FORSCHUNG
Intermittent brain network reconfigurations and
the resistance to social media influence
Italo’Ivo Lima Dias Pinto1, Nuttida Rungratsameetaweemana2, Kristen Flaherty1,3,
Aditi Periyannan1,4, Amir Meghdadi5, Christian Richard5, Chris Berka5,
Kanika Bansal1,6*, and Javier Omar Garcia1*
1US DEVCOM Army Research Laboratory, Aberdeen Proving Ground, MD, USA
2The Salk Institute for Biological Studies, La Jolla, CA, USA
3Cornell Tech, New York, New York, USA
4Tufts University, Medford, MA, USA
5Advanced Brain Monitoring, Carlsbad, CA, USA
6Abteilung für Biomedizintechnik, Columbia University, New York, New York, USA
*Co-senior authors.
Schlüsselwörter: Social media, Decision-making, EEG, Flexibility, Network reconfigurations
ABSTRAKT
Since its development, social media has grown as a source of information and has a significant
impact on opinion formation. Individuals interact with others and content via social media
platforms in a variety of ways, but it remains unclear how decision-making and associated
neural processes are impacted by the online sharing of informational content, from factual to
fabricated. Hier, we use EEG to estimate dynamic reconfigurations of brain networks and
probe the neural changes underlying opinion change (or formation) within individuals
interacting with a simulated social media platform. Our findings indicate that the individuals
who changed their opinions are characterized by less frequent network reconfigurations while
those who did not change their opinions tend to have more flexible brain networks with
frequent reconfigurations. The nature of these frequent network configurations suggests a
fundamentally different thought process between intervals in which individuals are easily
influenced by social media and those in which they are not. We also show that these
reconfigurations are distinct to the brain dynamics during an in-person discussion with
strangers on the same content. Zusammen, these findings suggest that brain network
reconfigurations may not only be diagnostic to the informational context but also the
underlying opinion formation.
ZUSAMMENFASSUNG DES AUTORS
Distinctive neural underpinnings of opinion formation and change during in-person and
online social interactions are not well understood. Hier, we analyze EEG recordings of the
participants interacting with a simulated social media platform and during an in-person
discussion by using a network-based analysis approach. We show that the structure of network
reconfigurations during these interactions is diagnostic of the opinion change and the context
in which information was received.
Keine offenen Zugänge
Tagebuch
Zitat: Lima Dias Pinto, ICH.,
Rungratsameetaweemana, N., Flaherty,
K., Periyannan, A., Meghdadi, A.,
Richard, C., Berka, C., Bansal, K., &
Garcia, J. Ö. (2022). Intermittent brain
network reconfigurations and the
resistance to social media influence.
Netzwerkneurowissenschaften, 6(3), 870–896.
https://doi.org/10.1162/netn_a_00255
DOI:
https://doi.org/10.1162/netn_a_00255
zusätzliche Informationen:
https://doi.org/10.1162/netn_a_00255
Erhalten: 23 November 2021
Akzeptiert: 10 Mai 2022
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Corresponding Authors:
Kanika Bansal
phy.kanika@gmail.com
Javier Omar Garcia
javier.o.garcia.civ@army.mil
Handling-Editor:
Vince Calhoun
Urheberrechte ©: © 2022
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
N
e
N
_
A
_
0
0
2
5
5
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network reconfigurations and social media
EINFÜHRUNG
Decision-making is the internal process by which information is reduced to a categorical and
actionable proposition (for review, see Gold & Shadlen, 2007). In the brain, the decision-
making process has been described as a nonlinear, context-dependent process that requires
a variety of brain areas to receive and interpret information (z.B., sensory), establish value of
this information, and then, based on prior experience and motivation, use a decision variable
to produce the proposition and subsequently act (Fellows, 2004). One context that is currently
and almost ubiquitously used as a source of information is social media, the suite of interactive
online technologies that have become a mainstay of not only our everyday interactions but
also current events and global happenings (Westerman et al., 2014). Because of the ubiquitous
nature of social media, the unbridled spread of information through it (Yoo et al., 2016), Und
the potentially negative consequences of it (Keles et al., 2020), it is important to understand
how it shapes our thoughts, influences our opinions, and impacts our future actions.
The neurological processes underlying the formation or changing of opinions due to social
media exposure have been studied from the perspective of the presence and nature of biased
content, and the way in which others interact with the information (z.B., likes, Kommentare,
retweets, usw.). Prior neuroscience work has specifically studied the effect of social influence
on opinion formation and opinion change within the social media environment, where a net-
work of brain regions including the striatum, orbitofrontaler Kortex, and temporoparietal junction
appear to have a critical role in this decision-making process (Baek et al., 2021; Casado-Aranda
et al., 2020; Cascio et al., 2015; Falk & Scholz, 2018; Falk et al., 2012; Izuma & Adolphs, 2013;
Kappes et al., 2020; Klucharev et al., 2011; Li et al., 2019; Nakao et al., 2016; Sherman et al.,
2016). Speziell, the neural mechanism of opinion change due to social media use has been
shown to integrate brain areas of the valuation, social pain/exclusion, and mentalizing systems
that include the ventro-medial prefrontal cortex, striatum, medial prefrontal cortex, dorsomedial
prefrontal cortex, temporo-parietal junction, posterior cingulate, medial tegmental gyrus, Und
anterior cingulate (Baek et al., 2021; Cascio et al., 2015; Falk et al., 2012; Kappes et al.,
2020). Other work has suggested that the popularity of content (Sherman et al., 2016) und das
valence of the content plays a significant role in swaying opinion on these platforms (Baek et al.,
2021). Due to the opportunity social media affords in rapidly disseminating information through-
out the globe, it also creates an interesting glimpse into the complex human decision-making
process that impacts our everyday lives (Schmälzle et al., 2017). In der Tat, with the intensity and
speed in which information spreads in this media convolved with the global scale, the contex-
tual impact on decisions derived from platforms like these have had demonstrably profound
impacts on society as a whole (Spinney, 2017).
Despite the understanding of the importance of these platforms in forming our decisions, Es
is still unclear how brain networks composed of regions, perhaps those associated with social
media informational processing and influence, interact to produce opinion change. Impor-
tantly, it is also unclear how this process may be unique to brain processes underlying in-
person interaction and free discussion. Network neuroscience provides a variety of tools to
understand the complex network properties of the brain and has proven successful in describ-
ing a variety of behaviors (z.B., Bassett & Spurns, 2017; Betzel & Bassett, 2017). Zum Beispiel,
the rate at which networks within the brain rapidly reconfigure to support cognition has been
found to be highly predictive of a variety of cognitive processes. Dynamic community detec-
tion, a technique used to distill complex connectivity patterns into time-varying labels of com-
munities (d.h., clusters of nodes) has been successful in capturing variability in a variety of
behaviors in fMRI studies primarily, but has also been used for understanding band-specific
Opinion change:
The act of a participant to change
their stance on some aspect of the
referred scenarios, as measured in
the paper through survey probes of
opinions.
Dynamic community detection:
The procedure used to define the
dynamics in community structure
whereby different EEG sensors are
grouped to determine the network
“state” in a given time layer.
Netzwerkneurowissenschaften
871
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
N
e
N
_
A
_
0
0
2
5
5
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network reconfigurations and social media
Network reconfigurations:
A modification in the community
Struktur (z.B., different nodal
affiliations) between at least two
different time layers.
EEG connectivity patterns (z.B., Garcia et al., 2020A). Hier, we have investigated the rapid
fluctuations in network connectivity while individuals are exposed to an interactive social
media platform containing factual and fake content, attempting to simulate the real-world
experience of social media and relate this to changes in opinions after exposure to this con-
tent, as we hypothesize that the flexible dynamics within the brain may be associated with
complex decision-making behind opinion change. Wichtig, we provide a comparison to
in-person discussion that allows us to disentangle the unique neural properties of this process.
Our results provide preliminary evidence of unique neural features marking the cognitive pro-
cesses supporting decision-making prompted by digital stimuli on a social media platform.
ERGEBNISSE
We have investigated the neural correlates of complex decision-making during online social
media and in-person social interactions and assessed opinion change with questionnaires that
asked participants for their opinions on several topical issues. Opinions on these topical issues
were gathered before and after the interaction with a simulated social media platform and after
in-person discussion of the content (Figur 1; see also Richard et al., 2021). EEG was concur-
rently collected (see Supporting Information Figure S1 for electrode montage) during the social
media and in-person interactions and was analyzed to understand the rapid reconfigurations
in EEG-derived brain connectivity matrices during the complex process of information gather-
ing and opinion change and/or formation. Hier, we used dynamic community detection, ein
algorithm that has previously been shown to successfully capture brain network reconfigura-
tions associated with the variability in human behavior across a variety of tasks. We extended
these findings by inspecting the temporal dynamics of node-pair community affiliations and
comparing this metric between individuals that changed their opinions and those that did not
across these social interaction conditions.
Characterizing Individuals by Opinion Change After Social Media and In-Person Interactions
Figure 1A shows the experimental timeline, Wo, after arriving in the laboratory, Fächer
were presented with questionnaires that asked their opinions on three particular real-world
topical issues, nämlich, (A) travel based on social awareness and volunteerism, (B) punishment
after a murder trial, Und (C) decisions to vaccinate from disease before and after interaction
with the social media platform as well as after the in-person discussion segment. EEG was
recorded during these two interactive conditions, das ist, social media interaction and in-
person discussion. These interactive conditions differed in several ways. Erste, during the social
media interaction interval, subjects were seated in front of a monitor and were allowed to freely
scroll through the simulated social media platform and interact (z.B., “like”, “share” posts) for no
mehr als 2 hours. During the in-person discussion segment, Fächer (three to four at a time)
were seated in another room where an experimenter moderated the conversation and asked
subjects to discuss the topics for no more than 20 minutes. Based on changes of the question-
naire answers before and after each interaction, we grouped the intervals into two segments,
those intervals in which subjects changed their opinions (“change,” C) and those that did not
(“no change,” NC), Und, for shorthand, we use the acronyms Cs and NCs for groups that change
and did not change their opinion after social media interaction. For in-person discussion, Wir
similarly use the shorthand Ci and NCi. Figure 1B–1D shows the distribution of responses and
the rate of opinion change across participants (N = 132) following both conditions. Erste, mit
the social media platform, a majority of individuals did not change their opinion from the initial
survey (N = 75); Jedoch, insgesamt 57 individuals changed their opinions, with the most indi-
viduals changing their opinion in the murder trial scenario (N = 39). A small proportion of the
Netzwerkneurowissenschaften
872
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
N
e
N
_
A
_
0
0
2
5
5
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Brain network reconfigurations and social media
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
N
e
N
_
A
_
0
0
2
5
5
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
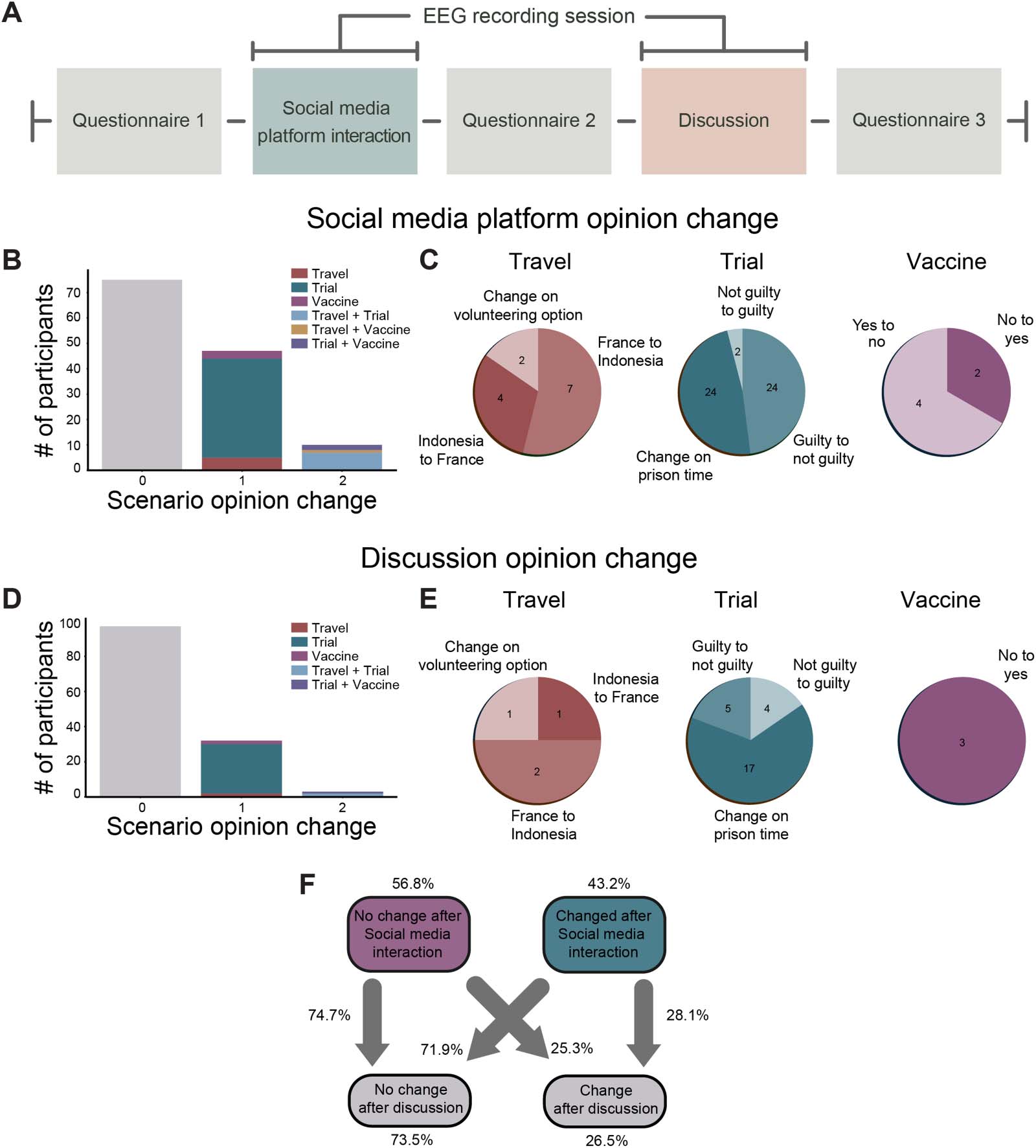
Figur 1. Experimental setup and opinion change quantification. (A) Timeline of the experimental design. The opinion changes of each
subject were assessed through the application of a questionnaire before and after the subject interaction with the social media platform,
in addition to in-person discussion. (B, D) Histogram of opinion change by scenario in social media interaction and in-person discussion,
jeweils; 0, 1, Und 2 indicate the number of scenarios in which individuals change their opinion. Color legend indicates the scenario.
(C, E) Pie charts indicate the direction of opinion change for all of the changes observed after the social media platform and in-person
interactions. (F) Flow chart shows the fraction of participants that changed their opinion from the social media platform (Cs) interaction to
in-person discussion (Ci).
individuals (N = 10) changed their opinions in two scenarios and were most likely to change
their opinion on the travel and murder scenarios (Figure 1B). Figure 1C displays a more granular
visualization of responses for each scenario, and similar to the design of the experiment that
presented equally positive and negative coverage on an issue, there was a large diversity in
opinion changes, validating the well-balanced affectual information within the platform. Für
Beispiel, even with the vaccination scenario, there were some individuals who changed their
opinion toward not vaccinating after the social media interaction. Wichtig, sowie, ist der
fact that the change in prison time in the murder trial contributed to the largest changes,
Netzwerkneurowissenschaften
873
Brain network reconfigurations and social media
mit 24 subjects changing the prison time after the social media interaction and 24 changing
from guilty to not guilty. This was significant for both the social media interaction (χ2(1, N =
132) = 7.75, p = .005) and in-person discussion segments (χ2(1, N = 132) = 48.48, P < .0001).
We observed some similarities and differences in opinion change after in-person discus-
sion. As shown in Figure 1D–1E, the overwhelming majority of individuals (73.5%, N = 97)
did not change their opinion, suggesting that the in-person discussion was less likely to affect
one’s opinion than social media interaction; however, the order of the questionnaires was the
same across all individuals. This limitation does not allow us to disentangle the effect ques-
tionnaire order may have on our effects. Similar to the social media interaction condition,
though, changes in opinion mostly occurred within the murder trial condition, accounting
for 80% of the total changes in opinion after the discussion. Very few individuals changed their
opinion in more than one scenario, accounting for only 8.6% of the total opinion change after
the discussion.
Finally, to characterize the opinion change, overall, we estimated transition probabilities
as presented in Figure 1F. These transition probabilities can give us a glimpse into the indi-
vidual subjects that did and did not change their opinions following the social media inter-
action and in-person interactive conditions. As is shown in Figure 1F, 56.8% of the subjects
did not change their responses after social media interaction, 74.7% of these individuals also
kept their responses after in-person discussion. From the 43.2% of the subjects that changed
their responses after the social media interaction, 28.1% changed their responses after in-
person discussion as well. Importantly, of all the participants, 73.5% did not change their
opinion after in-person discussion. Due to the distribution of those that did not change their
opinion and those that did, we next compared groups of subjects who did and did not
change their opinions. Critically, if an individual changed their opinion in any of the sce-
narios, they were included in the “change” (C) group, and only those who did not change
their opinion in any scenario were included in the “no change” group (NC) for all subse-
quent analyses.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
wPLI:
The weighted phase locking index
(wPLI), is a measure of statistical
dependency in the phase between
pairs of signals recorded by EEG
sensors.
Modularity:
A measure that aims to estimate how
far a given community structure is
from the case obtained by chance
(i.e., random linking of the network
nodes).
Louvain algorithm:
A greedy clustering algorithm used to
produce the dynamic community
structures used in the analysis;
briefly, it works by grouping the
networks nodes in communities such
that it optimizes the modularity.
Nodal Flexibility Distinguishes Individuals Who Changed Their Opinion Following Social
Media Interaction
We hypothesized that complex decision-making and information processing requires the
reconfiguration of underlying brain networks. To test this hypothesis, we applied a dynamic
community detection analysis to the EEG data and probed how network reconfigurations are
associated with opinion change by directly comparing the intervals in which subjects did and
did not change their opinion (Figure 2). This was accomplished in several steps. First, the
dynamic community structure requires an estimate of the underlying statistical dependency
between nodes. Here, we estimated this statistical dependence, or functional connectivity
of the EEG, by using the pair-wise weighted phase lag index (wPLI) separately for commonly
studied EEG oscillations (i.e., delta (1–3 Hz), theta (3–7 Hz), alpha (8–13 Hz), beta (21–30),
and gamma (25–40) bands) in nonoverlapping 10-s time windows. This data-driven approach
to functional connectivity exploits phase-based relationships within the data, yielding connec-
tivity matrices that are reliable and less susceptible to some expected artifacts without requir-
ing parameterization (Hardmeier et al., 2014). Once calculated, the wPLI matrices were used
to determine the community structure by modularity maximization using a Louvain algorithm
(see Methods). This distilled the connectivity time evolving matrices into an average of 315
(SD = 102) time windows of community labels that represent the band-specific community
affiliations of EEG sensors across time.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
874
Brain network reconfigurations and social media
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
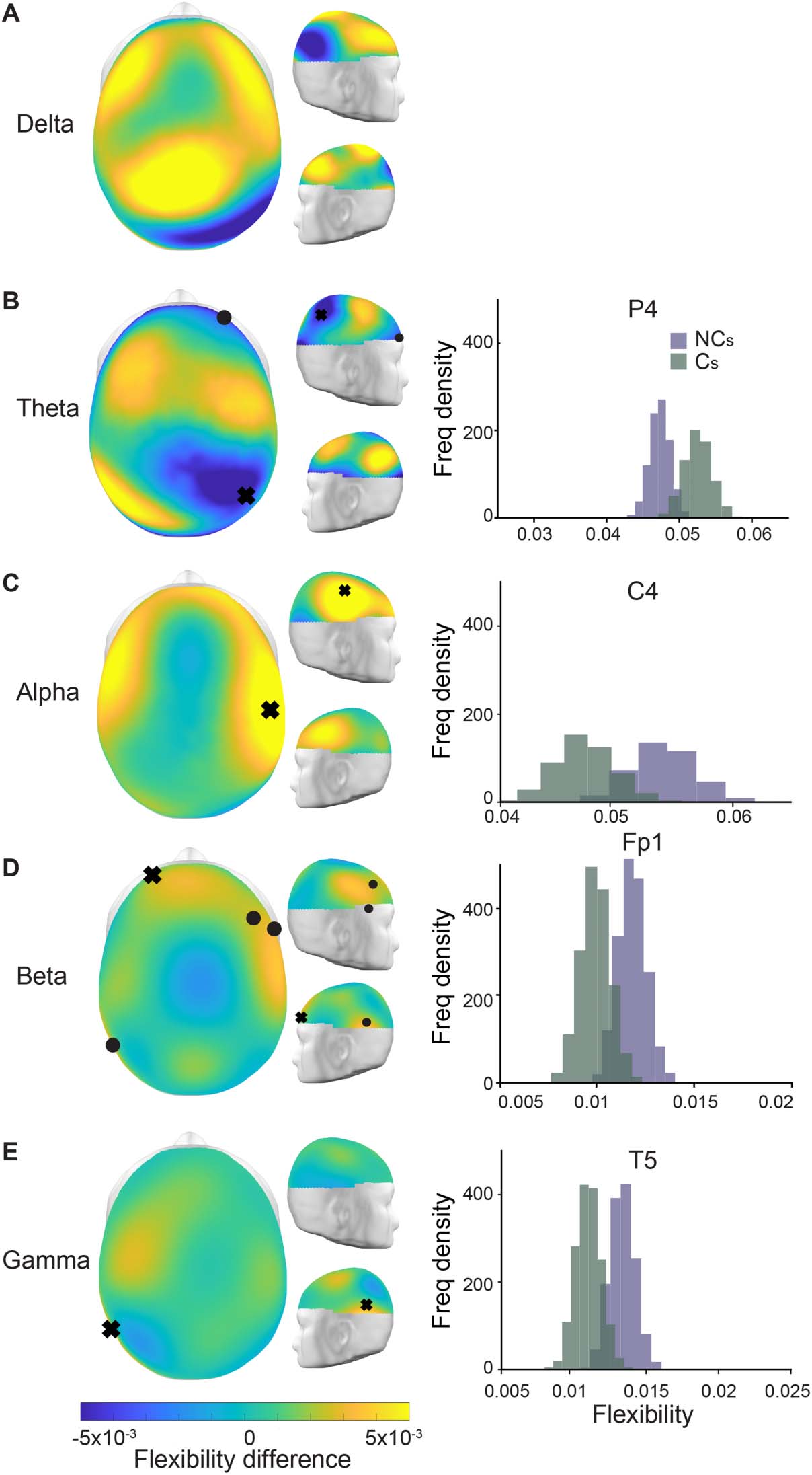
Figure 2. Flexibility differences between the intervals in which individuals did and did not change
their opinion. (A–E) Topographic plots of each frequency band showing the difference of the mean
flexibility between the two groups, such that positive values (in yellow) indicate an increased flex-
ibility for those that did not change their opinions. The black tokens (orbs, x’s) indicate sensors with
statistically significant differences in flexibility between the two groups as found via a bootstrap
procedure (see Methods). On the right, we show representative bootstrap distributions of the mean
flexibility of the sensors marked by an x for the two intervals in purple (NC, no change) and green
(C, change).
Network Neuroscience
875
Brain network reconfigurations and social media
Flexibility:
The number of instances in which a
node changes community affiliation,
normalized by the total possible
number of changes that could occur
across all the time layers.
From these affiliations (i.e., distilled connectivity matrices), we estimated sensor (node)
flexibility, which is a measure of how much each node changes its affiliation across time
between the opinion change groups. Since the data is not balanced between the groups,
we employed a bootstrap procedure to estimate the distributions of mean flexibility for each
of the EEG sensors for both groups (“change” vs. “no change”, see Methods) and subsequently
compared the node flexibility when individuals changed their opinion (or did not). We
observed that those intervals in which individuals did not change their opinion showed
significant increases in node flexibility in the alpha, beta, and gamma bands and a decrease
in flexibility in the theta band. Figure 2 shows the node flexibility differences; yellow (and
blue) shades indicate an increased flexibility on the group without change (and with change).
An increased node flexibility in those with no change in opinions was observed in the higher
frequency bands, with beta-band flexibility showing significant differences (bootstrap analysis,
p < 0.05) for sensors F8, F4, Fp1 and T5, T5 in the gamma band, and C4 in the alpha band. In
contrast, the theta band presents a statistically significant decrease in node flexibility in those
that did not change their opinion compared to those who did, sensors P4 and Fp2, differing
from the other frequency bands. These results indicate that the dynamics of the
synchronization-desynchronization processes, as measured by the wPLI, plays an important
role in the underlying mechanism of opinion change while interacting with social media.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Allegiance:
The ratio of total time in which two
nodes of a network share the same
community.
Intermittence:
A metric that estimates how
frequently the two nodes change
their affiliation from the same
community to different communities
and vice versa.
Assessing Dynamic Changes in Community Structure
We found that the node flexibility is informative as a neural marker of opinion change; how-
ever, it does not provide much information of the dynamic changes in the community structure
to further understand the underlying network reconfigurations leading to opinion change. For
example, one could ask how the links of the flexible nodes evolve with time and which other
nodes couple and decouple with them more often during the task. In this regard, allegiance is
a commonly used metric that captures the fraction of time two nodes share the same commu-
nity affiliation, 0 for a pair of nodes that never share a community and 1 for nodes that are
always in the same community. We estimated node allegiances for individuals with and with-
out a change in opinion and found that they do not differentiate the two groups (for further
details, see Supporting Information and Figure S2). However, it is also unclear from allegiances
alone whether more fine-grained temporal dynamics of network reconfigurations might differ-
entiate these groups. To more finely understand the temporal evolution of node-pair affiliation
change, we computed a new metric called intermittence.
Like allegiance, intermittence is a measure of the interaction between two nodes of the
network; however, while allegiance captures the fraction of time two nodes belong to the
same community, intermittence tracks how frequently the two nodes change their affiliation
from the same to different and vice versa. In other words, intermittence differentiates two
nodes’ affiliation changes that occur in rapid bursts from affiliation changes that occur in lon-
ger term after more static community affiliation similarity. Together, we may inspect allegiance
as the likelihood for two nodes to be in the same community, and intermittence can inform us
of the temporal nature of this relationship.
Intermittence Differentiates Changes in Opinion
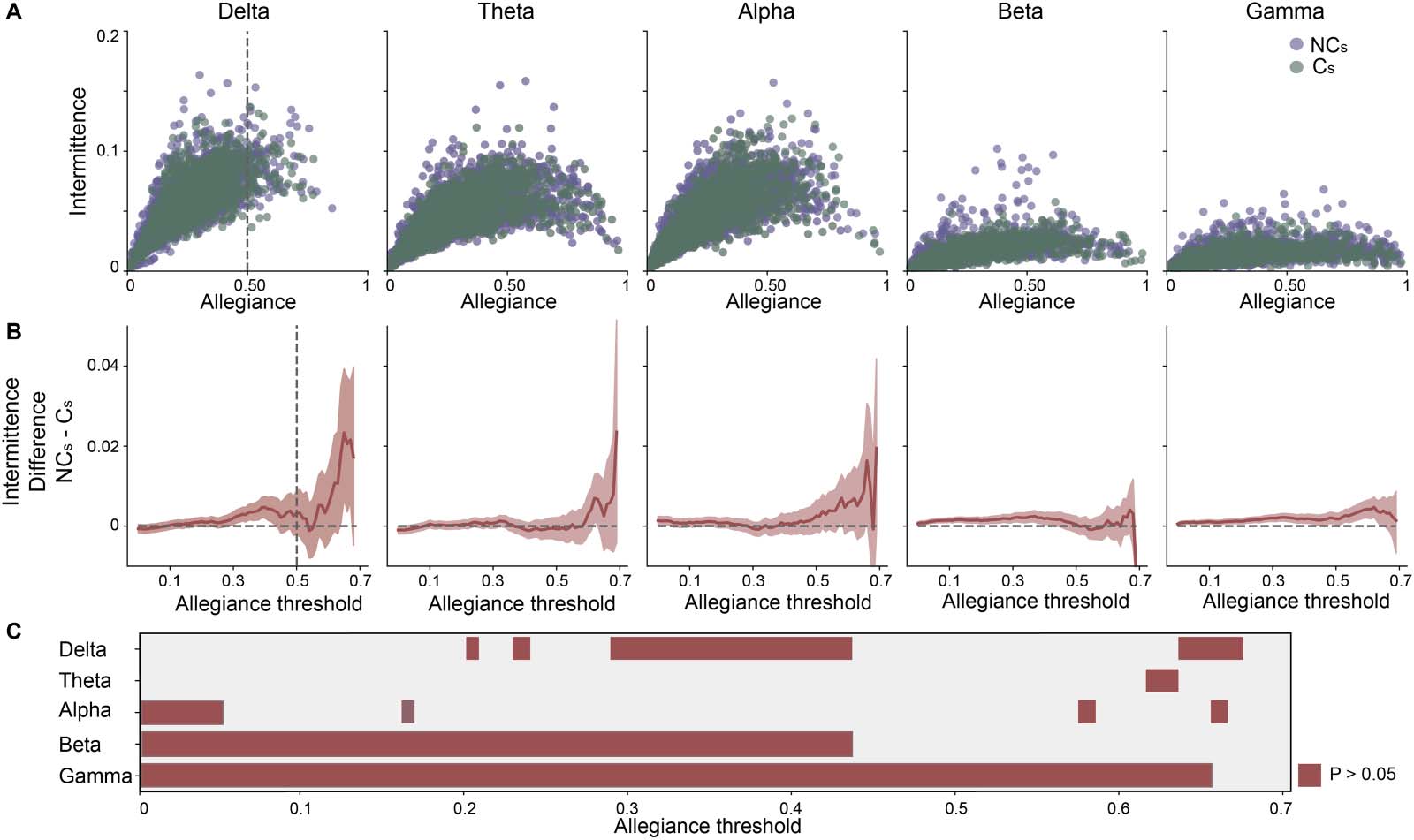
In exploring the intermittence metric, we first directly compare the allegiance and intermit-
tence metrics for both groups (Figure 3A). We observe for lower frequency bands (e.g., delta)
that intermittence is more variable, spanning a wider range of values than higher frequency
bands while the opposite is true for allegiance. Specifically, for the delta band, values of alle-
giance larger than 0.6 are less frequent than observed for the other frequency bands, and
Network Neuroscience
876
Brain network reconfigurations and social media
Figure 3. Comparing intermittence and allegiance in opinion change. (A) Scatter plots of the relationship between intermittence and alle-
giance for each frequency band of interest, where purple (or green) indicates channels of individuals who did not (and did) change their
opinion (C vs. NC). Dashed vertical lines indicate the middle value of allegiance, to visually anchor the plots. (B) Bootstrapped difference
(no change–change; NC-C ) plots of intermittence of the intervals for different threshold levels of allegiance for each frequency band of interest.
Shaded region is 95% confidence interval and the vertical dashed line indicates the midpoint of allegiance to visually anchor the midpoint. (C)
Allegiance thresholds that survive statistical comparison of opinion change for each frequency band.
values of intermittence above 0.1 are more frequent than for the beta and gamma bands. On
the other hand, inspecting higher frequency oscillations, we see intermittence is rarely above
0.05, but allegiance spans the entire range of possible values. This suggests that there is a
higher propensity for more static network reconfigurations at higher frequencies than lower
frequencies (e.g., compare Figure 3A gamma and delta). Importantly, with intermittence esti-
mation, simply by its calculation, allegiance imposes an inherent restriction on its range of
possible values. The maximum value of intermittence for a given pair of nodes is limited by
the value of allegiance between those nodes, as the reader should understand that there can-
not be more dynamic changes between nodes if they are rarely ever in the same community.
Thus, given our observation that higher frequency bands (beta, gamma) had higher allegiance
(that could allow for higher intermittence) in addition to the observed lower average intermit-
tence, the findings observed in higher frequency bands are even more striking, suggesting
higher frequencies display very static network dynamics across groups.
In Figure 3B, we show the differences between the means in intermittence of the two
groups. Critically, mean bootstrap distributions were calculated using only those points with
associated allegiance values higher than the allegiance threshold indicated on the x-axis. The
shaded area in Figure 3B represents a 95% confidence interval and was obtained by a boot-
strap procedure with 10,000 samples (see Methods). To summarize the statistical comparison
between those intervals with or without opinion change at each of the frequency bands,
Figure 3C displays those allegiance thresholds that display the statistical difference (p < 0.05).
Comparison of the mean intermittence between the two groups shows that the intermit-
tence metric can delineate between those in which there was or was not a change in opinion
in each frequency band, but to a highly variable extent. For example, our results show that
Network Neuroscience
877
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
within the delta band, we observed statistically significant differences for the allegiance thresh-
old range between 0.29 and 0.43 and a few other allegiance values accounting for more than
14% of the possible allegiance range values. For the theta band, there were minimal differ-
ences between groups observed, accounting for only 2.8% of the total allegiance range.
Within the alpha band, we observed, again, minimal differences between the groups account-
ing for less than 5% of allegiance thresholds; importantly, they were observed mostly at the
lowest allegiance thresholds. The most robust differences between opinion change groups
were observed within the beta and gamma bands. For the beta band, we observed significant
differences between the groups in approximately 43% of the allegiance threshold range. The
lowest p values we observed between those without a change in opinion (M = 0.018 a.u.) and
those with a change in opinion (M = 0.017 a.u.) was for the allegiance threshold of 0.12
(p = 2.7 × 10−5). The largest range of allegiance values with significant differences between
the two groups were observed in the gamma band, accounting for nearly 65% of the entire
allegiance range. The lowest p value was observed at an allegiance threshold of 0.29 for no
opinion change (M = 0.018 a.u.) and opinion change (M = 0.016 a.u., p = 3.6 × 10−6) inter-
vals. Thus, it appears that intermittence successfully delineates those individuals with and
without a change in opinion while interacting with social media, but does so in a frequency
band–specific manner where the beta and gamma bands show the most robust differences as
indicated by a wide range of allegiance thresholds for which the two groups have a significant
difference in intermittence. In other words, intermittence can be used to characterize opinion
change in band-specific oscillatory schemes, but it is still unclear whether this is a general
opinion change phenomenon or if this may be specific to the context in which information
is received (i.e., social media platform). Thus, we next explored how these findings could differ
in a different context, specifically during in-person discussion.
Social Media Interaction and In-Person Discussion Differences
To determine the specificity of our findings to the social interaction context (e.g., social media
vs. in-person discussion), we sought to determine if network dynamics between these opinion
change groups have similar structure during social media and in-person interactions. The
in-person discussion was completed after the social media interaction and was conducted
by an experimenter who acted as a moderator and prompted individuals to discuss the
topics that were probed by the surveys (see Methods). To estimate the difference in neural
dynamics between these two social interaction contexts, we calculated the coefficient of
variation (CoV) of the time series of the wPLI during social media interaction and in-person
discussion. This procedure is a computationally inexpensive and complementary approach
to capture dynamic reconfiguration of the synchronous patterns estimated from the EEG
recordings that we expect to be similarly sensitive to the intermittence metric but on the
node-level, like flexibility.
Similar to the EEG measurements during the social media interaction, the statistical depen-
dencies between nodes were estimated with the wPLI metric, and then, in a pair-wise fashion,
the temporal CoV was calculated for each of the node pairs. Finally, to aggregate the data for
each subject, the mean temporal CoV for each pair was estimated, and for each subject, we
calculated the mean across all the nodes. This procedure results in a single mean temporal
CoV for each session, and finally the group distribution and statistical comparisons were com-
pleted with a bootstrap procedure for each social context and opinion change (or no change)
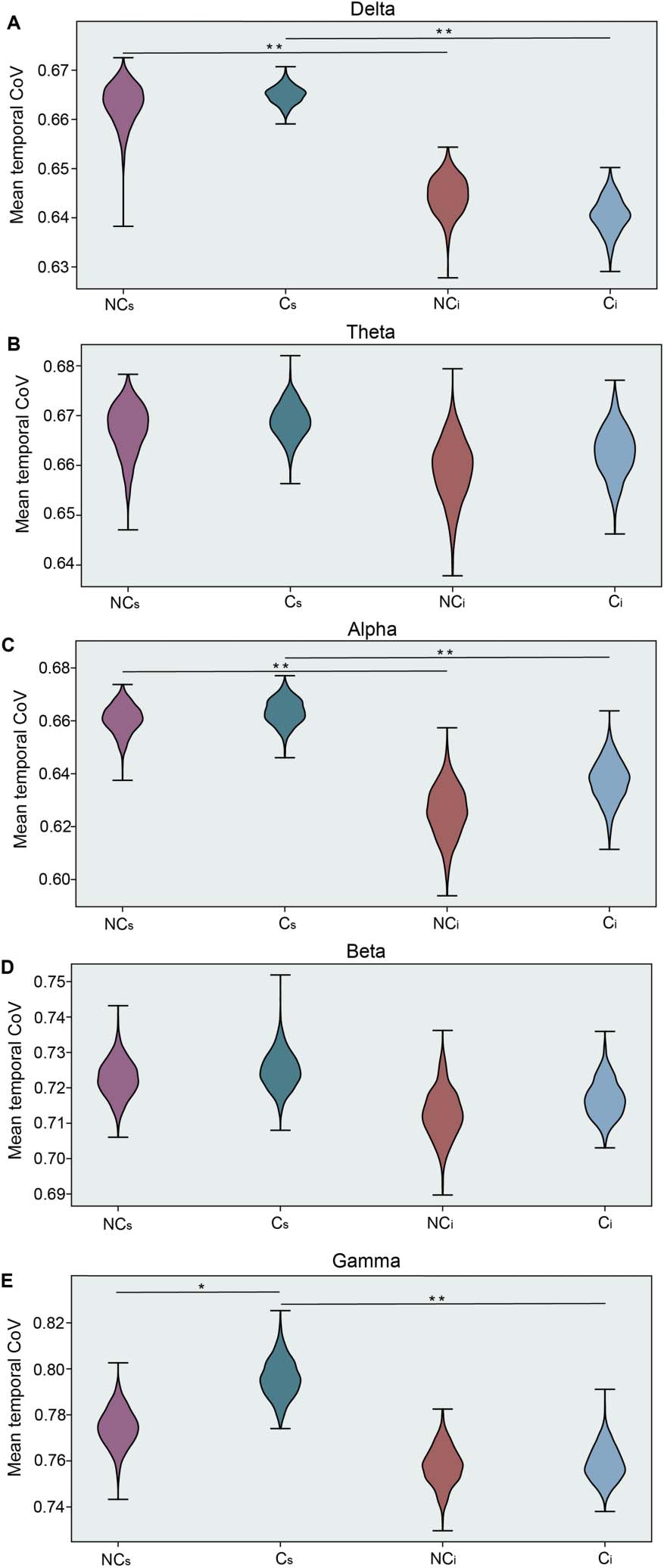
and are summarized in Figure 4. We observed clear group differences in the gamma band (p =
0.031) during the social media interaction, in accordance with the results obtained from the
intermittence analysis. For those subjects who were more likely to change their opinion, we
Network Neuroscience
878
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Social media and in-person discussion comparison. Mean wPLI temporal coefficient of
Figure 4.
variation (CoV) for each frequency band, including delta (A), theta (B), alpha (C), beta (D), and
gamma (E). Each panel presents results for the intervals with no opinion change (NC: purple-red)
and with a change in opinion (C: blue-green) interacting with the social media platform (Cs, NCs)
and during in-person discussion (Ci, NCi). Statistical differences were estimated through a bootstrap
procedure (1,000 replicas with N = 30 each) with significant results denoted with asterisks and are
shown both within group and across social interaction contexts (*p < 0.05, **p < 0.005).
Network Neuroscience
879
Brain network reconfigurations and social media
also observed a difference in context, where we observed more variability (temporal CoV) in
the social media interaction than in the in-person discussion (p < 0.05). Moreover, within the
delta and alpha bands, we observed this context effect, too, where temporal CoV of connec-
tivity in the social media intervals was significantly higher than in the in-person interaction
contexts. Interestingly and divergent from the previous findings, we observed no significant
differences in the beta and theta bands.
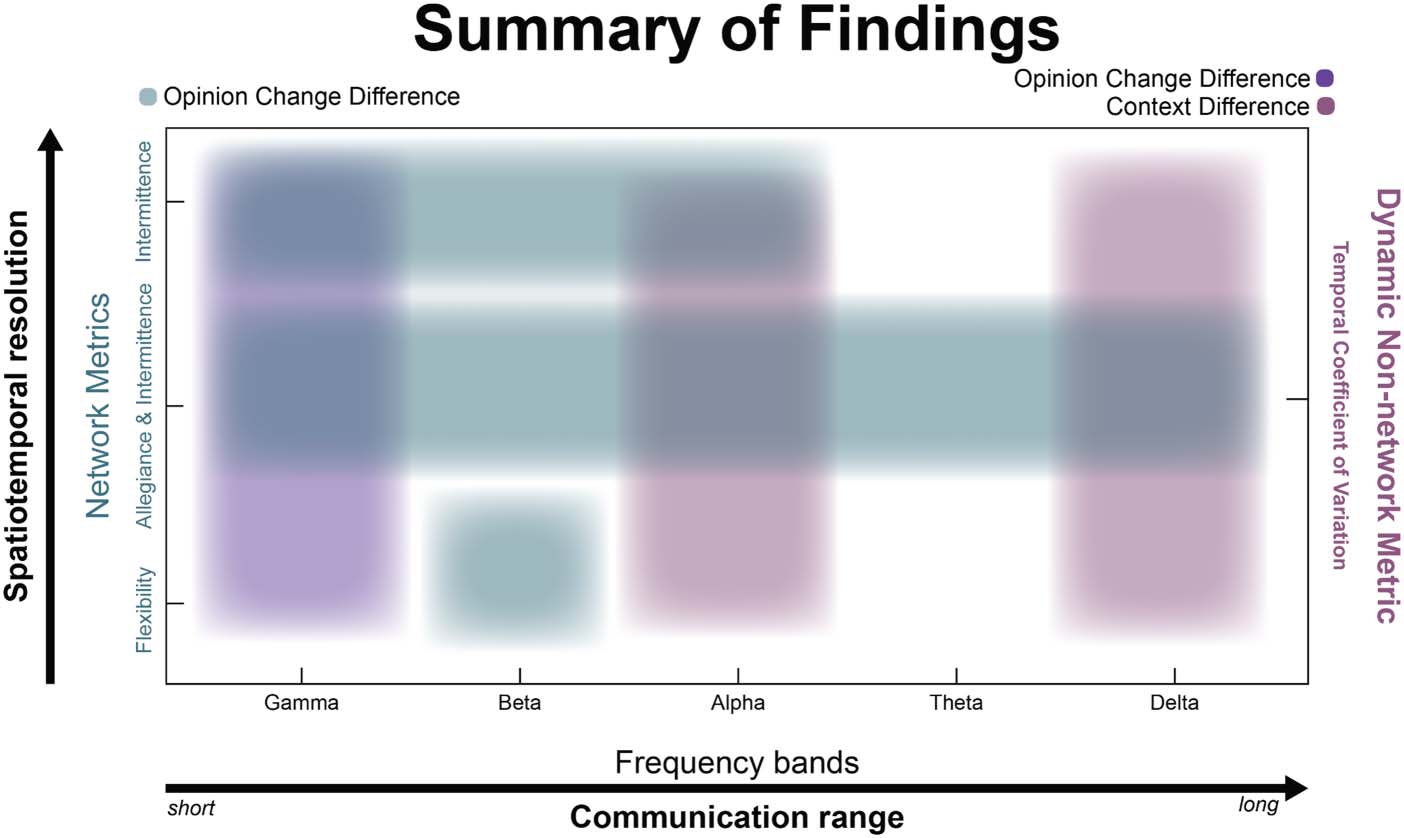
Summary of Findings
In aggregate, we have observed varying levels of sensitivity in the estimated metrics, attempt-
ing to describe the neural dynamics underlying social context and opinion change influences
within the brain. Due to the complex nature of the findings, the differences in context and the
cognitive processes driving our findings, we aggregate and visualize our results in Figure 5. We
present our findings along two continuous axes and one static axis. First, we showed results for
three different network metrics: flexibility, or the propensity of a node to change its affiliation
across time; allegiance, or the pair-wise likelihood that two nodes are in the same community;
and intermittence, or the rate at which pair-wise affiliation change (1 = constant and consistent
change, 0 = no change). These three metrics are ordered as a function of how spatiotemporally
resolved they are. Flexibility, a node-wise metric, does not differentiate between pair-wise sim-
ilarity and/or differences in community affiliation. Allegiance, on the other hand, is a pair-wise
measurement of “similarity” but does not take into account how node pairs change their affil-
iation across time. So, these three metrics are on an axis representing low spatiotemporal
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Summarizing the dynamics to opinion change. In aggregate, we summarize the findings
along three axes: (a) one in which the network metrics describe the spatiotemporal resolution of the
network dynamics ( y-axis, left); (b) an axis meant to describe the effects within the estimated fre-
quency bands, interpreted as short- to long-range communication within the brain (x-axis, bottom);
and finally, (c) a proxy for flexibility that estimates the overall dynamics across the scalp as mea-
sured with CoV ( y-axis, right). Network metrics on the left y-axis are sorted by increasing ability to
resolve the nature of the spatial specificity and temporal dynamics. On the x-axis, frequencies are
sorted in descending order in terms of communication range where gamma is sensitive to changes
in proximal neural ensembles and delta to long-range communication. Purple hues should be asso-
ciated with the right axis and teal hues are meant to be associated with the axis on the left.
Network Neuroscience
880
Brain network reconfigurations and social media
resolution at the origin and high temporal and spatial resolution at the top portion of the graph.
As our results indicate, these metrics must be interpreted within the context of the specificity in
frequency band, where each band has been associated with a variety of cognitive phenomena
but also represents communication across long distances (e.g., delta) or short (e.g., gamma).
For the least spatially resolved metric, flexibility, the differences between the C and NC groups
was most robust within the beta band, a frequency band often implicated in motor behavior
(e.g., Stancák & Pfurtscheller, 1996), but also in signaling the “status quo” (Engel & Fries,
2010). While allegiance, by itself, did not reveal significant pair-wise changes (see Supporting
Information Figure S2), considering intermittence at varying levels of allegiance revealed the
most robust changes across the groups at each frequency band of interest. Interestingly, inter-
mittence, by itself, was most robust in the highest frequency bands. Together, these results
indicate a specificity to these metrics but a robustness of the intermittence metric, suggesting
a unique importance to the nature in which networks reconfigure in a complex decision-
making task, like opinion formation and/or change.
As a final summary, we also inspected a proxy for these dynamic metrics, specifically
inspecting the mean CoV across time of the estimated functional connectivity (wPLI). Here,
a high CoV is associated with a fluctuating connectivity and may be associated with how often
a node changes its affiliation across time. Indeed, we see that this metric can capture the var-
iance associated with changing one’s mind after the social media interaction, but only within
the gamma band. Interestingly, this metric also captured variability in context (social media vs.
in-person discussion) within the alpha and delta bands, suggesting perhaps a different mech-
anism by which context shapes the way we encode information and make decisions.
DISCUSSION
Decision-making is a complex internal process by which information is consumed and an
action is executed, requiring the support of many interacting brain networks composed of a
variety of functionally diverse regions within the brain (for review, see Fellows, 2004; Rilling &
Sanfey, 2011; Wallace & Hofmann, 2021). The present study investigated the impact of infor-
mational context and its type on decision processes, specifically how social media and in-
person discussion influences one’s malleability to change one’s mind on “highly shared”
content in online platforms. Our findings have shown a large portion of individuals (e.g.,
five out of six in vaccination hesitancy) were susceptible to content displayed to them on
a simulated social media platform while the same individuals were not susceptible to free-
form and in-person discussions on the same topics. Using dynamic community detection
(see for review Garcia et al., 2018) and estimating the temporal dynamics of network recon-
figurations that occurred across several frequency bands, we found that the flexibility of spe-
cific sensors across the scalp could discriminate between those individuals who were and
were not influenced by content presented in a simulated social media platform (Figure 2).
Importantly, those with no change in their opinions showed higher flexibility in sensors
located within the frontal and posterior regions for the higher frequency bands (i.e., alpha,
beta, gamma), whereas the opposite effect is observed in theta band where higher flexibility
was observed for those in which there was a change in opinion within prefrontal and posterior
sensors (Figure 2). Our results extended the flexibility findings to show that the vigor in which
the network changes occurred was driving this effect with a new metric we call intermittence.
Interestingly, intermittence within the higher frequency bands was more robustly diagnostic for
different opinionators than lower frequency bands (Figure 3). Moreover, while our results did
not show substantial change in opinion after in-person discussion (only 35 people changed
Network Neuroscience
881
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
their mind after in-person discussion), we also did not find any of the observed brain network
reconfiguration changes during the in-person interaction (Figure 4).
Network Flexibility As a Marker of Opinion Change
The so-called flexibility metric has been used to describe the rate of motor learning (Bassett
et al., 2011; Bassett et al., 2013; Gerraty et al., 2018; Li et al., 2019; Reddy et al., 2018), has
been associated with multitasking (Alavash et al., 2015; Shafiei et al., 2020; Thomas Yeo et al.,
2015), pattern recognition (Telesford et al., 2016), language comprehension (Chai et al., 2016),
thought rumination (Han et al., 2020; Lydon-Staley et al., 2019), adaptations to new stimuli or
stress (Betzel & Bassett, 2017; Paban et al., 2019), and working memory (Braun et al., 2015;
Lauharatanahirun et al., 2020). Our findings support the increasing evidence suggesting the
importance of the rapid reconfigurations of brain networks in cognition, and specifically,
here, decision-making as it pertains to opinion change and/or formation (for review, see
Shine & Poldrack, 2018). We used flexibility as the probability a particular node (i.e., sensor)
changes its affiliation across time. Previous studies show that this type of network-defined
flexibility in frontal brain regions is associated with faster motor learning (Bassett et al.,
2011), psychological resilience (Paban et al., 2019), chronic behavior change in addiction
(Cooper et al., 2019), enhanced working memory performance (Braun et al., 2015)—which
is also moderated by sleep (see Lauharatanahirun et al., 2020)—and even improved adaptive
problem-solving skills (Barbey, 2018). Given the diversity in these behavioral findings and
our extension to even social media influence, it is reasonable to attempt to understand how
this metric may be highly sensitive to a variety of complex cognitive phenomena.
Indeed, the neuroscientific journey that has led to the importance of flexibility in neural
behavior may be understood from several different perspectives, and it is currently thought
to be the foundation to the human brain’s unique ability to rapidly adapt to task demands.
First, it should be noted that we have estimated network-based flexibility via a mathematically
defined dynamic network approach (see Methods), and on its surface it should not be con-
fused with concepts such as cognitive flexibility (Uddin, 2020) and neural flexibility (Yin &
Kaiser, 2021), but it can be complementary to both (Mattar et al., 2016). Cognitive flexibility
refers to the executive functions that allow an individual to rapidly transition from task to task
and has been found to be associated with improved performance in a variety of tasks and also
is reduced in certain pathologies (Hanes et al., 1995). In contrast, neural flexibility, while often
used in relation to cognitive flexibility, refers to the brain’s ability to rapidly shift across tasks
and be recruited for a variety of activities (Uddin, 2020). Dynamics in the neural signal have
previously been discarded as noise, but are now accepted as describing valuable variability in
human behavior and even psychopathology (Uddin, 2020). Our findings not only add to this
growing literature and support the network science approaches that can successfully capture
this variability, but, specifically, we have used dynamic community detection, an extension of
the network science approach of functional modularity, that is a theoretically derived (but indi-
vidualistic) technique to probe dynamic network changes via a distillation of dynamic connec-
tivity matrices (Garcia et al., 2018). Our research adds to the general nature of this technique
to capture the broadly cognitive, distributed, and adaptive nature of the brain, the primary
criteria for flexible brain regions and networks (Yin & Kaiser, 2021). Here, we speculate that
the process in which we are rigid in our opinions shows faster network reorganizations due to
an effort to accommodate conflicting information with previously held beliefs. Another possi-
ble explanation can be different social media browsing strategies that led them to be exposed
to more content for a smaller amount of time, being less prone to reevaluate previous beliefs,
while increasing the sensory input. Future studies may be designed to evaluate these options.
Network Neuroscience
882
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
Intermittent and Persistent Network Reconfigurations Are Diagnostic of Opinion Change
Our results expand on the pervasiveness of the flexibility results. We show that, at least for
complex, high-level decision-making, not only the rapidly evolving network reconfigurations
(as measured by flexibility) are important but also the fast dynamics of intermittent linking
(same nodes linking in an on-off fashion, aka intermittence) between modules are more diag-
nostic of social media influence on one’s opinion. Dynamic community detection has proved
to be an effective tool to explore temporal patterns in systems represented by complex net-
works, and a key aspect for this framework is the determination of the temporal resolution
of the dynamic communities (Telesford et al., 2016). A systematic way to determine a temporal
resolution that leads to behaviorally relevant network description of the brain can be achieved
by modularity maximization (Newman, 2006). Based on the dynamic communities obtained
through the modularity maximization algorithm, we explore the temporal patterns of network
links and how community allegiances of the network nodes change across time.
A critical feature of our findings is the fact that the temporal profiles of the estimated com-
munity structure is more diagnostic (e.g., intermittence) than simply the fact that dynamic net-
work reconfigurations occur (Figure 3). The temporal profile of interactions has a fundamental
importance on a wide range of phenomena such as the dynamics of neuron populations that
lead to seizures (Jirsa et al., 2014; Rungratsameetaweemana et al., 2021), weather models and
turbulent systems such as the Lorenz attractor (Ruelle, 1976) and the many synchronization
phenomena in which many units share the same temporal profile (Pikovsky et al., 2001;
Strogatz, 2000). From the point of view of dynamical systems, processes of opinion spreading
have been extensively studied using models such as the Voter (Holley & Liggett, 1975) and
majority rule models (Krapivsky & Redner, 2003), suggesting a complex interactive scheme
that gives rise to opinion formation and change. Interestingly, recent findings suggest that infor-
mation sharing and spreading can occur at different time scales in a consensus formation
model depending on the network connectivity and confidence of the agent involved (Ding
et al., 2019). With our approach, we explore this opinion change phenomenon at an individ-
ual level by using the complex networks framework to identify connectivity patterns of EEG
data that are diagnostic to an opinion change process during a social media interaction. In
aggregate, these findings coupled with our current results, suggest similar operations at both
the neural level and population level. Recent findings suggest the brain (as well as other com-
plex systems) operate outside of the boundaries of a particular spatial scale (Cocchi et al.,
2017). Perhaps, our results are the consequence of information spread, whether within a single
brain or across human interactions and are suggestive of a scale-free phenomenon (Mahmoodi
et al., 2017). Indeed, there are many complex systems that express this scale-free behavior;
however, it should be noted that recent findings have even shown that this universal principle
is flexible (Bansal et al., 2021).
Network Reconfigurations and Oscillatory Specificity Suggest a Complex Operation Within the Brain in
Opinion Change
Despite the increasing efforts to understand the neural processes that underlie deliberation and
decision-making, much on this subject remains unclear; however, important findings from the
literature in EEG oscillations, evidence accumulation, valuation, and identity may play critical
roles in understanding our results. First, we have shown that intermittence effects are more
robust at higher frequencies than lower frequencies in the observable EEG oscillatory scheme
(e.g., gamma vs. theta). We also show that the delta band is most diagnostic for social inter-
action context. Oscillations emanating from the brain, as measured with EEG, are a conse-
quence of short- and long-range connections within the brain that interact to give rise to
Network Neuroscience
883
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
cognitive capabilities (Buzsáki, 2006). Importantly, the slower oscillations mostly represent the
coordination of distal regions within cortex and sometimes even modulate higher frequency
oscillations within the brain (e.g., Bragin et al., 1995; Buzsáki & Wang, 2012; Canolty et al.,
2006; Chrobak & Buzsáki, 1998; Leopold et al., 2003; Schroeder & Lakatos, 2009). In other
words, oscillatory activity and the associated cognitive functions rely on the global coordina-
tion of local processes (Cavanagh & Frank, 2014; Rungratsameetaweemana et al., 2018).
Within this context, it would suggest that our results could be a consequence of both, where
the most critical “intermittence” effects were observed within the delta and gamma bands,
“flexibility” effects were most critical in the beta band, and social interaction context was most
observed within the delta band.
This broad coordination of neural communication in the brain gives rise to specific cogni-
tive functions, and our results could reflect several different operations at play. First, our find-
ings show the most significant results in flexibility within the beta band. The beta band is often
associated with motor behavior (Khanna & Carmena, 2015), but has also been proposed to
carry a more prominent role in maintaining motor or cognitive states (Engel & Fries, 2010);
interestingly, beta-band dynamics have even been associated with the accumulation of evi-
dence in the sensorimotor network in a vibrotactile decision task (Haegens et al., 2011). More-
over, fMRI and transcranial magnetic stimulation (TMS) studies have extensively implicated
the so-called value system in decision-making, a system that is engaged when weighing the
potential benefits of a particular decision route. Critical components of this system are thought
to include the ventromedial prefrontal cortex and ventral striatum. These regions, within social
contexts, have been linked to susceptibility to social influence from peers (for a comprehen-
sive review, see Falk & Scholz, 2018). More broadly in EEG, several frequency bands have
been implicated in decision-making (e.g., Nakao et al., 2019), but often show some specificity
in frontal and parietal regions (e.g., Golnar-Nik et al., 2019). Interestingly, a recent study
inspecting long-range temporal correlations in EEG recordings has shown a relationship
between theta to alpha bands and the abstract concept of self-identity and identity confusion
(Sugimura et al., 2021). Due to the highly complex decision, speculation, and potential action,
our results most likely indicate a complex coordination and reconfiguration of networks within
the brain, across several frequency bands, and reflect coordination of these processes includ-
ing evidence accumulation, valuation, and even internal reflection on identity. Future research
is needed to disentangle these processes and influences on decision-making context,
especially within the social media and in-person social contexts studied here.
General Properties of Network Reconfigurations Within the Brain
A somewhat unintuitive result is the finding that those individuals with more rapid network
reconfigurations are associated with a lack of change in opinion. We previously speculated
that this may be due to an effort to accommodate conflicting information with previously held
beliefs or mark different social media browsing strategies that led them to be exposed to more
content for a smaller amount of time. We hope that future research can differentiate between
these potential mechanisms, but when considering our findings within the ubiquitous nature of
these network reconfigurations and associations with behavior, we believe it may be a more
generalized process than previously understood. For example, let us consider a recent theory
of general intelligence based on how we navigate spatially through the world (Hawkins et al.,
2019). This theory is consistent with several principles that include generalized machinery of
the brain (Mountcastle, 1978) to navigate throughout whatever real (or abstract) space, prepare
predictions (Rao & Ballard, 1999), and compare to some reference frame built from previous
experiences (Lewis et al., 2019). It could be the case that metrics derived to estimate the
Network Neuroscience
884
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
dynamic reconfigurations within the brain are targeting this rapid navigation through possible
“observational” (sensorial) interpretations and possible actions, a foundational element to not
only critically think about a topic but also fundamental to general intelligence. Given that, we
hypothesize that those that are highly flexible are also navigating through this abstract sensory,
perception, decision, and/or action space more rigorously than those that are not.
Methodological Considerations and Future Directions
Within this manuscript, we prioritized the ecological validity of the experimentation rather
than distinct cognitive constructs. While the latter presents a very interesting avenue of scien-
tific pursuit, introducing controlled conditions or other contrived experimental manipulations
could potentially modify behavior in unexpected ways. Indeed, often with ecologically valid
research, there is a limitation in understanding all elements that have contributed to our find-
ings. For example, each subject was able to navigate freely through the social media content,
and there was no control on how much time was spent with the content, so variable input may
be a potential confound; however, traditional laboratory-based experiments are not only
defined by their highly precise neural findings, they could also shape the responses in
unnaturalistic ways. Similar to the trends in social (and general) neuroscience studies
(Osborne-Crowley, 2020), we believe that our results should be complimented by more
laboratory-specific effects. In other words, our results do not take into account any
content-specific processes or differences on individual social media interaction, nor do they
consider the controversial and emotional aspects of the different scenarios presented for the
subjects.
Within this experiment and the design that prioritized ecological validity, there are also
several considerations in the experimental measurements (e.g., EEG) and the specific type
of opinion change that was observed of note for future experimentation. For example, the type
of EEG connectivity that was deployed in this analysis prioritized the phase-based statistical
dependencies (for an additional related results, see Figure S5 in the Supporting Information)
within a priori and somewhat narrowly defined bands of interest, following traditional tech-
niques and to compare to the wide literature in oscillatory action as measured with EEG.
Moreover, as is common with EEG analyses, there is a potential that multiple sources
may contribute to the effects, as the spatial information from EEG is known to suffer from vol-
ume conduction. The high temporal resolution of EEG, however, creates an opportunity to
extend dynamic network measurements to more rapidly evolving network reconfigurations
(as compared with fMRI) and in a less constrained manner (e.g., at a desk, in a group discussion
rather than scanner); the inherent limitations in the spatial resolution and the potential contri-
butions to the signal should be considered in future studies. While the combination of flexibil-
ity, intermittence, and CoV analyses converge to a clear importance of the dynamic network
reconfigurations within the brain and their relationship with opinion change, different modules
within the brain may be sensitive to different cognitive phenomena. Future research may extend
these findings to other cognitive actions that contribute to this complex decision, perhaps even
further disentangling the sub-elements of this process, including perception, weighing alterna-
tives, belief consistency, social influence, and so forth. Moreover, within our study, the over-
whelming contribution to the type of opinion change involves determining the consequences of
a murder trial, of which, little information is directly known to the subject and created a wide
range of responses (see also Figure S4 in the Supporting Information). The generalizability of
these findings is yet to be known; perhaps, even, the nonpersonal nature of these results have a
substantial impact on the findings. This might be an aspect of the findings to be assessed with
further experimentation.
Network Neuroscience
885
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
Within this context, our results are merely the first step toward understanding the dynamic
reconfigurations within the brain and how different context and content interact to give rise to
opinion change, highlighting the difference between in-person discussion and social media
interaction. Future studies will investigate the unique aspects of opinion change that are gen-
eralizable beyond the scenarios presented here, include within-subject condition comparisons
to inspect general properties of opinion change within these two social contexts, and perhaps
even understand the interaction of human biases in their interaction. For example, self-
referential opinion changes may suffer from the interesting optimism bias (e.g., Sharot,
2011), thus requiring different neural resources than non-self-referential.
Other methodological limitations are related not to experimental design but to the analyt-
ical technique in which we estimated the dynamic network reconfigurations. Here, we use
dynamic community detection (for review, see Garcia et al., 2018) to distill connectivity pat-
terns derived from phase-based statistical dependencies into communities, or clusters of elec-
trodes, and then estimated shifts in communities across time. We have previously extended
this method to EEG on a limited number of channels (Garcia et al., 2020a), but there are lin-
gering questions on the robustness of the method to varying number of channels, the cognitive
aspects the chosen temporal window might capture, and the parameter search. Future studies
may explore other techniques to capture the processes underlying opinion formation, change,
and generally complex decision-making. Moreover, future studies are needed to understand
the full contributions to session and subject-level variability as well as disentangle the “inter-
mittence” results as potentially marking different internal processes while interacting with
social media or different strategies deployed for social media interaction.
Conclusion
The current study used a complex network-based framework (dynamic community detection)
to investigate the relationship of brain dynamics during social media interaction with the opin-
ion change and/or formation processes. Our results indicate that the rapidly evolving network
dynamics in delta, beta, and gamma bands are the markers of influence of social media plat-
form interaction on opinions in a range of scenarios, such that the slower dynamics are asso-
ciated with individuals who are more likely to change their opinion. We also introduced a new
metric called intermittence to assess differences in the observed faster or slower network
dynamics. Estimating the intermittent and persistent network changes (as measured via inter-
mittence), our results suggest that the functional brain network structure for opinionators with
opinion change also show differences when interacting with social media platforms compared
to in-person discussions. Together, our results suggest unique decision-making operations dur-
ing social media interaction and represent traitlike dynamics in individuals that change or not
in their opinions.
METHODS
Participants
The data was collected from a cohort of 123 healthy participants between the ages of 18 and
40 years. Subjects were screened and the ones diagnosed with sleep, psychiatric, neurological,
eating, behavioral (including attention-deficit disorder), or cardiopulmonary/vascular disorders,
uncontrolled blood pressure, heart disease, HIV+/AIDS, head trauma within the past 5 years,
regular use of prescription drugs that can alter EEG or impair their ability to participate, use of
illegal drugs (recreational and medical marijuana users were not excluded), excessive use of
nicotine, alcohol and/or caffeine, untreated vision or hearing issues, pregnant or nursing, and
Network Neuroscience
886
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
inadequate familiarity with the English language were not included. For more information on the
recruiting procedure, we refer the reader to Richard et al. (2021). The data acquisition for this
study using human participants was reviewed and approved by Alpha IRB and Air Force
Research Laboratory. The participants provided their written informed consent to participate
in this study.
The Social Media Platform
This study used an innovative platform known as the Social Media Analytic Replication Toolkit
(SMART). SMART, which was developed by the Intific Division of Cubic Defense Applications
Inc., and allowed users to interact within a closed social media environment for experimen-
tation and real-time exercises that were inspired by Facebook and Twitter. Social media
accounts were created for each of the group participants that allowed the participant to like,
share, or retweet posts as they normally would do on their personal social media accounts.
Participant interactions with SMART were not broadcasted to other group participants’
accounts, but all other aspects of the platform were meant to make the “feel” of the platform
as realistic as possible. For screen shots of the platform, please see Supporting Information
Figure S7.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Procedures
For the experimental timeline and descriptive analysis of behavioral results (e.g., opinion
change), see Figure 1. Briefly, after arriving in the laboratory, subjects were presented with
questionnaires that asked their opinions on particular real-world scenarios found commonly
on social media platforms. Questionnaires were presented before and after interaction with the
simulated social media platform as well as after the in-person discussion segment. EEG was
recorded during these two interactive contexts, that is, social media interaction and in-person
discussion. To briefly describe the procedures, participants were seated in front of a computer
after instrumented with EEG and allowed to freely browse through the three scenarios and
interact (e.g., like, share, etc.) with the content freely. While the platform contained four to
five articles that could either skew the opinion of the individual in two different directions
(e.g., to vaccinate or to not vaccinate), there were no restrictions or trials; however, the social
media interaction was limited to 2 hours and the in-person discussion segment was limited to
20 minutes. The in-person discussion was administered directly following the completion of
the second questionnaire and participants were directed to a room to discuss each scenario. A
research technician was present and acted as a moderator to ensure that the discussion was
efficient and appropriate, and all topics were discussed for approximately 5 minutes. Each
scenario was addressed in the same order for all groups during the discussion period. Partic-
ipants were asked the same two questions for each scenario by the moderator: “What did you
decide and why?” and “What, if any, social media posts influenced your decision?” When the
discussion ended, the participants were instructed to complete scenario-specific question-
naires one last time before completing their participation in the study.
Scenarios
Three hypothetical scenarios were presented to the subjects. During each session, subjects
were exposed to the contents of three scenarios. The opinions of each subject in the sce-
narios were accessed through questionnaires that were delivered before and after interaction
with a social media simulation software. The following details the three scenarios analyzed
in this study.
Network Neuroscience
887
Brain network reconfigurations and social media
Free travel destinations. Within this scenario, subjects could choose between two locations
(Paris, France, or Sulawesi Island, Indonesia) for an all-paid 1-week vacation where each loca-
tion was vulnerable to different dangers; Paris could have large protests and sporadic violence,
and Sulawesi Island had the potential for a destructive tsunami. Articles presented to the sub-
jects mentioned these dangers and mentioned how nice the locations were to visit, with equal
representation. The subjects were also given the opportunity to volunteer in support of the
rebuilding effort following the catastrophic damage; they were also asked how much time they
were willing to dedicate to the rebuilding effort.
In this scenario, subjects were asked to imagine themselves in the jury of a trial,
Murder trial.
which was based on a true story. After receiving information about a case in which a young
female college student was murdered, they were asked whether the young man accused of
murder should be considered guilty or not, and if guilty, the length of the sentence or death.
In this scenario the subjects were asked about vaccinating one of two hypothet-
Vaccinations.
ical children after the older one started to show development impairments after being vacci-
nated. The questionnaire presented only the binary question if the subject would vaccinate or
not their second child, and the online content was equally for or against vaccinations.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Behavioral Metrics
The subjects answered questions about each of the hypothetical scenarios, and opinion
changes on the topics of the three scenarios were evaluated to determine the likelihood of
opinion change. Of the three scenarios, the vaccination scenario was the only completely
binary response as the travel and murder scenarios included follow-up questions that were
not binary. Thus, several steps were completed to include all three scenarios in the analysis
and construct a behavioral metric of opinion change.
For the three scenarios analyzed in this study, the social media opinion change was coded
as either a 0 (no change) or a 1 (change), indicating a change in opinion relative to the pre-
viously answered questionnaire. In other words, for the social media interaction, change was
measured relative to the first questionnaire, and for the in-person discussion condition, opin-
ion change was assessed relative to the second questionnaire (completed after social media
interaction). Change was coded as follows: for the travel scenario, a response was considered
a change (1) in opinion if any of the following were true: (a) the subject changed the destina-
tion choice from France to Indonesia or vice versa or (b) the subject modified their decision to
volunteer. For the murder scenario, a response was considered a change if any of the following
were true: (a) the subject changed their opinion from guilty to not guilty and, vice versa, (b)
the subject changed their opinion on prison time or punishment. For the vaccination sce-
nario, the responses were either yes or no, so a response was considered a change if it did not
match the previous response. If the answers are all identical to the previous survey, then the
response was coded as a 0, or no change. The social media opinion change score for each
subject was estimated as the sum of the social media opinion change scores of each individual
scenario and could be 0, 1, 2, or 3, where a 3 represents a change in every scenario and 0 in
none. Critically, groups were defined as those that did not change their opinion in any scenario
(no change) and those that changed their opinion in at least one scenario.
EEG Analysis
Preprocessing. EEG was acquired using the B-Alert R X24 wireless sensor headset (Advanced
Brain Monitoring, Inc., Carlsbad, CA) placed on the subjects before the subjects interacted
Network Neuroscience
888
Brain network reconfigurations and social media
with the social media platform. The headset is composed of 20 EEG sensors located according
to the International 10–20 montage at Fz, Fp1, Fp2, F3, F4, F7, F8, Cz, C3, C4, T3, T4, T5, T6,
O1, O2, Pz, POz, P3, and P4. Reference sensors were linked and located behind each ear
along the mastoid bone region. The sampling rate was 256 Hz, and the signal was filtered
through a high band-pass filter at 0.1 Hz and a low band-bass, fifth-order filter, at 100 Hz.
To insure high-quality data was collected, a maximum allowable impedance was set to
40 kΩ. Next, the data was band-pass filtered within common frequency bands including delta
(1–3 Hz), theta (3–7 Hz), alpha (8–13 Hz), beta (21–30), and gamma (25–40) using a low order
(3) zero-phase forward and reverse digital IIR filter in Matlab (Mathworks, Inc.).
Temporal windowing. Temporal windowing is a common procedure in studies of EEG, where it
may capture (a) the temporal scale of a variety of phenomena, (b) may indicate the specificity
and robustness of the chosen temporal window (10 s) in relation to the cognitive phenomenon
in question, and (c) lies at the intersection of measurement sensitivity, environmental noise,
and the covert mental events we study.
First, when considering functional connectivity, EEG has been shown to be sensitive to a
variety of task effects (e.g., Nidal & Malik, 2014), but it is also well established that its feature
capture more rapid changes in the brain (Garcia et al., 2020a, 2020b), and even more stable
effects as may be understood with disease models and genetics (e.g., Smit et al., 2008), span-
ning a large variety of temporal scales and a variety of behavior.
Second, in relation to the chosen time window (10 s), there’s a large literature that has
sought to understand how temporal windowing choice can affect the ability to capture differ-
ences in relation to trait-based information (or individual differences) as measured with EEG.
For example, previous studies have suggested that graph metrics, as estimated via connectivity
patterns derived from EEG, are susceptible to bias in the actual value, showing, for example,
shorter path lengths for shorter windows (see Fraschini et al., 2016; compare 2 s vs. 10 s) but
more stability above 10-s window sizes. More critical to the analyses within this manuscript,
however, is the relative nature of our connectivity matrices as they inform the dynamic com-
munities that are estimated across time. In comparing the connectivity matrices of the individ-
uals associated with each behavioral outcome (i.e., change, no change), there is no reason to
believe differential noise or bias in the estimated connectivity, as we are comparing them
rather than interpreting, say, the value of the metric. Moreover, some research has shown
an increase in robustness to noise of connectivity above 5 s or so (see Bonita et al., 2014).
Importantly, as well, is that we use the genLouvain algorithm to estimate communities, which
has been shown to be more robust to noise, number of clusters, and number of layers than
other algorithms (Bonita et al., 2014). Thus, here we have sampled the functional connectivity
over a robust time period with robust methodology.
Despite the evidence supporting our windowed approach, our general philosophy lies in a
narrow interpretation of our results that accounts for the nuances of our measurement tech-
nique, our methodology, the environment (or context), and the phenomenon in question. For
example, often the aforementioned robustness studies are either simulated or constructed from
resting-state data, often trying to understand the stability of these metrics within an individual
or across a population. While this approach is sound, it should really only be considered
within the context of the resting-state events, which are associated with mentalizing (Hein
& Singer, 2008). Critically, as well, we interpret our results with specific frequency bands,
where we assume low-frequency band phenomena to be sensitive to long-range communica-
tion within the brain and high-frequency band effects are capturing localized and rapid com-
munication within the brain. This has large impacts when interpreting the window size and the
Network Neuroscience
889
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
frequency bands that show these effects. For an extended analysis of different window size,
please see Supporting Information Figure S3. Importantly, the dynamics in connectivity, over-
all, can differentiate those that change and do not change their opinions within the gamma
band at windows at 10 s and above.
Functional connectivity analysis. To estimate the functional connectivity of the EEG recordings
we calculated the pair-wise weighted phase lag index (wPLI) within each frequency band of
interest, which is known to be highly sensitive to linear and nonlinear interactions (Imperatori
et al., 2019). For each sensor, the EEG (already band-wise filtered) was partitioned in L win-
dows with duration 10 s. The dynamic changes of this 10-s window highlight differences
between the two intervals in which subjects did and did not change their opinion. As has been
observed, each time window was used to calculate a matrix in which each entry Aij accounts
for the weighted wPLI (Vinck et al., 2011) for the pair of sensors i and j, calculated as:
Φ ≡ E I Xf g
f
j
I Xf g
E
f
j
g
j
g
j
;
where E{.} denotes the expected value and I Xf g is the imaginary part of the cross-spectrum of
the EEG recordings of sensors i and j. The temporal layers obtained by the described procedure
were then used for the dynamic community detection analysis described in the next section.
Importantly, though, the number of windows (L) were variable across subjects, with a mean L
across subjects of 372 (SD = 100).
Community Detection and Network Dynamics Metrics
While human brain mapping efforts have demonstrated a relationship between spatial speci-
ficity and cognitive functions, techniques rooted in network science provide a useful frame-
work for characterizing and understanding the spatiotemporal dynamics of the functional
systems subserving cognition (Bassett & Sporns, 2017). One of the core concepts at the basis
of network science is network modularity, which is the idea that neural units are structurally or
functionally connected, forming modules or clusters (Garcia et al., 2018). This organization
allows for the system to perform both local-level exchanges of information, while maintaining
system-level performance. Here, we examine whether a particular node’s propensity to
change communities (i.e., flexibility) was related to change in opinions after interaction with
a social network platform. To measure such changes in network communities during the inter-
action with the social media platform, a multilayer community detection analysis was
employed (Bassett et al., 2011; Mucha et al., 2010) on the aforementioned wPLI estimates
calculated for 10-s nonoverlapping windows (see Temporal windowing section above), with
each social media interaction session accounting for 372 ± 100 temporal layers. This method
uses a Louvain algorithm to maximize modularity (Blondel et al., 2008) to define functional
communities and is completed in several steps. First, it relies on two parameters, ɣ and ω, so-
called structural and temporal parameters of the analysis. We swept the parameter space from
.5 to .4 for each parameter, subject, and segment and compared the mean estimated modu-
larity value Q to a shuffled null dataset. We chose a parameter set that on average produced
more than one community and was the highest difference in modularity from the estimated
modularity from the shuffled null dataset (for a similar procedure, see Garcia et al., 2020a,
2020b). This resulted in ɣ = 1.1364 and ω = 0.5. Due to the nondeterministic nature of the
analysis, the chosen optimization procedure was repeated 100 times, since the algorithm is
susceptible to multiple solutions (Good et al., 2010). From these multiple iterations, the fol-
lowing community metrics were computed: (a) flexibility, or proportion of time during which
each node switches to a different community assignment; (b) allegiance, related to how long
Network Neuroscience
890
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
two nodes are connected to each other during the task, and a new proposed metric (c) inter-
mittence, defined as how rapidly two nodes connect and disconnect through communities.
Those metrics were calculated for each of the 100 iterations, and our results used the mean
value for all the iterations. In more concrete terms, the flexibility of each node corresponds to
the number of instances in which a node changes community affiliation, g, normalized by the
total possible number of changes that could occur across the layers L. In other words, the
flexibility of a single node i, ξi, may be estimated by
ξ
i ¼
gi
L − 1
:
Allegiance is a metric calculated for each pair of nodes and accounts for the proportion of the
total time a pair of nodes belongs to the same community, and is defined as:
Allegiancejk ¼
X
L
t¼1
1
L
δCj tð ÞCk tð Þ;
where δ denotes the Kronecker delta and Cl (t) denotes the community, which contains the
node l at time t. Therefore, δCj(t )Ck(t ) equals 1 if the nodes j and k are in the same community
at time layer t and equals 0 otherwise.
Furthermore, to account for the temporal dynamics of allegiance, we proposed a new met-
ric, intermittence, which tracks how frequently the two nodes change their affiliation from the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Flexibility, allegiance, and intermittence. (A) Example of a community structure assign-
ment for eight time layers. (B) Representation for each time layer of the community structure in A.
Network Neuroscience
891
Brain network reconfigurations and social media
same to different and vice versa. Intermittence is defined as:
i
X
1
L − 1
h
δCj tð ÞCk tð Þ 1 − δCj tþ1
ð
L−1
t¼1
Intermittencejk ¼
ÞCk tþ1
ð
Þ
þ δCj tþ1
ð
ÞCk tþ1
ð
h
Þ 1 − δCj tð ÞCk tð Þ
i
:
To visualize the concept of intermittence consider the example in Figure 6. First observe
that the allegiance between nodes 1 and 4 is equal to the allegiance of nodes 2 and 3; how-
ever, the link between nodes 1 and 4 is present for two large continuous epochs while the link
between nodes 2 and 3 is connected for many short epochs; this characterizes the intermit-
tence between nodes 2 and 3 as larger than the intermittence between nodes 1 and 4. Con-
sider now the nodes 5 and 6 both have the same allegiance with node 1; however, since node
6 changes its community assignment more often, its flexibility is higher than the flexibility
of node 5. Observe that unlike intermittence, flexibility is a property of the node and is not
calculated for individual links of the nodes.
Statistical Comparisons
Two types of statistical comparisons were completed within the manuscript. Primary compar-
isons were between two groups, of unequal sizes: individuals who did and did not change
their opinions. Due to the unequal sizes, bootstrap distributions (Wehrens et al., 2000) were
estimated and used to estimate p value and 95% confidence intervals (Figures 2 and 3). For
this method, 10,000 drawings (with replacement) were made within each group (change and
no change) for each node (Figure 2) or across nodes (Figure 3) and means for each 10,000
replicas were calculated resulting in a distributions of the mean value (e.g., flexibility in
Figure 2) or distributions of differences between groups (e.g., Figure 3B) were estimated. This
process generated bootstrap distributions, from which 95% confidence intervals were then
estimated. For the analysis in Figure 4, we carried out the bootstrap procedure with the ran-
dom sorting on the individual level, calculating the CoV for each of the 1,000 replicas with 30
randomly selected individuals each, and estimated the probability of the differences observed
between the two groups for each condition, and for each group in the two conditions.
ACKNOWLEDGMENTS
The views and conclusions contained in this document are those of the authors and should not
be interpreted as representing the official policies, either expressed or implied, of the U.S.
DEVCOM Army Research Laboratory or the U.S. Government.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00255.
The raw data supporting the conclusions of this article and code will be made available by
the authors upon request, without undue reservation. Neural metrics and behavioral change
and scripts to reproduce the figures and analyses may be found in the Supporting Information.
AUTHOR CONTRIBUTIONS
Italo’Ivo Lima Dias Pinto: Project administration; Software; Validation; Writing – original draft;
Writing – review & editing. Nuttida Rungratsameetaweemana: Methodology; Visualization;
Writing – original draft; Writing – review & editing. Kristen Flaherty: Data curation; Project
administration; Software; Validation; Writing – original draft. Aditi Periyannan: Formal analy-
sis; Project administration; Visualization; Writing – original draft; Writing – review & editing.
Amir Mghdadi: Conceptualization; Investigation; Project administration; Validation; Writing –
review & editing. Christian Richard: Conceptualization; Data curation; Funding acquisition;
Network Neuroscience
892
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
Investigation; Project administration; Writing – review & editing. Chris Berka: Conceptualiza-
tion; Funding acquisition; Investigation; Project administration; Supervision; Writing – review
& editing. Kanika Bansal: Conceptualization; Formal analysis; Funding acquisition; Project
administration; Software; Supervision; Visualization; Writing – original draft; Writing – review
& editing. Javier Omar Garcia: Conceptualization; Formal analysis; Funding acquisition; Pro-
ject administration; Software; Supervision; Writing – original draft; Writing – review & editing.
FUNDING INFORMATION
This research was sponsored by the US DEVCOM Army Research Laboratory and was
completed under Cooperative Agreement Numbers W911NF-20-2-0067 (I.L.D.P.), and
W911NF-17-2-0158 (K.B.). The data collected for the work presented herein was originally
supported by DARPA Contract No. FA865019C68899 awarded to Advanced Brain Moni-
toring (Carlsbad, CA).
REFERENCES
Alavash, M., Hilgetag, C. C., Thiel, C. M., & Giessing, C. (2015).
Persistency and flexibility of complex brain networks underlie
dual-task interference. Human Brain Mapping, 36(9), 3542–3562.
https://doi.org/10.1002/hbm.22861, PubMed: 26095953
Baek, E. C., O’Donnell, M. B., Scholz, C., Pei, R., Garcia, J. O.,
Vettel, J. M., & Falk, E. B. (2021). Activity in the brain’s valuation
and mentalizing networks is associated with propagation of
online recommendations. Scientific Reports, 11(1), 1–11.
https://doi.org/10.1038/s41598-021-90420-2, PubMed:
34045543
Bansal, K., Garcia, J. O., Lauharatanahirun, N., Muldoon, S. F.,
Sajda, P., & Vettel, J. M. (2021). Scale-specific dynamics of
high-amplitude bursts in EEG capture behaviorally meaningful
variability. NeuroImage, 241, 118425. https://doi.org/10.1016/j
.neuroimage.2021.118425, PubMed: 34303795
Barbey, A. K. (2018). Network neuroscience theory of human intel-
ligence. Trends in Cognitive Sciences, 22(1), 8–20. https://doi.org
/10.1016/j.tics.2017.10.001, PubMed: 29167088
Bassett, D. S., & Sporns, O. (2017). Network neuroscience. Nature
Neuroscience, 20(3), 353–364. https://doi.org/10.1038/nn.4502,
PubMed: 28230844
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of human
brain networks during learning. Proceedings of the National
Academy of Sciences of the United States of America, 108(18),
7641–7646. https://doi.org/10.1073/pnas.1018985108,
PubMed: 21502525
Bassett, D. S., Wymbs, N. F., Rombach, M. P., Porter, M. A., Mucha,
P. J., & Grafton, S. T. (2013). Task-based core-periphery organi-
zation of human brain dynamics. PLoS Computational Biology,
9(9), 1–16. https://doi.org/10.1371/journal.pcbi.1003171,
PubMed: 24086116
Betzel, R. F., & Bassett, D. S. (2017). Multi-scale brain networks.
NeuroImage, 160, 73–83. https://doi.org/10.1016/j.neuroimage
.2016.11.006, PubMed: 27845257
of Statistical Mechanics: Theory and Experiment, 2008(10).
https://doi.org/10.1088/1742-5468/2008/10/P10008
Bonita, J. D., Ambolode, L. C. C., Rosenberg, B. M. et al. (2014).
Time domain measures of inter-channel EEG correlations: A
comparison of linear, nonparametric and nonlinear measures.
Cognitive Neurodynamics, 8(1), 1–15. https://doi.org/10.1007
/s11571-013-9267-8, PubMed: 24465281
Bragin, A., Jandó, G., Nádasdy, Z., Hetke, J., Wise, K., & Buzsáki, G.
(1995). Gamma (40–100 Hz) oscillation in the hippocampus of
the behaving rat. Journal of Neuroscience, 15(1), 47–60. https://
doi.org/10.1523/ JNEUROSCI.15-01-00047.1995, PubMed:
7823151
Braun, U., Schäfer, A., Walter, H., Erk, S., Romanczuk-Seiferth, N.,
Haddad, L., Schweiger, J. I., Grimm, O., Heinz, A., Tost, H.,
Meyer-Lindenberg, A., & Bassett, D. S. (2015). Dynamic recon-
figuration of frontal brain networks during executive cognition in
humans. Proceedings of the National Academy of Sciences of the
United States of America, 112(37), 11678–11683. https://doi.org
/10.1073/pnas.1422487112, PubMed: 26324898
Buzsáki, G. (2006). Rhythms of the brain. Oxford, UK: Oxford Uni-
versity Press. https://doi.org/10.1093/acprof:oso/9780195301069
.001.0001
Buzsáki, G., & Wang, X. J. (2012). Mechanisms of gamma oscillations.
Annual Review of Neuroscience, 35, 203–225. https://doi.org/10
.1146/annurev-neuro-062111-150444, PubMed: 22443509
Canolty, R. T., Edwards, E., Dalal, S. S., Soltani, M., Nagarajan,
S. S., Berger, M. S., Barbaro, N. M., & Knight, R. T. (2006). High
gamma power is phase-locked to theta oscillations in human
neocortex. Science, 313(5793), 1626–1628. https://doi.org/10
.1126/science.1128115, PubMed: 16973878
Casado-Aranda, L. A., Venkatraman, V., Sánchez-Fernández, J., &
Luque-Martínez, T. (2020). Does partisan bias modulate neural
processing of political information? An analysis of the neural cor-
relates of corruption and positive messages. Political Psychology,
41(1), 145–164. https://doi.org/10.1111/pops.12581
Blondel, V. D., Guillaume, J. L., Lambiotte, R., & Lefebvre, E.
(2008). Fast unfolding of communities in large networks. Journal
Cascio, C. N., Scholz, C., & Falk, E. B. (2015). Social influence and
the brain: Persuasion, susceptibility to influence and
Network Neuroscience
893
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
retransmission. Current Opinion in Behavioral Sciences, 3, 51–57.
https://doi.org/10.1016/j.cobeha.2015.01.007
Cavanagh, J., & Frank, M. J. (2014). Frontal theta as a mechanism for
cognitive control. Trends in Cognitive Sciences, 18(8), 414–421.
https://doi.org/10.1016/j.tics.2014.04.012, PubMed: 24835663
Chai, L. R., Mattar, M. G., Blank, I. A., Fedorenko, E., & Bassett,
D. S. (2016). Functional network dynamics of the language sys-
tem. Cerebral Cortex, 26(11), 4148–4159. https://doi.org/10
.1093/cercor/bhw238, PubMed: 27550868
Chrobak, J. J., & Buzsáki, G. (1998). Gamma oscillations in the
entorhinal cortex of the freely behaving rat. Journal of Neurosci-
ence, 18(1), 388–398. https://doi.org/10.1523/ JNEUROSCI.18
-01-00388.1998, PubMed: 9412515
Cocchi, L., Gollo, L. L., Zalesky, A., & Breakspear, M. (2017).
Criticality in the brain: A synthesis of neurobiology, models
and cognition. Progress in Neurobiology, 158, 132–152. https://
doi.org/10.1016/j.pneurobio.2017.07.002, PubMed: 28734836
Cooper, N., Garcia, J. O., Tompson, S. H., O’Donnell, M. B., Falk,
E. B., & Vettel, J. M. (2019). Time-evolving dynamics in brain
networks forecast responses to health messaging. Network Neu-
roscience, 3(1), 138–156. https://doi.org/10.1162/netn_a_00058,
PubMed: 30793078
Ding, Z., Chen, X., Dong, Y., & Herrera, F. (2019). Consensus
reaching in social network DeGroot model: The roles of the self-
confidence and node degree. Information Sciences, 486, 62–72.
https://doi.org/10.1016/j.ins.2019.02.028
Engel, A. K., & Fries, P. (2010). Beta-band oscillations—Signalling the
status quo? Current Opinion in Neurobiology, 20(2), 156–165.
https://doi.org/10.1016/j.conb.2010.02.015, PubMed: 20359884
Falk, E. B., Way, B. M., & Jasinska, A. J. (2012). An imaging genetics
approach to understanding social influence. Frontiers in Human
Neuroscience, 6, 1–13. https://doi.org/10.3389/fnhum.2012
.00168, PubMed: 22701416
Falk, E., & Scholz, C. (2018). Persuasion, influence, and value: Per-
spectives from communication and social neuroscience. Annual
Review of Psychology, 69(1), 329–356. https://doi.org/10.1146
/annurev-psych-122216-011821, PubMed: 28961060
Fellows, L. K. (2004). The cognitive neuroscience of human
decision-making: A review and conceptual framework. Behavioral
and Cognitive Neuroscience Reviews, 3(3), 159–172. https://doi.org
/10.1177/1534582304273251, PubMed: 15653813
Fraschini, M., Demuru, M., Crobe, A., Marrosu, F., Stam, C. J., &
Hillebrand, A. (2016). The effect of epoch length on estimated
EEG functional connectivity and brain network organisation.
Journal of Neural Engineering, 13(3), 036015. https://doi.org/10
.1088/1741-2560/13/3/036015, PubMed: 27137952
Garcia, J. O., Ashourvan, A., Muldoon, S., Vettel, J. M., & Bassett, D. S.
(2018). Applications of community detection techniques to brain
graphs: Algorithmic considerations and implications for neural
function. Proceedings of the IEEE, 106(5), 846–867. https://doi.org
/10.1109/JPROC.2017.2786710, PubMed: 30559531
Garcia, J. O., Ashourvan, A., Thurman, S. M., Srinivasan, R., Bassett,
D. S., & Vettel, J. M. (2020a). Reconfigurations within resonating
communities of brain regions following TMS reveal different
scales of processing. Network Neuroscience, 4(3), 611–636.
https://doi.org/10.1162/netn_a_00139, PubMed: 32885118
Garcia, J. O., Battelli, L., Plow, E., Cattaneo, Z., Vettel, J., &
Grossman, E. D. (2020b). Understanding diaschisis models of
attention dysfunction with rTMS. Scientific Reports, 10(1), 1–15.
https://doi.org/10.1038/s41598-020-71692-6, PubMed: 32913263
Gerraty, R. T., Davidow, J. Y., Foerde, K., Galvan, A., Bassett, D. S.,
& Shohamy, D. (2018). Dynamic flexibility in striatal-cortical cir-
cuits supports reinforcement learning. Journal of Neuroscience,
38(10), 2442–2453. https://doi.org/10.1523/JNEUROSCI.2084
-17.2018, PubMed: 29431652
Gold, J. I., & Shadlen, M. N. (2007). The neural basis of decision-making.
Annual Review of Neuroscience, 30(1), 535–574. https://doi.org/10
.1146/annurev.neuro.29.051605.113038, PubMed: 17600525
Golnar-Nik, P., Farashi, S., & Safari, M. S. (2019). The application of
EEG power for the prediction and interpretation of consumer
decision-making: A neuromarketing study. Physiology and
Behavior, 207, 90–98. https://doi.org/10.1016/j.physbeh.2019
.04.025, PubMed: 31047949
Good, B. H., De Montjoye, Y. A., & Clauset, A. (2010). Performance
of modularity maximization in practical contexts. Physical
Review E, 81(4), 1–20. https://doi.org/10.1103/ PhysRevE.81
.046106, PubMed: 20481785
Haegens, S., Händel, B. F., & Jensen, O. (2011). Top-down con-
trolled alpha band activity in somatosensory areas determines
behavioral performance in a discrimination task. Journal of Neu-
roscience, 31(14), 5197–5204. https://doi.org/10.1523
/JNEUROSCI.5199-10.2011, PubMed: 21471354
Han, S., Cui, Q., Wang, X., Li, L., Li, D., He, Z., Guo, X., Fan, Y.-S.,
Guo, J., Sheng, W., Lu, F., & Chen, H. (2020). Resting state
functional network switching rate is differently altered in bipolar
disorder and major depressive disorder. Human Brain Mapping,
41(12), 3295–3304. https://doi.org/10.1002/ hbm.25017,
PubMed: 32400932
Hanes, K., Andrewes, D., & Pantelis, C. (1995). Cognitive flexibility
and complex integration in Parkinson’s disease, Huntington’s
disease, and Schizophrenia. Journal of the International Neuro-
psychological Society, 1(6), 545–553. https://doi.org/10.1017
/S1355617700000679, PubMed: 9375241
Hardmeier, M., Hatz, F., Bousleiman, H., Schindler, C., Stam, C. J.,
& Fuhr, P. (2014). Reproducibility of functional connectivity and
graph measures based on the phase lag index (PLI) and weighted
phase lag index (wPLI) derived from high resolution EEG. PLoS
One, 9(10). https://doi.org/10.1371/journal.pone.0108648,
PubMed: 25286380
Hawkins, J., Lewis, M., Klukas, M., Purdy, S., & Ahmad, S. (2019).
A framework for intelligence and cortical function based on grid
cells in the neocortex. Frontiers in Neural Circuits, 12, 121.
https://doi.org/10.3389/fncir.2018.00121, PubMed: 30687022
Hein, G., & Singer, T. (2008). I feel how you feel but not always:
The empathic brain and its modulation. Current Opinion in Neu-
robiology, 18(2), 153–158. https://doi.org/10.1016/j.conb.2008
.07.012, PubMed: 18692571
Holley, R. A., & Liggett, T. M. (1975). Ergodic theorems for weakly
interacting infinite systems and the voter model. The Annals of
Probability, 3(4), 643–663. https://doi.org/10.1214/aop/1176996306
Imperatori, L. S., Betta, M., Cecchetti, L., Canales-Johnson, A.,
Ricciardi, E., Siclari, F., Pietrini, P., Chennu, S., & Bernardi,
G. (2019). EEG functional connectivity metrics wPLI and wSMI
account for distinct types of brain functional interactions. Scien-
tific Reports, 9(1), 1–15. https://doi.org/10.1038/s41598-019
-45289-7, PubMed: 31222021
Network Neuroscience
894
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
Izuma, K., & Adolphs, R. (2013). Social manipulation of preference
in the human brain. Neuron, 78(3), 563–573. https://doi.org/10
.1016/j.neuron.2013.03.023, PubMed: 23664619
Jirsa, V. K., Stacey, W. C., Quilichini, P. P., Ivanov, A. I., & Bernard,
C. (2014). On the nature of seizure dynamics. Brain, 137(8),
2210–2230. https://doi.org/10.1093/ brain/awu133, PubMed:
24919973
Kappes, A., Harvey, A. H., Lohrenz, T., Montague, P. R., & Sharot,
T. (2020). Confirmation bias in the utilization of others’ opinion
strength. Nature Neuroscience, 23(1), 130–137. https://doi.org
/10.1038/s41593-019-0549-2, PubMed: 31844311
Keles, B., McCrae, N., & Grealish, A. (2020). A systematic review:
The influence of social media on depression, anxiety and psycho-
logical distress in adolescents. International Journal of Adoles-
cence and Youth, 25(1), 79–93. https://doi.org/10.1080/02673843
.2019.1590851
Khanna, P., & Carmena, J. M. (2015). Neural oscillations: Beta
band activity across motor networks. Current Opinion in Neuro-
biology, 32, 60–67. https://doi.org/10.1016/j.conb.2014.11.010,
PubMed: 25528615
Klucharev, V., Munneke, M. A. M., Smidts, A., & Fernández, G.
(2011). Downregulation of the posterior medial frontal cortex
prevents social conformity. Journal of Neuroscience, 31(33),
11934–11940. https://doi.org/10.1523/ JNEUROSCI.1869-11
.2011, PubMed: 21849554
Krapivsky, P. L., & Redner, S. (2003). Dynamics of majority rule in
two-state interacting spin systems. Physical Review Letters, 90(23),
238701. https://doi.org/10.1103/ PhysRevLett.90.238701,
PubMed: 12857298
Lauharatanahirun, N., Bansal, K., Thurman, S. M., Vettel, J. M.,
Giesbrecht, B., Grafton, S., Elliott, J. C., Flynn-Evans, E., Falk,
E., & Garcia, J. O. (2020). Flexibility of brain regions during
working memory curtails cognitive consequences to lack of
sleep. arXiv. arXiv:2009.07233. https://doi.org/10.48550/arXiv
.2009.07233
Leopold, D. A., Murayama, Y., & Logothetis, N. K. (2003). Very
slow activity fluctuations in monkey visual cortex: Implications
for functional brain imaging. Cerebral Cortex, 13(4), 422–433.
https://doi.org/10.1093/cercor/13.4.422, PubMed: 12631571
Lewis, M., Purdy, S., Ahmad, S., & Hawkins, J. (2019). Locations in
the neocortex: A theory of sensorimotor object recognition using
cortical grid cells. Frontiers in Neural Circuits, 13, 22. https://doi
.org/10.3389/fncir.2019.00022, PubMed: 31068793
Li, L., Li, K. K., & Li, J. (2019). Private but not social information
validity modulates social conformity bias. Human Brain Map-
ping, 40(8), 2464–2474. https://doi.org/10.1002/ hbm.24536,
PubMed: 30697880
Lydon-Staley, D. M., Kuehner, C., Zamoscik, V., Huffziger, S.,
Kirsch, P., & Bassett, D. S. (2019). Repetitive negative thinking
in daily life and functional connectivity among default mode,
fronto-parietal, and salience networks. Translational Psychiatry,
9(1). https://doi.org/10.1038/s41398-019-0560-0, PubMed:
31534117
Mahmoodi, K., West, B. J., & Grigolini, P. (2017). Self-organizing
complex networks: Individual versus global rules. Frontiers in
Physiology, 8, 1–11. https://doi.org/10.3389/fphys.2017.00478,
PubMed: 28736534
Mattar, M. G., Betzel, R. F., & Bassett, D. S. (2016). The flexible
brain. Brain, 139(8), 2110–2112. https://doi.org/10.1093/brain
/aww151, PubMed: 27457229
Mountcastle, V. B. (1978). An organizing principle for cerebral
function: The unit module and the distributed system. In G. M.
Edelman & V. B. Mountcastle (Eds.), The mindful brain. Cambridge,
MA: MIT Press.
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Onnela,
J. P. (2010). Community structure in time-dependent, multiscale,
and multiplex networks. Science, 328(5980), 876–878. https://
doi.org/10.1126/science.1184819, PubMed: 20466926
Nakao, T., Kanayama, N., Katahira, K., Odani, M., Ito, Y., Hirata, Y.,
Nasuno, R., Ozaki, H., Hiramoto, R., Miyatani, M., & Northoff,
G. (2016). Post-response βγ power predicts the degree of
choice-based learning in internally guided decision-making. Sci-
entific Reports, 6(August), 1–9. https://doi.org/10.1038
/srep32477, PubMed: 27576670
Nakao, T., Miyagi, M., Hiramoto, R., Wolff, A., Gomez-Pilar, J.,
Miyatani, M., & Northoff, G. (2019). From neuronal to psycho-
logical noise—Long-range temporal correlations in EEG intrinsic
activity reduce noise in internally-guided decision-making. Neu-
roImage, 201, 116015. https://doi.org/10.1016/j.neuroimage
.2019.116015, PubMed: 31306772
Newman, M. E. (2006). Modularity and community structure in net-
works. Proceedings of the National Academy of Sciences of the
United States of America, 103(23), 8577–8582. https://doi.org/10
.1073/pnas.0601602103, PubMed: 16723398
Nidal, K., & Malik, A. S. (Eds.). (2014). EEG/ERP analysis: Methods
and applications. Boca Raton, FL: CRC Press. https://doi.org/10
.1201/b17605
Osborne-Crowley, K. (2020). Social cognition in the real world:
Reconnecting the study of social cognition with social reality.
Review of General Psychology, 24(2), 144–158. https://doi.org
/10.1177/1089268020906483
Paban, V., Modolo, J., Mheich, A., & Hassan, M. (2019). Psycholog-
ical resilience correlates with EEG source-space brain network
flexibility. Network Neuroscience, 3(2), 539–550. https://doi.org
/10.1162/netn_a_00079, PubMed: 30984906
Pikovsky, A., Rosenblum, M., & Kurths, J., & (2001). Synchroniza-
tion: A universal concept in nonlinear sciences. Cambridge, UK:
C a m b r i d g e U n i v e r s i t y P r e s s . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 7
/CBO9780511755743
Rao, R. P., & Ballard, D. H. (1999). Predictive coding in the visual
cortex: A functional interpretation of some extra-classical
receptive-field effects. Nature Neuroscience, 2(1), 79–87.
https://doi.org/10.1038/4580, PubMed: 10195184
Reddy, P. G., Mattar, M. G., Murphy, A. C., Wymbs, N. F., Grafton,
S. T., Satterthwaite, T. D., & Bassett, D. S. (2018). Brain state flex-
ibility accompanies motor-skill acquisition. NeuroImage, 171,
135–147. https://doi.org/10.1016/j.neuroimage.2017.12.093,
PubMed: 29309897
Richard, C., Karić, M. S., McConnell, M., Poole, J., Rupp, G., Fink,
A., Meghdadi, A., & Berka, C. (2021). Elevated inter-brain coher-
ence between subjects with concordant stances during discus-
sion of social issues. Frontiers in Human Neuroscience, 15,
1–11. https://doi.org/10.3389/fnhum.2021.611886, PubMed:
34054446
Network Neuroscience
895
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain network reconfigurations and social media
Rilling, J. K., & Sanfey, A. G. (2011). The neuroscience of social
decision-making. Annual Review of Psychology, 62, 23–48.
https://doi.org/10.1146/annurev.psych.121208.131647,
PubMed: 20822437
Ruelle, D. (1976). The Lorenz attractor and the problem of turbu-
lence. In Turbulence and Navier Stokes equations (pp. 146–158).
Berlin, Germany: Springer. https://doi.org/10.1007/BFb0091453
Rungratsameetaweemana, N., Itthipuripat, S., Salazar, A., &
Serences, J. T. (2018). Expectations do not alter early sensory
processing during perceptual decision-making. Journal of Neuro-
science, 38(24), 5632–5648. https://doi.org/10.1523/JNEUROSCI
.3638-17.2018, PubMed: 29773755
Rungratsameetaweemana, N., Lainscsek, C., Cash, S. S., Garcia,
J. O., Sejnowski, T. J., & Bansal, K. (2021). Brain network dynam-
ics codify heterogeneity in seizure propagation. bioRxiv. https://
doi.org/10.1101/2021.06.12.448205
Schmälzle, R., O’Donnell, M. B., Garcia, J. O., Cascio, C. N.,
Bayer, J., Bassett, D. S., Vettel, J. M., & Falk, E. B. (2017). Brain
connectivity dynamics during social interaction reflect social net-
work structure. Proceedings of the National Academy of Sciences
of the United States of America, 114(20), 5153–5158. https://doi
.org/10.1073/pnas.1616130114, PubMed: 28465434
Schroeder, C. E., & Lakatos, P. (2009). The gamma oscillation: Mas-
ter or slave? Brain Topography, 22(1), 24–26. https://doi.org/10
.1007/s10548-009-0080-y, PubMed: 19205863
Shafiei, S. B., Elsayed, A. S., Hussein, A. A., Iqbal, U., & Guru, K. A.
(2020). Evaluating the mental workload during robot-assisted sur-
gery utilizing network flexibility of human brain. IEEE Access, 8,
204012–204019. https://doi.org/10.1109/ACCESS.2020.3036751
Sharot, T. (2011). The optimism bias. Current Biology, 21(23),
R941–R945. https://doi.org/10.1016/j.cub.2011.10.030,
PubMed: 22153158
Sherman, L. E., Payton, A. A., Hernandez, L. M., Greenfield, P. M.,
& Dapretto, M. (2016). The power of the like in adolescence:
Effects of peer influence on neural and behavioral responses to
social media. Psychological Science, 27(7), 1027–1035. https://
doi.org/10.1177/0956797616645673, PubMed: 27247125
Shine, J. M., & Poldrack, R. A. (2018). Principles of dynamic net-
work reconfiguration across diverse brain states. NeuroImage,
180, 396–405. https://doi.org/10.1016/j.neuroimage.2017.08
.010, PubMed: 28782684
Smit, D. J., Stam, C. J., Posthuma, D., Boomsma, D. I., & de Geus,
E. J. (2008). Heritability of “small-world” networks in the brain: A
graph theoretical analysis of resting-state EEG functional connec-
tivity. Human Brain Mapping, 29(12), 1368–1378. https://doi.org
/10.1002/hbm.20468, PubMed: 18064590
Spinney, L. (2017). The shared past that wasn’t: How Facebook,
fake news and friends are altering memories and changing his-
tory. Nature, 543(9), 168–170. https://doi.org/10.1038/543168a,
PubMed: 28277522
Stancák, Jr., A., & Pfurtscheller, G. (1996). Event-related desynchro-
nisation of central beta-rhythms during brisk and slow self-paced
finger movements of dominant and nondominant hand. Cogni-
tive Brain Research, 4(3), 171–183. https://doi.org/10.1016
/S0926-6410(96)00031-6, PubMed: 8924046
Strogatz, S. H. (2000). From Kuramoto to Crawford: Exploring the
onset of synchronization in populations of coupled oscillators.
Physica D: Nonlinear Phenomena, 143(1–4), 1–20. https://doi
.org/10.1016/S0167-2789(00)00094-4
Sugimura, K., Iwasa, Y., Kobayashi, R., Honda, T., Hashimoto, J.,
Kashihara, S., Zhu, J., Yamamoto, K., Kawahara, T., Anno, M.,
Nakagawa, R., Hatano, K., & Nakao, T. (2021). Association
between long-range temporal correlations in intrinsic EEG activ-
ity and subjective sense of identity. Scientific Reports, 11(1), 422.
https://doi.org/10.1038/s41598-020-79444-2, PubMed:
33431948
Telesford, Q. K., Lynall, M. E., Vettel, J., Miller, M. B., Grafton, S. T.,
& Bassett, D. S. (2016). Detection of functional brain network
reconfiguration during task-driven cognitive states. NeuroImage,
142, 198–210. https://doi.org/10.1016/j.neuroimage.2016.05
.078, PubMed: 27261162
Thomas Yeo, B. T., Krienen, F. M., Eickhoff, S. B., Yaakub, S. N.,
Fox, P. T., Buckner, R. L., Asplund, C. L., & Chee, M. W. L.
(2015). Functional specialization and flexibility in human associ-
ation cortex. Cerebral Cortex, 25(10), 3654–3672. https://doi.org
/10.1093/cercor/bhu217, PubMed: 25249407
Uddin, L. Q. (2020). Bring the noise: Reconceptualizing spontaneous
neural activity. Trends in Cognitive Sciences, 24(9), 734–746.
https://doi.org/10.1016/j.tics.2020.06.003, PubMed: 32600967
Vinck, M., Oostenveld, R., Van Wingerden, M., Battaglia, F., &
Pennartz, C. M. A. (2011). An improved index of phase-
synchronization for electrophysiological data in the presence
of volume-conduction, noise and sample-size bias. NeuroImage,
55(4), 1548–1565. https://doi.org/10.1016/j.neuroimage.2011.01
.055, PubMed: 21276857
Wallace, K. J., & Hofmann, H. A. (2021). Decision-making in a
social world: Integrating cognitive ecology and social neurosci-
ence. Current Opinion in Neurobiology, 68, 152–158. https://doi
.org/10.1016/j.conb.2021.03.009, PubMed: 33915497
Wehrens, R., Putter, H., & Buydens, L. M. C. (2000). The bootstrap: A
tutorial. Chemometrics and Intelligent Laboratory Systems, 54(1),
35–52. https://doi.org/10.1016/S0169-7439(00)00102-7
Westerman, D., Spence, P. R., & Van Der Heide, B. (2014). Social
media as information source: Recency of updates and credibility
of information. Journal of Computer-Mediated Communication,
19(2), 171–183. https://doi.org/10.1111/jcc4.12041
Yin, D., & Kaiser, M. (2021). Understanding neural flexibility from a
multifaceted definition. NeuroImage, 235, 118027. https://doi.org
/10.1016/j.neuroimage.2021.118027, PubMed: 33836274
Yoo, E., Rand, W., Eftekhar, M., & Rabinovich, E. (2016). Evaluat-
ing information diffusion speed and its determinants in social
media networks during humanitarian crises. Journal of Opera-
tions Management, 45, 123–133. https://doi.org/10.1016/j.jom
.2016.05.007
Network Neuroscience
896
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
7
0
2
0
4
6
4
6
7
n
e
n
_
a
_
0
0
2
5
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3