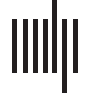FORSCHUNG
Edge-based network analysis reveals
frequency-specific network dynamics in aberrant
anxiogenic processing in rats
Yin-Shing Lam1,2
, Xiu-Xiu Liu1,2, Ya Ke1,2, and Wing-Ho Yung1,2
1School of Biomedical Sciences, Faculty of Medicine, The Chinese University of Hong Kong, Hongkong
2Gerald Choa Neuroscience Centre, The Chinese University of Hong Kong, Hongkong
Schlüsselwörter: Edge-based network analysis, Anxiety, Phase transfer entropy, Phase locking value,
Theta oscillation
Keine offenen Zugänge
Tagebuch
ABSTRAKT
Uncovering interactions between edges of brain networks can reveal the organizational
principle of the networks and also their dysregulations underlying aberrant behaviours such as
in neuropsychiatric diseases. In this study, we looked into the applicability of edge-based
network analysis in uncovering possible network mechanisms of aberrant anxiogenic
Verarbeitung. Utilizing a rat model of prodromal Parkinson’s disease we examined how a
dorsomedial striatum–tied associative network (DSAN) may mediate context-based
anxiogenic behaviour. Following dopamine depletion in the dorsomedial striatum, ein
exaggerated bottom-up signalling (posterior parietal-hippocampal-retrosplenial to anterior
prefrontal-cingulate-amygdala regions) and gradient specific to the theta frequency in this
network was observed. This change was accompanied by increased anxiety behaviour of the
Tiere. By employing an edge-based approach in correlating informational flow (Phase
transfer entropy) with functional connectivity of all edges of this network, we further explore
how the abnormal bottom-up signalling might be explained by alterations to the informational
flow-connectivity motifs in the network. Our results demonstrate usage of edge-based network
analysis in revealing concurrent informational processing and functional organization
dynamics across multiple pathways in a brain network. This approach in unveiling network
abnormalities and its impact on behavioural outcomes would be useful in probing the network
basis of neuropsychiatric conditions.
ZUSAMMENFASSUNG DES AUTORS
This study employs a unique approach to study the neural basis of anxiety by quantifying the
correlation of concurrent fluctuations in information flow and connectivity between all edges
of the brain network involved. This is the first time that such an edge-based technique is used
to analyse field potential signals of the brain in an animal model of a neuropsychiatric
disorder. We demonstrate how such edge-based analysis complements other analyses such as
local field potential power and gross network changes. This work helps lay the foundation
for future investigations into decoding the network aberrations of a wide spectrum of
neuropsychiatric disorders.
Zitat: Lam, Y.-S., Liu, X.-X., Ke, Y.,
& Yung, W.-H. (2022). Edge-based
network analysis reveals frequency-
specific network dynamics in aberrant
anxiogenic processing in rats. Netzwerk
Neurowissenschaften, 6(3), 816–833. https://doi
.org/10.1162/netn_a_00251
DOI:
https://doi.org/10.1162/netn_a_00251
zusätzliche Informationen:
https://doi.org/10.1162/netn_a_00251
Erhalten: 13 November 2021
Akzeptiert: 10 April 2022
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Corresponding Authors:
Ya Ke
yake@cuhk.edu.hk
Wing-Ho Yung
whyung@cuhk.edu.hk
Handling-Editor:
Richard Betzel
Urheberrechte ©: © 2022
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Network edges:
A measure of association or
interaction between two brain
regions or nodes of a broader
Netzwerk.
Phase locking value:
A measure of functional connectivity
between two brain regions by
measuring if there is a consistent
phase difference between the
oscillatory local field potential of the
two regions.
Phase transfer entropy:
A measure of directional information
flow via Shannon entropy of phase
distribution between oscillatory local
field potential of two brain regions.
Theta band:
Oscillations of local field potentials
at a frequency of 4 Zu 8 Hz.
EINFÜHRUNG
Analysis of brain functional connectivity at the network level in both health and disease is a
rapidly growing field (Bassett & Bullmore, 2009). Jedoch, current research lacks extensive
interrogation into the complex network coordination across multiple pathways, ergebend
incomplete understanding of physiological, psychiatric, and emotional disorders. Elucidation
of the interaction between the connections between two component regions, or edges, across
the entire network can reveal more extensive information on the functional coordination
between sets of pathways. Network configuration is well recognized to be nonstatic and exhibits
strong temporal fluctuations and functionally relevant patterns (Calhoun et al., 2014; Faskowitz
et al., 2020; Müller et al., 2020; Shine et al., 2015). Daher, to truly unveil network-wide coordi-
nation of complex cognitive processes, it is crucial to understand the relationship between net-
work edges via quantifying the temporal fluctuations in their properties. Interactions between
brain regions occur in a frequency-specific manner via oscillating field potentials (Buzsáki
et al., 2013). In den vergangenen Jahren, phase-dependent measures of brain oscillations used as network
parameters offer distinct advantages in quantifying functional and information flow relation-
ships between brain regions, primarily due to their independence on the amplitudes of the sig-
nal (Robinson et al., 2013; Siems & Siegel, 2020). While phase locking value (PLV) is a measure
of functional connectivity between two regions (Aydore et al., 2013), directed phase transfer
entropy (dPTE) can reveal directional information flow or transfer between them (Lobier
et al., 2014). Such metrics allow the interaction between network edges to be robustly quanti-
fied and compared.
Anxiety disorders constitutes the most common type of psychiatric disorder (Bandelow &
Michaelis, 2015). Anxiety is an innate emotional state induced by potentially threatening stim-
uli exhibited by almost all animals possessing higher cognitive capabilities (Steimer, 2002). Es
involves multicircuit brain-wide integration of environmental information from the sensory
cortices and the internal psychological state of the animal from higher cognitive centres of
das Gehirn (Barbot & Carrasco, 2018; Martin et al., 2009; Sussman et al., 2016), allowing indi-
viduals to respond to environmental threats and uncertainties in accordance with internal
goals and motivation appropriately. In rodents, fMRI studies have shown the recruitment of
multiple associative brain regions in anxious and introspective states (Becerra et al., 2011).
Fear and anxiety are also associated with multiple groups of neural pathways across associa-
tive regions (LeDoux, 2000). Theta band brain activity alterations of different brain regions
such as the hippocampal and frontal regions in anxious states are also well documented
(Buzsáki, 2004; Cavanagh & Frank, 2014; Jacinto et al., 2013; Soltani Zangbar et al., 2020).
From comprehensive tracing and functional studies, the dorsomedial striatum (DMS)
exhibits both close functional connectivity and receives broad excitatory projections from
associative brain regions encompassing the frontal cortex, cingulate cortex, posterior cingulate
Kortex, parietal associative cortex, and amygdala and hippocampus (Delcasso et al., 2014;
Greene et al., 2019; Hunnicutt et al., 2016). Such regions are recruited in brain processes such
as environmental context processing, memory retrieval, and social interaction (Cwik et al.,
2014; Delcasso et al., 2014; Glimcher & Fehr, 2014; Hunnicutt et al., 2016; Ritchey & Cooper,
2020). Studies have demonstrated that manipulating interactions between the medial pre-
frontal regions and the DMS can induce avoidance and approach behaviours of rodents
(Gunaydin et al., 2017; Loewke et al., 2020). daher, the striatum and closely associated
cortical and subcortical brain regions may constitute a network that bridge contextual infor-
mation and behavioural output, which in turn regulates the expression of anxiety. Specifi-
cally, we hypothesize that a network consisting of associative cortical regions demonstrated
to have significant projections and close functional associations to the DMS in rats (Delcasso
Netzwerkneurowissenschaften
817
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Posterior medial subnetwork:
A group of associative regions
located posteriorly of the brain,
encompassing the retrosplenial,
parietal associative cortices and
dorsal hippocampus that have close
functional ties.
6-Hydroxydopamine:
A neurotoxin that specifically targets
and depletes dopaminergic and
noradrenergic neurons.
et al., 2014; Genzel, 2020; Hunnicutt et al., 2016; Navarro-Lobato & Genzel, 2020) is under
the regulation of the DMS dopaminergic innervation and is critical in the genesis and
expression of anxiety. These areas include the medial prefrontal cortex (mPFC), the anterior
cingulate cortex (ACC), the central nucleus of amygdala (CeA), and components of the
default mode network posterior medial subnetwork encompassing the retrosplenial cortex
(RSC), the parietal associative cortex (PtA), and the dorsal hippocampus (dHip).
The function of the striatum is tightly regulated by dopamine, a key mediator in a multitude
of higher ordered cognitive and emotional states, such as motivation, cognitive flexibility, Und
Stimmungsregulierung (Cavanagh et al., 2017; Seo et al., 2008; Westbrook & Mutiger, 2016). Der
projection from the substantia nigra to the striatum, the nigrostriatal pathway, is the largest
dopaminergic pathway in the mammalian brain. DMS has been demonstrated to directly influ-
ence avoidance behaviours and anxiety in both rodents and humans via dopaminergic signal-
ling (Hilbert et al., 2015; Nguyen et al., 2019); In disorders of defunct dopaminergic signalling
such as Parkinson’s disease (PD), although motor symptoms are the major manifestations of the
Krankheit, anxiety is extremely prevalent during the prodromal phase of the disease. Anxiety and
depression are also regarded as preclinical risk factors in PD development (Chen & Marsh,
2014; Meireles & Massano, 2012; Pontone et al., 2009). daher, the loss of dopaminergic
signalling in nonmotor territories of the striatum could be a precipitating trigger in the devel-
opment of anxiety (Erro et al., 2012).
Hier, to investigate the network basis of anxiety induced by an artificial depletion of dopa-
minergic innervation of the DMS via direct 6-hydroxydopamine (6-OHDA) injection (Alvarez-
Fischer et al., 2008), we propose an analysis framework, similar in concept to cofluctuations of
connectivity patterns demonstrated recently by Esfahlani et al. (2020) and edge-centric func-
tional connectivity proposed by Faskowitz et al. (2020) that was applied to human fMRI con-
nectivity values. We integrate the information obtained from PLV and dPTE of the relevant
associative network, quantifying the relationship between network edges to observe multipath-
way and frequency-specific motifs of informational processing and functional connectivity.
We also observed their changes during the genesis of anxiety and investigated the relevance
of such motifs in the alterations of behaviour of rats in an anxiogenic environment and the
overall functional alterations of the network.
MATERIALS AND METHODS
Rats and Stereotaxic Surgery
Adult male Sprague-Dawley rats used in this study were bred and maintained by the Labora-
tory Animal Service Centre of The Chinese University of Hong Kong. The animal control room
was controlled at a temperature of 23°C. All animals were handled in strict accordance with
Chinese University of Hong Kong guidelines, and the procedures were approved by the
Animal Experimentations and Ethics Committee. For surgical procedures, rats were anesthe-
tized via intraperitoneal injection of ketamine (75 mg/kg, i.p.) and xylazine (6 mg/kg, i.p.)
before being placed gently and fixed on the stereotaxic apparatus.
6-OHDA and Saline Infusion Into the DMS
A total of 27 male Sprague-Dawley rats with mean initial weight of around 260 G (each rat not
exceeding 5 g from 260 G) underwent stereotaxic infusion of either 6-OHDA, in order to arti-
ficially deplete the dorsomedial striatum of its dopaminergic innervation, or infusion of saline
for the control group. Cranial openings were made on the skull above the striatal region. Four-
teen rats had 0.4 μl 6-OHDA of hydrobromide solution (40 μg 6-OHDA.HBr per 1 μl of sterile
Netzwerkneurowissenschaften
818
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Elevated plus maze:
An animal behavioral test that
measures level of innate anxiety by
measuring time, speed and distance
travelled on either the open arm,
closed arm (with walls) or the center
piece connecting the arms of a plus-
shaped maze elevated from the
Boden.
Dorsomedial striatum–tied
associative network:
A collection of cortical and
subcortical structures that have been
demonstrated to have direct neural
connections with the dorsomedial
striatum.
saline with 0.2% ascorbic acid) stereotaxically infused via a 1-μl Hamilton syringe into the
anterior dorsomedial striatum (AP: 1.0 mm; ML: 2.3 mm; DV: 5.5 mm) at a rate of
0.06 μl/min. Thirteen rats received stereotaxic infusion of equal volumes of sterile saline at
the same rate acting as the control group. The rats received subcutaneous anti-inflammatory,
analgesic, and antibiotic injection immediately following the surgery. Rats were handled
by the handler 5 min per day for 7 days after surgery.
Behavioral Tests
Rats were first habituated to the behavioural room for 30 min before any test took place. Der
elevated plus maze was elevated 50 cm from the ground, with the closed arms the dimension
von 15 × 50 cm (40-cm walls), the centre piece being 15 × 15 cm, the two open arms 15 ×
50 cm. The behavioural room was lit via a 220-W fluorescent ring lamp. As rats explored
the plus maze, AnyMaze software tracked the rats centre point on the maze and calculated
various behavioural parameters.
Electrode Implantation Into the Dorsomedial Striatum–Tied Associative Network Regions
Four rats treated with 6-OHDA and four rats treated with saline were allowed 1 week of
recovery from the injection surgery and then unilateral implantation of electrodes (left side)
were carried out. Electrodes constructed from twisting two strands of 0.00315-inch-diameter
stablohm 675 wire (California Fine Wire Co.) into a 2- to 3-cm-long electrode; 250 μm of the
recording tip of the electrode is deinsulated and 3 mm of the connector end of the electrode
deinsulated and welded to adaptors. Cranial openings were made above the target brain
Regionen; the electrodes were then lowered with the recording tips embedded into the target
Gehirnregionen: mPFC (AP: 2.52 mm; ML: −0.34 mm; DV: 4.9 mm), ACC (AP: 1.08 mm; ML:
−0.34 mm; DV: 2.5 mm), RSC (AP: −3.36 mm; ML: 0.34 mm; DV: 2.2 mm), CeA (AP:
−2.76 mm; ML: −4.5 mm; DV: 8.7 mm), PtA (AP: −4.08 mm; ML: −3.0 mm; DV: 1.10 mm),
and dHip (AP: −4.36 mm; ML: −1.6 mm; DV: 3.3 mm). The reference screw was screwed into
the cerebrospinal fluid above the cerebellum and connected to the adaptor. Superglue and
dental cement were used to stabilize the electrodes on the cranium. The adaptors were
inserted into housing pieces and fixed onto the dental cement. Rats received subcuta-
neous anti-inflammatory, analgesic, and antibiotic injection immediately, 24 hours and
48 hours after the surgery. Rats were handled by the handler for 5 min per day for 10 Tage
following surgery.
Histology
Rats underwent transcardial perfusion of 4% PFA. Histological sectioning and analysis were
performed after experiments were performed to confirm dopaminergic depletion of the DMS
via staining of tyrosine hydroxylase (Primary goat anti-TH antibody, secondary anti-goat rabbit
antibody) and to confirm correct placement of electrodes into the target brain regions.
Local Field Potential Recording and Analysis of Local Field Potential Readings
Local field potentials (LFPs) were recorded from the target brain regions referenced against
cerebellar activity with an acquisition rate of 1000 Hz. Ten days following electrode implan-
tation rats were habituated in the recording room for 30 min before being connected to the
Plexon electrophysiology acquisition system via a headstage and cable. Ten minutes of LFPs
were recorded for rats when they were awake and resting in their home cage, 10 min when the
rats were placed on the elevated plus maze to freely explore, Und 10 min when the rats were
Netzwerkneurowissenschaften
819
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
immediately placed back to their home cage in the same room. Power spectra of local field
potentials were calculated from the normalized raw 10-min voltage data notch filtered
0–0.01 Hz and 48–52 Hz. Relative power was calculated for each frequency from 0–100 Hz
bei 0.0244 Hz increments.
Theta oscillations were then isolated via band-pass filtering of 4–8 Hz oscillations from the
raw LFP, and Hilbert transformation was applied to identify the phase between 0 and 2π of the
signal at each recorded time point. Phase locking value was calculated between two LFP
epochs of the same rat via the following equation:
PLV X; Yð
Þ ¼ 1
N
XN
n¼1
D
Þ
ei θx nð Þ−θy nð Þ
where X and Y are the LFP time series of two different brain regions, N is the number of data
points of the LFP epoch (z.B., N = 1,000 für 1 s of LFP data), θx(N) and θy(N) is the phase of the
oscillations of X and Y at that particular time point.
For phase transfer entropy (PTE), this involves the calculation of Shannon entropy or uncer-
tainty of a particular stretch of time of LFP by observing the distribution of phases via the fol-
lowing equation:
H θx tð Þ
D
Þ ¼
P bð Þ (cid:2) log
(cid:2)
(cid:3)
1
P bð Þ
XB
b¼1
H(θx(T)) is the Shannon phase entropy of a time series X, where θx(T) is the phase of the oscil-
lations of X for any time point t, B is the number of bins in which phase from 0 to 2π is divided,
b is the numerical number assigned to a bin, with P(B) being the probability that at any time
point the time series would be in a particular phase bin. The size of each bin was determined
by the following equation first proposed by Scott (1992):
(cid:5)
(cid:4)
Binsize ¼ 3:49 (cid:2) mean σphase
(cid:2) N−1=3
N is the number of data points within the LFP epoch, and σ is the standard deviation of signal
phases within that instantaneous time. The phase of each data point is then fitted into bins
created from dividing 0 to 2π into bin size intervals, where the number of bins is equal to
2π/bin size. The above equation is referenced from the “brainstorm-tools” code in Fraschini
and Hillebrand (2016). Phase bin sizes of resting-state data (each segment and frequency
band) are shown in Supporting Information Figure S1; there was a consistent bin number of
22 for the vast majority of segments in all frequency bands (only some delta band segments
had deviations not exceeding four bins) and bin size of 0.22–0.31 for delta band and
0.28–0.30 rads for all other frequency bands in both saline and 6-OHDA treated rats.
As described by Lobier et al. (2014), in order to determine the information transfer between
two LFP time series X and Y, with the phase θx(T) and θy(T), jeweils, Shannon entropy of Y’s
phase at the present timepoint t conditioned on its own past at time t 0 is subtracted from the
Shannon entropy of Y’s phase conditioned on its own past and also the past of X’s phase, als
illustrated in the below equation:
(cid:6)
(cid:6)
(cid:6)θy t 0ð Þ; θx t 0ð ÞÞ
(cid:6)θy t 0ð ÞÞ − H θy tð Þ
PTEX→Y ¼ H θy tð Þ
(cid:4)
(cid:4)
Conditional entropy was calculated by this general formula:
H Bð jAÞ ¼ −
X
a2A;b2B
p a; bð
Þ log
Þ
p a; bð
p að Þ
Netzwerkneurowissenschaften
820
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Edge-based network analysis:
Analysis of networks with the
emphasis on edge properties and
interactions between edges.
Daher, the larger PTE is, the more θx(T 0) can reduce the entropy/uncertainty of θy(T), suggesting a
greater informational flow from the source origin of X to Y. The lag δ = t − t 0, is determined by
the number of full oscillation from 0 to 2π within a time series, daher, it is determined as:
δ ¼ N
C
N is the combined total data point from the two LFP time series compared, and C is the
combined number of times with which the phase goes from less than π to being larger than π.
PTE value is then normalized over direction to give directed PTE as described by Engels
et al. (2017):
directed PTEX→Y ¼
PTExy
PTExy þ PTEyx
Daher, when directed PTE is larger than 0.5, there is preferential flow from X to Y and vice
versa. In this manuscript, directed PTE value is further subjected to further adjustment by sub-
tracting the directed PTE by 0.5
directed PTEX→Y ¼ directed PTEX→Y − 0:5
Daher, when dPTE is larger than 0, there is preferential flow and net outflow of information from
X to Y and vice versa.
A Framework of Edge-Based Network Analysis Based on Correlated Information Flow and
Funktionale Konnektivität
We propose a framework of network analysis based on the correlation of phase-based
information flow and functional connectivity derived from LFP values. With respect to
the Dorsomedial striatum–tied associative network (DSAN), following injection of 6-OHDA
or saline into DMS, electrodes were implanted into target brain areas (Figure 1A). LFP were
recorded from these areas when the animals underwent behavioural assessment in resting state
Vor (RS), on an elevated plus maze (EPM), or post-EPM resting state (RS2) (Figure 1A). Für
each state, the LFP recording was first divided into fixed (10 S) Epochen, which was then band-
pass filtered through specific physiological frequencies (delta, theta, alpha, beta, gamma).
Instantaneous phase of LFP oscillations were obtained after Hilbert transformation (Figure 1B).
PLV and dPTE were then derived from the transformed LFP signals to quantify the functional
connectivity and information flow in different times (Abbildung 1C). Jedoch, just comparing
the differences between different states does not unveil much information regarding changes
to the underlying dynamics of a network. As networks are dynamic and exhibit evolving inter-
actions across time, especially during exploratory behaviour, the relationships between net-
work parameters between different edges can reveal how different pathways across a network
are associated with one another and whether bundles of pathways are regulated together. Von
correlating the dPTE values of each possible network edge direction against each other, a large-
scale multipathway informational processing relationship of the network could be revealed
(Figure 1D), in which changes in dPTE of individual pathways can be contextualized in a
broader network organizational framework of dPTE changes in other pathways. Zusätzlich,
by correlating dPTE values against PLV values of each brain pair and identifying significant,
concurrent changes in informational flow and function (Figure 1D), we can unveil how func-
tional connectivity within a network responds to information flow, which can give further
insight into the self-regulation of networks, as was observed in approach/avoidance behav-
iours (Spielberg et al., 2012).
Netzwerkneurowissenschaften
821
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
Figur 1. Workflow of edge-based network analysis of induced behavioral changes. (A) 6-OHDA or saline is infused into the dorsomedial
striatum of Sprague-Dawley rats. Rats were then implanted with six electrodes and LFP recorded during the resting (RS), elevated plus maze
(EPM), and resting 2 (RS2) state. (B) Due to the spontaneous and dynamic nature of informational flow and connectivity, LFP signals were
divided into 10-s epochs, in which for each segment a band-pass filter is applied and phase calculated via Hilbert transformation. dPTE and
PLV values were then calculated for each pair of brain regions for that particular LFP epoch. Epochs of LFP corresponding to rat transitioning
were also isolated, and for each epoch a dPTE and PLV value were calculated. (C) Network properties dynamically change in a temporal
dimension, with dPTE and PLV values fluctuating. (D) Network pathways that are strongly associated with each other can be unveiled by
correlating dPTE values of each edge and direction and observing the extent of their covariation throughout time via Pearson’s correlation.
Zusätzlich, information flow can be correlated with network organization via correlating dPTE with PLV values of various edges of the network
to reveal pathways involved in network arrangement.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
.
T
The use of 10-s epochs was a balance between having sufficient segments (around 60 pro
rat per state) for dynamic analysis and for each segment to have sufficient amount of phase
changes for robust dPTE measurements, even for the delta band at 1–4 Hz (für 1 Hz, there will
still be 10 full oscillations for that epoch with more than 20 phase bins). Fraschini et al. (2016)
has recommended, based on human EEG data, using epochs of more than 6 s for more stable
connectivity measurements, and their results showed that network parameters are more stable
with epoch lengths approaching 10 s or above.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
ERGEBNISSE
Increased Level of Anxiety in 6-OHDA-Treated Rats on the Elevated Plus Maze
Infusion of 6-OHDA into the DMS was performed to deplete dopaminergic fibres locally,
which was then verified post-mortem via tyrosine hydroxylase (TH) staining (Figure 2A). Fol-
lowing 7 days of recovery and habituation to the experimenter, the rats were then placed on
the EPM for 10 min of free exploration. Comparing the behavioral parameters with rats
treated with saline (n = 9) to the DMS, 6-OHDA-treated rats (n = 10) have a significantly
reduced time spent, total distance travelled, and average speed on the open arm of the EPM.
On the EPM, 6-OHDA-treated rats have a significantly reduced time spent, total distance
travelled, and average speed on the open arm. The differences were insignificant and less
Netzwerkneurowissenschaften
822
Edge-based network analysis of aberrant anxiogenic processing in rats
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
N
e
N
_
A
_
0
0
2
5
1
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
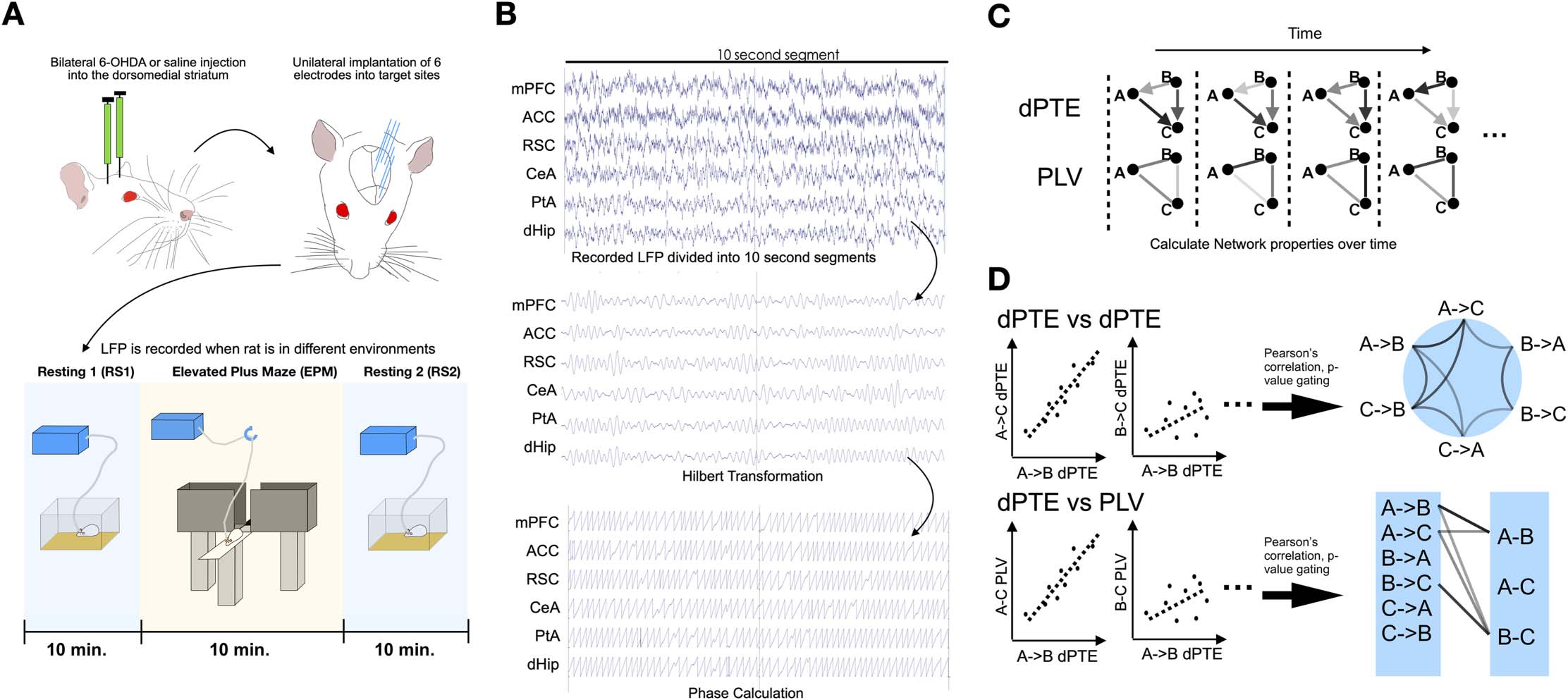
Figur 2. A 6-OHDA infusion and alterations to avoidance behaviors. (A) Schematic demonstrating the area in which 6-OHDA or saline will
be infused into and representative TH staining of a 6-OHDA-infused rat, showing drastic depletion of dopaminergic fibres in the targeted
dorsomedial striatum (bar = 1 mm). (B) Representative tracing of 6-OHDA- and saline-infused rat centre point on the elevated plus maze.
(C) Time, Distanz, and speed of 6-OHDA- (n = 10) and saline-infused rats (n = 9) in the closed, centre, and open zones of the elevated plus
maze. *P < 0.05, **p < 0.01 (two-sample t test).
pronounced in the centre zone and completely negligible in the closed arm (Figure 2B and
2C). Open arms are considered to be more anxiogenic to rats due to a natural fear of heights
and a tendency to stay in enclosed spaces (closed arm), and hence reduced spontaneous
motor/exploratory behaviour as indicated by speed, distance travelled, and time spent on
the open arm point toward increased anxiety (Walf & Frye, 2007). Such changes revealed
an increase in anxiogenic avoidance behaviours by rats with focal dopaminergic depletion
confined to the DMS.
Changes in Posterior-Anterior Theta Band Power in an Anxiogenic Context
Correct placement of recording electrodes targeting DSAN component regions in 6-OHDA or
saline-treated rats was verified histologically post-mortem (Supporting Information Figure S2).
Consistently, 6-OHDA-treated animals spent more time in both the transition zone and closed
arm (Supporting Information Figure S3A, B), and transitioning into the closed arm for a higher
number of times (Supporting Information Figure S3C). Similar changes related to transitioning
were also observed in nonelectrode-implanted rats such as a tendency to enter a closed arm
(Supporting Information Figure S4A), reduced time moving away from the closed arm
Network Neuroscience
823
Edge-based network analysis of aberrant anxiogenic processing in rats
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
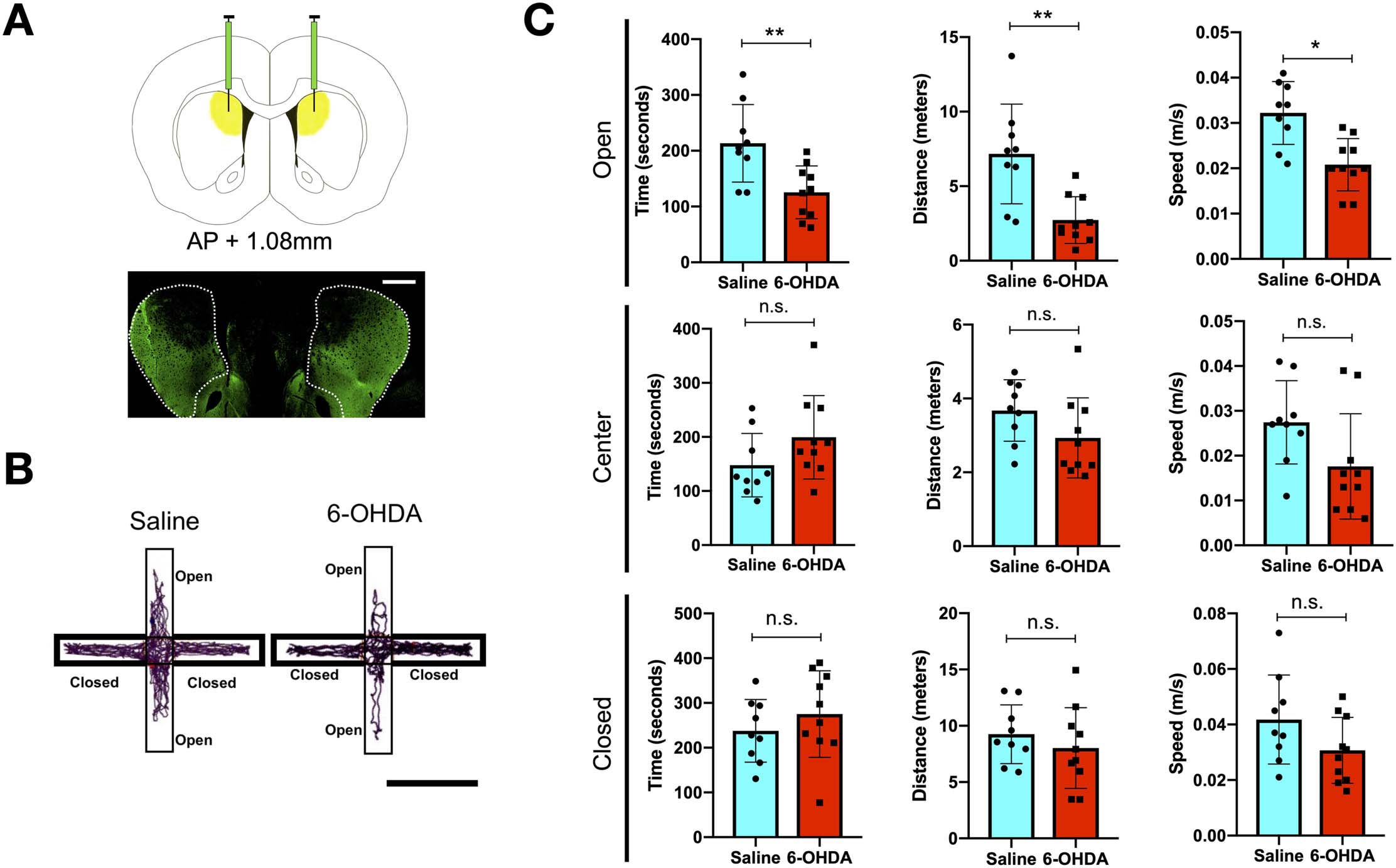
Figure 3. Theta band oscillatory changes in DSAN component brain regions. (A) Representative power spectra of control rat hippocampus
during rest and EPM: Following z-transformation of 10-min LFP signals for each state, theta peak were present on almost all brain regions for all
saline and 6-OHDA-infused rats during resting (one 6-OHDA-infused rat and one saline-infused rat do not possess a theta peak in the mPFC
during the resting state) and shifts when placed on the EPM. (B) The frequency of the maximum relative power theta peak for each DSAN
network brain region (n = 6 brain regions) for each individual rat in the resting state (RS) and on the EPM. (C) Relative theta peak power of each
brain region in the resting state (RS) and on the EPM for 6-OHDA (n = 4) and saline-infused rats (n = 4). *p <0.05, **p < 0.01, ***p < 0.001,
****p < 0.0001 (paired t test).
(Supporting Information Figure S4B), and higher immobility in the centre zone (Supporting
Information Figure S4C) in 6-OHDA-treated rats. Analysis of the power spectrum of the LFPs
revealed a peak in the theta rhythm in most recorded brain regions in all the animals (Figure 3A).
Notably, for both groups of animals, when comparing RS and EPM LFPs, there was a shift
in theta peak frequency across all brain regions examined. Furthermore, in the same rat,
the variation in the change of theta peak frequency among brain regions was very low
(Figure 3B). These findings are consistent with theta rhythm being enhanced during explor-
atory behaviour. In contrast to the consistent shift in peak frequency, a differential response
between control and 6-OHDA-treated rats was observed with respect to the changes in the
relative power of the theta rhythm. For saline-treated rats, there were no significant differ-
ence ( p = 0.1566, F = 1.835 one-way ANOVA test) in relative theta power between the
brain regions and no significant changes in power when the rats were transferred to the
EPM (Figure 3C). In contrast, in 6-OHDA-treated rats, we found that the relative power
across individuals for each brain region exhibited less variation. Furthermore, for ACC,
CeA, and dHip there was a significant increase in relative theta peak power when the rats
were placed on the EPM. Also notable is that the theta power of posterior regions (PtA and
dHip) on the EPM had a significantly higher value than the anterior regions (mPFC and ACC)
(Figure 3C).
Network Neuroscience
824
Edge-based network analysis of aberrant anxiogenic processing in rats
Changes in Theta Band Informational Flow and Functional Connectivity in the DSAN
We then asked whether there were frequency-specific alterations in information flow when the
rats were in the EPM compared with the resting states. To do so, the averaged dPTE values of
each frequency band on the EPM were subtracted with values obtained during RS and RS2.
Consistent with a dominant role of theta activities, theta band alterations were the most wide-
spread and prominent in terms of scale of change across the DSAN when the rats were on the
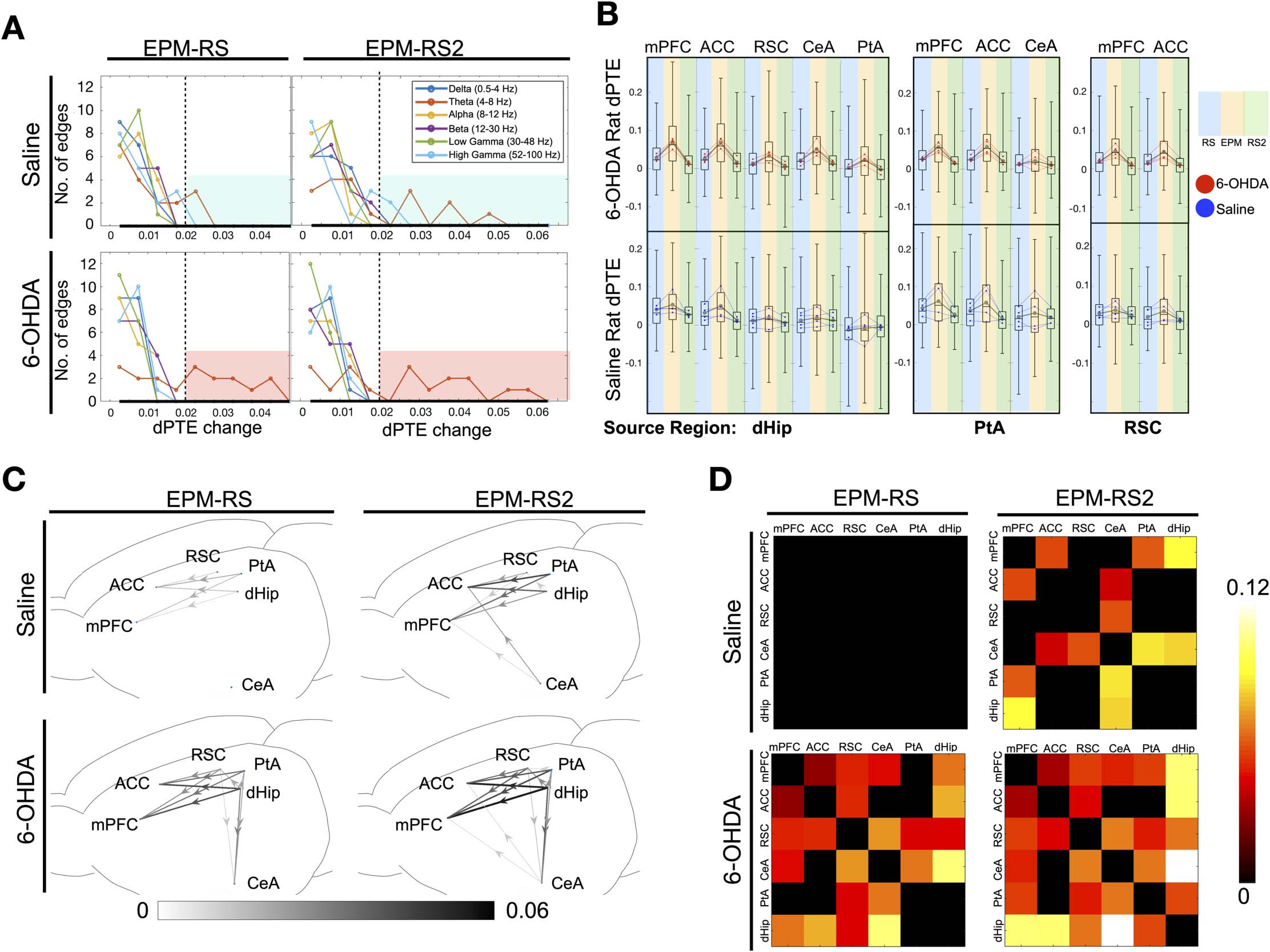
EPM than during the resting states (Figure 4A) as seen by the higher number of network edges
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Changes in dPTE and PLV values from resting to EPM and from EPM to resting. (A) Histogram of dPTE scalar changes of network
edges in various frequency bands when averaged dPTE values from EPM were subtracted with resting state (EPM-RS) and resting state 2 (EPM-
RS2) values in 6-OHDA- and saline-infused rats (average of n = 240 epochs per state from four rats). (B) Bee swarm plot and box plot of theta
band dPTE values of pooled individual epochs from the major bottom-up pathways with dHip, PtA, and RSC as the source region. Gray dot
and lines indicate changes in group mean (average of n = 240 epochs per state from four rats), whereas red or blue dot and lines indicate
changes in the mean of the epochs of each individual rat (60 epochs per state per individual rat) of 6-OHDA and saline-infused rats respec-
tively. (C) Network schematic revealing the specific connections, directionality, and magnitude of the dPTE alterations in the theta band when
averaged dPTE values from EPM were subtracted with resting-state (EPM-RS) and resting-state 2 (EPM-RS2) values in 6-OHDA- and saline-
infused rats (average of n = 240 epochs per state from four rats) with a threshold of p < 0.001. (D) Matrices demonstrating difference in theta
band PLV values across various the DSAN edges when rats were on the EPM and when in resting state (EPM-RS) and resting state 2 (EPM-RS2)
(average of n = 240 epochs for each state from four rats).
Network Neuroscience
825
Edge-based network analysis of aberrant anxiogenic processing in rats
having a large scalar change in dPTE. If by examining the number of edges with a dPTE change
of at least 0.02 (highlighted in red for 6-OHDA group, blue in saline group), for EPM-RS, saline
group had a total of two edges and 6-OHDA rats had 10; for EPM-RS2, saline group had a total
three edges and 6-OHDA rats had 10, showing that the theta dPTE increase was more prom-
inent for 6-OHDA-treated rats (Figure 4A). When looking at each rat individually, we can see
that 6-OHDA-treated rats when compared to saline-treated rats showed greater significance in
theta dPTE changes in terms of the total number of edges, with p < 0.01 (Supporting Informa-
tion Figure S5A, B)as well as overall p value of changes for each edge for EPM-RS and EPM-
RS2 combined (Supporting Information Figure S5C). Examining the raw dPTE values for each
LFP segment in each state revealed that, in 6-OHDA treated rats, dHip, PtA, and RSC outflow
to the mPFC, ACC, and CeA exhibited a clear increase in dPTE values on the EPM and a
decrease in RS2. In contrast, the theta band dPTE changes are much less consistent in the con-
trol animals (Figure 4B). When the magnitude of theta band dPTE change along with the direc-
tionality were plotted out, for 6-OHDA treated rats, the RSC-PtA-dHip region, that is, all the
components of the posterior medial subnetwork, are the major theta outflow region with the
PtA being the most prominent followed by the dHip (Figure 4C). In saline treated rats, there
were less significant increases in the theta band dPTE of the same RSC-PtA-dHip outflow
pathways (EPM-RS: 11 significant pathways in 6-OHDA group, 5 in saline; EPM-RS2: 12
in 6-OHDA, 7 in saline). The mean dPTE change of the outflow pathways were also lower
as compared to the changes in 6-OHDA-treated rats (EPM-RS: 0.0240 for 6-OHDA group,
0.0090 for saline; EPM-RS2: 0.0306 for 6-OHDA, 0.0166 for saline). When placed back to
their home cage in RS2, there is a corresponding decrease in theta band dPTE of roughly the
same pathways for both 6-OHDA- and saline-treated rats.
For PLV, low-frequency bands (delta, theta, alpha) showed a greater increase in overall net-
work connectivity, which was more prominent in 6-OHDA-treated rats (Supporting Informa-
tion Figure S6). Looking at the theta band specifically, significant increases in PLV value
between the CeA and RSC, PtA, dHip regions and also between the hippocampus and frontal
regions regions were observed in 6-OHDA-treated rats (Figure 4D). Such prominent increases
are not observed in saline-treated rats. Surrogate time series were generated via randomly shuf-
fling LFP time segments of each channel by 1,000 times. The dPTE and PLV values among
shuffled data and dPTE and PLV values approached zero, showing that the patterns of dPTE
and PLV were not the consequence of random noise within the time series (Supporting Infor-
mation Figure S7).
Loss of Striatal Dopamine Alters Multiedge Theta Band Motifs
As pointed out previously, network configuration and metrics (i.e., dPTE, PLV) are not static
and exhibit temporal fluctuations, transitioning through various network states. For example,
we observed that the dPTE values of the bottom-up pathways in the DSAN fluctuate over time,
with the fluctuations enhanced when the animals were on the plus maze (Supporting Informa-
tion Figure S8), reflecting the spontaneous nature of brain networks. In order to pursue the
question of the dynamics of theta band DSAN information flow, we took advantage of such
fluctuations by looking into the large-scale multipathway informational processing relationship
in the DSAN via correlating dPTE values of each possible edge direction against each other
(example in Supporting Information Figure S9), gating for positive and significant correlations.
When correlating dPTE values against each other, one can identify for 6-OHDA-treated rats
significant correlation between dHip, PtA, and RSC outflow toward the mPFC, ACC, and CeA.
Since the dPTE is antisymmetric and since Pearson’s correlation is nondirectional, reciprocal
increases in correlation were also observed in mPFC, ACC, and CeA outflow toward the
Network Neuroscience
826
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
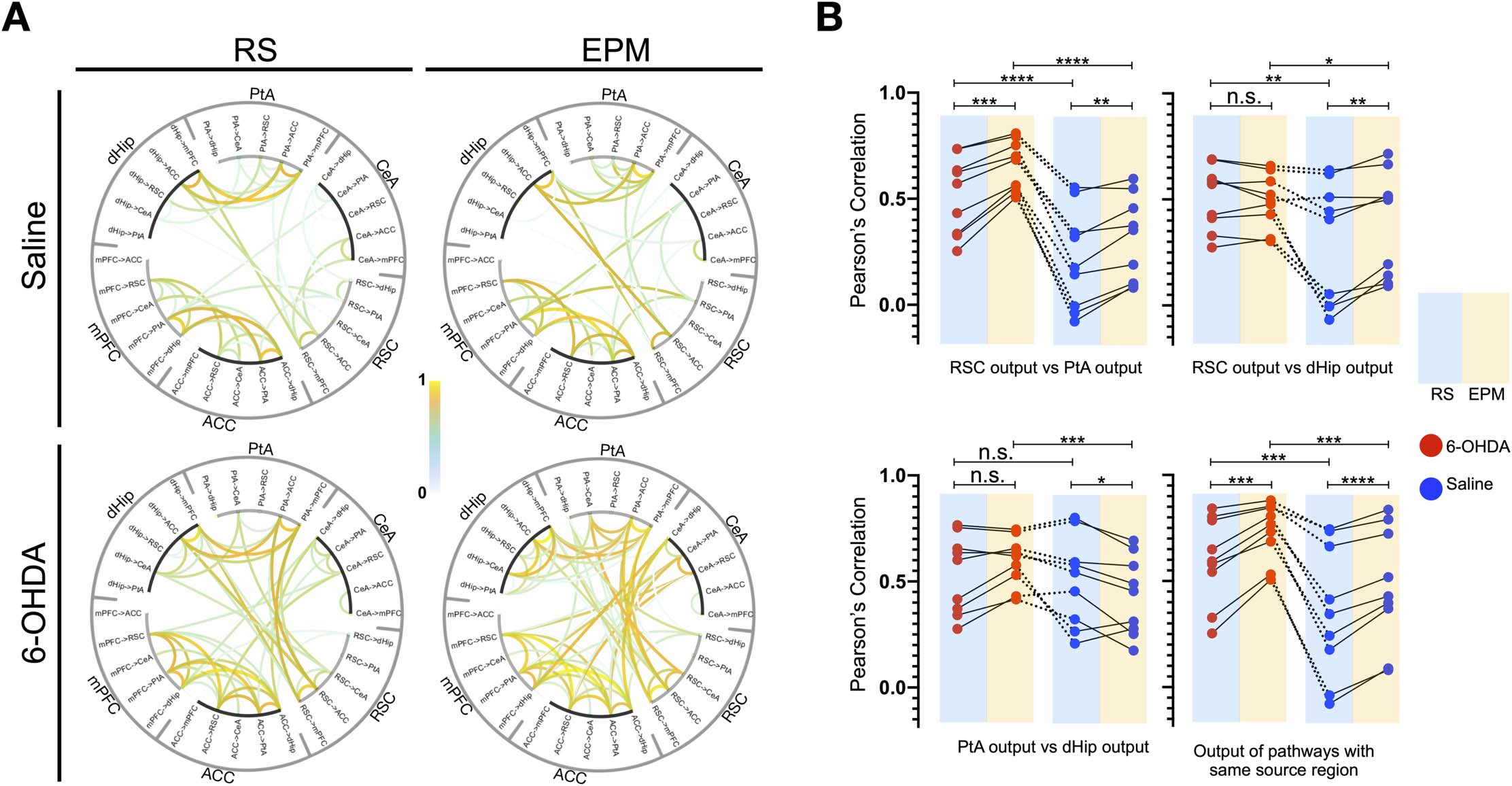
Figure 5. Altered theta band edge-based informational flow motifs in the DSAN following dopaminergic depletion. (A) Pearson’s correlation
of theta band dPTE between all possible brain pairs/directionality reveals large-scale coregulatory informational outflow pathways (average of
n = 240 epochs per state from four rats) in 6-OHDA- and saline-infused rats; correlations were presented at a threshold of p < 1 × 10−10
(Pearson’s correlation p value). (B) Differences between the correlation of RSC, PtA, and dHip outflow pathways to the CeA, ACC, and mPFC
in 6-OHDA- and saline-infused rats during a resting and EPM state. Dashed lines indicate the same pathway combination compared in
6-OHDA and saline rats. Nine pairings between RSC and PtA outflow pathways, nine pathway pairings between RSC and dHip outflow
pathways, and nine pathway pairings between PtA and dHip outflow pathways. Nine pairing between outflow pathways of the same source
region. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 (paired t test).
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
posterior regions (Figure 5A). However, since the correlation coefficients were enhanced
when the animals were placed on the EPM and from the gross changes in theta dPTE there
is an increased posterior to anterior flow, it is therefore more likely that dHip, PtA, and
RSC outflow toward the mPFC, ACC, and CeA is responsible for the higher correlations
observed.
To quantify the observed changes, correlation coefficients between RSC, PtA, and dHip
outflow edges to the mPFC, ACC, and CeA pathways are compared (Figure 5B). In both
saline- and 6-OHDA-treated rats, for correlations between pathways of the same source region
(e.g., dHip outflow to mPFC vs. dHip outflow to CeA) and between outflow pathways of the
RSC and PtA (e.g.. RSC outflow to the mPFC vs. PtA outflow to the mPFC), when the animals
were on the EPM, correlation values increased. However, 6-OHDA-treated rats had higher
correlation values in both resting and EPM state. For correlations between RSC and dHip out-
flow, 6-OHDA-treated rats had higher correlations in both resting and EPM state but only
saline-treated rats exhibited an increase on the EPM. For correlations between PtA and dHip
outflow, the EPM 6-OHDA-treated rats had higher correlations than saline-treated rats, but
only saline-treated rats exhibited a decrease in correlation on the EPM.
Network Neuroscience
827
Edge-based network analysis of aberrant anxiogenic processing in rats
Correlation Between Informational Flow and PLV Corresponds With Network Changes
During Transitioning
The relationship between informational flow and network connectivity is often overlooked in
network analysis, which neglects the fact that nodes of a neural network do not just merely
connect with one another, but also projects information. By correlating theta dPTE and PLV, we
can understand the dynamics underlying theta band informational transmission and its influ-
ence on network organization. In saline-treated rats, RSC, PtA, and dHip theta outflow to the
mPFC exhibits particularly strong correlations with mPFC theta band connectivity to the rest of
the network, which was further enhanced when the animals were placed on the EPM
(Figure 6A). This indicates that bottom-up informational flow is correlated with the centrality
of the mPFC in the network. For 6-OHDA-treated rats, there is a dramatic reduction in the
correlation between mPFC theta band connectivity with the rest of the network and RSC,
PtA, and dHip theta band outflow to the mPFC. The results correspond to network changes
in transitioning between maze arms, which is defined as the period of time starting when
the head of the rat begins to exit an arm and ends when the base of tail along with the entire
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
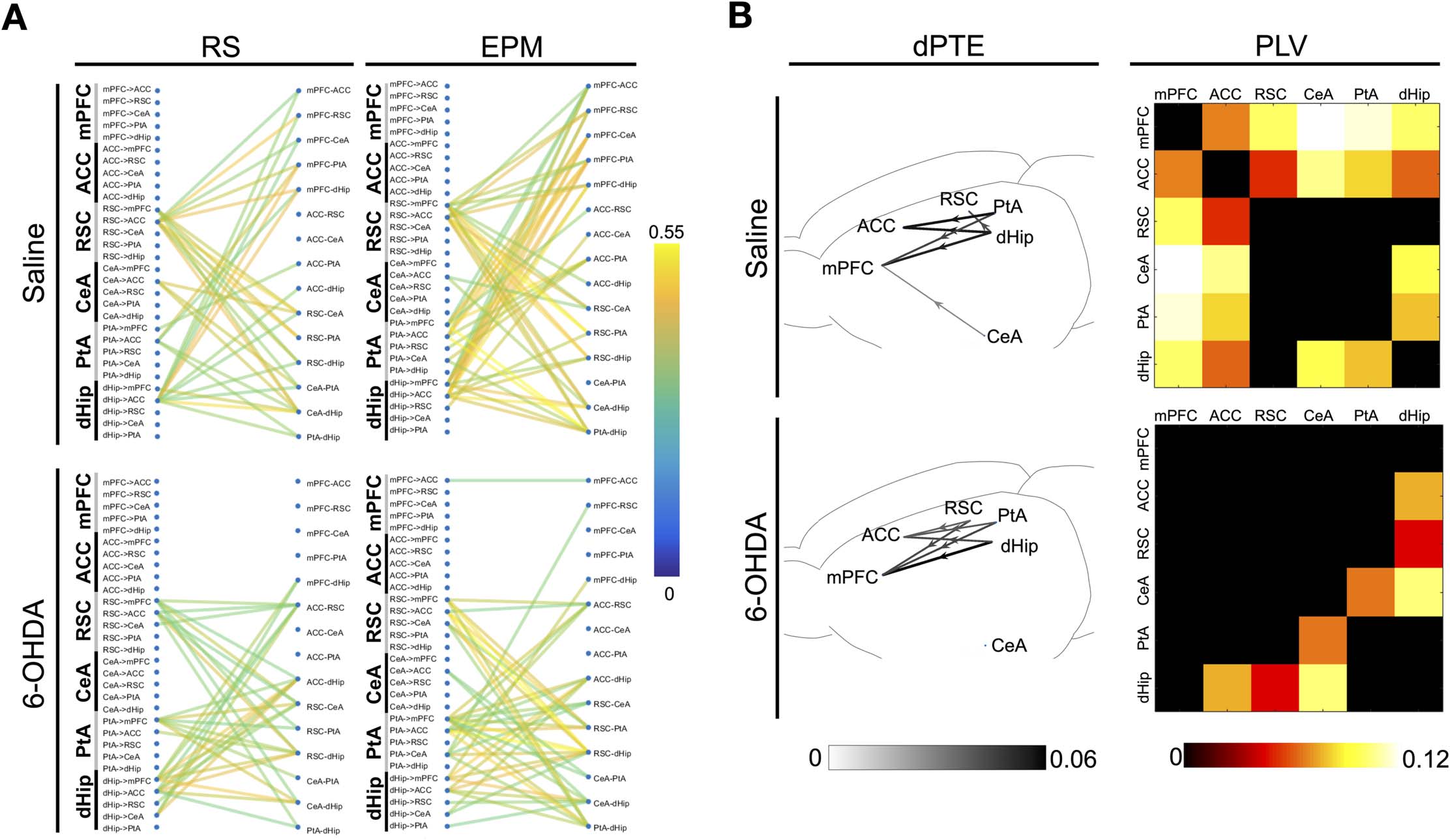
Figure 6. Theta band information flow-connectivity network regulatory motifs and theta band network response during transitioning.
(A) Correlations between fluctuations in theta band dPTE with fluctuations in theta band functional connectivity, revealing functional organi-
zational regulatory motifs across the DSAN (average of n = 240 epochs per state from four rats) in 6-OHDA- and saline-infused rats; correlations
were thresholded at p < 1 × 10−7 (Pearson’s correlation p value). (B) Network schematic unveiling the specific connections, directionality, and
magnitude of the dPTE alterations in the theta band when averaged dPTE values from transitioning (n = 77 epochs from four saline-infused
rats; n = 75 epochs from 4 6-OHDA-infused rats) subtracted from averaged EPM dPTE values (Transition-EPM) (average of n = 240 epochs
per state from four rats) with a threshold of p < 0.01 (two-sample t test) and matrices comparing differences in theta band PLV of transition-
ing epochs (average of n = 77 epochs from four saline-infused rats; n = 75 epochs from 4 6-OHDA-infused rats) versus the average PLV
values on the EPM (Transition-EPM) (average of n = 240 epochs for each state from four rats) in 6-OHDA and saline-infused rats.
Network Neuroscience
828
Edge-based network analysis of aberrant anxiogenic processing in rats
body enters another arm. During transitioning epochs, dPTE changes are also most prominent
in the theta band (Supporting Information Figure S10). As shown in Figure 6B, 6-OHDA- and
saline-treated rats have similar dPTE increases from the dHip and PtA to the mPFC and ACC.
The only difference here is that for saline-treated rats, RSC did not have significant increase in
informational flow toward the mPFC and ACC and even has net input from the dorsal hippo-
campus during transitioning. For functional connectivity, during transitioning, frontal regions
in particular with the mPFC connectivity with the entire network, especially the PtA and CeA,
are enhanced in saline-treated rats; the centrality of the network thus shifts toward the frontal
regions. Such changes are absent for 6-OHDA-treated rats.
DISCUSSION
In this study, we interrogated the changes in DMS-regulated associative network parameters in
response to anxiogenic environments in both focal dopamine-depleted and control rats via
multisite LFP recordings. As the functional divisions of the striatum and other brain network
architecture are largely conserved across mammals, it is reasonable to assume that anxiogenic
processing takes place through an evolutionarily conserved network mechanism (Adhikari,
2014; Kovner et al., 2019; Nikolova et al., 2018). We extended the utilization of the phase-
based metrics, dPTE and PLV, in tracking concomitant functional and informational flow rela-
tionship across multiple network edges, allowing for larger emerging network patterns to be
unveiled. Our study explored the possible role of dopaminergic innervation in the DMS in
regulating anxiogenic behaviours in a novel environment, where artificial depletion via
6-OHDA leads to an increase in avoidance behaviours and decrease in exploratory drive
in an anxiogenic environment. With the hippocampus being a major generator of theta oscil-
lations and being highly responsive to exploratory behaviour (Buzsáki, 2004; Ekstrom et al.,
2005; Whitlock et al., 2008), the major focus of this study was, when compared with saline-
treated rats, whether 6-OHDA-treated rats demonstrated abnormalities in terms of theta band
power and network organization changes between rest and when on the EPM.
First, we showed that there was a consistent network-wide increase in theta peak frequency
on the EPM in both groups. However, the loss of dopamine in DMS leads to a differential
network-wide theta power organization and response, with theta power showing lower vari-
ation, more consistent increases in power on the EPM, and is exaggerated in the posterior hip-
pocampal and parietal regions. Based on analysis of dPTE and PLV measures, the three brain
regions of the posterior medial subnetwork (Cwik et al., 2014; Ritchey & Cooper, 2020), that
is, RSC, PtA, and dHip, exhibit exaggerated theta band outflow when 6-OHDA-treated rats
were transferred to the EPM. The main recipients of these outflows were the mPFC and
ACC and the CeA. The parietal and hippocampal regions were most dominant in the subnet-
work and showed most significant outflow to the amygdala. Theta band functional connectiv-
ity following 6-OHDA infusion further showed that CeA increased its connectivity with the
posterior medial subnetwork, suggesting the amygdala was more functionally incorporated
into the subnetwork. We can hereby see an obvious posterior-anterior divide in terms of both
theta power and theta information flow that was exaggerated under dopaminergic depletion.
The predominance of the theta band in the posterior cortical regions is a well established phe-
nomenon in rats, where the coupling of the hippocampus and posterior midline cortical struc-
tures and neurons, especially during exploration, had been demonstrated in multiple studies
(Colom et al., 1988; Young & McNaughton, 2009). The posterior medial subnetwork has been
well characterized to be involved in sensory information processing such as navigation, mem-
ory, and vividness of emotion representation (Cwik et al., 2014; Ritchey & Cooper, 2020), and
the more anteriorly located mPFC and ACC are higher ordered affective and cognitive centres.
Network Neuroscience
829
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Network regulatory motifs:
Higher order associations
between edges as measured via
co-fluctuations in dynamic
information flow and functional
connectivity.
We speculate in 6-OHDA-infused rats, due to a reduction in frontal regulation, there was an
increase in bottom-up information transfer from the posterior medial subnetwork with a greater
sensory processing role to the higher ordered frontal mPFC/ACC regions and the amygdala.
From the results, we postulate that dopaminergic signalling of the dorsomedial striatum
contributes to long-distance top-down frontal to posterior medial subnetwork mediation of
informational flow. This may allow for higher ordered frontal regions to effectively regulate
contextual and spatial information to prevent an excessive anxiogenic response from environ-
mental stimuli. This notion relates with human studies where frontal midline theta rhythm has
been found to be weakened in PD patients (Singh et al., 2018).
Brain networks, in particular, associative cortical networks, can be characterized not only
in the spatial but also in the temporal domain (Pedersen et al., 2017; Vidaurre et al., 2017).
In this study, the focus on the correlation across multiple network edges enabled us to fur-
ther understand the aberrant theta outflow of the posterior medial subnetwork and identify
abnormal network-wide theta band processing and network regulatory motifs. From dPTE-
dPTE correlations, compared with saline-treated rats, 6-OHDA-treated rats had a hypersyn-
chrony (general increase in correlation) between posterior medial subnetwork outflow pathways
even during resting states that was further exaggerated in anxiogenic environments, forming a
single dominant outflow cluster. One possible interpretation is that in addition to an excessive
posterior medial subnetwork information outflow, dopaminergic depletion of the DMS may
change the subnetwork from providing differentiated and orderly information to becoming
more chaotic and unregulated. Thus, dopaminergic innervation to the DMS may be the key
in delineating different pathway bundles to ensure well-regulated and diverse informational
content across the DSAN.
From the results of correlating dPTE and PLV, in saline-treated rats, the posterior medial
subnetwork outflow to the frontal regions is key to mPFC centrality, which was enhanced in
an anxiogenic environment. The results seem to correspond well with theta band network pro-
cessing during transitioning between maze arms. Saline-treated rats exhibited increased
bottom-up theta band outflow and showed strong increase in mPFC connectivity with the
other components of the DSAN. However, for 6-OHDA treated rats, bottom-up outflow to
the frontal regions was also enhanced but at the same time showed an absence of increased
mPFC connectivity and centrality within the entire DSAN, which corresponded to the loss of
correlation between bottom-up outflow and mPFC centrality. This points toward the impor-
tance of frontal theta engagement in response to bottom-up outflow in initiating transitions
to an open arm. As transitioning is a process where an integration of contextual information
and internal anxiety bridges with decision-making, a robust frontal theta engagement with the
network may be necessary to suppress the intrinsic anxiogenic responses to contextual infor-
mation and generate the motivation to engage in exploratory behaviour. The loss of frontal
network engagement therefore may lead to dysregulation and hypersynchrony of bottom-up
pathways as seen in dPTE-dPTE correlations and hence an exaggerated context-dependent
anxiogenic response.
Previous studies have shown that especially band-passed LFP time series are autocorrelated
across time (Afyouni et al., 2019), which can lead to inflated connectivity values due to under-
sampling of state space. However, as our main results are not derived from the raw PLV and
dPTE values but from changes in connectivity values across different conditions and correlat-
ing the fluctuations of said connectivity values against each other, this effect is unlikely to be a
major confounder of the conclusions of this study. Nonetheless, given the relatively small sam-
ple size in this study, the conclusion obtained here warrants further investigation for confirma-
tion and extension.
Network Neuroscience
830
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Network dynamics:
The characterization of fluctuations
in network properties, in particular
edge properties or node properties,
temporally.
In conclusion, by integrating edge-based frequency-specific network dynamics with gross
network and regional changes, we showed that a dorsomedial striatal-tied associative network
in rats may be associated with the regulation of exploratory and avoidance behaviours in an
anxiogenic context via theta band–mediated information transfer. This suggests that behav-
ioural changes associated with anxiety may arise from degenerative neural disorders via spa-
tial and temporal changes in network signalling. This approach can be further applied to other
cognitive disease models to unearth the principles behind pathological behavioural alterations
and network regulatory mechanisms.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00251.
AUTHOR CONTRIBUTIONS
Yin-Shing Lam: Conceptualization; Data curation; Formal analysis; Investigation; Methodol-
ogy; Visualization; Writing – original draft; Writing – review & editing. Xiu-Xiu Liu: Formal
analysis; Methodology. Ya Ke: Conceptualization; Investigation; Methodology; Project admin-
istration; Supervision; Writing – review & editing. Wing-Ho Yung: Conceptualization; Investi-
gation; Methodology; Project administration; Supervision; Writing – review & editing.
FUNDING INFORMATION
Wing-Ho Yung, Hong Kong Research Grants Council Theme-Based Research Scheme
(T13-605/18-W). Ya Ke, Hong Kong Health and Medical Research Fund (07180906).
REFERENCES
Adhikari, A. (2014). Distributed circuits underlying anxiety. Fron-
tiers in Behavioral Neuroscience, 8, 112. https://doi.org/10
.3389/fnbeh.2014.00112, PubMed: 24744710
Afyouni, S., Smith, S. M., & Nichols, T. E. (2019). Effective degrees
of freedom of the Pearson’s correlation coefficient under autocor-
relation. NeuroImage, 199, 609–625. https://doi.org/10.1016/j
.neuroimage.2019.05.011, PubMed: 31158478
Alvarez-Fischer, D., Henze, C., Strenzke, C., Westrich, J., Ferger, B.,
Höglinger, G. U., Oertel, W. H., & Hartmann, A. (2008). Charac-
terization of the striatal 6-OHDA model of Parkinson’s disease in
wild type and α-synuclein-deleted mice. Experimental Neurol-
ogy, 210(1), 182–193. https://doi.org/10.1016/j.expneurol.2007
.10.012, PubMed: 18053987
Aydore, S., Pantazis, D., & Leahy, R. M. (2013). A note on the phase
locking value and its properties. NeuroImage, 74, 231–244.
https://doi.org/10.1016/j.neuroimage.2013.02.008, PubMed:
23435210
Barbot, A., & Carrasco, M. (2018). Emotion and anxiety potentiate
the way attention alters visual appearance. Scientific Reports, 8(1),
5938. https://doi.org/10.1038/s41598-018-23686-8, PubMed:
29651048
Bandelow, B., & Michaelis, S. (2015). Epidemiology of anxiety dis-
orders in the 21st century. Dialogues in Clinical Neuroscience,
17(3), 327–335. https://doi.org/10.31887/ DCNS.2015.17.3
/bbandelow, PubMed: 26487813
Bassett, D. S., & Bullmore, E. T. (2009). Human brain networks
in health and disease. Current Opinion in Neurology, 22(4),
340–347. https://doi.org/10.1097/ WCO.0b013e32832d93dd,
PubMed: 19494774
Becerra, L., Pendse, G., Chang, P.-C., Bishop, J., & Borsook, D.
(2011). Robust reproducible resting state networks in the awake
rodent brain. PLoS One, 6(10), e25701. https://doi.org/10.1371
/journal.pone.0025701, PubMed: 22028788
Buzsáki, G. (2004). Neuronal oscillations in cortical networks. Sci-
ence, 304(5679), 1926–1929. https://doi.org/10.1126/science
.1099745, PubMed: 15218136
Buzsáki, G., Logothetis, N., & Singer, W. (2013). Scaling brain size,
keeping timing: Evolutionary preservation of brain rhythms.
Neuron, 80(3), 751–764. https://doi.org/10.1016/j.neuron.2013
.10.002, PubMed: 24183025
Calhoun, V. D., Miller, R., Pearlson, G., & Adali, T. (2014). The
chronnectome: Time-varying connectivity networks as the next
frontier in fMRI data discovery. Neuron, 84(2), 262–274. https://
doi.org/10.1016/j.neuron.2014.10.015, PubMed: 25374354
Cavanagh, J. F., & Frank, M. J. (2014). Frontal theta as a mechanism for
cognitive control. Trends in Cognitive Sciences, 18(8), 414–421.
https://doi.org/10.1016/j.tics.2014.04.012, PubMed: 24835663
Cavanagh, J. F., Mueller, A. A., Brown, D. R., Janowich, J. R., Story-
Remer, J. H., Wegele, A., & Richardson, S. P. (2017). Cognitive
states influence dopamine-driven aberrant learning in Parkinson’s
disease. Cortex, 90, 115–124. https://doi.org/10.1016/j.cortex
.2017.02.021, PubMed: 28384481
Chen, J. J., & Marsh, L. (2014). Anxiety in Parkinson’s disease:
Identification and management. Therapeutic Advances in
Network Neuroscience
831
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Neurological Disorders, 7(1), 52–59. https://doi.org/10.1177
/1756285613495723, PubMed: 24409202
Colom, L. V., Christie, B. R., & Bland, B. H. (1988). Cingulate cell
discharge patterns related to hippocampal EEG and their modu-
lation by muscarinic and nicotinic agents. Brain Research, 460(2),
329–338. https://doi.org/10.1016/0006-8993(88)90377-0,
PubMed: 3224265
Cwik, J. C., Sartory, G., Schürholt, B., Knuppertz, H., & Seitz, R. J.
(2014). Posterior midline activation during symptom provocation
in acute stress disorder: An fMRI study. Frontiers in Psychiatry, 5, 49.
https://doi.org/10.3389/fpsyt.2014.00049, PubMed: 24847285
Delcasso, S., Huh, N., Byeon, J. S., Lee, J., Jung, M. W., & Lee, I.
(2014). Functional relationships between the hippocampus and
dorsomedial striatum in learning a visual scene-based memory
task in rats. Journal of Neuroscience, 34(47), 15534–15547.
https://doi.org/10.1523/ JNEUROSCI.0622-14.2014, PubMed:
25411483
Ekstrom, A. D., Caplan, J. B., Ho, E., Shattuck, K., Fried, I., &
Kahana, M. J. (2005). Human hippocampal theta activity during
virtual navigation. Hippocampus, 15(7), 881–889. https://doi.org
/10.1002/hipo.20109, PubMed: 16114040
Engels, M. M. A., Yu, M., Stam, C. J., Gouw, A. A., van der Flier,
W. M., Scheltens, P., van Straaten, E. C. W., & Hillebrand, A.
(2017). Directional information flow in patients with Alzheimer’s
disease. A source-space resting-state MEG study. NeuroImage:
Clinical, 15, 673–681. https://doi.org/10.1016/j.nicl.2017.06
.025, PubMed: 28702344
Erro, R., Pappatà, S., Amboni, M., Vicidomini, C., Longo, K.,
Santangelo, G., Picillo, M., Vitale, C., Moccia, M., Giordano,
F., Brunetti, A., Pellecchia, M. T., Salvatore, M., & Barone, P.
(2012). Anxiety is associated with striatal dopamine transporter
availability in newly diagnosed untreated Parkinson’s disease
patients. Parkinsonism & Related Disorders, 18(9), 1034–1038.
https://doi.org/10.1016/j.parkreldis.2012.05.022, PubMed:
22789824
Esfahlani, F. Z., Jo, Y., Faskowitz, J., Byrge, L., Kennedy, D. P.,
Sporns, O., & Betzel, R. F. (2020). High-amplitude cofluctuations
in cortical activity drive functional connectivity. Proceedings of
the National Academy of Sciences, 117(45), 28393–28401.
https://doi.org/10.1073/pnas.2005531117, PubMed: 33093200
Faskowitz, J., Esfahlani, F. Z., Jo, Y., Sporns, O., & Betzel, R. F. (2020).
Edge-centric functional network representations of human cere-
bral cortex reveal overlapping system-level architecture. Nature
Neuroscience, 23(12), 1644–1654. https://doi.org/10.1038
/s41593-020-00719-y, PubMed: 33077948
Fraschini, M., Demuru, M., Crobe, A., Marrosu, F., Stam, C. J., &
Hillebrand, A. (2016). The effect of epoch length on estimated
EEG functional connectivity and brain network organisation.
Journal of Neural Engineering, 13(3), 036015. https://doi.org/10
.1088/1741-2560/13/3/036015, PubMed: 27137952
Fraschini, M., & Hillebrand, A. (2016). “brainstorm-tools” [MATLAB
scripts]. https://github.com/brainstorm-tools/brainstorm3/
Genzel, L. (2020). Memory and sleep: Brain networks, cell dynam-
ics and global states. Current Opinion in Behavioral Sciences, 32,
72–79. https://doi.org/10.1016/j.cobeha.2020.02.003
Glimcher, P. W., & Fehr, E. (2014). Neuroeconomics: Decision
making and the brain. Amsterdam, the Netherlands: Elsevier Aca-
demic Press. https://doi.org/10.1016/C2011-0-05512-6
Greene, D. J., Marek, S., Gordon, E. M., Siegel, J. S., Gratton, C.,
Laumann, T. O., … Dosenbach, N. U. F. (2019). Integrative and
network-specific connectivity of the basal ganglia and thalamus
defined in individuals. Neuron, 105(4), 742–758. https://doi.org
/10.1016/j.neuron.2019.11.012, PubMed: 31836321
Gunaydin, L., Nelson, A., & Kreitzer, A. (2017). Fronto-striatal mod-
ulation of anxiety-like behaviors. Biological Psychiatry, 81(10),
S203. https://doi.org/10.1016/j.biopsych.2017.02.1108
Hilbert, K., Pine, D. S., Muehlhan, M., Lueken, U., Steudte-
Schmiedgen, S., & Beesdo-Baum, K. (2015). Gray and white
matter volume abnormalities in generalized anxiety disorder
by categorical and dimensional characterization. Psychiatry
Research: Neuroimaging, 234(3), 314–320. https://doi.org/10
.1016/j.pscychresns.2015.10.009, PubMed: 26490569
Hunnicutt, B. J., Jongbloets, B. C., Birdsong, W. T., Gertz, K. J.,
Zhong, H., & Mao, T. (2016). A comprehensive excitatory input
map of the striatum reveals novel functional organization. ELife,
5, e19103. https://doi.org/10.7554/eLife.19103, PubMed:
27892854
Jacinto, L. R., Reis, J. S., Dias, N. S., Cerqueira, J. J., Correia, J. H., &
Sousa, N. (2013). Stress affects theta activity in limbic networks
and impairs novelty-induced exploration and familiarization.
Frontiers in Behavioral Neuroscience, 7, 127. https://doi.org/10
.3389/fnbeh.2013.00127, PubMed: 24137113
Kovner, R., Oler, J. A., & Kalin, N. H. (2019). Cortico-limbic
interactions mediate adaptive and maladaptive responses relevant
to psychopathology. American Journal of Psychiatry, 176(12),
987–999. https://doi.org/10.1176/appi.ajp.2019.19101064,
PubMed: 31787014
LeDoux, J. E. (2000). Emotion circuits in the brain. Annual Review
of Neuroscience, 23(1), 155–184. https://doi.org/10.1146
/annurev.neuro.23.1.155, PubMed: 10845062
Lobier, M., Siebenhühner, F., Palva, S., & Palva, J. M. (2014). Phase
transfer entropy: A novel phase-based measure for directed con-
nectivity in networks coupled by oscillatory interactions. Neuro-
Image, 85, 853–872. https://doi.org/10.1016/j.neuroimage.2013
.08.056, PubMed: 24007803
Loewke, A. C., Minerva, A. R., Nelson, A. B., Kreitzer, A. C., &
Gunaydin, L. A. (2020). Fronto-striatal projections regulate
approach-avoidance conflict. bioRxiv. https://doi.org/10.1101
/2020.03.05.979708
Martin, E. I., Ressler, K. J., Binder, E., & Nemeroff, C. B. (2009). The
neurobiology of anxiety disorders: Brain imaging, genetics, and
psychoneuroendocrinology. Psychiatric Clinics of North Amer-
ica, 32(3), 549–575. https://doi.org/10.1016/j.psc.2009.05.004,
PubMed: 19716990
Meireles, J., & Massano, J. (2012). Cognitive impairment and
dementia in Parkinson’s disease: Clinical features, diagnosis,
and management. Frontiers in Neurology, 3, 88. https://doi.org
/10.3389/fneur.2012.00088, PubMed: 22654785
Müller, E. J., Munn, B. R., & Shine, J. M. (2020). Diffuse neural cou-
pling mediates complex network dynamics through the formation
of quasi-critical brain states. Nature Communications, 11(1), 6337.
https://doi.org/10.1038/s41467-020-19716-7, PubMed: 33303766
Navarro-Lobato, I., & Genzel, L. (2020). Anterior to posterior
whole-brain gradient for different types of memories? Trends in
Neurosciences, 43(7), 451–453. https://doi.org/10.1016/j.tins
.2020.04.011, PubMed: 32409016
Network Neuroscience
832
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Edge-based network analysis of aberrant anxiogenic processing in rats
Nguyen, D., Alushaj, E., Erb, S., & Ito, R. (2019). Dissociative
effects of dorsomedial striatum D1 and D2 receptor antagonism
in the regulation of anxiety and learned approach-avoidance
conflict decision-making. Neuropharmacology, 146, 222–230.
https://doi.org/10.1016/j.neuropharm.2018.11.040, PubMed:
30508508
Nikolova, Y. S., Misquitta, K. A., Rocco, B. R., Prevot, T. D., Knodt,
A. R., Ellegood, J., Voineskos, A. N., Lerch, J. P., Hariri, A. R.,
Sibille, E., & Banasr, M. (2018). Shifting priorities: Highly con-
served behavioral and brain network adaptations to chronic
stress across species. Translational Psychiatry, 8(1), 26. https://
doi.org/10.1038/s41398-017-0083-5, PubMed: 29353879
Pedersen, M., Omidvarnia, A., Walz, J. M., Zalesky, A., & Jackson,
G. D. (2017). Spontaneous brain network activity: Analysis of its
temporal complexity. Network Neuroscience, 1(2), 100–115.
https://doi.org/10.1162/NETN_a_00006, PubMed: 29911666
Pontone, G. M., Williams, J. R., Anderson, K. E., Chase, G., Goldstein,
S. A., Grill, S., Hirsch, E. S., Lehmann, S., Little, J. T., Margolis,
R. L., Rabins, P. V., Weiss, H. D., & Marsh, L. (2009). Prevalence
of anxiety disorders and anxiety subtypes in patients with
Parkinson’s disease. Movement Disorders, 24(9), 1333–1338.
https://doi.org/10.1002/mds.22611, PubMed: 19425086
Ritchey, M., & Cooper, R. A. (2020). Deconstructing the posterior
medial episodic network. Trends in Cognitive Sciences, 24(6),
451–465. https://doi.org/10.1016/j.tics.2020.03.006, PubMed:
32340798
Robinson, S. E., Mandell, A. J., & Coppola, R. (2013). Spatiotempo-
ral imaging of complexity. Frontiers in Computational Neurosci-
ence, 6, 101. https://doi.org/10.3389/fncom.2012.00101,
PubMed: 23355820
Scott, D. W. (1992). Multivariate density estimation: Theory, prac-
tice, and visualization (1st ed.). Hoboken, NJ: Wiley. https://doi
.org/10.1002/9780470316849
Seo, D., Patrick, C. J., & Kennealy, P. J. (2008). Role of serotonin
and dopamine system interactions in the neurobiology of impul-
sive aggression and its comorbidity with other clinical disorders.
Aggression and Violent Behavior, 13(5), 383–395. https://doi.org
/10.1016/j.avb.2008.06.003, PubMed: 19802333
Shine, J. M., Koyejo, O., Bell, P. T., Gorgolewski, K. J., Gilat, M., &
Poldrack, R. A. (2015). Estimation of dynamic functional connec-
tivity using multiplication of temporal derivatives. NeuroImage,
122, 399–407. https://doi.org/10.1016/j.neuroimage.2015.07
.064, PubMed: 26231247
Siems, M., & Siegel, M. (2020). Dissociated neuronal phase- and
amplitude-coupling patterns in the human brain. NeuroImage,
209, 116538. https://doi.org/10.1016/j.neuroimage.2020
.116538, PubMed: 31935522
Singh, A., Richardson, S. P., Narayanan, N., & Cavanagh, J. F.
(2018). Mid-frontal theta activity is diminished during cognitive
control in Parkinson’s disease. Neuropsychologia, 117, 113–122.
https://doi.org/10.1016/j.neuropsychologia.2018.05.020,
PubMed: 29802866
Soltani Zangbar, H., Ghadiri, T., Seyedi Vafaee, M., Ebrahimi Kalan,
A., Fallahi, S., Ghorbani, M., & Shahabi, P. (2020). Theta oscilla-
tions through hippocampal/prefrontal pathway: Importance in
cognitive performances. Brain Connectivity, 10(4), 157–169.
https://doi.org/10.1089/brain.2019.0733, PubMed: 32264690
Spielberg, J. M., Miller, G. A., Warren, S. L., Engels, A. S., Crocker,
L. D., Banich, M. T., Sutton, B. P., & Heller, W. (2012). A
brain network instantiating approach and avoidance motivation:
Network instantiating motivation. Psychophysiology, 49(9),
1200–1214. https://doi.org/10.1111/j.1469-8986.2012.01443.x,
PubMed: 22845892
Steimer, T. (2002). The biology of fear- and anxiety-related behav-
iors. Dialogues in Clinical Neuroscience, 4(3), 231–249. https://
doi.org/10.31887/DCNS.2002.4.3/tsteimer, PubMed: 22033741
Sussman, T. J., Jin, J., & Mohanty, A. (2016). Top-down and
bottom-up factors in threat-related perception and attention in
anxiety. Biological Psychology, 121, 160–172. https://doi.org/10
.1016/j.biopsycho.2016.08.006, PubMed: 27546616
Vidaurre, D., Smith, S. M., & Woolrich, M. W. (2017). Brain network
dynamics are hierarchically organized in time. Proceedings of the
National Academy of Sciences, 114(48), 12827–12832. https://doi
.org/10.1073/pnas.1705120114, PubMed: 29087305
Walf, A. A., & Frye, C. A. (2007). The use of the elevated plus maze
as an assay of anxiety-related behavior in rodents. Nature Proto-
cols, 2(2), 322–328. https://doi.org/10.1038/nprot.2007.44,
PubMed: 17406592
Westbrook, A., & Braver, T. S. (2016). Dopamine does double duty
in motivating cognitive effort. Neuron, 89(4), 695–710. https://
doi.org/10.1016/j.neuron.2015.12.029, PubMed: 26889810
Whitlock, J. R., Sutherland, R. J., Witter, M. P., Moser, M.-B., &
Moser, E. I. (2008). Navigating from hippocampus to parietal cor-
tex. Proceedings of the National Academy of Sciences, 105(39),
14755–14762. https://doi.org/10.1073/pnas.0804216105,
PubMed: 18812502
Young, C. K., & McNaughton, N. (2009). Coupling of theta oscil-
lations between anterior and posterior midline cortex and with the
hippocampus in freely behaving rats. Cerebral Cortex, 19(1), 24–40.
https://doi.org/10.1093/cercor/bhn055, PubMed: 18453538
Network Neuroscience
833
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
8
1
6
2
0
3
5
9
9
2
n
e
n
_
a
_
0
0
2
5
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3