) and were only identifiable when the target words were cued by pictures. Wir
further noted two additional complicating aspects of the task. Erste, even with the pictures
cueing the word category alternatives, the perceived ease of categorization of the synthesized
speech stimuli was enhanced with more exposure to the two competitors as the staircase run
proceeded during the relatively favorable signal-to-noise ratio trials; this effect was most notice-
able on the most ambiguous stimuli ( pie/buy). Mit anderen Worten, some degree of perceptual
differencing between time-separated items in the stimulus set was important for optimal task per-
Form. This differencing could tax pSTM. Zweite, the position of the two alternative pictures
(and therefore response buttons) was randomized: On some trials alternative A appeared on the
left with B on the right, and on other trials, it was reversed. This randomization places additional
short-term and working memory demands on the participant, who must not only categorize the
stimulus, but also match the decision to the correct response button, which varies from trial to
trial. Zusamenfassend, the adaptive task is nontrivial. In diesem Kontext, it is worth pointing out that in the
prior study of 24 healthy undergraduates (Stokes et al., 2019), for which the task was designed,
4 (16.7%) were excluded due to failure to converge on two or more staircase runs, indicating
that the task is indeed quite difficult for a sizeable fraction of even healthy listeners.
These observations suggest several explanations of the present speech-in-noise effect that
are alternatives to a motor-prediction or analysis-by-synthesis mechanism (Bever & Kacke,
2010). The first is that Case 1 may have fallen into the group of people who have difficulty with
this task even prior to her surgery. Das ist, based on available data (Stokes et al., 2019), there is
A 16.7% chance that she would have had a premorbid difficulty with the task that has nothing
to do with her current lesions. A second possible explanation is that her frontal lobe lesions
impaired not her ability to hear speech in noise, but her cognitive control ability (Brownsett
et al., 2014; Gläscher et al., 2012; Novick et al., 2005), which could interfere with the per-
formance of changing perception-response mappings. The third possible explanation is that
the degraded nature of the synthesized speech stimuli themselves, even without noise, makes
the task closer to a nonword discrimination task than a speech-in-noise task. The logic here is
that because the auditory stimuli on their own map poorly onto word categories, the listener
must first discriminate the two (nonword) alternatives and then map them onto word categories
rather than directly comprehend each stimulus in an ecologically natural way. This suggestion
is consistent with the well-known fact that the range of stimulus exemplars and comparison
categories in the stimulus set influences category judgements for speech (Holt & Lotto, 2010)
and implies some form of short-term memory of the stimulus set for task optimization. Daher,
Case 1’s apparent pSTM deficit, as revealed by her impaired performance on immediate serial
recall, sequential commands, object-relative sentence comprehension, and nonword discrim-
ination tasks, is consistent with this analysis of her failure on the adaptive task. While Case 2
did not exhibit a particularly large advantage on immediate serial recall tasks over Case 1, Die
advantage was consistent across stimulus conditions; the different STM ability levels between
these patients possibly straddled a critical threshold required for supporting speech processing
in other tasks.
We conclude that motor speech deficits impact pSTM as both of our cases had reduced
spans. If severely impaired, pSTM can impact performance on some speech perception tasks.
Sentence Comprehension
Both of the present cases showed well-preserved comprehension for simple sentences, showing
that receptive speech ability up to the level of basic sentences is not impeded by severe motor
Neurobiology of Language
349
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
speech dysfunction. Case 1, Jedoch, had notable difficulty with syntactically complex
Sätze, such as semantically reversible object-relative structures. She also had difficulty
with multistep sequential commands. Comprehension of such complex sentences is known to
activate frontal, motor-related language areas (for reviews see Friederici, 2003; Matchin &
Hickok, 2020; Rogalsky, Matchin, & Hickok, 2008; Segaert et al., 2012). Some have argued
that regions such as Broca’s area play a key role in syntax, including during comprehension
(Fedorenko & Kanwisher, 2011; Friederici, 2003; Segaert et al., 2012), while others argue that
frontal areas are only involved when working memory demands are high (Matchin & Hickok,
2020; Rogalsky, Matchin, & Hickok, 2008). The present findings do not strongly disentangle
these claims: It could be that Case 1’s pSTM deficit is the cause of her sentence comprehension
problems, or she might have both a pSTM deficit and a separate syntactic deficit due to her
frontal lesion. Case 2, with her relatively preserved complex sentence comprehension, might
have adjudicated, but her lesion appears to spare at least the pars triangularis of Broca’s area,
leaving open the possibility that this region is still available to support syntactic ability.
Audiovisual Speech
Dominant neural models of audiovisual speech integration hold that the posterior superior
temporal sulcus is a critical site (Beauchamp et al., 2004; Beauchamp et al., 2010; Venezia
et al., 2017). Other models implicate motor speech networks (Sams et al., 2005; Skipper
et al., 2005, 2007). The present cases enabled a test of the role of motor speech systems in
AV speech integration by assessing whether a McGurk effect would emerge when audio and
visual cues were in conflict. Both cases showed sensitivity to AV mismatched stimuli, in that they
deviated from their performance on audio-only perception. Case 2 showed a robust McGurk
Wirkung, reporting a fused percept on every trial. This demonstrates that the intact motor control
of speech is not necessary for McGurk fusion. A similar conclusion comes from a large-scale
study of unilateral lesions, which found that posterior, not anterior damage predicted AV
integration failures (Hickok et al., 2018). Case 1 reported fused syllables on 30% of the trials.
The interpretation of this pattern is unclear because not all healthy individuals exhibit McGurk
fusion (Basu Mallick et al., 2015).
Conclusions
The main finding from this study is that bilateral damage to the motor speech system has little
effect on the ability to recognize speech even when motor speech output is extremely impaired
Such damage can impair pSTM span and, in Case 1, produce an agrammatic-type comprehen-
sion pattern. Audiovisual speech integration is not necessarily impaired, consistent with a recent
large-scale lesion study, nor is speech-in-noise perception necessarily profoundly affected.
Typical of neuropsychological case reports, the neurological interventions studied here were
undertaken for medical, not scientific, Zwecke, Und, as such, the motor speech system was
not affected exclusively or completely. Außerdem, the continued presence of epileptic disor-
der, at least in one participant, and the considerable length of time between the surgical inter-
ventions and our assessments, allow for the possibility of reorganization of functional brain
Netzwerke. It is noteworthy, Jedoch, that despite all of the opportunity for reorganization of mo-
tor speech systems, this was not found for expressive function. The possibility remains open that
the motor system does play a nontrivial role in receptive speech processing under typical con-
ditions, but that when it is destroyed, sensory systems alone can ultimately achieve the same or
very nearly the same level of performance. But this is fundamentally what we are arguing: Der
motor system is not critical for speech perception. These findings confirm that a strong version of
Neurobiology of Language
350
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
the motor theory of speech perception is untenable and provide only weak support for weaker
versions of the motor theory, in which the motor system plays a small role at best in perceiving
speech under near threshold conditions for only some types of speech stimuli.
FUNDING INFORMATION
This research was supported by funding from the National Institute for Deafness and Other
Communication Disorders P50 DC014664-5340 (Hickok), the National Institute of Neurological
Disorders and Stroke R01 DC014589 (Tandon), and UT Health start-up for Texas Institute of
Restorative Neurotechnologies (Tandon).
BEITRÄGE DES AUTORS
Grant M. Walker: Datenkuration: Equal; Formale Analyse: Equal; Untersuchung: Lead;
Methodik: Equal; Projektverwaltung: Supporting; Visualisierung: Lead; Writing – original
Entwurf: Lead; Writing – review & Bearbeitung: Equal. Patrick Sarahan Rollo: Datenkuration: Equal;
Untersuchung: Supporting; Writing – review & Bearbeitung: Supporting. Nitin Tandon:
Konzeptualisierung: Equal; Formale Analyse: Equal; Akquise von Fördermitteln: Lead; Methodik:
Equal; Projektverwaltung: Supporting; Ressourcen: Lead; Aufsicht: Lead; Visualisierung:
Supporting; Writing – review & Bearbeitung: Equal. Gregory Hickok: Konzeptualisierung: Lead;
Akquise von Fördermitteln: Supporting; Methodik: Equal; Writing – original draft: Supporting;
Writing – review & Bearbeitung: Equal.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
VERWEISE
Baddeley, A.
(1992). Working memory. Wissenschaft, 255(5044),
556–559. DOI: https://doi.org/10.1126/science.1736359, PMID:
1736359
Bakar, M., Kirshner, H. S., & Niaz, F. (1998). The opercular-
subopercular syndrome: Four cases with review of the literature.
Behavioural Neurology, 11(2), 97–103. DOI: https://doi.org/10
.1155/1998/423645, PMID: 11568407
Basu Mallick, D., Magnotti, J. F., & Beauchamp, M. S. (2015).
Variability and stability in the McGurk effect: Contributions of
Teilnehmer, Reize, Zeit, and response type. Psychonomic
Bulletin & Rezension, 22(5), 1299–1307. DOI: https://doi.org/10.3758
/s13423-015-0817-4, PMID: 25802068, PMCID: PMC4580505
Beauchamp, M. S., Argall, B. D., Bodurka, J., Duyn, J. H., & Martin,
A. (2004). Unraveling multisensory integration: Patchy organiza-
tion within human STS multisensory cortex. Naturneurowissenschaften,
7(11), 1190–1192. DOI: https://doi.org/10.1038/nn1333, PMID:
15475952
Beauchamp, M. S., Nath, A. R., & Pasalar, S. (2010). fMRI-guided
transcranial magnetic stimulation reveals that the superior
temporal sulcus is a cortical locus of the McGurk effect. Der
Zeitschrift für Neurowissenschaften: The Official Journal of the Society for
Neurowissenschaften, 30(7), 2414–2417. DOI: https://doi.org/10.1523
/ JNEUROSCI.4865-09.2010, PMID: 20164324, PMCID:
PMC2844713
Bever, T. G., & Kacke, D. (2010). Analysis by synthesis: A (re-)emerging
program of research for language and vision. Biolinguistics, 4(2-3),
174–200.
Bishop, D. V. M., Braun, B. B., & Robson, J. (1990). The relation-
ship between phoneme discrimination, speech production, Und
language comprehension in cerebral-palsied individuals. Zeitschrift
of Speech, Language, and Hearing Research, 33(2), 210–219.
DOI: https://doi.org/10.1044/jshr.3302.210, PMID: 2359262
Boscariol, M., Garcia, V. L., Guimarães, C. A., Montenegro, M. A.,
Hage, S. R. V., Cendes, F., & Guerreiro, M. M. (2010). Auditiv
processing disorder in perisylvian syndrome. Brain and Development,
32(4), 299–304. DOI: https://doi.org/10.1016/j.braindev.2009.04
.002, PMID: 19410403
Brownsett, S. L. E., Warren, J. E., Geranmayeh, F., Woodhead, Z. V. J.,
Leech, R., & Wise, R. J. S. (2014). Cognitive control and its impact
on recovery from aphasic stroke. Gehirn, 137(1), 242–254. DOI:
https://doi.org/10.1093/brain/awt289, PMID: 24163248, PMCID:
PMC3891442
Buchsbaum, B. R., Baldo, J., Okada, K., Berman, K. F., Dronkers, N.,
D'Esposito, M., & Hickok, G. (2011). Conduction aphasia,
sensory-motor integration, and phonological short-term memory—
An aggregate analysis of lesion and fMRI data. Brain and Language,
119(3), 119–128. DOI: https://doi.org/10.1016/J.BANDL.2010.12
.001, PMID: 21256582, PMCID: PMC3090694
Buchsbaum, B. R., & D'Esposito, M. (2008). The search for the pho-
nological store: From loop to convolution. Zeitschrift für Kognition
Neurowissenschaften, 20(5), 762–778. DOI: https://doi.org/10.1162
/jocn.2008.20501, PMID: 18201133
Christen, H.-J., Hanefeld, F., Kruse, E., Imhäuser, S., Ernst, J. P., &
Finkenstaedt, M. (2007). Foix-Chavany-Marie (anterior opercu-
lum) syndrome in childhood: A reappraisal of Worster-Drought
syndrome. Developmental Medicine & Child Neurology, 42(2),
122–132. DOI: https://doi.org/10.1111/j.1469-8749.2000
.tb00057.x, PMID: 10698330
D’Ausilio, A., Bufalari, ICH., Salmas, P., & Ermüdung, L. (2012). Die Rolle
of the motor system in discriminating normal and degraded
speech sounds. Kortex, 48(7), 882–887. DOI: https://doi.org/10
.1016/J.CORTEX.2011.05.017, PMID: 21676385
D’Ausilio, A., Pulvermüller, F., Salmas, P., Bufalari, ICH., Begliomini,
C., & Ermüdung, L. (2009). The motor somatotopy of speech
Neurobiology of Language
351
Speech perception in opercular syndrome
perception. Aktuelle Biologie, 19(5), 381–385. DOI: https://doi.org
/10.1016/j.cub.2009.01.017, PMID: 19217297
Desai, S. D., Patel, D., Bharani, S., & Kharod, N. (2013). Opercular
syndrome: A case report and review. Journal of Pediatric
Neurosciences, 8(2), 123–125. DOI: https://doi.org/10.4103
/1817-1745.117842, PMID: 24082930, PMCID: PMC3783719
Eimas, P. D., Siqueland, E. R., Jusczyk, P., & Vigorito, J. (1971).
Speech perception in infants. Wissenschaft, 171(3968), 303–306.
DOI: https://doi.org/10.1126/science.171.3968.303, PMID:
5538846
Fedorenko, E., & Kanwisher, N. (2011). Some regions within
Broca’s area do respond more strongly to sentences than to
linguistically degraded stimuli: A comment on Rogalsky and
Hickok (2011). Zeitschrift für kognitive Neurowissenschaften, 23(10),
2632–2635. DOI: https://doi.org/10.1162/jocn_a_00043
Friederici, A. D. (2003). The role of left inferior frontal and superior
temporal cortex in sentence comprehension: Localizing syntactic
and semantic processes. Hirnrinde, 13(2), 170–177. DOI:
https://doi.org/10.1093/cercor/13.2.170, PMID: 12507948
Galantucci, B., Fowler, C. A., & Turvey, M. T. (2006). The motor
theory of speech perception reviewed. Psychonomic Bulletin &
Rezension, 13(3), 361–377. DOI: https://doi.org/10.3758
/BF03193857, PMID: 17048719, PMCID: PMC2746041
Walisisch, V., Gernsbacher, M. A., Heyes, C., Hickok, G., & Iacoboni, M.
(2011). Mirror neuron forum. Perspectives on Psychological Science,
6, 369–407. DOI: https://doi.org/10.1177/1745691611413392,
PMID: 25520744, PMCID: PMC4266473
Gläscher, J., Adolphs, R., Damasio, H., Bechara, A., Rudrauf, D.,
Calamia, M., Paul, L. K., & Tranel, D. (2012). Lesion mapping
of cognitive control and value-based decision making in the pre-
frontal cortex. Verfahren der Nationalen Akademie der Wissenschaften
of the United States of America, 109(36), 14681–14686. DOI:
https://doi.org/10.1073/pnas.1206608109, PMID: 22908286,
PMCID: PMC3437894
Groswasser, Z., Korn, C., Groswasser-Reider, ICH., & Solzi, P. (1988).
Mutism associated with buccofacial apraxia and bihemispheric
lesions. Brain and Language, 34(1), 157–168. DOI: https://doi
.org/10.1016/0093-934X(88)90129-0, PMID: 3382930
Hickok, G. (2014). The myth of mirror neurons: The real neuroscience
of communication and cognition. W. W. Norton & Unternehmen.
Hickok, G., Buchsbaum, B. R., Humphries, C., & Muftuler, T.
(2003). Auditory–motor interaction revealed by fMRI: Speech,
Musik, and working memory in area Spt. Zeitschrift für Kognition
Neurowissenschaften, 15(5), 673–682. DOI: https://doi.org/10.1162
/jocn.2003.15.5.673, PMID: 12965041
Hickok, G., Houde, J., & Rong, F. (2011). Sensorimotor integration
in speech processing: Computational basis and neural organiza-
tion. Neuron, 69(3), 407–422. DOI: https://doi.org/10.1016/j
.neuron.2011.01.019, PMID: 21315253, PMCID: PMC3057382
Hickok, G., Okada, K., Barr, W., Pa, J., Rogalsky, C., Donnelly, K.,
& Grant, A. (2008). Bilateral capacity for speech sound pro-
cessing in auditory comprehension: Evidence from Wada proce-
dures. Brain and Language, 107(3), 179–184. DOI: https://doi.org
/10.1016/ J.BANDL.2008.09.006, PMID: 18976806, PMCID:
PMC2644214
Hickok, G., & Kacke, D. (2000). Towards a functional neuroanatomy
of speech perception. Trends in den Kognitionswissenschaften, 4(4), 131–138.
DOI: https://doi.org/10.1016/S1364-6613(00)01463-7, PMID:
10740277
Hickok, G., & Kacke, D. (2004). Dorsal and ventral streams: A
framework for understanding aspects of the functional anatomy
of language. Cognition, 92(1–2), 67–99. DOI: https://doi.org/10
.1016/J.COGNITION.2003.10.011, PMID: 15037127
Hickok, G., & Kacke, D. (2007). The cortical organization of
speech processing. Nature Reviews Neurowissenschaften, 8(5), 393–402.
DOI: https://doi.org/10.1038/nrn2113, PMID: 17431404
Hickok, G., Rogalsky, C., Matchin, W., Basilakos, A., Cai, J., Pillay,
S. B.,Ferrill, M., Mickelsen, S., Anderson, S. W., Liebe, T., Binder, J.,
& Fridriksson, J. (2018). Neural networks supporting audiovisual
integration for speech: A large-scale lesion study. Kortex, 103,
360–371. DOI: https://doi.org/10.1016/J.CORTEX.2018.03.030,
PMID: 29705718, PMCID: PMC6223621
Holt, L. L., & Lotto, A. J. (2010). Speech perception as categoriza-
tion. Attention, Wahrnehmung, & Psychophysics, 72(5), 1218–1227.
DOI: https://doi.org/10.3758/APP.72.5.1218, PMID: 20601702,
PMCID: PMC2921848
Jansen, A. C., Leonard, G., Bastos, A. C., Esposito-Festen, J. E.,
Tampieri, D., Watkins, K., Andermann, F. & Andermann, E.
(2005). Cognitive functioning in bilateral perisylvian polymicro-
gyria (BPP): Clinical and radiological correlations. Epilepsy and
Behavior, 6(3), 393–404. DOI: https://doi.org/10.1016/j.yebeh
.2005.01.012, PMID: 15820349
Kertesz, A. (1982). The Western Aphasia Battery. Grune & Stratton.
Kuhl, P., & Müller, J. (1975). Speech perception by the chinchilla:
Voiced-voiceless distinction in alveolar plosive consonants.
Wissenschaft, 190(4209), 69–72. DOI: https://doi.org/10.1126
/science.1166301, PMID: 1166301
Liebenthal, E., & Möttönen, R. (2018). An interactive model of
auditory-motor speech perception. Brain and Language, 187
(Dezember), 33–40. DOI: https://doi.org/10.1016/j.bandl.2017
.12.004, PMID: 29268943, PMCID: PMC6005717
Londei, A., D’Ausilio, A., Basso, D., Sestieri, C., Gratta, C. Del,
Roma, G.-L., & Belardinelli, M. Ö. (2009). Sensory-motor brain
network connectivity for speech comprehension. Menschliches Gehirn
Mapping, 31(4), 567–580. DOI: https://doi.org/10.1002/ hbm
.20888, PMID: 19780042, PMCID: PMC6870571
Liebe, T., & Oster, E. (2002). On the categorization of aphasic typol-
ogies: The SOAP (a test of syntactic complexity). Zeitschrift für
Psycholinguistic Research, 31(5), 503–529. DOI: https://doi.org
/10.1023/A:1021208903394, PMID: 12528429
Marslen-Wilson, W. D. (1987). Functional parallelism in spoken
word-recognition. Cognition, 25(1–2), 71–102. DOI: https://doi
.org/10.1016/0010-0277(87)90005-9, PMID: 3581730
Marslen-Wilson, W., & Tyler, L. K. (1980). The temporal structure
of spoken language understanding. Cognition, 8(1), 1–71. DOI:
https://doi.org/10.1016/0010-0277(80)90015-3, PMID: 7363578
Massaro, D. W., & Chen, T. H. (2008). The motor theory of speech
perception revisited. Psychonomic Bulletin and Review, 15, 453–457.
DOI: https://doi.org/10.3758/PBR.15.2.453, PMID: 18488668
Matchin, W., & Hickok, G. (2020). The cortical organization of syntax.
Hirnrinde, 30(3), 1481–1498. DOI: https://doi.org/10.1093
/cercor/bhz180, PMID: 31670779, PMCID: PMC7132936
Milanlioglu, A., Aydın, M. N., Gökgül, A., Hamamcı, M., Erkuzu,
M. A., & Tombul, T. (2013). Ischemic bilateral opercular syn-
drome. Case Reports in Medicine, 2013, 513572. DOI: https://
doi.org/10.1155/2013/513572, PMID: 23476665, PMCID:
PMC3588394
Moineau, S., Dronkers, N. F., & Bates, E. (2005). Exploring the pro-
cessing continuum of single-word comprehension in aphasia.
Journal of Speech, Language, and Hearing Research, 48(4),
884–896. DOI: https://doi.org/10.1044/1092-4388(2005/061),
PMID: 16378480
Möttönen, R., & Watkins, K. E. (2012). Using TMS to study the role of
the articulatory motor system in speech perception. Aphasiology,
26(9), 1103–1118. DOI: https://doi.org/10.1080/02687038.2011
.619515, PMID: 22942513, PMCID: PMC3431548
Neurobiology of Language
352
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech perception in opercular syndrome
Novick, J. M., Trueswell, J. C., & Thompson-Schill, S. L. (2005).
Cognitive control and parsing: Reexamining the role of Broca’s
area in sentence comprehension. Kognitiv, Affektiv, & Behavioral
Neurowissenschaften, 5(3), 263–281. DOI: https://doi.org/10.3758/CABN
.5.3.263, PMID: 16396089
Pulvermüller, F., & Ermüdung, L. (2010). Active perception: Sensorimotor
circuits as a cortical basis for language. Nature Reviews Neurowissenschaften,
11(5), 351–360. DOI: https://doi.org/10.1038/nrn2811, PMID:
20383203
Rogalsky, C., LaCroix, A. N., Chen, K.-H., Anderson, S. W.,
Damasio, H., Liebe, T., & Hickok, G. (2018). The neurobiology of
agrammatic sentence comprehension: A lesion study. Zeitschrift für
Cognitive Neuroscience, 30(2), 234–255. DOI: https://doi.org/10
.1162/jocn_a_01200, PMID: 29064339, PMCID: PMC6434535
Rogalsky, C., Liebe, T., Driscoll, D., Anderson, S. W., & Hickok, G.
(2011). Are mirror neurons the basis of speech perception?
Evidence from five cases with damage to the purported human mirror
System. Neurocase, 17(2), 178–187. DOI: https://doi.org/10.1080
/13554794.2010.509318, PMID: 21207313, PMCID: PMC3681806
Rogalsky, C., Matchin, W., & Hickok, G. (2008). Brocas Gegend, Satz
comprehension, and working memory: Eine fMRT-Studie. Grenzen in
Human Neuroscience, 2(OCT), 14. DOI: https://doi.org/10.3389
/neuro.09.014.2008, PMID: 18958214, PMCID: PMC2572210
Rogalsky, C., Pitz, E., Hillis, A. E., & Hickok, G. (2008). Auditiv
word comprehension impairment in acute stroke: Relative
contribution of phonemic versus semantic factors. Brain and
Language, 107(2), 167–169. DOI: https://doi.org/10.1016/j
.bandl.2008.08.003, PMID: 18823655, PMCID: PMC2585413
Saletti, V., Bulgheroni, S., D’Incerti, L., Franceschetti, S., Molteni,
B., Airaghi, G., Pantaleoni, C., D’Arrigo, S., & Riva, D. (2007).
Verbal and gestural communication in children with bilateral
perisylvian polymicrogyria. Journal of Child Neurology, 22(9),
1090–1098. DOI: https://doi.org/10.1177/0883073807306247,
PMID: 17890406
Sams, M., Möttönen, R., & Sihvonen, T. (2005). Seeing and hearing
others and oneself talk. Kognitive Gehirnforschung, 23(2-3),
429–435. DOI: https://doi.org/10.1016/J.COGBRAINRES.2004
.11.006, PMID: 15820649
Sato, M., Tremblay, P., & Gracco, V. L. (2009). A mediating role of the
premotor cortex in phoneme segmentation. Brain and Language,
111(1), 1–7. DOI: https://doi.org/10.1016/J.BANDL.2009.03.002,
PMID: 19362734
Schomers, M. R., Kirilina, E., Weigand, A., Bajbouj, M., &
Pulvermüller, F. (2015). Causal influence of articulatory motor
cortex on comprehending single spoken words: TMS evidence.
Hirnrinde, 25(10), 3894–3902. DOI: https://doi.org/10
.1093/cercor/bhu274, PMID: 25452575, PMCID: PMC4585521
Schwartz, J.-L., Basirat, A., Ménard, L., & Sato, M. (2012). Der
Perception-for-Action-Control Theory (PACT): A perceptuo-
motor theory of speech perception. Journal of Neurolinguistics,
25(5), 336–354. DOI: https://doi.org/10.1016/ J.JNEUROLING
.2009.12.004
Segaert, K., Menenti, L., Weber, K., Petersson, K. M., & Hagoort, P.
(2012). Shared syntax in language production and language
comprehension—An fMRI study. Hirnrinde, 22(7),
1662–1670. DOI: https://doi.org/10.1093/cercor/ bhr249,
PMID: 21934094, PMCID: PMC3377967
Selnes, Ö. A., Jacobson, L., Machado, A. M., Becker, J. T., Wesch,
J., Müller, E. N., Visscher, B., & McArthur, J. C. (1991). Normative
data for a brief neuropsychological screening battery. Perceptual
and Motor Skills, 73(2), 539–550. DOI: https://doi.org/10.2466
/pms.1991.73.2.539, PMID: 1766784
Silveri, M. C., Incordino, F., Lo Monaco, R., Bizzarro, A., Masullo,
C., Piludu, F., & Colosimo, C. (2017). Neural substrates of the
‘low-level’ system for speech articulation: Evidence from pri-
mary opercular syndrome. Journal of Neuropsychology, 11(3),
450–457. DOI: https://doi.org/10.1111/jnp.12099, PMID:
26852905
Skipper, J. ICH., Devlin, J. T., & Lametti, D. R. (2017). The hearing ear
is always found close to the speaking tongue: Review of the role
of the motor system in speech perception. Brain and Language,
164, 77–105. DOI: https://doi.org/10.1016/j.bandl.2016.10.004,
PMID: 27821280
Skipper, J. ICH., Nusbaum, H. C., & Small, S. L. (2005). Listening to
talking faces: Motor cortical activation during speech perception.
NeuroImage, 25(1), 76–89. DOI: https://doi.org/10.1016/ J
.NEUROIMAGE.2004.11.006, PMID: 15734345
Skipper, J. ICH., van Wassenhove, V., Nusbaum, H. C., & Small, S. L.
(2007). Hearing lips and seeing voices: How cortical areas
supporting speech production mediate audiovisual speech per-
ception. Hirnrinde, 17(10), 2387–2399. DOI: https://doi
.org/10.1093/cercor/ bhl147, PMID: 17218482, PMCID:
PMC2896890
Snelbaker, A. J., Wilkinson, G. S., & Robertson, G. J., Glutting, J. J.
(2001). Wide Range Achievement Test 3 (wrat3).
In
(S. 259–274).
Understanding Psychological Assessment
Springer US. DOI: https://doi.org/10.1007/978-1-4615-1185-4
_13
Starkstein, S. E., Berthier, M., & Leiguarda, R. (1988). Bilateral oper-
cular syndrome and crossed aphemia due to a right insular lesion:
A clinicopathological study. Brain and Language, 34(2), 253–261.
DOI: https://doi.org/10.1016/0093-934X(88)90137-X, PMID:
3401694
Stokes, R. C., Venezia, J. H., & Hickok, G. (2019). The motor system’s
[modest] contribution to speech perception. Psychonomic
Bulletin and Review, 26(4), 1354–1366. DOI: https://doi.org/10
.3758/s13423-019-01580-2, PMID: 30945170
Venezia, J. H., Vaden, K. ICH., Rong, F., Maddox, D., Saberi, K., &
Hickok, G. (2017). Auditiv, visual and audiovisual speech
processing streams in superior temporal sulcus. Grenzen in
Human Neuroscience, 11, 174. DOI: https://doi.org/10.3389
/fnhum.2017.00174, PMID: 28439236, PMCID: PMC5383672
Watkins, K., & Paus, T. (2004). Modulation of motor excitability
during speech perception: The role of Broca’s area. Zeitschrift für
Cognitive Neuroscience, 16(6), 978–987. DOI: https://doi.org
/10.1162/0898929041502616, PMID: 15298785
Wilson, S. M. (2009). Speech perception when the motor system is
compromised. Trends in den Kognitionswissenschaften, 13(8), 329–330.
DOI: https://doi.org/10.1016/j.tics.2009.06.001, PMID:
19646917, PMCID: PMC4828932
Wilson, S. M., Saygin, A. P., Sereno, M. ICH., & Iacoboni, M. (2004).
Listening to speech activates motor areas involved in speech pro-
duktion. Naturneurowissenschaften, 7(7), 701–702. DOI: https://doi
.org/10.1038/nn1263, PMID: 15184903
Neurobiology of Language
353
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
2
3
3
3
5
1
9
2
8
8
9
9
N
Ö
_
A
_
0
0
0
3
7
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3