Professional Music Training and Novel Word Learning:
From Faster Semantic Encoding to Longer-lasting
Word Representations
Eva Dittinger1, Mylène Barbaroux1, Mariapaola D’Imperio1, Lutz Jäncke2,
Stefan Elmer2*, and Mireille Besson1*
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
Abstrakt
■ On the basis of previous results showing that music training
positively influences different aspects of speech perception and
Erkenntnis, the aim of this series of experiments was to test the
hypothesis that adult professional musicians would learn the
meaning of novel words through picture–word associations
more efficiently than controls without music training (d.h., fewer
errors and faster RTs). We also expected musicians to show fas-
ter changes in brain electrical activity than controls, insbesondere
regarding the N400 component that develops with word learn-
ing. In line with these hypotheses, musicians outperformed
controls in the most difficult semantic task. Darüber hinaus, although
a frontally distributed N400 component developed in both
groups of participants after only a few minutes of novel word
learning, in musicians this frontal distribution rapidly shifted
to parietal scalp sites, as typically found for the N400 elicited
by known words. Endlich, musicians showed evidence for better
long-term memory for novel words 5 months after the main ex-
perimental session. Results are discussed in terms of cascading
effects from enhanced perception to memory as well as in
terms of multifaceted improvements of cognitive processing
due to music training. To our knowledge, this is the first report
showing that music training influences semantic aspects of lan-
guage processing in adults. These results open new perspec-
tives for education in showing that early music training can
facilitate later foreign language learning. Darüber hinaus, the design
used in the present experiment can help to specify the stages of
word learning that are impaired in children and adults with
word learning difficulties. ■
EINFÜHRUNG
The aim of the present experiment was to examine the
influence of music training on word learning using both
behavioral and electrophysiological measures. On the ba-
sis of the evidence reviewed below, we tested the hy-
pothesis that musicians would be more efficient at
word learning than nonmusicians and that the differ-
ences would be reflected in their pattern of brain waves.
There is strong evidence from previous cross-sectional
studies comparing adult musicians and nonmusicians
that long-term music training promotes brain plasticity
(Münte, Altenmüller, & Jäncke, 2002) in modifying the
functional (Schneider et al., 2002; Pantev et al., 1998)
and structural (Elmer, Hänggi, Meyer, & Jäncke, 2013;
Gaser & Schlaug, 2003; Schneider et al., 2002) architec-
ture of the auditory pathway. Results of longitudinal stud-
ies, mostly in children, showed that music training can be
the cause of the observed effects (François, Chobert,
Besson, & Schön, 2013; Strait, Parbery-Clark, O’Connell,
& Kraus, 2013; Chobert, François, Velay, & Besson, 2012;
Moreno et al., 2011; Hyde et al., 2009; Moreno et al.,
2009). Most importantly for the present purposes, Dort
1CNRS and Université Aix Marseille, 2University of Zurich
*Shared last authorship.
© 2016 Massachusetts Institute of Technology
is also evidence that music training improves different
aspects of speech processing (for review, see Asaridou &
McQueen, 2013; Besson, Chobert, & Marie, 2011; Kraus &
Chandrasekaran, 2010). These transfer effects possibly
arise because speech and music are auditory signals rely-
ing on similar acoustic cues (d.h., Dauer, frequency, In-
tensity, and timbre) and because they share, at least in
Teil, common neuronal substrates for auditory perception
(Peretz, Vuvan, Lagrois, & Armony, 2015; Jäncke, 2009)
and for higher-order cognitive processing (Rogalsky, Rong,
Saberi, & Hickok, 2011; Patel, 2008; Maess, Kölsch,
Gunter, & Friederici, 2001). Zum Beispiel, music training fa-
cilitates the processing of a variety of segmental (Bidelmann,
Weiss, Moreno, & Alain, 2014; Kühnis, Elmer, & Jäncke,
2014; Elmer, Meyer, & Jäncke, 2012; Chobert, Marie,
François, Schön, & Besson, 2011; Musacchia, Sams, Skoe,
& Kraus, 2007) and suprasegmental speech attributes (Marie,
Delogu, Lampis, Olivetti Belardinelli, & Besson, 2011; Wong
& Perrachione, 2007) within native (Schön, Magne, &
Besson, 2004) and nonnative languages (Marques, Moreno,
Castro, & Besson, 2007). Darüber hinaus, both musically trained
Kinder (Jentschke & Kölsch, 2009) and adults (Fitzroy &
Sanders, 2013) are more sensitive to violations of linguistic
and music syntax than participants without music training.
Perhaps most importantly, recent results also showed that
Zeitschrift für kognitive Neurowissenschaften 28:10, S. 1584–1602
doi:10.1162/jocn_a_00997
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
Ö
5
C
4
N
9
_
6
A
/
_
J
0
Ö
0
C
9
N
9
7
_
A
P
_
D
0
0
B
9
j
9
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
/
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
long-term music training positively improves cognitive func-
tions such as auditory attention (Strait, Slater, O’Connell, &
Kraus, 2015), visual attention ( Wang, Ossher, & Reuter-
Lorenz, 2015), working and verbal memory (George &
Coch, 2011; Ho, Cheung, & Chan, 2003), executive func-
tionen (Zuk, Benjamin, Kenyon, & Gaab, 2014; Moreno
et al., 2011; Pallesen et al., 2010), and general intelligence
(Schellenberg, 2004). These findings are not surprising in-
sofar as playing an instrument at a professional level is a
multidimensional task that, together with specific motor
abilities, requires acute auditory perception and focused at-
Aufmerksamkeit, code switching between the visual information on
the score and the corresponding sounds, as well as the abil-
ity to maintain auditory information in short- and long-term
Erinnerung. Taken together, these results are in line with dy-
namic models of human cognition (Friederici & Singer,
2015; Hagoort, 2014) positing that language—and possibly
music—are processed in interaction with other cognitive
functions.
Similar to playing music, word learning is also a multi-
dimensional task requiring both perceptive and higher-
order cognitive abilities. Let us take the example of Thai.
Thai is a tonal and a quantitative language that mainly
comprises monosyllabic words (as Mandarin Chinese
and other tonal languages) and in which both tonal (d.h.,
five tones) and vowel length contrasts are linguistically rel-
evant for understanding word meaning (z.B., /pa1/ low
tone with a short vowel means “to find” and /pa:1/ niedrig
tone with a long vowel means “forest”; Gandour et al.,
2002). Daher, when it comes to learn novel words in Thai,
the learner has to focus attention on the acoustic stream
to discriminate spectral and temporal phonetic contrasts
and to build new phonological representations that can
then be associated with lexical meaning by recruiting
working, short-term, episodic, and semantic memory pro-
Prozesse. Daher, if music skills translate into improved audi-
tory perception and attention together with enhanced
working and verbal memory, it should be easier for musi-
cians to learn a language such as Thai.
The ERP method is one of the most eligible methods
to capture the fast temporal dynamics of word learning
and to examine brain plasticity, as reflected by changes
in the amplitude and/or latency of ERP components dur-
ing learning. Previous results in adults have shown that
the N400, a negative-going component that typically de-
velops between 300 Und 600 msec poststimulus onset
(Kutas & Hillyard, 1980), increases in amplitude when
meaningless items acquired meaning. Speziell, results
showed N400 enhancements in native English speakers
nach 14 hr of learning the meaning of novel French
Wörter (McLaughlin, Osterhout, & Kim, 2004) and after
45 min of learning the meaning of rare words (z.B., “clow-
der”; Perfetti, Wlotko, & Hart, 2005). Darüber hinaus, if a novel
word (Borovsky, Elman, & Kutas, 2012; Borovsky, Kutas,
& Elman, 2010; Mestres-Missé, Rodriguez-Fornells, &
Münte, 2007) or pseudoword (Batterink & Neville,
2011) is presented in a strongly constrained and mean-
ingful context, even a single exposure can be sufficient
to build up initial word representations, an effect referred
to as “fast mapping” (Carey, 1978). An incubation-like pe-
riod and further exposures are then required for consol-
idation and integration into existing lexical networks
(Dumay & Gaskell, 2007). Daher, the N400 is taken as a
reliable index of word learning, reflecting the formation
of semantic representations.
Note though that the N400 component at the core of
the above-mentioned experiments clearly showed a more
frontal scalp distribution (Borovsky et al., 2010; Mestres-
Missé et al., 2007) than the centroparietal N400 typically
elicited by already known words (Kutas, Van Petten, &
Besson, 1988). This frontal N400 distribution is compati-
ble with results showing that prefrontal and temporal
brain regions are associated with the maintenance of novel
information in working or short-term memory and the for-
mation of new associations (Hagoort, 2014) and/or with
the initial building-up of word representations in episodic
Erinnerung (Rodriguez-Fornells, Cunillera, Mestres-Missé, &
De Diego-Balaguer, 2009; Wagner et al., 1998).
Wie oben erwähnt, most studies of music-to-language
transfer effects have focused on segmental, suprasegmen-
tal, and syntactic processing levels. On the basis of the re-
sults on word learning reviewed above, this study aimed at
going a step further so as to determine whether profes-
sional music training would also influence the semantic
level of processing, most often considered as language-
specific (but see Koelsch et al., 2004) by facilitating the
learning process of novel word meaning. The general hy-
pothesis was that the optimization of perceptual and cog-
nitive functions in professional musicians would positively
influence the speed and quality of word learning as re-
flected by a behavioral advantage for musicians (d.h., lower
error rates [ERRs] and faster RTs). Darüber hinaus, basierend auf
ERPs and word learning literature (Borovsky et al., 2010,
2012; Batterink & Neville, 2011; Mestres-Missé et al.,
2007; Perfetti et al., 2005; McLaughlin et al., 2004), we ex-
pected a frontally distributed N400 component to develop
in all participants during the early stages of novel word
learning. Jedoch, if the perceptual and cognitive compu-
tations involved in word learning were facilitated in musi-
cians, the development of the N400 component should be
faster in musicians than in controls in the learning phase.
Im Gegensatz, we expected the N400 to show a centroparietal
distribution when novel word learning was consolidated.
To test these general hypotheses, we used an ecolog-
ically valid experimental design inspired by Wong and
Perrachione (2007) and based on a series of four experi-
ments that comprised several tasks performed during
the main experimental session (see Figure 1A–E). Erste, Zu
further test the hypothesis of improved auditory speech
discrimination in musicians compared with controls, Par-
ticipants performed a phonological categorization task at
the beginning and at the end of the main experimental
session (see Figure 1A and E). On the basis of previous
results showing that musicians are advantaged when
Dittinger et al.
1585
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
Ö
5
C
4
N
9
_
6
A
/
_
J
0
Ö
0
C
9
N
9
7
_
A
P
_
D
0
0
B
9
j
9
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
T
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
Ö
5
C
4
N
9
_
6
A
/
_
J
0
Ö
0
C
9
N
9
7
_
A
P
_
D
0
0
B
9
j
9
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
T
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
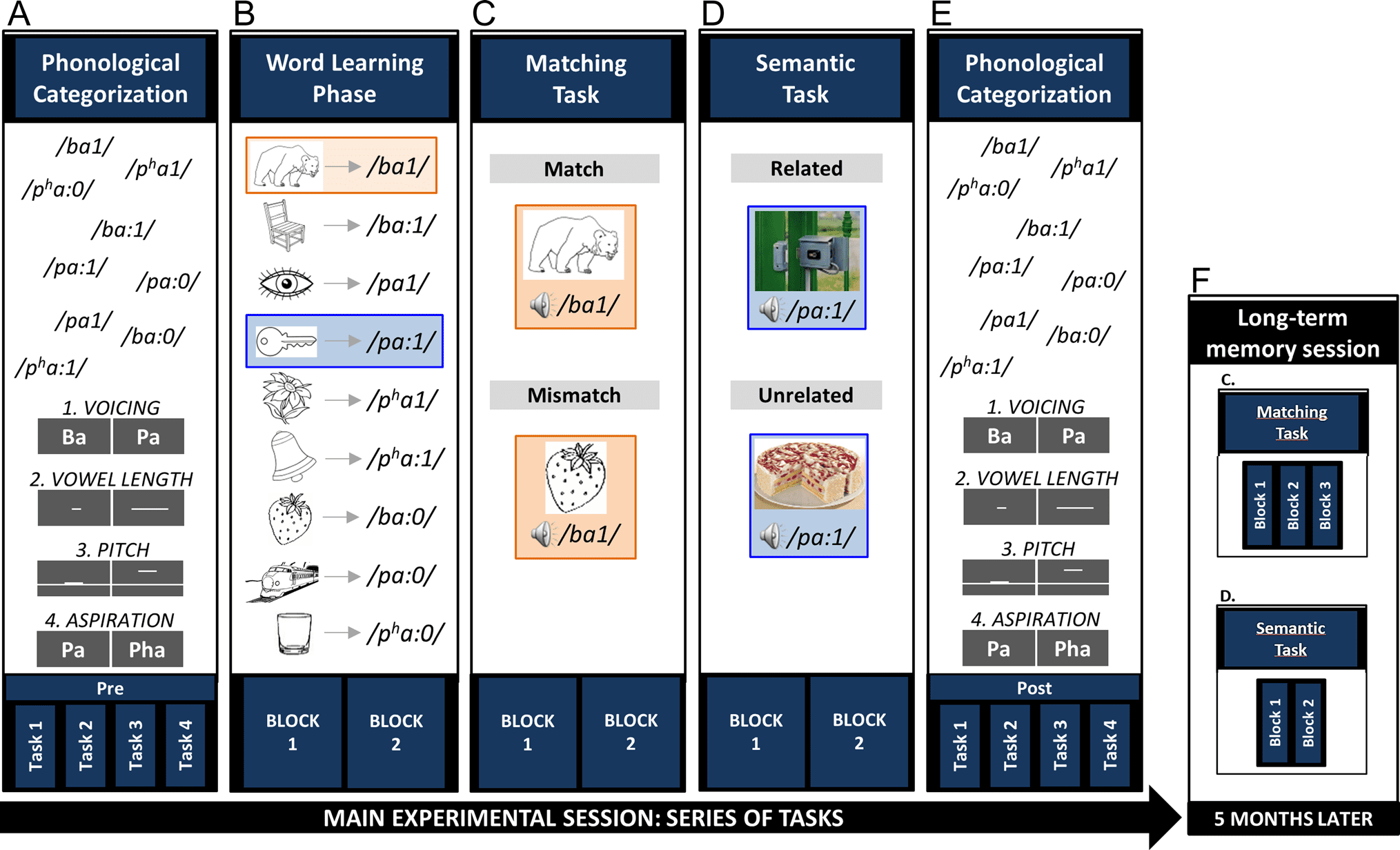
Figur 1. Experimentelles Design. Participants performed a series of tasks in the main experimental session (A–E): Erste, in the phonological categorization
Aufgabe (A), nine natural Thai monosyllabic words had to be categorized based on voicing (Task 1), vowel length (Task 2), pitch (Task 3), or aspiration
contrasts (Task 4). Zweite, in the word learning phase (B), each word was paired with its respective picture. This phase included two separate blocks of
Versuche. Dritte, in the matching task (C), the words were presented with one of the pictures, either matching or mismatching the previously learned
associations. This task included two separate blocks of trials. Vierte, in the semantic task (D), the words were presented with novel pictures that were
either semantically related or unrelated to the novel words. Wieder, this task included two separate blocks of trials. Fünfte, participants did again the four
tasks of the phonological categorization task (E). Endlich, participants came back 5 months after the main session to perform again the matching and
semantic tasks (F).
the discrimination is most difficult (Diamond, 2013; Schön
et al., 2004), we expected musicians to outperform controls
in identifying phonemic contrasts that are not relevant for
lexical discrimination in French. Darüber hinaus, based on previ-
ous literature reporting that the N100 component reflects
encoding of auditory cues in the auditory-related cortex
(Kühnis et al., 2014) and is influenced by auditory attention
and perceptual learning (Seppänen, Hämäläinen, Pesonen,
& Tervaniemi, 2012; Woldorff & Hillyard, 1991), we ex-
pected this behavioral advantage to be accompanied by
an increased N100 amplitude in musicians.
Zweite, participants were asked to learn the meaning
of the novel words through picture–word associations
(see Figure 1B), a design that has often been used in word
learning experiments in children (Friedrich & Friederici,
2008; Torkildsen et al., 2008) and in adults (Dobel,
Lagemann, & Zwitserlood, 2009). No behavioral response
was required during this word learning phase, but ERPs
were recorded to test the main hypothesis that frontally
distributed N400 components would develop in both
groups of participants (François et al., 2013; Borovsky
et al., 2010; Rodriguez-Fornells et al., 2009) but with faster
temporal dynamics in musicians than in controls.
Dritte, to test for the efficacy of the learning phase, Par-
ticipants performed a matching task and were asked to
decide whether a picture–word pair matched or mis-
matched the previously learned pairs (see Figure 1C).
Vierte, an important aspect was to determine whether
word learning was specific to the picture–word pairs
learned during the word learning phase or whether the
meaning of the newly learned words was already integrated
into semantic networks so that priming effects generalized
to new pictures. Zu diesem Zweck, participants performed a se-
mantic task during which novel pictures that had not been
seen in the previous tasks were presented. They were
asked to decide whether the picture and the word were
semantically related or unrelated (see Figure 1D). In both
the matching and the semantic tasks and in both groups
of participants, we predicted that N400 amplitudes would
be larger for mismatch and semantically unrelated words
than for match and semantically related words (d.h., the typ-
ical N400 effect; Kutas & Hillyard, 1980), thereby showing
that participants had learned the meaning of the novel
Wörter. Darüber hinaus, if the novel words’ meanings were
already integrated into existing semantic networks at the
end of the word learning phase (Borovsky et al., 2012;
1586
Zeitschrift für kognitive Neurowissenschaften
Volumen 28, Nummer 10
Batterink & Neville, 2011), we expected the N400 effect
(mismatching–matching and unrelated–related words) In
the matching and semantic tasks to show the centroparie-
tal distribution typically found for already known words
(Kutas et al., 1988). Of main interest was to specify the spa-
tiotemporal dynamics of the N400 effect in musicians and
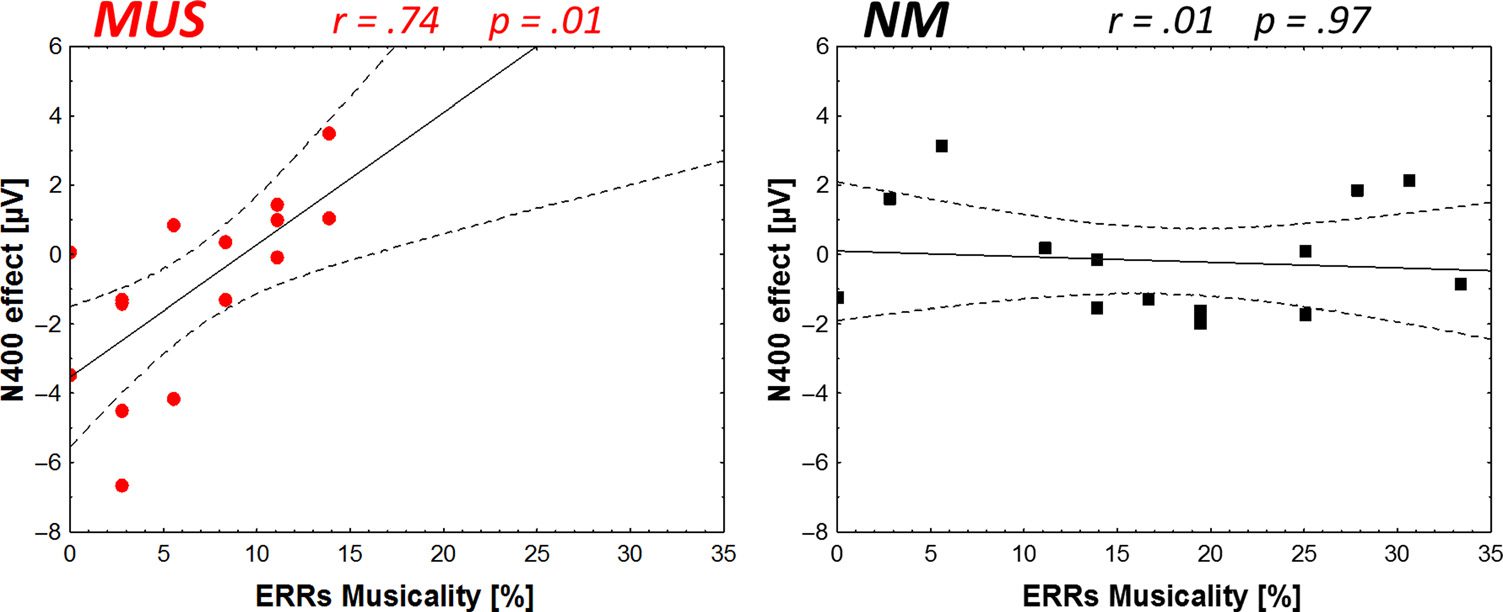
in nonmusicians. Endlich, if we were to find that music train-
ing influenced word learning, then we expected musical
ability to be positively correlated with word learning effi-
cacy, as reflected by behavioral measures and/or the
N400 effect in the matching and semantic tasks.
Endlich, a subset of the participants was behaviorally re-
tested after 5 months (see Figure 1F) in the matching and
semantic tasks to evaluate the maintenance of novel
words in long-term memory. It was of interest to deter-
mine whether the behavioral advantages of musicians in
a variety of cognitive domains, as reviewed above, extend
to long-term memory. To the best of our knowledge, Das
aspect has not yet been investigated.
Zusammenfassend, this experimental design is relevant for
specifying whether music expertise influences the se-
mantic level of speech processing, an issue that, to our
Wissen, has not been addressed before. By analyzing
ERPs, we aimed at better understanding the dynamics of
word learning, how fast semantic processes develop, Und
whether and how the N400 is influenced by music train-
ing. Showing that long-term music training with an early
start (as it is most often the case in professional musi-
cians) may facilitate foreign language learning later in life
should add evidence to the claim that music training has
important societal consequences for education (Besson
et al., 2011; Kraus & Chandrasekaran, 2010). Endlich, Das
experimental design is of potential interest for clinical re-
suchen: using several different tasks that call upon several
perceptual and cognitive functions (phonological catego-
rization, formation of picture–word associations, main-
taining these associations in short-term and long-term
memory and generalization of learning effects) innerhalb
the same patient may help specify the processing stages
that are deficient in adults or children with language
learning disorders.
METHODEN
Teilnehmer
A total of 30 participants with 15 professional musicians
(MUS, acht Frauen) Und 15 controls without formal mu-
sic training (nonmusicians, NM, acht Frauen) aber in-
volved in a regular leisure activity (z.B., sports, tanzen,
theater) were paid to participate in the experimental ses-
sion lasting for 2.5 Std (including the application of the
Electrocap, psychometric measurements, and experimen-
tal tasks). The two groups did not differ in age (MUS:
Durchschnittsalter = 25.1 Jahre, age range = 19–30, SD = 3.9;
NM: Durchschnittsalter = 25.7 Jahre, age range = 19–33, SD =
4.8; F(1, 28) = 0.02, p = .68). All participants were native
French speakers, had comparable education levels (uni-
versity degree) and socioeconomic background (Kriterien
of the National Institute of Statistics and Economic Stud-
ies; MUS: 4.4; NM: 4.9; F(1, 28) = 1.45, p = .24), and re-
ported no past or current audiological or neurological
deficits. MUS practiced their instruments for an average
von 17 Jahre (range = 11–24, SD = 4.1) and the musician
group included three pianists, two accordionists, four vio-
linists, one cellist, two guitarists, one hornist, one tubist,
and one flautist. None of the participants was bilingual,
but all spoke English as a second language and most par-
ticipants (except for 1 MUS and 3 NM) had a rudimentary
knowledge of a third language that was neither tonal nor
quantitative. The study was conducted in accordance with
the Helsinki declaration, and all participants gave their in-
formed consent before enrolling in the experiment.
Screening Measures
Cognitive Ability
Standardized psychometric tests were used to examine
short-term and working memory (forward and reverse
Digit Span, WISC-IV; Wechsler, 2003), visual attention
(NEPSY from Korkman, Kirk, & Kemp, 1998) and non-
verbal general intelligence (progressive matrices, PM47;
Raven, Corporation, & Lewis, 1962).
Musical Aptitude
Participants performed two musicality tests (adapted
from the MBEA battery; Peretz, Champod, & Hyde,
2003) consisting in judging whether pairs of piano melo-
dies were same or different, based either on melodic or
on rhythmic information.
Experimental Stimuli
Auditory Stimuli
Nine natural Thai monosyllabic words were selected for
the experiment: /ba1/, /pa1/, /pha1/, /ba:1/, /pa:1/, /pha:1/,
/ba:0/, /pa:0/, /pha:0/.1 These words varied in vowel dura-
tion, with short (/ba1/, /pa1/ and /pha1/; 261 msec on
average) and long vowels (/ba:1/, /pa:1/, /pha:1/, /ba:0/,
/pa:0/ and /p ha:0/; 531 msec on average), und in
fundamental frequency, with low-tone (/ ba1/, /pa1/,
/pha1/, /ba:1/, /pa:1/ and /pha:1/; F0 = 175 Hz on average)
and midtone vowels (/ba:0/, /pa:0/ and /pha:0/; F0 = 218 Hz
on average). Außerdem, words contained voicing
contrasts (/ba1/, /ba:1/ and /ba:0/, VOT = −144 msec vs.
/pa1/, /pa:1/ and /pa:0/, VOT = 3 ms) as well as aspiration
contrasts (/pa1/, /pa:1/ and /pa:0/, VOT = 3 msec vs. /pha1/,
/pha:1/ and /pha:0/, VOT = 77 ms).2 Stimuli were recorded
by a female Thai–French bilingual, ensuring that all words
were produced naturally. For each word, five versions were
digitally recorded to reproduce natural speech variability.
Sound pressure level was normalized across all words to
Dittinger et al.
1587
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
Ö
5
C
4
N
9
_
6
A
/
_
J
0
Ö
0
C
9
N
9
7
_
A
P
_
D
0
0
B
9
j
9
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
a mean level of 70 dB by using Praat software (Boersma
& Weenink, 2011).
than four successive same responses. Task order and re-
sponse side were counterbalanced across participants.
Visual Stimuli
For the learning phase, nine pictures representing famil-
iar objects (d.h., bear, flower, key, chair, bell, eye, strawberry,
train, glass) were selected based on the standardized set of
260 pictures (that are matched for name and image agree-
ment, Vertrautheit, and visual complexity) built by Snodgrass
and Vanderwart (1980).3 The same pictures as in the learn-
ing phase were then presented in the matching task. Für
the semantic task, 54 new pictures that the participants
had not seen before in the experiment and that were se-
mantically related or unrelated to the meaning of the newly
learned words were chosen from the Internet by two of the
authors (ED and MB). Students from our university (n =
60; age range = 19–25 years) were asked to rate the seman-
tic relatedness between new and old pictures (das ist, those
previously presented during the word learning phase). Half
of the presented pairs were semantically related and the
other half were semantically unrelated, and this was con-
firmed by the students’ ratings.
Experimental Tasks
Participants were tested individually in a quiet experimen-
tal room (d.h., Faraday cage), where they sat in a comfort-
able chair at about 1 m from a computer screen. Auditiv
stimuli were presented through HiFi headphones (HD590,
Sennheiser Electronic GmBH, Wedemark, Deutschland) bei
70-dB sound pressure level. Visual and auditory stimuli
presentation as well as the collection of behavioral data
were controlled by Presentation software ( Version 11.0,
Neurobehavioral Systems, Berkeley CA).
Main Experimental Session (See Figure 1A–E)
Phonological categorization task. Am Anfang
and at the end of the experiment, participants performed
four different phonological tasks that lasted for 2.3 min
jede. All nine Thai monosyllabic words were presented
in each task, but participants were asked to categorize
them based upon different features in each task: (1) Die
voicing contrast (z.B., /ba1/ vs. /pa1/), (2) the vowel length
(z.B., short: /ba1/ vs. long /ba:1/), (3) pitch (z.B., niedrig: /pa:1/
vs. hoch: /ba:0/), Und (4) the aspiration contrast (z.B., /pa1/
vs. /pha1/; see Figure 1A and E). For each task, the contrast
was visually represented on the left (z.B., “short” with a
short line) and right (z.B., “long” with a long line) half of
the screen and participants had to press one of two re-
sponse buttons according to the correct side (z.B., links
one for short and right one for long vowels), as quickly and
accurately as possible. Each word was presented 10 mal
in a pseudorandomized order with the constraints of no
immediate repetition of the same word and no more
Word learning phase. Participants were asked to learn
the meaning of each word previously presented in the
phonological categorization task using picture–word as-
sociations. Zum Beispiel, a drawing of a bear was followed
by the auditory presentation of the word /ba1/, and thus,
/ba1/ was the word for bear in our “foreign” language
(see Figure 1B). Each of the nine picture–word pairs
was presented 20 mal, ergebend 180 trials that were
pseudorandomly presented (d.h., no immediate repeti-
tion of the same association) in two blocks of 3 min each.
The picture was presented first and then followed after
750 msec by one of the nine words. Total trial duration
War 2000 ms. Two different lists were built, so that
across participants different pictures were associated
with different words. No behavioral response was re-
quired from the participants but they were told that sub-
sequent tests would evaluate whether they learned the
meaning of the novel words.
Matching task. One of the nine pictures was presented,
followed after 750 msec by an auditory word that
matched or mismatched the associations previously
learned in the word learning phase. Zum Beispiel, Wo-
as the drawing of a bear followed by /ba1/ (d.h., bear) War
a match, the drawing of a strawberry followed by /ba1/
was a mismatch (see Figure 1C). Participants were asked
to press one of two response keys accordingly, as quickly
and accurately as possible. Response hand was counter-
balanced across participants. At the end of the trial, a row
of XXXX appeared on the screen, and participants were
asked to blink during this time period (1000 ms; total
trial duration: 3750 ms) to minimize eye movement
artifacts during word presentation. Each word was pre-
gesendet 20 mal, half in match condition and half in mis-
match condition. The total of 180 trials was pseudorandomly
vorgeführt (d.h., no immediate repetition of the same asso-
ciation and no more than four successive same responses)
within two blocks of 5.6 min each.
Semantic task. One of the new pictures was presented,
followed after 1500 msec by an auditory word that was
semantically related or unrelated. Zum Beispiel, although
the picture of a lock was semantically related to the pre-
viously learned word /pa:1/ (d.h., “key”), the picture of a
strawberry cake was semantically unrelated to /pa:1/ (sehen
Figure 1D). Participants were asked to press one of two
response keys accordingly, as quickly and accurately as
möglich. A familiarization task including four trials was
administrated before starting the task. Response hand
was counterbalanced across participants. At the end of
the trial, a row of XXXX appeared on the screen, and par-
ticipants were asked to blink during this time period
(1000 ms; total trial duration = 4500 ms). Each word
was presented 12 mal, but none of the new pictures
1588
Zeitschrift für kognitive Neurowissenschaften
Volumen 28, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
Ö
5
C
4
N
9
_
6
A
/
_
J
0
Ö
0
C
9
N
9
7
_
A
P
_
D
0
0
B
9
j
9
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
were repeated, so that on each trial the word was associated
with a different related or unrelated picture. Half of the
picture–word pairs were semantically related, and half
were semantically unrelated. A total of 108 trials was pre-
sented pseudorandomly (d.h., no immediate repetition of
the same association and no more than four successive
same responses) within two blocks of 4 min each.
Long-term Memory Session (See Figure 1F)
To test for long-term memory effects on behavior (d.h.,
ERRs and RTs), participants performed again the matching
(always administered first) and semantic tasks 5 months af-
ter the main experimental session (no ERPs were recorded).
Because of a dropout rate of 33%, nur 10 Teilnehmer
were retested in each group. In the matching task, a total
von 270 trials were presented within three blocks. Im
semantic task, insgesamt 216 trials were presented in two
blocks (with a short pause within each block).
EEG Data Acquisition
The EEG was continuously recorded at a sampling rate of
512 Hz with a band-pass filter of 0–102.4 Hz by using a
Biosemi amplifier system (BioSemi Active 2, Amsterdam,
Die Niederlande) mit 32 active Ag/Cl electrodes (Biosemi
Pintype) located at standard positions according to the
International 10/20 System (Jasper, 1958). The EOG was re-
corded from flat-type active electrodes placed 1 cm to the
left and right of the external canthi and from an electrode
beneath the right eye. Two additional electrodes were
placed on the left and right mastoids. Electrode imped-
ance was kept below 5 kΩ. EEG data were analyzed using
Brain Vision Analyzer software ( Version 1.05.0005 &
Version 2.1.0; Brain Products, München, Deutschland). Alle
data were re-referenced offline to the averaged left and
right mastoids, filtered with a bandpass filter from 1 Zu
30 Hz (slope of 24 dB/oct), and independent component
analysis and inverse independent component analysis
were used to identify and remove components associated
with vertical and horizontal ocular movements. Endlich,
DC-detrend and removal of artifacts above a gradient cri-
terion of 10 μV/msec or a max–min criterion of 100 μV
over the entire epoch were applied automatically. Für
each participant, ERPs were time-locked to word onset,
segmented into 2700 msec epochs, including a 200-msec
baseline and averaged within each condition. Individuell
averages were then averaged together to obtain the grand
average across all participants.
task that was performed both at the beginning and at the
end of the experiment, Order (preexperiment vs. postex-
periment) was included as a within-subject factor together
with Task ( Voicing vs. Vowel length vs. Pitch vs. Aspira-
tion). The matching and semantic tasks were only per-
formed once but each experiment was divided into two
blocks of trials so that factors were Block (1 vs. 2) and Con-
dition (match vs. mismatch or related vs. unrelated). To
further increase clarity, factors were again specified at the
beginning of each task in the Results section. On the basis
of ERRs, four outliers (2 MUS and 2 NM, ±2 SD away from
der Mittelwert) were excluded from further analyses.
For ERPs, we analyzed the early stages of auditory pro-
cessing in the phonological categorization task using N100
peak amplitude measures. Im Gegensatz, during the word
learning phase, as well as in the matching and semantic
tasks, we focused on semantic processing and we analyzed
the mean amplitude of the N400 component. Effects on
the N200 were also analyzed using mean amplitudes. Sei-
cause ERPs were only analyzed for correct responses and
because the ERP traces of the four outliers that were elim-
inated from behavioral analyses were similar to the grand
average in each group, all participants (d.h., 15 MUS and
15 NM) were included in the ERP analyses. ANOVAs always
included Group (MUS vs. NM) as a between-subject factor
and Laterality (links: F3, C3, P3; midline: Fz, Cz, Pz; Rechts: F4,
C4, P4) and Anterior/Posterior (frontal: F3, Fz, F4; zentral:
C3, Cz, C4; parietal: P3, Pz, P4) as within-subject factors, Zu-
gether with specific factors for each task. As for behavior,
for the phonological categorization task, these factors were
Order (pre vs. post) and Task (Voicing vs. Vowel length vs.
Pitch vs. Aspiration). For the matching and semantic tasks,
factors were Block (1 vs. 2) and Condition (match vs. mis-
match or related vs. unrelated). Post hoc Tukey tests (Re-
ducing the probability of Type I errors) were used to
determine the origin of significant main effects and interac-
tionen. To simplify results presentation, we only report sig-
nificant results related to our hypotheses (full statistical
results can be seen in Table 1). Endlich, correlation analyses
(Pearson’s coefficient) were computed between error rates
in the musicality task with error rates or N400 effects in the
semantic task. General linear models (including Group as a
categorical factor, error rates in the musicality task as a con-
tinuous factor, and error rates or N400 effects in the seman-
tic task as a dependent factor) were used to test whether
the differences between the slopes and intercepts of the
two groups were significant.
ERGEBNISSE
Statistical Analyses
ANOVAs were computed using Statistica software
( Version 12.0, StatSoft, Inc., Tulsa, OK). For ERRs and
RTs, ANOVAs always included Group (MUS vs. NM) als
between-subject factor as well as specific factors for each
Aufgabe. As the phonological categorization task was the only
Results are presented first for the screening measures of
cognitive ability and musical aptitude and second for the ex-
perimental tasks. For each experimental task, behavioral da-
ta are presented first followed by the ERPs data, except for
the word learning phase in which no behavioral data were
recorded and for the long-term memory tasks in which no
ERPs were recorded. Endlich, for ERPs data (except in the
Dittinger et al.
1589
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
Ö
5
C
4
N
9
_
6
A
/
_
J
0
Ö
0
C
9
N
9
7
_
A
P
_
D
0
0
B
9
j
9
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
/
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Tisch 1. Results of ANOVAs on the ERPs Data in Different Tasks of the Main Experimental Session
Word Learning Phase
Matching Task
Semantic Task
ALL
MUS
NM
ALL
MUS
NM
ALL
MUS
NM
F
P
F
P
F
P
F
P
N400 (340–550 msec)
G
G × B
G × L
G × R
G × B × L
G × B × R
G × L × R
2.72 .11
0.12 .73
0.41 .67
0.13 .88
4.65 .01
1.87 .16
0.67 .62
G × B × L × R
0.82 .52
G × C
G × B × C
G × C × L
G × C × R
G × B × C × L
G × B × C × R
G × C × L × R
G × B × C × L × R
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
3.85
3.23
0.21
0.75
0.07
2.40
1.27
0.40
1.61
0.42
1.25
3.14
0.83
1.63
1.82
1.97
.06
.08
.82
.48
.93
.10
.29
.81
.21
.52
.29
.05
.44
.21
.13
.43
F
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
4.25 .05
1.57
.23
2.72 .12
3.13
.09 <.001
7.05 .002 16.38 <.001 0.10 .91
0.52
1.32 .27
0.42 .80
3.29
0.68
.05
.61
0.12 .89
1.46
0.54 .71
1.06
.60
.24
.38
.95
.29
0.28
0.08
0.84
0.53
0.07
0.01
1.17
9.72 <.001
5.36
B
B × L
B × R
B × L × R
C
C × B
C × L
C × R
C × B × L
C × B × R
C × B × L × R
N200 (230–340 msec)
G
G × B
G × L
G × R
G × B × L
G × B × R
G × L × R
–
–
–
–
–
–
–
–
–
–
–
–
–
–
1.24 .28
0.13 .73
0.60 .55
0.04 .96
1.05 .36
0.70 .50
0.99 .42
G × B × L × R
0.68 .61
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
p
F
p
F
p
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
.99
.76
.93
.50
.48
.79
.01
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
7.68 .02
0.31 .74
5.08 .01
0.54 .71
1.38 .26
2.06 .17
0.93
0.02
0.21
3.14
0.37
0.01
2.23
0.60
1.06
0.84
0.21
0.04
0.01
0.15
1.19
0.52
1.69
0.81
0.36
0.65
0.10
1.46
5.58 .009
1.41
.34
.89
.81
.05
.69
.99
.07
.67
.31
.37
.81
.96
.99
.86
.32
.72
.20
.45
.70
.63
.75
.24
.25
F
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
F
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
0.63
0.25
0.10
0.71
0.65
0.04
0.19
.44
.78
.90
.59
.43
.85
.83
1.11
0.91
0.32
0.51
0.42
2.43
2.25
.31
.42
.73
.73
.53
.14
.12
27.80 <.001 28.34 <.001 6.38 .005 19.18 <.001 8.98 <.001 10.22 <.001
0.47
2.43
1.29
5.56
0.04
0.27
0.87
0.03
0.69
1.55
0.20
.63
.10
.28
.03
.84
.77
.43
.97
.51
.19
.94
1.02
2.99
1.11
.38
.07
.36
0.39 .68
0.09 .91
1.15 .34
0.45
1.12
0.59
.64
.33
.67
0.43
1.36
0.57
.65
.27
.69
0.13
0.18
0.54
.88
.84
.71
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
1.49
7.26
2.41
2.56
0.29
0.85
1.76
0.69
.23
.01
.10
.09
.75
.43
.14
.60
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
1590
Journal of Cognitive Neuroscience
Volume 28, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. (continued )
Word Learning Phase
Matching Task
Semantic Task
ALL
MUS
NM
ALL
MUS
NM
ALL
MUS
NM
G × C
G × B × C
G × C × L
G × C × R
G × B × C × L
G × B × C × R
G × C × L × R
G × B × C × L × R
B
B × L
B × R
B × L × R
C
C × B
C × L
C × R
C × B × L
C × B × R
C × B × L × R
F
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
F
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
F
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
F
2.19
0.28
1.85
1.41
0.14
0.31
1.51
1.99
11.61 .002
6.50
4.44 .02
0.93 .40
0.51 .73
4.14
1.25
0.26
.02
.03
.30
.91
5.11 .04
2.28
0.82 .45
1.62
0.47 .63
0.30
1.33 .27
0.71
p
.15
.60
.17
.25
.87
.74
.21
.10
.14
.21
.74
.58
F
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
F
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
0.81
0.77
0.20
0.37
.38
.47
.82
.83
1.55 .23
0.90 .42
1.33 .28
0.62 .65
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
16.53 <.001 15.22
.002 3.38 .09
0.72
3.02
.40
.06
0.93
4.04
.35
.03
0.05 .82
1.18 .32
20.06 <.001 15.12 <.001 6.12 .006
6.83
.002 5.05
0.21
1.34
.81
.27
0.20
1.13
.82
.34
0.16 .85
0.26 .77
4.25
.003
3.98
.007 1.88 .13
0.28
5.25
1.53
.76
0.57
.008 4.97
.20
1.24
F
1.49
0.004
0.64
0.88
0.37
0.50
2.19
0.69
0.19
1.24
0.26
0.41
4.16
4.32
1.59
p
.23
.95
.53
.42
.70
.61
.08
.60
.67
.30
.77
.80
.05
.05
.21
F
–
–
–
–
–
–
–
–
5.36
0.78
0.58
1.40
4.87
2.00
1.60
p
–
–
–
–
–
–
–
–
.04
.47
.57
.25
.05
.18
.22
.01
.57
.01
.30
F
–
–
–
–
–
–
–
–
2.35
0.75
0.52
0.09
0.37
2.33
0.22
1.89
0.10
1.17
0.94
p
–
–
–
–
–
–
–
–
.15
.48
.60
.99
.55
.15
.80
.17
.90
.33
.45
Although the Condition × Anterior/posterior interactions are significant in musicians (MUS) and in nonmusicians (NM) for the N400 component, the effects are reversed
in both groups (typical N400 effect over parietal sites in MUS, inversed N400 effect over frontal sites in NM). Significant effects are printed in italics, and exact levels of
significance are indicated except when the p values are inferior to .001 (<.001). G = Group; B = Block; C = Condition; L = Laterality; R = Anterior/ Posterior.
phonological categorization task where the N100 compo-
nent is of main interest), analysis of the N400 component
is presented first, followed by analyses of the N200.
Screening Measures
Cognitive Ability
Psychometric data were evaluated by means of univariate
ANOVAs. Results showed no significant Group differences
regarding general reasoning abilities (i.e., progressive
matrices, PM47; F(1, 28) = 1.37, p = .25), verbal working
memory (reverse digit span; F(1, 28) = 2.88, p = .10),
nor visual attention (F(1, 28) = 3.17, p = .09). By con-
trast, MUS (mean = 7.6, SD = 0.30) showed better
short-term memory abilities than NM (mean = 6.7, SD =
0.30; direct digit span; F(1, 28) = 5.53, p = .03).
Musical Aptitude
A 2 × 2 ANOVA (i.e., 2 Groups × 2 Tasks) showed that
MUS made fewer errors (6.7%, SD = 2.0) than NM
(17.6%, SD = 2.0; main effect of Group: F(1, 28) =
14.71, p < .001), and all participants performed better
on the rhythmic (9.8%, SD = 1.6) than on the melodic
task (14.4%, SD = 2.0; main effect of Task: F(1, 28) =
4.19, p = .05) with no Group × Task interaction.
Experimental Tasks
Phonological Categorization Task
Behavioral data. Results of 2 × 2 × 4 ANOVAs [i.e., 2
Groups (MUS vs. NM) × 2 Orders (pre vs. post) × 4
Tasks ( Voicing vs. Vowel length vs. Pitch vs. Aspiration)]
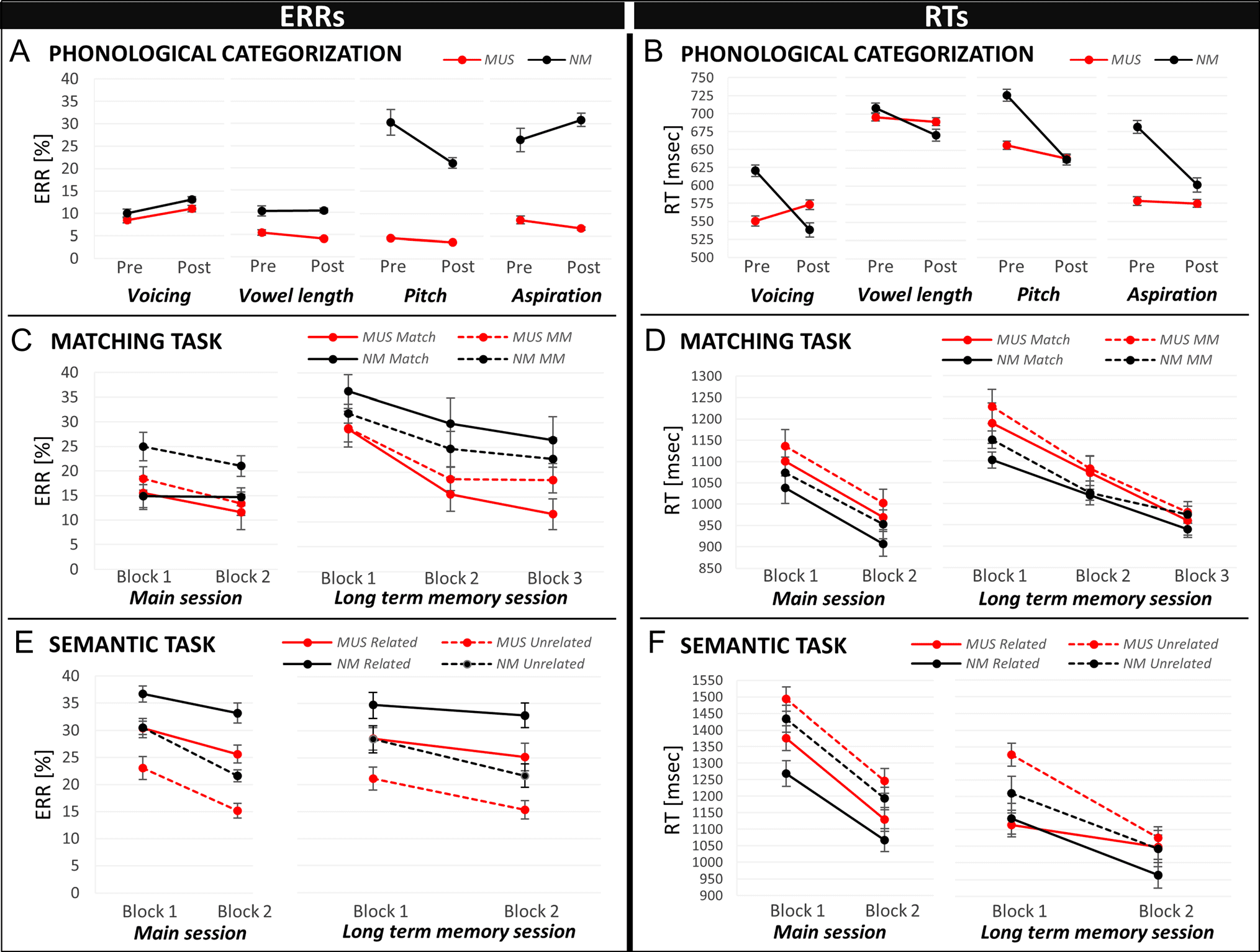
showed that MUS (6.6%, SD = 2.2) made overall fewer
errors compared with NM (19.1%, SD = 2.2; main effect
of Group: F(1, 24) = 16.29, p < .001; Figure 2A). The
influence of music training was largest in the pitch (MUS:
4.1%, SD = 4.8; NM: 25.8%, SD = 4.9; Tukey, p < .001) and
aspiration tasks (MUS: 7.6%, SD = 5.5; NM: 28.7%, SD =
5.5; Tukey, p < .001; Group × Task interaction: F(3,
72) = 11.82, p < .001). Finally, only NM improved their
level of performance from pre to post in the pitch task
(Group × Task × Order interaction: F(3, 72) = 3.31,
Dittinger et al.
1591
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
p = .03; NM: pre: 30.3%, SD = 4.1, post: 21.3%, SD = 3.3,
Tukey, p = .02; MUS: pre: 4.5%, SD = 4.1, post: 3.7%, SD =
3.3, Tukey, p = .99). Analyses of RTs did not reveal sig-
nificant Group differences (Figure 2B).
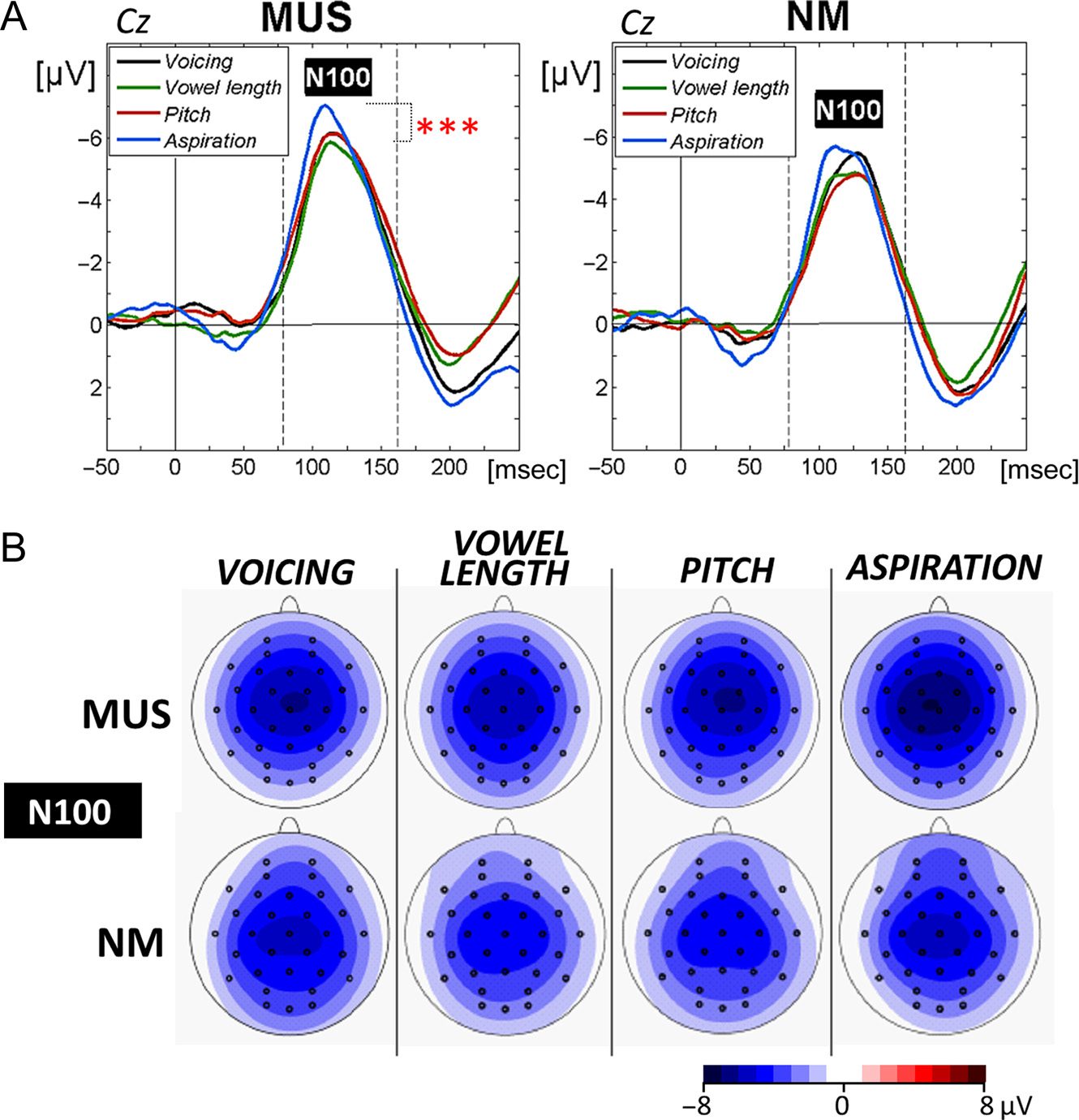
Electrophysiological data. N100 amplitudes were eval-
uated by means of a 2 × 2 × 4 × 3 × 3 ANOVA (i.e., 2
Groups × 2 Orders × 4 Tasks × 3 Laterality positions
[left vs. midline vs. right] × 3 Anterior/Posterior posi-
tions [frontal vs. central vs. parietal]). Results revealed
a significant Group × Task × Laterality interaction effect
(F(6, 168) = 3.19, p = .005). Separate ANOVAs for each
group showed that only MUS were characterized by a
larger N100 in the aspiration task (MUS: −6.50 μV, SD =
2.59 and NM: −5.57 μV, SD = 1.81) compared with
the other three tasks over the left hemisphere and at
midline electrodes (Task × Laterality interaction: MUS:
−5.47 μV, SD = 1.78; F(6, 84) = 3.13, p = .008, Tukey,
both ps < .001; NM: −5.01 μV, SD = 1.74; F < 1; Fig-
ure 3A and B).
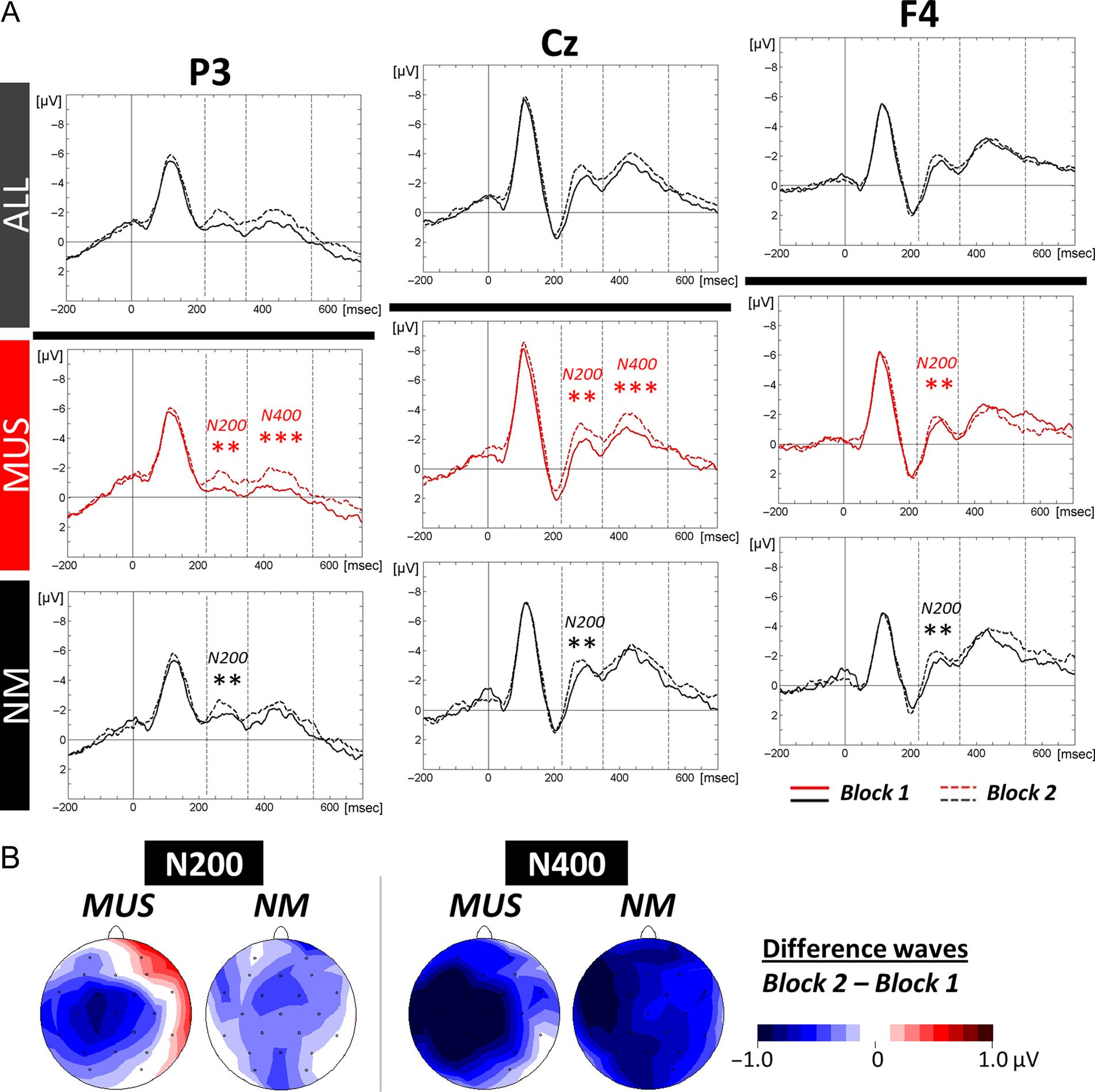
Word Learning Phase
Electrophysiological data. The N400 as well as the
N200 were evaluated by means of 2 × 2 × 3 × 3 ANOVAs
(i.e., 2 Groups × 2 Blocks [1 vs. 2] × 3 Laterality × 3
Anterior/Posterior positions).
For all participants and in line with previous results, the
N400 component was larger over frontal (−2.63 μV, SD =
0.87) and central (−2.52 μV, SD = 0.76) sites compared
with parietal sites (−1.29 μV, SD = 0.66; Tukey, both
ps < .001; main effect of Anterior/Posterior: F(2, 56) =
25.25, p < .001). In addition, the Group × Block × Later-
ality interaction effect was significant (F(2, 56) = 4.65,
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Percentages of errors (ERRs) and RTs in the different tasks are shown for musicians (MUS) in red and for nonmusician controls (NM) in
black. For the phonological categorization task (A and B), results are illustrated for the premeasurement (beginning) and postmeasurement (end
of main experimental session), separately for each task (voicing, vowel length, pitch, and aspiration). For the matching task (C and D), results
for Match (solid lines) and Mismatch (MM: dotted lines) words are illustrated in the two blocks of the main experimental session and in the
three blocks of the long-term memory session. For the semantic task (E and F), results for semantically Related (solid lines) and Unrelated
(dotted lines) words are illustrated in the two blocks of the main experimental session and of the long-term memory session.
1592
Journal of Cognitive Neuroscience
Volume 28, Number 10
Figure 3. Phonological
categorization. (A) N100
components at the Central (Cz)
electrode are compared between
tasks for musicians (MUS) and
for nonmusician controls (NM).
In this and subsequent figures,
time in milliseconds is in
abscissa and the amplitude of the
effects in microvolt is in ordinate.
Time zero corresponds to word
onset and negativity is plotted
upwards. Latency windows
for statistical analyses are
indicated with gray dotted lines
and the level of significance is
represented by stars with *p <
.05, **p < .01, and ***p < .001
(red stars for MUS and black
stars for NM). (B) Topographic
voltage distribution maps
illustrate the N100s to the
words separately for each task
and for MUS and NM. Voltage
values are scaled from −8 to
+8 μV.
p = .01). Separate group analyses showed that only
MUS showed significantly increased amplitudes from
Block 1 to Block 2. This effect was localized over the
left hemisphere and midline electrodes (MUS: Block 1:
−1.54 μV, SD = 0.76 and Block 2: −2.16 μV, SD =
0.79; Block × Laterality interaction: F(2, 28) = 16.38,
p < .001; Tukey, both ps < .001 and NM: Block 1:
−2.34 μV, SD = 1.36 and Block 2: −2.91 μV, SD = 1.38;
main effect of Block: F(1, 14) = 2.72, p = .12; Fig-
ure 4A and B).
Analyses of the N200 component did not reveal signif-
icant Group differences (main effect of Group: F(1, 28) =
1.24, p = .28), but all participants showed significantly
increased amplitudes from Block 1 (−1.13 μV, SD =
1.22) to Block 2 (−1.79 μV, SD = 0.99; main effect of
Block: F(1, 28) = 11.61, p = .002; Figure 4A and B).
Matching Task
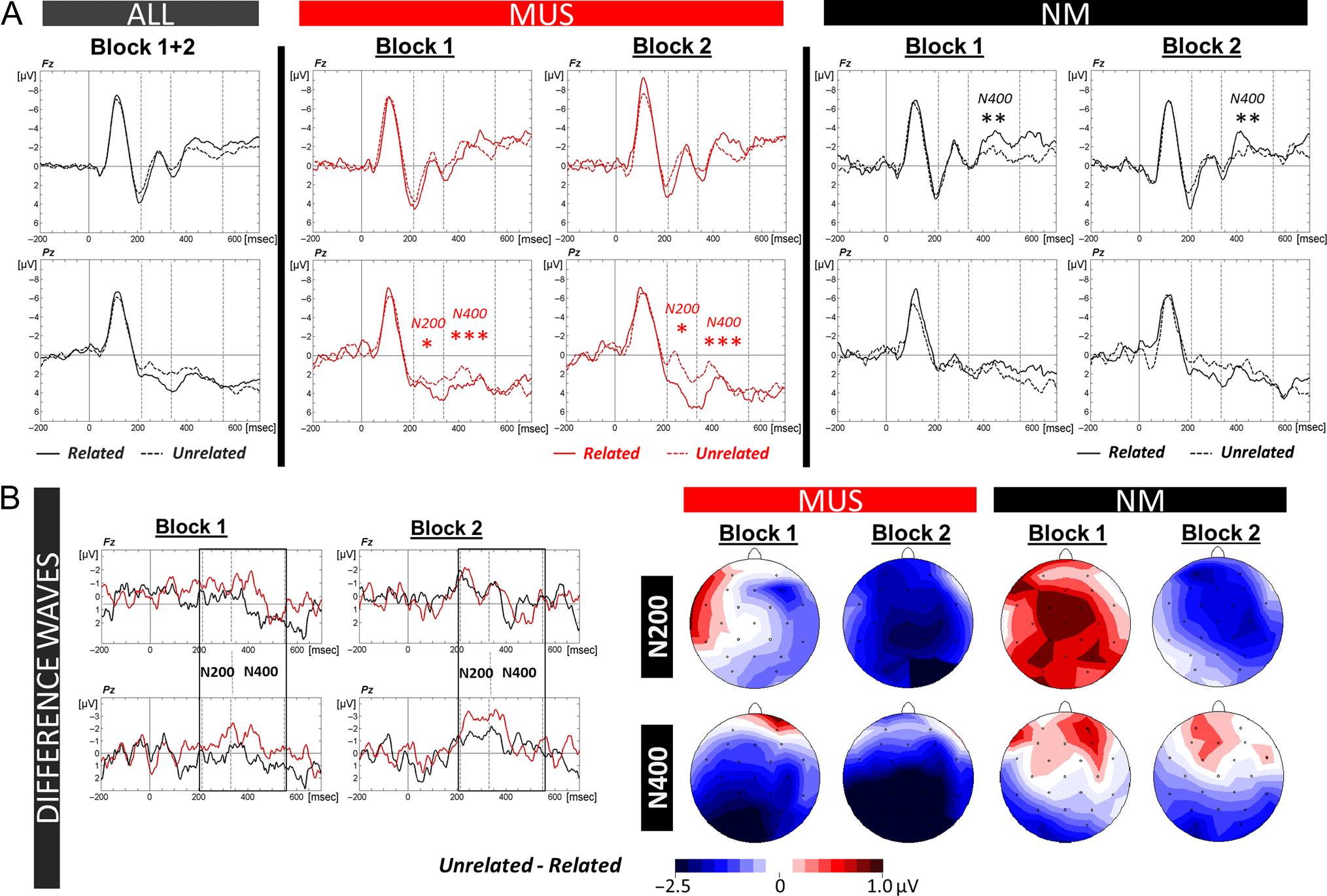
Behavioral data. Results of three-way ANOVAs (i.e., 2
Groups × 2 Blocks × 2 Conditions [match vs. mismatch])
showed that ERRs did not significantly differ between the
two groups (main effect of Group: F(1, 24) = 2.19, p =
.15). However, all participants committed overall fewer er-
rors for match (14.2%, SD = 2.6) compared with mismatch
words (19.4%, SD = 2.2) and fewer errors in Block 2
(15.2%, SD = 2.1) than in Block 1 (18.5%, SD = 2.2; main
effect of Condition: F(1, 24) = 7.68, p = .01; main effect of
Block: F(1, 24) = 9.27 p = .006; Figure 2C). In line with
ERRs, analyses of RTs did not reveal between-group differ-
ences (main effect of Group: F < 1), but overall faster RTs
for match (1041 msec, SD = 64) than for mismatch words
(1080 msec, SD = 70; main effect of Condition: F(1, 24) =
5.90, p = .02) and faster RTs in Block 2 (994 msec, SD =
61) than in Block 1 (1128 msec, SD = 73; main effect of
Block: F(1, 24) = 60.45, p < .001; Figure 2D).
Electrophysiological data. The N400 as well as the N200
component were evaluated by means of 2 × 2 × 2 × 3 × 3
ANOVAs (2 Groups × 2 Blocks × 2 Conditions × 3 Later-
ality × 3 Anterior/Posterior positions).
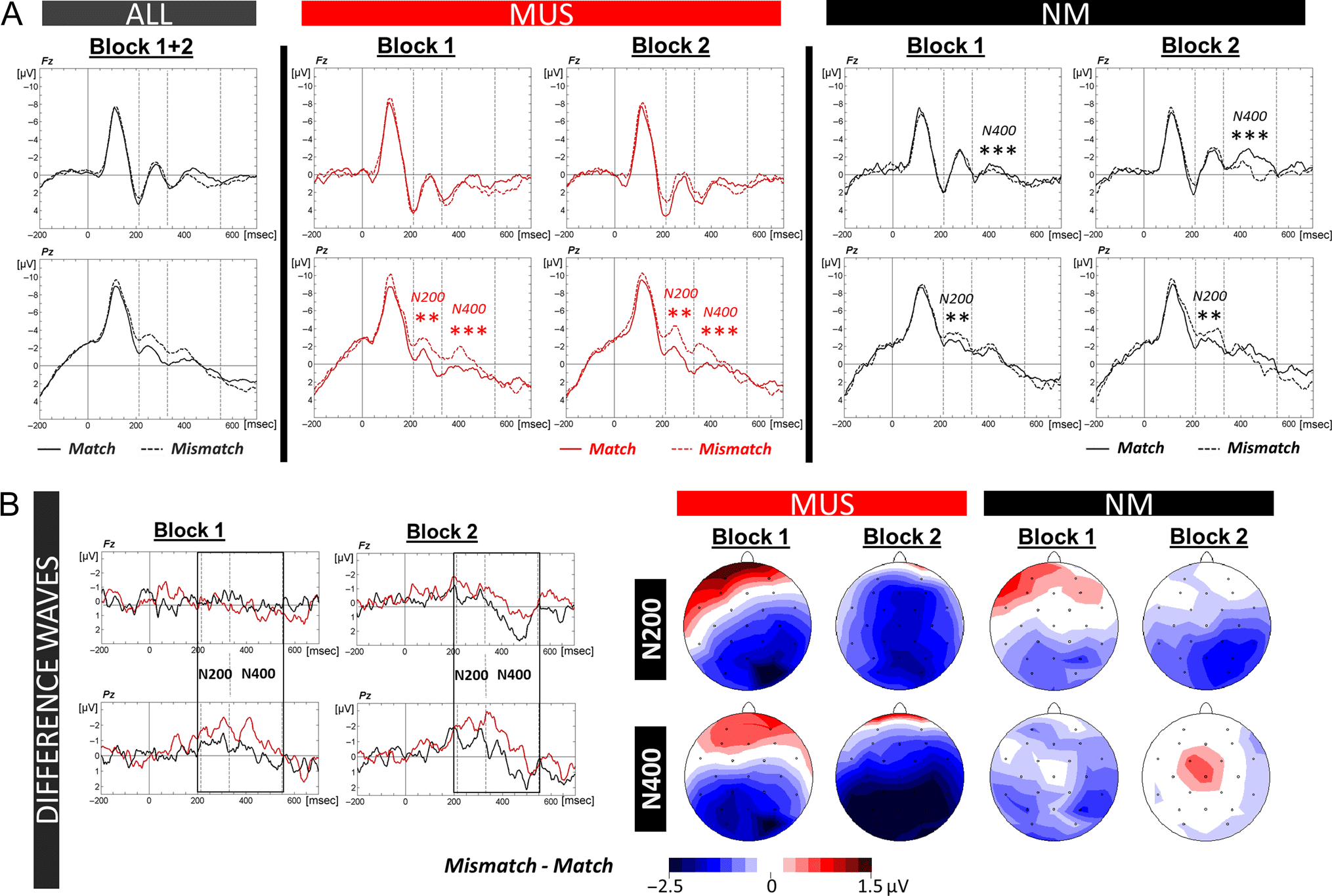
Analysis of the N400 revealed a significant Group × Con-
dition × Anterior/Posterior interaction effect (F(2, 56) =
3.14, p = .05). Results of separate group analyses showed
larger N400 amplitudes in MUS for mismatch (−0.10 μV,
SD = 1.82) compared with match words over centroparietal
regions (0.67 μV, SD = 1.58; Condition × Anterior/Posterior
interaction: F(2, 28) = 28.34, p < .001; Tukey, central: p = .02;
parietal: p < .001). The opposite pattern was found in NM
with larger N400 for match (−1.34 μV, SD = 1.42) than for
Dittinger et al.
1593
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
mismatch words over frontocentral sites (−0.89 μV, SD =
1.53; Condition × Anterior/Posterior interaction: F(2, 28)
= 6.38, p = .005; Tukey, frontal: p = .001; central: p =
.03; Figure 5A and B).
The N200 amplitude was overall smaller in MUS (−0.15 μV,
SD = 0.60) compared with NM (−2.14 μV, SD = 0.60; main
effect of Group: F(1, 28) = 5.56, p = .03) and the N200
effect (i.e., mismatch minus match words) was more widely
distributed in MUS compared with NM (Group × Block ×
Condition × Laterality × Anterior/Posterior interaction: F(4,
112) = 1.99, p = .10; Figure 5A and B). MUS showed larger
N200 amplitudes for mismatch (P4: −1.98 μV, SD = 0.76)
than for match words (P4: 0.01 μV, SD = 0.68) over centro-
parietal scalp sites with largest differences over midline and
right hemisphere (Condition × Anterior/Posterior interac-
tion: F(2, 28) = 15.12, p < .001; Condition × Laterality in-
teraction: F(2, 28) = 4.04, p = .03). In addition, the N200
effect was larger in Block 2 (P4: −2.22 μV, SD = 0.50) than
in Block 1 (P4: −1.66 μV, SD = 0.63) over midline and right
centroparietal sites (Condition × Block × Laterality ×
Anterior/Posterior interaction: F(4, 56) = 3.98, p = .007).
NM also showed an N200 effect that was localized over pa-
rietal sites (−1.06 μV, SD = 1.41; Condition × Anterior/
Posterior interaction: F(2, 28) = 6.12, p = .006).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Word learning phase. (A) ERPs recorded at left parietal (P3), central (Cz), and right frontal sites (F4) in Block 1 (solid line) and Block 2
(dotted line) are overlapped for all participants (ALL: black lines, top), and separately below for musicians (MUS: red lines) and nonmusician
controls (NM: black lines). (B) Topographic voltage distribution maps of the differences between the two blocks (Block 2 minus Block 1) are illustrated
for the ERP components of interest (N200, N400), separately for MUS and for NM. Voltage values are scaled from −1.0 to +1.0 μV.
1594
Journal of Cognitive Neuroscience
Volume 28, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Matching task. (A) Left: ERPs recorded at frontal (Fz) and parietal (Pz) sites are overlapped for Match (solid lines) and Mismatch (dotted lines)
words for all participants across the two blocks of trials (ALL: black lines). Central and right: ERPs are presented separately for Block 1 and Block 2 and
for musicians (MUS: red lines) and nonmusician controls (NM: black lines). (B) Difference waves (Mismatch minus Match) are overlapped for MUS (red)
and NM (black) separately for Block 1 and for Block 2 at Fz and Pz. Topographic voltage distribution maps of the Mismatch minus Match differences are
illustrated for N200 and N400 components separately and for MUS and NM in Block 1 and Block 2. Voltage values are scaled from −2.5 to +1.5 μV.
Semantic Task
Behavioral data. Results of three-way ANOVAs (i.e., 2
Groups × 2 Blocks × 2 Conditions [related vs. unrelated])
showed that MUS (23.6%, SD = 2.0) made overall fewer
errors than NM (30.5%, SD = 2.0; main effect of Group:
F(1, 24) = 5.82, p = .02), and all participants made fewer
errors for unrelated (22.6%, SD = 2.6) than for related
words (31.5%, SD = 2.9; main effect of Condition: F(1,
24) = 11.24, p = .003; Figure 2E). Moreover, all par-
ticipants made fewer errors in Block 2 (30.2%, SD = 2.3)
than in Block 1 (23.9%, SD = 2.4; main effect of Block:
F(1, 24) = 12.37, p = .002). RTs were faster for related
(1210 msec, SD = 72) than for unrelated words (1342 msec,
SD = 71; main effect of Condition: F(1, 24) = 41.32, p <
.001) and faster in Block 2 (1159 msec, SD = 70) than in
Block 1 (1393 msec, SD = 75; main effect of Block: F(1,
24) = 88.92, p < .001), with no between-group differences
(main effect of Group: F < 1; Figure 2F).
Electrophysiological data. The N400 as well as the
N200 component were evaluated by means of 2 × 2 ×
2 × 3 × 3 ANOVAs (i.e., 2 Groups × 2 Blocks × 2 Con-
ditions × 3 Laterality × 3 Anterior/Posterior positions).
N400 analyses revealed a significant Group × Anterior/
Posterior interaction effect (F(2, 56) = 3.14, p = .05). As
typically reported in the literature (Kutas et al., 1988), the
N400 was larger for semantically unrelated (2.17 μV, SD =
1.93) compared with related words (3.29 μV, SD = 1.66)
over parietal sites in MUS (Condition × Anterior/Posterior
interaction: F(2, 28) = 8.98, p < .001; Tukey, parietal: p <
.001). By contrast, a reversed N400 effect was found in NM
with larger N400 for related (−2.09 μV, SD = 1.60) than
for unrelated words (−1.19 μV, SD = 1.06) over frontal
sites (Condition × Anterior/Posterior; F(2, 28) = 10.22,
p < .001; Tukey, frontal: NM: p = .002, Figure 6A and B).
The N200 amplitude was larger in Block 2 than in
Block 1 in MUS only (Group × Block: (F(1, 28) = 7.26,
p = .01; MUS: Block 1: 1.58 μV, SD = 2.62 and Block 2:
0.89 μV, SD = 2.68; F(1, 14) = 5.35, p = .04 and NM:
Block 1: −0.11 μV, SD = 3.15 and Block 2: 0.39 μV,
SD = 2.58; F(1, 14) = 2.35, p = .15; Figure 6A and B).
In addition, the N200 was also larger for unrelated than
for related words in MUS but not in NM (main effect of
Dittinger et al.
1595
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
Figure 6. Semantic task. (A) Left: ERPs recorded at frontal (Fz) and parietal sites (Pz) are overlapped for semantically Related (solid lines) and
Unrelated (dotted lines) words for all participants across the two blocks of trials (ALL: black lines). Central and right panels: ERPs are presented
separately for Block 1 and Block 2 and for musicians (MUS: red lines) and nonmusician controls (NM: black lines). (B) Difference waves (Unrelated
minus Related) are overlapped for MUS (red) and NM (black) separately for Block 1 and Block 2 at Fz and Pz. Topographic voltage distribution maps
of the Unrelated minus Related differences are illustrated for N200 and N400 components separately and for MUS and NM in Block 1 and Block 2.
Voltage values are scaled from −2.5 to +1.0 μV.
Condition: MUS: Related: 1.88 μV, SD = 2.67 and Unre-
lated: 0.59 μV, SD = 3.03; F(1, 14) = 4.87, p = .05 and NM:
Related: 0.30 μV, SD = 3.63 and Unrelated: −0.02 μV,
SD = 2.23; F < 1; Group × Condition × Laterality ×
Anterior/Posterior interaction: F(4, 112) = 2.19, p = .08),
and this effect was larger over central and parietal sites
(Condition × Anterior/Posterior interaction: F(2, 28) =
5.05, p = .01; Tukey, both ps < .001).
Figure 7. Correlation analyses.
Correlations between the
percentages of error in the
musicality test (ERRs Musicality)
and the sizes of the N400 effect
(Unrelated minus Related) in
the semantic task are illustrated
for musicians (MUS: red) and
for nonmusician controls
(NM: black). Dotted lines
represent the 95% confidence
interval of the correlation line.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1596
Journal of Cognitive Neuroscience
Volume 28, Number 10
Long-term Memory: Matching Task (Behavior)
Results of three-way ANOVAs (i.e., 2 Groups × 3 Blocks ×
2 Conditions [match vs. mismatch]) showed that MUS
(20.3%, SD = 2.8) made fewer errors compared with NM
(28.6%, SD = 2.9; main effect of Group: F(1, 19) = 4.19,
p = .05; Figure 2C). Moreover, all participants improved
their level of performance from Block 1 (31.4%, SD =
2.6) to Blocks 2 and 3 (22.2%, SD = 3.4 and 19.8%, SD =
3.6, respectively; main effect of Block: F(2, 38) = 26.40, p <
.001). No significant between-group differences were
found on RTs but in both groups, RTs were faster in Block
3 (965 msec, SD = 67) than in Block 2 (1049 msec, SD =
72) and slowest in Block 1 (1168 msec, SD = 82; main ef-
fect of Block: F(2, 38) = 36.48, p < .001; Figure 2D).
Long-term Memory: Semantic Task (Behavior)
Results of three-way ANOVAs (i.e., 2 Groups × 2 Blocks ×
2 Conditions [related vs. unrelated]) showed that between-
group differences were not significant on ERRs nor on
RTs. However, all participants made fewer errors for un-
related (21.5%, SD = 6.0) than for related words (29.7%,
SD = 6.3; main effect of Condition; F(1, 19) = 6.60, p =
.02) and were faster for related (1055 msec, SD = 106)
than for unrelated words (1151 msec, SD = 119; main
effect of Condition; F(1, 19) = 14.54, p = .001; Figure 2E
and F).
Relationships between Musical Aptitude, Behavioral
Data, and Brain Activity
Error rates in the musicality test were correlated with the
size of the N400 effects in the semantic task for MUS (r =
.74, p = .01) but not for NM (r = .01, p = .97;
Figure 7A and B). Moreover, the slopes of the correla-
tions were significantly different for the two groups (main
effect of Group: F(1, 26) = 7.36, p = .01) as well as the cor-
relation intercepts (Group × Musicality interaction: F(2,
26) = 6.52, p = .005).
DISCUSSION
Summary of Results
By using an ecologic valid experimental design we tested
the general hypothesis that professional musicians would
learn the meaning of novel words more efficiently than
control participants. Overall, both behavioral and electro-
physiological data support this hypothesis. Behaviorally,
musicians performed better than controls in the musicality
and phonological categorization tasks. In addition, al-
though all participants performed similarly in the matching
task, musicians made significantly fewer errors in the se-
mantic task. Finally, after 5 months, musicians remem-
bered more words than controls as reflected by lower
error rates in the matching task.
The electrophysiological markers of word learning also
clearly differed between the two groups. Although con-
trol participants showed similar N100 amplitudes in all
phonological categorization tasks, musicians showed an
increase in N100 amplitude when categorizing the most
difficult aspiration contrast. Most importantly and in line
with the word learning literature, both groups showed
enhanced N400 amplitudes over frontal scalp sites when
learning the meaning of novel words. However, only
musicians were additionally characterized by larger left-
lateralized N400s in the second block compared with
the first block of the word learning phase. Finally, only
musicians showed the typical centroparietal distribution
of the N400 effect in the matching and semantic tasks.
By contrast, nonmusicians showed reversed N400 effects
in both tasks over frontal sites. These findings are dis-
cussed in detail below. It is, however, important to note
that cross-sectional studies, as the one reported here,
are necessary to first demonstrate differences between
musically trained and untrained participants before de-
signing longitudinal experiments to test for the causality
of the reported effects.
Spatiotemporal Dynamics in the Learning Phase
Results showed clear evidence for fast brain plasticity, as
reflected by the rapid development of the N400 in both
groups of participants after only 3 min of novel word
learning (Block 1), that is after 10 repetitions of each
picture–word association (see Figure 4). This finding
extends previous results on word learning showing
N400 enhancements when learning a second language
(McLaughlin et al., 2004), the meaning of rare words
(Perfetti et al., 2005), and when learning the meaning
of novel words or pseudowords from highly constrained
sentence contexts (Borovsky et al., 2010, 2012; Batterink
& Neville, 2011; Mestres-Missé et al., 2007). Importantly,
and in line with previous work in adults (Borovsky et al.,
2012; Mestres-Missé et al., 2007) and in children (François
et al., 2013; Friedrich & Friederici, 2008), in both musi-
cians and controls the N400 component to novel words
was larger frontocentrally than parietally. These results
are compatible with previous findings, suggesting that pre-
frontal and temporal brain regions are associated with the
maintenance of novel information in working memory
(Hagoort, 2014) and the acquisition of word meaning
(Rodriguez-Fornells et al., 2009).
Importantly, however, the N400 increase from the first
to the second block of novel word learning was only sig-
nificant in musicians (see Figure 4A). In line with our hy-
pothesis, this is taken to suggest faster encoding of novel
word meaning in musicians than in controls. In addition,
word learning in musicians was accompanied by a rapid
shift of the N400 component distribution from frontal to
centroparietal sites. This shift in distribution is in line with
the hypothesis that musicians have already integrated novel
words representations in semantic memory (Batterink &
Dittinger et al.
1597
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Neville, 2011) and that the N400 centroparietal distribution
in musicians reflects access to semantic memory (Kutas &
Federmeier, 2011). By contrast, in control participants, the
N400 component remained larger frontally throughout the
entire learning phase, suggesting that nonmusicians had
not yet integrated the novel words’ meaning into estab-
lished semantic networks. This interpretation can be di-
rectly tested in future experiments by increasing the
number of repetitions of picture–word associations. Under
such conditions, a frontal to centroparietal shift in N400
distribution should also be found in nonmusicians.
Evidence for Rapidly Established Representations
of Novel Words in the Matching and
Semantic Tasks
As expected, all participants, whether musicians or con-
trols, were able to learn the nine picture–word associa-
tions, as reflected by error rates below chance level (i.e.,
50% errors) in both the matching (mean = 17%) and se-
mantic tasks (mean = 27%; see Figure 2C and E). To follow
the dynamics of word learning within each task, results
were compared between the first and the second block
of trials. Interestingly, the level of performance increased
with repetition in both tasks, indicating a still ongoing
learning effect. Moreover, both musicians and controls
showed clear matching effects with lower error rates and fas-
ter RTs for match compared with mismatch words (Boddy
& Weinberg, 1981). However, and in contrast to typical se-
mantic priming effects (Meyer & Schvaneveldt, 1971), both
groups of participants made more errors for semantically
related words than for semantically unrelated words. Al-
though unexpected, this result may reflect a response bias
towards rejection (i.e., considering the word as unrelated
to the picture) as a consequence of task difficulty generat-
ing higher uncertainty (Gigerenzer, 1991). In other words,
when participants were not certain whether the pictures
and the words were semantically related (e.g., “honey”
and “bear”), they tended to respond that they were unre-
lated. By contrast, results for RTs conform to the literature
(i.e., faster RTs for semantically related than for unrelated
words; Meyer & Schvaneveldt, 1971). Although the pres-
ence of a speed–accuracy trade-off limits the interpretation
of this priming effect, faster RTs for semantically related
words are indicative that new pictures that had not been
seen before in the experiment did activate the representa-
tions of semantically related newly learned words.
Turning to the influence of music training, musicians
and nonmusician controls performed similarly in the
matching task but musicians outperformed controls in
the semantic task. This contrastive pattern of results sug-
gests that the two tasks tap into different memory sys-
tems. To decide whether the newly learned word
matched the picture in the matching task, participants
had to retrieve the specific picture–word associations
that were stored in episodic memory during the word
learning phase. By contrast, in the semantic task partici-
pants had to retrieve general information from semantic
memory because the novel pictures that were presented
before the newly learned words had not been seen be-
fore in the experiment. In line with the centroparietal
shift in N400 distribution observed at the end of the word
learning phase, the finding that musicians outperformed
nonmusicians in the semantic task is taken as evidence
that musicians had already integrated the novel words’
meanings into semantic memory so that priming effects
generalized to new pictures.
ERPs in the matching and semantic tasks also add sup-
port to this interpretation. In musicians and for both
tasks, the N400 over centroparietal sites was larger for
unexpected (mismatch/unrelated) than for expected
words (match/related; Figures 5 and 6). This sensitivity
to word characteristics and this scalp distribution corre-
spond to the N400 component, typically considered as the
electrophysiological marker of the integration of novel
words’ meanings into semantic memory (Borovsky et al.,
2012; Batterink & Neville, 2011; Mestres-Missé et al., 2007)
and “as reflecting the activity in a multimodal long-term
memory system that is induced by a given input stimulus
during a delimited time window as meaning is dynamically
constructed” (Kutas & Federmeier, 2011, p. 22; see also
Steinbeis & Koelsch, 2008, for N400s elicited by single
chords incongruous with the preceding context). By con-
trast, in nonmusician controls, the N400 effect was re-
versed over frontocentral sites in both tasks, with larger
N400 for expected (match/related) than for unexpected
words (mismatch/unrelated). This finding was surprising
based on previous results showing that the N400 is larger
for unrelated than for related words in both lexical deci-
sion tasks (Borovsky et al., 2012) and semantic priming ex-
periments (Mestres-Missé et al., 2007). Because the
amount of music training was not controlled in these ex-
periments, a possible explanation is that some of the par-
ticipants had musical skills, hence influencing the results.
However, stimulus and task differences are more likely to
account for this discrepancy. Most experiments on ERPs
and word learning in adults used similar designs with novel
words embedded in sentence contexts. Here and as in ex-
periments conducted with children (Friedrich & Friederici,
2008; Torkildsen et al., 2008) and with adults (Dobel et al.,
2009), participants learned novel words through picture–
word associations. It is thus possible that this learning
mode was more demanding than learning the meaning
of novel words from sentence contexts. Along these lines,
we interpret the reversed N400 effect in nonmusicians in
the matching and semantic tasks as showing that nonmu-
sicians had not yet fully learned the picture–word associa-
tions (as reflected by the frontal distribution of the N400 in
these tasks that is similar to the frontal N400 found during
word learning) and that they were still building up new ep-
isodic memory traces based on the correct information
provided by the matching words (as reflected by larger
N400 increase from the first to the second block for match
than for mismatch words; see Figure 5A). As mentioned
1598
Journal of Cognitive Neuroscience
Volume 28, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
above, these interpretations can be tested by increasing the
number of trials in the learning phase as well as in the
matching and semantic tasks and by comparing different
word learning designs (e.g., novel words in sentence con-
texts vs. picture–word associations).
Finally, although the main focus of this experiment was
on the modulations of N400 amplitude with novel word
learning, results also revealed interesting effects on the
N200 component. In both groups, the N200 components
in the matching task were larger for mismatching than for
matching words over parietal sites (see Figure 5). Insofar
as the N200 has been associated with categorization pro-
cesses (Friedrich & Friederici, 2008), mismatching words
possibly required more effortful categorization processes
than matching words. However, there is also evidence
that the N200 component reflects early contextual influ-
ences (van den Brink, Brown, & Hagoort, 2001) and pho-
nological processing (Connolly & Phillips, 1994). Thus,
mismatching words possibly elicited larger N200 than
matching words because they were unexpected based
on the picture context and/or at the phonological level.
In the semantic task, the increase in N200 amplitude
for semantically unrelated compared with related words
was only found in musicians, again suggesting that word
categorization was possibly more efficient in musicians
and/or that musicians were more sensitive to the context
(i.e., the picture) or to the phonological characteristics of
novel words than nonmusicians.
Evidence for Long-lasting Representations of
Novel Words
To our knowledge, this is the first experiment comparing
long-term memory (after 5 months) for novel words in
musicians and controls. Clearly, results need to be con-
sidered with caution because of the high dropout rate.
Nevertheless, they point to improved long-term memory
in musicians compared with controls in the matching
task (lower error rates) that was always performed first.
The memory traces of the words that have been learned
5 months before therefore seem stronger in musicians
than in nonmusicians. By contrast, no between-group dif-
ferences were found in the semantic task possibly be-
cause both groups of participants similarly benefited
from the reactivation of memory traces during the match-
ing task. Taken together, these results point to long-
lasting effects of rapidly established word representations
during word learning, and they open new perspectives
to further test for the influence of music training on
long-term memory.
Evidence for Transfer Effects from Music Training
to Word Learning
In summary, behavioral and electrophysiological data
showed that music training improved several aspects of
word learning. How can we account for the influence
of music training on novel word learning?
Cascading Effects from Perception to Word Learning
The first interpretation, in terms of cascading effects, is
that enhanced auditory perception and auditory atten-
tion (Strait et al., 2015) in musicians drive the facilitation
observed in word learning through different subsequent
steps (i.e., building up new phonological representations
and attaching meaning to them, storing this new informa-
tion in short- and long-term memory). In support of this
interpretation, the error rate in the musicality test was cor-
related with the size of the N400 effect in the semantic task
in musicians but not in controls (Figure 7), thereby clearly
pointing to a relationship between auditory perception/
attention and word learning.
Moreover, in line with the hypothesis of improved
speech discrimination in musicians when the task is most
difficult (Diamond, 2013; Schön et al., 2004), musicians
outperformed control participants in the phonological
categorization of tonal and aspiration contrasts, but both
groups performed equally well for simple voicing con-
trasts (/Ba/ vs. /Pa/). Thus, music training was most useful
to discriminate phonemes that are contrastive in Thai but
that do not belong to the French phonemic repertoire
(Figure 2A). It should be noted in this respect that, al-
though vowel length is not contrastive in French, controls
nevertheless performed as well as musicians in this task,
possibly because the natural difference in vowel duration
was large enough (270 msec) to be easily perceived by non-
musician controls. Taken together, these findings add sup-
port to the hypothesis of transfer effects between music
and speech (Asaridou & McQueen, 2013; Besson et al.,
2011), with near transfer for acoustic cues common to mu-
sic and speech such as frequency and with far transfer to
unfamiliar linguistic cues such as aspiration.
Note also that the N100 amplitude in musicians was en-
hanced for the nonnative aspiration contrast (see Figure 3).
Because this task was more difficult for French native
speakers than the other tasks, as revealed by behavioral
data, the increased N100 may reflect increased focused
attention (Strait et al., 2015) and mobilization of neuro-
nal resources in musicians. An alternative but com-
plementary interpretation is that this increase in N100
amplitude reflected increased neural synchronicity and
structural connectivity in musicians, who are typically more
sensitive to the acoustic-phonetic properties of speech
sounds than nonmusicians who showed similar N100 com-
ponents in all tasks (Bidelman et al., 2014; Elmer et al.,
2013; Chobert et al., 2012; Musacchia et al., 2007; Wong
& Perrachione, 2007).
Finally, and in line with the cascade hypothesis, it may
be that the very nature of the stimuli (monosyllabic Thai
words) is at least partly responsible of the musician’s ad-
vantage in the semantic task. Because musicians are
more sensitive than nonmusicians to variations in pitch,
Dittinger et al.
1599
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
duration, and VOT, this possibly helped them build pre-
cise phonological representations that were then more
easily associated to a novel word meaning. One way to
test for this interpretation would be to use a similar de-
sign but with written instead of spoken words thereby
potentially reducing the musicians’ acoustic-phonetic
advantage.
Multidimensional Facilitation Effects
The second interpretation, that is by no means contradic-
tory but rather complementary to the cascading interpre-
tation, is that the multidimensional nature of music
training independently improved the several functions
that are necessary for word learning. There is already
some evidence in the literature for enhanced working
and verbal memory (George & Coch, 2011; Ho et al.,
2003) and executive functions (Zuk et al., 2014; Moreno
et al., 2011; Rogalsky et al., 2011; Pallesen et al., 2010)
with musicianship. In line with these results, musicians
showed enhanced auditory short-term memory (digit
span; George & Coch, 2011). Moreover, as reported in
previous experiments, we also found significant be-
tween-group differences in the ERP component related
to semantic integration and memory (N400; Mestres-
Missé et al., 2007; Perfetti et al., 2005; McLaughlin
et al., 2004). However, in contrast to previous results,
we found no evidence for increased working memory
(reversed digit span; George & Coch, 2011) or for im-
proved nonverbal intelligence (PM47) in adult musicians.
Note that the assessment of general cognitive abilities
was quite limited in the present experiment because of
time constraints. It is thus possible that general cognitive
abilities other than the ones tested here could have influ-
enced the observed between-group differences in word
learning. Future experiments will aim at including more
tests targeting at elucidating different cognitive functions
(selective and sustained attention, working, short-term,
and long-term memory, executive functions). Finally, it
could also be argued that musicians performed better
than nonmusicians because they were generally more
motivated (Corrigall, Schellenberg, & Misura, 2013). Al-
though this is difficult to control for, it is important to
note that participants were not informed about the aim
of the experiment until the end of the session. Only that
we were interested in language, music, and the brain.
Thus, as they discovered task by task what the experi-
mental session was about, it is unlikely that general
motivation accounted for the present behavioral and
electrophysiological effects.
Conclusions
In summary, by recording ERPs during both learning and
test phases (matching and semantic tasks) of novel word
learning, results revealed fast changes in brain activity after
only a few minutes of exposure to picture–word associa-
tions. Moreover, these changes were stronger in musi-
cians than in controls. Specifically, the frontoparietal
shift of the N400 in the word learning phase (i.e., without
motor responses) only developed in musicians, which we
interpret as an electrophysiological signature of “fast map-
ping” (Carey, 1978). To our knowledge, this is the first re-
port showing that music training influences semantic
processing. As a future step, we plan to use a longitudinal
approach with nonmusician controls trained with music so
as to test for the causal influence of music training on
word learning. Finally, these results also open new per-
spectives to further investigate the influence of music
training on long-term memory for applications of music
training in the domain of native and second language
learning (Moreno, Lee, Janus, & Bialystok, 2015; Chobert
& Besson, 2013) and for using this type of experimental
design in clinical research to specify the stages of word
learning that are most deficient.
Acknowledgments
We would like to thank all the participants, Chotiga Pattamadilok
for registering the auditory stimuli, and Benjamin Furnari for
his help with data acquisition and analyses. The work of E. D.
M. Ba., M. D. I., and M. B., carried out within the Labex BLRI
(ANR-11-LABX-0036), has benefited from support from the
French government, managed by the French National Agency for
Research (ANR), under the program “Investissements d’Avenir”
(ANR-11-IDEX-0001-02). E. D. was supported by a doctoral
fellowship from the BLRI and M. Ba. by a doctoral fellowship
from the French Ministry of Research and Education.
Reprint requests should be sent to Eva Dittinger, Laboratoire de
Neuroscience Cognitives, Université Aix Marseille, Centre Saint
Charles, 3 Place Victor Hugo, Marseille, France, 13331, or via
e-mail: eva.dittinger@blri.fr.
Notes
1. Following phonetic transcription in Thai, 1 refers to low-
tone, 0 to midtone, ph to aspirated voicing, and the colon to
long vowel duration.
2. Voice Onset Time (VOT) is defined as the interval between
the noise burst produced at consonant release and the wave-
form periodicity associated with vocal cord vibrations (Lisker
& Abramson, 1967) .
3. Pictures were chosen from the Snodgrass and Vanderwart
(1980) pictures’ set but were found on the Internet to ensure
sufficient resolution and quality.
REFERENCES
Asaridou, S. S., & McQueen, J. M. (2013). Speech and music
shape the listening brain: Evidence for shared domain-general
mechanisms. Frontiers in Psychology, 4, 321.
Batterink, L., & Neville, H. (2011). Implicit and explicit
mechanisms of word learning in a narrative context: An
event-related potential study. Journal of Cognitive
Neuroscience, 23, 3181–3196.
Besson, M., Chobert, J., & Marie, C. (2011). Transfer of training
between music and speech: Common processing, attention,
and memory. Frontiers in Psychology, 2, 94.
1600
Journal of Cognitive Neuroscience
Volume 28, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Bidelman, G. M., Weiss, M. W., Moreno, S., & Alain, C. (2014).
Gandour, J., Wong, D., Lowe, M., Dzemidzic, M.,
Coordinated plasticity in brainstem and auditory cortex
contributes to enhanced categorical speech perception
in musicians. European Journal of Neuroscience,
40, 2662–2673.
Boddy, J., & Weinberg, H. (1981). Brain potentials, perceptual
mechanism and semantic categorization. Biological
Psychology, 12, 43–61.
Boersma, P., & Weenink, D. (2011). Praat: Doing phonetics
by computer [computer program]. Retrieved from
www.praat.org.
Borovsky, A., Elman, J., & Kutas, M. (2012). Once is enough:
N400 indexes semantic integration of novel word meanings
from a single exposure in context. Language Learning
and Development, 8, 278–302.
Borovsky, A., Kutas, M., & Elman, J. (2010). Learning to use
words: Event-related potentials index single-shot contextual
word learning. Cognition, 116, 289–296.
Carey, S. (1978). The child as word learner. In M. Halle,
J. Bresnan, & G. A. Miller (Eds.), Linguistic theory
and psychological reality (pp. 264–293). Cambridge,
MA: MIT Press.
Chobert, J., & Besson, M. (2013). Musical expertise and second
language learning. Brain Sciences, 3, 923–940.
Chobert, J., François, C., Velay, J. L., & Besson, M. (2012).
Twelve months of active musical training in 8- to 10-year-old
children enhances the preattentive processing of syllabic
duration and voice onset time. Cerebral Cortex, 24, 956–967.
Chobert, J., Marie, C., François, C., Schön, D., & Besson, M.
(2011). Enhanced passive and active processing of syllables
in musician children. Journal of Cognitive Neuroscience,
23, 3874–3887.
Connolly, J. F., & Phillips, N. A. (1994). Event-related potential
components reflect phonological and semantic processing
of the terminal words of spoken sentences. Journal of
Cognitive Neuroscience, 6, 256–266.
Corrigall, K. A., Schellenberg, E. G., & Misura, N. M. (2013).
Music training, cognition, and personality. Frontiers in
Psychology, 4, 222.
Diamond, A. (2013). Executive functions. Annual Review
Psychology, 64, 135–168.
Dobel, C., Lagemann, L., & Zwitserlood, P. (2009). Non-native
phonemes in adult word learning: Evidence from the N400m.
Philosophical Transactions of the Royal Society, Series B,
Biological Sciences, 364, 3697–3709.
Dumay, N., & Gaskell, M. G. (2007). Sleep-associated changes in
the mental representation of spoken words. Psychological
Science, 18, 35–39.
Elmer, S., Hänggi, J., Meyer, M., & Jäncke, L. (2013). Increased
cortical surface area of the left planum temporale in
musicians facilitates the categorization of phonetic and
temporal speech sounds. Cortex, 49, 2812–2821.
Elmer, S., Meyer, M., & Jäncke, L. (2012). Neurofunctional and
behavioral correlates of phonetic and temporal
categorization in musically trained and untrained subjects.
Cerebral Cortex, 22, 650–658.
Fitzroy, A. B., & Sanders, L. D. (2013). Musical expertise
modulates early processing of syntactic violations in
language. Frontiers in Psychology, 3, 603.
François, C., Chobert, J., Besson, M., & Schön, D. (2013). Music
training for the development of speech segmentation.
Cerebral Cortex, 23, 2038–2043.
Friederici, A. D., & Singer, W. (2015). Grounding language
processing on basic neurophysiological principles. Trends
in Cognitive Sciences, 19, 329–338.
Friedrich, M., & Friederici, A. D. (2008). Neurophysiological
correlates of online word learning in 14-month-old infants.
NeuroReport, 19, 1757–1761.
Satthamnuwong, N., Tong, Y., et al. (2002). A crosslinguistic
fMRI study of spectral and temporal cues underlying
phonological processing. Journal of Cognitive Neuroscience,
14, 1076–1087.
Gaser, C., & Schlaug, G. (2003). Brain structures differ
between musicians and nonmusicians. Journal of
Neuroscience, 23, 9240–9245.
George, E. M., & Coch, D. (2011). Music training and
working memory: An ERP study. Neuropsychologia,
49, 1083–1094.
Gigerenzer, G. (1991). How to make cognitive illusions
disappear: Beyond “heuristics and biases”. European
Review of Social Psychology, 2, 83–115.
Hagoort, P. (2014). Nodes and networks in the neural
architecture for language: Broca’s region and beyond.
Current Opinion in Neurobiology, 28, 136–141.
Ho, Y. C., Cheung, M. C., & Chan, A. S. (2003). Music training
improves verbal but not visual memory: Cross-sectional
and longitudinal explorations in children. Neuropsychology,
17, 439–450.
Hyde, K. L., Lerch, J., Norton, A., Forgeard, M., Winner, E.,
Evans, A. C., et al. (2009). Musical training shapes structural
brain development. Journal of Neuroscience, 29,
3019–3025.
Jäncke, L. (2009). Music drives brain plasticity. F1000 Biology
Reports, 1, 1–6.
Jasper, H. H. (1958). The ten–twenty system of the International
Federation. Clinical Neurophysiology, 10, 371–375.
Jentschke, S., & Koelsch, S. (2009). Musical training modulates
the development of syntax processing in children.
Neuroimage, 47, 735–744.
Koelsch, S., Kasper, E., Sammler, D., Schulze, K., Gunter, T., &
Friederici, A. D. (2004). Music, language and meaning: Brain
signatures of semantic processing. Nature Neuroscience,
7, 302–307.
Korkman, M., Kirk, U., & Kemp, S. (1998). NEPSY: A developmental
neuropsychological assessment. San Antonio, TX: The
Psychological Corporation.
Kraus, N., & Chandrasekaran, B. (2010). Music training for
the development of auditory skills. Nature Reviews
Neuroscience, 11, 599–605.
Kühnis, J., Elmer, S., & Jäncke, L. (2014). Auditory evoked
responses in musicians during passive vowel listening are
modulated by functional connectivity between bilateral
auditory-related brain regions. Journal of Cognitive
Neuroscience, 26, 2750–2761.
Kutas, M., & Federmeier, K. D. (2011). Thirty years and
counting: Finding meaning in the N400 component of the
event related brain potential (ERP). Annual Review of
Psychology, 62, 621–647.
Kutas, M., & Hillyard, S. A. (1980). Reading senseless sentences:
Brain potentials reflect semantic incongruity. Science,
207, 203–205.
Kutas, M., Van Petten, C., & Besson, M. (1988). Event-related
potential asymmetries during the reading of sentences.
Electroencephalogry and Clinical Neurophysiology,
69, 218–233.
Lisker, L., & Abramson, A. S. (1967). Some effects of context
on voice onset time in English stops. Language ans Speech,
10, 1–28.
Maess, B., Koelsch, S., Gunter, T. C., & Friederici, A. D. (2001).
“Musical syntax” is processed in the area of Broca: An
MEG-study. Nature Neuroscience, 4, 540–545.
Marie, C., Delogu, F., Lampis, G., Olivetti Belardinelli, M., &
Besson, M. (2011). Influence of musical expertise on
segmental and tonal processing in Mandarin Chinese.
Journal of Cognitive Neuroscience, 23, 2701–2715.
Dittinger et al.
1601
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Marques, C., Moreno, S., Castro, S. L., & Besson, M. (2007).
Musicians detect pitch violation in a foreign language better
than nonmusicians: Behavioural and electrophysiological
evidence. Journal of Cognitive Neuroscience, 19, 1453–1463.
McLaughlin, J., Osterhout, L., & Kim, A. (2004). Neural correlates
of second-language word learning: Minimal instruction
produces rapid change. Nature Neuroscience, 7, 703–704.
Mestres-Missé, A., Rodriguez-Fornells, A., & Münte, T. F. (2007).
Watching the brain during meaning acquisition. Cerebral
Cortex, 17, 1858–1866.
Meyer, D. E., & Schvaneveldt, R. W. (1971). Facilitation in
recognizing pairs of words: Evidence of a dependence
between retrieval operations. Journal of Experimental
Psychology, 20, 227–234.
Moreno, S., Bialystok, E., Barac, R., Schellenberg, E. G., Cepeda,
N. J., & Chau, T. (2011). Short-term music training enhances
verbal intelligence and executive function. Psychological
Science, 22, 1425–1433.
Moreno, S., Lee, Y., Janus, M., & Bialystok, E. (2015).
Short-term second language and music training induces
lasting functional brain changes in early childhood.
Child Development, 86, 394–406.
Moreno, S., Marques, C., Santos, A., Santos, M., Castro, S. L.,
& Besson, M. (2009). Musical training influences linguistic
abilities in 8-year-old children: More evidence for brain
plasticity. Cerebral Cortex, 19, 712–723.
Münte, T. F., Altenmüller, E., & Jäncke, L. (2002). The
musician’s brain as a model of neuroplasticity. Nature
Reviews Neuroscience, 3, 473–478.
Musacchia, G., Sams, M., Skoe, E., & Kraus, N. (2007). Musicians
have enhanced subcortical auditory and audiovisual
processing of speech and music. Proceedings of the National
Academy of Sciences, U.S.A., 104, 15894–15898.
Pallesen, K. J., Brattico, E., Bailey, C. J., Korvenoja, A., Koivisto, J.,
Gjedde, A., et al. (2010). Cognitive control in auditory working
memory is enhanced in musicians. PLoS One, 5, 11120.
Pantev, C., Oostenveld, R., Engelien, A., Ross, B., Roberts, L. E.,
& Hoke, M. (1998). Increased auditory cortical representation
in musicians. Nature, 23, 811–814.
Patel, A. D. (2008). Music, language, and the brain. New York:
Oxford University Press.
Peretz, I., Champod, A. S., & Hyde, K. (2003). Varieties of musical
disorders. The Montreal Battery of Evaluation of Amusia.
Annals of the New York Academy of Sciences, 999, 58–75.
Peretz, I., Vuvan, D., Lagrois, M.-É., & Armony, J. L. (2015).
Neural overlap in processing music and speech. Philosophical
Transactions of the Royal Society, Series B, Biological
Sciences, 370, 20140090.
Perfetti, C. A., Wlotko, E. W., & Hart, L. A. (2005). Word learning
and individual differences in word learning reflected in
event-related potentials. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 31, 1281–1292.
Raven, J. C., Corporation, P., & Lewis, H. K. (1962). Coloured
progressive matrices: Sets A, AB, B. London, UK: Oxford
Psychologist Press.
Rodriguez-Fornells, A., Cunillera, T., Mestres-Missé, A., &
De Diego-Balaguer, R. (2009). Neuropsychological mechanisms
involved in language learning in adults. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 364, 3711–3735.
Rogalsky, C., Rong, F., Saberi, K., & Hickok, G. (2011).
Functional anatomy of language and music perception:
Temporal and structural factors investigated using functional
magnetic resonance imaging. Journal of Neuroscience, 31,
3843–3852.
Schellenberg, E. G. (2004). Music lessons enhance IQ.
Psychological Science, 15, 511–514.
Schneider, P., Scherg, M., Dosch, H. G., Specht, H. J., Gutschalk,
A., & Rupp, A. (2002). Morphology of Heschl’s gyrus reflects
enhanced activation in the auditory cortex of musicians.
Nature Neuroscience, 5, 688–694.
Schön, D., Magne, C., & Besson, M. (2004). The music of speech:
Music training facilitates pitch processing in both music and
language. Psychophysiology, 41, 341–349.
Seppänen, M., Hämäläinen, J., Pesonen, A. K., & Tervaniemi, M.
(2012). Music training enhances rapid neural plasticity of N1
and P2 source activation for unattended sounds. Frontiers in
Human Neuroscience, 6, 43.
Snodgrass, J. G., & Vanderwart, M. (1980). A standardized
set of 260 pictures: Norms for name agreement, image
agreement, familiarity, and visual complexity. Journal of
Experimental Psychology. Human Learning and Memory,
6, 174–215.
Steinbeis, N., & Koelsch, S. (2008). Comparing the processing
of music and language meaning using EEG and fMRI provides
evidence for similar and distinct neural representations.
PLoS One, 3, 2226.
Strait, D. L., Parbery-Clark, A., O’Connell, S., & Kraus, N. (2013).
Biological impact of preschool music classes on processing
speech in noise. Developmental Cognitive Neuroscience, 6,
51–60.
Strait, D. L., Slater, J., O’Connell, S., & Kraus, N. (2015). Music
training relates to the development of neural mechanisms of
selective auditory attention. Developmental Cognitive
Neuroscience, 12, 94–104.
Torkildsen, J. V. K., Svangstu, J. M., Friis Hansen, H., Smith, L.,
Simonsen, H. G., Moen, I., et al. (2008). Productive
vocabulary size predicts event-related potential correlates of
fast mapping in 20-month-olds. Journal of Cognitive
Neuroscience, 20, 1266–1282.
van den Brink, D., Brown, C. M., & Hagoort, P. (2001).
Electrophysiological evidence for early contextual influences
during spoken-word recognition: N200 versus N400 effects.
Journal of Cognitive Neuroscience, 13, 967–985.
Wagner, A. W., Schacter, D. L., Rotte, M., Koutstaal, W., Maril, A.,
Dale, A. M., et al. (1998). Building memories: Remembering
and forgetting of verbal experiences as predicted by brain
activity. Science, 281, 1188–1190.
Wang, X., Ossher, L., & Reuter-Lorenz, P. A. (2015). Examining
the relationship between skilled music training and attention.
Consciousness and Cognition, 36, 169–179.
Wechsler, D. (2003). Wechsler Intelligence Scale for
children (4th ed.). San Antonio, TX: The Psychological
Corporation.
Woldorff, M., & Hillyard, S. A. (1991). Modulation of early
auditory processing during selective listening to rapidly
presented tones. Electroencephalography and Clinical
Neurophysiology, 79, 170–191.
Wong, P. C. M., & Perrachione, T. K. (2007). Learning pitch
patterns in lexical identification by native English-speaking
adults. Applied Psycholinguistics, 28, 565–585.
Zuk, J., Benjamin, C., Kenyon, A., & Gaab, N. (2014). Behavioral
and neural correlates of executive functioning in musicians
and non-musicians. PLoS One, 9, 99868.
1602
Journal of Cognitive Neuroscience
Volume 28, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
0
8
/
1
1
5
0
8
/
4
1
1
5
9
8
5
4
1
/
3
1
5
2
7
8
o
5
c
4
n
9
_
6
a
/
_
j
0
o
0
c
9
n
9
7
_
a
p
_
d
0
0
b
9
y
9
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1