RESEARCH ARTICLE
Neurophysiological Correlates of Frequency,
Concreteness, and Iconicity in American
Sign Language
Keine offenen Zugänge
Tagebuch
Karen Emmorey1, Kurt Winsler2
, Katherine J. Midgley3
Jonathan Grainger4
, and Phillip J. Holcomb3
1School of Speech, Language and Hearing Sciences, San Diego State University
2Abteilung für Psychologie, Universität von Kalifornien, Davis
3Abteilung für Psychologie, San Diego State University
4Laboratoire de Psychologie Cognitive, Aix-Marseille University, Centre National de la Recherche Scientifique
Schlüsselwörter: American Sign Language, event-related potentials, frequency, concreteness, iconicity,
lexical access
ABSTRAKT
To investigate possible universal and modality-specific factors that influence the
neurophysiological response during lexical processing, we recorded event-related potentials
while a large group of deaf adults (n = 40) viewed 404 signs in American Sign Language (ASL)
that varied in ASL frequency, concreteness, and iconicity. Participants performed a go/no-go
semantic categorization task (does the sign refer to people?) to videoclips of ASL signs (clips
began with the signer’s hands at rest). Linear mixed-effects regression models were fit with
per-participant, per-trial, and per-electrode data, allowing us to identify unique effects of each
lexical variable. We observed an early effect of frequency (greater negativity for less frequent
signs) beginning at 400 ms postvideo onset at anterior sites, which we interpreted as reflecting
form-based lexical processing. This effect was followed by a more widely distributed posterior
response that we interpreted as reflecting lexical-semantic processing. Paralleling spoken
Sprache, more concrete signs elicited greater negativities, beginning 600 ms postvideo onset
with a wide scalp distribution. Endlich, there were no effects of iconicity (except for a weak
effect in the latest epochs; 1,000–1,200 ms), suggesting that iconicity does not modulate
the neural response during sign recognition. Despite the perceptual and sensorimotoric
differences between signed and spoken languages, the overall results indicate very similar
neurophysiological processes underlie lexical access for both signs and words.
EINFÜHRUNG
Current theories in linguistics, Psychologie, and cognitive neuroscience have all been devel-
oped primarily from investigations of spoken languages. This focus has led theories to ignore
or downplay phenomena that are limited in speech but are pervasive in sign languages, solch
as iconicity (a nonarbitrary relation between phonological form and meaning) and observable
linguistic articulators (the vocal articulators for speech are largely hidden from view). By wid-
ening our scientific lens to include sign languages, we can distinguish neurobiological princi-
ples that are universal to human language processing from those that are modulated by the
specific sensorimotor systems within which language is instantiated. To investigate possible
universal factors in language processing, the present study used event-related potentials
Zitat: Emmorey, K., Winsler, K.,
Midgley, K. J., Grainger, J., & Holcomb,
P. J. (2020). Neurophysiological
correlates of frequency, concreteness,
and iconicity in American Sign
Language. Neurobiology of Language,
1(2), 249–267. https://doi.org/10.1162/
nol_a_00012
DOI:
https://doi.org/10.1162/nol_a_00012
zusätzliche Informationen:
https://doi.org/10.1162/nol_a_00012
Erhalten: 07 September 2019
Akzeptiert: 16 April 2020
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Karen Emmorey
kemmorey@sdsu.edu
Handling-Editor:
Mairéad MacSweeney
Urheberrechte ©: © 2020 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) Lizenz.
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
N
Ö
_
A
_
0
0
0
1
2
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurophysiological correlates of sign recognition
(ERPs) to determine the impact of lexical frequency and concreteness on the brain’s response
to a large set of signs (n = ∼400) from American Sign Language (ASL) in a large group of deaf
signers (n = 40). The frequency and semantic properties of lexical forms are likely to be repre-
sented and processed similarly for signed and spoken languages, although the time-course and
scalp distribution of these effects could differ due to differences between the visual-manual
and auditory-vocal modalities. Zusätzlich, we investigated whether iconicity, a phenomenon
influenced by the modality of sign language, affects the time course or amplitude of neural
responses when signers comprehend ASL signs.
One challenge to investigating the effects of frequency on language processing is that cur-
rently there are no ASL corpora available from which frequency counts can be obtained.
Psycholinguistic research has thus relied on sign familiarity ratings by deaf signers to estimate
lexical frequency (z.B., Carreiras, Guitiérrez-Sigut, Baquero, & Corina, 2008; Emmorey, 1991;
Emmorey, Petrich, & Gollan, 2013). Kürzlich, a database of ∼1,000 ASL signs (ASL-LEX) War
created that contains frequency ratings from 25 Zu 30 deaf signers per sign (Caselli, Sevcikova
Sehyr, Cohen-Goldberg, & Emmorey, 2017; Sevcikova Sehyr & Emmorey, 2019). For this da-
tabase, signers rated how often they felt a sign appears in everyday conversation on a scale of
1 (very infrequently) Zu 7 (very frequently). The sign videos for the present study were selected
from this database. For spoken language, familiarity ratings are highly correlated with corpora-
based frequency counts (Gilhooly & Logie, 1980) and are consistent across different groups of
Teilnehmer (Balota, Pilotti, & Cortese, 2001). For sign languages, Fenlon, Schembri, Rentelis,
Vinson, and Cormier (2014) found that subjective frequency ratings of British Sign Language
(BSL) from Vinson, Cormier, Denmark, Schembri, and Vigliocco (2008) were positively corre-
lated with objective frequency counts from the BSL Corpus, although the sample size for this
analysis was much smaller than for spoken languages.
Parallel to spoken languages, faster lexical decision times are reported for signs that are
rated as very frequent than for signs rated as infrequent (z.B., Carrieras et al., 2008; Caselli,
2015; Emmorey, 2002). Weiter, high-frequency signs are retrieved faster than low-frequency
signs in picture-naming tasks (Baus & Costa, 2015; Emmorey, Petrich, & Gollan, 2012;
Emmorey et al., 2013). Higher frequency signs are also acquired earlier by deaf children
(Caselli & Pyers, 2017), and later acquired signs tend to be lower frequency (Vinson et al.
2008). Zusätzlich, high-frequency signs, like high-frequency words, tend to be shorter in du-
ration (z.B., Börstell, Hörberg, & Östling, 2016) and are more likely to undergo coarticulation
processes, such as sign lowering (z.B., Russell, Wilkinson, & Janzen, 2011). To date, ähnlich
linguistic and behavioral effects of lexical frequency have been found for signed and spoken
languages.
In der vorliegenden Studie, we utilized ERPs to investigate the impact of lexical frequency on
sign comprehension. One limitation of using reaction times (RTs) to assess linguistic factors
that affect lexical processing is that RTs reflect the final outcome of lexical access, einschließlich
decision processes (z.B., Grainger & Jacobs, 1996). Im Gegensatz, ERPs continuously reflect
information processing in real time, providing insight into the temporal neural dynamics
of phonological (bilden) Verarbeitung, lexical access, and sign comprehension. No study to
our knowledge has examined how lexical frequency impacts the neural response in sign
comprehension; Jedoch, Baus and Costa (2015) investigated frequency effects in a sign-
production ERP study in which hearing bilinguals fluent in spoken Spanish or Catalan
and Catalan Sign Language (LSC) named pictures in either Spanish/Catalan or LSC. Der
authors reported that high-frequency signs elicited more negative amplitudes than low-
frequency signs in a 280–350-ms time window over occipital sites. This pattern parallels
Neurobiology of Language
250
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
N
Ö
_
A
_
0
0
0
1
2
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurophysiological correlates of sign recognition
the frequency effect for spoken word production, although the difference between high- Und
low-frequency words may emerge earlier for speech (z.B., Strijkers, Costa, & Thierry, 2009).
Im Gegensatz, for visual and auditory word comprehension, low-frequency words tend to elicit
more negative amplitudes than high-frequency words (z.B., Dufau, Grainger, Midgley, &
Holcomb, 2015; Dufour, Brunellière, & Frauenfelder, 2013; Kutas & Federmeier, 2011;
Winsler, Midgley, Grainger, & Holcomb, 2018). Winsler et al. (2018) conducted a large
megastudy of spoken word recognition (∼1,000 words; 50 Teilnehmer) using both a lexical
decision task and a semantic categorization task similar to the one used in the present study.
For the semantic decision task (detect an occasional animal word), Winsler et al. (2018)
reported greater ERP negativities for low-frequency words at frontal and central sites, begin-
ning 500 ms after word onset, which persisted into the final analyzed epoch, 800–900 ms.
Hier, we investigated whether the effects of sign frequency are parallel to the effects of word
frequency with respect to the polarity, scalp distribution, and timing of ERPs to visual-
manual signs.
We note that auditory word recognition is more parallel to sign recognition than visual
word recognition because both speech and sign unfold over time and written words are a sec-
ondary code derived from speech and acquired later through instruction. An early study by
Kutas, Neville, and Holcomb (1987) compared ERP responses to semantic anomalies in writ-
zehn, auditory, and signed sentences and found a strong similarity in the N400 component
across modalities (greater negativity for anomalous than expected lexical items; see also
Capek et al., 2009), but there were also differences, with a more prolonged ERP response
for both auditory words and signs compared to written words. Grosvald, Gutierrez, Hafer,
and Corina (2012) found that pseudosigns elicited a larger sentence-final N400 response com-
pared to semantically appropriate signs, while nonlinguistic grooming gestures (z.B., scratch-
ing one’s nose) elicited a large positivity. This result highlights the linguistic specificity of the
N400 component for signs. Weiter, Meade, Lee, Midgley, Holcomb, and Emmorey (2018)
reported both semantic and phonological priming effects in the N400 window for single signs.
Zusammen, these results indicate that the N400 elicited by signs is sensitive to both phonological
structure and lexical semantics. Based on these findings, we predict that lexical frequency will
modulate ERPs in the N400 window, with low-frequency signs eliciting greater negativity than
high-frequency signs, as found for spoken languages.
In addition to lexical frequency, the parallel megastudy by Winsler et al. (2018) examined
the effect of relative concreteness on ERPs during auditory word recognition. Concrete spo-
ken words elicited larger negativities than abstract words, with robust effects emerging after
400 ms that were widely distributed around central sites. Greater negativity for concrete than
abstract words within the N400 window has been interpreted as reflecting richer semantic
representations for concrete words that arise from associations with imagistic and sensorimo-
tor representations and from larger semantic networks (z.B., Holcomb, Kounios, Anderson, &
Westen, 1999; Kutas & Federmeier, 2011). Behaviorally, concrete words are typically recog-
nized faster than abstract words (z.B., Kroll & Merves, 1986), possibly due to their semantic
richness. For sign language, Emmorey and Corina (1993) found that concrete ASL signs were
recognized faster than abstract signs in a lexical decision task. No study to our knowledge
has investigated the effect of concreteness on ERPs during sign recognition. Jedoch, gegeben
the behavioral concreteness effects found by Emmorey and Corina (1993) and the sensitivity
of the N400 to semantic manipulations in sign language (Capek et al., 2009; Kutas et al.,
1987; Meade et al., 2018; Neville et al., 1997), we anticipate that concreteness effects within
the N400 window will pattern like spoken language, with greater negativity associated with
more concrete signs.
Neurobiology of Language
251
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
N
Ö
_
A
_
0
0
0
1
2
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurophysiological correlates of sign recognition
In addition to lexical frequency and concreteness, we examined the effect of iconicity on
ERPs to signs. Iconicity values were obtained from deaf signers who rated the iconicity of the
signs in the ASL-LEX database (Caselli et al., 2017; Sevcikova Sehyr & Emmorey, 2019). Der
number of deaf participants rating each sign varied between 26 Und 31. Parallel to the sub-
jective frequency ratings, participants were asked to rate each sign video on a 7-point scale
based on how much the sign looks like what it means (1 = not iconic at all, 7 = very iconic).
Several behavioral studies have used this type of rating to investigate the effect of iconicity on
sign comprehension and production. In picture-naming tasks, several studies have now found
that highly iconic signs are retrieved faster than noniconic signs (Baus & Costa, 2015;
McGarry, Mott, Midgley, Holcomb, & Emmorey, 2018; Navarrete, Peressotti, Lerose, &
Miozzo, 2017; Vinson, Thompson, Skinner, & Vigliocco, 2015). Jedoch, for comprehension
the effects of iconicity have been mixed. Bosworth and Emmorey (2010) found that iconic
signs were not recognized more quickly than noniconic signs in a lexical decision task. In einem
translation task, Baus, Carreiras, and Emmorey (2013) found that for proficient signers, iconic
signs were actually recognized more slowly than noniconic signs. In a picture-sign matching
Aufgabe, Thompson, Vinson, and Vigliocco (2009) and Vinson et al. (2015) reported faster deci-
sion times when the iconic properties of the sign were aligned with visual features in the pic-
tur (z.B., the ASL sign BIRD depicts a bird’s beak and matches a picture of a bird with a
prominent beak) compared to nonaligned pictures (z.B., a bird in flight where the beak is
not visible). Thompson, Vinson, and Vigliocco (2010) found that form decisions about hand-
shape (straight or curved fingers) were slower for more iconic signs, while Vinson et al. (2015)
found that decisions about movement direction (up or down) were faster for more iconic signs.
To date, the data suggest that iconicity does not have a clear, consistent impact on sign
recognition.
To our knowledge, the only ERP study to explicitly manipulate iconicity in a sign compre-
hension task with deaf signers is Mott, Midgley, Holcomb, and Emmorey (2020). Mott et al.
used ERPs and a translation priming paradigm (English word prime–ASL sign target) to inves-
tigate the effects of iconicity on sign recognition in proficient deaf signers and hearing L2
learners. Participants decided whether word-sign pairs were translation equivalents or not.
For hearing learners, iconic signs elicited earlier and more robust priming effects (d.h., greater
negativities for target signs preceded by unrelated word primes than by translation primes)
compared to noniconic signs. Im Gegensatz, for deaf signers, iconicity did not modulate transla-
tion priming effects either in RTs or in the ERPs within the N400 window. The fact that priming
effects did not begin earlier for iconic than noniconic signs suggests that iconicity does not
facilitate lexical access for deaf signers, in contrast to L2 learners. Here we explore whether
iconicity modulates ERPs to signs in a comprehension paradigm that does not involve priming
or a translation task.
In sum, the purpose of the present study was to use ERPs to investigate how lexical frequency,
concreteness, and iconicity affect the temporal neural dynamics of sign recognition. Following
the “megastudies” of auditory and visual word recognition by Winsler et al. (2018) and Dufau
et al. (2015), we gathered data from a large number of items and participants and treated these
lexical variables as continuous measures, rather than categorizing and factorially manipulating
ihnen. This method avoids potential experimenter bias in selecting cut-off boundaries when cat-
egorizing continuous variables and allows for statistical analyses that control for the effects of
other variables, such that results can clearly be attributed to the variable of interest (see Balota,
Yap, Hutchison, & Cortese, 2012, for a discussion of the advantages of this type of “megastudy”).
Following Winsler et al. (2018) and Emmorey, Midgley, Kohen, Sevcikova Sehyr, and Holcomb
(2017), we used linear mixed-effects regression (LMER) Techniken, rather than more traditional
Neurobiology of Language
252
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
N
Ö
_
A
_
0
0
0
1
2
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurophysiological correlates of sign recognition
Analysen, which allowed us to use single trial EEG data, rather than averaged ERP data. Wir
also used linear mixed-effects (LME) models to visualize the ERP effects by computing an LME
equivalent to scalp voltage maps using the t statistics at each electrode (see Data Analysis).
MATERIALS AND METHODS
Teilnehmer
Forty deaf ASL signers participated in this study (22 females; Durchschnittsalter = 28.9 Jahre; SD = 7.2
Jahre; range = 19–46 years). Thirty-one participants were native signers who were born into a
deaf signing family, eight participants had hearing parents and were exposed to ASL before
three years of age, and one participant learned ASL at age 12 Jahre. Four participants were
left-handed. Most participants were from San Diego or Riverside, Kalifornien, and were com-
pensated $15 per hour of participation. An additional eight participants were run but were not
included in the analyses due to high-artifact rejection rates, very noisy EEG data, or failure to
perform the task. Informed consent was obtained from all participants in accordance with the
institutional review board at San Diego State University.
Materials
The critical stimuli were 404 ASL sign videos from the ASL-LEX database (Caselli et al., 2017).
An additional 26 probe sign videos (also from ASL-LEX) that referred to people were also pre-
gesendet (z.B., MAN, NURSE, MOTHER). Each sign occurred twice for a total of 52 probe signs.
The critical stimuli varied in lexical class: nouns = 50%, verbs = 25%, adjectives = 16%, Anzeige-
verbs = 2%, and other (“minor” closed class signs) = 7%. All stimuli can be viewed on the ASL-
LEX website (http://asl-lex.org). The Entry IDs (English glosses) for the signs are provided in the
zusätzliche Informationen.
For frequency measures, we used the subjective frequency ratings from ASL-LEX, welche
used a scale of 1 (very infrequent) Zu 7 (very frequent). For the sample of critical signs, frequency
ratings ranged from 1.63 Zu 6.84, with a mean of 4.50 (SD = 1.07).
Because no database with concreteness ratings is available for ASL signs, we used ratings
from Brysbaert, Warriner, and Kuperman (2014) based on the English translations of the ASL
signs. Jedoch, there were 13 signs that did not have translation equivalents in Brysbaert et al.
(z.B., STARBUCKS, MCDONALDS, EUROPE), and therefore we gathered additional concrete-
ness ratings for these words from 39 students at San Diego State University, using the same
5-point scale as Brysbaert et al. and mixing these 13 words in with 37 other words that
varied in concreteness. Concreteness ratings for the English translation equivalents of the
ASL signs ranged from 1.22 Zu 5.0, with a mean of 3.42 (SD = 1.60).
Endlich, iconicity ratings were collected from deaf ASL signers (Sevcikova Sehyr &
Emmorey, 2019) on a scale of 1 (not iconic) Zu 7 (very iconic), as part of ASL-LEX 2.0 (Die
ratings will be publicly available on the website in 2020). Iconicity ratings ranged from 1.0
Zu 7.0, with a mean of 3.03 (SD = 1.60).
The mean length of the videos was 1,770 MS (SD = 260 MS; range = 934–2,903 ms). Der
mean onset of the sign was 497 ms after the start of the video (SD = 122 MS; range = 200–
1,168 MS). Sign onset is typically defined as when the hand(S) makes contact with the target
location on the body (see Caselli et al., 2017, for details on how sign onset is determined). Sign
offset is typically defined as the last video frame when the hand contacts the body before mov-
ing back to a resting position (see Caselli et al., 2017). The mean sign length was 506 MS (SD =
Neurobiology of Language
253
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
N
Ö
_
A
_
0
0
0
1
2
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurophysiological correlates of sign recognition
161 MS; range = 134–1,301 ms). We note that sign onset and length were related at least in
part to this particular model’s sign production (rather than inherent to the signs themselves).
Daher, they were not analyzed as experimental variables. Jedoch, given the variability of tim-
ing in the videos, sign length and sign onset were used as covariates in all analyses to control
for some of the possible differences in EEG signal due to timing differences in the videos.
Verfahren
Participants were seated in a comfortable chair, 150 cm from a 24-inch LCD stimulus monitor
in a sound-attenuating darkened room while engaging in a go/no-go semantic categorization
Aufgabe. The testing session began with a short practice block of 15 Versuche, followed by two exper-
imental blocks for 259 trials each. On each trial an ASL sign was presented as a video clip that
was centered on the LCD monitor. Trials were of varying duration depending on the length of
the individual video clips. Regardless of clip duration, a fixed blank-screen inter-stimulus-
interval of 620 ms was interspersed between the offset of one clip and the onset of the next
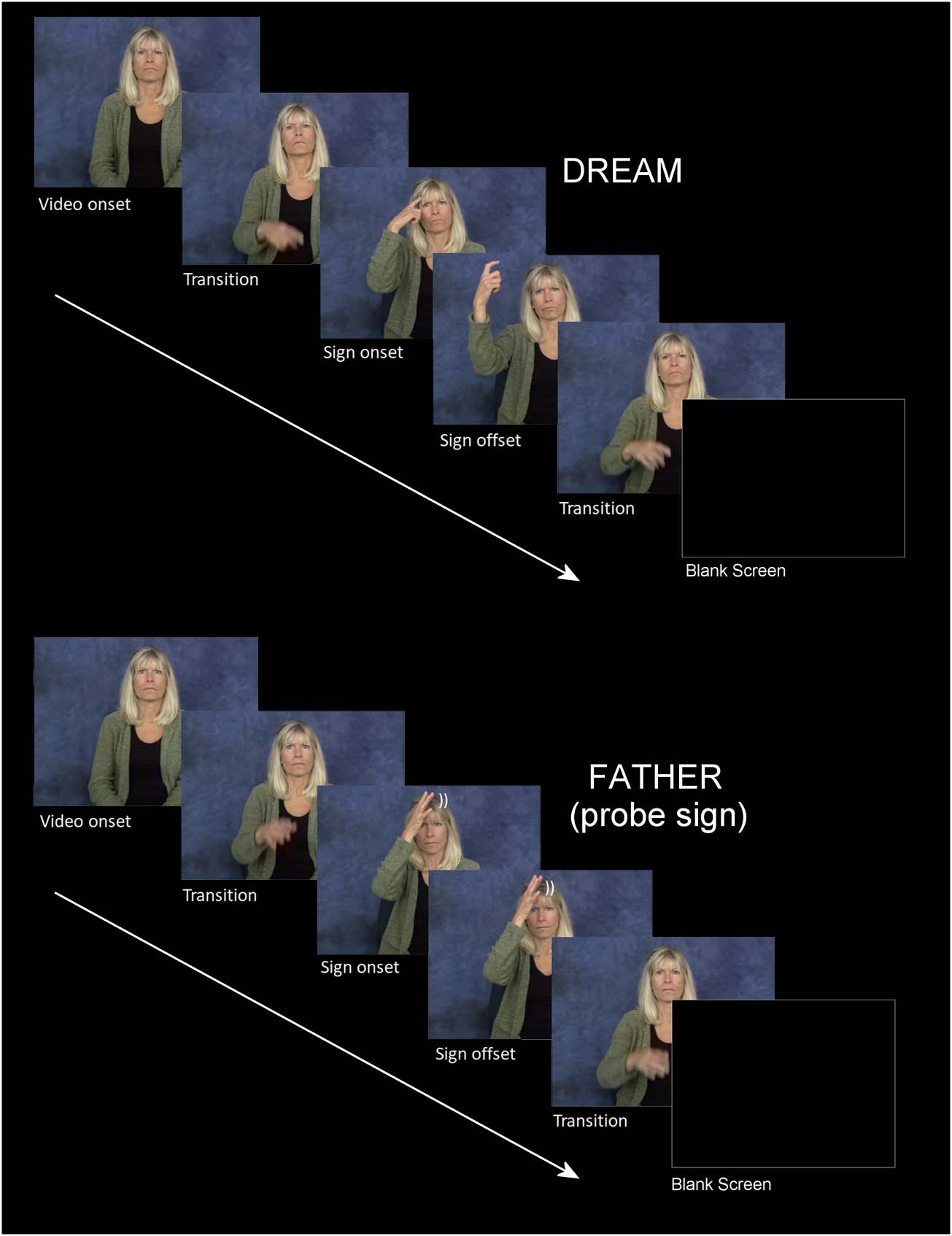
(siehe Abbildung 1 for a schematic of the paradigm). Each experimental block contained 202 critical
target signs and 26 randomly intermixed probe signs (so-called people signs [z.B., BOY,
NURSE]—12% of trials). Participants were instructed to press a button resting in their lap when-
ever they detected a people sign (d.h., a “go” stimulus) and to passively view all other “no-go”
signs. On average every 12 trials a visual “blink” stimulus was presented for 2.5 S. This indicated
that the participant could blink/rest their eyes, thus reducing the tendency for participants to
blink during the critical sign ERP epochs.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
N
Ö
_
A
_
0
0
0
1
2
P
D
.
/
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
EEG Recording
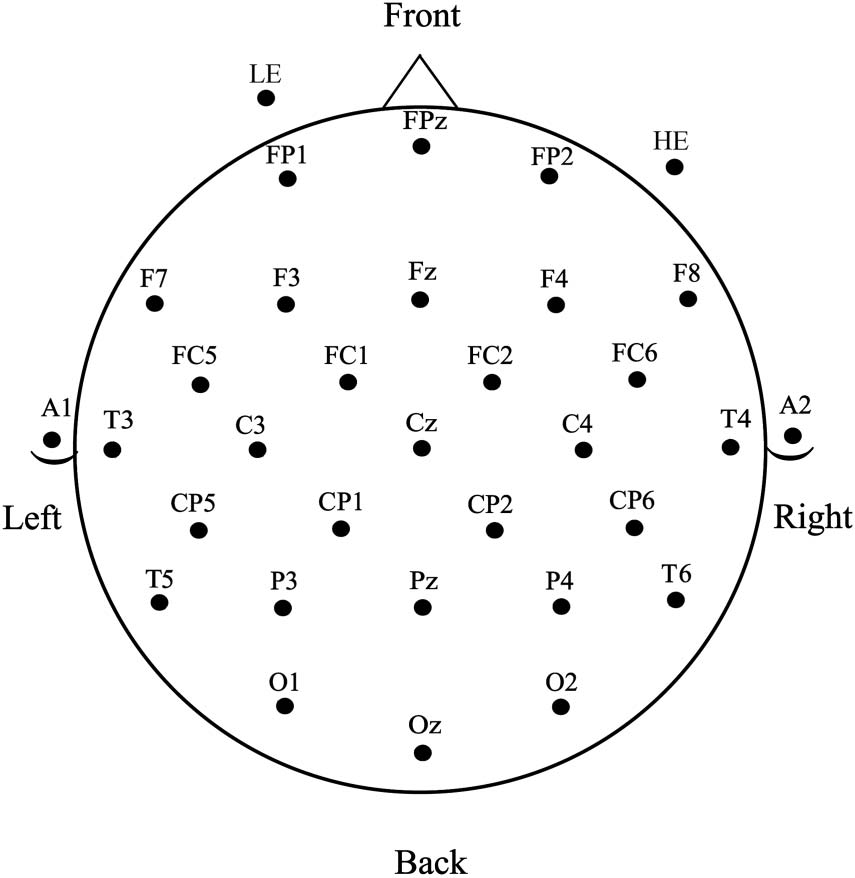
The EEG was collected using a 29-channel electrode cap containing tin electrodes (Electro-Cap
International, Inc., Eaton, OH), arranged in the International 10–20 system (siehe Abbildung 2).
Electrodes were also placed next to the right eye to monitor horizontal eye movements (HE)
and below the left eye (LE) to monitor vertical eye movements and blinks. Endlich, two elec-
trodes were placed behind the ears over the mastoid bones. The left mastoid site was used
as an online reference for the other electrodes, and the right mastoid site was used to evaluate
differential mastoid activity. Impedances were kept below 2.5 kΩ for all scalp and mastoid elec-
trode sites and below 5 kΩ for the two eye channels. The EEG signal was amplified by
SynAmpsRT amplifier (Neuroscan-Compumedics, Charlotte, NC) with a bandpass of DC to
200 Hz and was continuously sampled at 500 Hz. Prior to data analysis the raw EEG data were
corrected for blink and horizontal eye artifact using ICA (EEGLAB, Jung et al., 2000). Single-trial
ERPs were formed from artifact-free trials, beginnend 100 ms prior to the onset of each ASL sign
video and continuing for 1,200 MS. Der 100 ms pre-sign-onset period was used as a baseline.
Data Analysis
The data were analyzed using LMER, a relatively new approach to analyzing EEG data. LMER
modeling is particularly advantageous for designs such as the current one, where there are
multiple, potentially collinear, continuous variables. The correlation matrix for the variables
included in our analysis is given in the Supporting Information (Appendix A1). Weiter, LMER
allows for the model to simultaneously include random effects for both participant and item
(see Baayen, Davidson, & Bates, 2008; Barr, Erheben, Scheepers, & Tily, 2013). The models
described below were fit using the lme4 package (Bates, Mächler, Bolker, & Walker, 2015)
in R (R Core Team, 2014), and were structured based on models of Winsler et al. (2018).
Neurobiology of Language
254
Neurophysiological correlates of sign recognition
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
N
Ö
/
l
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
N
Ö
_
A
_
0
0
0
1
2
P
D
/
.
l
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 1. A schematic for two typical trials. Top is a critical sign trial and bottom is a “probe” trial
(people sign) requiring a button press. Note that in this figure the images are still frames extracted
from actual sign videos shown to participants.
EEG data were measured per participant, per item, and per electrode as average voltage
over 100-ms epochs, starting in a 100–200-ms epoch, and continuing through an 1,100–
1,200-ms epoch. Identical models were fit to predict mean amplitude for each of the 11 Zeit
windows. These models contained main effects for the three experimental variables,
Frequency, Concreteness, and Iconicity, as well as for the two covariates for Sign Length
and Sign Onset. Each of these five variables were standardized prior to analysis.
Interactions between experimental variables were not included in the model. Although it is
likely that these variables interact in ways that are detectable in the EEG signal, a full analysis
of interactions would greatly increase the complexity of the models and is outside the scope of
the present article. The questions of interest here relate to probing the broad patterns of effects
related to sign-level variables. Fewer exploratory experiments will be necessary to adequately
answer questions about how these variables interact with each other.
Neurobiology of Language
255
Neurophysiological correlates of sign recognition
Figur 2. Electrode montage used for EEG recordings.
To analyze the distribution of the effects in addition to their overall effects, all electrodes
were included in the models separately, each with three distributional variables corresponding
to the spatial location of the electrode. These dimensions (X-position, Y-position, and Z-position)
were included as interaction terms with each of the experimental variables and the covariates, als
well as included as covariates themselves. Daher, the models had four parameters for each
Variable, an overall effect across all electrode sites, one for how the effect differs from left to right
(X-position), one for how the effect differs between anterior and posterior sites ( Y-position), Und
one for how the effect differs across electrodes lower on the scalp (z.B., T3, Oz) versus higher,
central sites (z.B., Cz). See the Supporting Information (Appendix A2) for model code. Das war
the strategy used by Winsler et al. (2018) and was shown to appropriately analyze broad patterns
of ERP distributions. Given the exploratory nature of the present experiment and the low spatial
resolution of EEG signals, this approach was adopted to identify the general pattern of effects,
without strong a priori hypotheses. But it should be noted that this strategy has limited power to
detect especially focal or nonlinear interactions between effects and their distributions over the
scalp.
The random-effect structure included random intercepts for participant, item, and elec-
trode. Zusätzlich, there were by-participant random slopes for the effect of each experimen-
tal variable (Frequency, Concreteness, and Iconicity), as well as Sign Length and Sign Onset.
To assess significance of each effect, confidence intervals were generated for each param-
eter. Zusätzlich, p values were obtained for each parameter using type-two Wald tests,
which allowed us to test the partial effect (unique variance) of each variable of interest.
These p values were FDR (false discovery rate) corrected using the Mass Univariate
Analysis Toolbox (Groppe, Urbach & Kutas, 2011). Effects were only interpreted as significant
if they were significant by confidence interval (interval not containing 0) and by FDR-corrected
p value ( P < 0.05).
Data visualization
The confidence interval and t statistic for each parameter of interest are presented for each
time window in Figures 3A–5A. The effect is highlighted if it was significant with both the
Neurobiology of Language
256
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurophysiological correlates of sign recognition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
/
.
l
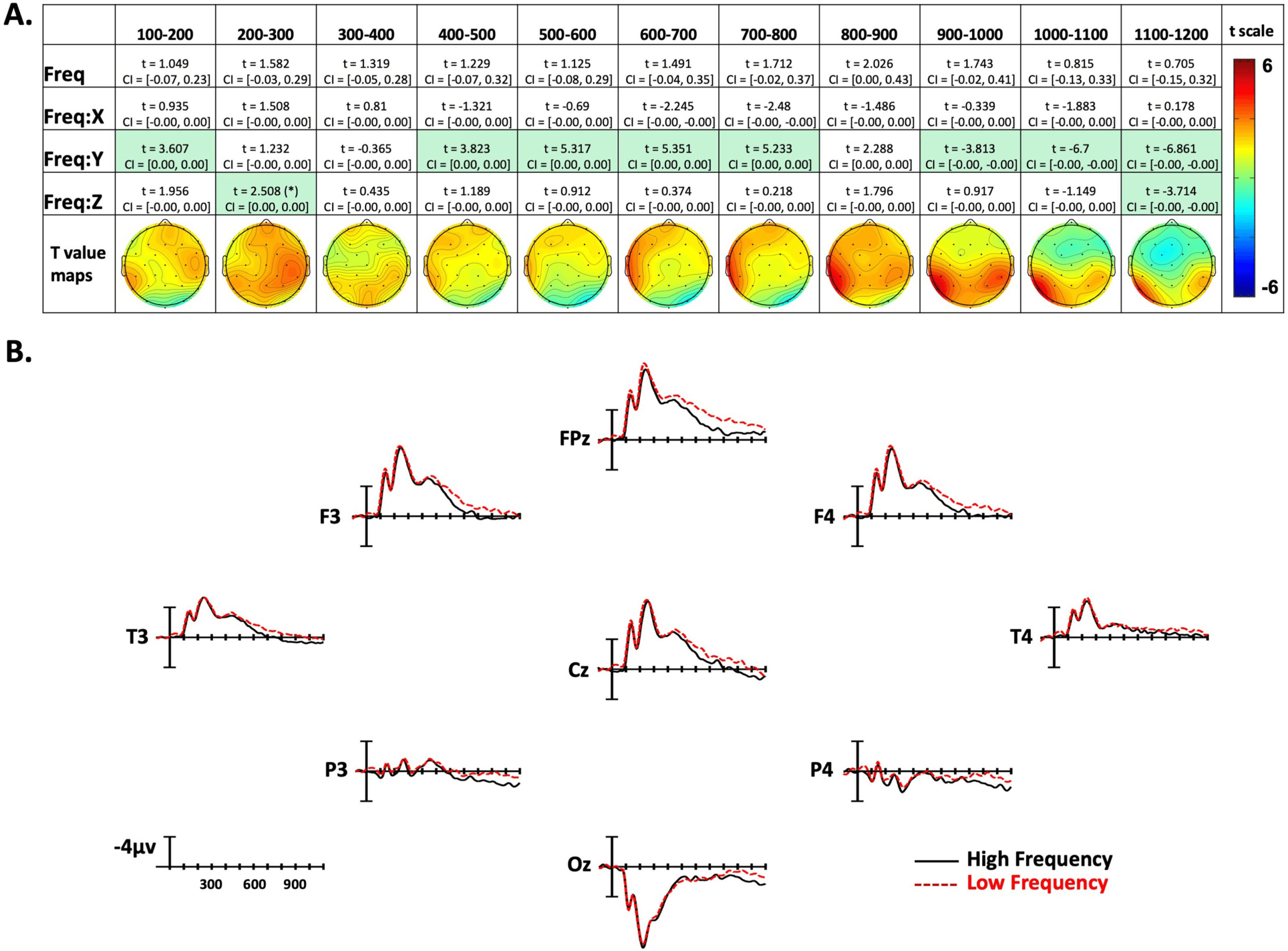
Figure 3. American Sign Language frequency effects. (A) linear mixed-effect t statistics, confidence intervals, and topographical t-statistic
maps for frequency effects. Effects are only highlighted if results were significant with both confidence intervals and false discovery rate
corrected p values; trend ( p < .06) indicated by (*). (B) Frequency ERP plots made using the top and bottom quartiles of items sorted by
frequency.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
confidence interval and the FDR-corrected p value. To visualize the distribution of the ef-
fects, models were constructed for each time epoch and electrode separately, and t values
for the effect of each variable of interest were plotted across the scalp as topographic maps
(maps in Figures 3A–5A). These models included the overall effects of Frequency,
Concreteness, and Iconicity, as well as Sign Length and Sign Onset as covariates. The electrode-
specific models also contained random intercepts for participants and items. Additionally, for
visualization, traditional ERPs were plotted by averaging EEG data from 50 representative signs
for the high and low conditions of each of the three experimental variables (see Figures 3B–
5B). These averages controlled for the other experimental variables such that each comparison
differed significantly only by the variable of interest, but not by the other two experimental
variables. Note that these ERPs are for visual reference only and have not been analyzed
statistically.
Neurobiology of Language
257
Neurophysiological correlates of sign recognition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
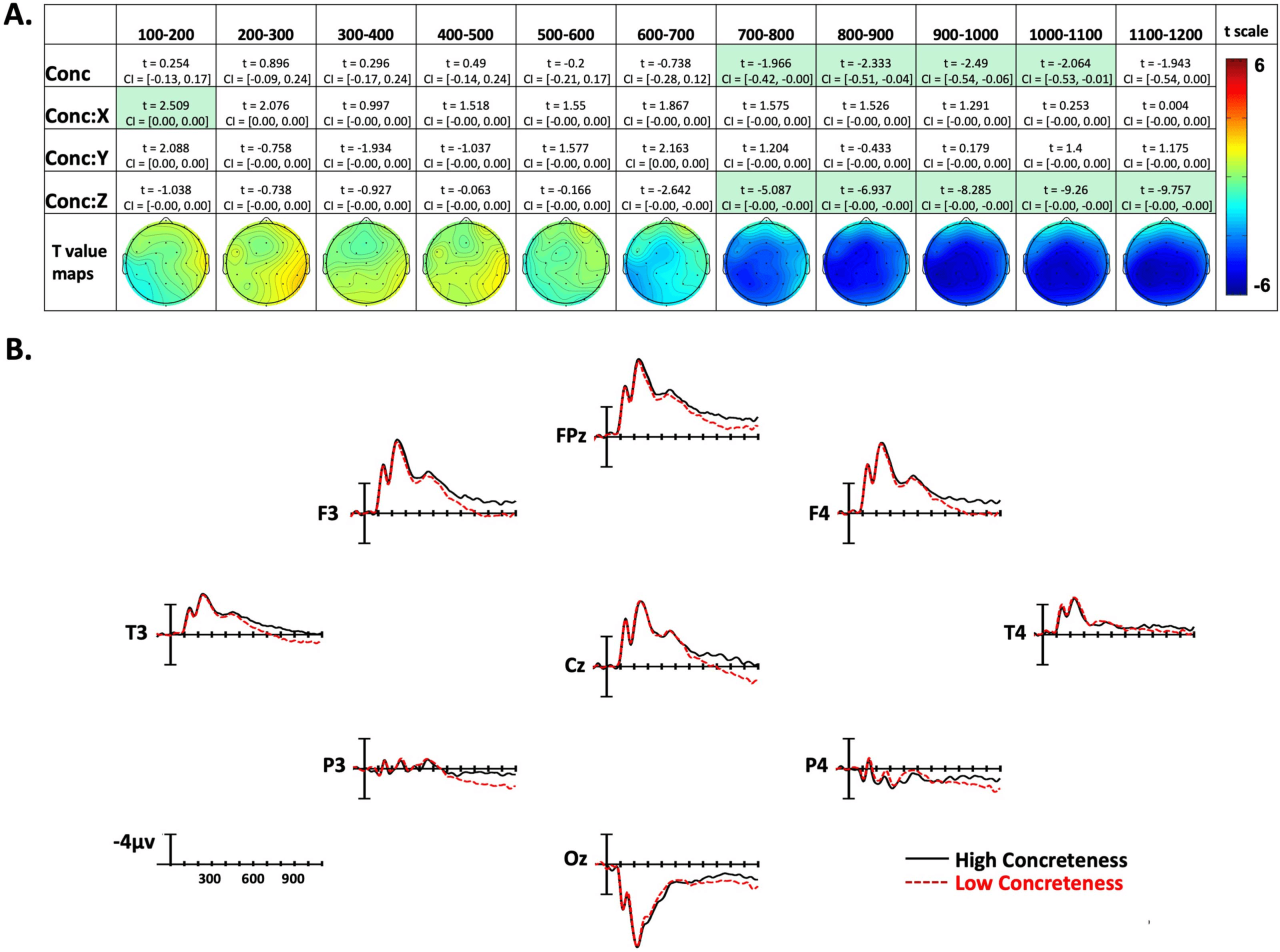
Figure 4. Concreteness effects. (A) Linear mixed-effect t statistics, confidence intervals, and topographical t-statistic maps for concreteness
effects. Effects are only highlighted if results were significant with both confidence intervals and false discovery rate corrected p values. (B)
Concreteness ERP plots made using the top and bottom quartiles of items sorted by frequency.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
RESULTS
Behavioral Results
Participants correctly detected an average of 87% of the probe signs that referred to people,
with an average false alarm rate of 4%. The mean RT for detecting the probe signs was 1,392 ms
(SD = 82 ms).
Linear Mixed-Effect Regression Results
Confidence intervals and t statistics for each parameter estimate are presented in a table for
each variable of interest (Frequency, Concreteness, and Iconicity) in Figures 3A–5A. Table
cells are highlighted if the effect is statistically significant both by its confidence interval,
Neurobiology of Language
258
Neurophysiological correlates of sign recognition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
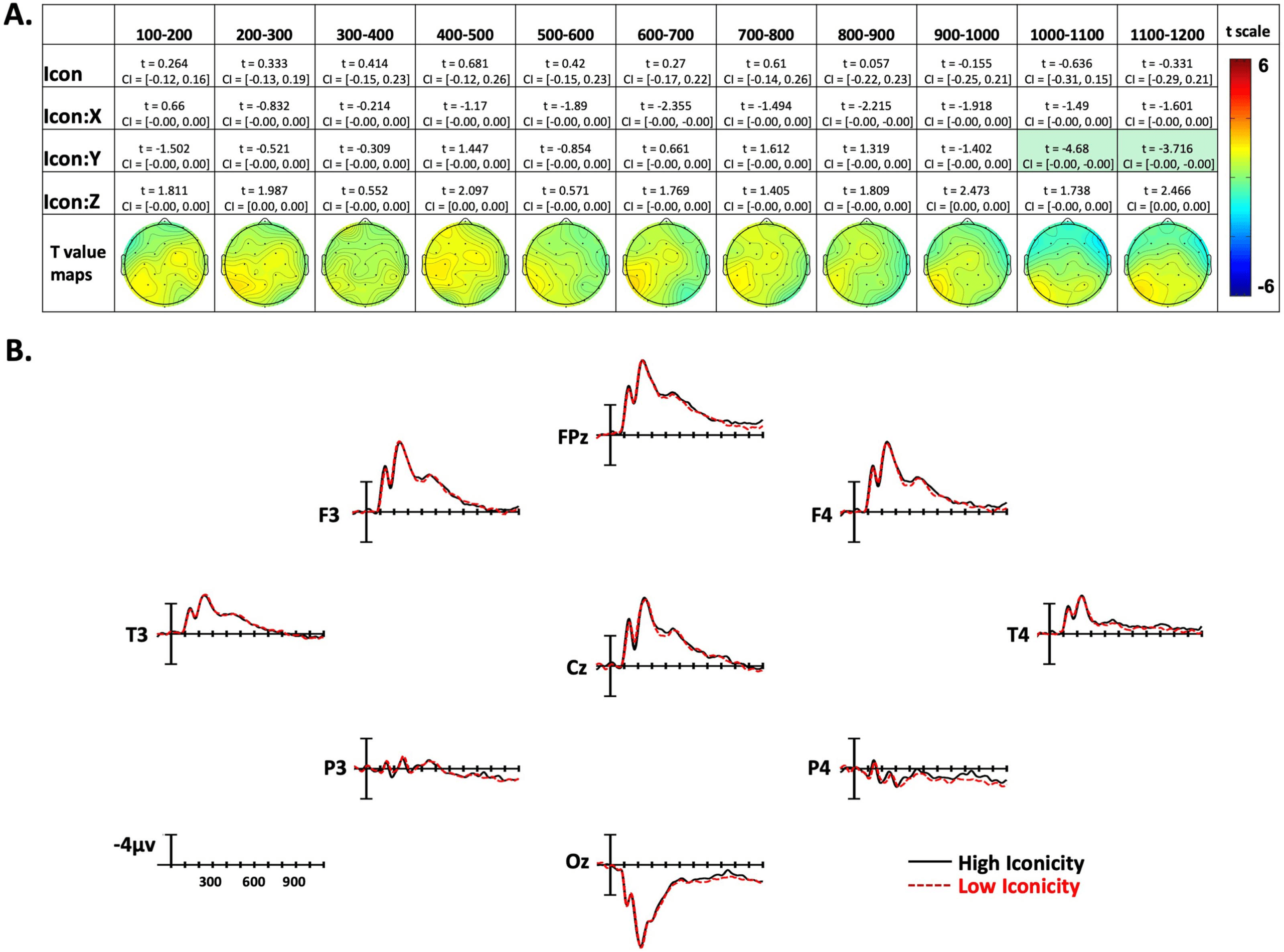
Iconicity effects. (A) Linear mixed-effect t statistics, confidence intervals, and topographical t-statistic maps for iconicity effects.
Figure 5.
Effects are only highlighted if results were significant with both confidence intervals and false discovery rate corrected p values. (B)
Iconicity ERP plots made using the top and bottom quartiles of items sorted by frequency.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
and FDR-corrected p value. To aid the visualization of the effects, for each time point there is a
topographical map made from t values obtained from per-electrode LMER models.
Additionally, Figures 3B–5B present averaged ERPs comparing each variable with 50 items
per average, balancing for the other two variables.
Frequency Effects
In the first epoch from 100–200 ms, there was a Frequency by Y-position interaction. As
shown in Figure 3A, this indicates that lower frequency signs tended to produce more nega-
tivity in frontal sites, and less negativity in posterior sites. In the following two time windows,
there were no significant effects. For the next four epochs, between 400 and 800 ms, there was
again an interaction with the Y dimension, with low-frequency signs generating greater
Neurobiology of Language
259
Neurophysiological correlates of sign recognition
negativity in anterior electrode sites (see Figure 3A). In the following 800–900-ms epoch there
were no significant effects. Based on the topographic maps in Figure 3A, the frequency effect
seems to be transitioning from the previous anterior distribution, to a more posterior distribu-
tion in the following epochs. In the 900-1,000-ms time window there was again a Frequency
by Y-position interaction, but now in the opposite direction as previously (note the flipped t
statistic), indicating lower frequency items elicited more negativity in posterior sites. This effect
remained through the final epoch (1,100–1,200 ms). Additionally, in the final epoch there was
a Frequency by Z-position interaction, showing more negativity to low-frequency signs in pe-
ripheral sites, and a slight positivity to low-frequency signs in central sites.
Concreteness Effects
In the 100–200-ms epoch there was a Concreteness by X-position interaction, with more con-
crete signs producing greater negativity on the left side of the montage, and less negativity on
the right side. However, for the next four epochs (200–600 ms), there were no significant ef-
fects of concreteness. Beginning in the 600–800-ms epoch and continuing through the final
epoch (1,100–1,200 ms), there was a Concreteness by Z-position interaction. This interaction
indicates that concrete items elicited more negativity than abstract items, and this effect was
distributed in the center of the scalp (see Figure 4A and Figure 4B). Additionally, in the four
time windows between 700 and 1,100 ms, there was an overall effect of concreteness, show-
ing that in these epochs the concreteness effect is distributed across the entire scalp.
Iconicity Effects
In the first nine time windows analyzed, there were no effects of Iconicity. In the final two
epochs (1,000–1,100 ms and 1,100–1,200 ms) there were Iconicity by Y-position interactions
showing greater negativity to low-iconicity signs in posterior sites, and less negativity in ante-
rior sites (see Figure 5A and Figure 5B).
DISCUSSION
This is the first ERP study to investigate the effects of lexical frequency, concreteness, and
iconicity on the temporal neural dynamics of sign comprehension. LMER models were fit in
100 ms-time epochs with per-participant, per-trial, per-electrode data to analyze the electro-
physiological effects of these lexical variables on sign recognition. The results revealed both
universal properties of lexical processing that are shared across signed and spoken languages,
as well as different patterns that may be attributable to characteristics of the auditory-oral and
visual-manual modalities. As predicted, lexical frequency and concreteness exhibited similar
electrophysiological effects for sign recognition as previously found for spoken word recogni-
tion, but the time-course and scalp distribution of these effects were somewhat different for
signs. No significant effects of iconicity were found, except for a weak effect in the late epochs.
Frequency
ERPs were time-locked to video onset, and sign onset occurred approximately 500 ms later.
Therefore, the very early effects of frequency observed in the first epoch (100–200 ms and
200–300 ms) are most likely associated with the transitional movement of the signer’s hand(s)
from the resting position on her lap to the target location of the sign (see Figure 1). In these
early epochs, lower frequency signs produced greater negativities than higher frequency signs
at frontal and central sites. Unlike spoken languages, the linguistic articulators for sign lan-
guages are fully visible, and psycholinguistic research has shown that signers are sensitive
Neurobiology of Language
260
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurophysiological correlates of sign recognition
to early linguistic cues that are visible in the transitional movement from a resting position to
sign onset, as well as in the transitional movement between signs. For example, in gating stud-
ies signers can often identify the handshape and location of the sign prior to the onset of the
sign, both when signs are presented in isolation (Emmorey & Corina, 1990; Grosjean, 1981)
and when presented in a sentence context (Clark & Grosjean, 1982). Further, in an ERP study
of sentence processing in German Sign Language, Hosemann, Herrmann, Steinbach,
Bornkessel-Schlesewsky, and Schlesewsky (2013) found that the onset of the N400 response
to sentence-final anomalous signs occurred prior to sign onset and thus had to be elicited by
information present during the transition phase. We suggest that the very early effect of lexical
frequency observed during the transition phase for isolated signs in the present study may re-
flect sensitivity to the frequency of sublexical properties, particularly handshape. Caselli et al.
(2017) reported that handshape frequency was positively correlated with lexical frequency
(i.e., higher frequency handshapes occurred in more frequent signs), but location frequency
was not correlated with lexical frequency. If signers recognize a sign’s handshape during the
transition phase, then it is possible that less frequent handshapes elicit a more negative re-
sponse compared to more frequent handshapes (that occur in more frequent signs).
Frequency effects next emerged in the 400–500-ms epoch at frontal sites (slightly left later-
alized), and then there was a later, more central-posterior frequency effect that began to
emerge in the 800–900-ms epoch. We suggest that the different timing and distribution of
these two effects may reflect sensitivity to frequency at two distinct levels: phonological form
and lexical-semantics. For spoken languages, frequency effects are known to occur at multiple
levels, including phonological encoding and lexical-semantic processing (e.g., Knobel,
Finkbeiner, & Caramazza, 2008; Winsler et al., 2018). Previous ERP studies investigating im-
plicit and explicit phonological priming in ASL indicate a frontal distribution for form priming,
with smaller negativities over anterior sites for sign pairs that overlap in form (e.g., share the
same handshape and location) compared to unrelated sign pairs (Meade, Midgley, Sevcikova
Sehyr, Holcomb, & Emmorey, 2017; Meade et al., 2018). These results lead us to hypothesize
that this earlier anteriorly distributed effect is related to accessing the phonological form of
signs.
The later central-posterior distribution is more typical of the frequency effect observed in
the N400 window for spoken language, which is usually associated with lexical-semantic pro-
cesses. Note that this later effect is significant in the 900–1,000-ms epoch, which is 400 ms
after the average sign onset (i.e., 500 ms after stimulus onset). It may be possible to observe
separate effects of phonological frequency and lexical-semantic frequency in the ERPs to signs
because phonological form encoding involves recognition of large movements of the hands
and arms and distinct body configurations. The neural regions involved in form processing
may be more neurally segregated from regions involved in lexical-semantic processing for sign
language compared to spoken language. For spoken language, temporal cortex is involved in
both phonological and lexical-semantic processing (e.g., Hickok & Poeppel, 2007), whereas
for sign language more distinct neural regions appear to be involved in phonological process-
ing (parietal cortex) and lexical-semantic processing (temporal cortex; see MacSweeney &
Emmorey, 2020). In addition, the timing of these processes may be more segregated for sign
language because the articulators are visible during the transition to sign onset. For speech,
word onset coincides with stimulus onset, whereas there is ∼500-ms delay between stimulus
(video) onset and sign onset that contains form information about the upcoming sign. Future
work that separately manipulates phonological and semantic variables will help to determine
whether the distinct timing and distribution of the frequency effects observed here are linked to
different processing levels involved in sign recognition.
Neurobiology of Language
261
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurophysiological correlates of sign recognition
Concreteness
A robust effect of concreteness began to emerge 700–800 ms after video onset (∼200 ms after
sign onset) and continued throughout all analyzed epochs. The polarity of the effect (more
negative for more concrete signs) and the wide distribution around central electrode sites par-
allel what has been found for spoken languages (e.g., Holcomb et al., 1999; Winsler et al.,
2018). ERP effects of concreteness on word recognition are typically interpreted as reflecting
richer semantic representations and denser links to associated semantic representations for
more concrete words compared to more abstract words (Holcomb et al., 1999; West &
Holcomb, 2000). Larger negativities to concrete words may result from increased neural ac-
tivation arising from the more extensive semantic networks of these words, although greater
N400 negativity does not appear to be monotonically associated with an increasing number of
semantic features (Amsel & Cree, 2013; Kounios et al., 2009). Abstract words presented in
isolation (as in the current study) may receive less semantic processing because they activate
a smaller number of associations that may not be easily integrated into a unified concept
(Barsalou & Wiemer-Hastings, 2005). In addition, semantic processing of concrete words en-
gages a larger number of neural networks that are linked to sensorimotor properties of the
concept (e.g., Barber, Otten, Kousta, & Vigliocco, 2013; Binder, Desai, Graves, & Conant,
2009). The parallel ERP results for signs and words indicate that language modality does
not impact the neural networks that underlie processing of concrete vs. abstract concepts.
The time course of the concreteness effect likely reflects how the perception of single signs
(produced in isolation) unfolds over time. A robust effect of concreteness emerges in the 700–
800-ms time window (see Figure 4A), which is ∼200 ms after the average sign onset (i.e., when
the hand reaches the target location on the face/body or in neutral space). We suggest that
some signs have already been recognized at this time window (Emmorey & Corina, 1990),
giving rise to the concreteness effect. There is a main effect of concreteness for the next four
epochs, and we suggest that this timing is consistent with the N400 concreteness effect ob-
served for spoken and written word recognition.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Iconicity
There were no significant effects of iconicity on ERPs until the final two epochs (1,000–
1,200 ms). In these late epochs, the effect of iconicity was relatively weak (compared to
the effects of frequency and concreteness) and consisted of a more negative response for
less iconic signs at posterior sites. This finding is consistent with the results of Mott et al.
(2020) who reported a late effect of iconicity when deaf signers performed a word-sign trans-
lation task. Specifically, noniconic signs exhibited a weaker translation priming effect (i.e.,
larger negativity for signs preceded by unrelated than by related English primes) compared to
iconic signs in time windows that followed lexical by access (i.e., after the N400 window
where translation priming was observed, but there was no interaction with iconicity).
Following Mott et al., we suggest that the weak, late effect of iconicity reflects postlexical
sensitivity to sign iconicity, perhaps reflecting a strategic effect when making the semantic
categorization judgment. The (weak) iconicity effect emerged about 200–300 ms prior to the
average RT for the “go” probe decision.
Our results indicate that the degree of form-meaning mapping does not impact the temporal
neural dynamics of sign recognition and lexical access. In contrast to lexical frequency and
concreteness, there does not appear to be a neural response that is modulated by lexical var-
iation in iconicity during sign comprehension. Although sign iconicity may impact
Neurobiology of Language
262
Neurophysiological correlates of sign recognition
performance on tasks such as picture naming (e.g., Navarrete et al., 2017) or picture-sign
matching (e.g., Thompson et al., 2009), iconicity does not appear to have a general impact
on the neural networks that support sign recognition (see also Bosworth & Emmorey, 2010).
For spoken language, Lockwood, Hagoort, and Dingemanse (2016) found that iconicity
(sound symbolism) impacted ERPs for new learners, specifically Dutch speakers who learned
Japanese ideophones (marked words that depict sensory imagery; Dingemanse, 2012) in either
a “real” condition (the correct Dutch translation) or in an “opposite” condition (the Dutch
translation had the opposite meaning of the ideophone). Ideophones (auditorily presented)
in the real condition elicited a larger P3 component and late positive complex compared to
ideophones in the opposite condition. Further, these effects were greater for individuals who
were more sensitive to sound symbolism (as assessed in a separate task). For native Japanese
speakers, Lockwood and Tuomainen (2015) compared ERPs to iconic adverbs (adverbial ideo-
phones) and arbitrary adverbs while participants made sensibility judgments to visually pre-
sented sentences that differed only in the type of adverb. Iconic adverbs elicited a greater P2
response than arbitrary adverbs, and there was a long-lasting late effect of iconicity, which the
authors interpreted as a late positive complex. The authors speculated that the P2 effect arises
from the integration of sound and sensory information associated with the distinctive phonol-
ogy of ideophones and the later effect may reflect facilitated lexical access for arbitrary ad-
verbs compared to ideophones.
However, ideophones differ from iconic signs because ideophones occur in sparse phono-
logical neighborhoods (due to their distinctive phonology; Dingemanse, 2012), whereas
iconic signs tend to be found in dense phonological neighborhoods (Caselli et al., 2017)
and are not phonologically marked. In addition, highly iconic ASL signs tend to be found
in dense semantic neighborhoods, whereas highly iconic English words are associated with
sparser semantic neighborhoods (Thompson, Perlman, Lupyan, Sevcikova Sehyr, & Emmorey,
2020). Thus, the effect of iconicity on ERPs does not appear to be parallel for signed and
spoken languages. However, no study that we know of has investigated whether continuous
lexical variation in iconicity as measured by iconicity ratings of spoken words (e.g., Perry,
Perlman, & Lupyan, 2015) modulates ERP components associated with spoken or written
word recognition.
The Temporal Neural Dynamics of Sign Recognition: Neurobiological Effects on Lexical Access
Our results revealed neurobiological principles that hold for both signed and spoken lan-
guages, as well as neural patterns that are modulated by language modality. The early wave-
forms shown in Figures 3B–5B (100–300 ms postvideo onset) reveal that signs elicit an
occipital P1 response followed by an N1 response—both components are typically elicited
by visual stimuli, including written words (Luck, 2014). Within these two early epochs, we
observed effects of sign frequency (Figure 3A), which we attributed to signers’ sensitivity to
the frequency of handshapes that are perceived during early transitional movements. This in-
terpretation is consistent with the results of a MEG study by Almeida, Poeppel, and Corina
(2016) in which deaf signers and hearing nonsigners were asked to discriminate between still
images of possible signs and anatomically impossible signs. The earliest visual cortical re-
sponses (M100 and M130) were sensitive to this distinction only for deaf signers who also
outperformed the nonsigners on the discrimination task. The authors concluded that extensive
sign language experience (and/or deafness) can “shape early neuronal mechanisms that un-
derlie the analysis of visual communication, likely on the basis of highly articulated, predictive
internal models of gesture and language processing” (p. 372).
Neurobiology of Language
263
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurophysiological correlates of sign recognition
As can be seen in Figures 3B–5B, the N1 component was followed by the N300, a com-
ponent that has been observed in studies using pictures or gestures (rather than written or spo-
ken words) and is hypothesized to be involved in processing early visual semantic features
(e.g., Hamm, Johnson, & Kirk, 2002; McPherson & Holcomb, 1999; Wu & Coulson, 2007,
2011). As found for picture and gesture processing, the N300 to signs has an anterior distri-
bution. Meade et al. (2018) found both phonological and semantic priming effects on the
N300 (reduced negativities for target signs preceded by related versus unrelated prime signs).
Here, we observed frequency effects emerging during this component (400–500 ms postvideo
onset), and we interpreted this early anterior effect as reflecting form-based lexical frequency,
that is, accessing visual-manual phonological representations. It is possible that the N300, like
the N250 for visual words, indexes the mapping between sublexical and lexical representa-
tions. Further research is needed to determine the functional significance of the N300 compo-
nent for sign recognition and the factors that modulate this response.
As found for spoken word recognition, the N400 response to signs tends to be prolonged,
compared to the N400 elicited by visually presented words. This is likely due to the fact that
both spoken words and signs are dynamic and unfold over time. During these later epochs
(∼400 ms postsign onset), the anterior frequency effect shifted to a more widely distributed
posterior effect that we interpreted as reflecting lexical-semantic frequency. Concreteness ef-
fects were also observed during these later epochs with the same polarity (i.e., more negative
for more concrete signs) and the same distribution as observed for spoken languages. Our find-
ings support the consensus that the N400 component is associated with amodal lexical-
semantic processing (Kutas & Federmeier, 2011). The results are also consistent with ERP
studies demonstrating N400 effects for lexical-level semantic violations in signed sentences
(Capek et al., 2009; Grosvald et al., 2012; Gutierrez, Williams, Grosvald, & Corina, 2012;
Kutas et al., 1987; Neville et al., 1997).
Finally, lexical variation in iconicity did not modulate the neural response during sign rec-
ognition, suggesting that this lexical variable is not represented in the brain in a manner that is
parallel to either frequency or concreteness. Despite the pervasiveness of iconicity in ASL
(Thompson et al., 2020), there does not appear to be a general neural response that is asso-
ciated with variation in iconicity. However, the present study was only designed to identify
general patterns of effects and may not have been able to detect particularly focal effects or
nonlinear interactions with iconicity. Thus, further work is necessary to determine under what
conditions (if any) sign iconicity impacts lexical access and sign recognition and/or if there are
particular types of iconicity that might modulate the neural response to signs, such as percep-
tual or motor iconicity (Perniss, Thompson, & Vigliocco, 2010) or highly transparent signs that
are “manual cognates” with gestures (Ortega, Özyürek, & Peeters, 2019; Sevcikova Sehyr &
Emmorey, 2019).
In sum, we used a large-scale, item-based analysis with LMER models which controlled
for the colinearity of lexical variables, and this approach allowed us to identify ERP effects
that were specific to the continuous variables of lexical frequency, concreteness, and ico-
nicity. Despite the perceptual and motoric differences between signed and spoken lan-
guages, the overall results indicate that very similar electrophysiological processes
underlie lexical access for signs and words. The findings provide a better understanding
of the timing and distribution of these lexical effects on sign recognition such that future
studies can analyze them more precisely. We expect that future studies will be able to un-
cover nuances in the temporal neural dynamics of sign recognition based on the broad pat-
tern of lexical effects presented here.
Neurobiology of Language
264
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurophysiological correlates of sign recognition
ACKNOWLEDGMENTS
The authors would like to thank Cindy O’Grady Farnady for help carrying out this study. We
also thank all of the participants without whom this research would not be possible.
FUNDING INFORMATION
Karen Emmorey, National Institute on Deafness and Other Communication Disorders (http://
dx.doi.org/10.13039/100000055), Award ID: DC010997. Phillip J. Holcomb, National
Institute of Child Health and Human Development (http://dx.doi.org/10.13039/100000071),
Award ID: HD25889.
AUTHOR CONTRIBUTIONS
Karen Emmorey: Conceptualization; Supervision - participant recruitment & data collection;
Writing - original draft; Writing - review & editing. Phillip J. Holcomb: Conceptualization;
Writing - review & editing. Katherine J. Midgley: Supervision - participant recruitment & data
collection; Writing - review & editing. Kurt Winsler: Data analysis; Writing - review & editing.
Jonathan Grainger: Writing - reviewing & editing.
REFERENCES
Almeida, D., Poeppel, D., & Corina, D. (2016). The processing of
biologically plausible and implausible forms in American Sign
Language: Evidence for perceptual tuning. Language, Cognition
and Neuroscience, 31, 361–374. https://doi.org/10.1080/
23273798.2015.1100315
Amsel, B. D., & Cree, G. S. (2013). Semantic richness, concrete-
ness, and object domain: An electrophysiological study.
Canadian Journal o f Experim ental P sychology/ Revue
Canadienne de Psychologie Expérimentale, 6, 117–129. https://
doi.org/10.1037/a0029807
Baayen, R. H., Davidson, D. J., & Bates, D. M. (2008). Mixed-
effects modeling with crossed random effects for subjects and
items. Journal of Memory and Language, 59, 390–412. https://
doi.org/10.1016/j.jml.2007.12.005
Balota, D. A., Pilotti, M., & Cortese, M. J. (2001). Subjective fre-
quency estimates for 2,938 monosyllabic words. Memory &
Cognition, 29, 639–647. https://doi.org/10.3758/BF03200465
Balota, D. A., Yap, M. J., Hutchison, K. A., & Cortese, M. J.
(2012). Megastudies. In J. S. Adelman (Ed.), Visual word recog-
nition Vol. 1: Models and methods, orthography and phonology
(pp. 90–115). New York, NY: Psychology Press.
Barber, H. A., Otten, L. J., Kousta, S. T., & Vigliocco, G. (2013).
Concreteness in word processing: ERP and behavioral effects in
a lexical decision task. Brain and Language, 125, 47–53. https://
doi.org/10.1016/j.bandl.2013.01.005
Barr, D. J., Levy, R., Scheepers, C., & Tily, H. J. (2013). Random
effects structure for confirmatory hypothesis testing: Keep it max-
imal. Journal of Memory and Language, 68, 255–278. https://doi.
org/10.1016/j.jml.2012.11.001
Barsalou, L. W., & Wiemer-Hastings, K. (2005). Situating abstract
concepts. In D. Pecher & R. Zwaan (Eds.), Grounding cognition:
The role of perception and action in memory, language, and thought
(pp.129–163). New York, NY: Cambridge University Press.
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2015). Fitting lin-
ear mixed-effects models using lme4. Journal of Statistical
Software, 67(1), 1–48. arXiv:1406.5823
Baus, C., Carreiras, M., & Emmorey, K. (2013). When does iconic-
ity in sign language matter? Language and Cognitive Processes,
28, 261–271. https://doi.org/10.1080/01690965.2011.620374
Baus, C., & Costa, A. (2015). On the temporal dynamics of sign
production: An ERP study in Catalan Sign Language (LSC).
Brain Research, 1609, 40–53. https://doi.org/10.1016/j.
brainres.2015.03.013
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19, 2767–2796. https://doi.org/10.1093/cercor/bhp055
Börstell, C., Hörberg, T., & Östling, R. (2016). Distribution and
duration of signs and parts of speech in Swedish Sign Language.
Sign Language & Linguistics, 19, 143–196. https://doi.org/
10.1075/sll.19.2.01bor
Bosworth, R., & Emmorey, K. (2010). Effects of iconicity and semantic
relatedness on lexical access in American Sign Language. Journal
of Experimental Psychology: Learning, Memory, and Cognition,
36, 11573–11581. https://doi.org/10.1037/a0020934
Brysbaert, M., Warriner, A. B., & Kuperman, V. (2014).
Concreteness ratings for 40 thousand generally known English
word lemmas. Behavior Research Methods, 46, 904–911.
https://doi.org/10.3758/s13428-013-0403-5
Capek, C. M., Grossi, G., Newman, A. J., McBurney, S. L., Corina,
D., Roeder, B., & Neville, H. J. (2009). Brain systems mediating
semantic and syntactic processing in deaf native signers: Bio-
logical invariance and modality specificity. Proceedings of the
National Academy of Sciences, 106, 8784–8789. https://doi.
org/10.1073/pnas.0809609106
Carreiras, M., Guitiérrez-Sigut, E., Baquero, S., & Corina, D. (2008).
Lexical processing in Spanish Sign Language (LSE). Journal of
Memory and Language, 58, 100–122. https://doi.org/10.1016/j.
jml.2007.05.004
Caselli, N. K. (2015). Language deprivation and the American Sign
Language lexicon (Unpublished doctoral dissertation). Tufts
University, Medford, MA.
Neurobiology of Language
265
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurophysiological correlates of sign recognition
Caselli, N. K., & Pyers, J. E. (2017). The road to language learning is
not entirely iconic: Iconicity, neighborhood density, and frequency
facilitate acquisition of sign language. Psychological Science, 28,
979–987. https://doi.org/10.1177/0956797617700498
Caselli, N., Sevcikova Sehyr, Z., Cohen-Goldberg, A., & Emmorey,
K. (2017). ASL-LEX: A lexical database of American Sign
Language. Behavioral Research Methods, 49, 784–801. https://
doi.org/10.3758/s13428-016-0742-0
Clark, L. & Grosjean, F. (1982). Sign recognition processes in American
Sign Language: The effect of context. Language and Speech, 25,
325–340. https://doi.org/10.1177/002383098202500402
Dingemanse, M. (2012). Advances in the cross-linguistic study of
ideophones. Language and Linguistics Compass, 6, 654–672.
https://doi.org/10.1002/lnc3.361
Dufau, S., Grainger, J., Midgley, K. J., & Holcomb, P. J. (2015). A
thousand words are worth a picture: Snapshots of printed-word
processing in an event-related potential megastudy. Psychological
Science, 26, 1887–1897. https://doi.org/10.1177/0956797615603934
Dufour, S., Brunellière, A., & Frauenfelder, U. H. (2013). Tracking
the time course of word-frequency effects in auditory word rec-
ognition with event-related potentials. Cognitive Science, 37,
489–507. https://doi.org/10.1111/cogs.12015
Emmorey, K. (1991). Repetition priming with aspect and agreement
J o u r n a l o f
m o r p h o l o g y i n A m e r i c a n S i g n L a n g u a g e .
Psycholinguistic Research, 20, 365–388.
Emmorey, K. (2002). Language, cognition, and the brain: Insights
from sign language research. Mahwah, NJ: Erlbaum.
Emmorey, K., & Corina, D. (1990). Lexical recognition in sign lan-
guage: Effects of phonetic structure and morphology. Perceptual
and Motor Skills, 71, 1227–1252. https://doi.org/10.2466/
pms.1990.71.3f.1227
Emmorey, K., & Corina, D. (1993). Hemispheric specialization for
ASL signs and English words: Differences between imageable
and abstract forms. Neuropsychologia, 31, 645–653. https://
doi.org/10.1016/0028-3932(93)90136-N
Emmorey, K., Midgley, K. J., Kohen, C., Sevcikova Sehyr, Z.,
& Holcomb, P. J. (2017). The N170 ERP component differs in la-
terality, distribution, and association with continuous reading
measures for deaf and hearing readers. Neuropsychologia, 106,
298–309. https://doi.org/10.1016/j.neuropsychologia.2017.10.001
Emmorey, K., Petrich, J. A. F., & Gollan, T. H. (2012). Bilingual pro-
cessing of ASL-English code-blends: The consequences of acces-
sing two lexical representations simultaneously. Journal of
Memory and Language, 67, 199–210. https://doi.org/10.1016/j.
jml.2012.04.005
Emmorey, K., Petrich, J. A. F., & Gollan, T. H. (2013). Bimodal bi-
lingualism and the frequency-lag hypothesis. Journal of Deaf
Studies and Deaf Education, 18, 1–11. https://doi.org/10.1093/
deafed/ens034
Fenlon, J., Schembri, A., Rentelis, R., Vinson, D., & Cormier, K.
(2014). Using conversational data to determine lexical frequency
in British Sign Language: The influence of text type. Lingua, 143,
187–202. https://doi.org/10.1016/j.lingua.2014.02.003
Gilhooly, K. J., & Logie, R. H. (1980). Age of acquisition, imagery,
concreteness, familiarity, and ambiguity measures for 1,944 words.
Behavior, Research, Methods & Instrumentation, 12, 395–427.
Grainger, J. & Jacobs, A. M. (1996). Orthographic processing in visual
word recognition: A multiple read-out model. Psychological Review,
103, 518–565. https://doi.org/10.1037/0033-295X.103.3.518
Groppe, D. M., Urbach, T. P., & Kutas, M. (2011). Mass univariate
analysis of event-related brain potentials/fields I: A critical tutorial
review. Psychophysiology, 48, 1711–1725. https://doi.org/
10.1111/j.1469-8986.2011.01273.x
Grosjean, F. (1981). Sign and word recognition: A first comparison.
Sign Language Studies, 32, 195–219.
Grosvald, M., Gutierrez, E., Hafer, S., & Corina, D. (2012).
Dissociating linguistic and non-linguistic gesture processing:
Electrophysiological evidence from American Sign Language.
Brain and Language, 121, 12–24. https://doi.org/10.1016/j.
bandl.2012.01.005
Gutierrez, E., Williams, D., Grosvald, M., & Corina, D. (2012).
Lexical access in American Sign Language: An ERP investiga-
tion of effects of semantics and phonology. Brain Research,
1468, 63–83. https://doi.org/10.1016/j.brainres.2012.04.029
Hamm, J. P., Johnson, B. W., & Kirk, I. J. (2002). Comparison of the
N300 and N400 ERPs to picture stimuli in congruent and incon-
gruent contexts. Clinical Neurophysiology, 113, 1339–1350.
https://doi.org/10.1016/S1388-2457(02)00161-X
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8, 393–402.
https://doi.org/10.1038/nrn2113
Holcomb, P. J., Kounios, J., Anderson, J. E., & West, W. C.
(1999). Dual-coding, context-availability, and concreteness
effects in sentence comprehension: An electrophysiological
investigation. Journal of Experimental Psychology: Learning,
Memory, and Cognition, 25, 721. https://doi.org/10.1037/
0278-7393.25.3.721
Hosemann, J., Herrmann, A., Steinbach, M., Bornkessel-
Schlesewsky, I., & Schlesewsky, M. (2013). Lexical prediction
via forward models: N400 evidence from German Sign
Language. Neuropsychologia, 51, 2224–2237. https://doi.org/
10.1016/j.neuropsychologia.2013.07.013
Jung, T. P., Makeig, S., Humphries, C., Lee, T. W., Mckeown, M. J.,
Iragui, V., & Sejnowski, T. J. (2000). Removing electroencepha-
lographic artifacts by blind source separation. Psychophysiology,
37, 163–178. https://doi.org/10.1111/1469-8986.3720163
Knobel, M., Finkbeiner, M., & Caramazza, A. (2008). The many
places of frequency: Evidence for a novel locus of the lexical fre-
quency effect in word production. Cognitive Neuropsychology,
25, 256–286. https://doi.org/10.1080/02643290701502425
Kounios, J., Green, D. L., Payne, L., Fleck, J. I., Grondin, R., &
McRae, K. (2009). Semantic richness and the activation of con-
cepts in semantic memory: Evidence from event-related poten-
tials. Brain Research, 1282, 95–102. https://doi.org/10.1016/j.
brainres.2009.05.092
Kroll, J. F., & Merves, J. S. (1986). Lexical access for concrete and
abstract words. Journal of Experimental Psychology: Learning,
Memory, and Cognition, 12(1), 92–107. https://doi.org/
10.1037/0278-7393.12.1.92
Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting:
Finding meaning in the N400 component of the event-related
brain potential (ERP). Annual Review of Psychology, 62, 621–647.
https://doi.org/10.1146/annurev.psych.093008.131123
Kutas, M., Neville, H. J., & Holcomb, P. J. (1987). A preliminary
comparison of the N400 response to semantic anomalies during
reading, listening and signing. Electroencephalography and
Clinical Neurophysiology Supplement, 39, 325–330.
Lockwood, G., Hagoort, P., & Dingemanse, M. (2016). How ico-
nicity helps people learn new words: Neural correlates and indi-
vidual differences in sound-symbolic bootstrapping. Collabra,
2(1), 7. https://doi.org/10.1525/collabra.42
Lockwood, G., & Tuomainen, J. (2015). Ideophones in Japanese
modulate the P2 and late positive complex responses. Frontiers
in Psychology, 6, 933. https://doi.org/10.3389/fpsyg.2015.00933
Luck, S. J. (2014). An introduction to the event-related potential
technique. Cambridge, MA: MIT Press.
Neurobiology of Language
266
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurophysiological correlates of sign recognition
MacSweeney, M., & Emmorey, K. (2020). The neurobiology of
sign language processing. In D. Poeppel, G. Mangun, & M.
Gazzaniga (Eds.), The cognitive neurosciences VI (pp. 851–858).
Cambridge, MA: MIT Press.
McGarry, M., Mott, M., Midgley, K. J., Holcomb, P. J., & Emmorey,
K. (2018). Picture naming in American Sign Language: An ERP
study of the effects of iconicity and alignment. Poster presented
at the Society for the Neurobiology of Language meeting,
Québec City, Canada.
McPherson, W. B., & Holcomb, P. J. (1999). An electrophysiolog-
ical investigation of semantic priming with pictures of real ob-
jects. Psychophysiology, 36, 53–65. https://doi.org/10.1017/
S0048577299971196
Meade, G., Lee, B., Midgley, K. J., Holcomb, P. J., & Emmorey, K.
(2018). Phonological and semantic priming in American Sign
Language: N300 and N400 effects. Language, Cognition and
Neuroscience, 33, 1092–1106. https://doi.org/10.1080/
23273798.2018.1446543
Meade, G., Midgley, K. J., Sevickova Sehyr, Z., Holcomb, P. J., &
Emmorey, K. (2017). Implicit co-activation of American Sign
Language in deaf readers: An ERP study. Brain and Language,
170, 50–61. https://doi.org/10.1016/j.bandl.2017.03.004
Mott, M., Midgley, K. J., Holcomb, P. J., & Emmorey, K. (2020).
Cross-modal translation priming and iconicity effects in deaf
signers and hearing learners of American Sign Language.
Bilingualism: Language and Cognition, 1–13. https://doi.org/
10.1017/S1366728919000889
Navarrete, E., Peressotti, F., Lerose, L., & Miozzo, M. (2017).
Activation cascading in sign production. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 43, 302–318.
https://doi.org/10.1037/xlm0000312
Neville, H. J., Coffey, S. A., Lawson, D. S., Fischer, A., Emmorey, K., &
Bellugi, U. (1997). Neural systems mediating American Sign
Language: Effects of sensory experience and age of acquisition.
Brain and Language, 57, 285–308. https://doi.org/10.1006/
brln.1997.1739
Ortega, G., Özyürek, A., & Peeters, D. (2019). Iconic gestures serve
as manual cognates in hearing second language learners of a sign
language: An ERP study. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 46, 403–415. https://doi.org/
10.1037/xlm0000729
Perniss, P., Thompson, R., & Vigliocco, G. (2010). Iconicity as a
general property of language: Evidence from spoken and signed
languages. Frontiers in Psychology, 1, 227. https://doi.org/
10.3389/fpsyg.2010.00227
Perry, L. K., Perlman, M., & Lupyan, G. (2015). Iconicity in English
and Spanish and its relation to lexical category and age of acqui-
sition. PLOS ONE, 10, e0137147. https://doi.org/10.1371/journal.
pone.0137147
R Core Team. (2014). nlme: linear and nonlinear mixed effects
models. R package version 3.1-117. https://cran.r-project.org/
web/packages/nlme/index.html
Russell, K., Wilkinson, E., & Janzen, T. (2011). ASL sign lowering as
undershoot: A corpus study. Laboratory Phonology, 2, 403–422.
https://doi.org/10.1515/labphon.2011.015
Sevcikova Sehyr, Z., & Emmorey, K. (2019). The perceived map-
ping between form and meaning in American Sign Language
depends on linguistic knowledge and task: Evidence from iconic-
ity and transparency judgments. Language and Cognition, 11,
208–234. https://doi.org/10.1017/langcog.2019.18
Strijkers, K., Costa, A., & Thierry, G. (2009). Tracking lexical access
in speech production: Electrophysiological correlates of word
frequency and cognate effects. Cerebral Cortex, 20, 912–928.
https://doi.org/10.1093/cercor/bhp153
Thompson, R., Vinson, D., & Vigliocco, G. (2009). The link
between form and meaning in American Sign Language:
Lexical processing effects. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 35, 550–557. https://doi.org/
10.1037/a0014547
Thompson, R. L., Vinson, D. P., & Vigliocco, G. (2010). The link
between form and meaning in British Sign Language: Effects of
iconicity for phonological decisions. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 36, 1017–1027.
https://doi.org/10.1037/a0019339
Thompson, W., Perlman, M., Lupyan, G., Sevcikova Sehyr, Z., &
Emmorey, K. (2020). A data-driven approach to the semantics
of iconicity in American Sign Language and English. Language
and Cognition, 12(1), 182–202. https://doi.org/10.1017/langcog.
2019.52
Vinson, D. P., Cormier, K., Denmark, T., Schembri, A., & Vigliocco,
G. (2008). The British Sign Language (BSL) norms for age
of acquisition, familiarity, and iconicity. Behavior Research
Methods, 40, 1079–1087. https://doi.org/10.3758/ BRM.
40.4.1079
Vinson, D., Thompson, R. L., Skinner, R., & Vigliocco, G. (2015).
A faster path between meaning and form? Iconicity facilitates sign
recognition and production in British Sign Language. Journal of
Memory and Language, 82, 56–85. https://doi.org/10.1016/j.
jml.2015.03.002
West, W. C., & Holcomb, P. J. (2000). Imaginal, semantic, and
surface-level processing of concrete and abstract words: An elec-
trophysiological investigation. Journal of Cognitive Neuroscience,
12, 1024–1037. https://doi.org/10.1162/08989290051137558
Winsler, K., Midgley, K. J., Grainger, J., & Holcomb, P. J. (2018). An
electrophysiological megastudy of spoken word recognition.
Language, Cognition and Neuroscience, 33, 1063–1082.
https://doi.org/10.1080/23273798.2018.1455985
Wu, Y. C., & Coulson, S. (2007). How iconic gestures enhance
communication: An ERP study. Brain and Language, 101, 234–245.
https://doi.org/10.1016/j.bandl.2006.12.003
Wu, Y. C., & Coulson, S. (2011). Are depictive gestures like pic-
tures? Commonalities and differences in semantic processing.
Brain and Language, 119, 184–195. https://doi.org/10.1016/j.
bandl.2011.07.002
Neurobiology of Language
267
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
2
4
9
1
8
6
7
7
0
4
n
o
_
a
_
0
0
0
1
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3