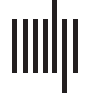FORSCHUNG
Reconfigurations within resonating
communities of brain regions following
TMS reveal different scales of processing
1,2, Arian Ashourvan1,2, Steven M. Thurman1,
Javier O. Garcia
Ramesh Srinivasan3,4, Danielle S. Bassett2,5, and Jean M. Vettel1,2,6
1UNS. Army CCDC Army Research Laboratory, Aberdeen Proving Ground, MD, USA
2Department of Bioengineering, University of Pennsylvania, Philadelphia, PA, USA
3Department of Cognitive Sciences, Universität von Kalifornien, Irvine, Irvine, CA, USA
4Abteilung für Biomedizintechnik, Universität von Kalifornien, Irvine, Irvine, CA, USA
5Department of Electrical and Systems Engineering, University of Pennsylvania, Philadelphia, PA, USA
6Psychological and Brain Sciences, Universität von Kalifornien, Santa Barbara, Santa Barbara, CA, USA
Schlüsselwörter: Neuroimaging, Network flexibility, Network allegiance, Local-global processing, TMS,
EEG
ABSTRAKT
An overarching goal of neuroscience research is to understand how heterogeneous neuronal
ensembles cohere into networks of coordinated activity to support cognition. To investigate
how local activity harmonizes with global signals, we measured electroencephalography
(EEG) while single pulses of transcranial magnetic stimulation (TMS) perturbed occipital and
parietal cortices. We estimate the rapid network reconfigurations in dynamic network
communities within specific frequency bands of the EEG, and characterize two distinct
features of network reconfiguration, flexibility and allegiance, among spatially distributed
neural sources following TMS. Using distance from the stimulation site to infer local and
global effects, we find that alpha activity (8–12 Hz) reflects concurrent local and global
effects on network dynamics. Pairwise allegiance of brain regions to communities on average
increased near the stimulation site, whereas TMS-induced changes to flexibility were
generally invariant to distance and stimulation site. Im Gegensatz, communities within the beta
(13–20 Hz) band demonstrated a high level of spatial specificity, particularly within a cluster
comprising paracentral areas. Zusammen, these results suggest that focal magnetic
neurostimulation to distinct cortical sites can help identify both local and global effects on
brain network dynamics, and highlight fundamental differences in the manifestation of
network reconfigurations within alpha and beta frequency bands.
ZUSAMMENFASSUNG DES AUTORS
TMS may be used to probe the causal link between local regional activity and global brain
Dynamik. Using simultaneous TMS-EEG and dynamic community detection, we introduce
what we call “resonating communities” or frequency band-specific clusters in the brain, as a
way to index local and global processing. These resonating communities within the alpha
and beta bands display both global (or integrating) behavior and local specificity, highlighting
fundamental differences in the manifestation of network reconfigurations.
Keine offenen Zugänge
Tagebuch
Zitat: Garcia, J. O., Ashourvan, A.,
Thurman, S. M., Srinivasan, R., Bassett,
D. S., & Vettel, J. M. (2020).
Reconfigurations within resonating
communities of brain regions following
TMS reveal different scales of
Verarbeitung. Netzwerkneurowissenschaften,
4(3), 611–636. https://doi.org/10.1162/
netn_a_00139
DOI:
https://doi.org/10.1162/netn_a_00139
zusätzliche Informationen:
https://doi.org/10.1162/netn_a_00139
Erhalten: 10 September 2019
Akzeptiert: 23 Marsch 2020
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Javier Omar Garcia
javier.o.garcia.civ@mail.mil
Handling-Editor:
Martijn van den Heuvel
Urheberrechte ©: No rights reserved. Das
work was authored as part of the
Contributors official duties as an
Employee of the United States
Government and is therefore the work
of the United States Government. In
accordance with 17 U.S.C. 105, NEIN
copyright protection is available for
such works under U.S. law.
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
N
e
N
_
A
_
0
0
1
3
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network reconfigurations after TMS
EINFÜHRUNG
The brain is an intricate collection of heterogeneous areas (Alivisatos et al., 2012), and a cen-
tral goal of neuroscientific research is to understand how the coordination of these different
regions supports cognition (Azevedo et al., 2009; Bressler & Menon, 2010; Gollo, Roberts, &
Cocchi, 2017). One theoretical approach encapsulates the coordinated activity into a frame-
work of scales, and research has examined how local regional activity harmonizes with global
Signale (Bressler & Kelso, 2001). Local activity refers to cortical or thalamocortical interactions
that reflect the transient coordination of inhibitory and excitatory neighboring neurons, con-
strained by basic neurophysiological factors such as refractory limitations and synaptic rising
(Fries, Nikoli´c, & Singer, 2007). Jedoch, research has shown that this local neural activity can
also be modulated by global activity in the brain (for review, see Buzsáki & Draguhn, 2004;
Buzsáki & Wang, 2012). Global activity arises from propagation delays in cortico-cortical
fibers and reflects the dynamic interactions and synchronization among distal networks. Das
conceptual framework of local and global networks interacting in cognitive processes is critical
to the interpretation of neurophysiological signals. Noch, how this activity coheres to manifest
cognition is still an active area of study (Bressler & Kelso, 2001; Cocchi, Gollo, Zalesky, &
Breakspear, 2017).
EEG affords a natural way to study the scales of processing by examining oscillatory dynam-
ics in different frequency bands (Buzsáki & Draguhn, 2004; Canolty & Ritter, 2010). Changes
in power in high frequencies, such as beta and gamma, have been used to infer local dynam-
ics arising from the synchronization of populations of neurons (Brunel & Wang, 2003; Geisler,
Brunel, & Wang, 2005). Ähnlich, the emergent activity in slower EEG frequencies, reichend
across delta, theta, and alpha, has been interpreted as global activity arising from long-distance
coordination of synchronized neural firing in disparate brain regions (Brunel & Wang, 2003;
Geisler et al., 2005); Jedoch, there are known examples of cross-frequency interactions that
challenge a strict local/global interpretation on frequency dynamics (Canolty & Ritter, 2010).
Zusammen, results from EEG studies have indicated the importance of both local and global ac-
tivity, indexed by high- and low-frequency oscillations, for understanding variability in human
behavior (Buzsáki, 2006; Nunez & Srinivasan, 2006; Nunez, Wingeier, & Silberstein, 2001;
Volberg, Kliegl, Hanslmayr, & Greenlee, 2009). Jedoch, EEG provides only an inferential
framework to study interactions across scales of neural activity. Advancements in neurostim-
ulation paradigms may provide an avenue to directly study the causal role of local changes
in oscillatory dynamics on global dynamics (Bergmann, Karabanov, Hartwigsen, Thielscher,
& Siebner, 2016; Pascual-Leone, Walsh, & Rothwell, 2000), a long-known property of neu-
rostimulation (Ilmoniemi et al., 1997).
Transcranial magnetic stimulation (TMS) has been proposed as a method to actively probe
the dynamic interplay between local processing and consequent global interactions with more
distal regions of the brain (Massimini, Tononi, & Huber, 2009; Romei, Thut, Mok, Schyns, &
Driver, 2012). Traditionell, single-pulse TMS is a technique used to induce a short, con-
trolled burst of activity in a predetermined local brain region, directly causing a change in
the local dynamics (Pascual-Leone et al., 2000). Research has identified behavioral outcomes
resulting from local stimulation for patients in clinical settings and healthy individuals in ex-
perimental tasks. Zum Beispiel, local stimulation in patients can successfully determine stroke
recovery (for review, see Auriat, Neva, Peters, Ferris, & Boyd, 2015), mitigate severe affective
disorders (z.B., Berman et al., 2000), and preserve motor and language functions in presur-
gical mapping (Eldaief, Drücken Sie, & Pascual-Leone, 2013). TMS has also been successfully em-
ployed to confirm the role of an individual brain region on task performance, ranging from
sensory attention (Herring, Thut, Jensen, & Bergmann, 2015; Romei, Murray, Cappe, & Thut,
2013; Taylor & Thut, 2012) to working memory performance (Brunoni & Vanderhasselt, 2014;
Elektroenzephalographie (EEG):
A method to monitor electrical
activity emanating from the brain
but recorded from the scalp.
Transcranial magnetic
stimulation (TMS):
A type of brain stimulation that
noninvasively introduces electrical
current into the brain to perturb
neural activity.
Netzwerkneurowissenschaften
612
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
N
e
N
_
A
_
0
0
1
3
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network reconfigurations after TMS
Rose et al., 2016). Noch, extant experimental research has also demonstrated that the brain alter-
limited to local perturbations (Ilmoniemi et al., 1997;
ations caused by TMS are not
Sale, Mattingley, Zalesky, & Cocchi, 2015). By pairing TMS with other concurrent imaging
modalities like EEG, it provides an innovative approach to study connectivity relationships
among disparate brain regions (Cocchi et al., 2015; Mancini et al., 2017; Siebner et al., 2009).
Multimodal studies have paired TMS with functional neuroimaging, such as fMRI (z.B.,
Bestmann et al., 2008; Bohning et al., 1999), EEG (z.B., Bortoletto, Veniero, Thut, & Miniussi,
2015; Garcia, Grossman, & Srinivasan, 2011), and PET (z.B., Paus, 1998), and measured
stimulation-induced responses in brain areas that are distal to the stimulation site, indicating
that stimulation can induce transient coordination between local and global activity (Bestmann
et al., 2008; Driver, Blankenburg, Bestmann, Vanduffel, & Ruff, 2009; Paus, 1998). Comple-
menting these findings, computational models of neurodynamics have demonstrated that re-
gional differences in structural connectivity may provide a mechanistic account for how local
network activity that is induced from a focal TMS pulse can propagate along cortico-cortical
fibers to influence global network synchronization (Gollo et al., 2017; Muldoon et al., 2016).
This idea is supported by neurostimulation research that shows a structure-function constraint
to the local stimulation and subsequent global (von)synchronization (Amico et al., 2017). Während
both experimental and modeling work have suggested the importance of interacting networks,
few studies have employed the rich set of tools of network science to understand the propa-
gation of TMS stimulation throughout the brain (Bortoletto et al., 2015). Network science not
only provides a mathematical language to describe complex connectivity patterns resulting
from stimulation; previous research has also proposed a variety of summary metrics in which
to characterize local and global connectivity in the brain (for review, see Garcia, Ashourvan,
Muldoon, Vettel, & Bassett, 2018). In this study, we address this existing gap in the literature
and employ a method recently developed in network science to study the interactions of local
connectivity and global network dynamics following TMS stimulation.
We investigated network reconfigurations from resting-state EEG following single pulses of
transcranial magnetic stimulation using a method from network science that reveals modular
architecture in the brain (Bassett & Bullmore, 2006; Bullmore & Spurns, 2012; Ercsey-Ravasz
et al., 2013). Participants received single pulses of TMS to occipital or parietal cortex, and we
computed functional connectivity using EEG data for a 2-s epoch surrounding stimulation (−1
Zu 1). Our theoretical question focused on the comparison of stimulations to spatially disparate,
large lobes of the brain, investigating how stimulation influenced network dynamics follow-
ing stimulation as indexed by the modular architecture of the functional connectivity patterns.
Each module is composed of regions with synchronized activity that are thought to be dynam-
ically linked for the purpose of cohesive processing (Achard, Salvador, Whitcher, Suckling, &
Bullmore, 2006; Bassett & Bullmore, 2006; Spurns, Chialvo, Kaiser, & Hilgetag, 2004). To
index local and global activity, we investigate two frequency bands that probe brain dynamics
across these scales. We separately characterize the modular architecture of resting-state EEG
within the alpha band and within the beta band, from which we define resonating commu-
nitäten, or communities of brain regions restricted to each frequency band. This delineation
was inspired, in part, by the theoretical proposal by Rosanova et al. (2009) that posits that
brain regions have a primary natural frequency: Resting-state activity is dominated by alpha
in the occipital cortex, whereas parietal activity is dominated by beta activity. Folglich,
we hypothesized that network changes in these frequency bands would both differentiate the
location of the stimulation site (occipital vs. parietal), and reveal the spatial scale (local vs.
global) of the propagation of network perturbations arising from TMS.
Resonating communities (or clusters):
Cluster of brain nodes representing a
network defined by band-specific
oscillatory activity (z.B., alpha
Aktivität).
Netzwerkneurowissenschaften
613
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
N
e
N
_
A
_
0
0
1
3
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network reconfigurations after TMS
Module allegiance:
A metric derived from a series of
node labels (d.h., network affiliation)
that indicates how often nodes are
within the same network of other
Knoten.
Network flexibility:
A metric derived from a series of
labels that reveals how often a node
changes its network affiliation across
Zeit.
Debiased weighted phase lag
index (dwPLI):
A specific type of functional
Konnektivität, often used in EEG
Analyse, that is specifically
designed to estimate the nodal
interrelationship with little artifact
interference.
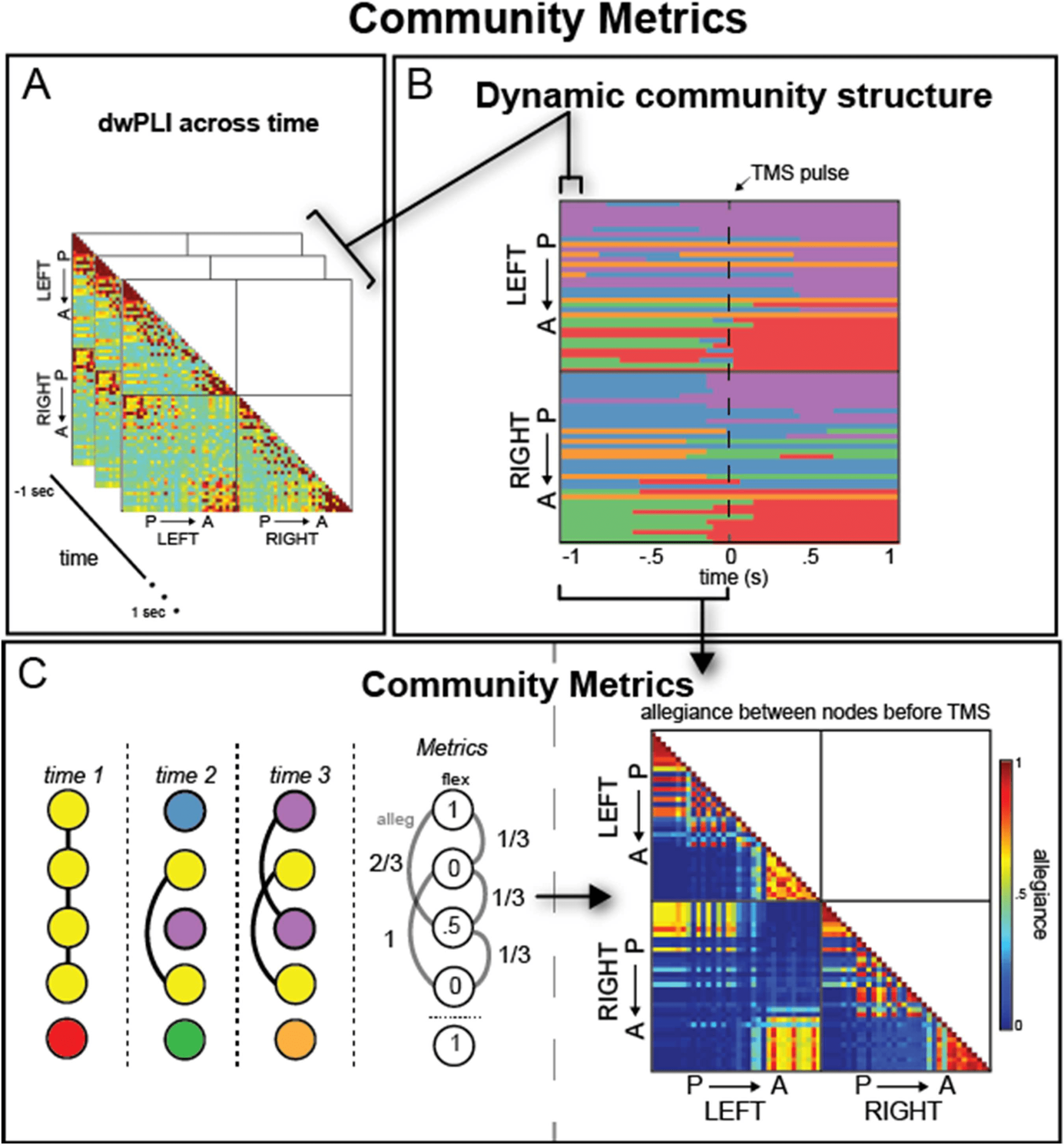
To characterize changes in functional network organization before and after stimulation, Wir
used two well-established metrics from network science: module allegiance and network flex-
ibility. Allegiance estimates how often regions are functionally connected with other regions,
capturing stable subnetworks in the community structure across time points. Flexibility, In
Kontrast, reveals the extent to which a region frequently (and flexibly) changes its assignment
across communities between time points. Daher, allegiance is a construct that can capture co-
ordinated activity of each node with every other node in the brain, thereby increasing the reso-
lution of community assignments, whereas flexibility reveals the propensity of brain regions to
change affiliations overall. We argue that these complementary metrics are uniquely suited to
investigate the scale of processing effects of stimulation since allegiance captures the unique
shifts between each pair of regions and flexibility identifies whether a node tends to shift its
community affiliation over time. Our analyses extend previous research that has found that network
flexibility successfully characterizes large-scale functional differences (z.B., Telesford et al., 2016),
for example in executive function (Braun et al., 2015) und Stimmung (Betzel, Satterthwaite, Gold,
& Bassett, 2017). Allegiance, andererseits, has been used to describe observed net-
work dynamics on a finer scale, estimating alignment with a predefined functional architecture
(Bassett et al., 2015) as well as identifying transitions among certain network configurations
(Ashourvan, Gu, Mattar, Vettel, & Bassett, 2017). Across the set of network science metrics
adapted for neuroscience application (for review, see Garcia et al., 2018), allegiance and flex-
ibility are the best suited to identify changes in scales of processing.
Using these measures, we report substantial differences between the alpha and beta band
communities. While activity in the alpha network revealed a dynamic interplay of local and
global connectivity, as hypothesized, communities within the beta band displayed a spatial
specificity across both metrics, suggesting a more local connectivity impact of stimulation. To-
gether, these results show how focal TMS to distinct cortical sites can help reveal both local
and global effects on dynamic network configurations, and demonstrate fundamental differ-
ences in the manifestation of network effects in alpha and beta frequency bands in different
areas of the brain.
ERGEBNISSE
Here we studied the brain dynamics following single pulses of TMS to occipital and parietal
cortex using recently developed approaches from network science. Erste, TMS was delivered
to two separate sites and we focused our analysis on two separate frequency bands. Nächste, Sei-
cause of the artifact-prone simultaneous technique of TMS-EEG (Rogasch & Fitzgerald, 2013),
we took several precautions and used strict criteria to reduce the measured artifact (see the
Materials and Methods section). We then reconstructed estimated neural sources on a volu-
metric brain mesh and then extracted time series for 68 Gehirnregionen (Figur 1). Using ep-
ochs surrounding the stimulation period, we computed functional connectivity between all
region pairs using the debiased weighted phase lag index (dwPLI)
that has shown robustness
to noise (Vinck, Oostenveld, van Wingerden, Battaglia, & Pennartz, 2011;Vindiola, Vettel,
Gordon, Franaszczuk, & McDowell, 2014). Our analysis focused on connectivity in the al-
pha and beta frequency bands since these bands have been suggested as resonant frequencies
within the stimulated regions, alpha in occipital and beta in parietal regions (Laufs et al., 2003;
Rosanova et al., 2009). We interpret our results within this context, and while our results con-
verge to a narrative of local/global dynamics across frequency bands, we also must consider
potential confound to these techniques (z.B., auditory responses, muscle contractions, und das
wie; for review, see Rogasch & Fitzgerald, 2013).
Netzwerkneurowissenschaften
614
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
N
e
N
_
A
_
0
0
1
3
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Network reconfigurations after TMS
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
N
e
N
_
A
_
0
0
1
3
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figur 1. Whole-brain connectivity changes following stimulation. (A) Average dwPLI across the
brain between 5 Hz and 25 Hz. (B, C) Debiased weighted phase lag index (dwPLI) differences be-
tween the second after TMS (post-TMS) and the second before TMS (pre-TMS) intervals across trials
averaged for occipital stimulation within the alpha band (B) and beta band (C). (D, E) Results sim-
ilar to Panels B and C, but for parietal stimulation. Brain insets display the significant connections
(P < 0.05, FDR adjusted) across the brain, providing a topographical illustration of the connectiv-
ity matrices where red lines indicate increased connectivity following stimulation and blue lines
indicate decreased connectivity following stimulation.
Stimulation Effects on Whole-Brain Connectivity
We began by examining patterns of functional connectivity in a whole-brain analysis (see
Figure 1). We observed slightly higher connectivity across the brain within the alpha band
(8–12 Hz) both before (black dotted line in Figure 1A) and after stimulation to either site in
bilateral occipital cortex (red line) or bilateral parietal cortex (blue line) compared with other
frequency bands. This dominant response in whole-brain alpha synchrony likely reflects its
role as a diffuse, communicative signal with multiple functions (Ba¸sar, Ba¸sar-Ero˘glu, Karaka¸s,
& Schürmann, 1999), serving as a global signal for sensory and information processing.
We next investigated changes in connectivity following stimulation by comparing changes
between pre- and post-TMS intervals. As shown in Figure 1A, we observed that the average
connectivity between all region pairs did not show much change within the alpha band after
Network Neuroscience
615
Network reconfigurations after TMS
stimulation to either occipital or parietal sites (occipital: t(9) = −0.95, p = 0.36; parietal:
t(9) = 1.05, p = 0.32, all uncorrected), and this was mirrored in the beta band with minimal
connectivity differences for both stimulation locations (occipital: t(9) = −1.39, p = 0.20;
parietal: t(9) = 0.41, p = 0.69, all uncorrected). However, there was a marked difference
between occipital and parietal stimulation sites when examining the directionality and spa-
tial specificity of the change following stimulation (Figure 1B–E). We submitted the difference
in the average dwPLI estimate 1 s after the TMS pulse to that before the TMS pulse using
a one-sample t test and then adjusted for multiple comparisons via false discovery rate (q;
Benjamini & Yekutieli, 2001). By subtracting the post-TMS dwPLI estimate from the pre-TMS
baseline, we observed a dispersed global decrease in connectivity for occipital stimulation
(Figure 1B–C) for the regional pairs with the largest differences within the alpha and beta
bands. Significant connections show some regional specificity, where the beta band shows
decreases in connectivity between lateral central locations and medial frontal sites ( q < 0.05).
The alpha band shows a similar connectivity pattern with an additional increase in connectivity
between lateral regions toward the center of the brain. In contrast, we observed a marked in-
crease within central and parietal sites as well as a frontal decrease in connectivity for parietal
stimulation (q < 0.05; Figure 1D–E). The alpha band shows a significant pattern of connec-
tivity increases along in parietal regions (q < 0.05), but this pattern is less robust within the
beta band. Collectively, these whole-brain connectivity results show some frequency speci-
ficity for the stimulation sites, as might be predicted based on theories that suggest that stim-
ulation could be facilitated or decremented by the inherent resonant frequency of the tissue
(Rosanova et al., 2009) and a difference between stimulation sites as well. Since these con-
nectivity results show both site and frequency specificity and generality from stimulation, it is
possible that a portion of these results (the pattern that is similar across the stimulation sites and
oscillations of interest) is driven by confounding variables (for review, see Rogasch & Fitzgerald,
2013), despite the extreme caution taken in artifact reduction (see the Materials and Methods
section). Nevertheless, this observation could reflect the global influence of these regions on
whole-brain connectivity rather than their targeted effects on subnetworks. Consequently, we
next employed recent methods from network science to examine the effect of stimulation at a
finer scale than average connectivity across nodes.
Community Organization in Resting Networks
To examine stimulation effects in brain communities, we capitalized on a network science
approach that has been used previously to study modularity in brain networks. To estimate
dynamic community structure, we optimize a multilayer modularity quality function, Q,
using a Louvain-like greedy algorithm (Blondel, Guillaume, Lambiotte, & Lefebvre, 2008;
Mucha, Richardson, Macon, Porter, & Onnela, 2010) to assign brain regions to communities,
where each layer is a separate time slice. With this optimization, we extract our experimental
communities by finding an optimal parameter scheme, which is composed of two parameters:
(a) a structural resolution γ parameter and (b) a temporal resolution ω parameter. These two
parameters determine the scale of the resulting graph, both structurally and temporally. As
described in Garcia et al. (2018), there are several heuristics we may use to determine the
optimal parameter for our dataset. We chose an unbiased “difference” heuristic because of
the unique properties of this stimulation dataset. With this method, we compare the estimated
Q from the pre-TMS interval to a Qnull derived from a shuffled null connectivity matrix where
we shuffle the pairwise dwPLI values, destroying the correlational structure observed in EEG
data for each subject and parameter pairing. Each Q was then subtracted for each parameter
pairing, comparing the observed model’s Q (from the unperturbed EEG connectivity patterns)
Network Neuroscience
616
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
and the null model’s Qnull (shuffled connectivity patterns) for each subject; our analysis found
a clear peak in the resulting Q matrix, suggesting that the range used was appropriate for this
dataset.
This data-driven approach showed more local granularity in the network landscape follow-
ing stimulation. Importantly, we defined network communities without stimulation during a
period of rest. This allowed us to interrogate the dynamics of community reconfigurations
following TMS, given a natural baseline, unbiased by the stimulation itself. Importantly, how-
ever, we interpret our results both within the confines of this community organization (Figures 3
and 4) and outside of these confines (Figure 5). We defined network communities separately
for both the alpha and beta bands, and used the most robust arrangement across the 100 it-
erations of modularity maximization as the final community structure. The 100 iterations of
the pre-TMS interval were remarkably robust and consistent, showing 100% agreement across
iterations for the alpha band and 98% agreement across the iterations within the beta band.
We also observed noteworthy similarity (approximately 97% spatial similarity) between them
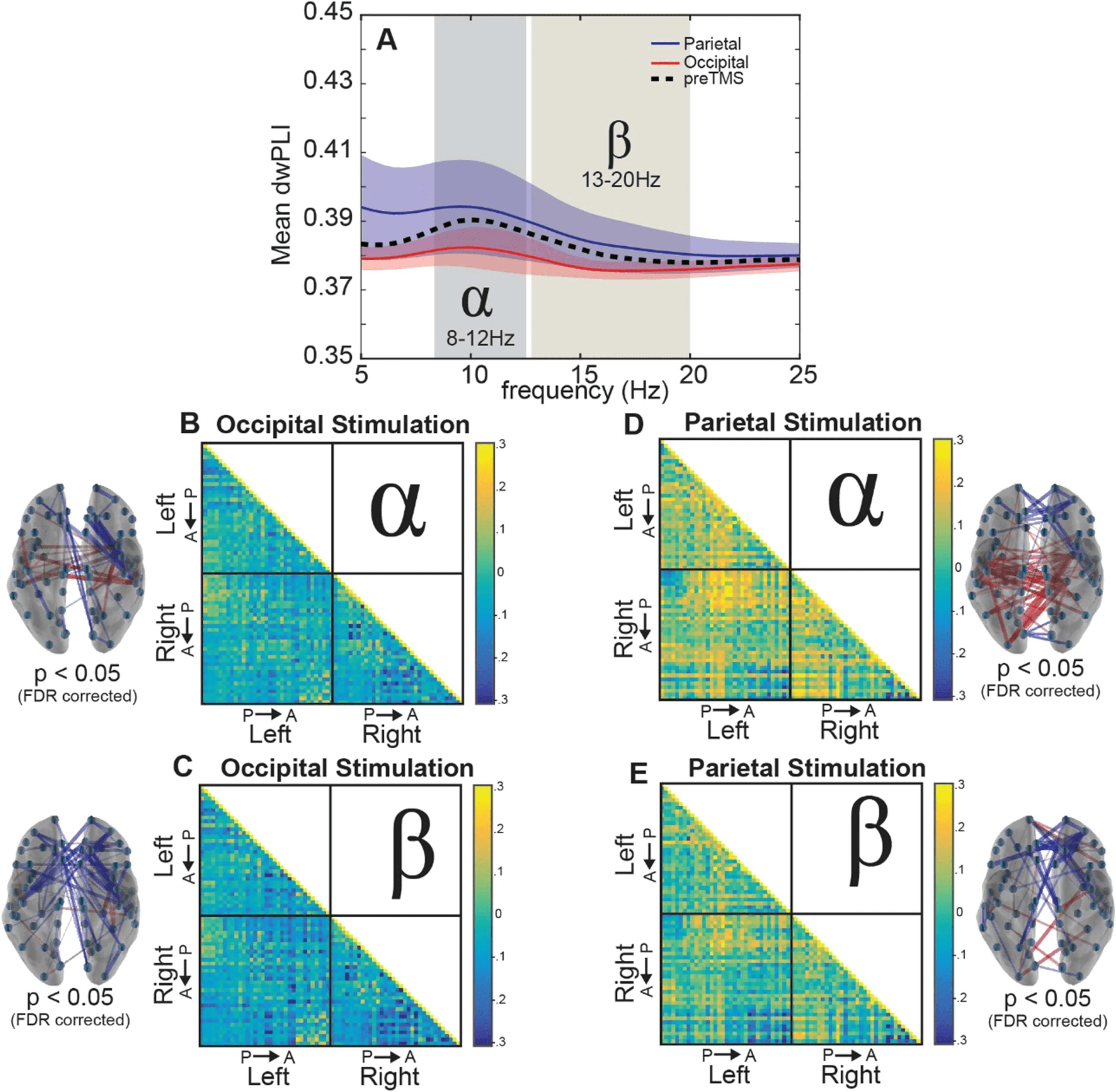
except for a small cluster of motor-related brain regions (Figure 2).
In the alpha band (Figure 2, left), five communities are illustrated: a bilateral occipital-
parietal network (Occ, blue); a right paracentral community (RPC, pink); a left temporal net-
work (LTem, yellow); a right temporal network (RTem, green); and a bilateral frontal network
(Fron, orange). This largest community (Occ) is a large cluster of regions in occipital and pari-
etal cortex, an organization that is perhaps unsurprising, given the commonly observed peak
of the alpha rhythm within occipital-parietal regions (Hari & Salmelin, 1997).
Interestingly,
five similar communities were also found within the beta band (Figure 2, right), and the only
observed difference was in the right paracentral community (Desikan-Killiany atlas regions: R
precentral, R postcentral). In addition to the two nodes of the pre- and postcentral sulcus in the
alpha RPC community, the beta band RPC community also contained regions of the medial
paracentral lobule, putative sources of motor-related planning (Desikan-Killiany atlas regions:
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Communities derived from the interregional allegiance matrix in the pre-TMS interval
for the alpha and beta bands. Inflated mesh visualizations of brain regions colored by community
organization. Orbs are plotted at the centroid of the regions of interest. Community organization
was found independently for the alpha band (left) and beta band (right) before stimulation with
TMS. Dotted lines surrounding nodes near medial portion of the brains indicate the only two nodes
unique to the different frequency bands.
Network Neuroscience
617
Network reconfigurations after TMS
R paracentral, R posterior cingulate). This RPC community in beta nicely aligns with previous
literature that implicates the beta band in motor-related activity (?), providing support that the
detected network communities captured frequency-specific effects.
Community Allegiance Differentiates Beta Band Communities From the Alpha Band
We next sought to characterize how stimulation influenced dynamic network reconfigurations
from the natural baseline resting state. We employed a metric of allegiance that captures
how often two nodes are present within the same community before and after stimulation.
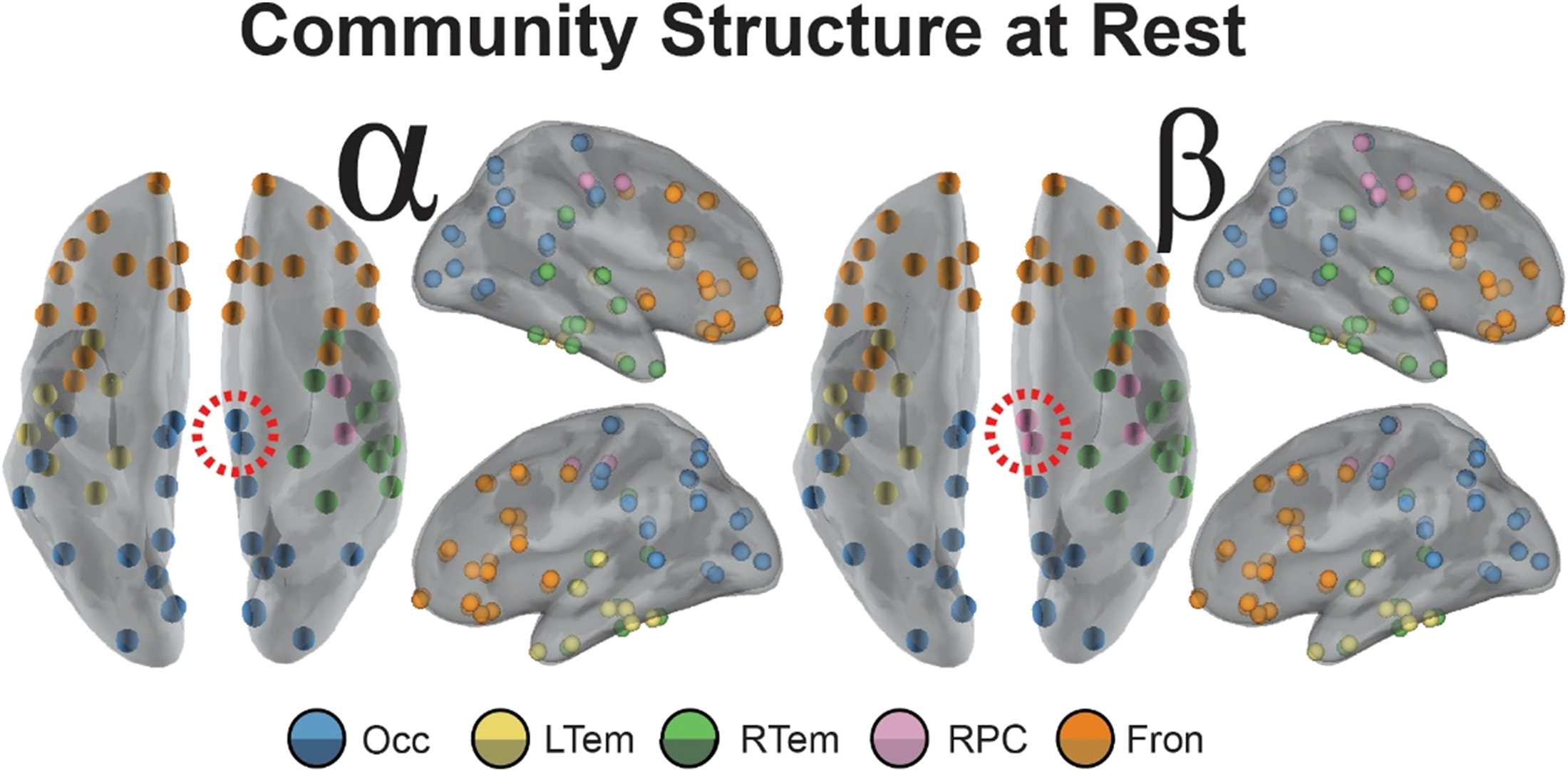
Figure 3 (A, C) shows the average pairwise difference in allegiance before and after stimula-
tion within each of the five communities identified from the pre-TMS resting-state connectivity
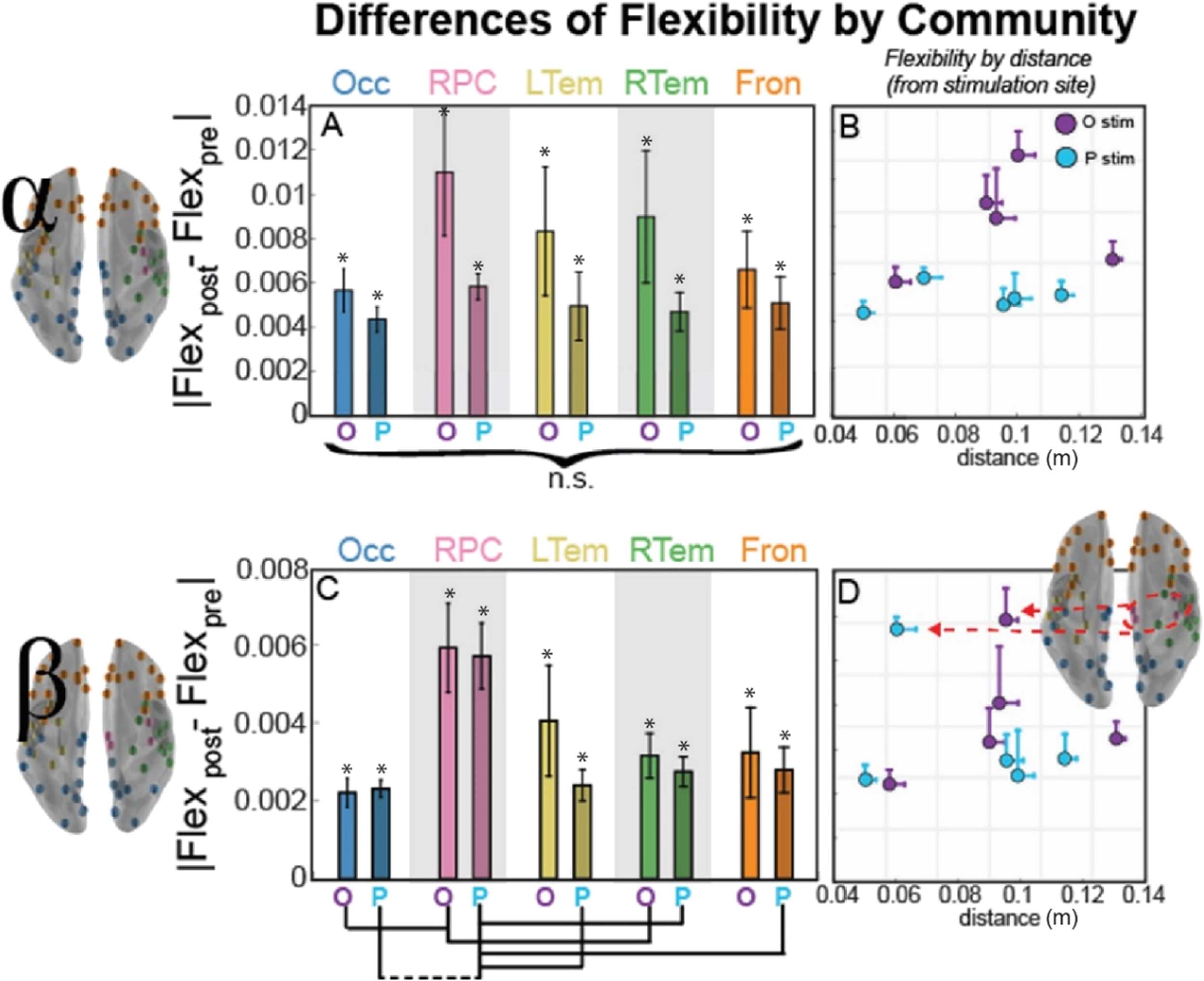
(Allegpost − Allegpre). Within the alpha band (top row), we observe some specificity to the stim-
ulation site. In comparing allegiance for each pairing of the communities, allegiance change
is highest for the occipital-parietal community (Occ) and lowest for the frontal community
(Fron; paired t test, t(9) = 3.8, p = 0.004, uncorrected), and this was true for each of the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Community allegiance changes within alpha (top) and beta (bottom) band network as a
function of distance from the stimulation site. (A, C) Bar plot of the mean magnitude change (SEM
across subjects) from the pre-TMS interval in pairwise allegiance from the stimulation site, with
the bar labeled O for occipital stimulation and P for parietal stimulation. For paired t test between
communities, dotted lines connecting communities indicate uncorrected significance (p < 0.05),
while solid lines indicate significance corrected for multiple comparisons (Bonferroni, p < 0.05).
(B, D) Scatter visualization of the mean magnitude allegiance change from the pre-TMS interval
shown in Panels A and C, but now plotted as a function of distance from the stimulation site. Error
bars indicate SEM across subjects (allegiance) or nodes within the community (distance), and the
color of the marker indicates stimulation site (occipital in purple and parietal in blue). Asterisk (*)
indicates a significant difference from 0, indicating a change from the pre-TMS interval (p < 0.05,
uncorrected). Brain inset for the beta band shows the nodes of the RPC community that are most
affected by TMS regardless of stimulation site.
Network Neuroscience
618
Network reconfigurations after TMS
subjects within our sample (see Supplemental Figure 6 in the Supporting Information to view
more about robustness of effects across subjects). The beta band (bottom row), however, shows
clear community specificity, where allegiance of the right paracentral (RPC) community is
significantly higher than the right temporal (RTem) and frontal (Fron) communities following
stimulation (paired t tests; RTem, t(9) = −2.6, p = 0.028; Fron, t(9) = −2.8, p = 0.020,
all uncorrected). To speak to robustness, the RPC showed the highest allegiance in 80% and
60% of subjects for occipital and parietal stimulation, respectively. Also, nearly each aver-
age change is significantly different from the pre-TMS resting-state estimate (labeled with * in
Figure 3A) with the exception of parietal stimulation effects in LTem and Fron.
We also examined whether the community allegiance depended on distance from the stim-
ulation site, which we operationalized as the Euclidean distance from the centroid of the com-
munity to the node closest to the stimulation site, estimated in meters from a reconstructed
3D mesh. The effects of stimulation within the alpha band revealed that the nodes closest to
the stimulation site are most susceptible to stimulation, and as Figure 3A shows, this effect is
reduced for the communities further from the stimulation site (see Supplemental Figure 7 in
the Supporting Information for a nonparametric correlational analysis with distance and the
graph metrics).
In contrast, the RPC community in the beta band was impacted more strongly by stim-
ulation with high specificity (Figure 3D, pink RPC nodes) by comparison to the other com-
munities. Thus, for the beta band, stimulation didn’t follow a simple rule based on distance
from the stimulation site as observed in alpha; instead, the stimulation effect was strongest in
the RPC community, suggestive of preferred resonant frequencies within the region targeted
frequencies account
(Rosanova et al., 2009).
of stimulation based on the strong role that beta band serves in motor-related activity
(Pfurtscheller et al., 1996b), and the prevalence of motor regions within the RPC commu-
nity. These results also indicate the importance of considering pairwise regional activity within
a community, so we next examined a network measure of flexibility to investigate regional
dynamics.
This observation aligns with the natural
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Flexibility Differences Indicate Whole-Brain Effects Within the Alpha Band
As a complement to allegiance, which measures the temporal consistency of community struc-
ture at the interregional level, we investigated flexibility, a metric that describes how often each
node changes the community to which it is allied. This analysis captures whether stimulation
drives certain brain regions to cohere with different communities in a manner that is different
from their community participation prior to stimulation (i.e., network reconfigurations).
Figure 4 shows the differences in flexibility, averaged across nodes within a community,
before and after stimulation (Flexpost − Flexpre). First, we compared flexibility in these com-
munities to 0, or no difference between Flexpost and Flexpre. We see that all communities have
a significant change in flexibility in both the alpha and the beta communities, suggesting a ro-
bust change in flexibility after stimulation reflecting the causal role of TMS pulses on dynamic
network reconfigurations. Also, overall, we see a large difference in effect size for the different
frequency bands, with alpha communities showing more flexibility overall.
We next compared each community pair to understand the specificity of these effects.
Within the alpha band (top row), we observed minimal difference between communities or
stimulation sites; rather, TMS is associated with a statistically equivalent change in flexibility
across communities. For the beta band (bottom row), a single community stands out. The RPC
community is again the most flexible following TMS. For occipital stimulation, flexibility of the
right paracentral (RPC) community is significantly higher than the occipital-parietal (Occ) and
Network Neuroscience
619
Network reconfigurations after TMS
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Community flexibility changes within alpha (top) and beta (bottom) band network as
a function of distance from the stimulation site.
(A, C) Bar plot of the mean magnitude change
(SEM across subjects) from the pre-TMS interval in flexibility as a function of the distance from
the stimulation site, with the bar labeled O for occipital stimulation and P for parietal stimulation.
For paired t test between communities, dotted lines connecting communities indicate uncorrected
significance (p < 0.05), while solid lines indicate significance corrected for multiple comparisons
(Bonferroni, p < 0.05). (B, D) Scatterplot of the mean change in flexibility from the pre-TMS interval
shown in Panels A and C, but now plotted as a function of distance from the stimulation site. Error
bars indicate SEM across subjects (flexibility) or nodes within the community (distance), and the
color of the marker indicates stimulation site (occipital in purple and parietal in blue). Asterisk (*)
indicates a significant difference from 0, indicating a change from the pre-TMS interval (p < 0.05,
uncorrected). Brain inset shows the nodes of the RPC community that are most affected by TMS
regardless of stimulation site.
trending for right temporal (RTem) communities (paired t tests; Occ, t(9) = −3.0, p = 0.016,
uncorrected; RTem, t(9) = −2.2, p = 0.057, uncorrected). This difference is even stronger for
parietal stimulation, where flexibility for RPC tends to be higher than that observed in the left
temporal (LTem), right temporal (RTem), frontal (Fron), and significant (Bonferroni corrected)
when compared with occipital-parietal (Occ) communities (paired t tests; LTem, t(9) = −3.1,
p = 0.013; RTem, t(9) = −2.6, p = 0.030; Fron, t(9) = −3.1, p = 0.014; Occ, t(9) = −4.0,
p = 0.003, all uncorrected). These results are reminiscent of the pairwise allegiance difference
showing an increase within a single community (Figure 5C).
We next examined whether flexibility depended on distance from the stimulation site. Al-
pha communities showed minimal dependence between distance and flexibility (Figure 4B),
but there was no significant difference across any of the communities (Figure 4A). In contrast,
when we considered flexibility within the beta band, we observed that the RPC community
Network Neuroscience
620
Network reconfigurations after TMS
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
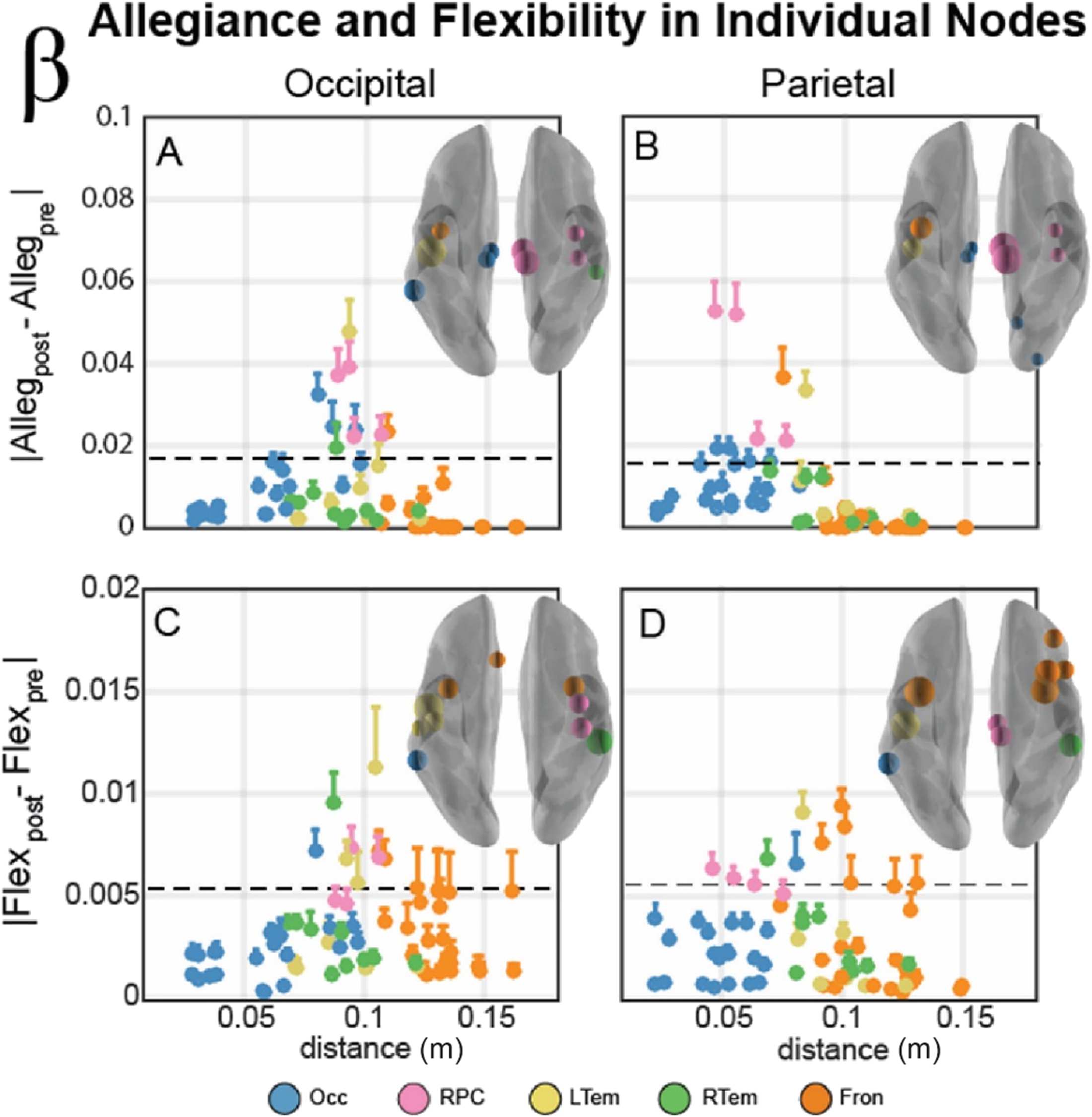
Figure 5.
Individual node allegiance (A, B) and flexibility (C, D) changes within the beta band
network. (A, B) Individual nodes module allegiance difference (Allegpost − Allegpre) plotted as a
function of distance from the stimulation site for occipital (A) and parietal (B) stimulation sites. Color
of node describes the community affiliation, and error bars indicate the standard error of the mean.
Brain insets display the 85th percentile of module allegiance across all nodes, with nodes scaled
by the relative magnitude of this allegiance change. The absolute magnitude of this percentile is
also indicated by a horizontal dotted line in each plot. (C, D) Individual nodes flexibility difference
(Flexpost − Flexpre) plotted as a function of distance from the stimulation site for occipital (A) and
parietal (B) stimulation sites. Color of nodes, error bars, and brain inset display the same properties
as above, but in this panel with flexibility rather than allegiance.
displayed the strongest effect of stimulation (Figure 4D). Combined, these flexibility results
demonstrate consistency with the allegiance results, suggesting an effect of resonant frequency
in the RPC community for the beta band. However, the communities have variable num-
bers of nodes, and a few nodes could substantially influence the means shown in Figures 4
and 5, so our final analysis examined individual node dynamics to determine whether the
smaller size of the RPC community could be the primary driver of beta band effects.
Individual Node Clusters Suggest a Reconfiguration of the Beta Band Network After TMS
Since all of the previous analyses examined only the overall community differences, our final
analysis examined the individual node allegiance to the stimulation site and flexibility changes
after stimulation. This analysis examines the spatial specificity of the TMS modification of the
Network Neuroscience
621
Network reconfigurations after TMS
graph metrics that may be masked by averaging across many nodes within the affiliated com-
munity. Figure 5 displays individual node magnitude allegiance (top row) and flexibility (bot-
tom row) differences in the five communities of the beta band network following stimulation
to occipital (left) and parietal (right) cortex. Overall, there is a change in allegiance within a
narrow range of distances from the stimulation site when considering both occipital (Figure 5A)
and parietal stimulation (Figure 5B). Although regions in the right paracentral community (RPC,
pink) show substantial change in allegiance, several nodes from other communities also have
a similar response profile. To examine the spatial topology of these effects, the brain insets
illustrate nodes corresponding to the top 15% of allegiance changes (nodes on or above the
threshold line in Figure 5A–B). The most influenced nodes surround the sensorimotor regions
of the brain, including the RPC community and nearby regions around the central sulcus, the
rolandic sites of the brain.
In contrast, the individual node flexibility changes are more diffuse (Figure 5, bottom row).
The nodes corresponding to the top 15% of flexibility changes are plotted above the threshold
line, and these effects span a larger range of distances than the allegiance changes. However,
the spatial topology is quite similar. Changes in flexibility after stimulation are strongest in a
cluster of nodes surrounding the central sulcus. Collectively, these results reveal that the nodes
in the RPC community were not uniquely influenced; instead, the network dynamics of the
RPC as well as the surrounding bilateral sensorimotor regions showed the largest flexibility
and allegiance changes within the beta band communities. Together these results suggest a
rapid reconfiguration of the resting beta community organization following TMS stimulation,
rather than enhancement of a single community.
DISCUSSION
We investigated network reconfiguration in resting-state EEG following single pulses of tran-
scranial magnetic stimulation (TMS) using a method from network science that allows for a
the brain’s modular architecture (Bassett & Bullmore, 2006;
quantitative description of
Bullmore & Sporns, 2012; Ercsey-Ravasz et al., 2013). Our analysis focused on connectivity
differences between the 1 s before stimulation and the 1 s after stimulation. More specifically,
we examined network differences in the alpha and beta frequency bands since these bands
have been suggested as resonant frequencies within the stimulated regions: alpha in occipital
and beta in parietal regions (Laufs et al., 2003; Rosanova et al., 2009).
Our results first examined whole-brain effects of stimulation, and we observed differen-
tial effects on connectivity according to stimulation site, although more subtle differentiation
was seen between the alpha and beta frequency bands as very similar data-driven network
structures were derived from each band. We employed a network theoretical approach to
identify communities from the resting-state EEG data, and importantly, we observed several
differences in the structure of functional connectivity in each frequency band following TMS.
Within the alpha band, stimulation produced local effects, but interestingly, also produced
more global effects as evidenced by modification of network flexibility across all occipital,
parietal, paracentral, and frontal communities when TMS was applied to either occipital or
parietal cortex. In stark contrast, beta band activity showed high specificity of TMS-induced
effects on allegiance and flexibility within a rather focal paracentral network near sensorimotor
cortex, regardless of stimulation site. These novel results using network science approaches
with TMS-EEG revealed an interesting interplay between local and global activity across fre-
quency bands that might underlie how network reconfigurations give rise to coordinated brain
activity.
Network Neuroscience
622
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
Global Effects Within the Alpha Band Manifest in Similar Flexibility Across Communities
We have shown that TMS to resonating communities constrained within the alpha band has
a similar impact on the overall flexibility of each community. This finding implies a global
impact of stimulation, regardless of stimulation site, to the alpha band networks. Since the first
observations of the alpha band by Berger (1929), alpha band activity (8–12 Hz), also known
as the Berger rhythm, has been a brain rhythm of frequent study because of its dominance in
resting EEG, and it is often the only visually observable pattern to the naked eye in the EEG
trace. Since its first observation, several hypotheses have been proposed ascribing a functional
role to its presence in EEG. The first theory was proposed by Adrian and Matthews (1934),
who found that the power within the alpha band increases when subjects are awake with eyes
closed. They interpreted this as alpha band activity reflecting a brain state of inactivity, prim-
ing the brain for incoming information. This theory has been expanded and revised to more
clearly represent “cortical idling” (Pfurtscheller, Stancák, & Neuper, 2006a). Recently, how-
ever, this theory has been somewhat abandoned because of the difficulty of reconciling it with
behavioral experiments that indicate a functional role for power within this band. For example,
alpha band activity is also associated with working memory load (Jensen, Gelfand, Kounios,
& Lisman, 2002; Klimesch, 1996; Tuladhar et al., 2007). Thus, beyond this “spontaneous”
alpha rhythm, researchers have discovered other forms of alpha, so-called functional alpha
(Ba¸sar, 2012), which is observed during many cognitive and motor processes. These theories
have been further broadened, suggesting that alpha activity may even be an access controller
to a knowledge system (Klimesch, 2012). Collectively, however, alpha activity consistently
represents a diffuse, communicative signal with multiple functions, an arguably global signal
in terms of its impact on sensory information.
Research using TMS to investigate the functional connectivity within the alpha band, how-
ever, provides a more limited view of alpha activity. For example, Rosanova et al. (2009)
showed that enhancement of the alpha band is primarily restricted to occipital cortex, regard-
less of stimulation site. These researchers noted that occipital cortex might even resonate at
the alpha frequency. Our results expand the theory proposed by Rosanova and colleagues by
showing that the brain may be parsed into separate resonating communities within the alpha
band and that each cluster is overall equally modified by TMS, as indicated by the similar
flexibility metrics across the resonating communities. Interestingly, however, nodal allegiance
to the stimulation site reveals a rather direct and localized impact within the occipital and
parietal cortex: specifically, the cluster that is the most spatially proximal to the stimulation
site and previously associated with alpha band activity (i.e., the bilateral occipital-parietal net-
work, Occ). Together, these results suggest a process whereby alpha connectivity can provide
both the specific visual effects shown in early studies while also serving many functional roles
across disparate brain regions.
Local Effects of Beta Band Manifest in the Specificity of Network Changes Within the Paracentral
Community
Two pieces of evidence in the current study converge to show that the paracentral network
plays a highly specific role in dynamic network reconfiguration within the beta band, and our
results support and extend the natural frequency theory of the brain following single pulses
of TMS (Rosanova et al., 2009). First, the network communities defined on resting-state ac-
tivity are identical between alpha and beta bands, except for the right paracentral network
(RPC). The beta band RPC consists of nodes within the paracentral lobule and two nodes
within the pre- and postcentral gyri that play a predominant role in sensorimotor processing,
consistent with previous research that has identified motor-related activity in the beta band
Network Neuroscience
623
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
(Pfurtscheller et al., 1996b). Second, following a single TMS pulse to either stimulation site,
we observe that the paracentral region uniquely displays both the highest flexibility and alle-
giance changes from the baseline state in the beta band, but not the alpha band. Together,
these findings suggest a unique specificity that would support the natural frequency theory of
TMS and are aligned with other network metrics suggesting beta band influence after parietal
stimulation (Amico et al., 2017). In fact, our results provide intuition at a level of granularity
that has not been previously explored, by capitalizing on recently developed methods from
network science (Bassett & Sporns, 2017) to investigate perturbations of brain networks fol-
lowing single pulses of TMS.
The granularity of this effect was further enhanced by inspection of the nodal allegiance and
flexibility across the brain. The RPC community was clearly involved and appeared to be an
isolated community with increased nodal allegiance (among its nodes) and global flexibility
in the beta band communities. However, when we inspected the single nodes contributing
to this effect, we found that this effect was not constrained by the boundaries of the beta
band RPC network as we defined from baseline activity. Instead, the effects spread to nodes
outside this RPC community and consisted of a cluster of nodes surrounding the central sulcus.
This granularity of the beta band network effects after TMS aligns well with the well-known
involvement of rolandic sites in sensorimotor processes and discharges of beta band activity
(Baker, 2007).
Clinical Implications
treating movement disorders
TMS has been used successfully in clinical settings for
(Pascual-Leone et al., 1994) and mitigating severe affective disorders (Berman et al., 2000);
however, some studies that have investigated the efficacy and efficiency of TMS treatment for
depression (for review, Loo & Mitchell, 2005) suggest that most treatment regimens are sub-
optimal, often stimulating for a duration of two weeks and having only a minor benefit. Here
we have shown a complex interplay between local and global neural processing, but more
generally, our results speak to the specificity of TMS, where particular resonating communities
of brain regions (e.g., beta band activity emanating from sensorimotor regions) or diffuse sets of
resonating communities (e.g., all regions in alpha band) may be modified by TMS regardless of
stimulation site. In other words, we show that stimulating a focal region can have distal effects
on many other brain regions. Future studies expanding on how individual variability in brain
connectivity impacts how TMS propagates through cortex may eventually reveal the specific
networks or brain regions that may predict successful treatment. We believe the methods and
initial results within this work hold promise in future studies to help determine stimulation
protocols for a variety of clinical settings and surrounding several cognitive domains.
Our results first examined whole-brain effects of stimulation, and we observed differential
effects on connectivity in both alpha and beta activity, although no stark differentiation was
seen between stimulation to occipital or parietal sites, globally. Next, we employed a network
theoretical approach to identify communities from the resting-state EEG data, and we observed
several differences in the structure of functional connectivity in both frequency bands after
TMS. Within the alpha band, stimulation produced local effects, but interestingly, stimulation
also produced more global effects, altering network flexibility across all communities when
applied to either occipital or parietal cortex.
Despite these general effects, our coarse-level results are merely the first step, as much
more must be completed to determine the robustness of much of these network dynamics that
may include any gender differences (70% of this sample were male), individual susceptibility
to stimulation, or state-based stimulation specificity (Thut et al., 2011), of which our current
Network Neuroscience
624
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
study does not tackle. We further expand on other methodological considerations that may
guide future studies within this domain in the following.
Methodological Considerations
Our use of community detection to understand functional connectivity in the brain and the
effects of TMS on specific brain networks focuses on two stimulation sites and two common fre-
quency bands. We use a phase-based undirected connectivity measurement and inspect graph
metric changes at a single snapshot of the available parameters within the dynamic modularity
framework we applied. This initial implementation of a graph theoretical approach on human
neurostimulation effects may be expanded in the future to investigate the directed commu-
nication between these resonating communities (e.g., Reimann et al., 2017), cross-frequency
communication (Canolty & Knight, 2010), and increased resolution across frequencies of the
brain. The methodological choices within this work also focused on merely one spatiotempo-
ral scale, which may not completely account for the spectral sensitivity across the regions of
the brain, and the results target a wakeful resting state in individuals, and it may not extend to
the active or sleeping brain (Hasson, Nusbaum, & Small, 2009; Massimini et al., 2005). Future
research may extend this work to take into account the spectral macroarchitecture of evoked
and induced oscillations in the brain.
Our experimental design did not employ neuronavigation or a control stimulation site;
instead, our participants completed four experimental sessions across four different days to
maximize the variability in the prestimulation period to identify stimulation effects robust to
state differences (boredom, fatigue, mind wandering, etc.). However, an interesting avenue
for future work would be to examine network changes that may be more closely tied to func-
tionally localized regions, where neuronavigation would serve a critical role in equating more
precise stimulation locations between individuals. Similarly, our analysis did not require a
control site since our investigation examined changes between baseline activity and activ-
ity following stimulation. The debate on how to “control” for neurostimulation techniques
has recently received increased attention. Research using simultaneous TMS-EEG studies
often have non-stimulation-related evoked activity (i.e., the auditory “click”; Conde et al.,
2019); however, there is also a debate on how the researchers implemented their controls
(Belardinelli et al., 2019). This debate is essential for studies that directly examine the neural
activity following stimulation; however, our design attempted to overcome the challenge of
non-stimulation-related evoked activity by comparing conditions where these nuisance sig-
nals are nearly equated. Thus, our investigation examined changes between baseline activity
and activity following stimulation and presents an alternative to this debate within the context
of two stimulation sites. The analytic logic we employed in our analysis follows the classic
comparison logic between conditions in traditional neuroimaging analysis: Conditions are
designed to only manipulate the factor of interest, so looking at their difference eliminates all
of the concomitant neural processing that occurs but is tangential to condition comparison of
interest. Here, the analysis statistically examined differences from a baseline period but also
qualitatively between occipital and parietal stimulation, so each served as the other’s control
for nuisance signals that are concurrent but tangential to the stimulation effects. A similar logic
could apply to the experimental design decision to not include an explicit control for the au-
ditory click sound from the TMS pulse. Although participants wore ear plugs to mitigate the
sensory response in auditory cortex, the focus on relative differences between stimulation sites
should help eliminate the effect of the auditory response on the results since it is expected
that the sensory effect is equivalent across the conditions. Our results, for the connectivity
measurement, clearly differentiate the stimulation sites; in contrast, it should be noted that the
Network Neuroscience
625
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
network metrics rarely differentiate stimulation sites. The auditory click could still contribute
to the observed network effects, but given the stimulation site specificity of the connectivity
matrices, it is likely to be a small contributor. However, future research could examine any of
these inferences/assumptions in greater detail. In particular, investigations may use more re-
gions and intensities to augment our understanding about differences in ongoing activity with
more functionally determined stimulation protocols.
Conclusion
Using a recently developed network-based methodology applied to EEG, we have investi-
gated the reconfigurations of naturally resonating communities of brain regions. While the
alpha network reveals the dynamic interplay of local and global activity, communities within
the beta band revealed a remarkable specificity, displaying more local connectivity changes.
Particularly important next steps include linking these observations with emerging theories
of the impact of stimulation on distributed networks in the form of network control theory
(Gu et al., 2015), which has begun to offer insights into the brain’s preference for certain low-
energy states (Betzel et al., 2017), the role that brain topology plays on the energy required for
brain state transitions (Kim et al., in press; Tang & Bassett, in press), and the predicted impact
of stimulation on distal areas (Gu et al., in press; Muldoon et al., 2016). Efforts to ground TMS
studies like the one we report here in a mechanistic theory could have lasting consequences in
studies linking behavioral changes to neural oscillations or neurostimulation (Medaglia et al.,
in press), but may also impact future protocols for clinical purposes, providing a means to
reconfigure resonating clusters in the brain.
MATERIALS AND METHODS
Participants
Ten individuals (seven men, three women, aged 20–33, M = 23.8, SD = 4.8) participated in
the TMS-EEG experiment. All gave informed, written consent as approved by the University of
California, Irvine, Institutional Review Board.
TMS-EEG Data Collection
Data collection occurred in the TMS-EEG Laboratory in the Human Neuroscience Lab at the
University of California, Irvine. The subjects were seated in a comfortable chair approximately
60 cm from the monitor, equipped with earplugs to attenuate the sound of the coil discharge,
and their heads were fixed in a chin rest to minimize movement while they continuously
fixated on the center of the monitor screen. No overt motor responses were required during
the 30-min experimental session.
Stimulation was applied with a MagStim Model 200 Monophasic Stimulator P/N 3010-00
equipped with a figure-of-eight coil at 55% stimulator intensity (E-field 297 V/m). We estab-
lished this intensity in a previous study that had a similar protocol (Garcia et al., 2011) by
systematically modulating the intensity threshold since motor thresholds are inappropriate for
occipital stimulation (Stewart, Walsh, & Rothwell, 2001). We found that only 37% of partici-
pants saw a phosphene at approximately 70%. Thus, we set the stimulation intensity for this
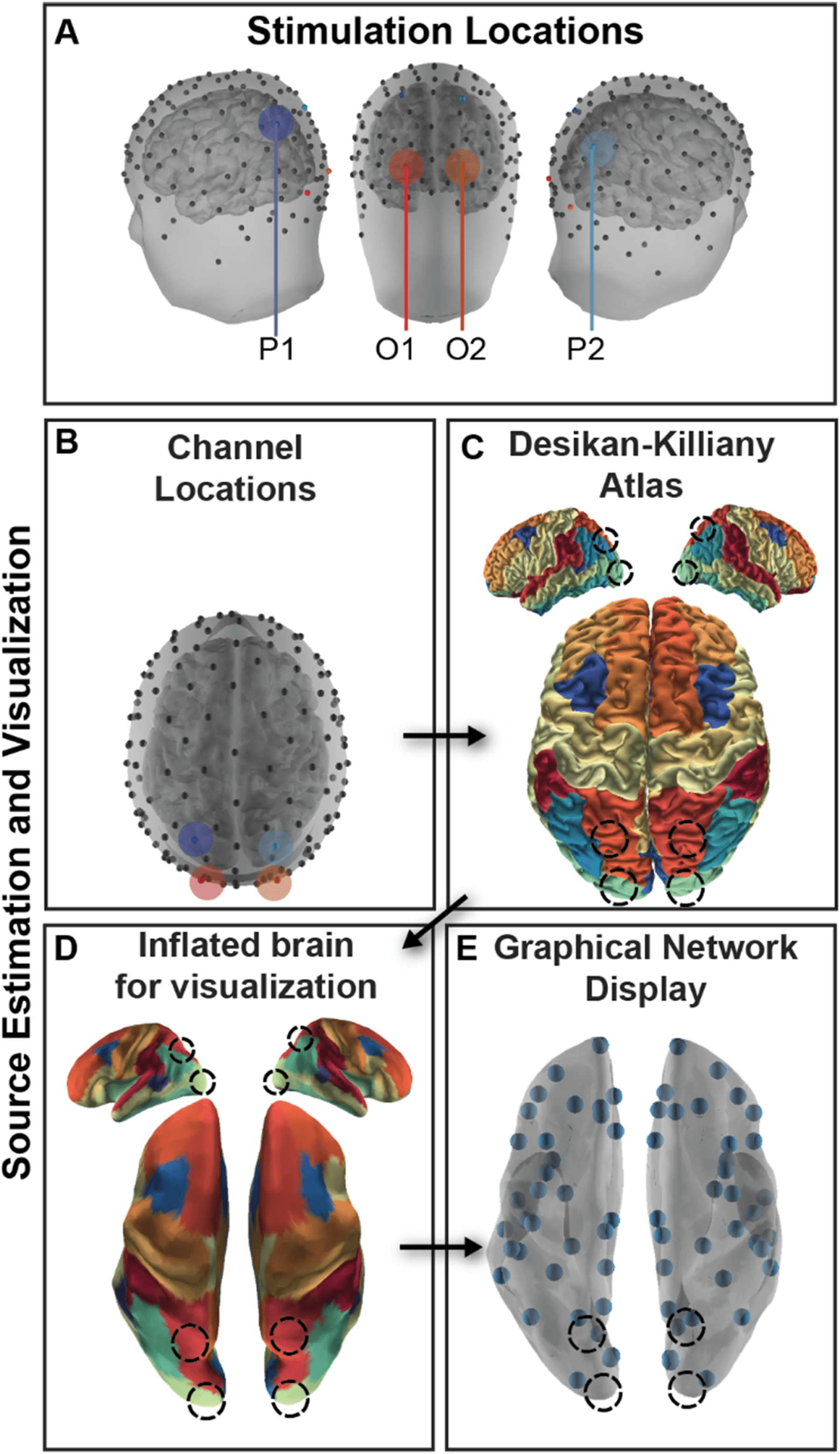
study to be at 55% of stimulation (20% less than the phosphene induction threshold). We tar-
geted four regions that included symmetric areas in occipital and parietal cortices (Figure 6),
and the location for these regions was estimated by electrodes O1/O2 and P1/P2 of the 10-20
international scheme for EEG. This method reliably targets a similar area across participants
within 2 cm of sulcal/gyral landmarks (Herwig, Satrapi, & Schönfeldt-Lecuona, 2003), the
Network Neuroscience
626
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(A) Participants received stimulation in symmetric
Figure 6.
Experimental design and analysis.
regions in occipital (O1, O2) and parietal (P1, P2) cortex.
(B) High-density EEG recorded from
128 channels was submitted to a cLORETA source analysis, and current source density (CSD) was
estimated for each vertex of a high-resolution mesh. (C) CSD was then averaged within a parcellation
of cortex following the Desikan-Killiany atlas parcellation to estimate regional brain activity. (D)
The cortex was inflated for visualization. (E) Each centroid of the region is plotted as a small orb.
Stimulation locations marked in each visualization with a shaded region or a dotted line.
resolution of high-density EEG used in this study. For occipital stimulation, the coil was ori-
ented parallel to the coronal view (i.e., paddle pointed up), and for parietal stimulation, the coil
was oriented approximately tangential to the curvature of the scalp at the electrode target lo-
cations (i.e., paddle pointed back and down). For each region, participants completed a block
of approximately 100 single pulses of stimulation that were no closer than 4 s apart (jittered up
to 6 s apart), following standard safety procedures (Rossi, Hallett, Rossini, & Pascual-Leone,
2009). Within a session, the 10 blocks were semi-randomly selected among the four stimulation
Network Neuroscience
627
Network reconfigurations after TMS
sites (O1, O2, P1, P2), ensuring that multiple blocks of each stimulation type occurred in each
session. Participants complete four sessions (one subject participated in five) on separate days,
providing an aggregate of approximately 4,000 total stimulation trials for each subject; how-
ever, one subject completed an additional session and another subject terminated one session
because of fatigue.
Throughout the session, 128 channels of EEG were recorded using a TMS-compatible EEG
system from Advanced Neuro Technology (ANT, The Netherlands). The Waveguard cap sys-
tem consisted of small Ag/AgCl electrode elements, specially designed to yield high-quality
and stable recordings with simultaneous TMS. EEG data were sampled at 1024 Hz for this
experiment and impedances were targeted to be kept below 10 kΩ.
EEG Analysis
The EEG data were preprocessed following an established pipeline
Segmentation of epochs.
(Garcia et al., 2011) for each of the four sessions for each of the 10 participants. First, raw EEG
were submitted to a principal component analysis (PCA) to identify the peak amplitude events
that correspond to the stimulation event and to eliminate any timing discrepancy between the
intended and actual timing of the stimulation pulse in the EEG data. After the timing of each
pulse was recovered, the decomposition was discarded and was not used in any subsequent
analysis (i.e., no components were removed). Using the maxima from the largest component
of each dataset as the onset of stimulation, the collected datasets were segmented into approx-
imately 40,000 epochs that were 6 s in duration, including 3 s (3,072 samples) before and after
the TMS pulse. After this segmentation procedure, the four samples before the pulse and 16
(15.6 ms) after the pulse were removed from the epochs to remove artifacts from the amplifier.
These samples were later replaced with a forward autoregressive moving average prediction of
the contaminated data from the intervals directly preceding the TMS pulse and a mild Savitzky-
Golay smoothing filter over the interval surrounding the pulse to remove any quick shifts in
amplitude due to the artifact editing procedure. Finally, each trial was normalized by the stan-
dard deviation of the 512 samples prior to the stimulation so that each subject contributed
similarly to grand mean TMS-evoked potentials (TEPs); however, this did not have an impact
on the overall shape of the TEP (see Supporting Information Figures). Because of the known
multiple sources of artifacts in work using simultaneous TMS-EEG (Rogasch & Fitzgerald, 2013)
beyond the amplifier artifacts within the 20 samples (19.5 ms), aggressive means were used to
ensure artifact was eliminated from our estimates. Automated artifact editing based on ampli-
tude thresholds was used. The thresholding procedure used a percentile rank of epochs within
the data. Ranks were calculated based on the maximum fluctuation found in the raw EEG re-
sponse on each trial for each electrode. In turn, trials were discarded if the maximum response
in 15% of electrodes was greater than the 95th percentile compared with all other trials. This
eliminated approximately 30% of trials likely contaminated with artifacts from the TMS pulse
or blink artifacts, leaving the following trial total for analysis in the participants: 3,129, 1,768,
3,535, 4,021, 2,269, 2,308, 2,931, 2,032, 2,840, and 3,088. After this preliminary prepro-
cessing, we largely followed the PREP approach, an artifact-removal procedure that has been
shown to be robust to artifacts within high-artifact environments (Bigdely-Shamlo, Mullen,
Kothe, Su, & Robbins, 2015). The following steps were completed: (a) line noise removal via a
frequency-domain (multitaper) regression technique to remove 60 Hz and harmonics present
in the signal; (b) a robust average reference with a Huber mean; (c) artifact subspace recon-
struction to remove residual artifacts with the standard deviation cutoff parameter set to 15;
and (d) band-pass filtering using a Butterworth filter with 2-dB attenuation at 2 and 50 Hz. TEPs
may be seen in Supplemental Figures 2, 4, and 5 in the Supporting Information, displaying how
the preprocessing pipeline has successfully removed these data.
Network Neuroscience
628
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
Distributed source reconstruction:
The method by which we may
estimate neural activity from specific
locations in the brain but measured
from the scalp.
Dynamic community detection:
A process that distills complex
connectivity matrices into a series of
cluster labels related to how nodes
coalesce into a network and change
across time.
From the preprocessed EEG data, we estimated current
Distributed source reconstruction.
source density (CSD) over a 5,003-vertex cortical mesh. A boundary element method (BEM)
forward model was derived from the Colin 27 anatomy (Holmes et al., 1998) and transformed
into MNI305 space (Evans et al., 1993) using standard electrode positions fit to the Colin 27
head surface in Brainstorm (Tadel, Baillet, Mosher, Pantazis, & Leahy, 2011). The BEM solution
was computed using OpenMEEG (Gramfort, Papadopoulo, Olivi, & Clerc, 2010; Kybic et al.,
2005), and the cLORETA approach was used for inverse modeling as described in detail in
Mullen et al. (2015) and implemented in the BCILAB (Kothe & Makeig, 2013) and Source In-
formation Flow (Mullen, 2014) toolboxes. Then CSD was averaged into one time course from
each of the 68 regions of the Desikan-Killiany atlas (Desikan et al., 2006) and downsampled to
128 Hz for use in the connectivity analysis. As a final step after parcellation, the surface mesh
of the Colin 27 brain (with DK atlas parcellated regions) was imported into Matlab and dis-
tance was estimated between regions closest to the stimulation site (bilateral lateral occipital
and superior parietal) and every other region in the DK atlas using pdist.m in Matlab. For com-
munity “distance,” the average distance from the stimulation site was estimated, corresponding
to centroid distance of the community.
Functional Connectivity Analysis
To estimate functional connectivity of brain networks before and after stimulation, an undi-
rected measure, known as the debiased weighted phase lag index (dwPLI), was computed
between each pair of the 68 estimated brain regions from the atlas. DwPLI is robust against
the influence of volume conduction, uncorrelated noise, and intersubject variations in sample
size (Vinck et al., 2011; Vindiola et al., 2014), and it has previously been proposed to be an
appropriate pairing with a source localization analysis to minimize the influence of these nui-
sance variables (Hillebrand, Barnes, Bosboom, Berendse, & Stam, 2012; van Diessen et al.,
2015).
The connectivity estimates were calculated using Matlab and FieldTrip (Oostenveld, Fries,
Maris, & Schoffelen, 2010). First, a multitaper spectral estimation was applied to the CSD
measurements, and then dwPLI was computed between all source pairs to estimate the func-
tional connectivity pattern of signals in the frequency range between 2 Hz and 25 Hz (step
= 0.5 Hz) in a 5-s window centered on the stimulation pulse (−2.5 s to 2.5 s). The dwPLI
connectivity estimates were calculated across trials, representing the trial-by-trial consistency
between regional CSD. Next the matrix of dwPLI estimates was reduced to the 51 time win-
dows corresponding to windows centered 38 ms apart, from 1 s before the TMS pulse to 1 s
after the TMS pulse. Since this estimate was done using a Hanning windowing method, the
windows are not independent and represent some smearing in time.
Dynamic Community Detection
In addition to looking at whole-brain connectivity patterns, we employed a community de-
tection algorithm (Bassett & Bullmore, 2006; Bullmore & Sporns, 2012; Ercsey-Ravasz et al.,
2013) to examine whether regions formed modular networks, and whether the regional com-
position of these networks changed before and after stimulation across the 51 time windows
in our 2-s epoch. The algorithm optimizes a multilayer modularity quality function, Q, us-
ing a Louvain-like greedy algorithm (Blondel et al., 2008; Mucha et al., 2010) to assign brain
regions to communities. The community assignments are dependent on two parameters: (a) a
structural resolution γ parameter and (b) a temporal resolution ω parameter. These two pa-
rameters determine the scale of the resulting graph, both structurally and temporally, and here,
Network Neuroscience
629
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
we sweep this parameter space to find the scale of the data that is most unlike that expected
in an appropriate random network null model. As described in Garcia et al. (2018), there are
several heuristics we may use to determine the optimal parameter for our dataset. We chose
an unbiased “difference” heuristic because of the unique properties of this stimulation dataset,
which we explain below.
Following our previous work on fMRI data (Bassett et al., 2013), the values for both pa-
rameters were determined by comparing the mean value of Q in the experimental data to the
mean value of Q in a shuffled null model of the data; we tested a very wide range of values
for each parameter since this algorithm has not yet been applied to EEG data, which have in-
herently different temporal and spatial scales of functional connectivity (Nunez & Srinivasan,
2006). Our analysis examined parameters for γ = 0.8 to γ = 1.6 and ω = 0.5 to ω = 35. The
null model of the data was created by randomly shuffling the pairwise dwPLI values, destroy-
ing the correlational structure observed in EEG data for each subject and parameter pairing.
Each Q was then subtracted for each parameter pairing, comparing the observed model’s Q
(from the unperturbed EEG connectivity patterns) and the null model’s Qnull (shuffled con-
nectivity patterns) for each subject. Our analysis found a clear peak in the resulting Q matrix,
suggesting that the range used was appropriate for this dataset. In fact, the largest difference
was found for γ = 1.025 and ω = 9, and these parameters were used in the reported analy-
ses, which suggests that the temporal parameter (ω) is the parameter that captures the unique
properties of the EEG signal. Since the community detection algorithm is nondeterministic
(Good, de Montjoye, & Clauset, 2010), 100 iterations of the hard partitions were estimated
with modularity maximization for each subject and stimulation condition (O1, O2, P1, P2),
yielding 100 sets of community labels for the 68 nodes for each of the four stimulation condi-
tions for each of the 10 subjects.
Community metrics. Within each of the 51 time windows of our 2-s stimulation epoch,
we examined the relationship among the brain regions within a community to characterize
the dynamic reconfiguration of spatially distributed neural sources before and after stimula-
tion. Our analysis investigated two community metrics, flexibility (Bassett et al., 2011) and
allegiance (Bassett et al., 2015). Figure 7 is a visual depiction of the steps needed to estimate
these community metrics.
The flexibility of each node corresponds to the number of instances in which a node changes
community affiliation, g, normalized by the total possible number of changes that could occur
across the layers L (Bassett et al., 2011), which represents each time slice within this dynamic
community detection algorithm. In other words, the flexibility of a single node i, ξi, may be
estimated with the following:
ξi =
gi
L − 1
,
(1)
where L is the total number of temporal windows.
Allegiance estimates how much regions communicate with subnetworks in the community
structure and demonstrate the same pattern of connectivity across time points. We define
allegiance matrix P, where edge weight Pij denotes the number of times a pair of nodes moves
to the same community together divided by L − 1 possible changes.
Thus, allegiance increases the resolution of community and captures coordinated activity
of each node with every other node in the brain, whereas flexibility examines whether a brain
region changes affiliations overall.
Each of these measures was calculated twice, once for the 25 windows of partitions before
TMS (pre-TMS) and once for the 25 windows following TMS (post-TMS), ignoring the cen-
ter window where stimulation occurred. Our analysis focused on the absolute difference in
Network Neuroscience
630
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Overview of analysis method and natural brain architecture. (A) DwPLI was estimated
across trials in 40-ms windows across the 2-s epoch, including 1 s before and 1 s after the TMS
onset (P: posterior, A: anterior nodes). (B) Dynamic network communities from a sample subject
derived from the dwPLI estimate for each window. Each color marks a different community label.
(C, left) From Panel B, community metrics were calculated that represent how often a node changes
over time (flexibility) or how often each node pair is in the same community over time (allegiance).
The cartoon networks show five hypothetical nodes that change communities over time (three time
windows shown). Connections are marked as black lines and metrics are given in the final column,
showing a range of flexibility and allegiance values. These metrics are used in subsequent analyses.
(C, right) Summary allegiance matrix for a sample subject for the period before TMS onset, indicating
the natural architecture of connectivity within the alpha band.
allegiance and flexibility between pre-TMS and post-TMS communities, emphasizing whether
stimulation influences local or global brain dynamics more strongly.
Finally, results were averaged across left and right stimulation sites since we were inter-
ested in the general magnitude of changes from functionally similar regions.
In support of
this data reduction primarily driven by our broad interest in coarse parietal/occipital stimula-
tion differences, follow-on analyses did not show any differences in consistency of network
changes between left and right stimulation, using a temporal consensus method inspired by
Doron, Bassett, & Gazzaniga (2012) in Supplemental Figure 8 (see the Supporting Information).
Statistical Analyses
To find the substantial changes in metrics across the pre-TMS and post-TMS intervals, tradi-
tional linear statistics were used, where the pre-TMS and post-TMS intervals were treated as
conditions and paired-sample t tests were applied to node or communities or a one-sample
Network Neuroscience
631
Network reconfigurations after TMS
t test of differences (for the dwPLI comparison) from 0, as indicated in the text. In cases where
multiple comparisons were carried out (e.g., Figures 1 and 3), a Bonferroni correction or false
discovery rate was used to determine significance. For the Bonferroni correction, each band-
specific metric was treated as a separate set of tests (10 comparisons within a set, Community
1 vs. Community 2, Community 1 vs. Community 3, etc.), so the corrected alpha value was
set to 0.005. For the false discovery rate, p values were adjusted within each dwPLI matrix
(Figure 1), and as reported q < 0.05 was used. Where appropriate, both the corrected and
uncorrected significant comparisons are shown (see Figures 3 and 4).
ACKNOWLEDGMENTS
The authors acknowledge thoughtful discussions with our colleagues at the University of Cal-
ifornia, Irvine, for study coordination and subject testing. This research was supported by
mission funding to the Army Research Laboratory as well as sponsored by the Army Research
Laboratory and accomplished under Cooperative Agreement Number W911NF-10-2-0022.
We would also like to acknowledge that the work was partially collected while J. O. G. was
funded by a National Research Service Award (F31-EY-019241) awarded by the National Insti-
tutes of Health. The views and conclusions contained in this document are those of the authors
and should not be interpreted as representing the official policies, either expressed or implied,
of the Army Research Laboratory or the U.S. Government.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00139.
AUTHOR CONTRIBUTIONS
Javier Omar Garcia: Conceptualization; Formal analysis; Methodology; Visualization; Writing-
Original Draft; Writing - Review & Editing. Arian Ashourvan: Formal analysis; Methodology;
Writing - Review & Editing. Steven M. Thurman: Validation; Writing - Original Draft. Ramesh
Srinivasan: Conceptualization; Data curation; Methodology; Resources; Writing - Review &
Editing. Danielle Bassett: Conceptualization; Formal analysis; Methodology; Writing - Original
Jean Vettel: Conceptualization; Methodology; Writing -
Draft; Writing - Review & Editing.
Original Draft; Writing - Review & Editing.
FUNDING INFORMATION
Army Research Laboratory (http://dx.doi.org/10.13039/100006754), Award ID: W911NF-10-
2-0022.
REFERENCES
Achard, S., Salvador, R., Whitcher, B., Suckling, J., & Bullmore, E.
(2006). A resilient, low-frequency, small-world human brain func-
tional network with highly connected association cortical hubs.
Journal of Neuroscience, 26, 63–72. https://doi.org/10.1523/
jneurosci.3874-05.2006
Adrian, E. D., & Matthews, B. H. C.
(1934). The interpretation
of potential waves in the cortex. Journal of Physiology, 81,
440–471.
Alivisatos, A. P., Chun, M., Church, G. M., Greenspan, R. J., Roukes,
M. L., & Yuste, R. (2012). The Brain Activity Map Project and the
challenge of functional connectomics. Neuron, 74, 970–974.
https://doi.org/10.1016/j.neuron.2012.06.006
Amico, E., Bodart, O., Rosanova, M., Gosseries, O., Heine, L.,
Van Mierlo, P., . . . , Laureys, S.
(2017). Tracking dynamic
interactions between structural and functional connectivity: A
TMS/EEG-dMRI study. Brain Connectivity, 7, 84–97. https://doi.
org/10.1089/brain.2016.0462
Ashourvan, A., Gu, S., Mattar, M. G., Vettel, J. M., & Bassett, D. S.
(2017). The energy landscape underpinning module dynamics
in the human brain connectome. NeuroImage, 157, 364–380.
https://doi.org/10.1016/j.neuroimage.2017.05.067
Auriat, A. M., Neva, J. L., Peters, S., Ferris, J. K., & Boyd, L. A.
(2015). A review of transcranial magnetic stimulation and multi-
modal neuroimaging to characterize post-stroke neuroplasticity.
Network Neuroscience
632
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
Frontiers in Neurology, 6. https://doi.org/10.3389/fneur.2015.
00226
Azevedo, F. A. C., Carvalho, L. R. B., Grinberg, L. T., Farfel, J. M.,
Ferretti, R. E. L., Leite, R. E. P., . . . , Herculano-Houzel, S. (2009).
Equal numbers of neuronal and nonneuronal cells make the hu-
Journal of
man brain an isometrically scaled-up primate brain.
Comparative Neurology, 513, 532–541. https://doi.org/10.1002/
cne.21974
Baker, S. N. (2007). Oscillatory interactions between sensorimotor
cortex and the periphery. Current Opinion in Neurobiology, 17,
649–655. https://doi.org/10.1016/j.conb.2008.01.007
Ba¸sar, E. (2012). A review of alpha activity in integrative brain func-
tion: Fundamental physiology, sensory coding, cognition and
pathology. International Journal of Psychophysiology, 86, 1–24.
https://doi.org/10.1016/j.ijpsycho.2012.07.002
Ba¸sar, E., Ba¸sar-Ero˘glu, C., Karaka¸s, S., & Schürmann, M.
(1999).
Are cognitive processes manifested in event-related gamma, alpha,
theta and delta oscillations in the EEG? Neuroscience Letters,
259, 165–168. https://doi.org/10.1016/S0304-3940(98)00934-3.
Bassett, D. S., & Bullmore, E.
(2006). Small-world brain net-
works. Neuroscientist, 12, 512–523. https://doi.org/10.1177/
1073858406293182
Bassett, D. S., Porter, M. A., Wymbs, N. F., Grafton, S. T., Carlson,
J. M., & Mucha, P. J.
(2013). Robust detection of dynamic
community structure in networks. Chaos, 23, 013142. https://
doi.org/10.1063/1.4790830
Bassett, D. S., & Sporns, O. (2017). Network neuroscience. Nature
Neuroscience, 20, 353–364. https://doi.org/10.1038/nn.4502
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
(2011). Dynamic reconfiguration of hu-
J. M., & Grafton, S. T.
man brain networks during learning. Proceedings of the National
Academy of Sciences, 108, 7641–7646. https://doi.org/10.1073/
pnas.1018985108
Bassett, D. S., Yang, M., Wymbs, N. F., & Grafton, S. T.
(2015).
Learning-induced autonomy of sensorimotor systems. Nature
Neuroscience, 18, 744–751. https://doi.org/10.1038/nn.3993
Belardinelli, P., Biabani, M., Blumberger, D. M., Bortoletto, M.,
Casarotto, S., David, O., . . . Ilmoniemi, R. J.
(2019). Repro-
ducibility in TMS–EEG studies: A call for data sharing, standard
procedures and effective experimental control. Brain Stimula-
tion: Basic. Translational, and Clinical Research in Neuromodu-
lation, 12, 787–790. https://doi.org/10.1016/j.brs.2019.01.010
Benjamini, Y., & Yekutieli, D. (2001). The control of the false discov-
ery rate in multiple testing under dependency. Annals of Statistics,
29, 1165–1188. https://doi.org/10.1214/aos/1013699998
Berger, H.
(1929). Electroencephalogram in humans. Archiv für
Psychiatrie und Nervenkrankheiten, 87, 527–570. https://doi.
org/10.1007/BF01797193
Bergmann, T. O., Karabanov, A., Hartwigsen, G., Thielscher, A., &
Siebner, H. R. (2016). Combining non-invasive transcranial brain
stimulation with neuroimaging and electrophysiology: Current
approaches and future perspectives. NeuroImage, 140, 4–19.
https://doi.org/10.1016/j.neuroimage.2016.02.012
Berman, R. M., Narasimhan, M., Sanacora, G., Miano, A. P.,
Hoffman, R. E., Hu, X. S., . . . Boutros, N. N. (2000). A random-
ized clinical trial of repetitive transcranial magnetic stimulation
in the treatment of major depression. Biological Psychiatry, 47,
332–337. https://doi.org/10.1016/S0006-3223(99)00243-7
Bestmann, S., Ruff, C. C., Blankenburg, F., Weiskopf, N., Driver, J.,
& Rothwell, J. C.
(2008). Mapping causal interregional influ-
ences with concurrent TMS–fMRI. Experimental Brain Research,
191, 383. https://doi.org/10.1007/s00221-008-1601-8
Betzel, R. F., Satterthwaite, T. D., Gold, J. I., & Bassett, D. S. (2017).
Positive affect, surprise, and fatigue are correlates of network
flexibility. Scientific Reports, 7, 520. https://doi.org/10.1038/
s41598-017-00425-z
Bigdely-Shamlo, N., Mullen, T., Kothe, C., Su, K.-M., & Robbins,
K. A. (2015). The PREP pipeline: Standardized preprocessing for
large-scale EEG analysis. Frontiers in Neuroinformatics, 9, 16.
https://doi.org/10.3389/fninf.2015.00016
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., & Lefebvre, E.
(2008). Fast unfolding of communities in large networks. Journal
of Statistical Mechanics, 2008, P10008. https://doi.org/10.1088/
1742-5468/2008/10/P10008
Bohning, D., Shastri, A., McConnell, K., Nahas, Z., Lorberbaum,
(1999). A combined TMS/
J., Roberts, D., . . . George, M. S.
fMRI study of
intensity-dependent TMS over motor cortex.
Biological Psychiatry, 45, 385–394. https://doi.org/10.1016/
S0006-3223(98)00368-0
Bortoletto, M., Veniero, D., Thut, G., & Miniussi, C.
(2015). The
contribution of TMS–EEG coregistration in the exploration of
the human cortical connectome. Neuroscience and Biobehav-
ioral Reviews, 49, 114–124. https://doi.org/10.1016/j.neubiorev.
2014.12.014
Braun, U., Schäfer, A., Walter, H., Erk, S., Romanczuk-Seiferth, N.,
Haddad, L., . . . Bassett,D. S. (2015). Dynamic reconfiguration of
frontal brain networks during executive cognition in humans. Pro-
ceedings of the National Academy of Sciences, 112, 11678–11683.
https://doi.org/10.1073/pnas.1422487112
Bressler, S. L., & Kelso, J. A. S.
(2001). Cortical coordination dy-
namics and cognition. Trends in Cognitive Sciences, 5, 26–36.
https://doi.org/10.1016/S1364-6613(00)01564-3
Bressler, S. L., & Menon, V. (2010). Large-scale brain networks in
cognition: Emerging methods and principles. Trends in Cogni-
tive Sciences, 14, 277–290. https://doi.org/10.1016/j.tics.2010.
04.004
Brunel, N., & Wang, X.-J. (2003). What determines the frequency
of fast network oscillations with irregular neural discharges? I.
Synaptic dynamics and excitation-inhibition balance. Journal of
Neurophysiology, 90, 415–430. https://doi.org/10.1152/jn.01095.
2002
Brunoni, A. R., & Vanderhasselt, M.-A. (2014). Working memory
improvement with non-invasive brain stimulation of the dorso-
lateral prefrontal cortex: A systematic review and meta-analysis.
Brain and Cognition, 86, 1–9. https://doi.org/10.1016/j.bandc.
2014.01.008
Bullmore, E., & Sporns, O.
(2012). The economy of brain net-
work organization. Nature Reviews Neuroscience, 13, 336–349.
https://doi.org/10.1038/nrn3214
Buzsáki, G. (2006). Rhythms of the brain. New York, NY: Oxford
University Press.
Network Neuroscience
633
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
Buzsáki, G., & Draguhn, A. (2004). Neuronal oscillations in corti-
cal networks. Science, 304, 1926–1929. https://doi.org/10.1126/
science.1099745
Buzsáki, G., & Wang, X. J.
(2012). Mechanisms of gamma oscil-
lations. Annual Review of Neuroscience, 35, 203–225. https://
doi.org/10.1146/annurev-neuro-062111-150444
Canolty, R. T., & Knight, R. T. (2010). The functional role of cross-
frequency coupling. Trends in Cognitive Sciences, 14, 506–515.
https://doi.org/10.1016/j.tics.2010.09.001
Cocchi, L., Gollo, L. L., Zalesky, A., & Breakspear, M. (2017). Criti-
cality in the brain: A synthesis of neurobiology, models and cog-
nition. Progress in Neurobiology, 158, 132–152. https://doi.org/
10.1016/j.pneurobio.2017.07.002
Cocchi, L., Sale, M. V., Lord, A., Zalesky, A., Breakspear, M., &
Mattingley, J. B. (2015). Dissociable effects of local inhibitory and
excitatory theta-burst stimulation on large-scale brain dynamics.
Journal of Neurophysiology, 113, 3375–3385. https://doi.org/10.
1152/jn.00850.2014
Conde, V., Tomasevic, L., Akopian, I., Stanek, K., Saturnino, G. B.,
Thielscher, A., . . . Siebner, H. R.
(2019). The non-transcranial
TMS-evoked potential is an inherent source of ambiguity in TMS-
EEG studies. NeuroImage, 185, 300–312. https://doi.org/10.
1016/j.neuroimage.2018.10.052
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson, B.
C., Blacker, D., . . . Killiany, R. J. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI scans
into gyral based regions of interest. NeuroImage, 31, 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021
Doron, K. W., Bassett, D. S., & Gazzaniga, M. S. (2012). Dynamic
network structure of interhemispheric coordination. Proceed-
ings of the National Academy of Sciences, 109, 18661–18668.
https://doi.org/10.1073/pnas.1216402109
Driver, J., Blankenburg, F., Bestmann, S., Vanduffel, W., & Ruff,
C. C. (2009). Concurrent brain-stimulation and neuroimaging for
studies of cognition. Trends in Cognitive Sciences, 13, 319–327.
https://doi.org/10.1016/j.tics.2009.04.007
Eldaief, M. C., Press, D. Z., & Pascual-Leone, A. (2013). Pascual-
leone, A Transcranial magnetic stimulation in neurology. Neu-
rology Clinical Practice, 3, 519–526. https://doi.org/10.1212/01.
CPJ.0000436213.11132.8e
Ercsey-Ravasz, M., Markov, N. T., Lamy, C., Van Essen, D. C.,
Knoblauch, K., Toroczkai, Z., & Kennedy, H.
(2013). A pre-
dictive network model of cerebral cortical connectivity based on
a distance rule. Neuron, 80, 184–197. https://doi.org/10.1016/
j.neuron.2013.07.036
Evans, A. C., Collins, D. L., Mills, S. R., Brown, E. D., Kelly, R. L., &
Peters, T. M. (1993). 3D statistical neuroanatomical models from
305 MRI volumes. 1993 IEEE Conference Record Nuclear Science
Symposium and Medical Imaging Conference, 3, 1813–1817.
https://doi.org/10.1109/NSSMIC.1993.373602
Fries, P., Nikoli´c, D., & Singer, W. (2007). The gamma cycle. Trends
in Neurosciences, 30, 309–316. https://doi.org/10.1016/j.tins.
2007.05.005
neural function. Proceedings of the IEEE, 106, 846–867. https://
doi.org/10.1109/JPROC.2017.2786710
Garcia, J. O., Grossman, E. D., & Srinivasan, R. (2011). Evoked po-
tentials in large-scale cortical networks elicited by TMS of the
Journal of Neurophysiology, 106, 1734–1746.
visual cortex.
https://doi.org/10.1152/jn.00739.2010
Geisler, C., Brunel, N., & Wang, X.-J.
(2005). Contributions of
intrinsic membrane dynamics to fast network oscillations with
Journal of Neurophysiology, 94,
irregular neuronal discharges.
4344–4361. https://doi.org/10.1152/jn.00510.2004
Gollo, L. L., Roberts, J. A., & Cocchi, L. (2017). Mapping how lo-
cal perturbations influence systems-level brain dynamics. Neuro-
Image, 160, 97–112. https://doi.org/10.1016/j.neuroimage.2017.
01.057
Good, B. H., de Montjoye, Y.-A., & Clauset, A.
(2010). Perfor-
mance of modularity maximization in practical contexts. Physi-
cal Review E, 81, 046106. https://doi.org/10.1103/PhysRevE.81.
046106
Gramfort, A., Papadopoulo, T., Olivi, E., & Clerc, M. (2010). Open-
MEEG: Opensource software for quasistatic bioelectromagnetics.
BioMedical Engineering OnLine, 9, 45. https://doi.org/10.1186/
1475-925X-9-45
Gu, S., Cieslak, M., Baird, B., Muldoon, S. F., Grafton, S. T.,
Pasqualetti, F., & Bassett, D. S. (in press). The energy landscape
of neurophysiological activity implicit in brain network structure.
NeuroImage. Retrieved from https://arxiv.org/abs/1607.01959
Gu, S., Pasqualetti, F., Cieslak, M., Telesford, Q. K., Yu, A. B., Kahn,
A. E., . . . Bassett, D. S. (2015). Controllability of structural brain
networks. Nature Communications, 6, ncomms9414. https://doi.
org/10.1038/ncomms9414
Hari, R. & Salmelin, R.
(1997). Human cortical oscillations: A
neuromagnetic view through the skull. Trends in Neurosciences,
20, 44–49. https://doi.org/10.1016/S0166-2236(96)10065-5
Hasson, U., Nusbaum, H. C., & Small, S. L. (2009). Task-dependent
organization of brain regions active during rest. Proceedings of
the National Academy of Sciences, 106, 10841–10846. https://
doi.org/10.1073/pnas.0903253106
Herring, J. D., Thut, G., Jensen, O., & Bergmann, T. O.
(2015).
Attention modulates TMS-locked alpha oscillations in the visual
cortex. Journal of Neuroscience, 35, 14435–14447. https://doi.
org/10.1523/JNEUROSCI.1833-15.2015
Herwig, U., Satrapi, P., & Schönfeldt-Lecuona, C.
(2003). Using
the international 10-20 EEG system for positioning of transcra-
nial magnetic stimulation. Brain Topography, 16(2), 95–99.
Hillebrand, A., Barnes, G. R., Bosboom, J. L., Berendse, H. W., &
Stam, C. J. (2012). Frequency-dependent functional connectivity
within resting-state networks: An atlas-based MEG beamformer
solution. NeuroImage, 59, 3909–3921. https://doi.org/10.1016/
j.neuroimage.2011.11.005
Holmes, C. J., Hoge, R., Collins, L., Woods, R., Toga, A. W., &
Evans, A. C. (1998). Enhancement of MR images using registra-
tion for signal averaging. Journal of Computer Assisted Tomogra-
phy, 22, 324–333.
Garcia, J. O., Ashourvan, A., Muldoon, S., Vettel, J. M., & Bassett,
D. S.
(2018). Applications of community detection techniques
to brain graphs: Algorithmic considerations and implications for
Ilmoniemi, R. J., Virtanen, J., Ruohonen, J., Karhu, J., Aronen,
H. J., Näätänen, R., & Katila, T. (1997). Neuronal responses to
magnetic stimulation reveal cortical reactivity and connectivity.
Network Neuroscience
634
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
Neuroreport, 8, 3537–3540. https://doi.org/10.1097/00001756-
199711100-00024
Jensen, O., Gelfand, J., Kounios, J., & Lisman, J. E. (2002). Oscil-
lations in the alpha band (9–12 Hz) increase with memory load
during retention in a short-term memory task. Cerebral Cortex,
12, 877–882.
Kim, J., Soffer, J. M., Kahn, A. E., Vettel, J. M., Pasqualetti, F., &
Bassett, D. S. (in press). Topological principles of control in dy-
namical network systems. Nature Physics. Retrieved from http://
arxiv.org/abs/1702.00354
Klimesch, W.
(1996). Memory processes, brain oscillations and
EEG synchronization. International Journal of Psychophysiology,
24, 61–100. https://doi.org/10.1016/S0167-8760(96)00057-8
Klimesch, W. (2012). Alpha-band oscillations, attention, and con-
trolled access to stored information. Trends in Cognitive Sciences,
16, 606–617. https://doi.org/10.1016/j.tics.2012.10.007
Kothe, C. A., & Makeig, S. (2013). BCILAB: A platform for brain-
computer interface development. Journal of Neural Engineering,
10, 056014. https://doi.org/10.1088/1741-2560/10/5/056014
Kybic, J., Clerc, M., Abboud, T., Faugeras, O., Keriven, R., &
Papadopoulo, T.
(2005). A common formalism for the integral
formulations of the forward EEG problem. IEEE Transactions on
Medical Imaging, 24, 12–28. https://doi.org/10.1109/TMI.2004.
837363
Laufs, H., Kleinschmidt, A., Beyerle, A., Eger, E., Salek-Haddadi,
(2003). EEG-correlated fMRI of human
A., Preibisch, C., et al.
alpha activity. NeuroImage, 19(4), 1463–1476.
Loo, C. K., & Mitchell, P. B.
(2005). A review of the efficacy of
transcranial magnetic stimulation (TMS) treatment for depression,
and current and future strategies to optimize efficacy. Journal of
Affective Disorders, 88(3), 255–267.
Mancini, M., Mastropasqua, C., Bonnì, S., Ponzo, V., Cercignani,
M., Conforto, S., . . . Bozzali, M. (2017). Theta burst stimulation
of the precuneus modulates resting state connectivity in the left
temporal pole. Brain Topography, 30, 312–319. https://doi.org/
10.1007/s10548-017-0559-x
Massimini, M., Ferrarelli, F., Huber, R., Esser, S. K., Singh, H., &
Tononi, G. (2005). Breakdown of cortical effective connectivity
during sleep. Science, 309, 2228–2232. https://doi.org/10.1126/
science.1117256
Massimini, M., Tononi, G., & Huber, R. (2009). Slow waves, synap-
tic plasticity and information processing: Insights from transcra-
nial magnetic stimulation and high-density EEG experiments.
European Journal of Neuroscience, 29, 1761–1770. https://doi.
org/10.1111/j.1460-9568.2009.06720.x
Medaglia, J. D., Huang, W., Karuza, E. A., Thompson-Schill, S. L.,
Ribeiro, A., & Bassett, D. S. (in press). Functional alignment with
anatomical networks is associated with cognitive flexibility. Na-
ture Human Behavior. Retrieved from https://arxiv.org/abs/1611.
08751
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Onnela,
J. P. (2010). Community structure in time-dependent, multiscale,
and multiplex networks. Science, 328, 876–878.
Muldoon, S. F., Pasqualetti, F., Gu, S., Cieslak, M., Grafton, S. T.,
Vettel, J. M., & Bassett, D. S. (2016). Stimulation-based control
of dynamic brain networks. PLoS Computational Biology, 12,
e1005076. https://doi.org/10.1371/journal.pcbi.1005076
Mullen, T. R. (2014). The dynamic brain: Modeling neural dynam-
ics and interactions from human electrophysiological recordings
(doctoral dissertation). Retrieved from https://escholarship.org/
uc/item/7kk2c4nd
Mullen, T. R., Kothe, C. A. E., Chi, Y. M., Ojeda, A., Kerth, T.,
Makeig, S., . . . Cauwenberghs, G. (2015). Real-time neuroimag-
ing and cognitive monitoring using wearable dry EEG. IEEE Trans-
actions on Biomedical Engineering, 62, 2553–2567. https://doi.
org/10.1109/TBME.2015.2481482
Nunez, P. L., & Srinivasan, R. (2006). Electric fields of the brain: The
neurophysics of EEG. New York, NY: Oxford University Press.
Nunez, P. L., Wingeier, B. M., & Silberstein, R. B. (2001). Spatial-
temporal structures of human alpha rhythms: Theory, micro-
current sources, multiscale measurements, and global binding
of local networks. Human Brain Mapping, 13, 125–164. https://
doi.org/10.1002/hbm.1030
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M.
(2010).
FieldTrip: Open source software for advanced analysis of MEG,
EEG, and invasive electrophysiological data. Computational In-
telligence and Neuroscience, 2011, e156869. https://doi.org/10.
1155/2011/156869
Pascual-Leone, A., Valls-Solé, J., Brasil-Neto, J. P., Cammarota, A.,
Grafman, J., & Hallett, M.
(1994). Akinesia in Parkinson’s dis-
ease. II. Effects of subthreshold repetitive transcranial motor cor-
tex stimulation. Neurology, 44, 892–898.
Pascual-Leone, A., Walsh, V., & Rothwell, J.
(2000). Transcra-
nial magnetic stimulation in cognitive neuroscience—virtual le-
sion, chronometry, and functional connectivity. Current Opinion
in Neurobiology, 10, 232–237. https://doi.org/10.1016/S0959-
4388(00)00081-7
Paus, T.
(1998).
Imaging the brain before, during, and after tran-
scranial magnetic stimulation. Neuropsychologia, 37, 219–224.
https://doi.org/10.1016/S0028-3932(98)00096-7
Pfurtscheller, G., Stancák, A., & Neuper, C. (2006a). Event-related
synchronization (ERS) in the alpha band—An electrophysiologi-
cal correlate of cortical idling: A review. International Journal of
Psychophysiology, 24, 39–46.
Pfurtscheller, G., Stancák, A., Jr., & Neuper, C.
(1996b). Post-
movement beta synchronization. A correlate of an idling motor
area? Electroencephalography and Clinical Neurophysiology, 98,
281–293. https://doi.org/10.1016/0013-4694(95)00258-8
Reimann, M. W., Nolte, M., Scolamiero, M., Turner, K., Perin, R.,
Chindemi, G., . . . Markram, H.
(2017). Cliques of neurons
bound into cavities provide a missing link between structure and
function. Front. Comput. Neurosci., 11. https://doi.org/10.3389/
fncom.2017.00048
Rogasch, N. C., & Fitzgerald, P. B. (2013). Assessing cortical net-
work properties using TMS-EEG. Human Brain Mapping, 34,
1652–1669. https://doi.org/10.1002/hbm.22016
Romei, V., Murray, M. M., Cappe, C., & Thut, G. (2013). The con-
tributions of sensory dominance and attentional bias to cross-
Journal of
modal enhancement of visual cortex excitability.
Cognitive Neuroscience, 25, 1122–1135. https://doi.org/10.1162/
jocn_a_00367
Romei, V., Thut, G., Mok, R. M., Schyns, P. G., & Driver, J. (2012).
Causal implication by rhythmic transcranial magnetic stimula-
tion of alpha frequency in feature-based local vs. global attention.
Network Neuroscience
635
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network reconfigurations after TMS
European Journal of Neuroscience, 35, 968–974. https://doi.org/
10.1111/j.1460-9568.2012.08020.x
Rosanova, M., Casali, A., Bellina, V., Resta, F., Mariotti, M., &
Massimini, M. (2009). Natural frequencies of human corticotha-
lamic circuits. Journal of Neuroscience, 29, 7679–7685. https://
doi.org/10.1523/jneurosci.0445-09.2009
Rose, N. S., LaRocque, J. J., Riggall, A. C., Gosseries, O., Starrett,
M. J., Meyering, E. E., & Postle, B. R. (2016). Reactivation of latent
working memories with transcranial magnetic stimulation. Science,
(354)1136–1139. https://doi.org/10.1126/science.aah7011
Rossi, S., Hallett, M., Rossini, P. M., & Pascual-Leone, A.
(2009).
Safety, ethical considerations, and application guidelines for the
use of transcranial magnetic stimulation in clinical practice and
research. Clinical Neurophysiology, 120, 2008–2039. https://
doi.org/10.1016/j.clinph.2009.08.016
Sale, M. V., Mattingley, J. B., Zalesky, A., & Cocchi, L. (2015). Im-
aging human brain networks to improve the clinical efficacy of
non-invasive brain stimulation. Neuroscience and Biobehav-
ioral Reviews, 57, 187–198. https://doi.org/10.1016/j.neubiorev.
2015.09.010
Siebner, H. R., Bergmann, T. O., Bestmann, S., Massimini, M.,
(2009).
Johansen-Berg, H., Mochizuki, H., . . . Rossini, P. M.
Consensus paper: Combining transcranial stimulation with neu-
roimaging. Brain Stimulation, 2, 58–80. https://doi.org/10.1016/
j.brs.2008.11.002
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C.
(2004).
Organization, development and function of complex brain net-
works. Trends in Cognitive Sciences, 8, 418–25. https://doi.org/
10.1016/j.tics.2004.07.008
Stewart, L. M., Walsh, V., & Rothwell, J. C.
(2001). Motor and
phosphene thresholds: A transcranial magnetic stimulation cor-
relation study. Neuropsychologia, 39, 415–419.
Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., & Leahy, R. M.
(2011). Brainstorm: A user-friendly application for MEG/EEG
analysis. Computational Intelligence and Neuroscience, 2011,
e879716. https://doi.org/10.1155/2011/879716
Tang, E., & Bassett, D. S.
(in press). Control of dynamics in brain
networks. Nature Communications. Retrieved from https://arxiv.
org/abs/1701.01531
Taylor, P. C. J., & Thut, G. (2012). Brain activity underlying visual
perception and attention as inferred from TMS–EEG: A review.
Brain Stimulation, 5, 124–129.
Telesford, Q. K., Lynall, M.-E., Vettel, J., Miller, M. B., Grafton, S. T.,
& Bassett, D. S.
(2016). Detection of functional brain network
reconfiguration during task-driven cognitive states. NeuroImage,
142, 198–210. https://doi.org/10.1016/j.neuroimage.2016.05.
078
Thut, G., Veniero, D., Romei, V., Miniussi, C., Schyns, P., & Gross,
J. (2011). hythmic TMS causes local entrainment of natural oscil-
latory signatures. Current Biology, 21, 1176–1185. https://doi.
org/10.1016/j.cub.2011.05.049
Tuladhar, A. M.,
ter Huurne, N., Schoffelen,
J.-M., Maris, E.,
Oostenveld, R., & Jensen, O.
(2007). Parieto-occipital sources
account for the increase in alpha activity with working memory
load. Human Brain Mapping, 28, 785–792. https://doi.org/10.
1002/hbm.20306
van Diessen, E., Numan, T., van Dellen, E., van der Kooi, A. W.,
Boersma, M., Hofman, D., . . . Stam, C. J.
(2015). Opportu-
nities and methodological challenges in EEG and MEG resting
state functional brain network research. Clinical Neurophysiol-
ogy, 126, 1468–1481. https://doi.org/10.1016/j.clinph.2014.11.
018
Vinck, M., Oostenveld, R., van Wingerden, M., Battaglia, F., &
Pennartz, C. M. A.
(2011). An improved index of phase-
synchronization for electrophysiological data in the presence
of volume-conduction, noise and sample-size bias. Neuro
Image, 55, 1548–65.
https://doi.org/10.1016/j.neuroimage.
2011.01.055
Vindiola, M. M., Vettel, J. M., Gordon, S. M., Franaszczuk, P. J.,
(2014). Applying EEG phase synchronization
& McDowell, K.
Journal
measures to non-linearly coupled neural mass models.
of Neuroscience Methods, 226, 1–14. https://doi.org/10.1016/j.
jneumeth.2014.01.025
Volberg, G., Kliegl, K., Hanslmayr, S., & Greenlee, M. W. (2009).
EEG alpha oscillations in the preparation for global and local pro-
cessing predict behavioral performance. Human Brain Mapping,
30, 2173–2183. https://doi.org/10.1002/hbm.20659
Network Neuroscience
636
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
6
1
1
1
8
6
7
2
8
1
n
e
n
_
a
_
0
0
1
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3