Repetitive Transcranial Magnetic Stimulation
Dissociates Working Memory Manipulation
from Retention Functions in the Prefrontal,
but not Posterior Parietal, Kortex
Bradley R. Postle, Fabio Ferrarelli, Massihullah Hamidi,
Eva Feredoes, Marcello Massimini, Michael Peterson,
Andrew Alexander, and Giulio Tononi
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
0
P
1
D
7
F
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
Ö
2
C
/
N
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
J
0
Ö
1
C
7
N
1
.
2
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
0
T
.
Ö
1
N
7
0
1
7
2
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
F
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Abstrakt
& Understanding the contributions of the prefrontal cortex
(PFC) to working memory is central to understanding the neu-
ral bases of high-level cognition. One question that remains
controversial is whether the same areas of the dorsolateral PFC
(dlPFC) that participate in the manipulation of information in
working memory also contribute to its short-term retention
(STR). We evaluated this question by first identifying, mit
functional magnetic resonance imaging (fMRT), brain areas
involved in manipulation. Nächste, these areas were targeted with
repetitive transcranial magnetic stimulation (rTMS) while sub-
jects performed tasks requiring only the STR or the STR
plus manipulation of information in working memory. fMRT
indicated that manipulation-related activity was independent
of retention-related activity in both the PFC and superior
parietal
lobule (SPL). rTMS, Jedoch, yielded a different
pattern of results. Although rTMS of the dlPFC selectively
disrupted manipulation, rTMS of the SPL disrupted manipu-
lation and STR to the same extent. rTMS of the postcentral
gyrus (a control region) had no effect on performance. Der
implications of these results are twofold. In the PFC, they are
consistent with the view that this region contributes more
importantly to the control of information in working memory
than to its STR. In the SPL, they illustrate the importance
of supplementing the fundamentally correlational data from
neuroimaging with a disruptive method, which affords stron-
ger inference about structure–function relations. &
EINFÜHRUNG
One perspective on the role of the prefrontal cortex (PFC)
is that it supports short-term retention (STR, alternatively
referred to as ‘‘storage’’ or ‘‘maintenance’’) functions (z.B.,
Courtney, 2004; Leung, Seelig, & Gore, 2004; Goldman-
Rakic & Leung, 2002; Pessoa, Gutierrez, Bandettini, &
Ungerleider, 2002; Constantinides, Franowicz, & Goldman-
Rakic, 2001; Funahashi, Chafee, & Goldman-Rakic, 1993),
and can thus be viewed as supporting the buffers of
a multiple-component working memory system such as
that proposed by Baddeley (2000) and Baddeley and Logie
(1999). An alternative perspective denies an important role
in STR for the PFC (z.B., Lebedev, Messinger, Kralik, &
Wise, 2004; Petride, 2000; D'Esposito & Postle, 1999),
ascribing these functions instead to activity in the non-
PFC regions that have evolved to accomplish sensory-,
representation-, and action-related functions (z.B., Postle,
2006; Jonides, Lacey, & Nee, 2005; Pasternak & Greenlee,
2005; Theeuwes, Olivers, & Chizk, 2005). One implica-
tion of the latter view is that working memory may be
University of Wisconsin
better understood as an emergent property of the mind
and brain, rather than as a system that can be readily
localized (as can, z.B., vision to the occipital cortex or
skeletomotor control to the precentral gyrus).
One way to operationalize working memory retention
processes is by varying the number of items that must
be retained on different trials—brain regions whose
delay-period activity is sensitive to such variations in
memory load are presumed to contribute to retention-
related processes. To date, the functional magnetic re-
sonance imaging (fMRT) literature on the load sensitivity
of the dorsolateral PFC (dlPFC) delay period activity
for verbal stimuli has been mixed, with some studies
finding evidence for (Narayanan et al., 2005; Zarahn,
Rakitin, Abela, Flynn, & Stern, 2005; Veltman, Rombouts,
& Dolan, 2003), and some finding evidence against
(Feredoes & Postle, 2005; Postle, Berger, & D'Esposito,
1999; Rypma & D'Esposito, 1999). The same study by
Postle et al. (1999) also reported evidence that some
load-insensitive regions of the dlPFC were nonetheless
sensitive to the requirement to reorder the contents
of working memory during the delay period—an oper-
im Gegenzug,
ationalization of manipulation. These results,
D 2006 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 18:10, S. 1712–1722
have been challenged by Veltman et al. (2003), WHO
reported that ‘‘maintenance’’ (what we refer to in this
article as retention) and manipulation ‘‘activate virtually
identical systems’’ (P. 247). Clearly, the neuroimaging
literature related to this issue is at an impasse.
The present study was intended to address the spe-
cific question of whether STR and manipulation func-
tions colocalize in the dlPFC and to do so with a method
that would support stronger inference than have the
neuroimaging studies performed to date. This approach
entailed supplementing fMRI with repetitive transcranial
magnetic stimulation (rTMS), a method that produces
the temporary disruption of the function of a local re-
gion of cortex. Although fMRI data are limited to testing
hypotheses about brain–behavior correlations, rTMS can
address hypotheses about the necessity of a brain area
to a particular aspect of cognitive performance ( Walsh
& Pascual-Leone, 2003). An additional goal of this study
was to broaden the investigation of manipulation in
working memory beyond the PFC, to reflect the general
consensus that most examples of high-level cognition, In-
cluding the control of working memory, are supported
by broadly distributed networks that extend beyond this
one brain region. Our two-step procedure entailed first,
acquiring fMRI data while subjects performed delayed
recognition with different loads and different manip-
ulation requirements, und zweitens, delivering rTMS to
fMRI-identified areas of the dlPFC and superior parietal
lobule (SPL) with rTMS while the same subjects per-
formed the same task. (The SPL has also been implicated
in executive control, z.B., Garavan, Ross, Li, & Stein, 2000,
and has shown manipulation-related activity in previous
Studien, unpublished observation). We predicted that
delay-period rTMS of manipulation-sensitive regions of
the dlPFC and SPL would produce selective deficits on
trials requiring the manipulation, but not the simple re-
Aufmerksamkeit, of items in working memory.
METHODEN
Subjects
Der 12 Erwachsene (6 men and 6 Frauen; Durchschnittsalter = 22 Jahre,
SD = 2.7) whose data are presented here had no psy-
chiatric or neurologic disorders, as determined by phys-
ical examination, a structured psychiatric diagnostic
screening interview (Mini-International Neuropsychiatric
Interview; Sheehan et al., 1998), and a mood assessment
(Hamilton Depression Rating Scale; Hamilton, 1960), alle
administered by a psychiatrist.
Behavioral Task
The task, requiring delayed recognition of item position,
was identical to that used by Postle et al. (1999). Jede
trial began with the simultaneous presentation of two or
five consonant letters (all in a single row), followed by
instructions (‘‘forward’’ or ‘‘alphabetize’’), followed by
an 8-sec delay period, followed by a memory probe
comprising an item from the memory set and a digit.
On forward trials, subjects were to retain a memory of
the two or five letters in the order in which they were
vorgeführt. On these trials, the probe digit represented
(with p = .5) the ordinal position in which the probed
letter had appeared in the initial stimulus display. An
alphabetize trials, subjects were to reorder the letters
into alphabetical order. On these trials, the probe digit
represented (also with p = .5) the alphabetical position
of the probed letter with respect to the other four letters
in the memory set.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
0
P
1
D
7
F
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
Ö
2
C
/
N
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
J
0
Ö
1
C
7
N
1
.
2
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
0
T
.
Ö
1
N
7
0
1
7
2
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
F
.
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Functional Magnetic Resonance Imaging
Data Acquisition and Preprocessing
Whole-brain images were acquired with a 3-T scanner (GE
Signa VH/I, Waukesha, WI). High-resolution T1-weighted
Bilder (30 axial slices, 0.9375 (cid:1) 0.9375 (cid:1) 4 mm) were ob-
tained in all participants, and a gradient-echo, echoplanar
sequence (TR = 2000 ms, TE = 50 ms) was used to
acquire data sensitive to the blood oxygen level depen-
dent (BOLD) signal (Kwong et al., 1992; Ogawa et al.,
1992) within a 64 (cid:1) 64 Matrix (30 axial slices coplanar
with the T1 acquisition, 3.75 (cid:1) 3.75 (cid:1) 4 mm, no skip).
Scans of the delayed-recognition task were preceded by
a scan in which we derived an estimate of the hemody-
namic response function (HRF) für jeden Teilnehmer.
During this scan, each participant performed a simple
reaction-time task that required a bimanual button press
once every 20 sec in response to a brief change in shape
of the fixation stimulus. A partial F test associated with a
Fourier basis covariate set ( Josephs, Turner, & Friston,
1997) was used to evaluate the significance of task-
correlated activity in each voxel of primary somatosensory
and motor cortical regions of interest (ROIs). An HRF
estimate was extracted from the suprathreshold voxels
of these ROIs by spatially averaging their time series, fil-
tering the resultant averaged fMRI time series to remove
hoch (> 0.244 Hz) and low (< 0.05 Hz) frequencies,
adjusting it to remove the effects of nuisance covari-
ates (Friston, Holmes, Poline, Heather, & Frackowiak,
1995), and trial averaging. The HRF characterizes the
fMRI response resulting from a brief impulse of neural
activity (Boynton, Engel, Glover, & Heeger, 1996) and can
vary markedly across subjects (Handwerker, Ollinger, &
D’Esposito, 2004; Aguirre, Zarahn, & D’Esposito, 1998).
The subject-specific HRFs were used to convolve inde-
pendent variables entered into the modified general
linear model (GLM; Worsley & Friston, 1995) that we
used to analyze the data from the scans of the working
memory task. The eight scans of the working memory
task each lasted 6 min 20 sec (6 min of task preceded by
20 sec of dummy pulses to achieve a steady state of tissue
magnetization).
Postle et al.
1713
Analyses
The fMRI time series analysis modeled the signal change
associated with each discrete epoch of the trial with a
covariate comprised of a BOLD HRF shifted along the
timeline of the task in order best model the trial epoch
in question (Postle, Zarahn, & D’Esposito, 2000; Zarahn,
Aguirre, & D’Esposito, 1997). The least-squares solu-
tion of the GLM of the fMRI time series data yielded
parameter estimates that were associated with each co-
variate of interest. The smoothness of the fMRI response
to neural activity allows fMRI evoked responses that
arise from temporally dependent events to be resolved
on the order of 4 sec (Zarahn et al., 1997). Load-sensitive
and alphabetization-sensitive voxels were identified
with the contrasts [DelayForward 5–DelayForward 2] and
[DelayAlphabetize 5–DelayForward 5], respectively, thresh-
olded at a mapwise level of p = .05, Bonferroni-corrected
for multiple comparisons.
Because the principal function of the fMRI data for
this study was to provide activation maps that would
guide the rTMS, the principal analyses were performed
as single-subject analyses. (This first step was a precise
replication of the procedure from Postle et al., 1999.)
Transforming a subject’s anatomical and functional data
into a ‘‘normalized’’ atlas space would not be appropri-
ate with this approach for the simple reason that rTMS
can only be applied to a subject’s brain in its ‘‘native’’
configuration (i.e., it is not possible to apply rTMS to a
composite, group-normalized statistical volume; for a
similar approach, see the work of Herwig et al., 2003).
Before the rTMS session, the whole-brain alphabetiza-
tion statistical map was coregistered and merged with a
high-resolution T1-weighted anatomical scan. The three-
dimensional reconstruction of this merged image would
be used to guide rTMS.
To provide a sense of aggregate trends in activity
produced by our task, we also performed a spatial
normalization-based group analysis. This analysis was
performed by first warping unthresholded statistical
volumes from each subject to a template in MNI space,
smoothing them to 8-mm full width half maximum, then
evaluating the reliability of these statistical maps across
subjects with a ‘‘second-level’’ analysis implemented
with a GLM that treated subject as a random variable.
The resultant group maps were thresholded at p = .01,
uncorrected for multiple comparisons.
Transcranial Magnetic Stimulation
Procedure
The behavioral task used in the rTMS session only in-
cluded Alphabetize 5 and Forward 5 trials that occurred
with equal probability in a randomly determined or-
der. An intertrial interval of 10 sec separated each trial.1
Orthogonal to the factor of instructions was that of rTMS
(present, absent; each also occurring randomly with p =
.5). An entire rTMS study comprised 12 twelve-trial
blocks, with four consecutive blocks performed for each
stimulation site: middle frontal gyrus (MFG) of the dlPFC,
SPL, and postcentral gyrus (PCG). Order of stimulation
site was counterbalanced across subjects. Within each
block, the orthogonal factors of instructions (alphabetize,
forward) and rTMS (present, absent) were randomized
such that each trial type occurred three times during each
block. Probe validity also varied independently of the two
principal factors of interest, such that an equal number of
valid and invalid probes occurred during each block, no
more than two trials of any type (e.g., an ‘‘alphabetize,
rTMS absent’’ trial) featured a valid probe within a single
block, and an equal number of valid and invalid probes
(i.e., three) had occurred for each trial type upon com-
pletion of each even-numbered block.
Apparatus
TMS was delivered with a Magstim Standard Rapid mag-
netic stimulator fit with a 70-mm figure-8 stimulating coil
(Magstim, Whitland, Wales, UK). The first step of the
TMS session was to determine the minimal intensity at
which a single pulse through the TMS coil, positioned
over the motor cortex, reliably produced a motor-evoked
potential of (cid:2)50 AV in the abductor pollicis brevis in 5
of 10 successive stimuli. This ‘‘motor threshold’’ was the
intensity at which the subsequent rTMS was performed.2
As stated in the Introduction section, the logic of
the experiment was to target portions of the dlPFC
and SPL that showed alphabetization sensitivity (i.e.,
DelayAlphabetize 5 > DelayForward 5) in the fMRI scan. Das
was accomplished via coregistration, with infrared-based
frameless stereotaxy (eXimia Navigated Brain Stimulation
[NBS]; Nexstim, Helsinki, Finland), of the subject’s head
with his/her MRI data. The TMS coil was also fitted with
infrared-reflecting beacons, thereby permitting us to tar-
get regions identified in the fMRI data with rTMS. NBS
works from the understanding that TMS preferentially
stimulates neurons located in the area where the induced
current is strongest (Thielscher & Kammer, 2002). Der
system displays the cortical area likely to be maximally
stimulated by TMS by displaying the electric field maxi-
mum in the cortex after calculating the estimated distri-
bution and strength of the intracranial electric field. Das
computation takes into account the exact shape of the
copper wiring inside the TMS coil, the three-dimensional
position and orientation of the coil, and the overall shape
of the head and the brain.
Target Selection
In each subject, only one site was stimulated in each of
three regions: dlPFC, SPL, and a control site in the PCG.
We opted to target the ‘‘hotspots’’ from each individual
subject’s fMRI data, rather than areas defined by compos-
ite, group-averaged statistical maps, because of growing
1714
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
0
P
1
D
7
F
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
Ö
2
C
/
N
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
J
0
Ö
1
C
7
N
1
.
2
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
0
T
.
Ö
1
N
7
0
1
7
2
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
F
.
.
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
evidence for high levels of intersubject topographical
variability in many domains of cognition (z.B., Swallow,
Mutiger, Snyder, Speer, & Zacks, 2003; Tsao, Freiwald,
Knutsen, Mandeville, & Tootell, 2003; Miller et al., 2002),
including the STR of information (Feredoes & Postle,
2005), but relatively lower intrasubject variability over time
(d.h., good test–retest reliability, Feredoes & Postle, 2005;
Peeling & Downing, 2005; Tsao et al., 2003; Miller et al.,
2002). (One manifestation of high intersubject topograph-
ic variability in the present data set was that the mean
alphabetization effect size from the dlPFC was an order
of magnitude larger in the single-subject analyses, 2% sig-
nal change, Figur 1, than in the group-averaged analysis,
0.33 % signal change, averaged across the two hemi-
spheres; Tisch 1.) Based on this, it may be that a single-
subject statistical map provides a better estimate of the
true anatomical location of task-related activity for that
subject than would a map derived from a group average.
(If it were the case that we were applying rTMS to sub-
jects for whom we did not have fMRI data, Jedoch, unser
best estimate would come from group-averaged data.)
In some instances, there were multiple foci of alpha-
betization-sensitive activity in a particular region, und in
these cases the focus selected was the one whose stim-
ulation was judged by the experimenters to be most
likely to be tolerated by the subject. Insbesondere, target-
ing regions of the MFG located relatively ventrally and
anteriorly increases the likelihood of stimulating the su-
perior auricularis muscle, which can produce involuntary
wincing and discomfort. In such instances, if a different
focus of alphabetization sensitivity were located in a more
dorsal and/or posterior portion of the MFG, we would
target this latter focus. Insbesondere, we found that stim-
ulation sites near or overlapping the inferior bank of
the superior frontal sulcus were well tolerated by sub-
jects.3 Another constraint was that, for each individual
Thema, all three stimulation sites were in the same hemi-
Kugel. This meant that, for each subject, the hemi-
sphere to be stimulated was determined by the location
of alphabetization-sensitive activity in the dlPFC and SPL.
PCG was chosen as the control region for this study be-
cause it was presumed to have no direct involvement in
any component of the working memory task, and thus
could serve as a region to control for nonspecific effects
of cortical rTMS. daher, PCG stimulation sites were
expressly selected for the absence of either alphabetiza-
tion- or load-sensitive activity. At each stimulation site,
the stimulating coil was oriented with the handle point-
ing posteriorly with respect to the subject’s head, Und
roughly parallel to the midline, so as to induce current in
the brain in the posterior-to-anterior direction.
Stimulation Parameters
On stimulation-present trials, the system was programmed
to deliver 30 equally spaced pulses during a 6-sec epoch,
beginning 2 sec after the offset of the instructions and
lasting for the remaining 6 sec of the delay period (d.h., A
6-sec-long train of 5 Hz rTMS). Jedoch, a programming
error was detected after the experiment was completed,
and inspection of the stimulation logs indicated that for
three subjects, Die 30 pulses were actually delivered over
Figur 1. Group load and
alphabetization effects from
the fMRI data. Effects were
extracted in a two step
Verfahren. Erste, load-sensitive
and alphabetization-sensitive
voxels were identified in each
of the three types of region.
Zweite, the magnitude of load
and alphabetization sensitivity
was determined for the voxels
identified in Step 1. Der
topography of load sensitivity
varied across subjects
(see text), and the effects
displayed here were drawn
from the functionally defined
load sensitive region of
each subject. Because load
sensitivity was only observed
in two subjects in the dlPFC
(Subjects 16 Und 21) Und
in two subjects in the SPL
(Subjects 21 Und 27), diese
results are plotted as single
data points, rather than as
means. Error bars represent
95% confidence intervals.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
0
P
1
D
7
F
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
Ö
2
C
/
N
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
J
0
Ö
1
C
7
N
1
.
2
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
0
T
.
Ö
1
N
7
0
1
7
2
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
F
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Postle et al.
1715
Tisch 1. Tabulation of Activity Identified in Spatial Normalization-based Group Analyses
Region of Activation
X
j
z
MNI Coordinates (mm)
Effect Size (Mean Percent
Signal Change)
Volume of Activation
(mm3)
Alphabetization contrast
Right PFC
Left PFC
Anterior cingulate cortex
Superior frontal gyrus
Right SPL
Right intraparietal sulcus
Left SPL
Left SPL
Left inferior parietal lobule
31.88
26.3
(cid:3)2
7.7
3.8
33.8
(cid:3)15
(cid:3)7.5
(cid:3)37.5
Right head of the caudate nucleus
11.25
Left head of the caudate nucleus
(cid:3)11.25
Load contrast
Left inferior frontal gyrus/MFG
Right central sulcus
Right PCG
Right SPL
Right inferior frontal gyrus
Right fusiform gyrus
Right amygdala
Right medial temporal lobe
(cid:3)45
30
18.75
26.25
67.5
48.75
22.5
18.75
22.5
28.1
10.5
1.9
(cid:3)63.8
(cid:3)60
(cid:3)56.25
(cid:3)60
(cid:3)45
11.25
7.5
(cid:3)7.5
(cid:3)41.25
(cid:3)33.75
(cid:3)71.25
7.5
(cid:3)45
(cid:3)3.75
(cid:3)18.75
36
34
40
66
48
44
56
44
56
12
14
44
56
64
48
32
(cid:3)24
(cid:3)20
(cid:3)16
0.37
0.29
0.34
0.46
0.63
0.42
0.39
0.38
0.47
0.45
0.13
0.14
0.21
0.23
0.73
0.42
0.41
0.43
3825.0
2137.5
562.5
956.3
900.0
506.3
506.3
281.3
900.0
1012.5
5.6
5.6
5.6
56.3
56.3
56.3
56.3
56.3
The coordinates reported here indicate the centers of clusters of activity identified within each anatomical region. Identification of anatomical
regions was confirmed via conversion of MNI coordinates to Talairach coordinates with the mni2tal Matlab routine of Matthew Brett (http://
www.mrc-cbu.cam.ac.uk/Imaging/mnispace.html) and looking them up in the atlas of Talairach and Tournoux (1988).
a 6.3-sec epoch (daher, at a rate of 4.8 Hz), und für
the remaining nine subjects, Die 30 pulses were deliv-
ered over a 6.8-sec epoch (daher, at a rate of 4.3 Hz).
The result was that, for each subject, rTMS was delivered
during the final 6 sec of the delay period and continued
during the first few hundred msec after the onset of the
probe. This ‘‘spillover’’ of rTMS into the probe portion
of the trial, although unfortunate, does not complicate
our interpretation of our results, for reasons summa-
rized in the Results section.
ERGEBNISSE
Functional Magnetic Resonance Imaging
Single-subject Analyses
Alphabetization-sensitive voxels were identified in the
dlPFC in 11 subjects and in the SPL in all 12 Fächer.
Load-sensitive voxels were identified in 11 Fächer, im
following regions (with n corresponding to the number
of subjects in which it was detected):
left superior
temporal gyrus and/or inferior parietal lobule, n = 5;
right inferior parietal lobule, n = 1; SPL, n = 2; left dorsal
extrastriate cortex, n = 1; left central sulcus, n = 2; Rechts
central sulcus, n = 1; anterior cingulate gyrus, n = 1; links
dlPFC, n = 2; right temporal pole, n = 1. To accommo-
date this topographical variability, the ‘‘load sensitivity
region’’ was defined exclusively from functional proper-
ties and permitted to vary topographically across sub-
Projekte. To address the question of whether alphabetization
sensitivity and load sensitivity are seen in the same vox-
els, we extracted estimates of these two effects (sehen
Methods section, Functional Magnetic Resonance Imag-
ing, Analyses) from alphabetization-sensitive voxels found
in the three ROIs (load-sensitive, dlPFC, and SPL) sowie
as from load-sensitive voxels found in these ROIs (z.B.,
1716
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
0
P
1
D
7
F
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
Ö
2
C
/
N
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
J
0
Ö
1
C
7
N
1
.
2
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
0
T
.
Ö
1
N
7
0
1
7
2
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
F
.
.
Ö
N
1
8
M
A
j
2
0
2
1
from alphabetization-sensitive voxels in the dlPFC, Wir
extracted an estimate of the alphabetization sensitivity
and of the load sensitivity.) Inspection of mean effect
sizes and their associated 95% confidence intervals indi-
cated that in none of the ROIs did alphabetization-
sensitive voxels display significant load sensitivity or, In
any ROI, did load-sensitive voxels demonstrate alphabet-
ization sensitivity (Figur 1). In der Tat, in the dlPFC and
SPL, alphabetization-sensitive voxels showed reliably neg-
ative load effects (mean and 95% confidence interval <
0). These results replicate and extend the findings of
Postle et al. (1999) and stand in contrast to the conclu-
sions of Veltman et al. (2003).
Group Analyses
Alphabetization-sensitive activity identified by the spatial
normalization-based group analyses was extensive and
spanned large extents, bilaterally, of the PFC, anterior
cingulate cortex, superior frontal cortex, SPL, and the
caudate nucleus (Table 1). In both hemispheres of the
PFC, these voxels showed a significant negative effect of
load [right: t(11) = (cid:3)3.4, p < .01; left: t(11) = (cid:3)4.4,
p < .005]. Load-sensitive activity identified by this analy-
sis was markedly more sparse, consisting only of indi-
vidual voxels in the right and left frontal cortex, right
central sulcus and parietal cortex, and three locations in
the right temporal lobe (Table 1). In neither hemisphere
of the PFC did these load-sensitive voxels show a sig-
nificant alphabetization effect [right: t(11) = (cid:3)0.5, ns;
left: t(11) = 1.5, ns]. Therefore, the results of the group
analyses also demonstrated a dissociation of retention
from manipulation effects (as with the single-subject
analyses, consistent with Postle et al., 1999, and incon-
sistent with Veltman et al., 2003).
Repetitive Transcranial Magnetic Stimulation
rTMS was performed in the left hemisphere in seven
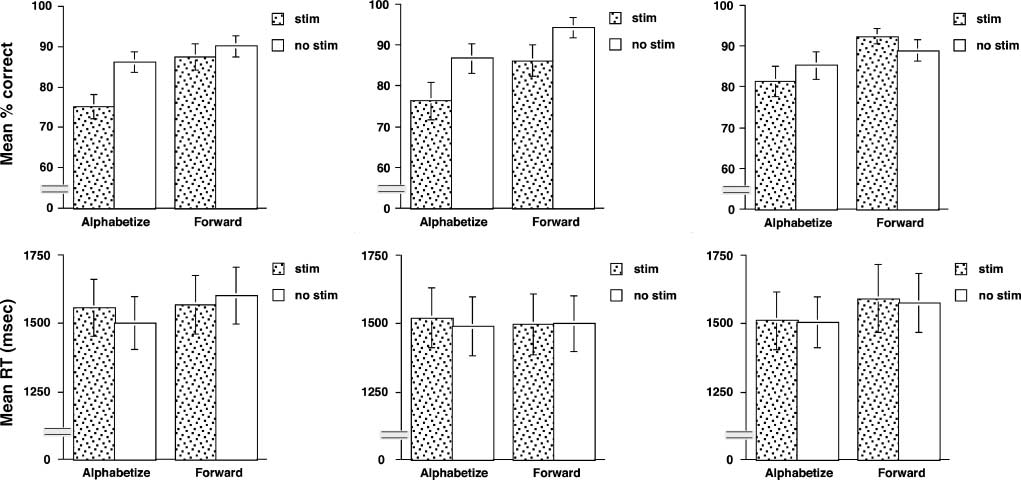
subjects and in the right hemisphere in five (Figure 2).
Analyses of variance (ANOVAs) indicated that rTMS had
its greatest effects on Alphabetize 5 performance at the
dlPFC and SPL sites and on Forward 5 performance at
the SPL (Figure 3). An initial omnibus ANOVA found no
effect of hemisphere of stimulation, F(1,10) = 0.1, ns,
and so all subsequent analyses collapsed across this varia-
ble. Omnibus ANOVA of the accuracy data revealed main
effects of trial type, F(1,11) = 11.2, p < .005, and rTMS,
F(1,11) = 7.6, p < .05, but not of region, F(2,22) < 1.0,
ns, interactions Region (cid:1) rTMS, F(2,22) = 4.9, p < .05,
and Trial Type (cid:1) rTMS, F(1,11) = 5.42, p < .05 (no other
interactions achieved significance, Fs < 1.0). ANOVA of
the accuracy data from dlPFC alone confirmed a selective
effect of rTMS on alphabetization performance, with sig-
nificant main effects of trial type, F(1,11) = 10.4, p < .01,
and rTMS, F(1,11) = 9.5, p < .05, and a Trial type (cid:1) rTMS
interaction, F(1,11) = 5.1, p < .05. In contrast, ANOVA of
the accuracy data from the PCG control region did not
show evidence for comparable selectivity of rTMS. In-
stead, it revealed a main effect of trial type, F(1,11) = 5.7,
p < .05, but no main effect of rTMS or Trial type (cid:1) rTMS
interaction (Fs < 3.5). To confirm the differential effects
of rTMS on these two regions, ANOVA directly comparing
the dlPFC with PCG was performed. It revealed a main
effect of trial type, F(1,11) = 10.6, p < .001, Region (cid:1)
rTMS interaction, F(1,11) = 7.1, p < .05, and Trial Type (cid:1)
rTMS interaction, F(1,11) = 9.3, p < .05.
In contrast with the results from the dlPFC, ANOVA of
the accuracy data from SPL stimulation revealed main
effects of trial type, F(1,11) = 5.1, p < .05, and rTMS,
F(1,11) = 5.1, p < .05, but no Trial Type (cid:1) rTMS
interaction, F(1,11) < 1, ns. The absence of this inter-
action indicated that rTMS effects on Alphabetize 5 and
Forward 5 performance were comparable.
Analyses of RT data (Figure 3) confirmed that there
were no effects of interest in these data (including no
effect of hemisphere of stimulation; Fs (cid:4) 2.0, ns). Note
that the mean RT, collapsed across all trial types and
regions, was roughly double that of the 800-msec intru-
sion of rTMS, experienced by nine of the subjects, into
the probe epoch of the trial. This, and the fact that in no
region was RT sensitive to rTMS, gives us confidence
that the error effects reported above were because of
the disruption of processes engaged during the delay
period.
DISCUSSION
The results from the dlPFC are clear: Alphabetization-
related fMRI activity is independent of (indeed, perhaps
negatively correlated with) load-related activity, and
delay-period rTMS of
loci of alphabetization-related
activity produces a decrement on Alphabetize 5 trials,
but not on Forward 5 trials. This portion of the results
therefore confirms our hypothesis that disruption
of manipulation-sensitive regions of the dlPFC would
disrupt manipulation-dependent, but not retention-
dependent, performance. (Note that these dlPFC results
cannot be attributed to the disparity in difficulty be-
tween the two tasks because of the results from the
SPL.) Our preferred interpretation of this aspect of our
results is that they were produced by rTMS disruption
of delay-period manipulation processes. We cannot rule
out, however, the possibility that the spillover of rTMS
into the first 300–800 msec of the probe epoch may
have interfered with probe perception and/or evaluation-
related processes. Nonetheless, this ambiguity does not
lessen the result of principle theoretical import, which
is that rTMS manipulation-sensitive regions of the dlPFC
did not significantly disrupt the STR of information.
The results from the SPL are more complex and,
indeed, illustrate the value that disruptive techniques
Postle et al.
1717
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
0
p
1
d
7
f
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
o
2
c
/
n
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
j
0
o
1
c
7
n
1
.
2
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
0
t
.
o
1
n
7
0
1
7
2
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
0
p
1
d
7
f
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
o
2
c
/
n
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
j
0
o
1
c
7
n
1
.
2
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
0
t
.
o
1
n
7
0
1
7
2
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
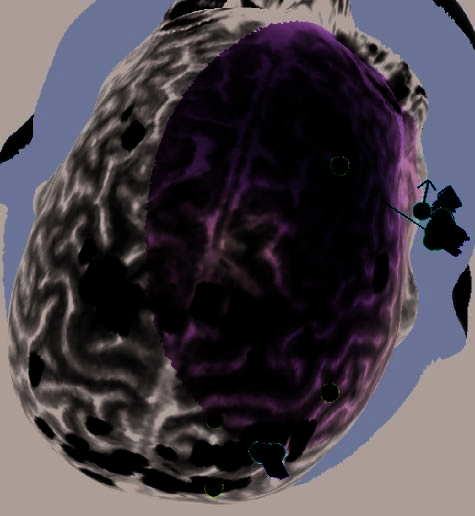
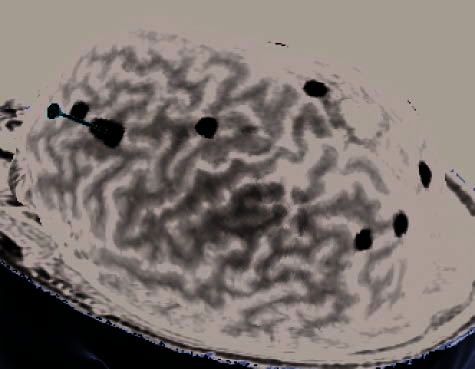
Figure 2. (A) Illustration of an rTMS experiment, from Subject 14 as displayed by the NBS system. The brain is displayed as though looking down
from above, with the nose at the top of the image. The right hemisphere appears on the right side of the image. Skin and bone have been removed
to below the level of the Sylvian fissure, and the cortex has been ‘‘peeled’’ to the depth that best displays the fMRI information (white blobs
indicate regions showing alphabetization sensitivity). Each red sphere indicates the location on the scalp at which an rTMS train was delivered; the
corresponding yellow spike indicates the orientation of the induced magnetic field for that stimulation train; the corresponding orange arrow indicates
the estimated direction of current induction. Purple spheres indicate the targeted portions of the dlPFC and SPL. For this subject, PCG was the last
region stimulated, and the splash of purple color at the end of the red spike indicates the estimated area of maximal intensity of the single rTMS
train that is captured in this image. (B–H) Analogous images, highlighting rTMS of the dlPFC, are presented for seven additional subjects. Images for
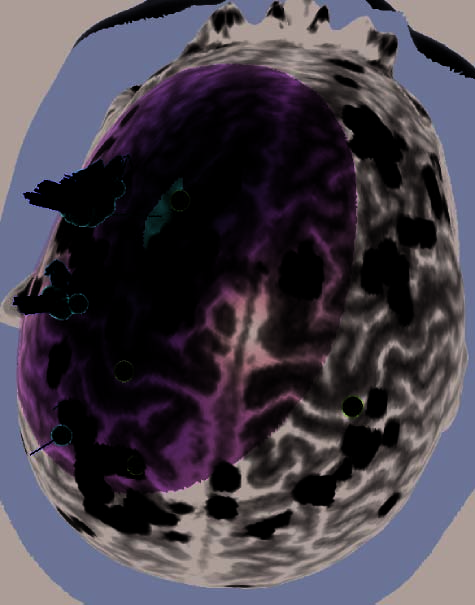
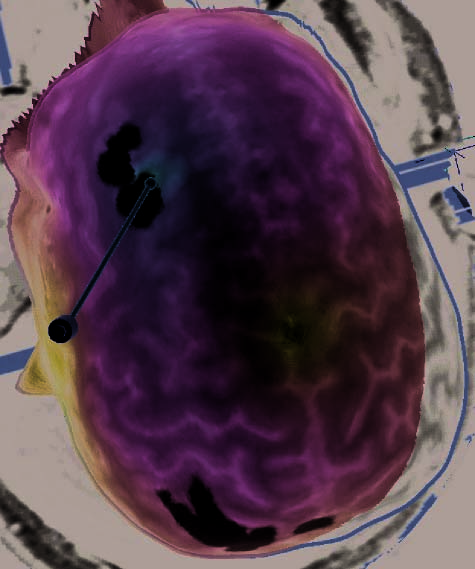
the rTMS sessions of the remaining four subjects were lost during a software upgrade. (B) Illustration of the left dlPFC rTMS of Subject 3. Display
conventions are the same as those in (A). (C) Illustration of the right dlPFC rTMS of Subject 8. Display conventions are the same as those in (A).
(D) Illustration of the three rTMS targets of Subject 16, including the left dlPFC. Display conventions are the same as those in (A).
1718
Journal of Cognitive Neuroscience
Volume 18, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
0
p
1
d
7
f
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
o
2
c
/
n
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
j
0
o
1
c
7
n
1
.
2
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
0
t
.
o
1
n
7
0
1
7
2
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
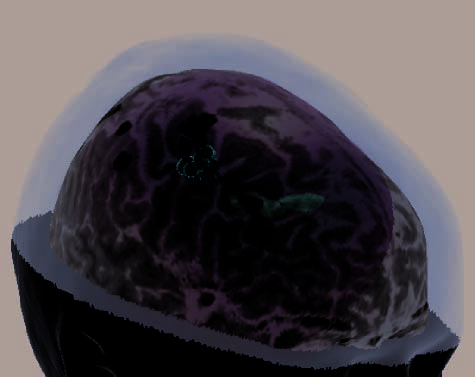
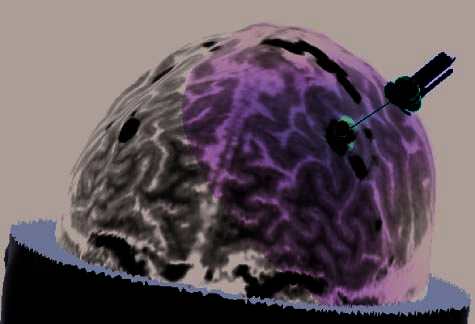
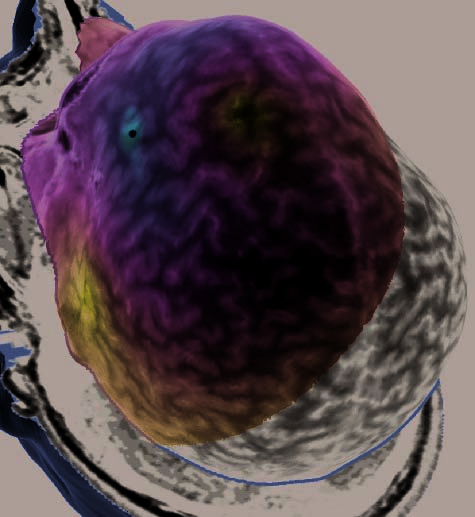
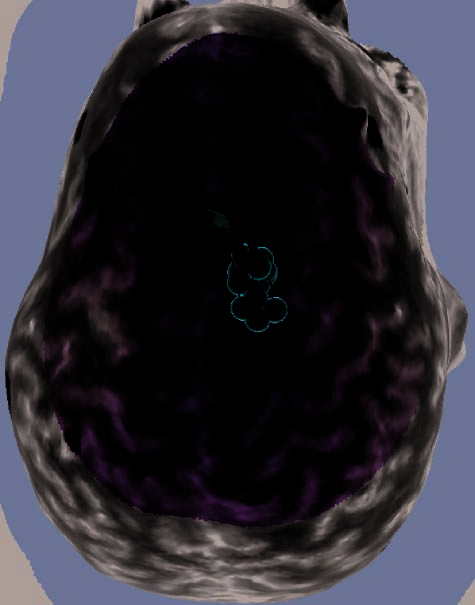
Figure 2. (continued ) (E) Illustration of frontal-lobe rTMS target of Subject 20. This is the subject mentioned in footnote 3, who could
not tolerate MFG rTMS. Display conventions are the same as those in (A). (F ) Illustration of the left dlPFC rTMS of Subject 25. Display
conventions differ from those in (A) in that an orange sphere represents the targeted portion of cortex and information relating to the NBS
system’s estimates of stimulation parameters is not shown. (G) Illustration of the left dlPFC rTMS of Subject 27. Display conventions are
the same as those in (F), except that the line emanating from the targeted portion of cortex illustrates the maximal energy vector of the
rTMS-induced magnetic field. (H) Illustration of the left dlPFC rTMS of Subject 17. Display conventions are the same as those in (G).
Postle et al.
1719
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
0
p
1
d
7
f
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
o
2
c
/
n
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
j
0
o
1
c
7
n
1
.
2
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
0
t
.
o
1
n
7
0
1
7
2
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Figure 3. Results from rTMS session. Error bars indicate SEM.
can bring to systems-level analyses of cognitive func-
tions. As with the dlPFC,
fMRI data from the SPL
indicated that alphabetization-related activity was inde-
pendent of retention-related activity in the SPL. The
rTMS data, however,
led to the opposite conclusion:
Disruption of activity at alphabetization-sensitive sites in
the SPL yielded comparable levels of impairment on
retention-requiring trials as on manipulation-requiring
trials. There are at least two possible explanations for
this divergence of the rTMS from the fMRI data. One is
that the fMRI techniques that we employed are not
sufficiently sensitive to detect retention-related effects
in the SPL that are nonetheless sensitive to rTMS, a
possibility that would suggest that manipulation- and
retention-related functions are not independent in the
SPL. This is difficult to reconcile, however, with the fact
that the SPL load effects in alphabetization-sensitive
voxels were not simply ‘‘not different from 0,’’ but were
actually significantly less than 0. A second explanation,
is that the SPL voxels identified by
which we favor,
fMRI as alphabetization-specific are just that, but that
the SPL sites that we stimulated were more proximal
to retention-related areas than were the analogous
dlPFC sites. The proximity invoked here might be either
topographical or synaptic. By the former account, the
spread of the rTMS-induced electric field might be
such that rTMS targeting the SPL also has disruptive
effects on topographically adjacent areas, such as the
angular and/or supramarginal gyri of the inferior parietal
lobule, that may, themselves, support the retention of
phonologically encoded information in working mem-
ory. By the latter account, the region(s) supporting
retention may be only one or two synapses distant from
the SPL. It is known, for example, that TMS produces
PET activation in regions that are mono- and disynapti-
cally connected to the region that is directly targeted
with TMS (Ferrarelli et al., 2004).
The absence of disruptive effects of the PFC rTMS
on Forward 5 performance is consistent with the previ-
ously existing literature. Most notably, Herwig et al.
(2003) used a delayed letter recognition task quite sim-
ilar to our Forward 5 trials, and a similar method for
guiding rTMS with fMRI data. They found no effect with
a memory load of six items of delay-period rTMS target-
ing the dlPFC or parietal cortex (SPL or inferior parietal
lobule, depending on the subject). With a load of seven
items, however, delay-period rTMS targeting both the lat-
eral premotor and parietal cortex and had disruptive ef-
fects. (They did not stimulate the dlPFC at load 7.) Other
studies that have disrupted working memory perform-
ance with rTMS of the PFC have used tasks that leave
ambiguous the type of information and/or process that is
being disrupted. For example, delay-period activity during
spatial delayed response (Brandt, Ploner, Meyer, Leistner,
& Villringer, 1998; Pascual-Leone & Hallett, 1994) might
be sensory, attentional, motoric, or some combination of
these. The n-back task (Mottaghy, Gangitano, Krause, &
Pascual-Leone, 2003; Mottaghy, Doring, Muller-Gartner,
Topper, & Krause, 2002; Mottaghy et al., 2000) requires
the simultaneous engagement of several retention- and
control-related operations. The absence of laterality effect
in our rTMS results is also consistent with the existing
literature, which documents disruption of verbal working
memory performance with right hemisphere as well as
left hemisphere rTMS (e.g., Herwig et al., 2003; Mottaghy
et al., 2002). This may be because of the complex and
1720
Journal of Cognitive Neuroscience
Volume 18, Number 10
distributed nature of the networks that support verbal
working memory (e.g., Mottaghy et al., 2002, 2003) and/
or that rTMS of a right hemisphere target is expected to
have strong effects on the homologous contralateral re-
gion (Ferrarelli et al., 2004; Bestmann, Baudewig, Siebner,
Rothwell, & Frahm, 2003).
The results of the present study are consistent with a
model of segregation of executive control from STR
functions in the PFC. They are also consistent with the
idea that posterior regions may be more important for
the STR of information than is the PFC. The contribu-
tions of the PFC to working memory may be via general-
purpose control processes that are not specialized for
working memory (Postle, in press).
Acknowledgments
We thank Steve Esser, Olufunsho Faseyitan, Cassandra Jones,
and Christopher Jordan for technical and experimental assist-
ance. This work was supported by NIH MH064498 (B. R. P.)
and NARSAD (G. T.).
Reprints requests should be sent to Bradley R. Postle, 1202
West Johnson Street, Madison, WI 53726, USA, or via e-mail:
postle@wisc.edu.
Notes
1. The logic of this portion of the study was that Forward 2
trials had only been included in the fMRI study to permit eval-
uation of whether alphabetization-sensitive voxels (i.e., voxels
significant for the [DelayAlphabetize 5–DelayForward 5] contrast)
also showed load sensitivity (as assessed with the contrast
[DelayForward 5–DelayForward 2]). For rTMS, however, Forward 5
trials were sufficient, because if Forward 5 performance were
disrupted by delay-period rTMS, one could infer that processes
necessary for the STR of 5 items were disrupted. (Note that this
inference would hold whether or not Forward 2 performance
was disrupted by rTMS, a fact that makes clear why Forward 2
trials were not needed for the rTMS portion of this study.)
The converse would be true if Forward 5 performance were
not disrupted by delay-period rTMS.
2. The motor threshold offers a means of normalizing stim-
ulation intensity across subjects, because stimulation of the
(contralateral) motor cortex and PFC at motor threshold pro-
duces positively correlated evoked responses (Kahkonen et al.,
2004).
3. One subject (Subject 20) could not tolerate rTMS of the
MFG, because of excessive stimulation of the superior auricularis
muscle. For this subject, a cluster of alphabetization-sensitive
voxels along the midline, in a region judged to correspond to
Brodmann’s area 8, was selected as the alternative target for
dlPFC stimulation (Figure 2E). The remaining two regions for
this subject were stimulated in the right hemisphere.
REFERENCES
Aguirre, G. K., Zarahn, E., & D’Esposito, M. (1998). The
variability of human, BOLD hemodynamic responses.
Neuroimage, 8, 360–369.
Baddeley, A. D. (2000). The episodic buffer: A new component of
working memory? Trends in Cognitive Sciences, 4, 417–423.
Baddeley, A. D., & Logie, R. H. (1999). Working memory:
The multiple-component model. In A. Miyake & P. Shah
(Eds.), Models of working memory (pp. 28–61). Cambridge,
UK: Cambridge University Press.
Bestmann, S., Baudewig, J., Siebner, H. R., Rothwell, J. C.,
& Frahm, J. (2003). Subthreshold high-frequency TMS of
human primary motor cortex modulates interconnected
frontal motor areas as detected by interleaved fMRI-TMS.
Neuroimage, 20, 1685–1696.
Boynton, G. M., Engel, S. A., Glover, G. H., & Heeger, D. J.
(1996). Linear systems analysis of functional magnetic
resonance imaging in human V1. Journal of Neuroscience,
16, 4207–4221.
Brandt, S. A., Ploner, C. J., Meyer, B.-U., Leistner, S., &
Villringer, A. (1998). Effects of repetitive transcranial
magnetic stimulation over dorsolateral prefrontal and
posterior parietal cortex on memory-guided saccades.
Experimental Brain Research, 118, 197–204.
Constantinides, C., Franowicz, M. N., & Goldman-Rakic, P. S.
(2001). The sensory nature of mnemonic representation
in the primate prefrontal cortex. Nature Neuroscience, 4,
311–316.
Courtney, S. M. (2004). Attention and cognitive control as
emergent properties of information representation in
working memory. Cognitive, Affective, & Behavioral
Neuroscience, 4, 501–516.
D’Esposito, M., & Postle, B. R. (1999). The dependence of
span and delayed-response performance on prefrontal
cortex. Neuropsychologia, 37, 1303–1315.
Feredoes, E., & Postle, B. R. (2005). Does size matter?
Quantitatively and qualitatively discrepant results
yielded by single-subject and spatial normalization
approaches to fMRI group analysis. Poster presented at
the annual meeting of the Society for Neuroscience 2005.
Ferrarelli, F., Haraldsson, H. M., Barnhart, T. E., Roberts,
A. D. F., Oakes, T. R., Massimini, M., Stone, C. K., Kalin,
N. H., & Tononi, G. (2004). A [17F]-fluoromethane PET/TMS
study of effective connectivity. Brain Research Bulletin,
64, 103–113.
Friston, K. J., Holmes, A. P., Poline, J.-B., Heather, J. D., &
Frackowiak, R. S. J. (1995). Analysis of fMRI time-series
revisited. Neuroimage, 2, 45–53.
Funahashi, S., Chafee, M. V., & Goldman-Rakic, P. S. (1993).
Prefrontal neuronal activity in rhesus monkeys performing
a delayed anti-saccade task. Nature, 365, 753–756.
Garavan, H., Ross, T. J., Li, S.-J., & Stein, E. A. (2000). A
parametric manipulation of central executive functioning.
Cerebral Cortex, 10, 585–592.
Goldman-Rakic, P. S., & Leung, H.-C. (2002). Functional
architecture of the dorsolateral prefrontal cortex in
monkeys and humans. In D. T. Stuss & R. T. Knight (Eds.),
Principles of frontal lobe function (pp. 85–95). Oxford:
Oxford University Press.
Hamilton, M. (1960). A rating scale for depression. Journal
of Neurology, Neurosurgery, and Psychiatry, 23, 56–62.
Handwerker, D. A., Ollinger, J. M., & D’Esposito, M. (2004).
Variation of BOLD hemodynamic responses across subjects
and brain regions and their effects on statistical analyses.
Neuroimage, 21, 1639–1651.
Herwig, U., Abler, B., Schonfeldt-Lecuona, C., Wunderlich, A.,
Grothe, J., Spitzer, M., & Walter, H. (2003). Verbal storage
in a premotor-parietal network: Evidence from fMRI-guided
magnetic stimulation. Neuroimage, 20, 1032–1041.
Jonides, J., Lacey, S. C., & Nee, D. E. (2005). Processes of
working memory in mind and brain. Current Directions
in Psychological Science, 14, 2–5.
Josephs, O., Turner, R., & Friston, K. (1997). Event-related
fMRI. Human Brain Mapping, 5, 243–248.
Postle et al.
1721
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
0
p
1
d
7
f
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
o
2
c
/
n
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
j
0
o
1
c
7
n
1
.
2
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
0
t
.
o
1
n
7
0
1
7
2
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Kahkonen, S., Wilenius, J., Komssi, S., & Elmoniemi, R. J.
Pessoa, L., Gutierrez, E., Bandettini, P. A., & Ungerleider,
(2004). Distinct differences in cortical reactivity of motor
and prefrontal cortices to magnetic stimulation. Clinical
Neurophysiology, 115, 583–588.
Kwong, K. K., Belliveau, J. W., Chesler, D. A., Goldberg, I. E.,
Weisskoff, R. M., Poncelet, B. P., Kennedy, D. N., Hoppel,
B. E., Cohen, M. S., Turner, R., Cheng, H. M., Bradly, T. J.,
& Rosen, B. R. (1992). Dynamic magnetic resonance
imaging of human brain activity during primary sensory
stimulation. Proceedings of the National Academy of
Sciences, U.S.A., 89, 5675–5679.
Lebedev, M. A., Messinger, A., Kralik, J. D., & Wise, S. P.
(2004). Representation of attended versus remembered
locations in prefrontal cortex. PloS Biology, 2, 1919–1935.
Leung, H.-C., Seelig, D., & Gore, J. C. (2004). The effect of
memory load on cortical activity in the spatial working
memory circuit. Cognitive, Affective, & Behavioral
Neuroscience, 4, 553–563.
Miller, M. B., Van Horn, J. D., Wolford, G. L., Handy, T. C.,
Valsangkar-Smyth, M., Inati, S., Grafton, S., & Gazzaniga,
M. S. (2002). Extensive individual differences in brain
activations associated with episodic retrieval are reliable over
time. Journal of Cognitive Neuroscience, 14, 1200–1214.
Mottaghy, F. M., Doring, T., Muller-Gartner, H. W., Topper, R.,
& Krause, B. J. (2002). Bilateral parietofrontal network for
verbal working memory: An interference approach using
repetitive transcranial magnetic stimulation. European
Journal of Neuroscience, 16, 1627–1632.
Mottaghy, F. M., Gangitano, M., Krause, B. J., & Pascual-Leone, A.
(2003). Chronometry of parietal and prefrontal activations
in verbal working memory revealed by transcranial
magnetic stimulation. Neuroimage, 18, 565–575.
Mottaghy, F. M., Krause, B. J., Kemna, L. J., Topper, R.,
Tellmann, L., Beu, M., Pascual-Leone, A., & Muller-Gartner,
H. W. (2000). Modulation of the neuronal circuitry
subserving working memory in healthy human subjects by
repetitive transcranial magnetic stimulation. Neuroscience
Letters, 280, 167–170.
Narayanan, N., Prabhakaran, V., Bunge, S. A., Christoff, K.,
Fine, E. M., & Gabrieli, J. D. (2005). The role of prefrontal
cortex in the maintenance of verbal working memory
information: An event-related fMRI analysis.
Neuropsychology, 19, 223–232.
Ogawa, S., Tank, D. W., Menon, R., Ellermann, J. M., Kim, S. G.,
Merkle, H., & Ugurbil, K. (1992). Intrinsic signal changes
accompanying sensory stimulation: Functional brain
mapping using MRI. Proceedings of the National Academy
of Sciences, U.S.A., 89, 5951–5955.
Pascual-Leone, A., & Hallett, M. (1994). Induction of errors
in a delayed response task by repetitive transcranial
magnetic stimulation of the dorsolateral prefrontal
cortex. NeuroReport, 5, 2517–2520.
L. G. (2002). Neural correlates of visual working memory:
fMRI amplitude predicts task performance. Neuron, 35,
975–987.
Petrides, M. (2000). Dissociable roles of mid-dorsolateral
prefrontal and anterior inferotemporal cortex in
visual working memory. Journal of Neuroscience, 20,
7496–7503.
Postle, B. R. (2006). Working memory as an emergent property
of the mind and brain. Neuroscience, 139, 23–38.
Postle, B. R., Berger, J. S., & D’Esposito, M. (1999). Functional
neuroanatomical double dissociation of mnemonic and
executive control processes contributing to working
memory performance. Proceedings of the National
Academy of Sciences, U.S.A., 96, 12959–12964.
Postle, B. R., Zarahn, E., & D’Esposito, M. (2000). Using
event-related fMRI to assess delay-period activity during
performance of spatial and nonspatial working memory
tasks. Brain Research Protocols, 5, 57–66.
Rypma, B., & D’Esposito, M. (1999). The roles of prefrontal
brain regions in components of working memory: Effects
of memory load and individual differences. Proceedings of
the National Academy of Sciences, U.S.A., 96, 6558–6563.
Sheehan, D. V., Lecrubier, Y., Sheehan, K. H., Amorim, P.,
Janavs, J., Weiller, E., Hergueta, T., Baker, R., & Dunbar, G. C.
(1998). The Mini-International Neuropsychiatric Interview
(MINI): The development and validation of a structured
diagnostic psychiatric interview for DSM-IV and ICD-10.
Journal of Clinical Psychiatry, 59, 22–33.
Swallow, K. M., Braver, T. S., Snyder, A. Z., Speer, N. K., &
Zacks, J. M. (2003). Reliability of functional localization
using fMRI. Neuroimage, 20, 1561–1577.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. New York: Thieme.
Theeuwes, J., Olivers, C. N. L., & Chizk, C. L. (2005).
Remembering a location makes the eyes curve away.
Psychological Science, 16, 196–199.
Thielscher, A., & Kammer, T. (2002). Linking physics with
physiology in TMS: A sphere field model to determine
the cortical stimulation site in TMS. Neuroimage, 17,
1117–1130.
Tsao, D. Y., Freiwald, W. A., Knutsen, T. A., Mandeville, J. B.,
& Tootell, R. B. H. (2003). Faces and objects in macaque
cerebral cortex. Nature Neuroscience, 6, 989–995.
Veltman, D. J., Rombouts, S. A., & Dolan, R. J. (2003).
Maintenance versus manipulation in verbal working memory
revisited: An fMRI study. Neuroimage, 18, 247–256.
Walsh, V., & Pascual-Leone, A. (2003). Transcranial magnetic
stimulation: A neurochronometrics of mind. Cambridge:
MIT Press.
Worsley, K. J., & Friston, K. J. (1995). Analysis of fMRI
time-series revisited—again. Neuroimage, 2, 173–182.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
0
p
1
d
7
f
1
/
2
1
8
1
/
9
1
3
5
0
6
/
0
1
3
7
1
o
2
c
/
n
1
2
7
0
5
0
5
6
9
1
4
8
8
/
1
j
0
o
1
c
7
n
1
.
2
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
0
t
.
o
1
n
7
0
1
7
2
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
f
.
.
.
Pasternak, T., & Greenlee, M. W. (2005). Working memory
Zarahn, E., Aguirre, G. K., & D’Esposito, M. (1997). A trial-based
in primate sensory systems. Nature Reviews Neuroscience,
6, 97–107.
Peelen, M. V., & Downing, P. E. (2005). Within-subject
reproducibility of category-specific visual activation with
functional MRI. Human Brain Mapping, 25, 402–408.
experimental design for fMRI. Neuroimage, 6, 122–138.
Zarahn, E., Rakitin, B., Abela, D., Flynn, J., & Stern, Y. (2005).
Positive evidence against human hippocampal involvement
in working memory maintenance of familiar stimuli.
Cerebral Cortex, 15, 303–316.
o
n
1
8
M
a
y
2
0
2
1
1722
Journal of Cognitive Neuroscience
Volume 18, Number 10