FORSCHUNG
Dynamic rewiring of electrophysiological brain
networks during learning
Paolo Ruggeri1
, Jenifer Miehlbradt1, Aya Kabbara2,4, and Mahmoud Hassan3,4
1Brain Electrophysiology Attention Movement Laboratory, Institute of Psychology, University of Lausanne, Schweiz
2Lebanese Association for Scientific Research, Tripoli, Libanon
3School of Engineering, University of Reykjavik, Reykjavik, Iceland
4MINDig, F-35000 Rennes, Frankreich
Schlüsselwörter: Human learning, Brain network dynamics, Elektroenzephalographie
Keine offenen Zugänge
Tagebuch
ABSTRAKT
Human learning is an active and complex process. Jedoch, the brain mechanisms underlying
human skill learning and the effect of learning on the communication between brain regions,
at different frequency bands, are still largely unknown. Hier, we tracked changes in large-scale
electrophysiological networks over a 6-week training period during which participants
practiced a series of motor sequences during 30 home training sessions. Our findings showed
that brain networks become more flexible with learning in all the frequency bands from
theta to gamma ranges. We found consistent increase of flexibility in the prefrontal and limbic
areas in the theta and alpha band, and over somatomotor and visual areas in the alpha band.
Specific to the beta rhythm, we revealed that higher flexibility of prefrontal regions during
the early stage of learning strongly correlated with better performance measured during home
Trainingseinheiten. Our findings provide novel evidence that prolonged motor skill practice
results in higher, frequency-specific, temporal variability in brain network structure.
ZUSAMMENFASSUNG DES AUTORS
We investigated the large-scale organization of electrophysiological brain networks of a cohort
von 30 participants practicing a series of motor sequences during 6 weeks of training. Mit
learning, we observed a progressive modulation of the dynamics of prefrontal and limbic
regions from theta to alpha frequencies, and of centro-parietal and occipital regions within
visuomotor networks in the alpha band. Zusätzlich, higher prefrontal regional flexibility
during early practice correlated with learning occurring during the 6 weeks of training. Das
provides novel evidence of a frequency-specific reorganization of brain networks with
prolonged motor skill learning and an important neural basis for noninvasive research into the
role of cortical functional interactions in (visuo)motor learning.
EINFÜHRUNG
Learning is a multidimensional concept with various definitions depending on the realm and
the level of investigation. At the most fundamental level, learning results in brain architecture
modifications through modulation of neural synapses (Smolen et al., 2016) causing changes in
neural information processing (Dayan & Cohen, 2011). Behaviorally, such adaptability allows
transforming an initially cognitively demanding and slow activity into a more spontaneous and
Zitat: Ruggeri, P., Miehlbradt, J.,
Kabbara, A., & Hassan, M.
(2023). Dynamic rewiring of
electrophysiological brain networks
during learning. Netzwerk
Neurowissenschaften, 7(2), 578–603.
https://doi.org/10.1162/netn_a_00289
DOI:
https://doi.org/10.1162/netn_a_00289
zusätzliche Informationen:
https://doi.org/10.1162/netn_a_00289
Erhalten: 4 Juli 2022
Akzeptiert: 2 November 2022
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Paolo Ruggeri
paolo.ruggeri@unil.ch
Handling-Editor:
Cornelis Jan Stam
Urheberrechte ©: © 2022
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network dynamics underlying motor skill learning
MRT:
Medical imaging technique used to
assess the anatomical structure of the
Körper, including the brain.
fMRT:
Technique used to measure the brain
activity through the assessment of
cerebral blood flow’s changes.
EEG:
A recording of the electrical activity
of the brain using electrodes placed
on the subject’s scalp.
Graphentheorie:
Mathematical investigation of the
structural organization of graphs.
Brain network dynamics:
Evolution of the brain’s functional
networks over time.
automatic process (Grafton et al., 2008). The last two decades have been characterized by a
growing interest in the scientific community in the area of learning, particularly motor skill
learning. Several previous learning experiments with MRI produced in the context of pre-
and posttraining regimes were able to clearly demonstrate that macroscale investigations
through brain structural and functional connectivity analyses can successfully identify
learning-related changes in cortical and subcortical brain networks. Genauer,
structural investigations (z.B., Draganski et al., 2006; Sampaio-Baptista et al., 2014; Taubert
et al., 2010, 2011) have consistently reported associations between gray matter increase
and learning in task-relevant areas including motor, parietal, and prefrontal cortex. Auf der
andererseits, following the practice of various motor learning tasks, functional connectivity
(z.B., Mehrkanoon et al., 2016; Sami et al., 2014; Sampaio-Baptista et al., 2015; Sun et al.,
2006; Taubert et al., 2011; Tung et al., 2013) was seen to be modulated within numerous
cortical (motor, fronto-parietal, sensorimotor, and visual) and subcortical (within and between
cerebellar nuclei, thalamus, and basal ganglia) networks as well as between cortical and sub-
cortical areas.
Jedoch, the entire field still remains underrepresented, probably because exploring
learning-related aspects requires long-term longitudinal designs (from weeks to months of
üben) Und, consequently, considerable deployment of research material and human
know-how. Zusätzlich, there is a need for studies dedicated to the temporal dynamics of inter-
actions between brain regions during learning phases that can surpass and augment knowl-
edge provided by—mostly resting-state—pre- and postinvestigation protocols. In this regard,
the most significant advances have been recently achieved using fMRI techniques and the
joint development and use of innovative data analysis methodologies (Mucha et al., 2010)
perfectly tailored to characterize brain dynamic processes such as those underlying learning.
Surprisingly, studies using electrophysiological neuroimaging techniques such as magneto/
electro-encephalography (M/EEG) are rare, and most of the few that do exist have investigated
motor adaptation (z.B., Gentili et al., 2015; Mehrkanoon et al., 2016; Miraglia et al., 2018)
Und, to a lesser extent, motor skill learning (z.B., Schubert et al., 2021; Tzvi et al., 2016,
2018) during very short periods of practice (from single sessions to very few days). Das ist
despite the flexibility and ease of use of these neuroimaging techniques and the recent devel-
opment and adaptation of existing brain dynamics methodologies to M/EEG data (Hassan &
Wendling, 2018; O’Neill et al., 2017; Tabbal et al., 2021).
The combination of fMRI techniques and the development of powerful mathematical tools
from network science and graph theory (Barabási, 2013; Stam, 2014) to identify metrics asses-
sing brain network dynamics have allowed for the characterization of changes in region-to-
region interactions in the brain (Bassett & Spurns, 2017; Kivelä et al., 2014; Mucha et al.,
2010). This approach provided a breakthrough in the description of adaptive fundamental
brain processes supporting learning (Bassett et al., 2011, 2013B, 2015; Mattar et al., 2018;
Reddy et al., 2018). Most of the investigations were based on the use of learning tasks involv-
ing motor and visual systems and the acquisition of new motor skills in particular. Results of
these studies have shown that learning is accompanied by large-scale dynamic variations over
time scales of seconds to minutes (Allen et al., 2014; Betzel et al., 2017; Zalesky et al., 2014)
across distributed networks associated with executive functions, visual processing, and motor
execution during task performance (Bassett et al., 2011, 2013B, 2015). Compared to classical
approaches focused on the activation strength of isolated brain regions, these studies brought
specific insights into how broad-scale interconnected systems accommodate learning. Im Par-
besonders, they showed that the recruitment of core visual and motor areas did not vary with prac-
tice, while their integration decreased with training. Zusätzlich, they revealed that individual
Netzwerkneurowissenschaften
579
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network dynamics underlying motor skill learning
Network community structures:
Set of brain regions (or nodes)
displaying dense functional
connections between each other and
relatively fewer connections with
other regions.
Brain region flexibility:
Quantitative measure assessing the
frequency with which each region
changes community affiliation over
Zeit.
learning was related to the extent of the decrease in the integration across medio-frontal brain
areas required in higher order cognitive processes (Bassett et al., 2015). These findings were
accompanied by evidence that brain network community structures (Mattar & Bassett, 2016)
(d.h., densely connected regions within a network) are not stable over time but evolve dynam-
isch. In the context of motor skill learning, several studies have assessed brain network
dynamics by quantifying the flexibility of brain regions in changing their community alle-
giance over time (Bassett et al., 2011, 2013B; Betzel et al., 2017; Reddy et al., 2018). Aus
a computational point of view, flexibility is commonly used as a tool to determine the temporal
variability of community structures and quantifies the frequency that a brain region changes its
modular affiliation over time. High values of flexibility are interpreted as indicative of contin-
uous change in community affiliation, while lower values are indicative of stable affiliation
im Laufe der Zeit (Bassett et al., 2011, 2013B; Betzel et al., 2017). Zusätzlich, it has been demon-
strated that regions with low (hoch) flexibility tend to be strongly (weakly) connected network
Knoten (Bassett et al., 2013B). For motor skill learning, this approach was particularly suited to
describe the transition from controlled toward automatic processes (Bassett et al., 2011,
2013B; Reddy et al., 2018). Bassett et al. (2011) were the first to highlight how brain region
flexibility was temporally correlated with subsequent performance during the acquisition of a
simple motor skill, and to suggest a possible modulation of flexibility with learning. Later, verwenden
a longitudinal motor skill learning protocol with fMRI, the same authors (Bassett et al., 2013B;
Reddy et al., 2018) confirmed the relationship between performance and flexibility and the
increase of flexibility with learning. Brain network flexibility has also been shown to correlate
with cognitive flexibility (Mattar & Bassett, 2016), positive affect, surprise, and fatigue (Betzel
et al., 2017). Taken together, this complementary evidence highlights the advantage of using
brain network flexibility to characterize the complex dynamics of brain networks.
The EEG literature on motor skill learning lacks brain network studies exploring the time
hierarchy and dynamics of learning-related brain functions over longer period of motor skill
Erwerb, but provides important research showing that the specific recruitment of brain
regions during performance of visuomotor adaptation and motor skill learning tasks takes
place in frequency bands from theta to gamma ranges. Several studies have shown EEG power
modulations in areas relevant to the performance of these tasks over frontal, motor, and visual
Bereiche. Genauer, the involvement of prefrontal areas, often implicated in aspects of
executive control with activity modulated by attentional and cognitive load (Klimesch,
1999; Langer et al., 2013; Scheeringa et al., 2008) has been mainly observed in the theta
(Aliakbaryhosseinabadi et al., 2021; Crivelli-Decker et al., 2018; Koch et al., 2020) frequency
band and, to a lesser extent, in the alpha (Meyer et al., 2014), beta (Aliakbaryhosseinabadi
et al., 2021; Jahani et al., 2020), and gamma (Aliakbaryhosseinabadi et al., 2021) bands.
Others have shown the involvement of sensorimotor and motor areas in the beta
(Aliakbaryhosseinabadi et al., 2021; Andres & Gerloff, 1999; Aoki et al., 2001; Boonstra
et al., 2007; Cunha et al., 2006; Heinrichs-Graham et al., 2016; Jahani et al., 2020; Jerbi
et al., 2004; Pollok et al., 2014; Rilk et al., 2011) and alpha (Andres & Gerloff, 1999;
Jerbi et al., 2004; Meyer et al., 2014; Rilk et al., 2011; Rueda-Delgado et al., 2019; Schubert
et al., 2021; Tzvi et al., 2016, 2018; Zhuang et al., 1997) bands, while the implication of the
theta (Koch et al., 2020; Perfetti et al., 2011; Studer et al., 2010; Tzvi et al., 2016) and gamma
(Aliakbaryhosseinabadi et al., 2021; Aoki et al., 2001; Perfetti et al., 2011) bands was less
consistently reported. A number of studies have also highlighted the recruitment of visual pro-
cessing areas during adaptation and visuomotor sequence tasks in the alpha (Meyer et al.,
2014; Rilk et al., 2011; Tzvi et al., 2016, 2018) band and, less consistently, in the beta (Rilk
et al., 2011) and gamma (Tzvi et al., 2016) bands. Important evidence is provided by
Netzwerkneurowissenschaften
580
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network dynamics underlying motor skill learning
connectivity studies during visuomotor practice. With respect to an executive network repre-
sented by functional interactions between prefrontal, zentral, and parietal regions, Rilk et al.
(2011) revealed an association between frontocentral coupling in the alpha band and
increased errors in a tracking task. In line with this finding, a decrease in coherence between
prefrontal and central regions was observed with ongoing learning and improved performance
in the theta, alpha, and beta bands (Gentili et al., 2015). Along the same lines, it was suggested
that this network becomes less relevant with acquired encoding (Tzvi et al., 2018). Mit
respect to a visuomotor network formed by functional interactions between central, parietal,
and occipital regions, it was observed that the coherence between motor and visual regions in
the alpha and beta band increased during visuomotor task performance (Classen et al., 1998;
Erla et al., 2012). Consistently, it was shown that coherence in these regions increased during
task execution in the beta band and that it was related to better tracking task performance in
the alpha band (Rilk et al., 2011). Endlich, few brain network studies showed an increase of
modularity and transitivity metrics with learning, in the theta and alpha bands (Miraglia et al.,
2018), and a relationship between small-worldness and better learning in the alpha band
(Vecchio et al., 2018).
Taken together, this substantial evidence from neuroimaging studies highlights how there is
still much to be clarified about the mechanisms underlying motor skill learning and how func-
tional communication between different brain regions is modulated by learning. Insbesondere,
to provide a more detailed description of the dynamics of these functional brain networks,
much work is needed to reveal how functional connectivity modulations at different frequen-
cies occur over extended periods of practice. Hier, we aimed to close this gap by leveraging
on an EEG-based time-varying connectivity analysis in the context of a longitudinal motor skill
protocol. Zu diesem Zweck, inspired by previous fMRI studies on this topic (z.B., Bassett et al., 2013A,
2013B), we investigated whether learning a new motor skill from visual cues was accompa-
nied by a reorganization of brain networks also visible in the EEG. Insbesondere, we assessed
whether extended practice was accompanied by frequency-specific modulations of flexibility
across brain regions and whether the dynamics of brain networks in early stages of learning
was related to motor skill performance (Bassett et al., 2011, 2013B; Mattar et al., 2018). Wir
studied a cohort of healthy adult human subjects (N = 30) who performed 30 sessions of a
discrete sequence-production task (DSP) über 6 weeks (Figur 1). Während dieser Zeit, partici-
pants underwent four high-density (HD) EEG sessions while practicing the same DSP task, fol-
lowing an experimental protocol used in several previous works (Bassett et al., 2013B, 2015;
Mattar et al., 2018; Reddy et al., 2018; Wymbs & Grafton, 2015). We examined network
reconfiguration using dynamic network measures involving community detection and flexibil-
ity analysis (Bassett et al., 2013A, 2013B; Mucha et al., 2010).
We hypothesized a (ich) global increase in flexibility (Bassett et al., 2013B; Reddy et al., 2018)
over successive EEG sessions. Genauer, Wir (ii) expected that a reduced involvement
of prefrontal regions in cognitive control processes with learning (Bassett et al., 2015) würde
manifest in an increased availability of interaction with other regions and thus in the form of
increased flexibility of regions of this network in theta (Aliakbaryhosseinabadi et al., 2021;
Gentili et al., 2015; Miraglia et al., 2018), alpha (Gentili et al., 2015; Miraglia et al., 2018;
Rilk et al., 2011), Und, to a lesser extent, beta (Gentili et al., 2015) frequency bands. Weiter-
mehr, we hypothesized that (iii) the previously observed learning-related reduction in integra-
tion between regions in the somatomotor and visual networks (Bassett et al., 2015) may
manifest in the form of increased flexibility of regions belonging to these networks predomi-
nantly in the alpha and beta frequency bands (Classen et al., 1998; Erla et al., 2012; Rilk et al.,
2011). zuletzt, based on evidence that both the functional disengagement of prefrontal regions
Netzwerkneurowissenschaften
581
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network dynamics underlying motor skill learning
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
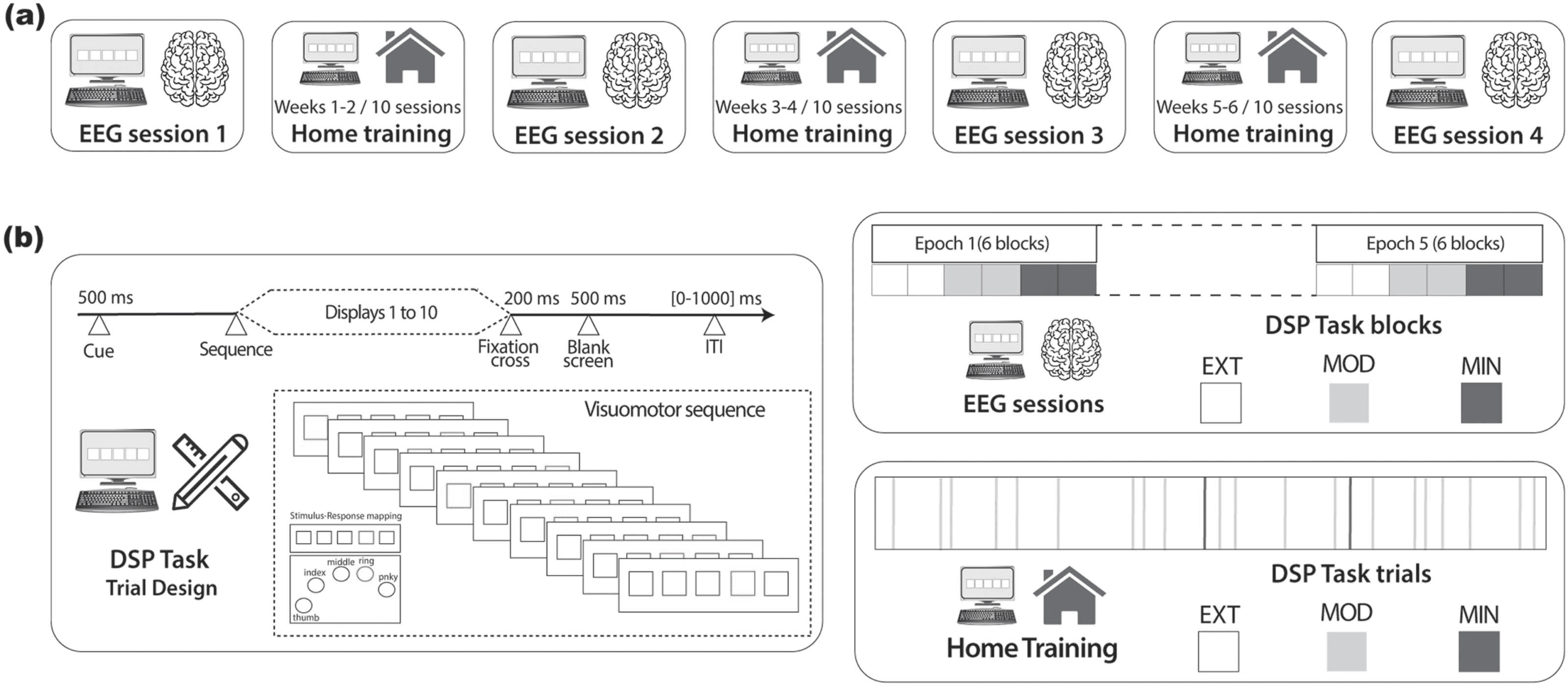
Experimental protocol and task design. (A) EEG sessions in the laboratory were interleaved with training sessions at home. Study
Figur 1.
participants practiced the DSP task for the first time during EEG session 1 and returned for another EEG session every 2 weeks (EEG sessions 2
Zu 4) nach 10 home training sessions of the DSP task. (B) Auf der Linken, the trial design of the DSP task. The numbers represent the duration of the
presented stimuli consisting of a 500-ms cue preceding the practice of the visuomotor sequence made of 10 consecutive displays, followed by
a fixation cross (200 MS), a blank screen (500 MS), and an intertrial interval of varying duration (0–1,000 ms). To terminate the sequence,
participants had to select the correct stimulus-response mapping. Auf der rechten Seite, the DSP task practiced during EEG and home training sessions.
The former consisted of five epochs made of six blocks (two for EXT, MOD, and MIN). Within each block, five sequences of each type (EXT 1
Und 2, MOD1 and 2, and MIN 1 Und 2) were practiced in respective trials. The appearance of the sequences was randomized within each
block and the appearance of the blocks was randomized within each epoch. At the end of each EEG session, participants practiced 100 EXT
Versuche (50 EXT1 and EXT2), 100 MOD trials (50 MOD1 and MOD2), Und 100 MIN trials (50 MIN1 and MIN2). The DSP task practiced during
the home sessions consisted of 150 Versuche, divided into 64 trials for each EXT sequence, 10 trials for each MOD sequence, and one trial for each
MIN sequence. Trials were randomized.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
Ruhezustand:
Periods during which
(electro)physiological activity is
recorded in the absence of stimuli
evoked by the experimental protocol.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
(Bassett et al., 2015) and the configuration of brain networks during resting state (Mattar et al.,
2018) and early phases of learning (Bassett et al., 2011, 2013B) are related to the amount of
learning in subsequent practice sessions, (iv) we expected a positive relationship between
early prefrontal regions flexibility and the learning rate over 6 weeks of practice in the theta,
alpha, and beta frequency bands. Endlich, it is important to note that the methods used in this
arbeiten, although extensively used and representing the methodological reference of the totality
of the works in the field, are still undergoing continuous improvement. To shed light on pos-
sible confounding factors (such as the signal’s power and the task design) that could influence
the extracted metrics, these methods have been put in perspective and further explored in
section 4 of the Supporting Information.
ERGEBNISSE
We investigated the dynamics of brain network reconfiguration underlying prolonged training
of a simple motor skill (Figur 1 and Figure 2). We focused on functional connectivity between
different brain regions at four frequency bands (theta: 4–8 Hz; alpha: 8–12 Hz; beta: 12–28 Hz;
gamma: 28–45 Hz) measured during four EEG sessions. These sessions were executed every
2 weeks as part of an experimental protocol quantifying learning of a simple sensorimotor
Netzwerkneurowissenschaften
582
Brain network dynamics underlying motor skill learning
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
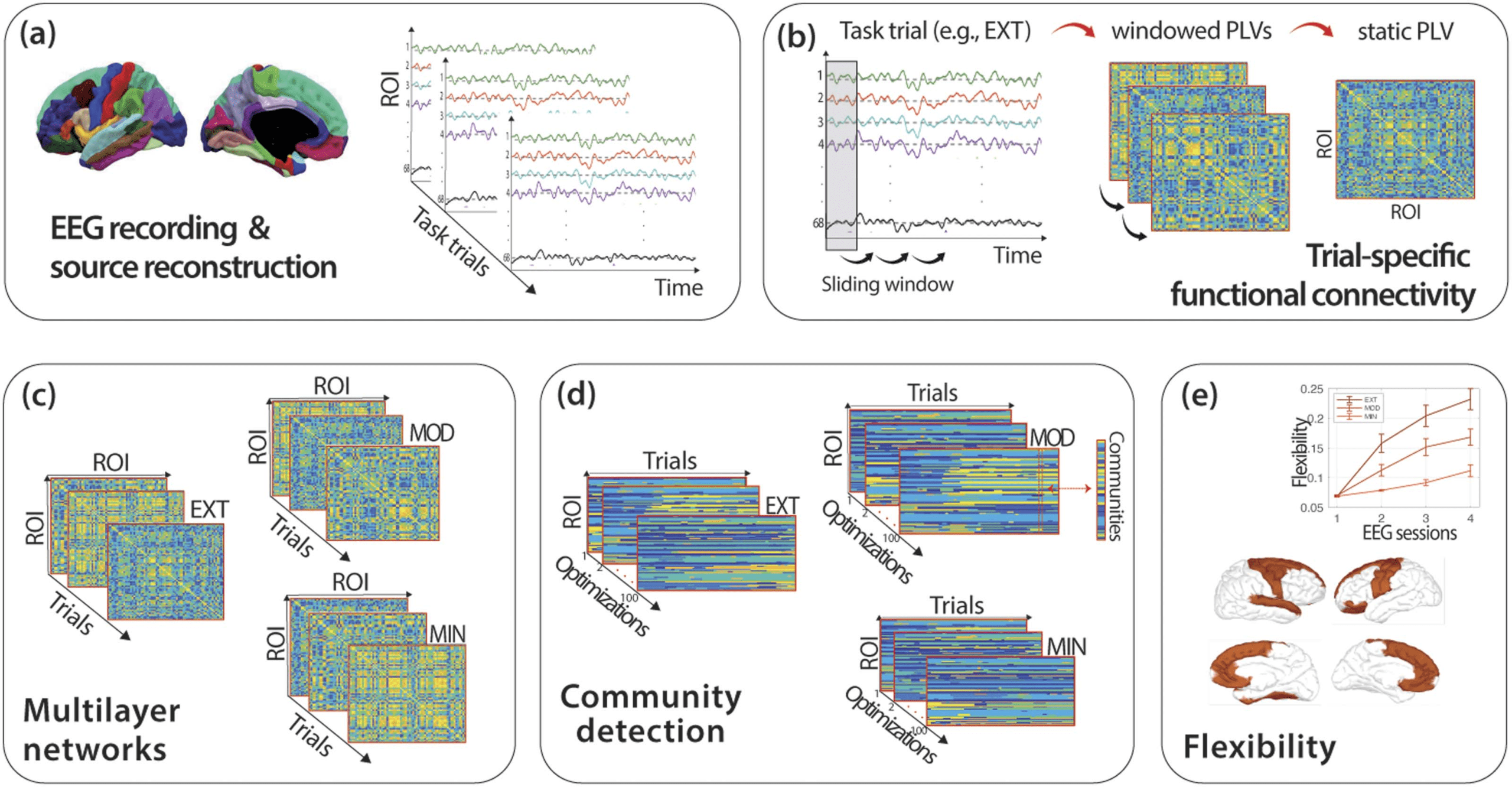
Figur 2. Methodological framework to compute flexibility. (A) EEG data were recorded, and the source activity of 68 ROIs of the Desikan–
Killiany atlas in each trial of the DSP task was computed using the wMNE algorithm. (B) Trial-specific functional connectivity was obtained as a
static PLV matrix obtained by averaging the windowed PLVs obtained with sliding window approach where connectivity was computed within
each temporal window. (C) Multilayer network tensors for each type of sequence (EXT, MOD, and MIN) were built by concatenating static
PLVs from consecutive trials of the same type. (D) Community detection was implemented on each multilayer network tensor through the
modularity maximization method. Modularity maximization was run 100 mal, leading to 100 optimized community assignments for each
sequence type. (E) Global and regional flexibility was computed for each run and then averaged over the 100 runs. The procedure outlined
above was applied to the EEG signal of each participant in each frequency band (theta to gamma) and experimental session (1 Zu 4).
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
sequence learning task practiced during 30 home-based sessions over 6 weeks (Figure 1A).
Während dieser Zeit, 30 participants learned six 10-element sequences, practiced in pairs at
an intensive (two EXT sequences: EXT1 and EXT2), moderate (two MOD sequences: MOD1
and MOD2), and minimal (two MIN sequences: MIN1 and MIN2) pace (Figure 1B). To char-
acterize brain dynamics (Figur 2), functional connectivity matrices were computed from the
EEG source-localized signals extracted during the execution of each trial of the task (corre-
sponding to the execution of a sequence). Multilayer temporal networks (Holme & Saramäki,
2012) were then separately built for each training intensity (EXT, MOD, and MIN) von
concatenating, in successive layers, connectivity matrices corresponding to consecutive trials
of the same type (EXT, MOD, or MIN). These multilayer temporal networks were then analyzed
using computational tools for dynamic community detection (Bassett et al., 2013A; Mucha
et al., 2010) consisting in the maximization of a multilayer modularity quality function Q
describing the within- and across-layers relationships between brain regions. This was done
to identify functional modules (d.h., clusters of communities of brain regions sharing correlated
brain dynamics) across the entire course of learning. Each community of brain regions can be
thought of as segregated network coding for specific cognitive or motor functions. To quantify
the dynamic properties of community structure and capture changes over the 6 weeks of con-
tinuous learning, we used flexibility (Bassett et al., 2011, 2013B) as network diagnostics. Endlich,
behavioral task performance was quantified by the time required to complete an entire
Netzwerkneurowissenschaften
583
Brain network dynamics underlying motor skill learning
sequence (movement time, MT). Following the approach validated in previous studies (Bassett
et al., 2013B, 2015; Mattar et al., 2018), the reduction in MT with ongoing practice was used as
an indicator of learning.
Behavioral Performance
We first sought to determine, from a behavioral point of view, the effectiveness of the experimen-
tal manipulation as practice progressed. Zu diesem Zweck, changes in MT (d.h., time required to prac-
tice a sequence correctly) across the six weeks of training were assessed with repeated measures
ANOVAs (with training intensity—EXT, MOD, and MIN—and laboratory session—1 to 4—as
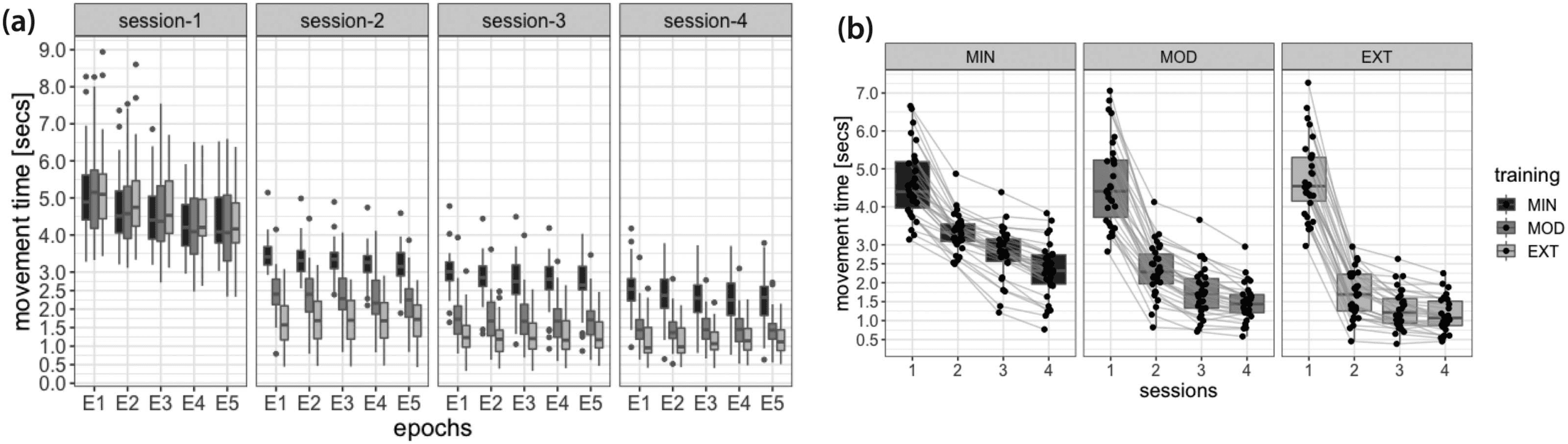
within-subject factors; Figure 3A and B). MT significantly decreased across sessions (P < .001)
and differed between EXT, MOD, and MIN sequences (p < .001) with lowest values observed
during the execution of the EXT sequences (EXT: M = 2.22 s, SE = 0.11 s; MOD: M = 2.54 s,
SE = 0.13 s; MIN: M = 3.26 s, SE = 0.12 s). MT decreased more rapidly across scan sessions
for sequences that were extensively practiced (EXT) as compared to less practiced ones (inter-
action effect, p < .001). Taken together, these analyses showed that the MT recorded during EEG
sessions consistently decreased within-subjects for all practiced sequences (see also Figure 3B
for a qualitative appreciation of this reduction as the subject-specific level), corroborating
previous findings (Bassett et al., 2015; Wymbs & Grafton, 2015).
Following previous works using the same experimental procedure (Bassett et al., 2013b,
2015; Mattar et al., 2018; Reddy et al., 2018), we quantified learning by assessing the decrease
in the MT required to correctly perform each sequence as practice progressed. The learning
rate K of each participant was computed as the exponential drop-off of MT (r2: M = 0.96; SD =
0.05) required to perform the two EXT sequences during the 30 home sessions (see Methods
section and Figure S1). Indeed, these sequences were extensively practiced (1,920 trials) over
30 consecutive sessions evenly spaced over the 6 weeks of training. The learning rate, quan-
tifying how fast each participant converged to a relatively steady performance, varied signif-
icantly across participants (min = −0.96, max = −0.07; 25th percentile = −0.36, median =
−0.31, 75th percentile = −0.23), highlighting a substantial interindividual difference in the
sample tested (see Table S1 for the descriptive of the initial and final MT, and other parameters
Figure 3. Behavioral indicators of performance. (A) MT recorded during EEG sessions and grouped by training intensity (EXT, MOD, and
MIN), revealing different improvements as a function of practice exposure during home sessions. During each EEG session the EXT, MOD,
and MIN sequences are equally practiced during five consecutive epochs. (B) Paired observations highlighting the distribution of MT for EXT,
MOD, and MIN sequences recorded during EEG sessions 1–4, and revealing that all participants decreased their MT during the 6 weeks of
practice. Lower and upper box boundaries represent the 25th and 75th percentiles (Q1 and Q3); the difference between Q1 and Q3 is the
interquartile range (IQR); the horizontal line inside the box represents the median of the distribution; the lower and upper extreme lines show
Q1 − 1.5xIQR and Q3 + 1.5xIQR, respectively; finally, the filled gray circles are data points falling outside the lower and upper extreme lines.
Network Neuroscience
584
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
computed from the fitting procedure). Results and analyses of this section are further detailed
in section 1 of the Supporting Information.
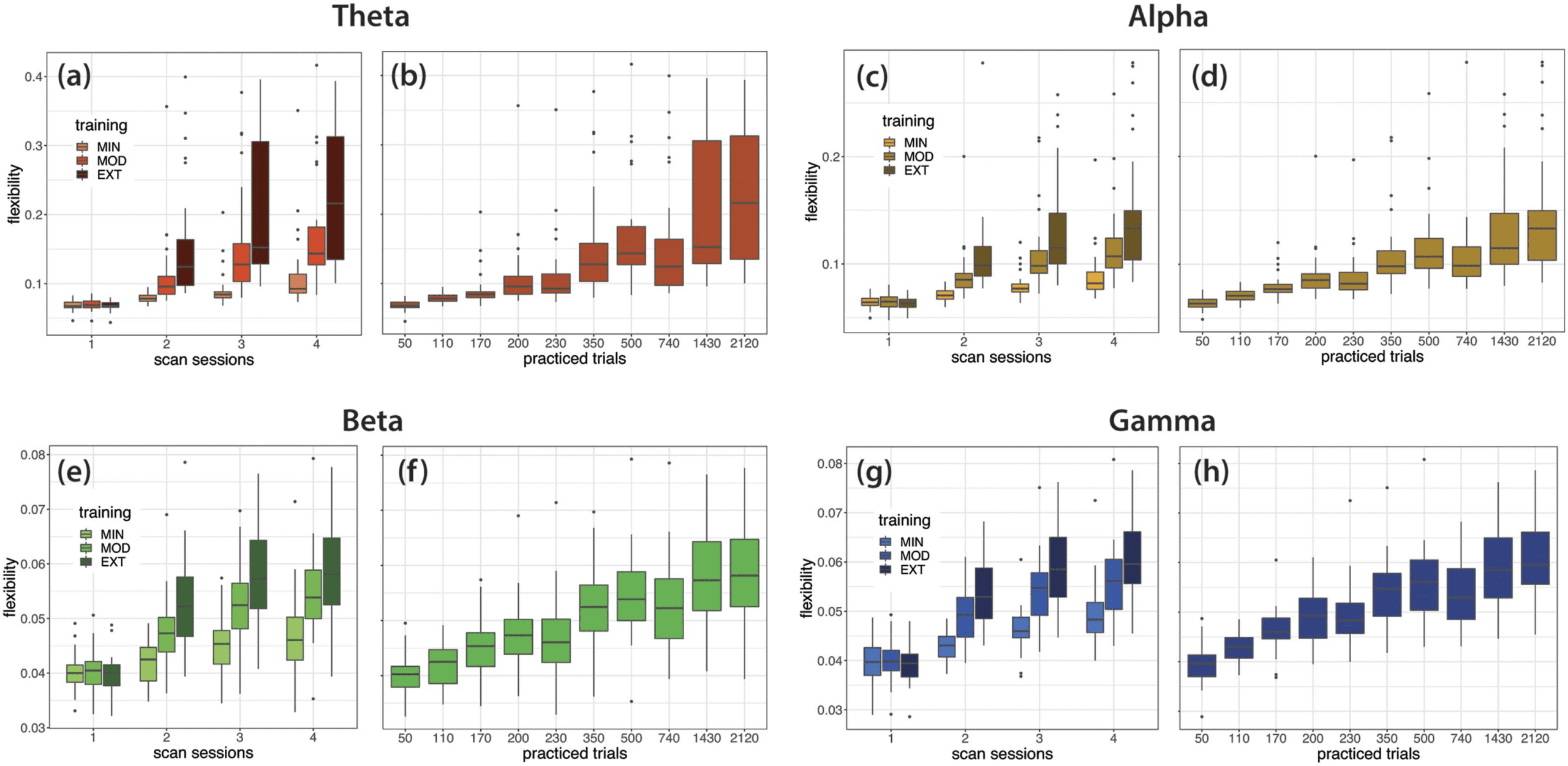
Global and Regional Flexibility Variation With Ongoing Practice
Based on the detected community assignments obtained for each participant and EEG session,
we computed the flexibility of each of the 68 brain regions defined by the Desikan–Killiany
anatomical atlas for each training intensity (EXT, MOD, and MIN) and frequency band of inter-
est. Flexibility was used to quantify the frequency at which brain regions change their com-
munity affiliation with time (i.e., across layers), normalized by the total possible changes. An
assessment of this quantity at different levels of practice offers the advantage of quantifying the
functional reorganization of specific brain regions throughout learning.
Quantification of global flexibility. We first sought to determine how brain network dynamics
were configured as practice progresses, at a global level. To this end, changes in global flex-
ibility (i.e., average of flexibility across brain regions) over the 6 weeks of training were eval-
uated with repeated measure ANOVAs (with training intensity—EXT, MOD, and MIN—and
laboratory session—1 to 4—as within-subject factors) for each frequency band (Figure 4A, C,
E, and G). In all frequency bands, flexibility increased across sessions (p < .001) and significantly
differed between EXT, MOD, and MIN trials (p < .001), with highest values observed during
the execution of the EXT sequences. Global flexibility increased more rapidly across scan
sessions for sequences that were extensively practiced (interaction effect, p < .001). When
data are sorted according to the number of trials performed (Figure 4B, D, F, and H; refer
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Global flexibility changes with learning. (A, C, E, G) Global flexibility, grouped by training intensity (EXT, MOD, and MIN), com-
puted in the theta, alpha, beta, and gamma bands from EEG data recorded at consecutive EEG sessions. (B, D, F, H) Global flexibility computed
as a function of the number of trials completed after a given EEG session in the theta, alpha, beta, and gamma bands. Lower and upper box
boundaries represent the 25th and 75th percentiles (Q1 and Q3); the difference between Q1 and Q3 is the interquartile range (IQR); the
horizontal line a inside the box represents the median of the distribution; the lower and upper extreme lines show Q1 − 1.5xIQR and
Q3 + 1.5xIQR, respectively; finally, the filled gray circles are data points falling outside the lower and upper extreme lines.
Network Neuroscience
585
Brain network dynamics underlying motor skill learning
to Table S2 to see how cumulative practice trials were computed), they qualitatively replicate
those shown in Bassett et al. (2013b; see Figure 2c in their paper) where flexibility estimates
were obtained from fMRI data recorded with the same experimental protocol used in this
study. Consistently, we also observed an increasing number of communities with learning in
all frequency bands (Figure S2E–H), although the quality of partitions in functional communi-
ties with increasing practice seems to increase in the theta and alpha bands and decrease in
the beta and gamma bands (Figure S2A–D). Results are detailed in section 2 of the Supporting
Information.
Quantification of regional flexibility. To investigate the contribution of each brain region in
increasing flexibility with practice, we contrasted flexibility values obtained from successive
laboratory sessions for each frequency band. Significant differences were quantified with a
one-tailed Wilcoxon test, testing the assumption that flexibility measured at a given session
is bigger than flexibility at previous sessions. We included Bonferroni correction for multiple
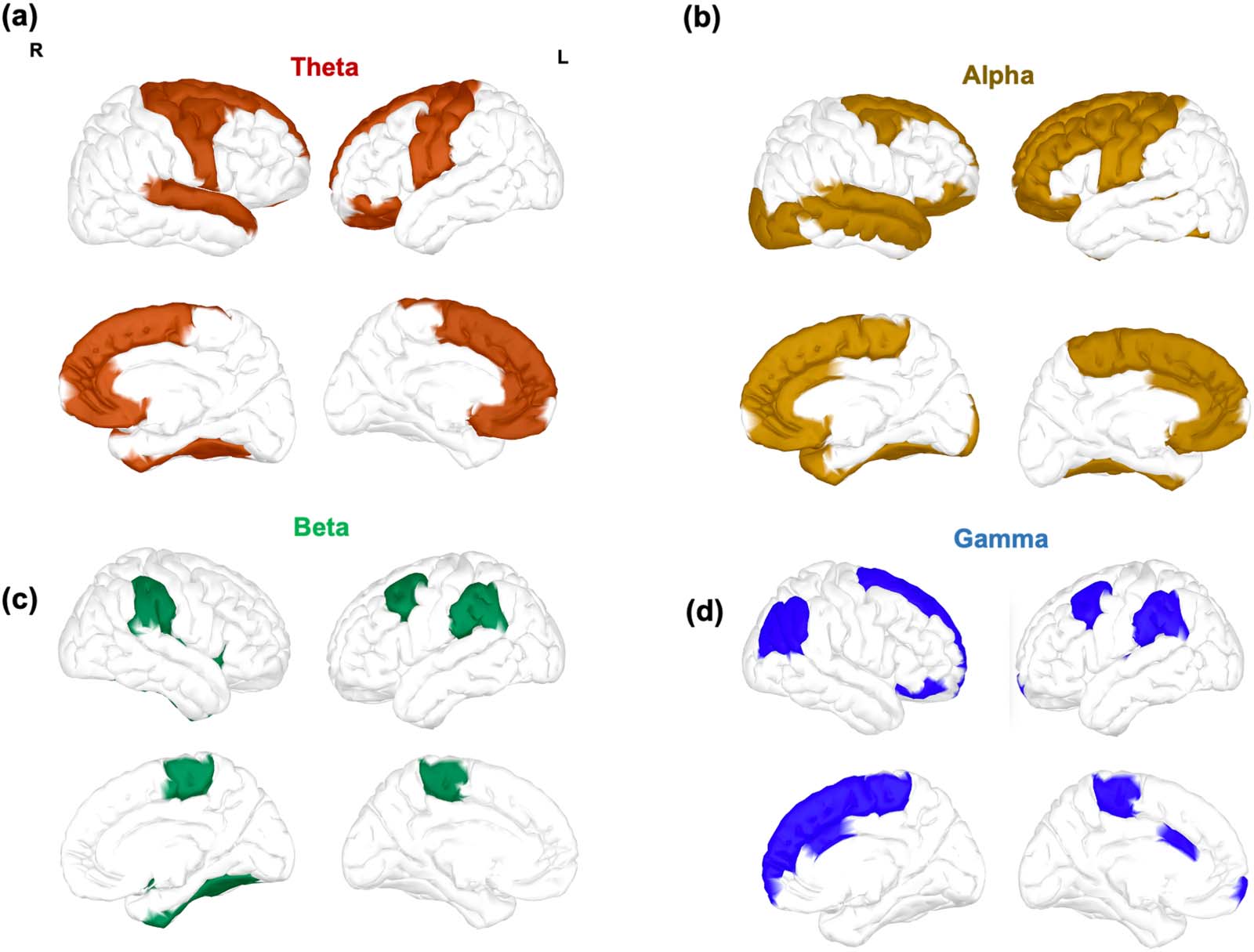
comparisons across brain regions. In Figure 5 we display regions with the most significant
increase in flexibility between session 1 and 4.
The theta and alpha bands (Figure 5A and B, respectively) were characterized by a similar
increase of flexibility in prefrontal and central regions within default mode network (DMN)
areas (rostral anterior cingulate, superior frontal, caudal middle frontal, and superior temporal),
in central regions within somatomotor network areas (precentral, postcentral, and superior
temporal), in prefrontal regions within limbic network areas (medial-orbitofrontal and rostral
anterior cingulate), and in frontal regions over dorsal attention (superior frontal) and fronto-
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Brain regions displaying the most significant increase in flexibility between EEG recordings in session 1 and 4 for the (A) theta, (B)
alpha, (C) beta, and (D) gamma frequency bands. Contrasts were assessed using a Wilcoxon test with Bonferroni correction, statistical results
were divided in quintiles, and only statistically significant brain regions in the uppermost two quintiles are shown.
Network Neuroscience
586
Brain network dynamics underlying motor skill learning
parietal (caudal middle frontal) network areas. Specific to the alpha band (Figure 5B), we
observed an increase in prefrontal regions within fronto-parietal and ventral attention network
areas (caudal anterior cingulate and left rostral middle frontal), over few more frontal regions
within the DMN (caudal anterior cingulate and right middle temporal), and over posterior
regions in visual network areas (right lateral occipital). Unlike for the theta and alpha bands,
we did not observe a diffused significant increase in flexibility across brain regions in the beta
frequency band (Figure 5C). Instead, flexibility specifically increased in the caudal middle
frontal (covering areas within default mode and fronto-parietal networks), in the parietal areas
of the supra-marginal (within areas specific to dorsal, somatomotor, and ventral networks), and
in the paracentral within the somatomotor network. Finally, the gamma band (Figure 5D) was
characterized by an increase in prefrontal (right superior frontal, caudal middle frontal, and
anterior cingulate, and pars orbitalis) and parietal (right inferior parietal) regions within the
DMN areas, and in regions within areas of the fronto-parietal (caudal middle frontal, caudal
anterior cingulate, and right inferior parietal), ventral (caudal anterior cingulate and left supra-
marginal) and dorsal attention (right superior frontal, left supra-marginal, and right inferior pari-
etal), and somatomotor (left supra-marginal and paracentral) networks.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
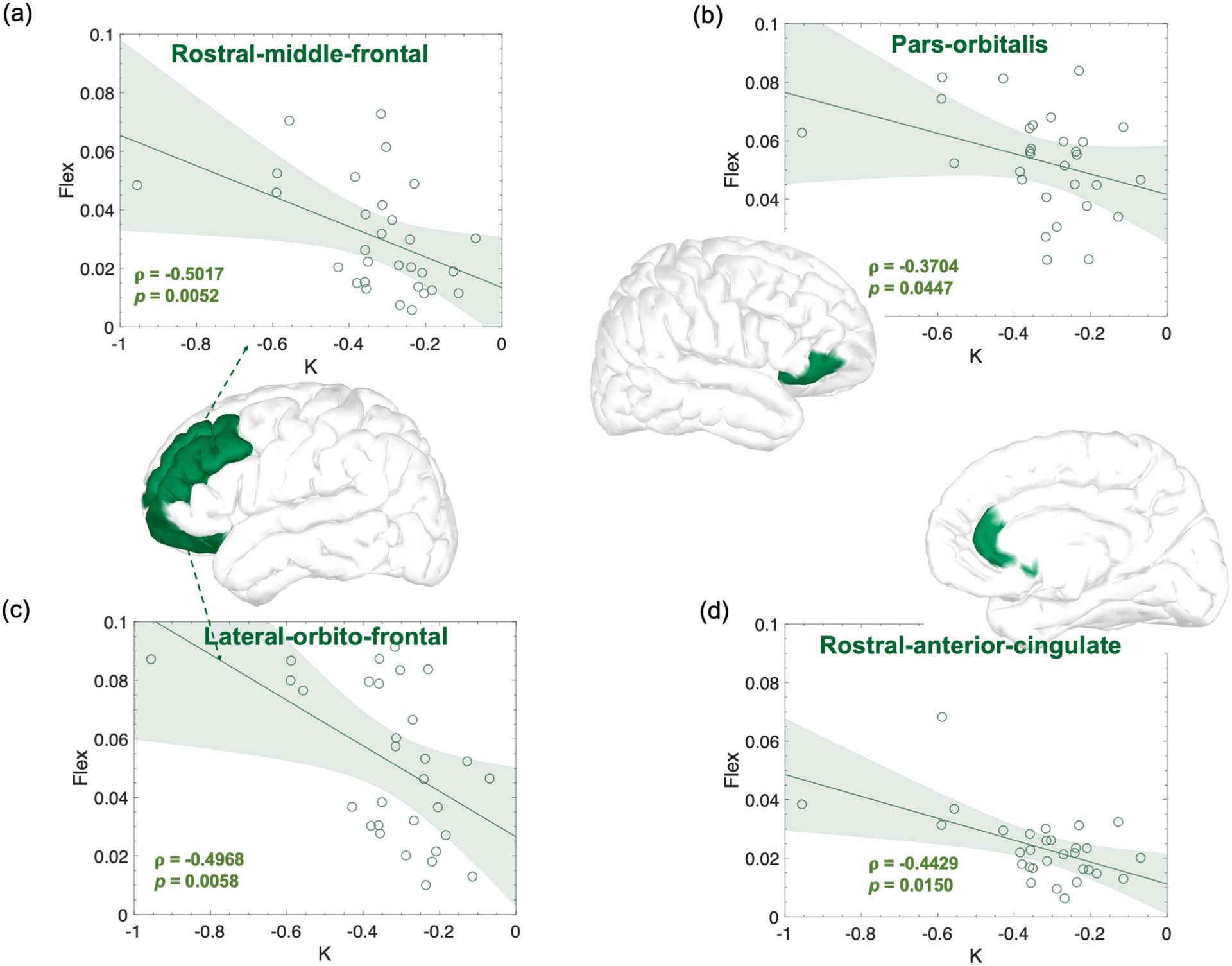
Significant correlations between regional flexibility in the beta frequency band computed from the EEG recorded in the first lab-
Figure 6.
oratory session and the learning rate K obtained from performance of the EXT sequences during home training sessions. Effect sizes (r and p) are
also shown on the figure as well as the confidence intervals.
Network Neuroscience
587
Brain network dynamics underlying motor skill learning
Functional Correlates of Performance
Next, we asked whether the flexibility of brain regions prior to extensive practice could be
indicative of the individual learning profile observed during the following 6 weeks of training.
To this end, we focused exclusively on the EEG signal recorded during the first EEG session. As
described above, during this session participants practiced the six motor sequences for the
very first time, and the flexibility measures extracted during this recording could be indicative
of the prelearning functional state of each individual. To have consistent measures of flexibility
across subjects, we thresholded the task trials length of each subject according to the length of
the fastest recorded trial across subjects during the first session. As for the previous analysis, we
built and analyzed the multilayer temporal networks through multilayer modularity optimiza-
tion, and we used the computed flexibility as network diagnostics. We then correlated, for
each region, the flexibility scores in each frequency band with the learning rate K obtained
from practice during the following 6 weeks. Significant relationships—evaluated with Spear-
man’s rank correlations—were found in the beta frequency bands (Figure 6) in the left orbito-
frontal (ρ = −0.50; p value = 0.005) and rostral middle frontal (ρ = −0.50; p value = 0.005)
regions, and in the right parsorbitalis (ρ = −0.37; p value = 0.044) and rostral anterior cingulate
(ρ = −0.44; p value = 0.014) regions. All the relationships remained significant after controlling
for the initial and final MT, except for the right parsorbitalis region (Table S3).
DISCUSSION
In this study, we aimed to assess whether extended practice of a new motor skill from visual
cues was accompanied by frequency-specific changes of brain network dynamics. We pre-
dicted (i) a global increase of cortical flexibility with learning. Specifically, we expected (ii)
increased flexibility over prefrontal regions in the theta, alpha, and beta frequency bands,
and (iii) over sensorimotor and occipital regions in the alpha and beta bands. Lastly, (iv) spe-
cific to the theta, alpha, and beta bands, we expected a positive relationship between partic-
ipants’ prefrontal flexibility recorded before extended sequence practice and their learning rate
from 6 weeks of training. Analyses in the gamma band were included to provide exploratory
evidence of the dynamics of brain networks in these frequency ranges.
Thanks to the superior (subsecond) temporal accuracy of EEG compared to fMRI and the
use of a longitudinal experimental setup (Figures 1 and 2), we were able to explore frequency-
dependent aspects of learning by characterizing brain networks in specific EEG rhythms. We
found that all participants improved performance of all sequences, with a strong interindivid-
ual variability characterized by variable rates of improvement (Figure 3 and Figure S1). We
showed that the advancement of motor skill learning through repeated and sustained sequence
practice is accompanied by a robust reorganization of functional brain networks as shown by
global flexibility increase in all studied frequency bands from theta to gamma ranges (Figure 4).
As learning progresses, a considerable number of brain regions change more frequently their
modular affiliation (Figure 5). Most of the increase was detected in the theta, alpha, and
gamma bands and, to a lesser extent, in the beta band. In particular, increases in numerous
prefrontal and centro-parietal areas within DMN, fronto-parietal, and dorsal attention net-
works were observed in the theta, alpha, and gamma bands, and over occipital regions within
the visual network in the alpha band. Increases in prefrontal areas of the limbic network were
specific to the theta and alpha bands. A consistent modulation of regions within the somato-
motor network was instead revealed in all frequency bands. Finally, few regions within the
ventral attention network increased their flexibility, mostly over frontal (alpha and gamma)
and parietal (beta and gamma) areas. Finally, we have shown that individuals with highly
Network Neuroscience
588
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
flexible prefrontal regions in the beta spectrum at the onset of learning are those who learn
more rapidly in the following 6 weeks of training (Figure 6).
The global increase of flexibility with practice is in line with previous fMRI studies (Bassett
et al., 2011, 2013b; Reddy et al., 2018). Bassett et al. (2011) monitored performance on a
simple motor skill task during three consecutive fMRI sessions and observed an increase in
global flexibility during the early stages of learning followed by a relative decrease during
the third recording. The lack of consistent flexibility increases as observed in our results could
be explained by the three fMRI scans occurring at initial stages of learning. This did not allow
an assessment of learning and flexibility over a longer period, as was instead possible in two
subsequent fMRI studies (Bassett et al., 2013b; Reddy et al., 2018) assessing motor skill
learning using the same longitudinal protocol used in our work. Consistent with our results
(Figure 4B, D, F, and H), Bassett et al. (2013b) showed an increase in global flexibility (see
Figure 2c in their work) with the number of trials practiced, which is consistent with an
increased specificity of functional connectivity patterns with extended learning. On the same
line, Reddy et al. (2018) recently identified two “canonical” brain states made by sensorimotor
and fronto-temporal subcortical regions during practice and observed that the rate of switching
between these states constantly increased during the 6 weeks of training. Overall, the global
flexibility increase observed in our study is coherent with a transition from greater behavioral
adaptability toward more automatic performance with greater freedom of cognitive resources
for other processes (Bassett et al., 2015; Shamloo & Helie, 2016).
As expected, we observed increased flexibility over prefrontal and limbic areas in the theta
and alpha band, but we did not find a significant increase over these regions in the beta band.
In terms of the brain areas involved and their dynamics with learning, our results are consistent
with one of the core findings of Bassett et al. (2015) who highlighted a diffused decrease in the
recruitment of prefrontal regions particularly relevant to the fronto-cingulate network. In line
with their work, we observed variations of flexibility over hub regions (rostral anterior cingu-
late, caudal middle frontal, medial orbito-frontal, and superior frontal) of cognitive control
systems like the fronto-parietal and cingulo-opercular networks (Elton & Gao, 2014). It was
speculated (Bassett et al., 2015) that with learning these hubs become disengaged with the
rest of the network as cognitive control is particularly critical during early skill acquisition
(Hikosaka et al., 2002; Petersen et al., 1998). Our observed increase in flexibility supports this
possibility. The fact that the increased flexibility with learning in these regions is specific to the
theta and alpha rhythms is coherent with several lines of research. Our results are consistent
with previous work highlighting a reduced coherence between prefrontal and central areas in
both bands during movement planning and execution (Gentili et al., 2015) at later stages of
learning. Similarly, our findings agree with those of Miraglia et al. (2018) showing a global
increase of modularity and transitivity measures after motor adaptation training in the theta
and alpha band, and with those of Vecchio et al. (2018) relating increased network segregation
and integration with better learning in the alpha band. Importantly, Rilk et al. (2011) showed in
a visuomotor tracking task that high tracking error was associated with enhanced frontocentral
coupling in the alpha band, suggesting additional activation of a fronto-parietal control
network. Taken together, these are all evidence supporting the possibility that the release of
specific hubs in the fronto-parietal and cingulo-opercular networks (Bassett et al., 2015)
occur within the theta and alpha frequency ranges. Finally, we observed unpredicted increases
in few prefrontal regions of the right hemisphere in the gamma band. Although there is very
little evidence for this frequency band, our findings seem coherent with a recently observed
positive correlation between visuomotor tracking tasks difficulty and fronto-central connectivity
(Aliakbaryhosseinabadi et al., 2021).
Network Neuroscience
589
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
Coherent with our hypothesis, we found increased flexibility in the alpha band over soma-
tomotor (pre- and postcentral gyrus) and visual areas (lateral occipital regions). In the beta
band, flexibility also increased over somatomotor areas but was only specific to paracentral
and supra-marginal regions. Past research on how somatomotor areas change their activity
during motor skill learning is divided into studies monitoring task-evoked BOLD activity
(Dayan & Cohen, 2011) and connectivity (Bassett et al., 2015) during different phases of learn-
ing. Several studies showed a reduction in BOLD activity during the early stages of learning
(Floyer-Lea & Matthews, 2005; Sakai et al., 1999) followed by an increase in activity during
the late stages (Floyer-Lea & Matthews, 2005; Grafton et al., 2008; Honda et al., 1998).
However, real insight into the dynamics of the undergoing process has been provided by
connectivity studies and particularly by the work of Bassett et al. (2015). Consistent with
the functional requirements of skill acquisition, they suggested that with extended practice
motor and visual systems transition from being tightly integrated to operating as independent
units. Our results of increased flexibility over somatomotor and occipital areas are coherent
with these findings and go further suggesting that this late learning independence could be
expressed by the increased frequency with which these regions change allegiance to func-
tional communities. Only in the alpha band the modulations of flexibility covered areas
related to a visuomotor network, whereas this was less evident in the beta band. Studies have
shown that the coherence between motor and visual areas over centro-parietal and occipital
regions in the alpha and beta band increases during the execution of visuomotor tasks (Classen
et al., 1998; Erla et al., 2012), suggesting a functional link between these areas subserving
sensorimotor integration. In addition, a higher coherence between these regions was associ-
ated with better performance during visuomotor tracking tasks (Rilk et al., 2011). However,
these works say nothing about how connectivity varies with long-term learning but only pro-
vide information about the frequency ranges in which these modulations are supposed to
occur. In contrast to these studies, we uncovered network adaptations across a continuum
of learning and showed how the transition from more constrained toward more flexible
dynamics happens in the alpha and, although less consistently, beta ranges. Finally, we
observed unpredicted increases over somatomotor and centro-parietal areas in the theta
and gamma bands. There is no specific evidence for the implication of somatomotor regions
in the theta band, but our results are coherent with previous findings pointing to a reduced
connectivity between frontal and central areas with ongoing visuomotor learning (Gentili
et al., 2015). As for the gamma band, there is no concrete evidence highlighting a change
in the dynamics of centro-parietal areas with visuomotor learning and our findings are not
consistent with a recent study showing a gamma-related decrease in modularity following
visuomotor adaptation training (Miraglia et al., 2018).
An important result of our work shows that some features of brain network dynamics mea-
sured in the very early stages of practice correlated very well with performance measured in
independent sessions recorded over six weeks of practice. The most consistent results were
found over prefrontal areas in the beta frequency band, including orbitofrontal, parsorbitalis,
and rostral middle frontal and anterior cingulate. Higher flexibility values were strongly cor-
related with faster reduction of MT and thus, as suggested in previous work (Bassett et al.,
2015; Wymbs & Grafton, 2015), faster learning. Previous fMRI studies have consistently shown
that brain network flexibility correlates well with cognitive flexibility and learning (Bassett
et al., 2011, 2013b; Braun et al., 2015; Reddy et al., 2018). In the context of simple motor
skill learning, Bassett et al. (2011) showed that the amount of flexibility measured in one
specific session correlated well with the amount of learning in the following practice session.
Similarly, Bassett et al. (2013b) showed that the core-periphery geometry derived from each
Network Neuroscience
590
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
individual flexibility distribution during early learning correlated well with learning in the fol-
lowing 10 days of training. Later, not directly addressing brain network flexibility, Bassett et al.
(2015) showed that a consistent and gradual disengagement of prefrontal regions correlated
with better learning. Our results are in line with these findings and go further by suggesting that
early characteristics of prefrontal dynamics may be related to learning quality. We have
noticed an absence of long-term longitudinal studies addressing (visuo)motor skill learning
with EEG and lack of flexibility analysis that could be directly related to our work. However,
our findings seem consistent with several lines of evidence from studies of visuomotor learning
during periods of resting state. Consistent with our results and specific to the beta frequency
range, Wu et al. (2014) have shown that lower resting-state connectivity between motor and
left prefrontal areas was related to greater skill acquisition during subsequent training on a
pursuit motor task. Although difficult to confirm without direct analysis, we speculate that
lower connectivity between regions in these areas could translate into a more flexible modular
composition over time and thus higher flexibility. In a similar study investigating arm reaching
in a force mediated field, Faiman et al. (2018) evidenced a positive relationship between
resting-state motor-prefrontal connectivity in the alpha band and performance. Their findings
seem inconsistent with our results and with previous work (Wu et al., 2014). However, a point
that is important to emphasize is the fact that the outcomes of Faiman and colleagues were
obtained from analyses at scalp level, a factor that could explain their inconsistency. The
implication of orbitofrontal and middle frontal areas in the beta frequency range is supported
by evidence from other studies assessing brain dynamics during performance. A reduction in
medial frontal beta band activity (e.g., higher activation of the underlying area) during visual
rotation was interpreted as the inhibition of automatic motor responses in favor of cognitively
controlled movements (Jahani et al., 2020). We can speculate that a bias toward more cogni-
tively controlled action may increase activity and connectivity between the areas involved,
reducing temporal variability and thus flexibility. Finally, other studies have also shown rela-
tionships between brain dynamics at rest and visuomotor performance in other frequency
bands, for example, alpha (Manuel et al., 2018) and theta (Miraglia et al., 2018) bands,
although these were not revealed by our study.
In conclusion, this work is the first attempt to characterize brain network dynamics in the
context of motor skill learning by leveraging on a longitudinal design made up of several EEG
recordings interspersed with long training periods. We confirmed previous evidence suggest-
ing that with ongoing learning, cortical regions’ activity shifts from a constrained toward a less
constrained, more flexible dynamics. In addition, we complemented traditional fMRI studies
by providing access to the frequencies at which these changes occurred. Specifically, we have
shown that the gradual disengagement of prefrontal regions highlighted in previous work is
specific to cross-region interactions within frequencies from theta to alpha ranges, and that
previous evidence on learning-related reduction of integration between sensorimotor and
visual areas finds consistent support from the dynamics of these regions within the alpha fre-
quency band. Importantly, we highlighted a relationship between the early dynamics of some
prefrontal regions in the beta band and motor skill learning. This suggests some basic organi-
zational principles and constraints of the brain already visible at the onset of learning.
Methodological Considerations and Limitations
Several important methodological and conceptual considerations are pertinent to this work.
First, some details need to be discussed regarding the choice of experimental protocol used
in this study, and the possibility that the observed behavioral and physiological outcomes are
(at least in part) not directly linked to the experimental manipulation (in our case, practice of
Network Neuroscience
591
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
specific sequences and their learning). The experimental protocol used in this work is the same
as that used by Bassett et al. (2013a, 2013b) and by other authors following their first publi-
cation (Bassett et al., 2015; Mattar et al., 2018; Reddy et al., 2018; Wymbs & Grafton, 2015).
This choice was based on the compelling need to replicate recording procedures and analyses
as consistently as possible with previous studies using the same experimental manipulation.
Although the experimental protocol was not designed to include active control group or an
additional experimental control condition, its design includes the presence of a within-
participant factor through which the learning intensity is manipulated (on three levels: MIN,
MOD, and EXT) in order to control for nonspecific familiarity effects due to the amount of time
spent performing the experiment. For this reason, two sequences were practiced extensively,
two occasionally, and two rarely throughout the training regime. This choice is justified by the
evidence that the total amount of prior practice, rather than chronologic time, is the primary
determinant of the magnitude and location of sequence-specific representations (Wymbs &
Grafton, 2015). Therefore, there are valid reasons to assume that dynamics of changes in
flexibility was largely related to learning and not necessarily due to the participant or the
experimenter merely becoming accustomed to the experimental setting, or due to other
changes relating to the passage of time or to general exposure to task practice. Indeed, the
interaction effect between EEG session and training intensity level (MIN, MOD, and EXT)
revealed a higher increase in flexibility with EEG sessions for the extensively practiced
sequences than for the less practiced ones. During the last EEG scan, higher flexibility was
observed during the practice of the EXT sequences compared to the MIN sequences, although
the participants in the preceding weeks had received the same exposure to the task and exper-
imental settings. However, the absence of an active group/control condition (designed in such
a way as not to require learning—but with similar difficulties and manipulations of the training
regime as in the experimental condition) poses the need to maintain relative caution regarding
the effects of the experimental intervention presented in this and previous studies. To defini-
tively exclude the possibility that the observed effects are due to factors external to learning,
future studies should consider the inclusion of an additional control condition/group (e.g., via
the deployment of simple motor tasks). Nevertheless, we can already anticipate the difficulty in
reaching such a compromise given that the additional control group/condition should be
designed to solicit cognitive-motor processes comparably to the experimental condition, as
well as match in difficulty and motivation for the participants, all of which are extremely dif-
ficult factors to control.
Second, a further point of discussion relates to the use of flexibility as a quantitative mea-
sure to describe functional brain dynamics and its link with learning. The use of this parameter
allowed us to compare the results obtained in this study with those obtained in previous fMRI
studies with similar protocols. However, an inherent limitation of this measure is that it does
not provide the level of detail required to infer the functional mechanisms that generate the
observed variations in flexibility. For instance, the same flexibility value can be generated by a
region changing, with a certain frequency, affiliation to only two or more communities. This is
complicated by the fact that the size and spatial location of each community obtained through
modularity maximization is dynamic and therefore varies over time. Future studies could focus
effort on research and implementation of quantitative measures possibly complementary to the
approach followed in this work such as metrics related to network segregation (such as
clustering coefficient and recruitment), integration (such as participation coefficient), and
hubness (such as betweenness centrality). Moreover, measures such as global and nodal
efficiency could also be implemented to test a plausible hypothesis that learning favors faster
communication and less effort in transferring information. A final consideration must be made
Network Neuroscience
592
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
with respect to the scale resolution of analysis presented in this work, as this study focused on
dynamic variations occurring at a larger time scale corresponding to the time period of task
execution. However, the available dataset would in principle allow adapting existing analysis
procedures to enable a finer scale analysis of the dynamics occurring at the scale of the indi-
vidual trial. This possibility might uncover additional features that would enhance understand-
ing of functional network-based predictors of learning phenomena.
Third, with respect to the computation of functional connectivity to build multilayer mod-
ularity tensors, it is important to note that the dynamic connectivity matrices computed in each
trial (windowed phase locking values, PLVs) were not used in our work other than to calculate
the values of the static PLVs in each trial. It is in fact the latter that were used as inputs for the
construction of the multilayer network tensors for community detection. An equivalent and
computationally more advantageous alternative would have been to calculate the static PLVs
directly using the entire signal within each trial, thus avoiding the intermediate step involving
the calculation of the wind owed PLVs. Nevertheless, our choice to go through the additional
step of calculating the windowed PLVs has the advantage that the database that we created
could be used as a basis for more detailed analyses (e.g., for questions specific to temporal
dynamics at the level of the individual trial) in future studies, with the possibility of comparing
in a more consistent manner future findings with the results obtained in this work.
Fourth, with respect to the algorithm used to calculate global and regional flexibility, we
replicated in this work the methodological steps described in previous works that used the
same or similar protocols to the one used in Bassett et al. (2011, 2013b) and Betzel et al.
(2017). Specifically, as described in the Construction of Dynamic Networks, Community
Detection, and Flexibility Estimates section, the flexibility at the subject level was computed
by averaging the flexibility values obtained from different partitions obtained by multiple max-
imizations of the multilayer modularity function (this was done for each session, training inten-
sity level, and frequency band). This approach made it possible to limit near degeneracies in
the modularity landscape and to obtain flexibility estimates that were consistent and compa-
rable with those obtained in the previous studies cited above. However, several valid alterna-
tives may exist to the procedure implemented here. These include the possibility of using a
consensus partition from the realizations obtained from the maximization process. In this
sense, a recent study has proposed the use of a generative modeling approach called weighted
stochastic clock models as an effective tool to describe different types of community structure
in brain networks that goes beyond simple modularity (Faskowitz et al., 2018). Consensus
partitions and flexibility measures can be estimated using this method. This method was suc-
cessfully applied to study changes in the human connectome that occur across the life-span,
and has indeed great potential to be implemented on longitudinal datasets like ours to identify
how community structure regimes relate to aspects of behavior and cognition, such as
learning.
Fifth, our sample of young adults has, out of 30 participants, 25 female volunteers. Testing
mainly females makes the generalization of our results slightly less robust, although there are
premises in the literature suggesting no advantage for either female or male adults. First,
previous studies using the same task as ours reported no gender differences in the variables
studied (Bassett et al., 2011, 2013b, 2015; Mattar et al., 2018; Reddy et al., 2018; Wymbs &
Grafton, 2015). Second, and with respect to various contexts of motor performance, evidence
in the literature has reported an advantage for women in fine-motor tasks (e.g., pegboard tasks,
handwriting: Berninger et al., 2008; Bornstein, 1986), an advantage for men in speed execu-
tion but not in accuracy of purely motor tasks (e.g., finger tapping: Gur et al., 2010; Hausmann
et al., 2004; Nicholson & Kimura, 1996; Ruff & Parker, 1993), but no clear advantage for either
Network Neuroscience
593
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
male or female adults in performance of sensorimotor tasks (e.g., Gur et al., 2010, 2012). In
addition, evidence on performance of professional musicians during sequential motor skill
tasks like our DSP task (e.g., Kincaid et al., 2002) did not reveal any gender differences in
either professional musicians or controls. Finally, control on gender by introducing it as a
between-subject factor in our analyses of variance did not reveal any significant difference
in MT and learning parameter estimates. However, these analyses should be interpreted con-
sidering the small sample and the unequal number of subjects in each factor’s level.
Finally, we explored the possible confounders related to two additional factors—EEG signal
strength and task design—and were able to put some elements into perspective (see section 4
of the Supporting Information). First, we have shown that the flexibility results obtained
through phase-related analyses of the EEG signal are independent of the amplitude content
of the signal itself (Supporting Information section 4.1). Second, we have highlighted that flex-
ibility was correlated with variables related to the implicit structure of the task (e.g., trial
length). We have suggested that properties of multilayer networks and flexibility may have a
common origin due to reduced signal length with learning, although it was impossible to infer
a causal role between those variables. In this regard, we have provided additional analysis
(Supporting Information sections 4.2.1 and 4.2.2) to take these variables into account but
are aware of the multiple limitations of this approach. Among these, the most critical is the
loss of a significant amount of EEG signal due to the proposed thresholding procedure to resize
the trials to the same length. Nevertheless, despite these limitations, the results obtained are
in partial overlap with the main findings, which is reassuring considering previous work on
this topic (Bassett et al., 2013b, 2015; Reddy et al., 2018). However, we argue that the ele-
ments raised here must be seriously taken into consideration, as they represent an interesting
opening for future work: because of the difficulty to disentangle mechanisms that compete
with the type of tasks typically developed and used in the learning paradigms, we flag as a
priority the development and use of new experimental protocols that are tailored to the meth-
odology used.
MATERIALS AND METHODS
Participants
Thirty healthy volunteers (25 females; mean age = 22.2 years, SD = 6.2 years, range 18 to
50 years) gave written informed consent to participate in the experiment and were compen-
sated with credits for their participation. All participants reported normal or corrected-to-
normal vision acuity and a right-hand preference (84 ± 22) (Oldfield, 1971). Research was
approved by the Cantonal Ethics Committee for Human Research ( Vaud, Switzerland; protocol
N°2019/02345) and was in accordance with the code of ethics of the World Medical Associ-
ation (Declaration of Helsinki) for experiments involving human subjects in research.
Experimental Setup and Procedure
We used the same experimental protocol developed by Bassett et al. (2013b) and previously
employed in a large variety of studies (Bassett et al., 2015; Mattar et al., 2018; Wymbs &
Grafton, 2015). The protocol (Figure 1A) is a longitudinal study consisting of home and
laboratory sessions. Participants practiced 30 home training sessions of a discrete sequence-
production task (DSP) over 42 days at a rate of 10 sessions every 14 days. In addition, they
were tested on the same DSP task during four laboratory sessions where EEG was recorded.
The first laboratory session was planned at the beginning of the study, and the remaining three
every 14 days after 10 consecutive home sessions.
Network Neuroscience
594
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
During home sessions (Figure 1B) participants practiced a total of 150 DSP task trials, consist-
ing of two sequences (EXT1 and EXT2) each presented for 64 trials (extensively trained
sequences; EXT), two sequences (MOD1 and MOD2) for 10 trials (moderately trained
sequences; MOD), and two additional sequences (MIN1 and MIN2) for 1 trial (minimally
trained sequences; MIN). Each sequence was made of 10 sequential visual displays. Each visual
display contained a horizontal array of five square stimuli, each corresponding to one finger (from
left to right, the thumb and the pinky were placed on the keyboard’s spacebar and on the letter L
and corresponded to the leftmost and rightmost square, respectively; the index, middle, and ring
fingers on the letters H, J, and K). In each display, one of the five squares had a red outline, indi-
cating which key to press. Immediately after pressing the correct key, the display changed, and
another square switched its outline to red. Participants could only advance the sequence by
pressing the correct keypress and had unlimited time to complete it as accurately and quickly
as possible. In case of error, an error message appeared on the screen for 100 ms, and the
sequence was paused at the error and restarted upon the appropriate key press. Each sequence
was conceived such that each of the five square stimuli switched twice its outline to red, and to
avoid immediate repetitions (e.g., “1-1”) and regularities (e.g., “4-5-4” or “4-3-2”) across consec-
utive visual displays. Each trial started with the presentation of a cue at the center of the screen for
500 ms (red and blue circle for the EXT sequences, green and yellow triangles for the MOD
sequences, white and black stars for the MIN sequences), followed by the first of the 10 visual
displays of the sequence. The end of the sequence was followed by a 200-ms screen-centered
fixation cross, and an intertrial blank screen varying in duration between 500 and 1,500 ms. For
every block of 10 trials, feedback about the number of error-free sequences and mean time to
complete the error-free sequences was presented for 3,000 ms right after the fixation cross. In
these trials, the feedback was followed by a random intertrial interval lasting between 0 and
1,000 ms. EXT, MOD, and MIN trials were randomly distributed throughout the task.
During EEG sessions (Figure 1B) participants practiced EXT, MOD, and MIN sequences for
50 trials each, for a total of 300 trials. The trial design was the same as in the home sessions.
However, unlike the home sessions, the trials were not presented randomly but evenly distrib-
uted over five epochs of 60 trials each (i.e., each epoch contained the same number of EXT,
MOD, and MIN trials). Each epoch contained two blocks of 10 EXT trials (five trials for each of
the two EXT sequences), two blocks of MOD trials (five trials for each of the two MOD
sequences), and two blocks of MIN trials (five trials for each of the two MIN sequences). Trials
were randomized within each block and blocks randomized within each epoch.
The DSP task practiced during home sessions was coded with PsychoPy (Peirce et al., 2019)
in JavaScript and was made available in the form of a Web-based task that participants could
access from their personal computer. Participants were asked not to perform close sessions
and to keep the same daily schedule and environment as much as possible for home practice.
The DSP task practiced during laboratory sessions was coded with Psychopy and run on the
laboratory computers. Stimuli were synchronized with the EEG hardware by means of TTL-
triggers marking the EEG signal with task-related events (e.g., visual displays onsets). During
these sessions, participants were seated in a quiet room sheltered from electromagnetic distur-
bances (Faraday cage). Stimulus size and participant’s distance from the screen were adjusted so
that the five square stimuli of the DSP task covered a horizontal length of 7° visual angle. To limit
EEG artifacts due to head movements, the head of participants was placed on a chin rest.
Behavioral Estimates of Learning
Previous works using the same experimental procedure (Bassett et al., 2013b, 2015; Mattar
et al., 2018; Wymbs & Grafton, 2015) have quantified behavioral learning with movement
Network Neuroscience
595
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
time (MT), defined as the time that elapses between the first and last button press during the
execution of a sequence. The decrease in MT with practice has often been used to quantify
learning (Crossman, 1959; Stratton et al., 2007). Various functional forms have been used to
model this decrease and the most robust choices have often been made in favor of (variant of )
exponential functions (Heathcote et al., 2000). To quantify the decrease of MT with practice,
we used in this work an approach similar to previous studies which modeled the MTs associ-
ated to EXT sequences using a double exponential function (Bassett et al., 2013b, 2015; Mattar
et al., 2018; Wymbs & Grafton, 2015). This choice was motivated by the vastly superior
number of practiced trials in EXT sequences. First, MTs were calculated for each sequence
performed in the home sessions during the 42 days of practice. The MT was computed from
the first to the last button press of the 10 displays. In case of error, the time spent visualizing the
error display was not included into the computation of MT. The MTs of the two EXT sequences
were then averaged together for each of the 30 home-based sessions, resulting in 30 MT values
for each participant. These data were then fitted with a double exponential function of the
form (Equation 1):
MT ¼ aetK þ betλ;
(1)
where t is time, K 2 (−∞, 0] is the exponential drop-off parameter describing the fast rate of
improvement (typically occurring during the first days of practice), λ 2 (−∞, 0] is the exponen-
tial drop-off parameter describing the slow, sustained rate of improvement, and a and b 2 [0,
+∞) are constants. The parameter K, which is independent of the initial and final individual’s
performance, was used as an indicator of the speed of learning as in previous works (Bassett
et al., 2015; Dayan & Cohen, 2011): curves with high absolute values of K are characterized
by a higher improvement of MT per unit of time (i.e., decrease more rapidly) than those
described by smaller values. In addition to the learning rate K it was also computed, for each
participant, the initial and final MT (average MT computed from EXT sequences in the first and
last home training session).
EEG Recording, Preprocessing, and Functional Connectivity Estimation
Continuous EEG was obtained with a Biosemi ActiveTwo system using 128 active electrodes
placed at the international radial ABC system location and two pairs of bipolar electrodes
recorded electrooculography in both vertical and horizontal directions. Two additional elec-
trodes (active CMS: common mode sense; and passive DRL: driven right leg) were used as
reference and ground to compose a feedback loop for amplifier reference. Data sampling
was set at 1024 Hz with 24-bit A/D conversion. EEG data were acquired at each EEG session
during performance on the DSP task, and during two 5-min resting-state periods occurring at
the beginning and at the end of the experimental session (the reader should refer to Figure 2 for
a visual representation of the methodological steps described in the following paragraphs).
We first band-pass filtered the EEG signals (1–45 Hz) and we extracted trial epochs from the
onset of the first display to the offset of the last sequence display, with a 500-ms prestimulus
baseline. We removed eye blinks using the EOG regression method (i.e., use the EOG chan-
nels to clean the 128 EEG channels) (Parra et al., 2005). Channels with high standard deviation
and flat waveform were identified as bad channels and interpolated using the spherical spline
method as implemented in eeglab (Delorme & Makeig, 2004). Signals were then re-referenced
to an average reference montage. We then computed several quality check parameters to rank
the preprocessed epochs from high to lower quality, namely the Overall High Amplitude, the
Time High Variance, and the Channel High Variance. All these metrics are described in Auto-
magic (Pedroni et al., 2019). This was used as an additional step to remove bad trials with very
Network Neuroscience
596
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
low-quality metrics. We finally performed an additional visual inspection of the trials based on
topographies displayed at the max of the global field power within each trial.
After preprocessing the EEGs, the functional connectivity matrices were estimated in each
trial using the EEG source-connectivity method (Hassan & Wendling, 2018; Schoffelen &
Gross, 2009). First, the dynamics of cortical brain sources were reconstructed by solving the
inverse problem. To do so, EEG channel locations and MRI template (ICBM152) were coregis-
tered, and using the OpenMEEG toolbox (Gramfort et al., 2010), a realistic head model based
on the boundary element method with three layers (scalp, skull, and brain) was built. The
weighted minimum norm estimate (wMNE) was used to estimate the brain sources on a surface
mesh of 15,002 vertices. The regularization parameter is set to 0.3 in our study. The noise
covariance matrix was computed from a 500-ms prestimulus baseline. In this work, we used
the MATLAB function implemented in Brainstorm toolbox (Tadel et al., 2019) to compute
wMNE, with the signal to noise ratio set to 3 and depth weighting value to 0.5 (default values).
The regional time series of the 68 cortical regions of interest of the Desikan–Killiany atlas were
computed by averaging the activity of sources included in each region of interest (Figure 2A).
Afterward, we filtered the regional time series in different EEG frequency bands: theta: 4–
8 Hz; alpha: 8–12 Hz; beta: 12–28 Hz; gamma: 28–45 Hz. For each frequency band, we com-
puted functional connectivity using the phase locking value (PLV), a measure for assessing
phase synchrony between two signals in a particular frequency band, formed from estimates
of the instantaneous phase of the signal. More specifically, we inferred connectivity at each
trial by applying a sliding window approach where connectivity was computed within each
temporal window (dynamic or windowed PLVs) and then averaged across windows to obtain a
single connectivity matrix (static PLV). This was done by taking into account the recommen-
dations of Lachaux et al. (1999) concerning the choice of smallest window length to guarantee
a sufficient number of cycles per window at the given frequency band. This minimum window
length equals 6 divided by the central frequency of each frequency band, where the number 6
corresponds to the smallest number of cycles recommended by Lachaux et al. (1999). We
therefore ended up, for each frequency band, with one static PLV for each trial (Figure 2B).
These static PLVs from the task trials of each session were used in the following analyses as
inputs for the construction of the multilayer network tensors for community detection
(Figure 2C).
Construction of Dynamic Networks, Community Detection, and Flexibility Estimates
We characterized brain networks in terms of global cohesive functional structures that could
capture relevant dynamics underlying motor skill learning occurring at time scales of several
weeks of practice. We first constructed multilayer network tensors A (Mucha et al., 2010),
representing time-dependent networks (Figure 2C), by temporally combining the static con-
nectivity matrices (static PLVs) from L consecutive trials of a given sequence type (EXT,
MOD, and MIN). This was done, in each frequency band, for each participant and EEG ses-
sion. No thresholding was applied to the static PLVs. Starting from these multilayer network
tensors, we then identified functional communities (or modules) composed of brain regions
exhibiting similar functional activity through time. To this end, we performed community
detection (Fortunato, 2010; Porter et al., 2009) on each multilayer network tensor in order
to identify groups of brain regions that have a strong connection to each other (and preserved
across temporal layers) with respect to brain regions assigned to other groups (Figure 2D).
To obtain functional communities we opted for the method of modularity maximization
(Mucha et al., 2010) through the use of the Louvain-like locally greedy algorithm (Blondel
Network Neuroscience
597
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
et al., 2008), recently adapted and used in the context of studies linking multilayer networks to
motor skill learning (Bassett et al., 2011, 2013a, 2013b, 2015). Modularity maximization con-
sists in the optimization of a multilayer modularity quality function Q (Equation 2) defined as
in Mucha et al. (2010):
Q ¼
1
2μ
X
(cid:4)
(cid:2)
ijlr Aijl − γlMijl
(cid:3)
δlr þ δij ωjlr
(cid:5)
(cid:2)
(cid:2)
δ gil; gjr
(cid:3)
(cid:3)
;
(2)
where μ is the sum of edge weights across nodes and layers, δij equals 1 when i = j and equals
0 otherwise, and δlr equals 1 when l = r and equals 0 otherwise. Aijl is an element of the mul-
tilayer network tensor A, representing the strength of the connectivity edge between node i and
j in layer l, while Mijl is the strength expected from a Newman–Girvan null model (Bassett
et al., 2013a); γl is the structural resolution parameter of layer l (intralayer coupling parameter),
the quantity gil and gjr give the community assignment of node i and j in layer l and r, respec-
tively; (δ( gil, gjr) equals 1 when gil = gjr, 0 otherwise). And ωjlr is the interlayer coupling param-
eter (often referred as temporal resolution parameter) representing the connection strength
from node j in layer l to node j in layer r.
We performed multilayer modularity maximization using the generalized Louvain package
in MATLAB implemented with the modularity probability method (MPM) algorithm (for details
on how the MPM algorithm is implemented to detect and merge communities, see Bazzi et al.,
2016; Yang et al., 2021), with γ = ω = 1 (Bassett et al., 2011, 2013a, 2015). To overcome near
degeneracies in the modularity landscape, which is expressed in a different community par-
tition at each run of the algorithm optimizing Q, we avoid focusing on a single algorithmic run
(Bassett et al., 2013a). Instead, following the approach proposed in previous works that have
also used flexibility as a quantitative metric for their analyses (Bassett et al., 2013b; Betzel
et al., 2017), we optimized each multilayer modularity quality function Q 100 times and com-
puted regional and global network flexibility for each algorithmic run and subsequently aver-
aged over the 100 runs. We computed regional network flexibility for each of the brain regions
defined by the Desikan–Killiany template (Figure 2E). It is an indicator of the stability of func-
tional communities, derived from the optimization of the quality function Q, across temporal
layers. Specifically, it quantifies the fraction of times that a specific brain region changes
assignment to a community in successive temporal layers, normalized on the total number
of possible changes. A value close to 0 indicates great consistency in community assignment
over time (i.e., over the course of a scan session for a specific sequence type: EXT, MOD, or
MIN), while a value close to 1 indicates great variability. Global network flexibility was com-
puted by averaging regional flexibility across all brain regions. Regions in which significant
changes in flexibility are observed are named using the Desikan–Killiany template. Through-
out the text, an indication of the possible brain networks with which they are associated is also
provided (Thomas Yeo et al., 2011).
Statistical Modeling
Using a two-way repeated measures analysis of variance (ANOVA), we tested the effect of
laboratory sessions (session 1 to 4) and training intensity (MIN, MOD, EXT) on the MT (Behav-
ioral Performance section and Supporting Information section 1), the global flexibility estimates
(Global and Regional Flexibility Variation With Ongoing Practice section and Supporting Infor-
mation section 2: quantification of global flexibility) and power estimates (Supporting Infor-
mation section 4: relationship between EEG signal power and flexibility estimates) in each
frequency band. We applied a correction of the F values in case of violation of sphericity
assumption (Mauchly’s test) using a Greenhouse–Geisser (if Greenhouse–Geisser epsilon <
Network Neuroscience
598
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
n
e
n
_
a
_
0
0
2
8
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network dynamics underlying motor skill learning
0.75) or a Huynh–Feldt (if Greenhouse–Geisser epsilon > = 0.75) correction. We interpreted
significant interactions using pairwise comparisons and applied Bonferroni corrections to
address the problem of multiple comparisons. To identify the cortical regions showing signif-
icant regional flexibility values, we used a one-tailed Wilcoxon test and ran Bonferroni and
false discovery rate corrections to address the problem of multiple comparisons. Endlich, Wir
used Spearman’s rank correlation as a measure of statistical relationship between flexibility
and behavior (Functional Correlates of Performance section), power estimates (Supporting
Information section 4), and variables used to characterize task design (zusätzliche Informationen
section 4). This nonparametric statistic measures the monotonic relationship between two var-
iables without a requirement for linearity. ANOVAs were performed with The Jamovi Project
(2021), Jamovi ( Version 1.0.7.0) (Computer Software) retrieved from https://www.jamovi.org.,
n.d.; all other analyses were performed in MathWorks MATLAB R2020b implementing custom
Code.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00289.
Data and code are made available on the open access repository Zenodo (Ruggeri, 2022).
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
BEITRÄGE DES AUTORS
Paolo Ruggeri: Konzeptualisierung; Datenkuration; Formale Analyse; Akquise von Fördermitteln; Inves-
tigation; Methodik; Projektverwaltung; Software; Aufsicht; Visualisierung; Writing –
original draft; Writing – review & Bearbeitung. Jenifer Miehlbradt: Datenkuration; Untersuchung;
Methodik; Projektverwaltung; Software; Writing – review & Bearbeitung. Aya Kabbara: Für-
mal analysis; Writing – review & Bearbeitung. Mahmoud Hassan: Konzeptualisierung; Formal anal-
ysis; Methodik; Projektverwaltung; Software; Visualisierung; Writing – original draft;
Writing – review & Bearbeitung.
FUNDING INFORMATION
Paolo Ruggeri, Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen
Forschung (https://dx.doi.org/10.13039/501100001711), Award ID: CRSK-1_190830.
VERWEISE
Aliakbaryhosseinabadi, S., Lontis, R., Farina, D., & Mrachacz-
Kersting, N. (2021). Effect of motor learning with different
complexities on EEG spectral distribution and performance
improvement. Biomedical Signal Processing and Control, 66,
102447. https://doi.org/10.1016/j.bspc.2021.102447
Allen, E. A., Damaraju, E., Plis, S. M., Erhardt, E. B., Eichele, T., &
Calhoun, V. D. (2014). Tracking whole-brain connectivity
dynamics in the resting state. Hirnrinde, 24(3), 663–676.
https://doi.org/10.1093/cercor/bhs352, PubMed: 23146964
Andres, F. G., & Gerloff, C. (1999). Coherence of sequential move-
ments and motor learning. Journal of Clinical Neurophysiology,
16(6), 520–527. https://doi.org/10.1097/00004691-199911000
-00004, PubMed: 10600020
Aoki, F., Fetz, E. E., Shupe, L., Lettich, E., & Ojemann, G. A. (2001).
Changes in power and coherence of brain activity in human sen-
sorimotor cortex during performance of visuomotor tasks.
Biosystems, 63(1-3), 89–99. https://doi.org/10.1016/S0303
-2647(01)00149-6, PubMed: 11595332
Barabási, A.-L. (2013). Network science. Philosophische Transaktionen
of the Royal Society A: Mathematical, Physical, and Engineering
Wissenschaften, 371(1987), 20120375. https://doi.org/10.1098/rsta
.2012.0375, PubMed: 23419844
Bassett, D. S., Porter, M. A., Wymbs, N. F., Grafton, S. T., Carlson,
J. M., & Mucha, P. J. (2013A). Robust detection of dynamic com-
munity structure in networks. Chaos: An Interdisciplinary Journal
of Nonlinear Science, 23(1), 013142. https://doi.org/10.1063/1
.4790830, PubMed: 23556979
Bassett, D. S., & Spurns, Ö. (2017). Network neuroscience. Natur
Neurowissenschaften, 20(3), 353–364. https://doi.org/10.1038/nn.4502,
PubMed: 28230844
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of human
Netzwerkneurowissenschaften
599
Brain network dynamics underlying motor skill learning
brain networks during learning. Verfahren des Nationalen
Akademie der Wissenschaften, 108(18), 7641–7646. https://doi.org/10
.1073/pnas.1018985108, PubMed: 21502525
Bassett, D. S., Wymbs, N. F., Rombach, M. P., Porter, M. A., Mucha,
P. J., & Grafton, S. T. (2013B). Task-based core-periphery organi-
zation of human brain dynamics. PLoS Computational Biology,
9(9), e1003171. https://doi.org/10.1371/journal.pcbi.1003171,
PubMed: 24086116
Bassett, D. S., Yang, M., Wymbs, N. F., & Grafton, S. T. (2015).
Learning-induced autonomy of sensorimotor systems. Natur
Neurowissenschaften, 18(5), 744–751. https://doi.org/10.1038/nn.3993,
PubMed: 25849989
Bazzi, M., Porter, M. A., Williams, S., McDonald, M., Fenn, D. J., &
Howison, S. D. (2016). Community detection in temporal multi-
layer networks, with an application to correlation networks.
Multiscale Modeling & Simulation, 14(1), 1–41. https://doi.org
/10.1137/15M1009615
Berninger, V. W., Nielsen, K. H., Abbott, R. D., Wijsman, E., &
Raskind, W. (2008). Gender differences in severity of writing
and reading disabilities. Journal of School Psychology, 46(2),
151–172. https://doi.org/10.1016/j.jsp.2007.02.007, PubMed:
19083355
Betzel, R. F., Satterthwaite, T. D., Gold, J. ICH., & Bassett, D. S. (2017).
Positive affect, surprise, and fatigue are correlates of network
flexibility. Wissenschaftliche Berichte, 7(1), 520. https://doi.org/10.1038
/s41598-017-00425-z, PubMed: 28364117
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., & Lefebvre, E.
(2008). Fast unfolding of communities in large networks. Zeitschrift
of Statistical Mechanics: Theory and Experiment, 2008(10),
P10008. https://doi.org/10.1088/1742-5468/2008/10/P10008
Boonstra, T. W., Daffertshofer, A., Breakspear, M., & Beek, P. J.
(2007). Multivariate time-frequency analysis of electromagnetic
brain activity during bimanual motor learning. NeuroImage,
36(2), 370–377. https://doi.org/10.1016/j.neuroimage.2007.03
.012, PubMed: 17462913
Bornstein, R. A. (1986). Normative data on intermanual differences
on three tests of motor performance. Journal of Clinical and
Experimental Neuropsychology, 8(1), 12–20. https://doi.org/10
.1080/01688638608401293, PubMed: 3944242
Braun, U., Schäfer, A., Walter, H., Erk, S., Romanczuk-Seiferth, N.,
Haddad, L., Schweiger, J. ICH., Grimm, O., Heinz, A., Tost, H.,
Meyer-Lindenberg, A., & Bassett, D. S. (2015). Dynamic recon-
figuration of frontal brain networks during executive cognition in
humans. Verfahren der Nationalen Akademie der Wissenschaften,
11 2 ( 3 7 ) , 11 6 7 8 –11 6 8 3 . h t t p s : / / d o i . o r g / 1 0 . 1 0 7 3 / p n a s
.1422487112, PubMed: 26324898
Classen, J., Gerloff, C., Honda, M., & Hallett, M. (1998). Integrative
visuomotor behavior is associated with interregionally coherent
oscillations in the human brain. Journal of Neurophysiology,
79(3), 1567–1573. https://doi.org/10.1152/jn.1998.79.3.1567,
PubMed: 9497432
Crivelli-Decker, J., Hsieh, L.-T., Clarke, A., & Ranganath, C. (2018).
Theta oscillations promote temporal sequence learning. Neurobi-
ology of Learning and Memory, 153, 92–103. https://doi.org/10
.1016/j.nlm.2018.05.001, PubMed: 29753784
Crossman, E. R. F. W. (1959). A theory of the acquisition of
speed-skill. Ergonomics, 2, 153–166. https://doi.org/10.1080
/00140135908930419
Cunha, M., Machado, D., Bastos, V. H., Ferreira, C., Cagy, M.,
Basile, L., Piedade, R., & Ribeiro, P. (2006). Neuromodulatory
effect of bromazepam on motor learning: An electroencephalo-
graphic approach. Neurowissenschaftliche Briefe, 407(2), 166–170.
https://doi.org/10.1016/j.neulet.2006.08.028, PubMed:
16959410
Dayan, E., & Cohen, L. G. (2011). Neuroplasticity subserving motor
skill learning. Neuron, 72(3), 443–454. https://doi.org/10.1016/j
.neuron.2011.10.008, PubMed: 22078504
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source tool-
box for analysis of single-trial EEG dynamics including indepen-
dent component analysis. Journal of Neuroscience Methods,
134(1), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009,
PubMed: 15102499
Draganski, B., Gaser, C., Kempermann, G., Kuhn, H. G., Winkler,
J., Büchel, C., & Mai, A. (2006). Temporal and spatial dynamics
of brain structure changes during extensive learning. Zeitschrift für
Neurowissenschaften, 26(23), 6314–6317. https://doi.org/10.1523
/JNEUROSCI.4628-05.2006, PubMed: 16763039
Elton, A., & Gao, W. (2014). Divergent task-dependent functional
connectivity of executive control and salience networks. Kortex,
51, 56–66. https://doi.org/10.1016/j.cortex.2013.10.012,
PubMed: 24315034
Erla, S., Faes, L., Nollo, G., Arfeller, C., Braun, C., & Papadelis, C.
(2012). Multivariate EEG spectral analysis evidences the func-
tional link between motor and visual cortex during integrative
sensorimotor tasks. Biomedical Signal Processing and Control,
7(3), 221–227. https://doi.org/10.1016/j.bspc.2011.08.002
Faiman, ICH., Pizzamiglio, S., & Turner, D. L. (2018). Resting-state
functional connectivity predicts the ability to adapt arm reaching
in a robot-mediated force field. NeuroImage, 174, 494–503.
https://doi.org/10.1016/j.neuroimage.2018.03.054, PubMed:
29588227
Faskowitz, J., Yan, X., Zuo, X.-N., & Spurns, Ö. (2018). Weighted
stochastic block models of the human connectome across the life
Spanne. Wissenschaftliche Berichte, 8(1), 12997. https://doi.org/10.1038
/s41598-018-31202-1, PubMed: 30158553
Floyer-Lea, A., & Matthews, P. M. (2005). Distinguishable brain
activation networks for short- and long-term motor skill learning.
Journal of Neurophysiology, 94(1), 512–518. https://doi.org/10
.1152/jn.00717.2004, PubMed: 15716371
Fortunato, S. (2010). Community detection in graphs. Physics Reports,
486(3–5), 75–174. https://doi.org/10.1016/j.physrep.2009.11.002
Gentili, R. J., Bradberry, T. J., Oh, H., Costanzo, M. E., Kerick, S. E.,
Contreras-Vidal, J. L., & Hatfield, B. D. (2015). Evolution of
cerebral cortico-cortical communication during visuomotor
adaptation to a cognitive-motor executive challenge. Biologisch
Psychologie, 105, 51–65. https://doi.org/10.1016/j.biopsycho
.2014.12.003, PubMed: 25530479
Grafton, S. T., Schmitt, P., Van Horn, J., & Diedrichsen, J. (2008).
Neural substrates of visuomotor learning based on improved
feedback control and prediction. NeuroImage, 39(3), 1383–1395.
https://doi.org/10.1016/j.neuroimage.2007.09.062, PubMed:
18032069
Gramfort, A., Papadopoulo, T., Olivi, E., & Clerc, M. (2010). Open-
MEEG: Opensource software for quasistatic bioelectromagnetics.
BioMedical Engineering OnLine, 9(1), 45. https://doi.org/10.1186
/1475-925X-9-45, PubMed: 20819204
Netzwerkneurowissenschaften
600
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network dynamics underlying motor skill learning
Gur, R. C., Richard, J., Calkins, M. E., Chiavacci, R., Hansen, J. A.,
Bilker, W. B., Loughead, J., Connolly, J. J., Qiu, H., Mentch, F. D.,
Abou-Sleiman, P. M., Hakonarson, H., & Gur, R. E. (2012). Alter
group and sex differences in performance on a computerized
neurocognitive battery in children age 8−21. Neuropsychology,
26(2), 251–265. https://doi.org/10.1037/a0026712, PubMed:
22251308
Gur, R. C., Richard, J., Hughett, P., Calkins, M. E., Macy, L., Bilker,
W. B., Brensinger, C., & Gur, R. E. (2010). A cognitive
neuroscience-based computerized battery for efficient measure-
ment of individual differences: Standardization and initial con-
struct validation. Journal of Neuroscience Methods, 187(2),
254–262. https://doi.org/10.1016/j.jneumeth.2009.11.017,
PubMed: 19945485
Hassan, M., & Wendling, F. (2018). Electroencephalography source
Konnektivität: Aiming for high resolution of brain networks in time
and space. IEEE Signal Processing Magazine, 35(3), 81–96.
https://doi.org/10.1109/MSP.2017.2777518
Hausmann, M., Kirk, ICH., & Corballis, M. (2004). Influence of task
complexity on manual asymmetries. Kortex, 40(1), 103–110.
https://doi.org/10.1016/S0010-9452(08)70923-7, PubMed:
15070005
Heathcote, A., Braun, S., & Mewhort, D. J. K. (2000). The power
law repealed: The case for an exponential law of practice. Psy-
chonomic Bulletin & Rezension, 7(2), 185–207. https://doi.org/10
.3758/BF03212979, PubMed: 10909131
Heinrichs-Graham, E., Arpin, D. J., & Wilson, T. W. (2016). Cue-
related temporal factors modulate movement-related beta oscil-
latory activity in the human motor circuit. Zeitschrift für Kognition
Neurowissenschaften, 28(7), 1039–1051. https://doi.org/10.1162/jocn
_a_00948, PubMed: 26967947
Hikosaka, O., Nakamura, K., Sakai, K., & verstopft, H. (2002).
Central mechanisms of motor skill learning. Aktuelle Meinung in
Neurobiology, 12(2), 217–222. https://doi.org/10.1016/S0959
-4388(02)00307-0, PubMed: 12015240
Holme, P., & Saramäki, J. (2012). Temporal networks. Physik
Reports, 519(3), 97–125. https://doi.org/10.1016/j.physrep.2012
.03.001
Honda, M., Deiber, M. P., Ibáñez, V., Pascual-Leone, A., Zhuang,
P., & Hallett, M. (1998). Dynamic cortical involvement in
implicit and explicit motor sequence learning. A PET study.
Gehirn, 121(11), 2159–2173. https://doi.org/10.1093/ brain/121
.11.2159, PubMed: 9827775
Jahani, A., Schwey, A., Bernier, P.-M., & Malfait, N. (2020). Spa-
tially distinct beta-band activities reflect implicit sensorimotor
adaptation and explicit re-aiming strategy. Journal of Neurosci-
enz, 40(12), 2498–2509. https://doi.org/10.1523/JNEUROSCI
.1862-19.2020, PubMed: 32034068
Jerbi, K., Lachaux, J., Baillet, S., & Garnero, L. (2004). Imaging cor-
tical oscillations during sustained visuomotor coordination in
MEG. 2004 2nd IEEE International Symposium on Biomedical
Imaging: Nano to Macro (IEEE Cat No. 04EX821), 1, 380–383.
https://doi.org/10.1109/ISBI.2004.1398554
Kincaid, A. E., Duncan, S., & Scott, S. A. (2002). Assessment of fine
motor skill in musicians and nonmusicians: Differences in timing
versus sequence accuracy in a bimanual fingering task. Percep-
tual and Motor Skills, 95, 245–257. https://doi.org/10.2466/pms
.2002.95.1.245, PubMed: 12365261
Kivelä, M., Arenas, A., Barthelemy, M., Gleeson, J. P., Moreno, Y., &
Porter, M. A. (2014). Multilayer networks. Journal of Complex
Netzwerke, 2(3), 203–271. https://doi.org/10.1093/comnet
/cnu016
Klimesch, W. (1999). EEG alpha and theta oscillations reflect cog-
nitive and memory performance: A review and analysis. Gehirn
Research Reviews, 29(2-3), 169–195. https://doi.org/10.1016
/S0165-0173(98)00056-3, PubMed: 10209231
Koch, G., Esposito, R., Motta, C., Casula, E. P., Di Lorenzo, F.,
Bonnì, S., Cinnera, A. M., Ponzo, V., Maiella, M., Picazio, S.,
Assogna, M., Sallustio, F., Caltagirone, C., & Pellicciari, M. C.
(2020). Improving visuo-motor learning with cerebellar theta
burst stimulation: Behavioral and neurophysiological evidence.
NeuroImage, 208, 116424. https://doi.org/10.1016/j.neuroimage
.2019.116424, PubMed: 31794855
Lachaux, J.-P., Rodriguez, E., Martinerie, J., & Varela, F. J. (1999).
Measuring phase synchrony in brain signals. Human Brain Map-
ping, 8(4), 194–208. https://doi.org/10.1002/(WISSENSCHAFT )1097
-0193(1999)8:4<194::AID-HBM4>3.0.CO;2-C, PubMed: 10619414
Langer, N., von Bastian, C. C., Wirz, H., Oberauer, K., & Jäncke, L.
(2013). The effects of working memory training on functional
brain network efficiency. Kortex, 49(9), 2424–2438. https://doi
.org/10.1016/j.cortex.2013.01.008, PubMed: 23489778
Manuel, A. L., Guggisberg, A. G., Thézé, R., Turri, F., & Schnider, A.
(2018). Resting-state connectivity predicts visuo-motor skill
learning. NeuroImage, 176, 446–453. https://doi.org/10.1016/j
.neuroimage.2018.05.003, PubMed: 29730496
Mattar, M. G., & Bassett, D. S. (2016). Brain network architecture:
Implications for human learning. ArXiv:1609.01790. https://doi
.org/10.48550/arXiv.1609.01790
Mattar, M. G., Wymbs, N. F., Bock, A. S., Aguirre, G. K., Grafton,
S. T., & Bassett, D. S. (2018). Predicting future learning from
baseline network architecture. NeuroImage, 172, 107–117.
https://doi.org/10.1016/j.neuroimage.2018.01.037, PubMed:
29366697
Mehrkanoon, S., Boonstra, T. W., Breakspear, M., Hinder, M., &
Summers, J. J. (2016). Upregulation of cortico-cerebellar func-
tional connectivity after motor learning. NeuroImage, 128,
252–263. https://doi.org/10.1016/j.neuroimage.2015.12.052,
PubMed: 26767943
Meyer, T., Peters, J., Zander, T. O., Schölkopf, B., & Grosse-
Wentrup, M. (2014). Predicting motor learning performance from
electroencephalographic data. Journal of NeuroEngineering and
Rehabilitation, 11(1), 24. https://doi.org/10.1186/1743-0003-11
-24, PubMed: 24594233
Miraglia, F., Vecchio, F., & Rossini, P. M. (2018). Brain electroen-
cephalographic segregation as a biomarker of learning. Neuronal
Netzwerke, 106, 168–174. https://doi.org/10.1016/j.neunet.2018
.07.005, PubMed: 30075353
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Utopie, J.-P.
(2010). Community structure in time-dependent, multiscale, Und
multiplex networks. Wissenschaft, 328(5980), 876–878. https://doi.org
/10.1126/science.1184819, PubMed: 20466926
Nicholson, K. G., & Kimura, D. (1996). Sex differences for speech
and manual skill. Perceptual and Motor Skills, 82(1), 3–13.
https://doi.org/10.2466/pms.1996.82.1.3, PubMed: 8668494
Oldfield, R. C. (1971). The assessment and analysis of handedness:
The Edinburgh inventory. Neuropsychologie, 9(1), 97–113.
Netzwerkneurowissenschaften
601
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network dynamics underlying motor skill learning
https://doi.org/10.1016/0028-3932(71)90067-4, PubMed:
5146491
O’Neill, G. C., Tewarie, P. K., Colclough, G. L., Gascoyne, L. E.,
Hunt, B. A. E., Morris, P. G., Woolrich, M. W., & Brookes, M. J.
(2017). Measurement of dynamic task related functional networks
using MEG. NeuroImage, 146, 667–678. https://doi.org/10.1016/j
.neuroimage.2016.08.061, PubMed: 27639354
Parra, L. C., Spence, C. D., Gerson, A. D., & Sajda, P. (2005). Rec-
ipes for the linear analysis of EEG. NeuroImage, 28(2), 326–341.
https://doi.org/10.1016/j.neuroimage.2005.05.032, PubMed:
16084117
Pedroni, A., Bahreini, A., & Langer, N. (2019). Automagic: Stan-
dardized preprocessing of big EEG data. NeuroImage, 200,
460–473. https://doi.org/10.1016/j.neuroimage.2019.06.046,
PubMed: 31233907
Peirce, J., Gray, J. R., Simpson, S., MacAskill, M., Höchenberger, R.,
Sogo, H., Kastman, E., & Lindeløv, J. K. (2019). PsychoPy2:
Experiments in behavior made easy. Behavior Research Methods,
51(1), 195–203. https://doi.org/10.3758/s13428-018-01193-y,
PubMed: 30734206
Perfetti, B., Moisello, C., Landsness, E. C., Kvint, S., Lanzafame, S.,
Onofrj, M., Di Rocco, A., Tononi, G., & Ghilardi, M. F. (2011).
Modulation of gamma and theta spectral amplitude and phase
synchronization is associated with the development of visuo-
motor learning. Zeitschrift für Neurowissenschaften, 31(41), 14810–14819.
https://doi.org/10.1523/ JNEUROSCI.1319-11.2011, PubMed:
21994398
Petersen, S. E., van Mier, H., Fiez, J. A., & Rachel, M. E. (1998).
The effects of practice on the functional anatomy of task perfor-
Mance. Verfahren der Nationalen Akademie der Wissenschaften, 95(3),
853–860. https://doi.org/10.1073/pnas.95.3.853, PubMed:
9448251
Pollok, B., Latz, D., Krause, V., Butz, M., & Schnitzler, A. (2014).
Changes of motor-cortical oscillations associated with motor
learning. Neurowissenschaften, 275, 47–53. https://doi.org/10.1016/j
.neuroscience.2014.06.008, PubMed: 24931763
Porter, M. A., Utopie, J.-P., & Mucha, P. J. (2009). Communities in
Netzwerke (SSRN Scholarly Paper ID 1357925). Social Science
Research Network. https://papers.ssrn.com/abstract=1357925
Reddy, P. G., Mattar, M. G., Murphy, A. C., Wymbs, N. F., Grafton,
S. T., Satterthwaite, T. D., & Bassett, D. S. (2018). Brain state flex-
ibility accompanies motor-skill acquisition. NeuroImage, 171,
135–147. https://doi.org/10.1016/j.neuroimage.2017.12.093,
PubMed: 29309897
Rilk, A. J., Soekadar, S. R., Sauseng, P., & Plewnia, C. (2011). Alpha
coherence predicts accuracy during a visuomotor tracking task.
Neuropsychologie, 49(13), 3704–3709. https://doi.org/10.1016/j
.neuropsychologia.2011.09.026, PubMed: 21964200
Rueda-Delgado, L. M., Heise, K. F., Daffertshofer, A., Mantini, D.,
& Swinnen, S. P. (2019). Age-related differences in neural spec-
tral power during motor learning. Neurobiologie des Alterns, 77,
44–57. https://doi.org/10.1016/j.neurobiolaging.2018.12.013,
PubMed: 30776651
Ruff, R. M., & Parker, S. B. (1993). Gender- and age-specific
changes in motor speed and eye-hand coordination in adults:
Normative values for the Finger Tapping and Grooved Pegboard
Tests. Perceptual and Motor Skills, 76(3 suppl), 1219–1230.
https://doi.org/10.2466/pms.1993.76.3c.1219, PubMed: 8337069
Ruggeri, P. (2022). Dynamic rewiring of electrophysiological brain
networks during learning [Data set]. Zenodo. https://doi.org/10
.5281/zenodo.6394946
Sakai, K., Hikosaka, O., Miyauchi, S., Sasaki, Y., Fujimaki, N., &
Pütz, B. (1999). Presupplementary motor area activation during
sequence learning reflects visuo-motor association. Zeitschrift für
Neurowissenschaften, 19(10), RC1. https://doi.org/10.1523/JNEUROSCI
.19-10-j0002.1999, PubMed: 10234047
Sami, S., Robertson, E. M., & Miall, R. C. (2014). The time course of
task-specific memory consolidation effects in resting state net-
funktioniert. Zeitschrift für Neurowissenschaften, 34(11), 3982–3992. https://doi
.org/10.1523/JNEUROSCI.4341-13.2014, PubMed: 24623776
Sampaio-Baptista, C., Filippini, N., Stagg, C. J., Near, J., Scholz, J.,
& Johansen-Berg, H. (2015). Changes in functional connectivity
and GABA levels with long-term motor learning. NeuroImage,
106, 15–20. https://doi.org/10.1016/j.neuroimage.2014.11.032,
PubMed: 25463472
Sampaio-Baptista, C., Scholz, J., Jenkinson, M., Thomas, A. G.,
Filippini, N., Smit, G., Douaud, G., & Johansen-Berg, H.
(2014). Gray matter volume is associated with rate of subsequent
skill learning after a long term training intervention. NeuroImage,
96, 158–166. https://doi.org/10.1016/j.neuroimage.2014.03
.056, PubMed: 24680712
Scheeringa, R., Bastiaansen, M. C. M., Petersson, K. M., Oostenveld,
R., Norris, D. G., & Hagoort, P. (2008). Frontal theta EEG activity
correlates negatively with the default mode network in resting
state. International Journal of Psychophysiology, 67(3),
242–251. https://doi.org/10.1016/j.ijpsycho.2007.05.017,
PubMed: 17707538
Schoffelen, J.-M., & Brutto, J. (2009). Source connectivity analysis
with MEG and EEG. Kartierung des menschlichen Gehirns, 30(6), 1857–1865.
https://doi.org/10.1002/hbm.20745, PubMed: 19235884
Schubert, C., Dabbagh, A., Classen, J., Krämer, U. M., & Tzvi, E.
(2021). Alpha oscillations modulate premotor-cerebellar connec-
tivity in motor learning: Insights from transcranial alternating cur-
rent stimulation. NeuroImage, 241, 118410. https://doi.org/10
.1016/j.neuroimage.2021.118410, PubMed: 34303797
Shamloo, F., & Helie, S. (2016). Changes in default mode network
as automaticity develops in a categorization task. Behavioural
Gehirnforschung, 313, 324–333. https://doi.org/10.1016/j.bbr
.2016.07.029, PubMed: 27457134
Smolen, P., Zhang, Y., & Byrne, J. H. (2016). The right time to learn:
Mechanisms and optimization of spaced learning. Natur
Reviews Neuroscience, 17(2), 77–88. https://doi.org/10.1038
/nrn.2015.18, PubMed: 26806627
Stam, C. J. (2014). Modern network science of neurological disor-
ders. Nature Reviews Neurowissenschaften, 15(10), 683–695. https://doi
.org/10.1038/nrn3801, PubMed: 25186238
Stratton, S. M., Liu, Y.-T., Hong, S. L., Mayer-Kress, G., & Newell,
K. M. (2007). Snoddy (1926) revisited: Time scales of motor
learning. Journal of Motor Behavior, 39(6), 503–515. https://doi
.org/10.3200/JMBR.39.6.503-516, PubMed: 18055356
Studer, B., Koeneke, S., Blum, J., & Jäncke, L. (2010). Die Auswirkungen von
practice distribution upon the regional oscillatory activity in
visuomotor learning. Behavioral and Brain Functions, 6(1), 8.
https://doi.org/10.1186/1744-9081-6-8, PubMed: 20205755
Sun, F. T., Müller, L. M., Rao, A. A., & D'Esposito, M. (2006). Func-
tional connectivity of cortical networks involved in bimanual
Netzwerkneurowissenschaften
602
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network dynamics underlying motor skill learning
motor sequence learning. Hirnrinde, 17(5), 1227–1234.
https://doi.org/10.1093/cercor/bhl033, PubMed: 16855008
Tabbal, J., Kabbara, A., Khalil, M., Benquet, P., & Hassan, M.
(2021). Dynamics of task-related electrophysiological networks:
A benchmarking study. NeuroImage, 231, 117829. https://doi.org
/10.1016/j.neuroimage.2021.117829, PubMed: 33549758
Tadel, F., Bock, E., Niso, G., Mosher, J. C., Cousineau, M., Pantazis,
D., Leahy, R. M., & Baillet, S. (2019). MEG/EEG group analysis
with Brainstorm. Frontiers in Neuroscience, 13, 76. https://doi.org
/10.3389/fnins.2019.00076, PubMed: 30804744
Taubert, M., Draganski, B., Anwander, A., Müller, K., Horstmann,
A., Villringer, A., & Ragert, P. (2010). Dynamic properties of
human brain structure: Learning-related changes in cortical areas
and associated fiber connections. Zeitschrift für Neurowissenschaften, 30(35),
11670–11677. https://doi.org/10.1523/ JNEUROSCI.2567-10
.2010, PubMed: 20810887
Taubert, M., Lohmann, G., Margulies, D. S., Villringer, A., & Ragert,
P. (2011). Long-term effects of motor training on resting-state
networks and underlying brain structure. NeuroImage, 57(4),
1492–1498. https://doi.org/10.1016/j.neuroimage.2011.05.078,
PubMed: 21672633
The Jamovi Project. (2021). Jamovi ( Version 1.0.7.0) [Computer
Software]. Retrieved from https://www.jamovi.org.
Thomas Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R.,
Lashkari, D., Hollinshead, M., Roffman, J. L., Smoller, J. W.,
Zöllei, L., Polimeni, J. R., Fischl, B., Liu, H., & Buckner, R. L.
(2011). The organization of the human cerebral cortex estimated
by intrinsic functional connectivity. Journal of Neurophysiology,
106(3), 1125–1165. https://doi.org/10.1152/jn.00338.2011,
PubMed: 21653723
Tung, K.-C., Uh, J., Mao, D., Xu, F., Xiao, G., & Lu, H. (2013). Ändern-
ations in resting functional connectivity due to recent motor task.
NeuroImage, 78, 316–324. https://doi.org/10.1016/j.neuroimage
.2013.04.006, PubMed: 23583747
Tzvi, E., Bauhaus, L. J., Kessler, T. U., Liebrand, M., Wöstmann, M.,
& Krämer, U. M. (2018). Alpha-gamma phase amplitude cou-
pling subserves information transfer during perceptual sequence
learning. Neurobiology of Learning and Memory, 149, 107–117.
https://doi.org/10.1016/j.nlm.2018.02.019, PubMed: 29474959
Tzvi, E., Verleger, R., Münte, T. F., & Krämer, U. M. (2016). Reduziert
alpha-gamma phase amplitude coupling over right parietal cor-
tex is associated with implicit visuomotor sequence learning.
NeuroImage, 141, 60–70. https://doi.org/10.1016/j.neuroimage
.2016.07.019, PubMed: 27403869
Vecchio, F., Miraglia, F., Quaranta, D., Lacidogna, G., Marra, C., &
Rossini, P. M. (2018). Learning processes and brain connectivity
in a cognitive-motor task in neurodegeneration: Evidence from
EEG network analysis. Zeitschrift für Alzheimer-Krankheit, 66(2),
471–481. https://doi.org/10.3233/ JAD-180342, PubMed:
30282357
Wu, J., Srinivasan, R., Kaur, A., & Cramer, S. C. (2014). Resting-state
cortical connectivity predicts motor skill acquisition. Neuro-
Image, 91, 84–90. https://doi.org/10.1016/j.neuroimage.2014
.01.026, PubMed: 24473097
Wymbs, N. F., & Grafton, S. T. (2015). The human motor system
supports sequence-specific representations over multiple training-
dependent timescales. Hirnrinde, 25(11), 4213–4225.
https://doi.org/10.1093/cercor/bhu144, PubMed: 24969473
Yang, Z., Telesford, Q. K., Franco, A. R., Lim, R., Gu, S., Xu, T., Ai,
L., Castellanos, F. X., Yan, C.-G., Colcombe, S., & Milham, M. P.
(2021). Measurement reliability for individual differences in
multilayer network dynamics: Cautions and considerations. Neu-
roImage, 225, 117489. https://doi.org/10.1016/j.neuroimage
.2020.117489, PubMed: 33130272
Zalesky, A., Fornito, A., Cocchi, L., Gollo, L. L., & Breakspear, M.
(2014). Time-resolved resting-state brain networks. Verfahren
der Nationalen Akademie der Wissenschaften, 111(28), 10341–10346.
https://doi.org/10.1073/pnas.1400181111, PubMed: 24982140
Zhuang, P., Toro, C., Grafman, J., Manganotti, P., Leocani, L., &
Hallett, M. (1997). Event-related desynchronization (ERD) In
the alpha frequency during development of implicit and explicit
learning. Electroencephalography and Clinical Neurophysiology,
102(4), 374–381. https://doi.org/10.1016/S0013-4694(96)96030-7,
PubMed: 9146500
Netzwerkneurowissenschaften
603
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
2
5
7
8
2
1
1
8
4
2
2
N
e
N
_
A
_
0
0
2
8
9
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3