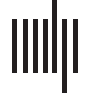FORSCHUNG
Sex differences in multilayer functional network
topology over the course of aging in 37543
UK Biobank participants
Mite Mijalkov1, Dániel Veréb1, Oveis Jamialahmadi2, Anna Canal-Garcia1,
Emiliano Gómez-Ruiz3, Didac Vidal-Piñeiro4, Stefano Romeo2,5,6,
Giovanni Volpe3, and Joana B. Pereira1,7
1Abteilung für Neurobiologie, Care Sciences and Society, Karolinska Institutet, Stockholm, Schweden
2Department of Molecular and Clinical Medicine, Goteborg University, Goteborg, Schweden
3Department of Physics, Goteborg University, Goteborg, Schweden
4Abteilung für Psychologie, University of Oslo, Oslo, Norwegen
5Cardiology Department, Sahlgrenska University Hospital, Gothenburg, Schweden
6Clinical Nutrition Unit, University Magna Graecia, Catanzaro, Italien
7Memory Research Unit, Department of Clinical Sciences Malmö, Lund University, Lund, Schweden
Schlüsselwörter: Altern, Sex differences, Multilayer networks, Funktionale Konnektivität, Anti-correlations
ABSTRAKT
Aging is a major risk factor for cardiovascular and neurodegenerative disorders, with considerable
societal and economic implications. Healthy aging is accompanied by changes in functional
connectivity between and within resting-state functional networks, which have been
associated with cognitive decline. Jedoch, there is no consensus on the impact of sex on
these age-related functional trajectories. Hier, we show that multilayer measures provide
crucial information on the interaction between sex and age on network topology, allowing for
better assessment of cognitive, strukturell, and cardiovascular risk factors that have been shown
to differ between men and women, as well as providing additional insights into the genetic
influences on changes in functional connectivity that occur during aging. In a large cross-
sectional sample of 37,543 individuals from the UK Biobank cohort, we demonstrate that such
multilayer measures that capture the relationship between positive and negative connections
are more sensitive to sex-related changes in the whole-brain connectivity patterns and their
topological architecture throughout aging, when compared to standard connectivity and
topological measures. Our findings indicate that multilayer measures contain previously
unknown information on the relationship between sex and age, which opens up new avenues
for research into functional brain connectivity in aging.
ZUSAMMENFASSUNG DES AUTORS
Alterations in the resting-state functional connectivity patterns are a hallmark of the aging
Verfahren. Jedoch, there is little agreement on how sex affects these age-related functional
trajectories. Hier, we propose novel multilayer measures that capture the relationship
between positive and negative connections. We show that they are more sensitive to sex-
related changes in the whole-brain connectivity patterns and their topology throughout aging,
when compared to standard connectivity and topological measures. Our results suggest that
multilayer measures provide information on the interaction between sex and age that has not
been previously explored, which leads to a more accurate evaluation of cognitive, strukturell,
Keine offenen Zugänge
Tagebuch
Zitat: Mijalkov, M., Veréb, D.,
Jamialahmadi, O., Canal-Garcia, A.,
Gómez-Ruiz, E., Vidal-Piñeiro, D.,
Romeo, S., Volpe, G., & Pereira, J. B.
(2023). Sex differences in multilayer
functional network topology over the
course of aging in 37543 UK Biobank
Teilnehmer. Netzwerkneurowissenschaften,
7(1), 351–376. https://doi.org/10.1162
/netn_a_00286
DOI:
https://doi.org/10.1162/netn_a_00286
zusätzliche Informationen:
https://doi.org/10.1162/netn_a_00286
Erhalten: 21 April 2022
Akzeptiert: 6 Oktober 2022
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Corresponding Authors:
Joana B. Pereira
joana.pereira@ki.se
Mite Mijalkov
mite.mijalkov@ki.se
Handling-Editor:
Alex Fornito
Urheberrechte ©: © 2022
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
and cardiovascular risk factors, and provides new insights into the genetic influences on
changes in functional connectivity that occur throughout aging.
EINFÜHRUNG
Although the human life span has increased from 50 Zu 80 years of age in the past two cen-
turies, this has not been matched by an improvement in health span (Crimmins, 2015). Tatsächlich,
age is one of the major risk factors for debilitating conditions such as cardiovascular and neu-
rodegenerative diseases, having a large societal and economic impact (Dhingra & Vasan,
2012; Hou et al., 2019). Jedoch, not all individuals age in the same way. Insbesondere,
sex seems to be responsible for a substantial interindividual variability during aging, mit
women displaying a higher probability of developing certain age-related disorders such as
Alzheimer’s disease (Mazure & Swendsen, 2016) and multiple sclerosis (Golden & Voskuhl,
2017), whereas men are more likely to develop Parkinson’s disease (K. M. Schmied & Dahodwala,
2014). These differences in vulnerability to distinct diseases suggest that men and women have
a distinct underlying brain network organization that might predispose them to develop spe-
cific pathological processes.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Negative connections:
Anticorrelations (negative
associations) between the nodes in
the whole-brain connectome.
Positive connections:
Correlations (positive associations)
between the nodes in the whole-
brain connectome.
The functional network organization of the brain can be assessed using the correlations of
spontaneous fluctuations in brain activity across brain regions by measuring the blood oxygen
level–dependent signals on resting-state functional magnetic resonance imaging (rs-fMRI)
(B. Biswal, Zerrin Yetkin, Haughton, & Hyde, 1995). Using this technique, several studies have
identified highly reproducible resting-state networks in the brain such as the sensorimotor,
dorsal attention, and default-mode networks, which play an important role in motor and cog-
nitive functions (van den Heuvel & Pol, 2010). The communication between these networks is
particularly important for brain function and has been shown to change during the course of
Altern (Betzel et al., 2014; Speisesaal, Bartrés Faz, & Junqué, 2015), with older individuals
showing a loss of anticorrelations (negative connections) (Keller et al., 2015; Spreng, Stevens,
Viviano, & Schacter, 2016) and increases in positive correlations (positive connections)
(Damoiseaux, 2017; Ferreira et al., 2016) between resting-state networks. These changes
reflect the tendency of older individuals to over-recruit functional networks, needing to acti-
vate more brain networks than younger individuals, thus decreasing functional specialization
and spending more neural resources (Goldstone et al., 2016).
Jedoch, the impact of sex on the communication between functional brain networks during
aging is still not well understood (Ritchie et al., 2018; Scheinost et al., 2015), in part due to the small
number of participants included in previous studies and their limited statistical power (Ritchie et al.,
2018; Ruigrok et al., 2014). Assessing sex differences in functional connectivity is important for
several reasons. Zum Beispiel, because functional networks are closely associated with cognitive
and sensorimotor functions (van den Heuvel & Pol, 2010), understanding how their communica-
tion deteriorates with aging might provide important clues on why men and women are vulner-
able to different diseases (Golden & Voskuhl, 2017; Mazure & Swendsen, 2016; K. M. Schmied &
Dahodwala, 2014) and why they show differences in other important health aspects such as brain
Struktur, cardiovascular risk factors, and cognitive function (McCarrey, Ein, Kitner-Triolo,
Ferrucci, & Resnick, 2016; Ramirez & Sullivan, 2018; Ritchie et al., 2018; Sachdev, Parslow,
Wen, Anstey, & Easteal, 2009; Weiss, Kemmler, Deisenhammer, Fleischhacker, & Delazer, 2003).
From a methodological point of view, studies analyzing functional connectivity have
mainly focused on positive connections (Chan, Park, Savalia, Petersen, & Wig, 2014; Tomasi
& Volkow, 2012A). While this approach is more straightforward to assess the organization or
Netzwerkneurowissenschaften
352
Multilayer functional networks identify sex differences in aging
topology of brain networks (Fornito, Zalesky, & Breakspear, 2013), negative connections are
commonly found between brain networks or areas and seem to play an important role in brain
communication (Fox et al., 2005; Hampson, Driesen, Roth, Gore, & Polizist, 2010) Und
Erkenntnis (Barber, Caffo, Pekar, & Mostofsky, 2013). Since the negative connections or anti-
correlations carry behaviorally relevant information (Barber et al., 2013), an integrative
approach that incorporates information from negative correlations as well as positive
correlations may reveal unique insights on sex differences throughout aging.
In this study, we developed this approach by combining the positive and negative func-
tional connections between networks as separate layers in a complex multilayer network.
We demonstrated that the multilayer measures provide novel insights about the impact of
age and sex on the different functional connectivity trajectories in men and women between
47 Und 79 years old in a large cross-sectional cohort of 37,543 individuals. Darüber hinaus, together
with other measures of network organization, they were associated with structural brain imag-
ing markers, cardiovascular risk factors, and cognitive functions, which typically differ
between men and women (McCarrey et al., 2016; Ramirez & Sullivan, 2018; Ritchie et al.,
2018; Sachdev et al., 2009; Weiss et al., 2003), as well as genes involved in physiological
processes associated with aging. These findings open new avenues for the study of functional
brain connectivity in aging by using multilayer network measures.
ERGEBNISSE
Sample
We included 19,975 women and 17,568 men with resting-state functional MRI from the UK
Brain Biobank cohort (Miller et al., 2016) (Methoden, section Participants and Supporting
Information Figure S1). A subset of these individuals had available structural brain imaging data
such as T1-weighted imaging and underwent a comprehensive assessment of cognitive func-
tions and cardiovascular risk factors (Methoden, sections Structural Imaging Preprocessing,
Cardiovascular Risk Factors, and Cognitive Tests). Using permutation testing to compare the
demographic characteristics between women and men, we found that men showed higher
scores than women in the executive cognitive domain during middle ages and in the visuospatial
cognitive domain across all ages (Supporting Information Figure S2C and D). In line with previ-
ous research showing that men are more susceptible to cardiovascular problems during middle
Erwachsensein (Anand et al., 2008; Gillis & Sullivan, 2016), we observed a significantly greater prev-
alence of high blood pressure, heart attack and white matter hyperintensities in men between the
ages of 51 Und 76 compared to women (Supporting Information Figure S2E, F, K). Endlich, men
had larger subcortical volumes across all ages (Supporting Information Figure S2I–K), konsistent
with previous studies (Ritchie et al., 2018; Wang, Xu, Luo, Hu, & Zuo, 2019). There were no
significant differences between sexes in professional qualifications and years of education.
Brain Connectivity Analysis
Functional brain connectivity was assessed for each participant using the negative and positive
correlations between 21 nodes that correspond to the resting-state functional MRI networks
shown in Figure 1A and Supporting Information Figure S3 (Methoden, section Functional Image
Vorverarbeitung). Erste, we computed classical single-layer connectivity measures, nämlich, Die
average connectivity for the whole correlation network, followed by the average negative con-
nectivity, the average positive connectivity, and the number of negative correlations (Methoden,
section Connectivity Measures). Dann, the negative and positive correlations of each functional
network were separated (Figure 1B.1 and 1B.2) and analyzed as two independent layers. To
Connectivity measures:
Measures summarizing the average
functional connectivity strength of
the whole-brain connectome.
Netzwerkneurowissenschaften
353
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
Figur 1. Analysis workflow. (A) Example of the 21 resting-state networks used as nodes and their positive (Rot) and negative connections
(Blau) for one of the subjects included in the analyses. Each network was identified based on previous descriptions (Miller et al., 2016). (B) Der
positive and negative connections were split into two networks: one negative (b.1) and the other one positive (b.2). The topology of these two
networks was evaluated using the clustering coefficient and global efficiency. (C and D) The positive and negative networks were then inte-
grated as two separate layers in a multiplex network (C) (where each node in one layer is connected to the same node in the other layer) und ein
multilayer network (D) (where each node in one layer is connected to all other nodes in the other layer). We evaluated the topology of the
multiplex network using the clustering and participation coefficients, whereas the topology of the multilayer network was assessed using the
novel multilayer global efficiency and multilayer clustering coefficient. In all graphs, thicker connections represent stronger positive or neg-
ative functional connections.
Multiplex network:
A two-layer multilayer network
where interlayer connections are
allowed only between corresponding
nodes in the positive and negative
layers.
Multilayer network:
A two-layer network where interlayer
connections are allowed between all
pairs of nodes in the two layers.
σ:
The fraction of the strongest
functional connection in the
corresponding multilayer network
that is used to calculate the weight of
the interlayer connections.
Multilayer global efficiency:
Measure that assesses the level of
functional integration in a multilayer
Netzwerk.
Netzwerkneurowissenschaften
evaluate the topological organization of each single layer, we used two measures: the clustering
coefficient and the global efficiency. The clustering coefficient is a measure of segregation that
increases with the number of local connections and represents the average clustered connec-
tivity around all nodes in the network. The global efficiency is a measure of integration that
increases as the paths connecting any two nodes in the network become shorter and estimates
the average capability with which different nodes communicate with each other (Mijalkov
et al., 2017) (Methoden, section Single-Layer Network Measures). To assess the relationship
between these two layers, we integrated them into a multiplex network (Abbildung 1C) and a mul-
tilayer network (Figure 1D). In the multiplex network approach (Abbildung 1C), each node in the
positive layer was connected with the same node in the negative layer. We computed two mul-
tiplex measures: the multiplex clustering coefficient, a measure that increases with the local
connections in neighboring nodes between the two layers, and the multiplex participation,
which is a measure of integration that assesses how evenly a node is connected in the two layers
(Battiston, Nicosia, & Latora, 2014) (Methoden, section Multiplex Network Measures). A disad-
vantage of the multiplex approach is that the relation between the two layers is local being only
allowed between the same nodes. To address this limitation, in the multilayer network
approach we connected each node in one layer to every node in the other layer (Figure 1D).
The strength of the relationship between the two layers can be changed by adjusting the
weight of the interlayer connections, σ. For each multilayer network, we define σ as a fraction
of the strongest functional connections in the corresponding network, and evaluate the mea-
sures’ ability to characterize sex differences across the wide range of σ values. We developed
two new measures to assess the integration and segregation properties of these multilayer net-
funktioniert. Speziell, we calculated the multilayer global efficiency, which compares the global
354
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
Multilayer clustering coefficient:
Measure that assesses the level of
functional segregation in a multilayer
Netzwerk.
efficiency differences due to the intra- and interlayer connections. Ähnlich, we also calculated
the multilayer clustering coefficient, which compares the clustering coefficients or triangles in
all nodes between the two layers (Methoden, section Multilayer Network Measures).
Women Have Less Negative Connections Than Men
In a first step, to identify which simple connectivity measures showed the greatest differences
between sexes over the course of aging, we compared men and women at all ages using a
permutation test and used separate linear models that included whole-brain average connec-
tivity, average negative connectivity, average positive connectivity, and the number of nega-
tive correlations as the outcome and age, Sex, age2, age × sex, and age2 × sex interactions as
predictors (the models were fit on the average data by sex and within each age; Methoden,
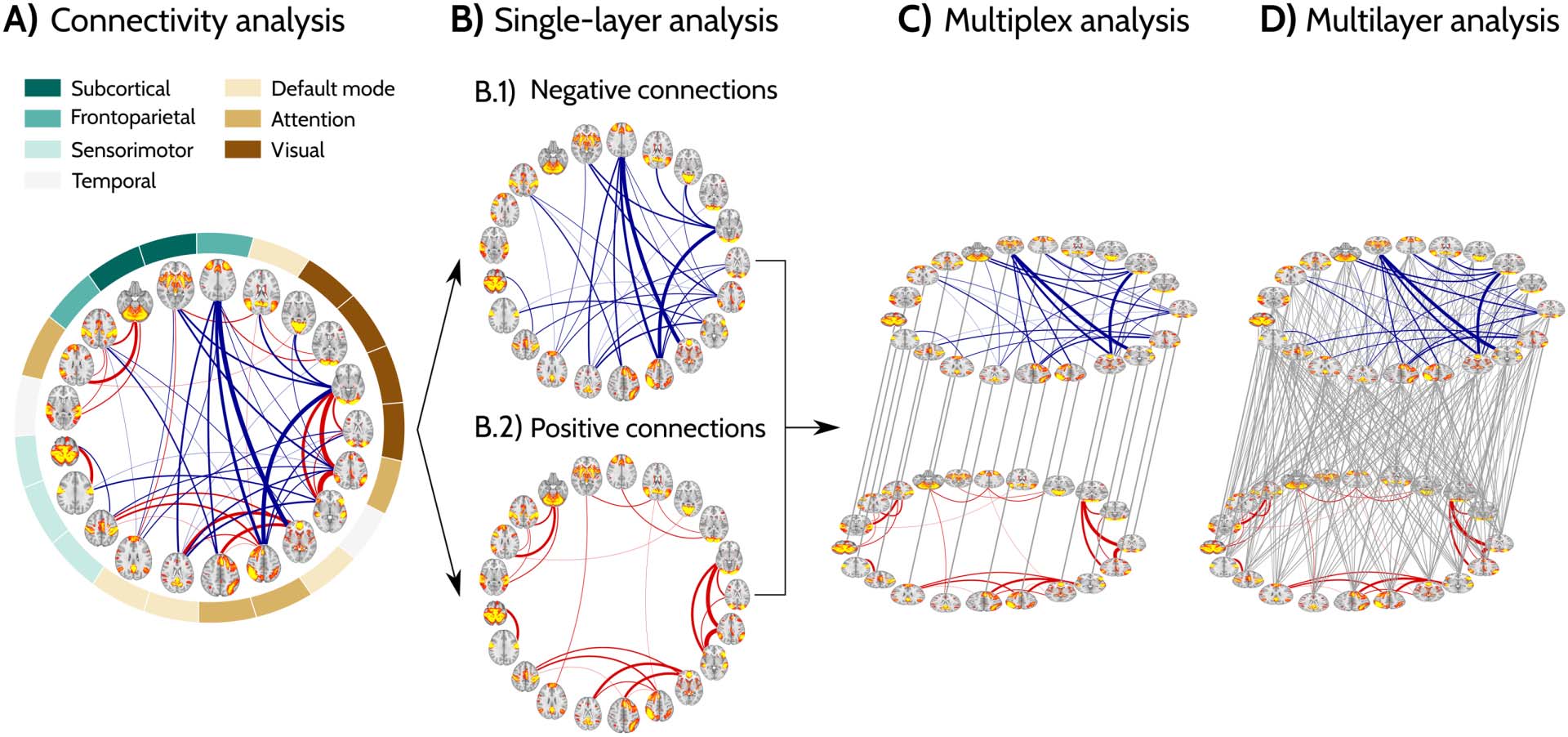
section Statistical Analysis). These models showed that women had significantly higher aver-
age functional connectivity than men (R2 = 0.769; AIC = 244.651; MSE = 2.184; Figure 2A and
Supporting Information Figure S4A), whereas men had a significantly higher number of neg-
ative connections compared to women (R2 = 0.778; AIC = 501.106; MSE = 108.420;
Figure 2D and Supporting Information Figure S4D) across a broad age range (50 Zu 71 Jahre).
These differences in connectivity between sexes diminished with increasing age, to the point
where there were almost no differences in mean connectivity strength or number of negative
connections between men and women after 75 Jahre (age × sex interaction: average connec-
tivity p = 0.01; number of negative connections p < 0.001). On the other hand, there were no
significant differences between sexes in the average positive and negative connectivity
strength (R2 = 0.403; AIC = 281.229; MSE = 3.947 and R2 = 0.408; AIC = 281.229; MSE =
2.013, respectively; Figure 2B and C and Supporting Information Figure S4B and C). These
results suggest that women have higher connectivity strengths than men due to a lower num-
ber of negative connections, but these differences dissipate with increasing age.
Men Have Shorter Network Paths Than Women
To identify which single-layer topological measures showed the greatest differences between
sexes over the course of aging, we repeated the above analysis by including them as depen-
dent variables in separate linear models with age, sex, age2, age × sex, and age2 × sex inter-
actions as predictors. These models showed that men had higher global efficiency than
women in the positive and negative layers (R2 = 0.924; AIC = −1.051; MSE = 0.055 and
R2 = 0.649; AIC = 88.603; MSE = 0.209, respectively; Figure 2G–H and Supporting Informa-
tion Figure S4G–H), indicating that their functional connectomes were characterized by
shorter paths in the networks with negative and positive connections. Interestingly, these
sex differences remained constant across different ages, suggesting that they were independent
of age (age × sex or age2 × sex interaction not significant in global efficiency measures; Sup-
porting Information: Linear Models). In contrast, no significant differences in the clustering
coefficients in the positive and negative networks were observed between women and men
(R2 = 0.459; AIC = 226.729; MSE = 1.728 and R2 = 0.533; AIC = −96.294; MSE = 0.013,
respectively; Figure 2E and F and Supporting Information Figure S4E and F).
Men Have a Greater Balance Between Positive and Negative Connections
Our linear regression models showed that the multiplex participation coefficient was signifi-
cantly higher in men than in women across all ages (R2 = 0.891; AIC = −172.848; MSE =
0.004; Figure 2J, Supporting Information Figure S4J and Supporting Information: Linear
Models), indicating that the men’s functional connectomes were characterized by nodes with
a similar number of connections in the negative and positive connectivity layers. However,
Network Neuroscience
355
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Multilayer functional networks identify sex differences in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
.
t
Figure 2. Functional connectivity dependence on age and sex. Results of the linear models with simple connectivity measures (A–D), single-
layer topological measures (E–H) and multi-layer topological measures (I–J, multiplex; K–L, multilayer) as the outcomes and age, sex, age2,
age × sex and age2 × sex interactions as predictors. The areas show the 95% confidence intervals (CI) for the predictions and the solid lines
show the best line fit. The stars indicate points that showed significant differences between men and women after correction for multiple
comparisons across the different age groups (FDR at q < 0.05). The dots show the average values for men and women at the corresponding
age. For each measure, only the best fitting model containing a subset of the predictors is shown. Details about the best fitting, as well as the
full model which includes all predictors, are shown in Supporting Information: Linear Models. Detailed report of statistics are shown in
Supporting Information: Statistical Comparisons and Supporting Information: Power Analysis, see also Methods, section Statistical Analysis
and Supporting Information Figure S4.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
these differences decreased with age, similarly to the average connectivity and number of neg-
ative connections (age × sex interaction: p = 0.036). The multiplex clustering coefficient did
not reveal any significant differences between men and women (R2 = 0.096; AIC = −106.989;
MSE = 0.282; Figure 2I and Supporting Information Figure S4I). We note that caution needs to
be applied when considering multiplex clustering coefficients as their interpretation can be
cumbersome for multiplex correlation networks (Masuda, Sakaki, Ezaki, & Watanabe, 2018;
Zalesky, Fornito, & Bullmore, 2012).
Network Neuroscience
356
Multilayer functional networks identify sex differences in aging
Multilayer Topological Measures Are Lower in Women Than in Men
The differences between men and women were robust for the complete range of interlayer
weights σ for the multilayer clustering and for smaller values of σ in the case of multilayer
global efficiency. However, the strongest differences were observed for the σ = 0.7 and σ =
0.2 in the case of multilayer clustering and multilayer global efficiency, respectively (Support-
ing Information Figures S5 and S6). Whereas the multilayer clustering coefficient differences
between men and women remained stable with aging (R2 = 0.799; AIC = −130.497; MSE =
0.008; age × sex interaction not significant; Figure 2K and Supporting Information: Linear
Models), the differences in multilayer global efficiency decreased with aging (R2 = 0.795;
AIC = −82.061; MSE = 0.016; age × sex interaction: p < 0.001; Figure 2L and Supporting
Information: Linear Models). Up to 80% of the variance in both multilayer measures was
associated with sex differences over time, outperforming all previous models for simple con-
nectivity, single-layer measures, and multiplex topological measures. For a more consistent
comparison of the different measures, we have repeated our analysis when the functional con-
nectivity metrics were fitted to a full model with age, sex, age2, age × sex and age2 × sex) as
predictors (Supporting Information: Linear Models). These results further confirm that multi-
layer measures and multiplex participation are the best performing measures and suggest that
complex network measures that account for the relationship between positive and negative
functional connections are more sensitive to sex differences across aging.
Finally, we have conducted several analyses to assess the reproducibility of our findings.
Specifically, we assessed the reproducibility of the multilayer measures as a function of sample
size (Marek et al., 2022) (Supporting Information: Figures, Section VII), whether they were repli-
cable at individual network densities (Supporting Information: Figures, Section VIII), as well as
their test-retest reliability using the intraclass correlation coefficient (ICC) (McGraw & Wong,
1996) (Supporting Information: Test-Retest). Our findings showed that multilayer measures can
uncover similar patterns of between-sex differences for different analysis parameters. Specifically,
at most ages, they can detect the differences observed in the original sample in subsamples with
sizes as low as 55%–60% of the original sample, as well as at individual network densities higher
than 15%. Finally, their test-retest reliability was greatest in younger individuals and higher than
that of test-retest reliability of single-layer and connection measures; their ICC values ranged from
0.4 to 0.65, suggesting a fair to good degree of clinical significance (Cicchetti & Sparrow, 1981).
Multilayer and Multiplex Measures Are Significant Explainers of Structural, Cognitive and
Cardiovascular Differences Between Men and Women
Next, we assessed whether the observed sex differences in functional connectivity throughout
aging were associated with the cognitive functions, structural brain measures and vascular risk
factors that differed between men and women in our cohort (executive functions, visuospatial
functions, blood pressure and heart attack prevalence, subcortical volumes, white matter
hyperintensities; see Results, section Sample, and Supporting Information Figure S2). Due to
the high collinearity between the functional connectivity measures (Supporting Information
Figure S7), we examined these associations using partial least squares (PLS) regressions. The
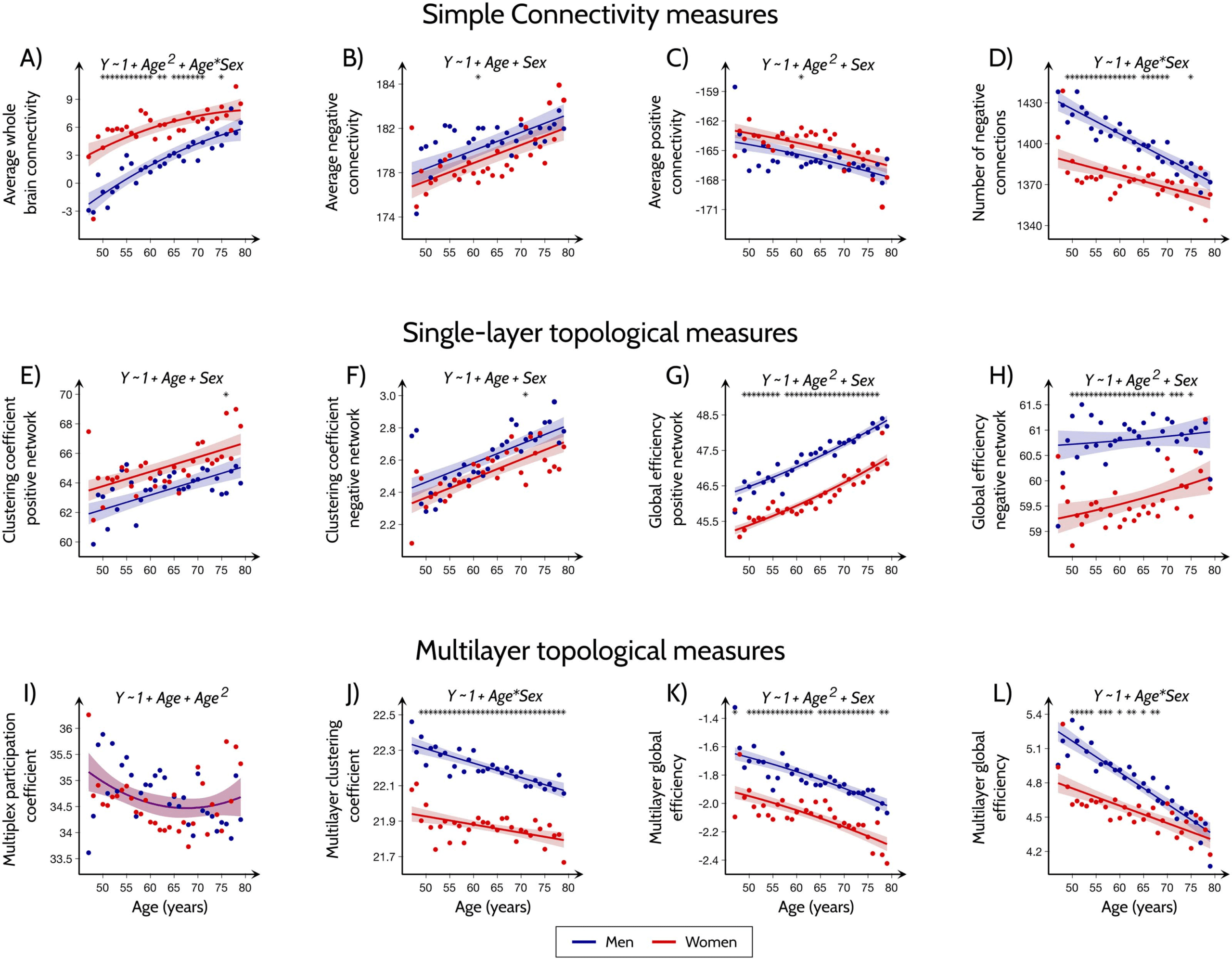
executive cognitive scores were best explained by the single-layer positive global efficiency,
followed by the single-layer negative clustering, average positive connectivity and negative
connectivity. The visuospatial cognitive scores were best explained by the multilayer func-
tional connectivity measures, followed by the average connectivity, number of negative con-
nections, and the positive single-layer global efficiency. Heart attack and high blood pressure
prevalence were best predicted by multiplex participation and single-layer global efficiency of
the positive connections, with blood pressure additionally being predicted by multilayer
Single-layer measures:
Classical network measures of
clustering coefficient and global
efficiency calculated for the networks
of positive and negative connections
separately.
Partial least squares (PLS) regression:
A multivariate regression analysis
that projects sets of dependent and
explanatory variables to a lower
dimensional subspace consisting of
latent variables that are maximally
correlated.
Network Neuroscience
357
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Multilayer functional networks identify sex differences in aging
clustering coefficient, number of negative connections, average connectivity, and single-layer
global efficiency of the positive network. Multilayer global efficiency was a significant predic-
tor of subcortical brain volumes and white matter hyperintensities, in addition to the average
connectivity, number of negative connections, multilayer clustering coefficient, multiplex par-
ticipation, and single-layer clustering coefficient in the case of subcortical volumes and single-
layer clustering coefficient in the case of white matter hyperintensities. The detailed VIP scores
for each predictor and the PLS model cumulative explained variance for the predicted vari-
ables are shown in Figure 3 and Supporting Information: Linear Models. Altogether, these
results indicate that, compared to the other measures, the multilayer and multiplex measures
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Relationship between functional connectivity measures and brain structure, cognitive measures and cardiovascular disease factors.
Plots showing the VIP scores for all functional connectivity measures and the amount of variance (for predicted and predictor variables)
explained by the corresponding latent variables. The PLS analysis was performed for measures showing significant differences between
men and women in (A) executive and (B) visuospatial domains; (C) prevalence of high blood pressure and (D) heart attack; (E) subcortical
volumes and (F) white matter hyperintensities. Abbreviations: LV: latent variables; Conn-Ave: Average connectivity; PosConn-Ave: Average
positive connectivity; NegConn-Ave: Average negative connectivity; NegConn-No: Number of negative connections; SLpos-CC and SLneg-CC:
Single-layer clustering coefficient for networks of positive and negative connections; SLpos-Ge and SLneg-Ge: Single-layer global efficiency for
networks of positive and negative connections; MP-CC: Multiplex clustering coefficient; MP-Pt: Multiplex participation coefficient; ML-CC and
ML-Ge: Multilayer clustering coefficient and global efficiency.
Network Neuroscience
358
Multilayer functional networks identify sex differences in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
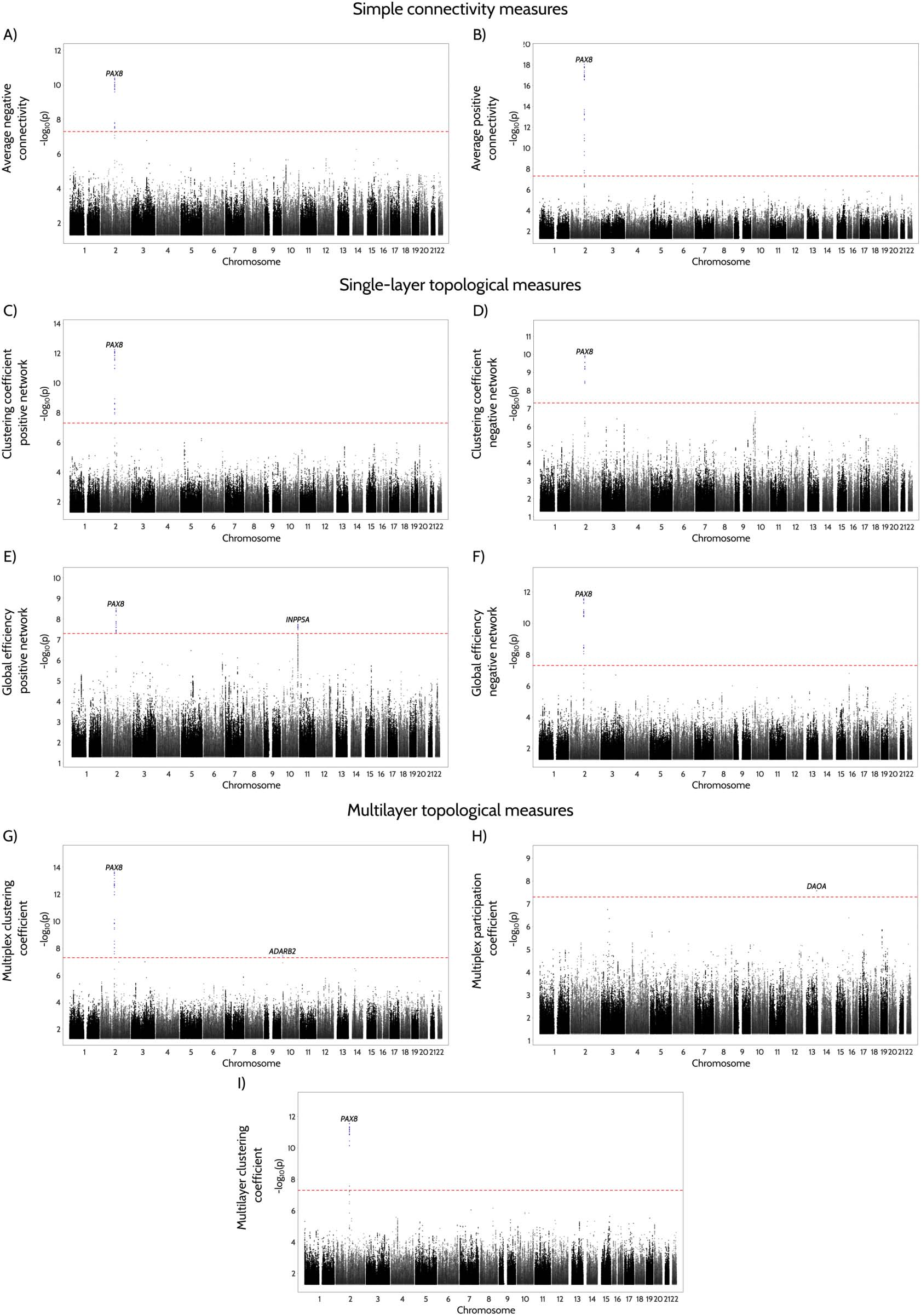
Figure 4. Manhattan plots for GWAS analysis of functional connectivity measures. The results from the GWAS analysis of simple connectivity
measures (A–B), single-layer topological measures (C–F) and multi-layer topological measures (G–H, multiplex; I, multilayer) in a total sample
of 33,773 individuals. The y-axis shows the p values for the association tests on a −log10 scale, while the different chromosomes (differentiated
by the black and gray colors) are ordered on the x-axis. The lead variants that surpass the genome-wide significance threshold (indicated by the
red line) are highlighted as blue circles.
Network Neuroscience
359
Multilayer functional networks identify sex differences in aging
were the best predictors of cognitive functions, structural brain measures, and vascular risk
factors that differed between men and women during aging.
Multilayer and Multiplex Measures Are Associated With Genes Involved in Aging-Related
Physiological Processes
To identify genetic variation associated with the 12 multilayer, single-layer and connectivity
functional measures we examined 9,356,431 genetic variants with minor allele frequency >
1% in a total of 33,773 European participants (Methoden, section Genetic Association Analyses).
A total of 4 loci exceeded a genome-wide significance level among one or more of these traits
(Figur 4 and Supporting Information Figure S8). A locus near Paired Box 8 (PAX8), a gene
involved in sleep efficiency, diastolic blood pressure, Entwicklung, and vulnerability to neu-
rodegenerative diseases (Elliott et al., 2018; Foo et al., 2021B; Jones et al., 2019), showed an
association with the majority of functional connectivity measures, including multilayer and
multiplex clustering, as well as all single-layer measures. Colocalization analysis with eQTL
summary statistics of 49 tissues in GTEx project and brain tissues in BrainSeq, ROSMAP,
Braineac2, and CommonMind datasets (Methoden, section Colocalization) (GTEx Consortium,
2020; Guelfi et al., 2020; Jaffe et al., 2018; Kerimov et al., 2021; Ng et al., 2017; Sieberts et al.,
2020), suggests a consistent strong colocalization between this locus and gene expression
patterns of Immunoglobulin Kappa Variable 1/OR2-108 (IGKV1OR2-108), COBW domain-
containing protein 2 (CBWD2), and Forkhead Box D4 Like 1 (FOXD4L1) over the multilayer
and multiplex clustering coefficients, average positive and negative connectivity, and all single-
layer functional measures (zusätzliche Informationen: Genetic Results). Außerdem, a locus near
Inositol Polyphosphate-5-Phosphatase A (INPP5A) gene showed significant associations with
the single-layer positive global efficiency. Interessant, the genome-wide association study
(GWAS) lead variant at this locus colocalized with eQTL for INPP5A in brain dorsolateral pre-
frontal cortex (DLPFC) from CommonMind dataset. Endlich, loci near Adenosine Deaminase
RNA specific B2 (ADARB2) and D-Amino Acid Oxidase Activator (DAOA) genes, welche haben
previously been linked with processes involved in normal memory functioning and ATP metab-
olism (Prata et al., 2012), were associated with multiplex clustering and multiplex participation
coefficients respectively.
DISKUSSION
Complex network measures are becoming increasingly popular in the field of network science
(van den Heuvel & Pol, 2010). In combination with large samples, these measures can
improve our understanding of brain connectivity (Fornito et al., 2013), Und, insbesondere,
our ability to predict sex differences over the course of aging (Stumme, Jockwitz, Hoffstaedter,
Amunts, & Caspers, 2020). Previous studies of brain connectivity and network topology have
mainly focused on measures that exclude the negative connections or average the effects of
positive and negative connections (Chan et al., 2014; Ferreira et al., 2016; Foo et al., 2021A;
Tomasi & Volkow, 2012B; Zhang et al., 2016). Jedoch, changes in the balance between pos-
itive and negative connectivity could be a more sensitive marker of abnormalities that occur in
men and women in middle and late adulthood. Here we show that measures that assess this
balance are indeed better predictors of sex differences during aging. Außerdem, we also
show that these measures are associated with genes implicated in aging-related physiological
processes as well as cognition, brain structure, and vascular disease, which have been previ-
ously shown to differ between men and women (McCarrey et al., 2016; Ramirez & Sullivan,
2018; Ritchie et al., 2018; Sachdev et al., 2009; Weiss et al., 2003). Altogether these findings
highlight the importance of integrating the information from positive and negative connections
Netzwerkneurowissenschaften
360
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
in a multiplex or multilayer network approach to provide a more holistic view of functional
brain connectivity changes.
Increasing age is associated with lower physical fitness and worse cognitive abilities, Sein
one of the greatest risk factors for the development of neurodegenerative diseases (Deary et al.,
2009; Wyss-Coray, 2016). Jedoch, there are several factors that determine how an individual
ages and his/her predisposition to develop certain diseases. Sex is one of these factors and it
has recently received a lot of attention due to increasing recognition that precision medicine
approaches should take into account biological sex to treat neurodegenerative diseases
(Ferretti et al., 2018). Sex was found to affect the functional connectivity (Allen et al., 2011;
B. B. Biswal et al., 2010; Ritchie et al., 2018); Jedoch, the location and nature of the functional
connectivity differences between men and women vary across studies (Scheinost et al., 2015).
Between-sex differences were most consistently observed in the default mode network (Allen
et al., 2011), but also reported in other resting-state networks (Allen et al., 2011; B. B. Biswal
et al., 2010; Filippi et al., 2013; Scheinost et al., 2015), as well as in the functional connectivity
patterns between them (Goldstone et al., 2016; Satterthwaite et al., 2015; Stumme et al., 2020).
These differences between men and women change with aging (Cosgrove, Mazure, & Staley,
2007), with studies showing different age-related trajectories in functional connectivity that
can be either diverging or converging in different resting-state networks (Goldstone et al.,
2016; Scheinost et al., 2015). Jedoch, other studies have reported that between-sex differences
persist also in older individuals (Stumme et al., 2020). Such differing results suggest that the
understanding of sex differences in the adult brain is still in its infancy and utilizing larger sam-
ples or more sensitive methods are needed to address these discrepancies.
Functional brain connectivity plays a crucial role in how brain networks communicate
with each other, being closely associated with behavior and cognition (van den Heuvel &
Pol, 2010). Studies have shown that aging is accompanied with an increase in the connec-
tions between these networks (Betzel et al., 2014; Chan et al., 2014; Damoiseaux, 2017;
Ferreira et al., 2016). These increases in connectivity are thought to be due to a less efficient
use of neural resources in older individuals, who tend to over-recruit brain networks to com-
pensate for the detrimental effects of aging (Geerligs, Renken, Saliasi, Maurits, & Lorist, 2015;
Goldstone et al., 2016; Park & Reuter-Lorenz, 2009). Hier, we confirm these findings in a
larger sample of middle-age and old adults by showing that aging is associated with increases
in the average whole-brain connectivity and decreases in the number of negative connec-
tionen. Darüber hinaus, our findings revealed that men displayed lower average connectivity and
a greater number of negative connections than women at young ages, but these differences
dissipated with increasing age, to the point that no differences between sexes were observed
any longer at old ages. These findings agree with results from previous studies showing
different functional trajectories between men and women (Satterthwaite et al., 2015;
Scheinost et al., 2015). Jedoch, in contrast to other studies (Stumme et al., 2020), Hier
we demonstrate that, although the functional connectomes of younger men and women
are different, they become increasingly similar with older age, possibly due to a faster rate
of functional changes observed in the brains of men (Foo et al., 2021A), which is in line with
studies showing that men have lower resilience to age-related cognitive decline compared to
Frauen (McCarrey et al., 2016).
Regarding measures of network organization, we found that the positive global efficiency
was higher in men compared to women, but these differences remained stable across different
Alter. Similar differences were found in the negative global efficiency; Jedoch, in diesem Fall,
the differences were not stable and seemed to decrease with aging. The global efficiency is
used to assess integration or the ability for an efficient processing to occur between distant
Netzwerkneurowissenschaften
361
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
brain networks (Fornito et al., 2013); an excessive integration is thought to impede the ability
of the brain to process information in a meaningful way (Lord, Stevner, Deco, & Kringelbach,
2017). Außerdem, global efficiency is associated with the presence of long range connec-
tions between networks. The healthy brain connectome is characterized by a low number of
such connections because long range connections are associated with higher metabolic costs
and minimizing these costs is essential for evolution (Bullmore & Spurns, 2012). daher,
these differences between sexes might provide clues on why men are more prone to develop
specific diseases with age, Zum Beispiel, Parkinson’s disease and epileptic seizures, welche sind
associated with abnormal organization in the functional connectivity networks as a result of
increased integration (Lord et al., 2017; Mijalkov, Volpe, & Pereira, 2022).
To evaluate whether the relationship between the positive and negative connections can
reveal additional insights into sex differences over the course of aging, we integrated these
connections as two separate layers in a multiplex and a multilayer networks. This relationship
estimates the brain’s ability to inhibit certain functional connections in order to modulate the
coactivation between different resting-state networks. Because the positive and negative con-
nections can arise from different neurovascular mechanisms (Goelman, Gordon, & Bonne,
2014), the presence of both connection types is necessary to reach a more balanced commu-
nication between resting-state networks and consequently a better cognitive performance
(Saberi, Khosrowabadi, Khatibi, Misic, & Jafari, 2021); in fact, the lack of anticorrelation or
negative connections has been associated with lower cognitive control and working memory
performance during neurodevelopment (Chai, Ofen, Gabriela, & Whitfield-Gabrieli, 2014).
This balance is disrupted in older adults, which over-recruit the functional networks as a
compensatory mechanism to maintain or improve function (Goldstone et al., 2016; Park &
Reuter-Lorenz, 2009), Zum Beispiel, by engaging both the left and right hemispheres (HAROLD
Modell) (Cabeza, Anderson, Locantore, & McIntosh, 2002) or showing more distributed
activation when compared to younger adults in performing verbal working and long-term
memory as well as category-learning tasks (Park & Reuter-Lorenz, 2009).
In the multiplex network, where only connections between the same nodes in the two
layers are allowed, we measured the balance between the positive and negative connections
by the multiplex participation coefficient, which quantifies the heterogeneity of the connec-
tivity patterns of a given node in both layers. We found that men had higher multiplex par-
ticipation coefficients compared to women, indicating that they had a higher balance
between positive and negative connections in the two layers that decreased with aging at a
faster rate compared to women. Consistent with earlier findings, our results suggest that the
less efficient connectomes reported in older persons may be related to an increase in the num-
ber of positive connections at the expense of negative connections (Ferreira et al., 2016),
thereby disrupting the balance between coactivation and inhibition connectivity between
functional networks.
The multilayer network approach, where connections are allowed between all nodes in the
two layers, extends the multiplex approach by providing an estimation of the degree to which the
between-layer balance affects the topological organization of the network. We found that men
had higher multilayer clustering and multilayer global efficiency compared to women, welche
both decreased with aging. These findings indicate that the relation between positive and neg-
ative layers is stronger in men than in women. This might come at the expense of greater neural
resources and metabolic costs, which could predispose the male connectome to the effects of
increased oxidative stress and poor antioxidant defense mechanisms, which have been sug-
gested to accompany higher brain connectivity (Griffa, Baumann, Thiran, & Hagmann, 2013)
and could potentially lead to steeper rates of cognitive decline (McCarrey et al., 2016).
Netzwerkneurowissenschaften
362
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
When the different connectivity and topological network measures were compared to each
andere, we found that the multiplex and multilayer network measures were the variables that
were best predicted by sex differences over aging. Insbesondere, age and sex were able to pre-
dict multiplex participation coefficient, multilayer clustering and multilayer global efficiency
by explaining up to 89.1%, 79.9%, Und 79.5% of the variance. These findings indicate that the
integration of positive and negative connections as separate layers in a complex network
approach is sensitive to important age- and sex-related variability not captured by conven-
tional measures. This approach could thus be used to understand why men and women age
differently. Zum Beispiel, we found that cardiovascular risk factors such as hypertension and
heart attack prevalence, which were higher in men between 51 Und 79 Alter, were best pre-
dicted by these measures. Zusätzlich, differences between sexes that remained stable over
aging such as lower visuospatial cognitive scores and lower subcortical volumes in women
compared to men were also best explained by multilayer measures.
Regarding the results of the GWAS analysis, a locus near the PAX8 gene showed an association
with the majority of functional connectivity measures. Interessant, this locus has been associated
with sleep efficiency, diastolic blood pressure, insomnia, and sleep length (Jones et al., 2019).
These data are consistent with the aging process resulting in changes in sleep habits and high
blood pressure (Ancoli-Israel, 2009; Ramirez & Sullivan, 2018), both of which have been linked
to functional connectivity changes (Foo et al., 2021B; Neitzel et al., 2021; Tagliazucchi et al.,
2012). The PAX8 protein has also been associated to the regulation of multiple genes involved
in thyroid hormone synthesis, which is necessary for brain development and function, for exam-
Bitte, through processes such as neuronal differentiation, synaptogenesis, and dendritic prolifer-
ation (Foo et al., 2021B). All functional measures associated with the PAX8 gene demonstrated
substantial colocalization with the FoxD4L1 gene, which is similarly implicated in processes
that promote the onset of neural differentiation (Klein et al., 2013). They also demonstrated colo-
calization with CBWD2, which has been linked to sleep duration (Foo et al., 2021B), Und
IGKV1OR2-108, which has been found to be elevated in the livers of type 2 diabetes patients
(Li, Pan, & Yang, 2019) and can lead to abnormal functional connectivity (Chen et al., 2014).
The rs4309079 locus, associated with single-layer positive global efficiency, has been
linked to functional connectivity measures in previous research (Elliott et al., 2018). Es ist
located adjacent to the INPP5A gene, which is involved in calcium signaling. As a fundamen-
tal cellular mechanism, inositol calcium signaling is expected to play a role in a range of neu-
rological pathways that underlie functional connectivity (Neitzel et al., 2021). The INPP5A
gene has been further associated with brain age explained by changes in functional connec-
tivity and decreased system segregation (Neitzel et al., 2021; S. M. Smith et al., 2020), welches ist
consistent with our current findings.
Multiplex participation showed a strong association in a locus near the DAOA gene, welches ist
known to play an important role in the control of glutamatergic transmission (Prata et al., 2012).
Glutamate is the most common excitatory neurotransmitter in mammals, and glutamate’s acti-
vation of NMDA receptors is critical for normal memory function. Glutamatergic antagonists
(z.B., ketamine) have been demonstrated to lower performance on tests of declarative memory,
verbal fluency, and problem solving, all of which have been linked to aging (Prata et al., 2012).
We nevertheless could not find any striking colocalization evidence in the examined gene
expression QTL datasets. Endlich, for multiplex clustering, we found a locus near the ADARB2
(index variant: rs2152237) gene, which is involved in the ATP/ITP metabolic pathway. As a
Ergebnis, our findings add to our understanding of the genetic influences on functional connectivity
and provide a link between the functional and genetic architecture of the brain, which might be
relevant in explaining the changes in a variety of biological processes throughout healthy aging.
Netzwerkneurowissenschaften
363
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
This study has some limitations. Erste, a longitudinal study design would have been more
appropriate to assess age-related differences between sexes in functional brain connectivity. In
besondere, recent studies have shown that results regarding aging-related changes obtained
from cross-sectional and longitudinal designs can be different from each other (Vidal-Piñeiro
et al., 2021; Xing, 2021); daher, our findings should be interpreted with this limitation in
Geist. Jedoch, multiple efforts to collect large longitudinal samples are currently underway
(Liu et al., 2021; Miller et al., 2016), which opens the opportunity for future studies to over-
come this limitation. Zweite, there was a considerable overlap between the values for indi-
vidual men and women in functional connectivity measures. This overlap has also been
observed in previous studies (Joel & Fausto-Sterling, 2016; Ritchie et al., 2018; Stumme
et al., 2020; Zhang et al., 2016; Zhang, Dougherty, Baum, White, & Michael, 2018), indicat-
ing that the prediction of an individual’s sex from a small number of functional connectivity
measures is challenging (Zhang et al., 2018). dennoch, our findings revealed consistent
group differences between men and women across a wide age range (47–79 ages), vorschlagen
Das, although there is considerable variability, some changes seem to be quite robust
(Chekroud, Station, Rosenberg, & Holmes, 2016). Endlich, since in this study the nodes of the
networks corresponded to 21 resting-state networks derived from independent component
Analysen (Miller et al., 2016), the negative connections are related to anticorrelations between
resting-state networks, which have been consistently observed and demonstrated to have a
neurophysiological basis (Chai, Castañón, Öngür, & Whitfield-Gabrieli, 2012; Fox et al.,
2005; Fuchs, Zhang, Snyder, & Rachel, 2009). Daher, these connections should be interpreted
differently than the negative correlations between brain regions analyzed in studies using
brain areas as nodes, which are still not clearly interpretable (Chai et al., 2012; Fornito
et al., 2013).
To summarize, in this study we developed novel multilayer connectivity measures in order
to assess the connectivity patterns and topological architecture between resting-state networks
in a large cohort of middle age and old adults. We showed that these multilayer measures are
superior at capturing sex-related effects during aging when compared to simpler connectivity
measures that do not account for the relationship between positive and negative connections.
The multilayer measures were also significant predictors of sex differences in cognitive, struc-
tural, and cardiovascular measures, and they were associated with genes that have previously
been implicated in aging-related processes. Daher, our findings highlight the importance of
studying the balance between positive and negative functional connections to understand
the effects of sex over aging, which should be included in future studies.
METHODEN
Teilnehmer
The UK Biobank cohort is a large population-based study with more than 500,000 Teilnehmer
from the United Kingdom (https://www.ukbiobank.ac.uk/). Following an initial visit for collec-
tion of medical and other clinical information, 37,704 individuals underwent MRI. To achieve
robust group comparisons, we limited our analyses to age groups with at least 50 Teilnehmer,
resulting in a sample size of 37,543 people in the age range 47–79 years (17,568 men: mean
age = 64.75; SD = 7.58 Und 19,975 Frauen: Durchschnittsalter = 63.49; SD = 7.33).
Image Acquisition
The functional MRI scans were performed on a standard Siemens Skyra 3T scanner using an
echo-planar imaging (EPI) sequence with the following parameters: duration ∼ 6 min; 490
Netzwerkneurowissenschaften
364
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
time points; repetition time = 735 MS; Echozeit = 39 MS; Sichtfeld = 88 × 88 × 64; voxel
size = 2.4 mm3; flip angle = 52°. Participants were instructed to relax and think of nothing in
particular while focusing their eyes on a crosshair during the scan. Regarding structural MRI,
the T1-weighted images were obtained using a 3D magnetization-prepared rapid gradient-
echo imaging sequence with the following parameters: 208 Scheiben, Echozeit = 880 MS; rep-
etition time = 2,000 MS; Sichtfeld = 208 × 256 × 256; Voxelgröße = 1 mm3 (Alfaro-Almagro
et al., 2018).
Image Preprocessing
All images obtained by the different imaging modalities were analyzed using an image-
preprocessing pipeline run by the UK Biobank imaging core, who also performed quality
assessment of the images (Alfaro-Almagro et al., 2018). Im Folgenden, we summarize the
procedure for the different modalities; full details of the preprocessing and quality control
are available elsewhere (Alfaro-Almagro et al., 2018; Miller et al., 2016).
Functional Image Preprocessing
After data acquisition, a number of preprocessing steps were carried out using FSL, einschließlich
motion correction using MCFLIRT, grand-mean intensity normalization by a single multiplica-
tive factor, high-pass temporal filtering with a Gaussian-weighted least-squares straight line
fitting (σ = 50.0 S), EPI unwarping by using a field map obtained before data collection, gra-
dient distortion correction (GDC) unwarping, and removal of all artifacts by an ICA-based X-
noiseifier. Endlich, all datasets underwent temporal demeaning and variance normalization.
The preprocessed data of 4,100 participants was used for a Group-ICA analysis. Using FSL’s
MELODIC tool and FSLNets toolbox (https://fsl.fmrib.ox.ac.uk/fsl/fslwiki), a spatial-ICA with a
dimensionality of 25 components was applied and the resulting ICA maps were mapped onto
each subject’s resting-state fMRI time series data to generate one representative timeseries per
ICA component. During this procedure, four networks were identified as artifacts and were
discarded from further analysis. This resulted in a 21 × 21 connectivity matrix for each partic-
ipant, where the functional connectivity between each pair of ICA spatial maps is character-
ized by full normalized temporal correlation. Analyzed networks included the default mode,
fronto-parietal, sensorimotor, visuell, attention, subcortical, and temporal networks (Figure 1A);
the group-ICA spatial maps can be found at https://biobank.ctsu.ox.ac.uk/crystal/refer.cgi?id
=9028, and the average functional connectomes for several representative age groups are
shown in Supporting Information Figure S2. The complete description of the preprocessing
procedures can be found elsewhere (Alfaro-Almagro et al., 2018; Miller et al., 2016) (online
Dokumentation: https://biobank.ctsu.ox.ac.uk/crystal/docs/brain_mri.pdf).
Structural Imaging Preprocessing
T1-weighted scans were preprocessed using the standard procedures of the FreeSurfer pipe-
Linie (Ausführung 6.0; https://surfer.nmr.mgh.harvard.edu/). We calculated the cortical thickness
for each individual by averaging the regional cortical thicknesses from 68 regions from
the Desikan–Killiany atlas (Desikan et al., 2006). Zusätzlich, we calculated the subcortical
volumes for each individual by averaging the volumes of all subcortical gray matter struc-
tures (cerebellar cortex, thalamus, Schwanz, putamen, pallidum, hippocampus, amygdala,
accumbens), which were corrected for total intracranial volume using a regression approach
(O’Brien et al., 2011). This data was available for a subsample of 17,317 men and 19,842
Frauen.
Netzwerkneurowissenschaften
365
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Multilayer functional networks identify sex differences in aging
White Matter Hyperintensities
The white matter hyperintensities were segmented from a combined T1 and T2-FLAIR images,
using a fully automated supervised method based on the k-nearest neighbors algorithm
(Griffanti et al., 2016).
Cardiovascular Risk Factors
At their initial visit, individuals were questioned about their medical history, including whether
or not they had a high blood pressure (subsample of 17,517 men and 19,911 Frauen), a heart
Attacke (subsample of 15,204 men and 16,918 Frauen), a heart attack–related angina episode
(subsample of 14,224 men and 15,520 Frauen), or a stroke (subsample of 14,224 men and
15,520 Frauen). We calculated the percentage of men and women diagnosed with the
previous conditions at all age groups, and used these percentages as dependent variables in
further analyses. Since the diagnosis of cardiovascular diseases was performed at a different
time point than the brain scanning, we included the time difference between diagnosis and
scanning as a covariate in the analyses with cardiovascular variables. Außerdem, as there
were several age groups that did not have any individuals with heart attack and hypertension,
the models for these variables were corrected for age prior to the PLS regression.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive Tests
The cognitive assessments were administered on a touch screen and took place at the same
visit as the brain scans. We averaged the standardized z-scores of 10 cognitive tests (Fawns-
Ritchie & Deary, 2020) into four different cognitive domains: attention/psychomotor speed
(reaction time, trail making – numeric path, symbol digit substitution tests; subsample of
11,531 men and 13,125 Frauen), Erinnerung (numeric memory, paired associative learning, Profi-
spective memory, pairs matching tests; subsample of 11,657 men and 13,284 Frauen), exec-
utiv (fluid intelligence/reasoning, trail making – alphanumeric path tests; subsample of 11,535
men and 13,159 Frauen), and visuospatial/visuoconstructional (matrix pattern completion,
tower rearranging tests; subsample of 11,452 men and 13,013 Frauen). More details about
the cognitive tests are available at https:// biobank.ndph.ox.ac.uk/showcase/ label.cgi?id
=100026.
Connectivity Measures
We assessed the functional connectomes using four different connectivity measures. A net-
work’s average connectivity (Conn-Ave) is defined as the average functional strength of all
its connections. Ähnlich, we estimated the average positive connectivity (PosConn-Ave)
and average negative connectivity (NegConn-Ave) as the mean strength of the network’s pos-
itive and negative connections, jeweils. Endlich, we calculated the number of negative
connections NegConn-No in the network. These measures can be evaluated as:
Conn-Ave ¼ 1
N
X
wi;
i2A
PosConn-Ave ¼ 1
Npos
NegConn-Ave ¼ 1
Nneg
X
i2Apos
X
wi;
wi;
i2Aneg
Netzwerkneurowissenschaften
366
Multilayer functional networks identify sex differences in aging
where N is the total number of connections in the network A. The network A can be expressed
as a sum of Apos and Aneg, which denote the networks consisting of only positive and negative
connections. The total number of connections in Apos and Aneg are denoted by Npos and Nneg,
jeweils.
The connectivity measures were calculated on the weighted connectivity networks consist-
ing of positive and negative connections (shown in Figure 1A). For each weighted connectivity
Netzwerk, we calculated a corresponding binary network in which the individual connections
retained their weight if they exceeded a certain threshold and were set to zero if they did not.
In this process, the absolute value of each connection was compared to the threshold;
Jedoch, their sign was preserved in the resulting binary matrix (d.h., negative connections
in the weighted network remained negative in the binary network). As there are multiple
thresholding approaches and there is currently no consensus as to which network density
should be used (Fornito et al., 2013), we performed the thresholding at a density range of
6% Zu 33%, in steps of 1%. For densities below 6%, the networks became largely discon-
nected with fewer edges than nodes, wohingegen 33% was the maximum density that could be
reached by all men’s and women’s networks of positive and negative connections (Supporting
Information Figure S12).
After calculating all measures at each density within the complete density range, we eval-
uated the corresponding area under the curve (AUC) value, which was used to assess the
between-sex differences (Supporting Information Figure S13). The AUC value was obtained
by numerically integrating the measure values over the density range; this procedure resulted
in a single numerical value for each network measure across the range of densities. Als dies
analysis takes into account the entire density range, it is considered to be less susceptible to
the thresholding process (Fornito et al., 2013).
Topological Network Analysis
Single-layer network measures. We split the weighted connectivity networks into networks con-
sisting of positive connections and negative connections (Figure 1B). As network measures are
not defined for negative weights, the weights in the network of negative connections were
substituted by their absolute values. daher, the topological analysis was equivalent for both
positive and negative connections networks; in the following we omit the subscript (pos/neg)
for clarity. Both networks were independently binarized at the density range 6%–33%, und das
AUC value for all measures was obtained across this range, as outlined above. The topology of
these networks was evaluated by calculating the global efficiency (SL-Ge) and clustering coef-
ficient (SL-CC).
In a weighted network, the distance dij between nodes i and i is
Single-layer global efficiency.
the total sum of individual connection lengths along the shortest path that connects the two
Knoten. Because large weights often imply strong relationships and close proximity, connection
lengths are inversely proportional to connection weights. While the shortest paths have the
smallest weighted distance, this does not necessarily equate to having the fewest number of
edges (Mijalkov et al., 2017). The regional global efficiency of a node i, denoted by SL-Gei, Ist
defined as the average inverse distance from to the other nodes in the network. The global
efficiency of a network, SL-Ge, is determined as the average of the global efficiency of all
Knoten:
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
SL-Ge ¼ 1
N
X
i2N
SL-Gei ¼ 1
N
P
X
i2N
j2N;j≠id−1
n − 1
ij
:
Netzwerkneurowissenschaften
367
Multilayer functional networks identify sex differences in aging
Single-layer clustering coefficient. The clustering coefficient of a given node represents the frac-
tion of the total number of triangles that are present around it. The network clustering coeffi-
cient is derived by averaging the clustering coefficients of all nodes. Using the definitions of
the number of triangles and nodal degree, the network clustering coefficient can be repre-
sented as (Mijalkov et al., 2017):
SL-CC ¼ 1
N
X
i2N
SL-CCi ¼ 1
N
X
i2N
2ti
ki ki − 1
D
Þ
:
ki, the degree of a node i, is defined as the total number of connections i has with other nodes
in the network, regardless of their weight. If the neighbors of node i are connected with each
andere, a triangle can be constructed around i. In weighted networks, the total number of trian-
gles around node i, ti, is calculated by summing the contributions of each individual triangle,
defined as the geometric mean of the triangle’s edge weights:
ti ¼ 1
2
X
J;h2N
ij w1=3
w1=3
jk w1=3
ki
:
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
Multiplex network measures. For each individual, we built a multiplex network with two layers,
one with positive connections and the other with negative connections (Abbildung 1C). The mul-
tiplex networks were calculated at densities ranging from 6% Zu 33% by combining the two
single-layer networks at the respective densities. At each density the multiplex networks can
be represented by a supra-adjacency matrix, W, which consists of the intralayer adjacency
matrices on the main diagonal and the interlayer connections in the off-diagonal entries.
The interlayer connections are formed only among the node’s replicas, das ist, Wj2β
i2α = 1 ⇔
i = j, where α and β denote layer 1 and layer 2, jeweils. We evaluated the topology of
these networks by calculating the participation and clustering coefficients and evaluating the
AUC over the complete density range.
Multiplex participation coefficient. This measure, MP-Pt, is used to quantify the heterogeneity in
the connectivity patterns of a node across the different layers. It is calculated as:
MP-Pti ¼ Nl
Nl − 1
”
1 −
!
2
α½
(cid:2)
S
ich
oi
#
;
XM
α¼1
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
where Nl is the number of layers and s
(cid:2)
α½
ich
is the strength of node i at the α-th layer, defined as
P
the sum of the weights of all edges connected to i. Endlich, oi =
is the overlapping
strength of the node i. MP-Pti determines whether i’s connections are evenly spread throughout
the layers or are largely concentrated in one or a few layers. MP-Pti has values in the range 0 Zu
1 Und, in general, bigger MP-Pti values suggest a more equal distribution of node i’s
connections in all layers of the multiplex network. The global participation coefficient
MP-Pt of the multiplex network is defined as the average of nodal MP-Pti coefficients over
all nodes.
α s
(cid:2)
α½
ich
Multiplex clustering coefficient. The clustering coefficient can be used to assess whether the
clustering features of the aggregated multiplex network differed from those of the individual
layers. In contrast to single-layer clustering, a triangle in a multiplex network is formed by one
Netzwerkneurowissenschaften
368
Multilayer functional networks identify sex differences in aging
edge from one layer and the remaining two edges from the other layer. daher, the multi-
plex clustering coefficient of a node i (MP-CCi) is defined as:
P
P
MP-CCi ¼
α
β≠α
D
M − 1
P
(cid:2)
α½
(cid:2)
i≠m;j w
ij w
(cid:2)
P
α½
(cid:2)
α½
(cid:2)
Þ
αk
ich
ich
k
(cid:3)1
3
;
(cid:2)
α½
β½
(cid:2)
jmw
mi
(cid:3)
− 1
(cid:2)
α½
where w
ij denote the connection between the nodes i and j in the layer α and ki denotes the
degree of a node i. The global multiplex clustering coefficient is calculated as the average of
nodal clustering coefficients across all nodes.
Multilayer network measures. We constructed two-layer multilayer networks for each individ-
ual following an analogous procedure as the one of multiplex networks. Jedoch, in contrast
to the multiplex approach, we established weighted interlayer connections between all node
pairs to quantify the degree of relationship between the two layers. In diesem Rahmen, Die
strength of the between-layer relationship is an independent variable that can be controlled
by the weight of the interlayer connections. Due to the variability of the intralayer connection
weights in all subjects, we define the interlayer weights, wij, as a fraction of each participant’s
biggest absolute functional connection:
wij ¼ σ (cid:3) max wα
maxwβ
max
(cid:4)
(cid:5)
; ∀i 2 α and ∀j 2 β;
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
max and wβ
where wα
max denote the maximum weight within layers 1 Und 2 jeweils, and σ is
the fraction of the maximum weight. In our analyses, we have evaluated the multilayer in the σ
range of 0.05 Zu 1, in steps of 0.05. Previous studies have explored the possibility of including
interlayer connections with variable strengths in the case of magnetoencephalographic record-
ings (Buldú & Porter, 2018), which could open possibilities for future studies to extend these
Maßnahmen. In our study we fixed these strengths to have the same weight for an easier inter-
pretation of our method and the corresponding results.
Multilayer global efficiency. Consider a node i, which is part of layer 1. The multilayer global
efficiency of this node, denoted by ML-Gei, reflects the difference between the average inverse
distances from i to all nodes in layer 2 and layer 1. Speziell, we first calculate the “same-
layer” global efficiency, SameL-Gei, using the methods defined in the section Single-Layer
Global Efficiency, which considers only the shortest paths between i and all nodes belonging
to layer 1. Note that the SameL-Gei does not depend on the interlayer weights as it considers
only the shortest distances between nodes within the same layer:
SameL-Gei ¼
P
j2αd−1
ij
n − 1
;
where n denotes the number of nodes and dij denotes the distance between the nodes i and j
as before. In the second step, we calculate the “opposite-layer” global efficiency, OppL-Gei,
which assesses the average inverse distances from i to all nodes in layer 2. Zu diesem Zweck, Wir
build a new network of size n + 1, which includes i and all nodes and connections of layer 2.
Dann, OppL-Gei can be calculated as:
OppL-Gei
σð Þ ¼
D
P
j2βd−1
ij
Þ − 1
n þ 1
:
Netzwerkneurowissenschaften
369
Multilayer functional networks identify sex differences in aging
We emphasize that OppL-Gei(σ) has an explicit dependence on σ since node i is connected
with the nodes in layer 2 via the interlayer connections of weight wij, which depend explicitly
on σ. Dann, the multilayer global efficiency of node i, ML-Gei, can be defined as:
ML-Gei σð Þ ¼ OL-Gei σð Þ − SL-Gei:
daher, ML-Gei measures the difference in global efficiency between the layer of negative
and positive connections at different strengths of interlayer relation that depend on the value of
σ. Higher values of ML-Gei suggest that the two layers have more divergent topology in terms
of information integration. In this case, the interlayer weights have larger impact of the topol-
ogy of the multilayer network, which leads to a stronger relationship between the layers.
Multilayer clustering coefficient. We defined this measure, ML-CCi(σ), analogously to the
multilayer global efficiency, in order to quantify the difference in the clustering properties
between the two layers at different strengths of between-layer relationship. We first calculated
SameL-CCi and OppL-CCi(σ) by following the procedures outlined in the section Single-Layer
Clustering Coefficient. Dann, the ML-CCi(σ) was calculated as:
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
N
e
N
_
A
_
0
0
2
8
6
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
ML-CCi σð Þ ¼ OppL-CCi
σð Þ − SameL-CCi;
and the network multilayer clustering coefficient was calculated as the average of ML-CCi(σ)
over all nodes in the network.
Partial least squares regression analysis (PLS). PLS performs a linear decomposition of the predic-
tor and predicted variable matrix into latent variables (LVs) optimized so that the covariance
between the resulting predictor and predicted matrix components (called factors and loadings,
jeweils) is maximal (Abdi & Williams, 2013). We fit a PLS regression model for each pre-
dicted variable (cognitive tests, structural measures, and prevalence of cardiovascular dis-
erleichtert) unabhängig. The optimal component number for the decomposition was determined
in each case by the Bayesian information criterion (BIC) based on the estimated degree of
freedom of the PLS model (Krämer & Sugiyama, 2011). Insbesondere, predictor and predicted
variable matrices were decomposed into 2–12 LVs, with the optimal number of LVs based on
BIC scores being defined as follows: attention = 5, executive = 6, memory = 2, thickness = 12,
visuospatial = 6, volumes = 5, WMH = 5, heart attack = 3, hypertension = 4. The contribution
of each variable to the prediction was quantified via variable importance in the projection
( VIP) scores, calculated as the sum of PLS weights over LVs, weighted by the variance
explained by each LV. Variables were defined as significant contributors to the prediction
based on a VIP score of > 1 (Chong & Jun, 2005).
Genetic Association Analyses
We examined the association between 9 million high-quality imputed common variants (MAF
> 1%, imputation INFO score > 0.8 and Hardy–Weinberg equilibrium P > 10−10) from UK
Biobank and each derived phenotype using linear mixed-effects models as implemented in
REGENIE (Jamialahmadi et al., 2021; Mbatchou et al., 2021). All traits were rank-based inverse
normal transformed before the analysis, and adjusted for age at MRI, Sex, age2, Alter * Sex,
age2 * Sex, body mass index (BMI), the first 10 principal components of ancestry, and genotyping
array. For step 1, we used a subset of high-quality directly genotyped variants as described
Vor (Jamialahmadi et al., 2021). We next performed a stringent physical LD clumping
(PLINK parameters: –clump-p1 5e-8 –clump-r2 0.05 –clump-kb 1000, after excluding
Netzwerkneurowissenschaften
370
Multilayer functional networks identify sex differences in aging
individuals with third-degree or closer relatives) (Bycroft et al., 2018; Chang et al., 2015),
followed by an approximate step-wise model selection in conditional and joint multiple-SNP
Analyse (COJO-GCTA) (Yang et al., 2012), with a window of 1 Mb and using 50,000 randomly
selected unrelated Europeans (in-sample LD structure) as described before (Jamialahmadi
et al., 2021).
Colocalization
Colocalization was performed between independent genetic loci from COJO-GCTA and
summary statistics of gene expression quantitative trait loci (eQTL) von 49 tissues in GTEx v8,
BrainSeq, ROSMAP, Braineac2 and CommonMind from eQTL cataolgue release 4 (GTEx
Konsortium, 2020; Guelfi et al., 2020; Jaffe et al., 2018; Kerimov et al., 2021; Ng et al.,
2017; Sieberts et al., 2020). All genes with at least one significant association (FDR adjusted
p value < 0.1) within a window of 1 Mb around each index variant were tested using coloc R
package (Giambartolomei et al., 2014) with default priors, and H4 posterior probability (PP) >
0.8 was considered as a strong evidence that both traits share the same causal variant.
Statistical Analysis
The statistical significance of the differences between men and women was assessed by per-
forming nonparametric permutation tests with 10,000 permutations, which were considered
significant for a two-tailed test of the null hypothesis at p < 0.05. These results were adjusted
for multiple comparisons by applying false discovery rate (FDR) corrections at q < 0.05 using
the Benjamini–Hochberg procedure (Benjamini & Hochberg, 1995). All results remained
statistically significant after adjusting for average cortical thickness and subcortical volumes,
suggesting that the observed between-sex differences were independent of brain atrophy (data
not shown). To detect sex differences over aging, we used linear regression models where all
functional connectivity measures for men and women at each age were summarized by their
average value due to the high overlap between brain measures across different ages. Then,
they were included as the dependent variables in the linear regression models, whereas
age, sex, age2, age * sex, and age2 * sex were included as independent variables. The best
model for each measure (Supporting Information: Linear Models) consisted of a combination
of predictors that resulted in the minimum value of the Akaike information criterion (AIC) for
that model. The significance of the overall model and the independent coefficients was eval-
uated by an F test, which was considered significant at p < 0.05. To compare the performance
of the different models, we used the AIC and the mean squared error of all models.
Data Availability
The authors did not participate in data collection. The primary data source used in the study
was the UK Biobank, which requires an application for access (https://www.ukbiobank.ac.uk/).
This study was conducted under application number 37142. All data from eQTL Catalogue
project are freely available at https://www.ebi.ac.uk/eqtl/.
Ethics Declarations
“UK Biobank has approval from the North West Multi-centre Research Ethics Committee
(MREC) as a Research Tissue Bank (RTB) approval.” (from https://www.ukbiobank.ac.uk
/learn-more-about-uk-biobank/about-us/ethics). Informed consent was obtained from all UK
Biobank participants. The current study is covered by this approval as we did not use any
additional data or re-contacted the participants.
Network Neuroscience
371
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Multilayer functional networks identify sex differences in aging
ACKNOWLEDGMENTS
We thank the following: Swedish Research Council; Swedish Alzheimer Foundation; Swedish
Brain Foundation; Strategic Research Area Neuroscience (StratNeuro); Center for Medical
Innovation (CIMED); Foundation for Geriatric Diseases at Karolinska Institutet; Gamla
Tjänarinnor; Stohnes Foundation; Lars Hierta Memorial Foundation.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00286.
AUTHOR CONTRIBUTIONS
Mite Mijalkov: Conceptualization; Formal analysis; Funding acquisition; Methodology; Visu-
alization; Writing – original draft; Writing – review & editing. Dániel Veréb: Formal analysis;
Methodology; Writing – original draft; Writing – review & editing. Oveis Jamialahmadi: Formal
analysis; Writing – original draft; Writing – review & editing. Anna Canal-Garcia: Software;
Visualization; Writing – review & editing. Emiliano Gómez-Ruiz: Software; Writing – review
& editing. Didac Vidal-Piñeiro: Writing – review & editing. Stefano Romeo: Formal analysis;
Writing – original draft; Writing – review & editing. Giovanni Volpe: Conceptualization;
Methodology; Supervision; Writing – original draft; Writing – review & editing. Joana Pereira:
Conceptualization; Funding acquisition; Methodology; Supervision; Writing – original draft;
Writing – review & editing.
FUNDING INFORMATION
Joana Pereira, Vetenskapsrådet (https://dx.doi.org/10.13039/501100004359). Joana Pereira,
Alzheimerfonden (https://dx.doi.org/10.13039/501100008599).
Joana Pereira, Hjärnfonden
(https://dx.doi.org/10.13039/501100003792). Joana Pereira, Strategic Research Area Neuro-
science (StratNeuro). Joana Pereira, Center for Medical Innovation (CIMED). Mite Mijalkov,
Foundation for Geriatric Diseases at Karolinska Institutet. Mite Mijalkov, Stiftelsen för Gamla
Tjänarinnor (https://dx.doi.org/10.13039/100010815). Mite Mijalkov, Gun och Bertil Stohnes
Stiftelse (https://dx.doi.org/10.13039/100009673). Mite Mijalkov, Stiftelsen Lars Hiertas Minne
(https://dx.doi.org/10.13039/501100004722).
REFERENCES
Abdi, H., & Williams, L. J. (2013). Partial least squares methods:
Partial least squares correlation and partial least square regres-
sion. In Computational toxicology (pp. 549–579). Springer.
https://doi.org/10.1007/978-1-62703-059-5_23, PubMed:
23086857
Alfaro-Almagro, F., Jenkinson, M., Bangerter, N. K., Andersson,
J. L. R., Griffanti, L., Douaud, G., … Smith, S. M. (2018). Image
processing and quality control for the first 10,000 brain imaging
datasets from UK Biobank. NeuroImage, 166, 400–424. https://
doi.org/10.1016/j.neuroimage.2017.10.034 , PubMed:
29079522
Allen, E. A., Erhardt, E. B., Damaraju, E., Gruner, W., Segall, J. M.,
Silva, R. F., … Calhoun, V. D. (2011). A baseline for the multivar-
iate comparison of resting-state networks. Frontiers in Systems
Neuroscience, 5, 2. https://doi.org/10.3389/fnsys.2011.00002,
PubMed: 21442040
Anand, S. S., Islam, S., Rosengren, A., Franzosi, M. G., Steyn, K.,
Yusufali, A. H., … Yusuf, S. (2008). Risk factors for myocardial
infarction in women and men: Insights from the interheart study.
European Heart Journal, 29(7), 932–940. https://doi.org/10.1093
/eurheartj/ehn018, PubMed: 18334475
Ancoli-Israel, S. (2009). Sleep and its disorders in aging popula-
tions. Sleep Medicine, 10, S7–S11. https://doi.org/10.1016/j
.sleep.2009.07.004, PubMed: 19647483
Barber, A. D., Caffo, B. S., Pekar, J. J., & Mostofsky, S. H. (2013).
Developmental changes in within-and between-network connec-
tivity between late childhood and adulthood. Neuropsychologia,
51(1), 156–167. https://doi.org/10.1016/j.neuropsychologia.2012
.11.011, PubMed: 23174403
Battiston, F., Nicosia, V., & Latora, V. (2014). Structural measures for
multiplex networks. Physical Review E, 89(3), 032804. https://doi
.org/10.1103/PhysRevE.89.032804, PubMed: 24730896
Network Neuroscience
372
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Multilayer functional networks identify sex differences in aging
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society: Series B (Methodological),
57(1), 289–300. https://doi.org/10.1111/j.2517-6161.1995
.tb02031.x
Betzel, R. F., Byrge, L., He, Y., Goñi, J., Zuo, X.-N., & Sporns, O.
(2014). Changes in structural and functional connectivity among
resting-state networks across the human lifespan. NeuroImage,
102, 345–357. https://doi.org/10.1016/j.neuroimage.2014.07
.067, PubMed: 25109530
Biswal, B., Zerrin Yetkin, F., Haughton, V. M., & Hyde, J. S. (1995).
Functional connectivity in the motor cortex of resting human
brain using echo-planar MRI. Magnetic Resonance in Medicine,
34(4), 537–541. https://doi.org/10.1002/mrm.1910340409,
PubMed: 8524021
Biswal, B. B., Mennes, M., Zuo, X.-N., Gohel, S., Kelly, C., Smith,
S. M., … Milham, M. P. (2010). Toward discovery science of
human brain function. Proceedings of the National Academy of
Sciences, 107(10), 4734–4739. https://doi.org/10.1073/pnas
.0911855107, PubMed: 20176931
Buldú, J. M., & Porter, M. A. (2018). Frequency-based brain net-
works: From a multiplex framework to a full multilayer descrip-
tion. Network Neuroscience, 2(4), 418–441. https://doi.org/10
.1162/netn_a_00033, PubMed: 30294706
Bullmore, E., & Sporns, O. (2012). The economy of brain network
organization. Nature Reviews Neuroscience, 13(5), 336–349.
https://doi.org/10.1038/nrn3214, PubMed: 22498897
Bycroft, C., Freeman, C., Petkova, D., Band, G., Elliott, L. T.,
Sharp, K., … Marchini, J. (2018). The UK Biobank resource with
deep phenotyping and genomic data. Nature, 562(7726),
203–209. https://doi.org/10.1038/s41586-018-0579-z, PubMed:
30305743
Cabeza, R., Anderson, N. D., Locantore, J. K., & McIntosh, A. R.
(2002). Aging gracefully: Compensatory brain activity in
high-performing older adults. NeuroImage, 17(3), 1394–1402.
https://doi.org/10.1006/nimg.2002.1280, PubMed: 12414279
Chai, X. J., Castañón, A. N., Öngür, D., & Whitfield-Gabrieli, S.
(2012). Anticorrelations in resting state networks without global
signal regression. NeuroImage, 59(2), 1420–1428. https://doi.org
/10.1016/j.neuroimage.2011.08.048, PubMed: 21889994
Chai, X. J., Ofen, N., Gabrieli, J. D. E., & Whitfield-Gabrieli, S.
(2014). Selective development of anticorrelated networks in the
intrinsic functional organization of the human brain. Journal of
Cognitive Neuroscience, 26(3), 501–513. https://doi.org/10
.1162/jocn_a_00517, PubMed: 24188367
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig, G. S.
(2014). Decreased segregation of brain systems across the
healthy adult lifespan. Proceedings of the National Academy of
Sciences, 111(46), E4997–E5006. https://doi.org/10.1073/pnas
.1415122111, PubMed: 25368199
Chang, C. C., Chow, C. C., Tellier, L. C., Vattikuti, S., Purcell, S. M.,
& Lee, J. J. (2015). Second-generation PLINK: Rising to the chal-
lenge of larger and richer datasets. Gigascience, 4(1), s13742-
015-0047-8. https://doi.org/10.1186/s13742-015-0047-8,
PubMed: 25722852
Chekroud, A. M., Ward, E. J., Rosenberg, M. D., & Holmes, A. J.
(2016). Patterns in the human brain mosaic discriminate males
from females. Proceedings of the National Academy of Sciences,
113(14), E1968. https://doi.org/10.1073/pnas.1523888113,
PubMed: 26984491
Chen, Y.-C., Jiao, Y., Cui, Y., Shang, S.-A., Ding, J., Feng, Y., … Teng,
G.-J. (2014). Aberrant brain functional connectivity related to
insulin resistance in type 2 diabetes: A resting-state fMRI study.
Diabetes Care, 37(6), 1689–1696. https://doi.org/10.2337/dc13
-2127, PubMed: 24658392
Chong, I.-G., & Jun, C.-H. (2005). Performance of some variable
selection methods when multicollinearity is present. Chemo-
metrics and Intelligent Laboratory Systems, 78(1–2), 103–112.
https://doi.org/10.1016/j.chemolab.2004.12.011
Cicchetti, D. V., & Sparrow, S. A. (1981). Developing criteria for
establishing interrater reliability of specific items: Applications
to assessment of adaptive behavior. American Journal of Mental
Deficiency, 86(2), 127–137. PubMed: 7315877
Cosgrove, K. P., Mazure, C. M., & Staley, J. K. (2007). Evolving
knowledge of sex differences in brain structure, function, and
chemistry. Biological Psychiatry, 62(8), 847–855. https://doi.org
/10.1016/j.biopsych.2007.03.001, PubMed: 17544382
Crimmins, E. M. (2015). Lifespan and healthspan: Past, present, and
promise. The Gerontologist, 55(6), 901–911. https://doi.org/10
.1093/geront/gnv130, PubMed: 26561272
Damoiseaux, J. S. (2017). Effects of aging on functional and struc-
tural brain connectivity. NeuroImage, 160, 32–40. https://doi.org
/10.1016/j.neuroimage.2017.01.077, PubMed: 28159687
Deary, I. J., Corley, J., Gow, A. J., Harris, S. E., Houlihan, L. M.,
Marioni, R. E., … Starr, J. M. (2009). Age-associated cognitive
decline. British Medical Bulletin, 92(1), 135–152. https://doi.org
/10.1093/bmb/ldp033, PubMed: 19776035
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., … Killiany, R. J. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI scans
into gyral based regions of interest. NeuroImage, 31(3), 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021, PubMed:
16530430
Dhingra, R., & Vasan, R. S. (2012). Age as a risk factor. Medical
Clinics, 96(1), 87–91. https://doi.org/10.1016/j.mcna.2011.11
.003, PubMed: 22391253
Elliott, L. T., Sharp, K., Alfaro-Almagro, F., Shi, S., Miller, K. L.,
Douaud, G., … Smith, S. M. (2018). Genome-wide association
studies of brain imaging phenotypes in UK Biobank. Nature,
562(7726), 210–216. https://doi.org/10.1038/s41586-018-0571
-7, PubMed: 30305740
Fawns-Ritchie, C., & Deary, I. J. (2020). Reliability and validity of the
UK Biobank cognitive tests. PLoS One, 15(4), e0231627. https://
doi.org/10.1371/journal.pone.0231627, PubMed: 32310977
Ferreira, L. K., Regina, A. C. B., Kovacevic, N., Martin, M. d. G. M.,
Santos, P. P., Carneiro, C. d. G., … Busatto, G. F. (2016). Aging
effects on whole-brain functional connectivity in adults free of
cognitive and psychiatric disorders. Cerebral Cortex, 26(9),
3851–3865. https://doi.org/10.1093/cercor/ bhv190, PubMed:
26315689
Ferretti, M. T., Iulita, M. F., Cavedo, E., Chiesa, P. A., Dimech, A. S.,
Chadha, A. S., … Women’s Brain Project and the Alzheimer Pre-
cision Medicine Initiative. (2018). Sex differences in Alzheimer
disease—The gateway to precision medicine. Nature Reviews
Neurology, 14(8), 457–469. https://doi.org/10.1038/s41582-018
-0032-9, PubMed: 29985474
Network Neuroscience
373
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Multilayer functional networks identify sex differences in aging
Filippi, M., Valsasina, P., Misci, P., Falini, A., Comi, G., & Rocca,
M. A. (2013). The organization of intrinsic brain activity differs
between genders: A resting-state fMRI study in a large cohort
of young healthy subjects. Human Brain Mapping, 34(6),
1330–1343. https://doi.org/10.1002/ hbm.21514, PubMed:
22359372
Foo, H., Thalamuthu, A., Jiang, J., Koch, F. C., Mather, K. A., Wen,
W., & Sachdev, P. S. (2021a). Age- and sex-related topological
organization of human brain functional networks and their relation-
ship to cognition. Frontiers in Aging Neuroscience, 13, 758817.
https://doi.org/10.3389/fnagi.2021.758817, PubMed: 34975453
Foo, H., Thalamuthu, A., Jiang, J., Koch, F. C., Mather, K. A., Wen,
W., & Sachdev, P. S. (2021b). Novel genetic variants associated
with brain functional networks in 18,445 adults from the UK
Biobank. Scientific Reports, 11(1), 14633. https://doi.org/10
.1038/s41598-021-94182-9, PubMed: 34272439
Fornito, A., Zalesky, A., & Breakspear, M. (2013). Graph analysis of
the human connectome: Promise, progress, and pitfalls. Neuro-
Image, 80, 426–444. https://doi.org/10.1016/j.neuroimage.2013
.04.087, PubMed: 23643999
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences, 102(27),
9673–9678. https://doi.org/10.1073/pnas.0504136102,
PubMed: 15976020
Fox, M. D., Zhang, D., Snyder, A. Z., & Raichle, M. E. (2009). The
global signal and observed anticorrelated resting state brain net-
works. Journal of Neurophysiology, 101(6), 3270–3283. https://
doi.org/10.1152/jn.90777.2008, PubMed: 19339462
Geerligs, L., Renken, R. J., Saliasi, E., Maurits, N. M., & Lorist,
M. M. (2015). A brain-wide study of age-related changes in func-
tional connectivity. Cerebral Cortex, 25(7), 1987–1999. https://
doi.org/10.1093/cercor/bhu012, PubMed: 24532319
Giambartolomei, C., Vukcevic, D., Schadt, E. E., Franke, L., Hingorani,
A. D., Wallace, C., & Plagnol, V. (2014). Bayesian test for colo-
calisation between pairs of genetic association studies using
summary statistics. PLoS Genetics, 10(5), e1004383. https://doi
.org/10.1371/journal.pgen.1004383, PubMed: 24830394
Gillis, E. E., & Sullivan, J. C. (2016). Sex differences in hypertension:
Recent advances. Hypertension, 68(6), 1322–1327. https://doi
.org/10.1161/ HYPERTENSIONAHA.116.06602, PubMed:
27777357
Goelman, G., Gordon, N., & Bonne, O. (2014). Maximizing nega-
tive correlations in resting-state functional connectivity MRI by
time-lag. PLoS One, 9(11), e111554. https://doi.org/10.1371
/journal.pone.0111554, PubMed: 25396416
Golden, L. C., & Voskuhl, R. (2017). The importance of studying sex
differences in disease: The example of multiple sclerosis. Journal
of Neuroscience Research, 95(1–2), 633–643. https://doi.org/10
.1002/jnr.23955, PubMed: 27870415
Goldstone, A., Mayhew, S. D., Przezdzik, I., Wilson, R. S., Hale,
J. R., & Bagshaw, A. P. (2016). Gender specific re-organization
of resting-state networks in older age. Frontiers in Aging Neuro-
science, 8, 285. https://doi.org/10.3389/fnagi.2016.00285,
PubMed: 27932978
Griffa, A., Baumann, P. S., Thiran, J.-P., & Hagmann, P. (2013).
Structural connectomics in brain diseases. NeuroImage, 80,
515–526. https://doi.org/10.1016/j.neuroimage.2013.04.056,
PubMed: 23623973
Griffanti, L., Zamboni, G., Khan, A., Li, L., Bonifacio, G., Sundaresan,
V., … Jenkinson, M. (2016). BIANCA (Brain Intensity AbNormality
Classification Algorithm): A new tool for automated segmentation
of white matter hyperintensities. NeuroImage, 141, 191–205.
https://doi.org/10.1016/j.neuroimage.2016.07.018, PubMed:
27402600
GTEx Consortium. (2020). The GTEx Consortium atlas of genetic
regulatory effects across human tissues. Science, 369(6509),
1318–1330. https://doi.org/10.1126/science.aaz1776, PubMed:
32913098
Guelfi, S., D’Sa, K., Botía, J. A., Vandrovcova, J., Reynolds, R. H.,
Zhang, D., … Ryten, M. (2020). Regulatory sites for splicing in
human basal ganglia are enriched for disease-relevant informa-
tion. Nature Communications, 11(1), 1041. https://doi.org/10
.1038/s41467-020-14483-x, PubMed: 32098967
Hampson, M., Driesen, N., Roth, J. K., Gore, J. C., & Constable,
R. T. (2010). Functional connectivity between task-positive and
task-negative brain areas and its relation to working memory per-
formance. Magnetic Resonance Imaging, 28(8), 1051–1057.
https://doi.org/10.1016/j.mri.2010.03.021, PubMed: 20409665
Hou, Y., Dan, X., Babbar, M., Wei, Y., Hasselbalch, S. G., Croteau,
D. L., & Bohr, V. A. (2019). Ageing as a risk factor for neurode-
generative disease. Nature Reviews Neurology, 15(10), 565–581.
https://doi.org/10.1038/s41582-019-0244-7, PubMed:
31501588
Jaffe, A. E., Straub, R. E., Shin, J. H., Tao, R., Gao, Y., Collado-
Torres, L., … Weinberger, D. R. (2018). Developmental and
genetic regulation of the human cortex transcriptome illuminate
schizophrenia pathogenesis. Nature Neuroscience, 21(8),
1117–1125. https://doi.org/10.1038/s41593-018-0197-y,
PubMed: 30050107
Jamialahmadi, O., Mancina, R. M., Ciociola, E., Tavaglione, F.,
Luukkonen, P. K., Baselli, G., … Romeo, S. (2021). Exome-wide
association study on alanine aminotransferase identifies
sequence variants in the GPAM and APOE associated with fatty
liver disease. Gastroenterology, 160(5), 1634–1646. https://doi
.org/10.1053/j.gastro.2020.12.023, PubMed: 33347879
Joel, D., & Fausto-Sterling, A. (2016). Beyond sex differences: New
approaches for thinking about variation in brain structure and
function. Philosophical Transactions of the Royal Society B: Bio-
logical Sciences, 371(1688), 20150451. https://doi.org/10.1098
/rstb.2015.0451, PubMed: 26833844
Jones, S. E., van Hees, V. T., Mazzotti, D. R., Marques-Vidal, P.,
Sabia, S., van der Spek, A., … Wood, A. R. (2019). Genetic stud-
ies of accelerometer-based sleep measures yield new insights
into human sleep behaviour. Nature Communications, 10(1),
1585. https://doi.org/10.1038/s41467-019-09576-1, PubMed:
30952852
Keller, J. B., Hedden, T., Thompson, T. W., Anteraper, S. A.,
Gabrieli, J. D., & Whitfield-Gabrieli, S. (2015). Resting-state
anticorrelations between medial and lateral prefrontal cortex:
Association with working memory, aging, and individual differ-
ences. Cortex, 64, 271–280. https://doi.org/10.1016/j.cortex
.2014.12.001, PubMed: 25562175
Kerimov, N., Hayhurst, J. D., Peikova, K., Manning, J. R., Walter, P.,
Kolberg, L., … Alasoo, K. (2021). A compendium of uniformly
Network Neuroscience
374
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Multilayer functional networks identify sex differences in aging
processed human gene expression and splicing quantitative trait
loci. Nature Genetics, 53(9), 1290–1299. https://doi.org/10.1038
/s41588-021-00924-w, PubMed: 34493866
Klein, S. L., Neilson, K. M., Orban, J., Yaklichkin, S., Hoffbauer, J.,
Mood, K., … Moody, S. A. (2013). Conserved structural domains
in FoxD4L1, a neural forkhead box transcription factor, are
required to repress or activate target genes. PLoS One, 8(4),
e61845. https://doi.org/10.1371/journal.pone.0061845,
PubMed: 23610594
Krämer, N., & Sugiyama, M. (2011). The degrees of freedom of par-
tial least squares regression. Journal of the American Statistical
Association, 106(494), 697–705. https://doi.org/10.1198/jasa
.2011.tm10107
Li, L., Pan, Z., & Yang, X. (2019). Key genes and co-expression net-
work analysis in the livers of type 2 diabetes patients. Journal of
Diabetes Investigation, 10(4), 951–962. https://doi.org/10.1111
/jdi.12998, PubMed: 30592156
Liu, S., Wang, Y.-S., Zhang, Q., Zhou, Q., Cao, L.-Z., Jiang, C., …
Chinese Color Nest Consortium. (2021). Chinese Color Nest Pro-
ject: An accelerated longitudinal brain-mind cohort. Develop-
mental Cognitive Neuroscience, 52, 101020. https://doi.org/10
.1016/j.dcn.2021.101020, PubMed: 34653938
Lord, L.-D., Stevner, A. B., Deco, G., & Kringelbach, M. L. (2017).
Understanding principles of integration and segregation using
whole-brain computational connectomics: implications for neu-
ropsychiatric disorders. Philosophical Transactions of the Royal
Society A: Mathematical, Physical, and Engineering Sciences,
375(2096), 20160283. https://doi.org/10.1098/rsta.2016.0283,
PubMed: 28507228
Marek, S., Tervo-Clemmens, B., Calabro, F. J., Montez, D. F., Kay,
B. P., Hatoum, A. S., … Dosenbach, N. U. F. (2022). Reproducible
brain-wide association studies require thousands of individuals.
Nature, 603(7902), 654–660. https://doi.org/10.1038/s41586
-022-04492-9, PubMed: 35296861
Masuda, N., Sakaki, M., Ezaki, T., & Watanabe, T. (2018). Cluster-
ing coefficients for correlation networks. Frontiers in Neuroinfor-
matics, 12, 7. https://doi.org/10.3389/fninf.2018.00007,
PubMed: 29599714
Mazure, C. M., & Swendsen, J. (2016). Sex differences in Alzheimer’s
disease and other dementias. The Lancet Neurology, 15(5),
451–452. https://doi.org/10.1016/S1474-4422(16)00067-3,
PubMed: 26987699
Mbatchou, J., Barnard, L., Backman, J., Marcketta, A., Kosmicki,
J. A., Ziyatdinov, A., … Marchini, J. (2021). Computationally effi-
cient whole-genome regression for quantitative and binary traits.
Nature Genetics, 53(7), 1097–1103. https://doi.org/10.1038
/s41588-021-00870-7, PubMed: 34017140
McCarrey, A. C., An, Y., Kitner-Triolo, M. H., Ferrucci, L., &
Resnick, S. M. (2016). Sex differences in cognitive trajectories
in clinically normal older adults. Psychology and Aging, 31(2),
166–175. https://doi.org/10.1037/pag0000070, PubMed:
26796792
McGraw, K. O., & Wong, S. P. (1996). Forming inferences about
some intraclass correlation coefficients. Psychological Methods,
1(1), 30–46. https://doi.org/10.1037/1082-989X.1.1.30
Mijalkov, M., Kakaei, E., Pereira, J. B., Westman, E., Volpe, G., &
Alzheimer’s Disease Neuroimaging Initiative. (2017). BRAPH:
A graph theory software for the analysis of brain connectivity.
PLoS One, 12(8), e0178798. https://doi.org/10.1371/journal
.pone.0178798, PubMed: 28763447
Mijalkov, M., Volpe, G., & Pereira, J. B. (2022). Directed brain con-
nectivity identifies widespread functional network abnormalities
in Parkinson’s disease. Cerebral Cortex, 32(3), 593–607. https://
doi.org/10.1093/cercor/bhab237, PubMed: 34331060
Miller, K. L., Alfaro-Almagro, F., Bangerter, N. K., Thomas, D. L.,
Yacoub, E., Xu, J., … Smith, S. M. (2016). Multimodal population
brain imaging in the UK Biobank prospective epidemiological
study. Nature Neuroscience, 19(11), 1523–1536. https://doi.org
/10.1038/nn.4393, PubMed: 27643430
Neitzel, J., Malik, R., Muetzel, R., Knol, M. J., Zonneveld, H.,
Georgakis, M. K., … Ewers, M. (2021). Genetic variants link
lower segregation of brain networks to higher blood pressure
and worse cognition within the general aging population.
medRxiv. https://doi.org/10.1101/2021.08.12.21261975
Ng, B., White, C. C., Klein, H.-U., Sieberts, S. K., McCabe, C.,
Patrick, E., … De Jager, P. L. (2017). An xQTL map integrates
the genetic architecture of the human brain’s transcriptome
and epigenome. Nature Neuroscience, 20(10), 1418–1426.
https://doi.org/10.1038/nn.4632, PubMed: 28869584
O’Brien, L. M., Ziegler, D. A., Deutsch, C. K., Frazier, J. A., Herbert,
M. R., & Locascio, J. J. (2011). Statistical adjustments for brain
size in volumetric neuroimaging studies: Some practical implica-
tions in methods. Psychiatry Research: Neuroimaging, 193(2),
113–122. https://doi.org/10.1016/j.pscychresns.2011.01.007,
PubMed: 21684724
Park, D. C., & Reuter-Lorenz, P. (2009). The adaptive brain: Aging
and neurocognitive scaffolding. Annual Review of Psychology,
60, 173–196. https://doi.org/10.1146/annurev.psych.59.103006
.093656, PubMed: 19035823
Prata, D. P., Papagni, S. A., Mechelli, A., Fu, C. H. Y., Kambeitz, J.,
Picchioni, M., … McGuire, P. K. (2012). Effect of D-amino acid
oxidase activator (DAOA; G72) on brain function during verbal
fluency. Human Brain Mapping, 33(1), 143–153. https://doi.org
/10.1002/hbm.21198, PubMed: 21391259
Ramirez, L. A., & Sullivan, J. C. (2018). Sex differences in hyperten-
sion: Where we have been and where we are going. American
Journal of Hypertension, 31(12), 1247–1254. https://doi.org/10
.1093/ajh/hpy148, PubMed: 30299518
Ritchie, S. J., Cox, S. R., Shen, X., Lombardo, M. V., Reus, L. M.,
Alloza, C., … Deary, I. J. (2018). Sex differences in the adult
human brain: Evidence from 5216 UK Biobank participants.
Cerebral Cortex, 28(8), 2959–2975. https://doi.org/10.1093
/cercor/bhy109, PubMed: 29771288
Ruigrok, A. N. V., Salimi-Khorshidi, G., Lai, M.-C., Baron-Cohen,
S., Lombardo, M. V., Tait, R. J., & Suckling, J. (2014). A
meta-analysis of sex differences in human brain structure. Neu-
roscience & Biobehavioral Reviews, 39, 34–50. https://doi.org/10
.1016/j.neubiorev.2013.12.004, PubMed: 24374381
Saberi, M., Khosrowabadi, R., Khatibi, A., Misic, B., & Jafari, G.
(2021). Topological impact of negative links on the stability of
resting-state brain network. Scientific Reports, 11(1), 2176.
https://doi.org/10.1038/s41598-021-81767-7, PubMed:
33500525
Sachdev, P. S., Parslow, R., Wen, W., Anstey, K., & Easteal, S.
(2009). Sex differences in the causes and consequences of white
matter hyperintensities. Neurobiology of Aging, 30(6), 946–956.
Network Neuroscience
375
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Multilayer functional networks identify sex differences in aging
https://doi.org/10.1016/j.neurobiolaging.2007.08.023, PubMed:
17950492
Sala-Llonch, R., Bartrés-Faz, D., & Junqué, C. (2015). Reorganiza-
tion of brain networks in aging: A review of functional connec-
tivity studies. Frontiers in Psychology, 6, 663. https://doi.org/10
.3389/fpsyg.2015.00663, PubMed: 26052298
Satterthwaite, T. D., Wolf, D. H., Roalf, D. R., Ruparel, K., Erus, G.,
Vandekar, S., … Gur, R. C. (2015). Linked sex differences in
cognition and functional connectivity in youth. Cerebral Cortex,
25(9), 2383–2394. https://doi.org/10.1093/cercor/ bhu036,
PubMed: 24646613
Scheinost, D., Finn, E. S., Tokoglu, F., Shen, X., Papademetris, X.,
Hampson, M., & Constable, R. T. (2015). Sex differences in nor-
mal age trajectories of functional brain networks. Human Brain
Mapping, 36(4), 1524–1535. https://doi.org/10.1002/ hbm
.22720, PubMed: 25523617
Sieberts, S. K., Perumal, T. M., Carrasquillo, M. M., Allen, M.,
Reddy, J. S., Hoffman, G. E., … Mangravite, L. M. (2020). Large
eQTL meta-analysis reveals differing patterns between cerebral
cortical and cerebellar brain regions. Scientific Data, 7(1), 340.
https://doi.org/10.1038/s41597-020-00642-8, PubMed:
33046718
Smith, K. M., & Dahodwala, N. (2014). Sex differences in Parkin-
son’s disease and other movement disorders. Experimental Neu-
rology, 259, 44–56. https://doi.org/10.1016/j.expneurol.2014.03
.010, PubMed: 24681088
Smith, S. M., Elliott, L. T., Alfaro-Almagro, F., McCarthy, P., Nichols,
T. E., Douaud, G., & Miller, K. L. (2020). Brain aging comprises
many modes of structural and functional change with distinct
genetic and biophysical associations. Elife, 9, e52677. https://
doi.org/10.7554/eLife.52677, PubMed: 32134384
Spreng, R. N., Stevens, W. D., Viviano, J. D., & Schacter, D. L.
(2016). Attenuated anticorrelation between the default and dor-
sal attention networks with aging: Evidence from task and rest.
Neurobiology of Aging, 45, 149–160. https://doi.org/10.1016/j
.neurobiolaging.2016.05.020, PubMed: 27459935
Stumme, J., Jockwitz, C., Hoffstaedter, F., Amunts, K., & Caspers, S.
(2020). Functional network reorganization in older adults:
Graph-theoretical analyses of age, cognition and sex. Neuro-
Image, 214, 116756. https://doi.org/10.1016/j.neuroimage.2020
.116756, PubMed: 32201326
Tagliazucchi, E., von Wegner, F., Morzelewski, A., Borisov, S.,
Jahnke, K., & Laufs, H. (2012). Automatic sleep staging using
fMRI functional connectivity data. NeuroImage, 63(1), 63–72.
https://doi.org/10.1016/j.neuroimage.2012.06.036, PubMed:
22743197
Tomasi, D., & Volkow, N. D. (2012a). Aging and functional brain
networks. Molecular Psychiatry, 17(5), 549–558. https://doi.org
/10.1038/mp.2011.81, PubMed: 21727896
Tomasi, D., & Volkow, N. D. (2012b). Gender differences in brain
functional connectivity density. Human Brain Mapping, 33(4),
849–860. https://doi.org/10.1002/ hbm.21252, PubMed:
21425398
van den Heuvel, M. P., & Pol, H. E. H. (2010). Exploring the brain
network: A review on resting-state fMRI functional connectivity.
European Neuropsychopharmacology, 20(8), 519–534. https://
doi.org/10.1016/j.euroneuro.2010.03.008, PubMed: 20471808
Vidal-Piñeiro, D., Wang, Y., Krogsrud, S. K., Amlien, I. K., Baaré,
W. F. C., Bartres-Faz, D., … Fjell, A. (2021). Individual variations
in ‘brain age’ relate to early-life factors more than to longitudinal
brain change. Elife, 10, e69995. https://doi.org/10.7554/eLife
.69995, PubMed: 34756163
Wang, Y., Xu, Q., Luo, J., Hu, M., & Zuo, C. (2019). Effects of age
and sex on subcortical volumes. Frontiers in Aging Neuroscience,
11, 259. https://doi.org/10.3389/fnagi.2019.00259, PubMed:
31616285
Weiss, E. M., Kemmler, G., Deisenhammer, E. A., Fleischhacker,
W. W., & Delazer, M. (2003). Sex differences in cognitive func-
tions. Personality and Individual Differences, 35(4), 863–875.
https://doi.org/10.1016/S0191-8869(02)00288-X
Wyss-Coray, T. (2016). Ageing, neurodegeneration and brain reju-
venation. Nature, 539(7628), 180–186. https://doi.org/10.1038
/nature20411, PubMed: 27830812
Xing, X.-X. (2021). Globally aging cortical spontaneous activity
revealed by multiple metrics and frequency bands using
resting-state functional MRI. Frontiers in Aging Neuroscience,
13, 803436. https://doi.org/10.3389/fnagi.2021.803436,
PubMed: 35027890
Yang, J., Ferreira, T., Morris, A. P., Medland, S. E., Madden, P. A. F.,
Heath, A. C., … Visscher, P. M. (2012). Conditional and joint
multiple-SNP analysis of GWAS summary statistics identifies
additional variants influencing complex traits. Nature Genetics,
44(4), 369–375. https://doi.org/10.1038/ng.2213, PubMed:
22426310
Zalesky, A., Fornito, A., & Bullmore, E. (2012). On the use of
correlation as a measure of network connectivity. NeuroImage,
60(4), 2096–2106. https://doi.org/10.1016/j.neuroimage.2012
.02.001, PubMed: 22343126
Zhang, C., Cahill, N. D., Arbabshirani, M. R., White, T., Baum,
S. A., & Michael, A. M. (2016). Sex and age effects of functional
connectivity in early adulthood. Brain Connectivity, 6(9),
700–713. https://doi.org/10.1089/ brain.2016.0429, PubMed:
27527561
Zhang, C., Dougherty, C. C., Baum, S. A., White, T., & Michael,
A. M. (2018). Functional connectivity predicts gender: Evidence
for gender differences in resting brain connectivity. Human Brain
Mapping, 39(4), 1765–1776. https://doi.org/10.1002/ hbm
.23950, PubMed: 29322586
Network Neuroscience
376
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
3
5
1
2
0
7
2
1
1
1
n
e
n
_
a
_
0
0
2
8
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3