RESEARCH ARTICLE
Age-Related Differences in Auditory Cortex
Activity During Spoken Word Recognition
Chad S. Rogers1
Kristin J. Van Engen3
, Michael S. Jones2
, Sarah McConkey2
, Mitchell S. Sommers3, and Jonathan E. Peelle2
, Brent Spehar2
,
Keine offenen Zugänge
Tagebuch
1Abteilung für Psychologie, Union College, Schenectady, New York, USA
2Department of Otolaryngology, Washington University in St. Louis, St. Louis, MO, USA
3Department of Psychological and Brain Sciences, Washington University in St. Louis, St. Louis, MO, USA
Schlüsselwörter: speech perception, kognitives Altern, speech production
ABSTRAKT
Understanding spoken words requires the rapid matching of a complex acoustic stimulus
with stored lexical representations. The degree to which brain networks supporting spoken
word recognition are affected by adult aging remains poorly understood. In the current study
we used fMRI to measure the brain responses to spoken words in two conditions: an attentive
listening condition, in which no response was required, and a repetition task. Listeners were
29 young adults (aged 19–30 years) Und 32 older adults (aged 65–81 years) without self-reported
hearing difficulty. We found largely similar patterns of activity during word perception for
both young and older adults, centered on the bilateral superior temporal gyrus. Wie erwartet,
the repetition condition resulted in significantly more activity in areas related to motor planning
und Ausführung (including the premotor cortex and supplemental motor area) compared to
the attentive listening condition. Wichtig, Jedoch, older adults showed significantly
less activity in probabilistically defined auditory cortex than young adults when listening
to individual words in both the attentive listening and repetition tasks. Age differences in
auditory cortex activity were seen selectively for words (no age differences were present for
1-channel vocoded speech, used as a control condition), and could not be easily explained
by accuracy on the task, movement in the scanner, or hearing sensitivity (available on a subset
of participants). These findings indicate largely similar patterns of brain activity for young and
older adults when listening to words in quiet, but suggest less recruitment of auditory cortex
by the older adults.
EINFÜHRUNG
Understanding spoken words requires mapping complex acoustic signals to a listener’s stored
lexical representations. Evidence from neuropsychology and cognitive neuroscience provides
increasingly converging evidence about the roles of the bilateral temporal cortex (insbesondere
the superior temporal gyrus and the middle temporal gyrus) in processing speech acoustics
and recognizing single words (Binder et al., 2000; Hickok & Kacke, 2007; Peelle, Johnsrude,
& Davis, 2010). Jedoch, the degree to which the networks supporting spoken word recognition
change over our lifetime remains unclear. The goals of the current study were to test whether
young and older adults relied on different brain networks during successful spoken word recog-
Nation, and whether any age differences were related to the specific task.
Zitat: Rogers, C. S., Jones, M. S.,
McConkey, S., Spehar, B., Van Engen,
K. J., Sommers, M. S., & Peelle, J. E.
(2020). Age-related differences in
auditory cortex activity during spoken
word recognition. Neurobiology of
Language, 1(4), 452–473. https://doi.
org/10.1162/nol_a_00021
DOI:
https://doi.org/10.1162/nol_a_00021
zusätzliche Informationen:
https://doi.org/10.1162/nol_a_00021
Erhalten: 5 Marsch 2020
Akzeptiert: 11 August 2020
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Chad S. Rogers
rogersc@union.edu
Handling-Editor:
Ingrid Johnsrude
Urheberrechte ©: © 2020 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) Lizenz.
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
Important themes when considering older adults’ language processing include the degree to
which linguistic processing is preserved, and whether older adults may adopt different strategies
when understanding language compared to young adults (Peelle, 2019; Wingfield & Stine-
Morrow, 2000). Particularly important for spoken word recognition is that adult aging frequently
brings changes to both hearing sensitivity (Peelle & Wingfield, 2016) and cognitive ability (Park
et al., 2002). Daher, it is not surprising that older adults’ spoken word perception differs from that
of young adults, particularly in the presence of background noise (Humes, 1996). Older adults
tend to take longer to recognize words (Lash, Rogers, Zoller, & Wingfield, 2013; Wingfield,
Aberdeen, & Stine, 1991), make more recognition errors than young adults, and show increased
sensitivity to factors such as the number of phonological neighbors (competitors) associated with
a given target word (Sommers & Danielson, 1999). An open question centers on the brain
networks on which older adults rely during spoken word recognition. Of particular interest is
whether additional regions may be recruited to support successful recognition, compared to
those engaged by young adults.
A number of studies have investigated neural activity during older adults’ speech processing in
noise or other acoustic degradation, using an assortment of tasks and testing participants with
different levels of hearing (Bilodeau-Mercure, Lortie, Sato, Guitton, & Tremblay, 2015; Hwang,
Li, Wu, Chen, & Liu, 2007; Manan, Yusoff, Franz, & Mukari, 2017; Manan, Franz, Yusoff, &
Mukari, 2015; Wong et al., 2009). Harris, Dubno, Keren, Ahlstrom, and Eckert (2009), for exam-
Bitte, examined spoken word recognition in young and older adults. They varied the intelligibility
of the target items using low-pass filtering of the acoustic signal. During scanning, Teilnehmer
were asked to repeat back the word they heard. The authors found increased activity in regions
associated with word processing, including the auditory cortex and the premotor cortex, Wann
words were more intelligible; these intelligibility-related changes did not statistically differ
between young and older adults. Older adults did show more activation in the anterior cingulate
cortex and the supplemental motor area than the young adults did, suggesting a possible increase
in top-down executive control.
Age differences in speech understanding have also been studied in the context of sentence
comprehension. One common finding is that during successful sentence processing, older adults
show additional activity compared to young adults (z.B., in contralateral homologs to regions
seen in young adults, or in regions beyond the network activated by young adults; Peelle,
Troiani, Wingfield, & Grossman, 2010; Tyler et al., 2010). These findings have been interpreted
in a compensation framework in which older adults are less efficient using a core speech network
and need to recruit additional regions to support successful comprehension (Wingfield &
Grossman, 2006). Jedoch, at least some of this additional activity has been shown to be related
to the tasks performed by participants in the scanner, which frequently contain metalinguistic
decisions not required during everyday conversation (Davis, Zhuang, Wright, & Tyler, 2014).
Daher, it may be that core language computations are well-preserved in aging (Campbell et al.,
2016; Shafto & Tyler, 2014).
The role of executive attention in older adults’ spoken word recognition has also been of sig-
nificant interest. Listening to speech that is acoustically degraded can result in perception errors,
after which listeners must re-engage attention systems to support successful listening. The cingulo-
opercular network, an executive attention network (Neta et al., 2015; Power & Petersen, 2013),
shows increased activity following perception errors (similar to error-related activity in other
domains). Crucially, when listening to spoken words in background noise, increased cingulo-
opercular activity following one trial is associated with recognition success on the following trial
(Vaden et al., 2016; Vaden et al., 2013), consistent with a role in maintaining task-related attention
(Eckert et al., 2009).
Neurobiology of Language
453
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
An important challenge when considering the performance of listeners with hearing loss is
that words may not be equally intelligible to all listeners. A common measure of accuracy in
spoken word recognition is to ask listeners to repeat each word after hearing it; Jedoch, this type
of task requires motor responses, which may obscure activations related to speech perception
and increase participant motion in the scanner (Gracco, Tremblay, & Pike, 2005). Zusätzlich,
differences in the brain regions coordinating speech production in older adults (Bilodeau-
Mercure & Tremblay, 2016; Tremblay, Sato, & Deschamps, 2017) may interfere with clear
measurements of activity during perception and recognition. The degree to which motor effects
resulting from word repetition may obscure activity related to speech perception is unclear. In
sentence processing tasks, task effects can be significant (Davis et al., 2014), and if not accounted
for may obscure what are actually consistent patterns of language-related activity across the life-
Spanne (Campbell et al., 2016).
In the current study we investigated spoken word processing in young and older adult listeners
in the absence of background noise. We compared paradigms requiring words to be repeated
with “attentive listening” (no motor response required). Our interest is, Erste, whether age differ-
ences exist in the brain networks supporting spoken word recognition, und zweitens, whether these
differences are affected by the choice of task. Daher, our primary analyses will focus on activity
seen for words (greater than noise) in the experimental conditions.
The influence of psycholinguistic factors on spoken word recognition has long been appre-
ciated. In a secondary set of analyses, we will investigate whether word frequency or phono-
logical neighborhood density modulate activity during spoken word recognition. Obwohl
behavioral and electrophysiological studies suggest that high frequency words are processed
more quickly than low frequency words, the degree to which this might be captured in fMRI is
unclear. Ähnlich, although neighborhood density effects are widely reported in behavioral
Studien (with words from dense neighborhoods typically being more difficult to process), Die
degree to which lexical competition effects may differ with age is unclear.
MATERIALS AND METHODS
Stimuli, Daten, and analysis scripts are available from https://osf.io/vmzag/.
Teilnehmer
We recruited two groups of participants (young and older adults) for this study. The young
adults were 29 self-reported healthy, right-handed adults, aged 19–30 years (M = 23.8, SD =
2.9, 19 weiblich), and were recruited via the Washington University in St. Louis Department of
Psychological and Brain Sciences Subject Pool. Older adult participants were 32 self-reported
healthy, right-handed adults, aged 65–81 years (M = 71.0, SD = 5.0, 17 weiblich). All participants
self-reported themselves to be native speakers of American English with no history of neurolog-
ical difficulty, and with normal hearing (and no history of a diagnosed hearing problem).
Participants were compensated for their participation, and all provided informed consent com-
mensurate with practices approved by the Washington University in St. Louis Institutional
Review Board.
Audiograms were collected on a subset of eight young and nine older participants using
pure-tone audiometry (Figure 1a). We summarized hearing ability using a better-ear pure tone
average (PTA) bei 1, 2, Und 4 kHz. PTAs in participants’ better hearing ears ranged from −3.33
Zu 8.33 dB HL in young adults (M = 2.92, SD = 4.15), Und 8.33 Zu 23.3 dB HL in older adults
(M = 23.3, SD = 9.17).
Neurobiology of Language
454
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
Figur 1.
Experiment overview. (A) Audiograms for the subset of participants on whom hearing was available for left and right ears. Individuell
participants are shown in thin lines, group means in thick lines. (B) Frequency of occurrence and phonological neighborhood density for the
240 experimental items. (C) Task design for attentive listening and word repetition tasks. (D) Behavioral accuracy for the repetition condition for
young and older adults. HAL = Hyperspace Analogue to Language, EPI = echo planar imaging.
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Materials
Stimuli for this study were 375 monosyllabic consonant-vowel-consonant words. The auditory
stimuli were recorded at 48 kHz using a 16-bit digital-to-analog converter with an Audio
Technica 2035 microphone in a quiet room. Words were spoken by a female speaker with
a standard American dialect. Root-mean-square amplitude of the stimuli was equated.
Out of the full set of words, 75 words were vocoded using a single channel with white noise as
a carrier signal (Shannon, Zeng, Kamath, Wygonski, & Ekelid, 1995) using jp_vocode.m from
http://github.com/jpeelle/jp_matlab. These stimuli were used for an unintelligible baseline
“noise” condition. The remaining 300 words were divided into five lists of 60 Wörter, verwenden
MATCH software (Van Casteren & Davis, 2007), and were balanced for word frequency (als
measured by the log of the Hyperspace Analogue to Language dataset), orthographic length,
concreteness (Brysbaert, Warriner, & Kuperman, 2014), and familiarity (Balota et al., 2007).
The distribution of word frequency and phonological neighborhood density are shown in
Figure 1b.
One of these lists was combined with 15 of the noise vocoded words and used for word
repetition task practice outside of the scanner. The remaining four lists of 60 words served as
Neurobiology of Language
455
Spoken word recognition in older adults
the critical items inside the scanner, with half of the lists used for attentive listening (120 total
Wörter) and the other half for word repetition (120 total words). Word lists were counterba-
lanced such that each word was presented in both “listen” and “repeat” conditions across
Teilnehmer.
Verfahren
Prior to scanning, participants were taken to a quiet room. (The room was not sound isolated and
low frequency noise from the building heating, ventilation, and air conditioning system was typi-
cally present.) During that time participants provided informed consent, completed demographic
questionnaires, and a subset had their hearing tested using a calibrated Maico MA40 portable
audiometer (Maico Diagnostics, Inc., Eden Prairie MN) by an audiologist-trained researcher.
Participants were then instructed for the two tasks they would perform in the scanner:
attentive listening and word repetition. During attentive listening, participants were asked to
stay alert, still, and keep their eyes focused on a fixation cross while listening to a sequence of
auditory sounds, including words, silence, and noise (single-channel noise vocoded words).
During word repetition, participants were asked to do the same as in attentive listening, mit
the addition of repeating the word they just heard aloud. Participants were instructed to repeat
the words following the volume acquisition after each word (Figure 1c). Participants were told
to give their best guess if they could not understand a word. Participants practiced a simulation
of the word repetition task until the experimenter was confident that the participant understood
the pacing and the nature of the task. Sound levels were adjusted to achieve audible presen-
tations at the beginning of the study and thereafter not adjusted.
Functional MRI scanning took place over the course of four scanning blocks, where partic-
ipants alternated between blocks of attentive listening and word repetition (Figure 1c). Der
order of blocks was counterbalanced such that participants were equally likely to begin with
a word repetition or an attentive listening block. During word repetition, participants’ spoken
responses were recorded using an in-bore Fibersound optical microphone. These responses
were scored for accuracy offline by a research assistant (Figure 1d).
MRI Data Acquisition and Processing
The MRI data collected in this study are available from https://openneuro.org/datasets/ds002382
(Poldrack et al., 2013). MRI data were acquired using a Siemens Prisma scanner (Siemens
Medical Systems) bei 3 T equipped with a 32-channel head coil. Scan sequences began with a
T1-weighted structural volume using an MPRAGE sequence (repetition time [TR] = 2.4 S, echo
Zeit [DER] = 2.2 MS, flip angle = 8°, 300 × 320 Matrix, Voxelgröße = 0.8 mm isotropic). Blood
oxygenation level-dependent fMRI images were acquired using a multiband echo planar imag-
ing sequence (Feinberg et al., 2010; TR = 3.07 S, TA = 0.770 S, TE = 37 MS, flip angle = 37°, voxel
size = 2 mm isotropic, multiband factor = 8). (The flip angle was suboptimal due to an error
setting up the sequences; although discovered partway through the study, we left it unchanged
to maintain consistent data quality. With a TR of ~3 s we would expect a better signal-to-noise ratio
with a flip angle of 90°.) We used a sparse imaging design in which there was a 2.3 s delay between
scanning acquisitions and the TR was longer than the acquisition time to allow for minimal scanning
noise during stimulus presentation and audio recording of participant responses (Edmister,
Talavage, Ledden, & Weisskoff, 1999; Hall et al., 1999).
Neurobiology of Language
456
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
Analysis of the MRI data was performed using Automatic Analysis version 5.4.0 (Cusack et al.,
2015; RRID:SCR_003560), which scripted a combination of SPM12 ( Wellcome Trust Centre for
Neuroimaging) Ausführung 7487 (RRID:SCR_007037) and FMRIB Software Library (FSL; FMRIB
Analysis Group; Jenkinson, Beckmann, Behrens, Woolrich, & Schmied, 2012) Ausführung 6.0.1
(RRID:SCR_002823).
Data were realigned using rigid-body image registration, and functional data were coregis-
tered with the bias-corrected T1-weighted structural image. Spatial and functional images were
normalized to MNI space using a unified segmentation approach (Aschenbrenner & Friston, 2005),
and resampled to 2 mm. Endlich, the functional data were smoothed using an 8 mm full width at
half maximum Gaussian kernel.
For the attentive listening condition, we did not have measures of accuracy, so we analyzed
all trials. For the repetition condition, we analyzed only trials associated with correct responses.
For both tasks, we modeled the noise condition in addition to words. Endlich, we included three
parametric modulators for word events: word frequency, phonological neighborhood density,
and their interaction. To avoid order effects (Mumford, Polina, & Poldrack, 2015), these were not
orthogonalized.
Motion effects were of particular importance given that participants were speaking during the
repetition condition. To mitigate the effects of motion, we used a thresholding approach in
which high motion frames were individually modeled for each subject using a delta function
in the general linear model (sehen, z.B., Siegel et al., 2014). Motion was quantified using framewise
displacement (FD), calculated from the six motion parameters estimated during realignment,
assuming the head is a sphere having a radius of 50 mm (Power, Barnes, Snyder, Schlaggar,
& Petersen, 2012). We then chose an FD threshold (0.561) that we used for all participants.
Our rationale was that some participants move more, and thus produce worse data; we therefore
wanted to use a single threshold for all participants, resulting in more data exclusion from high-
motion participants. This threshold resulted in 2.2–19.4% (M = 6.21, SD = 4.45) data exclusion
for the young adults and 2.8–58.4% (M = 22.6, SD = 15.3) data exclusion for the older adults. Für
each frame exceeding this threshold, we added a column to that participant’s design matrix con-
sisting of a delta function at the time point in question, which effectively excludes the variance of
that frame from the model.
Contrast images from single subject analyses were analyzed at the second level using permu-
tation testing (FSL randomise; 5,000 permutations; https://fsl.fmrib.ox.ac.uk/fsl/fslwiki/FSL), mit
a cluster-forming threshold of p < 0.001 (uncorrected) and results corrected for multiple com-
parisons based on cluster extent ( p < 0.05). Images (contrast images and unthresholded t maps)
are available from https://identifiers.org/neurovault.collection:6735 (Gorgolewski et al., 2015).
Anatomical localization was performed using converging evidence from author experience
(Devlin & Poldrack, 2007) viewing statistical maps overlaid in MRIcroGL (Rorden & Brett,
2000), supplemented by atlas labels (Tzourio-Mazoyer et al., 2002).
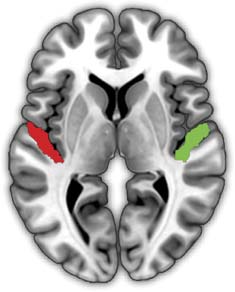
For region of interest (ROI) analysis of primary auditory cortex, we used probabilistic maps
based on postmortem human histological staining (Morosan et al., 2001), available in the SPM
Anatomy toolbox (Eickhoff et al., 2005; RRID:SCR_013273). We created a binary mask for re-
gions Te1.0 and Te1.1 and then extracted parameter estimates for noise and word contrasts for
the attentive listening and repetition conditions from each participant’s first-level analyses by
averaging over all voxels in each ROI (left auditory, right auditory).
Outputs from analysis stages used for quality control are available from https://osf.io/vmzag/
in the aa_report folder.
Neurobiology of Language
457
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
n
o
_
a
_
0
0
0
2
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spoken word recognition in older adults
RESULTS
Behavioral Data
We analyzed the accuracy data using a linear mixed effects analysis, implemented using the
lme4 and lmerTest packages in R version 3.6.2 (Bates, Mächler, Bolker, & Walker, 2015;
Kuznetsova, Brockhoff, & Christensen, 2017; RRID:SCR_001905). Because trial-level accuracy
data was binary, we used logistic regression. We first tested for age differences using a model that
included age group as a fixed factor and subject as a random factor:
m0 <- glmer(accuracy ~ age_group + (1 | subject),
data = df, family = "binomial"> older adults
Region
Left superior temporal gyrus
Left Heschl’s gyrus
Left Heschl’s gyrus
Size (μl)
8,472
Right superior temporal gyrus
3,400
Right superior temporal gyrus
t score
6.29
4.18
4.08
4.84
3.37
X
−62
−40
−42
52
62
Coordinates
j
−16
−30
−24
−16
8
z
8
10
12
10
2
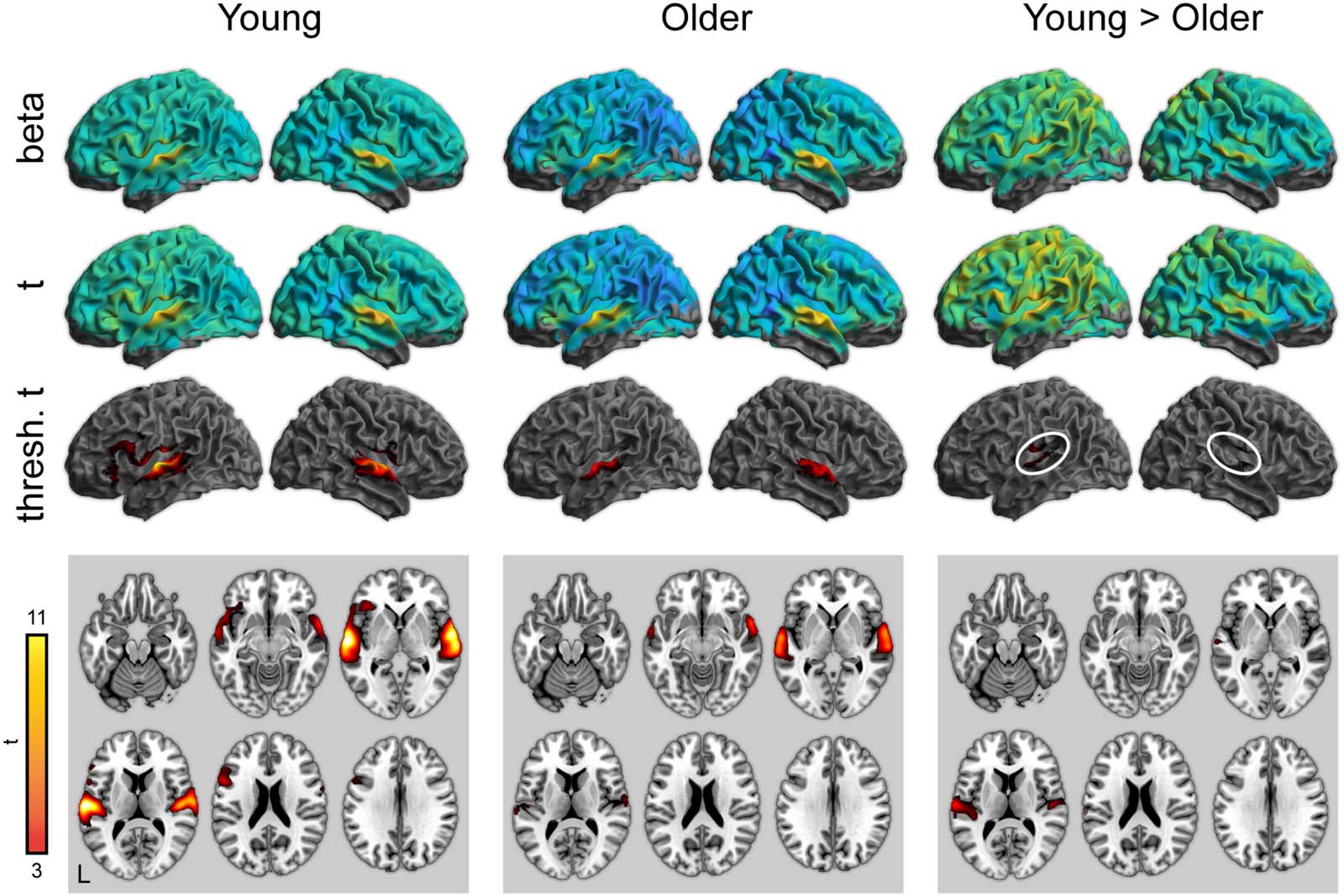
DISKUSSION
We used fMRI to examine neural activity during spoken word recognition in quiet for young
and older adult listeners. In both ROI and whole-brain analyses, we found converging evi-
dence for reduced activity in the auditory cortex for the older adults. The age differences in
auditory cortex activation were present in both the attentive listening task and the word rep-
etition task: Although the repetition task resulted in more widespread activation overall, pat-
terns of age-related differences in the auditory cortex were comparable.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
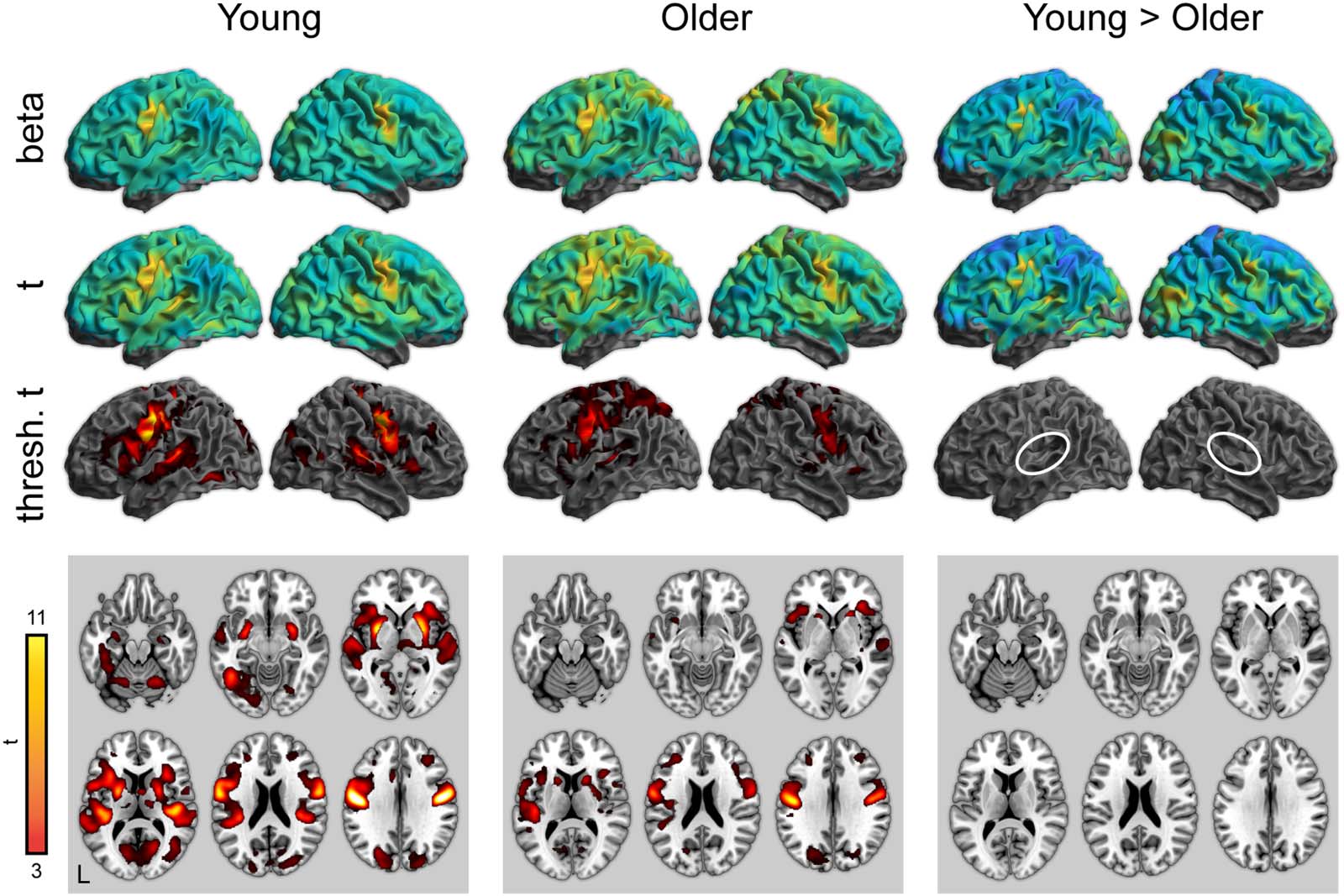
Figur 4. Whole-brain activity for the repetition condition (correct responses only). Top: Unthresholded parameter estimates. Mitte:
Unthresholded t maps. Bottom: Thresholded t maps ( P < 0.05, cluster corrected). White ovals highlight left and right auditory cortex.
Neurobiology of Language
463
Spoken word recognition in older adults
Table 6.
Peak activations for repetition condition greater than noise, young adults
Region
Left superior temporal gyrus
Size (μl)
581,432
t score
14
Left postcentral gyrus
Left postcentral gyrus
Right postcentral gyrus
Left postcentral gyrus
Left putamen
Right superior temporal gyrus
Supplemental motor area
Right superior temporal gyrus
Right superior temporal gyrus
Right putamen
Right precentral gyrus
Left paracentral lobule
Left Heschl’s gyrus
Left inferior frontal gyrus
Left inferior temporal gyrus
Left insula
Left inferior parietal cortex
Right insula
Dorsal anterior cingulate
13.9
13.8
13.7
13.4
12.9
12.5
12.4
12.3
11.4
10.6
10.6
10.5
9.89
9.58
9.13
8.84
8.41
8.19
8.11
x
−60
−42
−48
44
−52
−24
56
0
52
66
28
20
−18
−36
−52
−44
−32
−38
34
−8
Coordinates
y
−14
−16
−14
−12
−8
0
−10
0
−16
−20
0
−28
−30
−30
10
−56
22
−36
20
12
z
4
38
40
36
30
4
4
58
6
2
−4
60
60
14
22
−10
4
42
6
38
There are a number of possible explanations for older adults’ reduced activity during spo-
ken word recognition. One possibility is that age differences in intelligibility might play a role.
Intelligible speech is associated with increased activity in a broad network of frontal and tem-
poral regions (Davis & Johnsrude, 2003; Kuchinsky et al., 2012), and in prior studies of older
adults, intelligibility has correlated with auditory cortex activity (Harris et al., 2009). We re-
stricted our analyses to correct responses in the repetition condition, and found no statistical
support for a relationship between intelligibility and auditory cortex activation (although nu-
merically, participants with better accuracy showed more activity than participants with worse
accuracy).
The fact that young and older adults showed comparable activity in the auditory cortex
during noise trials, with age differences emerging for word recognition trials, is significant.
Group differences in activation could be driven not only by neural processing, but also by
such factors as neurovascular coupling, goodness-of-fit of a canonical hemodynamic re-
sponse, or movement within the scanner—in other words, artifacts that might differentially
Neurobiology of Language
464
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
n
o
_
a
_
0
0
0
2
1
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spoken word recognition in older adults
Table 7.
Peak activations for repetition condition greater than noise, older adults
Region
Left postcentral gyrus
Size (μl)
278,528
t score
10.1
9.49
9.35
9.26
9.19
8.14
8.01
7.84
7.68
7.51
7.39
6.87
6.76
6.72
6.48
6.44
6.44
6.32
6.25
5.97
4.13
3.98
3.6
3.56
3.41
3.35
3.33
3.3
3.04
3,784
Supplemental motor area
Right postcentral gyrus
Left postcentral gyrus
Right superior temporal gyrus
Right postcentral gyrus
Left superior temporal gyrus
Left superior temporal gyrus
Left superior parietal cortex
Left inferior frontal gyrus
Left superior temporal gyrus
Left precentral gyrus
Right insula
Right putamen
Left insula
Left postcentral
Left caudate
Right insula
Left inferior parietal cortex
Fornix
Left thalamus
Superior cerebellar pedunculus
Left superior cerebellar pedunculus
Right superior cerebellar pedunculus
Right thalamus
Right thalamus
Left superior cerebellar pedunculus
Right thalamus
Left superior cerebellar pedunculus
Neurobiology of Language
x
−44
−2
42
−56
64
56
−60
−44
−26
−44
−62
20
32
18
−30
−18
−16
36
−46
6
−12
0
−4
2
12
16
−6
14
−10
Coordinates
y
−14
4
−12
−4
−18
−4
−14
−22
−66
8
−28
−28
26
16
24
−30
14
18
−32
0
−18
−24
−28
−14
−20
−18
−34
−18
−34
z
34
56
36
24
0
28
2
10
52
26
4
60
0
0
4
58
8
8
40
6
0
2
−14
−8
2
−2
−2
8
−20
465
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
n
o
_
a
_
0
0
0
2
1
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spoken word recognition in older adults
Table 8.
Peak activations for repetition condition greater than noise, young > older adults
Size (μl)
5,224
t score
5.22
Region
Right Heschl’s gyrus
Right Heschl’s gyrus
Right superior temporal sulcus
Right superior temporal gyrus
Right superior temporal gyrus
Right superior temporal gyrus
Left Heschl’s gyrus
4,600
Left superior temporal gyrus
Left superior temporal gyrus
Left superior temporal gyrus
Left Heschl’s gyrus
Left superior temporal gyrus
Left superior temporal gyrus
Left postcentral gyrus
4,248
Left postcentral gyrus
Left postcentral gyrus
Left postcentral gyrus
4.99
4.18
4.08
3.79
3.62
4.92
4.17
3.78
3.66
3.47
3.29
3.19
5.26
5.21
5.12
3.88
X
48
40
60
68
56
48
−36
−64
−62
−60
−46
−62
−52
−50
−46
−42
−54
Coordinates
j
−20
−26
−30
−26
−8
−34
−30
−18
−32
−18
−24
−38
−16
−14
−16
−18
−8
z
10
16
0
8
4
10
14
8
14
−4
6
8
2
44
42
40
28
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
impact model parameter estimates in young and older adults but are not of theoretical interest
in diesem Zusammenhang. Although impossible to completely rule out, the selective age differences for
Rede (but not noise) are consistent with a condition-specific—and thus we argue, neural—
Deutung.
Recent evidence suggests age-related changes in temporal sensitivity in auditory regions
can be detected with fMRI (Erb, Schmitt, & Obleser, 2020). Although our current stimuli do
not allow us to explore specific acoustic features, one possibility is that the age-related differ-
ences in auditory activity we observed reflect well-known changes in auditory cortical pro-
cessing that occur in normal aging (Peelle & Wingfield, 2016). Given the increased
acoustic complexity of the words relative to noise, acoustic processing differences might drive
overall response differences. Such changes may also reflect decreased stimulation as a result of
hearing loss; we had insufficient data to rule out this possibility. It is important to note that we
cannot completely rule out audibility effects. Even though we limited our responses to correct
identification trials, specific acoustic features may still have been less audible for the older
Erwachsene. It remains an open question whether varying the presentation level of the stimuli would
change the age effects we observed.
Age differences in auditory processing are not the only explanation for our results. The au-
ditory cortex is positioned in a hierarchy of speech processing regions that include both
Neurobiology of Language
466
Spoken word recognition in older adults
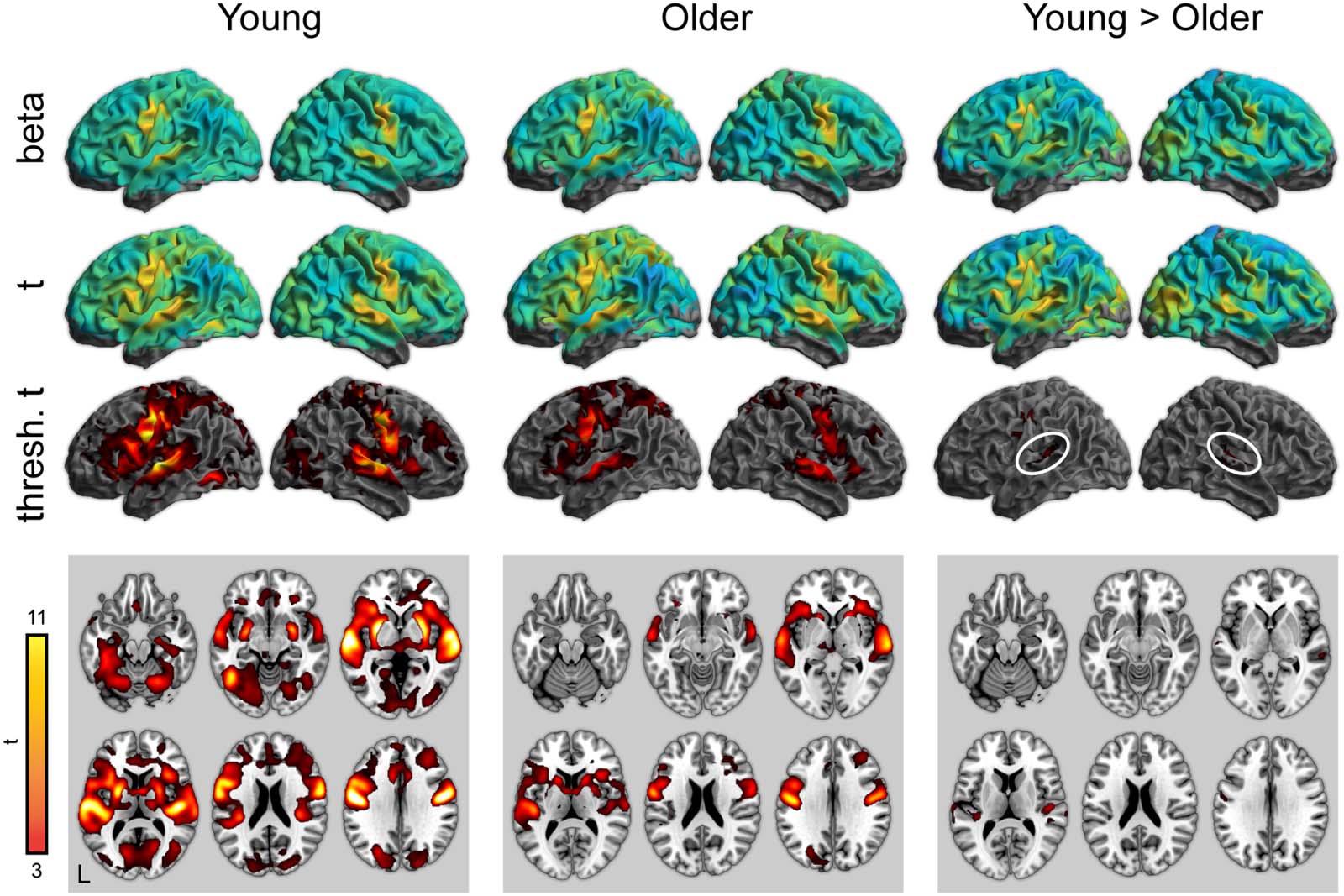
Figur 5. Whole-brain activity for the repetition condition > attentive listening. Top: Unthresholded parameter estimates. Mitte:
Unthresholded t maps. Bottom: Thresholded t maps ( P < 0.05, cluster corrected). White ovals highlight left and right auditory cortex.
There were no significant differences between young and older adults in the repetition > listening contrast.
ascending and descending projections (Davis & Johnsrude, 2007; Peelle, Johnsrude, et al.,
2010). The auditory cortex not only is sensitive to changes in acoustic information, but also
reflects top-down effects of expectation and prediction (Signoret, Johnsrude, Classon, &
Rudner, 2018; Sohoglu, Peelle, Carlyon, & Davis, 2012; Wild et al., 2012). Daher, the observed
age differences in the auditory cortex may reflect differential top-down modulation of auditory
activity in young and older adult listeners.
In der Tat, prior to conducting this study, we expected to observe increased activity (z.B., In
the prefrontal cortex) for older adults relative to young adults, reflecting top-down compensa-
tion for reduced auditory sensitivity. Such activity would be consistent with increased cogni-
tive demand during speech perception in listeners with hearing loss or other acoustic
Herausforderungen (Peelle, 2018; Pichora-Fuller et al., 2016). Although we were somewhat surprised
not to see this, in retrospect, perhaps it would be expected. The stimuli in the current study
were presented in quiet, and thus may not have challenged perception sufficiently to robustly
engage frontal brain networks. We conclude that during perception of acoustically clear
Wörter, older adults do not seem to require additional resources from the frontal cortex;
whether this changes with increasing speech demands (either acoustic or linguistic) remains
an open question.
We did not observe significant effects of either word frequency or phonological neighbor-
hood density on activity during spoken word recognition. These results stand in contrast to prior
studies showing frequency effects in visual word perception in fMRI (Hauk, Davis, &
Pulvermüller, 2008; Kronbichler et al., 2004), and word frequency effects in electrophysiolog-
ical responses (Embick, Hackl, Schäffer, Kelepir, & Marantz, 2001). Prior fMRI studies of lexical
Neurobiology of Language
467
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
Tisch 9.
Peak activations for word recognition in the repetition condition greater than listening condition, young adults
Size (μl)
311,408
Region
Left postcentral gyrus
Right postcentral gyrus
Left putamen
Supplemental motor area
Right postcentral gyrus
Left Heschl’s gyrus
Right superior temporal gyrus
Left postcentral gyrus
Right Heschl’s gyrus
Right putamen
Left insula
Left inferior frontal gyrus
Right insula
Left superior temporal sulcus
Anterior cingulate
Right superior temporal gyrus
Left inferior parietal cortex
Right calcarine sulcus
Right cerebellum
t score
13.8
13.8
10.4
9.97
9.81
9.36
9.11
9.06
8.96
8.91
7.16
7.13
7.11
6.23
6
5.61
5.6
5.47
5.31
X
−44
44
−24
0
20
−36
46
−18
38
28
−32
−46
34
−52
−8
64
−38
18
24
Coordinates
j
−14
−12
0
−2
−28
−32
−20
−30
−26
0
20
8
22
−42
12
−24
−36
−66
−60
z
36
36
4
60
60
14
8
60
14
−4
8
20
4
6
38
4
42
8
−22
competition (including phonological neighborhood density) have been mixed, with some
studies finding effects (Zhuang, Randall, Stamatakis, Marslen-Wilson, & Tyler, 2011) and others
nicht (Binder et al., 2003). It could be that a wider range of frequency or density or a greater num-
ber of stimuli would be needed to identify such effects.
Endlich, we found largely comparable age differences in the attentive listening and repetition
conditions in the auditory cortex. The similarity of the results suggests that using a repetition task
may be a reasonable choice in studies of spoken word recognition: Although repetition tasks
necessarily engage regions related to articulation and hearing one’s own voice, in our data these
were not differentially affected by age. An advantage of using a repetition task, Natürlich, is that trial-
by-trial accuracy measures can be obtained, which are frequently useful. It is worth noting that our
finding of comparable activity in young and older adults for attentive listening and repetition tasks
may not generalize to other stimuli or tasks (Campbell et al., 2016; Davis et al., 2014).
A significant limitation of our current study is that we only collected hearing sensitivity data
on a minority of our participants. Daher, although we saw a trend toward poorer hearing being
associated with reduced auditory cortex activation, it is challenging to draw any firm conclu-
sions regarding the relationship between hearing sensitivity and brain activity. Prior studies
using sentence-level materials have found relationships between hearing sensitivity and brain
Neurobiology of Language
468
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
Tisch 10. Peak activations for word recognition in the repetition condition greater than listening condition, older adults
Region
Left postcentral gyrus
Size (μl)
238,712
t score
10.21
Right postcentral gyrus
Supplemental motor area
Right postcentral gyrus
Left Heschl’s gyrus
Left superior parietal cortex
Left postcentral gyrus
Right precentral gyrus
Left precentral gyrus
Right putamen
Right caudate
Right insula
Left inferior parietal cortex
Anterior cingulate
Left postcentral gyrus
Left insula
Right superior parietal cortex
Left insula
Left caudate
Left precentral gyrus
Right superior temporal gyrus
4,632
Right Heschl’s gyrus
Right superior temporal gyrus
Right superior temporal gyrus
Right posterior insula
9.04
7.56
7.43
6.45
6.43
6.03
6.02
6
5.88
5.84
5.73
5.69
5.49
5.44
5.38
5.29
5.28
5.16
5.13
4.12
4.11
4.1
4.09
3.14
X
−44
42
−2
54
−42
−28
−18
20
−46
20
18
34
−44
−6
−26
−46
18
−32
−14
−32
58
34
64
52
32
Coordinates
j
−14
−12
2
−4
−24
−64
−30
−28
2
16
14
28
−32
14
−40
14
−68
24
14
−2
−22
−24
−16
−16
−24
z
34
36
56
26
10
54
60
60
38
0
6
0
40
42
62
−2
56
4
8
64
0
4
0
4
16
activity in both young (Lee et al., 2018) und älter (Peelle, Troiani, Grossman, & Wingfield,
2011) Erwachsene. Future investigations with a larger sample of participants with hearing data will
be needed to further explore the effects of hearing in spoken word recognition.
From a broader perspective, the link between spoken word recognition and everyday commu-
nication is not always straightforward. Much of our everyday communication occurs in the context
of semantically meaningful, coherent sentences, frequently with the added availability of visual
Neurobiology of Language
469
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
speech and gesture cues. Given potential age differences in reliance on many of these cues—
including older adults’ seemingly greater reliance on semantic context (Rogers, 2016; Rogers,
Jacoby, & Sommers, 2012; Wingfield & Lindfield, 1995)—it seems likely that our findings using
isolated spoken words cannot be extrapolated to richer naturalistic settings.
Zusammenfassend, we observed largely overlapping brain regions supporting spoken word recogni-
tion in young and older adults in the absence of background noise. Older adults showed less
activity than young adults in the auditory cortex when listening to words, but not noise. Diese
patterns of age difference were present regardless of the task (attentive listening vs. repetition).
ACKNOWLEDGMENTS
Research reported here was funded by grant R01 DC014281 from the US National Institutes of
Health. The multiband echo planar imaging sequence was provided by the University of
Minnesota Center for Magnetic Resonance Research. We are grateful to Linda Hood for assis-
tance with data collection, and to Henry Greenstein, Ben Muller, Olivia Murray, Connor
Perkins, and Tracy Zhang for help with data scoring.
FUNDING INFORMATION
Jonathan E. Peelle, National Institute on Deafness and Other Communication Disorders (http://
dx.doi.org/10.13039/100000055), Award ID: R01 DC014281.
BEITRÄGE DES AUTORS
Chad S. Rogers: Konzeptualisierung: Equal; Datenkuration: Equal; Untersuchung: Equal; Project
administration: Equal; Aufsicht: Supporting; Validierung: Equal; Writing–Review & Editing:
Equal. Michael S. Jones: Formale Analyse: Lead; Methodik: Equal; Software: Lead; Validierung:
Lead; Writing–Review & Editing: Equal. Sarah McConkey: Untersuchung: Equal; Project administra-
tion: Equal; Writing–Review & Editing: Equal. Brent Spehar: Konzeptualisierung: Equal;
Untersuchung: Supporting; Ressourcen: Supporting; Writing–Review & Editing: Equal. Kristin J. Van
Engen: Konzeptualisierung: Equal; Akquise von Fördermitteln: Supporting; Projektverwaltung: Equal;
Writing–Review & Editing: Equal. Mitchell S. Sommers: Konzeptualisierung: Equal; Funding acqui-
sition: Supporting; Projektverwaltung: Supporting; Writing–Review & Editing: Equal. Jonathan
E. Peelle: Konzeptualisierung: Equal; Datenkuration: Equal; Formale Analyse: Equal; Funding acqui-
sition: Lead; Projektverwaltung: Equal; Aufsicht: Lead; Visualisierung: Lead; Writing–
Original Draft: Lead; Writing–Review & Editing: Equal.
VERWEISE
Aschenbrenner, J., & Friston, K. J. (2005). Unified segmentation.
NeuroImage, 26, 839–851. DOI: https://doi.org/10.1016/j.neuro-
image.2005.02.018, PMID: 15955494
Balota, D. A., Yap, M. J., Cortese, M. J., Hutchison, K. A., Kessler,
B., Loftis, B., … Treiman, R. (2007). The English lexicon project.
Behavior Research Methods, 39, 445–459. DOI: https://doi.org/
10.3758/BF03193014, PMID: 17958156
Bates, D., Mächler, M., Bolker, B. M., & Walker, S. C. (2015). Fitting
linear mixed-effects models using lme4. Journal of Statistical
Software, 67(1), 1–48. DOI: https://doi.org/10.18637/jss.v067.i01
Bilodeau-Mercure, M., Lortie, C. L., Sato, M., Guitton, M., &
Tremblay, P. (2015). The neurobiology of speech perception
decline in aging. Brain Structure & Function, 220, 979–997. DOI:
https://doi.org/10.1007/s00429-013-0695-3, PMID: 24402675
Bilodeau-Mercure, M., & Tremblay, P. (2016). Age differences in
sequential speech production: Articulatory and physiological factors.
Journal of the American Geriatrics Society, 64(11), e177–e182. DOI:
https://doi.org/10.1111/jgs.14491, PMID: 27783395
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S.,
Springer, J. A., Kaufman, J. N., & Possing, E. T. (2000). Human
temporal lobe activation by speech and nonspeech sounds.
Hirnrinde, 10(5), 512–528. DOI: https://doi.org/10.1093/
cercor/10.5.512, PMID: 10847601
Binder, J. R., McKiernan, K. A., Parsons, M. E., Westbury, C. F.,
Possing, E. T., Kaufman, J. N., & Buchanan, L. (2003). Neuronal
correlates of lexical access during visual word recognition.
Zeitschrift für kognitive Neurowissenschaften, 15(3), 373–393. DOI: https://
doi.org/10.1162/089892903321593108, PMID: 12729490
Neurobiology of Language
470
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
Brysbaert, M., Warriner, A. B., & Kuperman, V. (2014). Concreteness
ratings for 40 thousand generally known English word lemmas.
Behavior Research Methods, 46(3), 904–911. DOI: https://doi.
org/10.3758/s13428-013-0403-5, PMID: 24142837
Campbell, K. L., Samu, D., Davis, S. W., Geerligs, L., Mustafa, A.,
Tyler, L. K., & for Cambridge Centre for Aging and Neuroscience.
(2016). Robust resilience of the frontotemporal syntax system to
Altern. Zeitschrift für Neurowissenschaften, 36, 5214–5227. DOI: https://doi.
org/10.1523/JNEUROSCI.4561-15.2016, PMID: 27170120,
PMCID: PMC4863058
Cusack, R., Vicente-Grabovetsky, A., Mitchell, D. J., Wild, C. J., Auer,
T., Linke, A. C., & Peelle, J. E. (2015). Automatic analysis (aa):
Efficient neuroimaging workflows and parallel processing using
Matlab and XML. Frontiers in Neuroinformatics, 8, 90. DOI: https://
doi.org/10.3389/fninf.2014.00090, PMID: 25642185, PMCID:
PMC4295539
Davis, M. H., & Johnsrude, ICH. S. (2003). Hierarchical processing in
spoken language comprehension. Zeitschrift für Neurowissenschaften, 23(8),
3423–3431. DOI: https://doi.org/10.1523/JNEUROSCI.23-08-
03423.2003, PMID: 12716950, PMCID: PMC6742313
Davis, M. H., & Johnsrude, ICH. S. (2007). Hearing speech sounds:
Top-down influences on the interface between audition and
speech perception. Hörforschung, 229, 132–147. DOI:
https://doi.org/10.1016/j.heares.2007.01.014, PMID: 17317056
Davis, S. W., Zhuang, J., Wright, P., & Tyler, L. K. (2014). Altersbedingt
sensitivity to task-related modulation of language-processing
Netzwerke. Neuropsychologie, 63, 107–115. DOI: https://doi.org/
10.1016/j.neuropsychologia.2014.08.017, PMID: 25172389,
PMCID: PMC4410794
Devlin, J. T., & Poldrack, R. A. (2007). In praise of tedious anatomy.
NeuroImage, 37(4), 1033–1041. DOI: https://doi.org/10.1016/j.
neuroimage.2006.09.055, PMID: 17870621, PMCID: PMC1986635
Eckert, M. A., Menon, V., Walczak, A., Ahlstrom, J., Denslow, S.,
Horwitz, A., & Dubno, J. R. (2009). At the heart of the ventral
attention system: The right anterior insula. Kartierung des menschlichen Gehirns,
30, 2530–2541. DOI: https://doi.org/10.1002/hbm.20688, PMID:
19072895, PMCID: PMC2712290
Edmister, W. B., Talavage, T. M., Ledden, P. J., & Weisskoff, R. M.
(1999). Improved auditory cortex imaging using clustered volume
acquisitions. Kartierung des menschlichen Gehirns, 7, 89–97. DOI: https://doi.
org/10.1002/(WISSENSCHAFT)1097-0193(1999)7:2<89::AID-HBM2>3.0.
CO;2-N, PMID: 9950066, PMCID: PMC6873308
Eickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C., Fink, G. R.,
Amunts, K., & Zilles, K. (2005). A new SPM toolbox for combining
probabilistic cytoarchitectonic maps and functional imaging data.
NeuroImage, 25(4), 1325–1335. DOI: https://doi.org/10.1016/j.
neuroimage.2004.12.034, PMID: 15850749
Embick, D., Hackl, M., Schäffer, J., Kelepir, M., & Marantz, A. (2001).
A magnetoencephalographic component whose latency reflects
lexical frequency. Kognitive Gehirnforschung, 10(3), 345–348. DOI:
https://doi.org/10.1016/s0926-6410(00)00053-7, PMID: 11167059
Erb, J., Schmitt, L.-M., & Obleser, J. (2020). Temporal selectivity
declines in the aging human auditory cortex. In bioRxiv, 2020.01.
24.919126. DOI: https://doi.org/10.1101/2020.01.24.919126
Feinberg, D. A., Moeller, S., Schmied, S. M., Auerbach, E., Ramanna,
S., Glasser, M. F., … Yacoub, E. (2010). Multiplexed echo planar
imaging for sub-second whole brain FMRI and fast diffusion
Bildgebung. PLOS One, 5, e15710. DOI: https://doi.org/10.1371/journal.
pone.0015710, PMID: 21187930, PMCID: PMC3004955
Gorgolewski, K. J., Varoquaux, G., Rivera, G., Schwarz, Y., Ghosh,
S. S., Maumet, C., … Margulies, D. S. (2015). NeuroVault.org: A
web-based repository for collecting and sharing unthresh-olded
statistical maps of the human brain. Frontiers in Neuroinformatics,
9, 8. DOI: https://doi.org/10.3389/fninf.2015.00008, PMID:
25914639, PMCID: PMC4392315
Gracco, V. L., Tremblay, P., & Pike, B. (2005). Imaging speech
production using fMRI. NeuroImage, 26(1), 294–301. DOI: https://
doi.org/10.1016/j.neuroimage.2005.01.033, PMID: 15862230
Hall, D. A., Haggard, M. P., Akeroyd, M. A., Palmer, A. R.,
Summerfield, A. Q., Elliott, M. R., … Bowtell, R. W. (1999).
“Sparse” temporal sampling in auditory fMRI. Menschliches Gehirn
Mapping, 7(3), 213–223. DOI: https://doi.org/10.1002/(WISSENSCHAFT)
1097-0193(1999)7:3<213::AID-HBM5>3.0.CO;2-N, PMID:
10194620, PMCID: PMC6873323
Harris, K. C., Dubno, J. R., Keren, N. ICH., Ahlstrom, J. B., & Eckert,
M. A. (2009). Speech recognition in younger and older adults: A de-
pendency on low-level auditory cortex. Zeitschrift für Neurowissenschaften,
29, 6078–6087. DOI: https://doi.org/10.1523/JNEUROSCI.0412-
09.2009, PMID: 19439585, PMCID: PMC2717741
Hauk, O., Davis, M. H., & Pulvermüller, F. (2008). Modulation of
brain activity by multiple lexical and word form variables in visual
word recognition: A parametric fMRI study. NeuroImage, 42(3),
1185–1195. DOI: https://doi.org/10.1016/j.neuroimage.2008.
05.054, PMID: 18582580
Hickok, G., & Kacke, D. (2007). The cortical organization of
speech processing. Nature Reviews Neurowissenschaften, 8, 393–402.
DOI: https://doi.org/10.1038/nrn2113, PMID: 17431404
Humes, L. E. (1996). Speech understanding in the elderly. Journal of the
American Academy of Audiology, 7, 161–167. PMID: 8780988
Hwang, J.-H., Li, C.-W., Wu, C.-W., Chen, J.-H., & Liu, T.-C.
(2007). Aging effects on the activation of the auditory cortex during
binaural speech listening in white noise: Eine fMRT-Studie. Audiology
& Neuro-Otology, 12(5), 285–294. DOI: https://doi.org/10.1159/
000103209, PMID: 17536197
Jenkinson, M., Beckmann, C. F., Behrens, T. E. J., Woolrich, M. W.,
& Schmied, S. M. (2012). FSL. NeuroImage, 62(2), 782–790. DOI:
https://doi.org/10.1016/j.neuroimage.2011.09.015, PMID:
21979382
Kronbichler, M., Hutzler, F., Wimmer, H., Mair, A., Staffen, W., &
Ladurner, G. (2004). The visual word form area and the frequency
with which words are encountered: Evidence from a parametric
fMRT-Studie. NeuroImage, 21(3), 946–953. DOI: https://doi.org/
10.1016/j.neuroimage.2003.10.021, PMID: 15006661
Kuchinsky, S. E., Vaden, K. ICH., Jr., Keren, N. ICH., Harris, K. C., Ahlstrom,
J. B., Dubno, J. R., & Eckert, M. A. (2012). Word intelligibility and
age predict visual cortex activity during word listening. Zerebral
Kortex, 22, 1360–1371. DOI: https://doi.org/10.1093/cercor/
bhr211, PMID: 21862447, PMCID: PMC3357178
Kuznetsova, A., Brockhoff, P., & Christensen, R. (2017). lmerTest
Package: Tests in linear mixed effects models. Zeitschrift für
Statistical Software, 82(13), 1–26. DOI: https://doi.org/10.18637/
jss.v082.i13
Lash, A., Rogers, C. S., Zoller, A., & Wingfield, A. (2013).
Expectation and entropy in spoken word recognition: Effects of age
and hearing acuity. Experimental Aging Research, 39, 235–253.
DOI: https://doi.org/10.1080/0361073X.2013.779175, PMID:
23607396, PMCID: PMC3668645
Lee, Y. S., Wingfield, A., Min, N. E., Kotloff, E., Grossman, M., &
Peelle, J. E. (2018). Differences in hearing acuity among “normal-
hearing” young adults modulate the neural basis for speech com-
prehension. eNeuro, 5(3), e0263–17.2018. DOI: https://doi.org/
10.1523/ENEURO.0263-17.2018, PMID: 29911176, PMCID:
PMC6001266
Manan, H. A., Franz, E. A., Yusoff, A. N., & Mukari, S. Z.-M. S.
(2015). The effects of aging on the brain activation pattern during
a speech perception task: Eine fMRT-Studie. Aging Clinical and
Neurobiology of Language
471
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
Experimental Research, 27(1), 27–36. DOI: https://doi.org/
10.1007/s40520-014-0240-0, PMID: 24906677
Manan, H. A., Yusoff, A. N., Franz, E. A., & Mukari, S. Z.-M. S.
(2017). Effects of aging and background babble noise on speech
perception processing: Eine fMRT-Studie. Neurophysiologie, 49(6),
441–452. DOI: https://doi.org/10.1007/s11062-018-9707-5
Morosan, P., Rademacher, J., Schleicher, A., Amunts, K.,
Schormann, T., & Zilles, K. (2001). Human primary auditory cor-
tex: Cytoarchitectonic subdivisions and mapping into a spatial
reference system. NeuroImage, 13, 684–701. DOI: https://doi.
org/10.1006/nimg.2000.0715, PMID: 11305897
Mumford, J. A., Polina, J.-B., & Poldrack, R. A. (2015).
Orthogonalization of regressors in fMRI models. PLOS One, 10,
e0126255. DOI: https://doi.org/10.1371/journal.pone.0126255,
PMID: 25919488, PMCID: PMC4412813
Neta, M., Miezin, F. M., Nelson, S. M., Dubis, J. W., Dosenbach,
N. U. F., Schlaggar, B. L., & Petersen, S. E. (2015). Spatial and
temporal characteristics of error-related activity in the human
Gehirn. Zeitschrift für Neurowissenschaften, 35, 253–266. DOI: https://doi.
org/10.1523/JNEUROSCI.1313-14.2015, PMID: 25568119,
PMCID: PMC4287146
Park, D. C., Lautenschlager, G., Hedden, T., Davidson, N. S.,
Schmied, A. D., & Schmied, P. K. (2002). Models of visuospatial and
verbal memory across the adult life span. Psychologie und Altern,
17(2), 299–320. DOI: https://doi.org/10.1037/0882-7974.
17.2.299, PMID: 12061414
Peelle, J. E. (2018). Listening effort: How the cognitive consequences
of acoustic challenge are reflected in brain and behavior. Ear and
Hearing, 39(2), 204–214. DOI: https://doi.org/10.1097/AUD.
0000000000000494, PMID: 28938250, PMCID: PMC5821557
Peelle, J. E. (2019). Language and aging. In G. ICH. de Zubicaray &
N. Ö. Schiller (Hrsg.), The Oxford handbook of neurolinguistics
(S. 295–316). Oxford: Oxford University Press.
Peelle, J. E., Johnsrude, ICH. S., & Davis, M. H. (2010). Hierarchical
processing for speech in human auditory cortex and beyond.
Grenzen der menschlichen Neurowissenschaften, 4, 51. DOI: https://doi.org/10.
3389/fnhum.2010.00051, PMID: 20661456, PMCID: PMC2907234)
Peelle, J. E., Troiani, V., Grossman, M., & Wingfield, A. (2011). Hearing
loss in older adults affects neural systems supporting speech compre-
hension. Zeitschrift für Neurowissenschaften, 31(35), 12638–12643. DOI:
https://doi.org/10.1523/JNEUROSCI.2559-11.2011, PMID:
21880924, PMCID: PMC3175595
Peelle, J. E., Troiani, V., Wingfield, A., & Grossman, M. (2010).
Neural processing during older adults’ comprehension of spoken
Sätze: Age differences in resource allocation and connec-
tivity. Hirnrinde, 20, 773–782. DOI: https://doi.org/
1 0 . 1 0 9 3 / c e r c o r / b h p 1 4 2, P M I D : 1 9 6 6 6 8 2 9 , P M C I D :
PMC2837088)
Peelle, J. E., & Wingfield, A. (2016). The neural consequences of age-
related hearing loss. Trends in den Neurowissenschaften, 39(7), 486–497.
DOI: https://doi.org/10.1016/j.tins.2016.05.001, PMID:
27262177, PMCID: PMC4930712
Pichora-Fuller, M. K., Kramer, S. E., Eckert, M. A., Edwards, B.,
Hornsby, B. W. Y., Humes, L. E., … Wingfield, A. (2016).
Eriksholm workshop on hearing impairment and cognitive energy:
The framework for understanding effortful listening (FUEL).
Ear and Hearing, 37, 5S–27S. DOI: https://doi.org/10.1097/
AUD.0000000000000306, PMID: 27355766
Poldrack, R. A., Barch, D. M., Mitchell, J. P., Wager, T. D., Wagner,
A. D., Devlin, J. T., … Milham, M. P. (2013). Toward open sharing
of task-based fMRI data: The OpenfMRI project. Grenzen in
Neuroinformatics, 7, 12. DOI: https://doi.org/10.3389/fninf.2013.00012,
PMID: 23847528, PMCID: PMC3703526
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., &
Petersen, S. E. (2012). Spurious but systematic correlations in
functional connectivity MRI networks arise from subject motion.
NeuroImage, 59, 2142–2154. DOI: https://doi.org/10.1016/j.
neuroimage.2011.10.018, PMID: 22019881, PMCID: PMC3254728
Power, J. D., & Petersen, S. E. (2013). Control-related systems in the
human brain. Aktuelle Meinung in der Neurobiologie, 23, 223–228.
DOI: https://doi.org/10.1016/j.conb.2012.12.009, PMID:
23347645, PMCID: PMC3632325
Rogers, C. S. (2016). Semantic priming, not repetition priming, Ist
to blame for false hearing. Psychonomic Bulletin & Rezension,
24, 1194–1204. DOI: https://doi.org/10.3758/s13423-016-
1185-4, PMID: 27844295, PMCID: PMC5429986
Rogers, C. S., Jacoby, L. L., & Sommers, M. S. (2012). Frequent false
hearing by older adults: The role of age differences in metacog-
Nation. Psychologie und Altern, 27, 33–45. DOI: https://doi.org/
10.1037/a0026231, PMID: 22149253, PMCID: PMC3319693
Rorden, C., & Brett, M. (2000). Stereotaxic display of brain lesions.
Behavioural Neurology, 12, 191–2000. DOI: https://doi.org/
10.1155/2000/421719, PMID: 11568431
Shafto, M. A., & Tyler, L. K. (2014). Language in the aging brain:
The network dynamics of decline and preservation. Wissenschaft,
346, 583–587. DOI: https://doi.org/10.1126/science.1254404,
PMID: 25359966
Shannon, R. V., Zeng, F.-G., Kamath, V., Wygonski, J., & Ekelid, M.
(1995). Speech recognition with primarily temporal cues. Wissenschaft,
270, 303–304. DOI: https://doi.org/10.1126/science.270.
5234.303, PMID: 7569981
Siegel, J. S., Power, J. D., Dubis, J. W., Vogel, A. C., Church, J. A.,
Schlaggar, B. L., & Petersen, S. E. (2014). Statistical improvements
in functional magnetic resonance imaging analyses produced
by censoring high-motion data points. Kartierung des menschlichen Gehirns, 35,
1981–1996. DOI: https://doi.org/10.1002/hbm.22307, PMID:
23861343, PMCID: PMC3895106
Signoret, C., Johnsrude, ICH., Classon, E., & Rudner, M. (2018).
Combined effects of form- and meaning-based predictability on
perceived clarity of speech. Journal of Experimental Psychology:
Human Perception and Performance, 44(2), 277–285. DOI:
https://doi.org/10.1037/xhp0000442, PMID: 28557490
Sohoglu, E., Peelle, J. E., Carlyon, R. P., & Davis, M. H. (2012).
Predictive top-down integration of prior knowledge during
speech perception. Zeitschrift für Neurowissenschaften, 32(25), 8443–8453.
DOI: https://doi.org/10.1523/JNEUROSCI.5069-11.2012, PMID:
22723684, PMCID: PMC6620994
Sommers, M. S., & Danielson, S. M. (1999). Inhibitory processes
and spoken word recognition in young and older adults: The inter-
action of lexical competition and semantic context. Psychologie
and Aging, 14, 458–472. DOI: https://doi.org/10.1037/0882-
7974.14.3.458, PMID: 10509700
Tremblay, P., Sato, M., & Deschamps, ICH. (2017). Age differences in the
motor control of speech: An fMRI study of healthy aging. Human
Brain Mapping, 38(5), 2751–2771. DOI: https://doi.org/10.1002/
hbm.23558, PMID: 28263012, PMCID: PMC6866863
Tyler, L. K., Shafto, M. A., Randall, B., Wright, P., Marslen-Wilson,
W. D., & Stamatakis, E. A. (2010). Preserving syntactic processing
across the adult life span: The modulation of the frontotemporal
language system in the context of age-related atrophy. Zerebral
Kortex, 20, 352–364. DOI: https://doi.org/10.1093/cercor/
bhp105, PMID: 19505991, PMCID: PMC2803734
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., … Joliot, M. (2002). Automated ana-
tomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neurobiology of Language
472
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Spoken word recognition in older adults
NeuroImage, 15, 273–289. DOI: https://doi.org/10.1006/
nimg.2001.0978, PMID: 11771995
Vaden, K. ICH., Jr., Kuchinsky, S. E., Ahlstrom, J. B., Teubner-Rhodes,
S. E., Dubno, J. R., & Eckert, M. A. (2016). Cingulo-opercular
function during word recognition in noise for older adults with
hearing loss. Experimental Aging Research, 42, 67–82. DOI:
https://doi.org/10.1080/0361073X.2016.1108784, PMID:
26683042, PMCID: PMC4899824
Vaden, K. ICH., Jr., Kuchinsky, S. E., Cute, S. L., Ahlstrom, J. B., Dubno,
J. R., & Eckert, M. A. (2013). The cingulo-opercular network
provides word-recognition benefit. Zeitschrift für Neurowissenschaften, 33,
18979–18986. DOI: https://doi.org/10.1523/JNEUROSCI.1417-
13.2013, PMID: 24285902, PMCID: PMC3841458
Van Casteren, M., & Davis, M. H. (2007). Match: A program to
assist in matching the conditions of factorial experiments.
Behavioral Research Methods, 39(4), 973–978. DOI: https://
doi.org/10.3758/BF03192992, PMID: 18183914
Wild, C. J., Yusuf, A., Wilson, D. E., Peelle, J. E., Davis, M. H., &
Johnsrude, ICH. S. (2012). Effortful listening: The processing of degraded
speech depends critically on attention. Zeitschrift für Neurowissenschaften, 32
(40), 14010–14021. DOI: https://doi.org/10.1523/JNEUROSCI.
1528-12.2012, PMID: 23035108, PMCID: PMC6704770
Wingfield, A., Aberdeen, J. S., & Stine, E. A. (1991). Word onset
gating and linguistic context in spoken word recognition by
young and elderly adults. Journals of Gerontology, 46(3), 127–129.
DOI: https://doi.org/10.1093/geronj/46.3.P127, PMID: 2030278
Wingfield, A., & Grossman, M. (2006). Language and the aging
Gehirn: Patterns of neural compensation revealed by functional
brain imaging. Journal of Neurophysiology, 96, 2830–2839.
DOI: https://doi.org/10.1152/jn.00628.2006, PMID: 17110737
Wingfield, A., & Lindfield, K. C. (1995). Multiple memory systems
in the processing of speech: Evidence from aging. Experimental
Aging Research, 21(2), 101–121. DOI: https://doi.org/10.1080/
03610739508254272, PMID: 7628506
Wingfield, A., & Stine-Morrow, E. A. L. (2000). Language and
Rede. In F. ICH. M. Craik & T. A. Salthouse (Hrsg.), The handbook
of aging and cognition (2nd ed., S. 359–416). Mahwah, NJ:
Lawrence Erlbaum.
Wong, P. C. M., Jin, J. X., Gunasekera, G. M., Abel, R., Lee, E. R., &
Dhar, S. (2009). Aging and cortical mechanisms of speech
perception in noise. Neuropsychologie, 47(3), 693–703. DOI:
https://doi.org/10.1016/j.neuropsychologia.2008.11.032, PMID:
19124032, PMCID: PMC2649004
Zhuang, J., Randall, B., Stamatakis, E. A., Marslen-Wilson, W. D., &
Tyler, L. K. (2011). The interaction of lexical semantics and
cohort competition in spoken word recognition: Eine fMRT-Studie.
Zeitschrift für kognitive Neurowissenschaften, 23, 3778–3790. DOI: https://
doi.org/10.1162/jocn_a_00046, PMID: 21563885
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
4
4
5
2
1
8
6
7
7
6
9
N
Ö
_
A
_
0
0
0
2
1
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurobiology of Language
473