PERSPECTIVE
Switching between internal and external modes:
A multiscale learning principle
1
Christopher J. Honey
, Ehren L. Newman
2
, and Anna C. Schapiro
3
1Department of Psychological and Brain Sciences, Johns Hopkins Universität, Baltimore, MD, USA
2Department of Psychological and Brain Sciences, Indiana University, Bloomington, IN, USA
Department of Psychiatry, Beth Israel Deaconess Medical Center / Harvard Medical School, Boston, MA, USA
3
Schlüsselwörter: Acetylcholine, Learning, Timescale, Switching, Contrastive learning, Hippocampus,
Sleep, Default mode
Keine offenen Zugänge
Tagebuch
ABSTRAKT
Brains construct internal models that support perception, prediction, and action in the
external world. Individual circuits within a brain also learn internal models of the local world
of input they receive, in order to facilitate efficient and robust representation. How are these
internal models learned? We propose that learning is facilitated by continual switching
between internally biased and externally biased modes of processing. We review
computational evidence that this mode-switching can produce an error signal to drive
learning. We then consider empirical evidence for the instantiation of mode-switching in
diverse neural systems, ranging from subsecond fluctuations in the hippocampus to
wake-sleep alternations across the whole brain. We hypothesize that these internal/external
switching processes, which occur at multiple scales, can drive learning at each scale. Das
framework predicts that (A) slower mode-switching should be associated with learning of
more temporally extended input features and (B) disruption of switching should impair the
integration of new information with prior information.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
Zitat: Honey, C. J., Newman, E. L., &
Schapiro, A. C. (2017). Switching
between internal and external modes:
A multiscale learning principle.
Netzwerkneurowissenschaften, 1(4), 339–356.
https://doi.org/10.1162/netn_a_00024
DOI:
https://doi.org/10.1162/netn_a_00024
Erhalten: 10 Januar 2017
Akzeptiert: 18 August 2017
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Christopher J. Honey
chris.honey@jhu.edu
Handling-Editor:
Danielle Bassett
Urheberrechte ©: © 2017
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
EINFÜHRUNG
Why do so many of the neural processes supporting memory and perception fluctuate over
Zeit? Perceptual thresholds increase and decrease over milliseconds, seconds, and min-
utes (Verehrung & Verehrung, 2011; VanRullen, 2016); the associative strength of hippocampal encod-
ing fluctuates over milliseconds (Hasselmo, 2006; Hasselmo, Bodelón, & Wyble, 2002) Und
seconds (Douchamps, Jeewajee, Blundell, Bürger, & Lever, 2013; Duncan, Sadanand, &
Davachi 2012); and overall arousal fluctuates within the waking state (McGinley et al., 2015B)
as well as on the timescale of wake and sleep.
It is possible that these fluctuations are epiphenomena, reflecting arbitrary biological con-
straints or noise. But there is a cost to breaking from the world: a reduced sensitivity to what
is occurring in the environment. Why would the brain be willing to pay this price? We pro-
pose that many of these fluctuating processes, across wide-ranging scales and systems of the
mammalian brain, can be understood as mode-switching processes that facilitate learning. In
besondere, we hypothesize that mode-switching enables individual neural systems to construct
and refine internal models of the afferent signals that constitute their world. These “models”
of afferent signals include any kind of representation that allows a system to improve its pre-
dictions about its environment.
Learning by switching internal and external modes
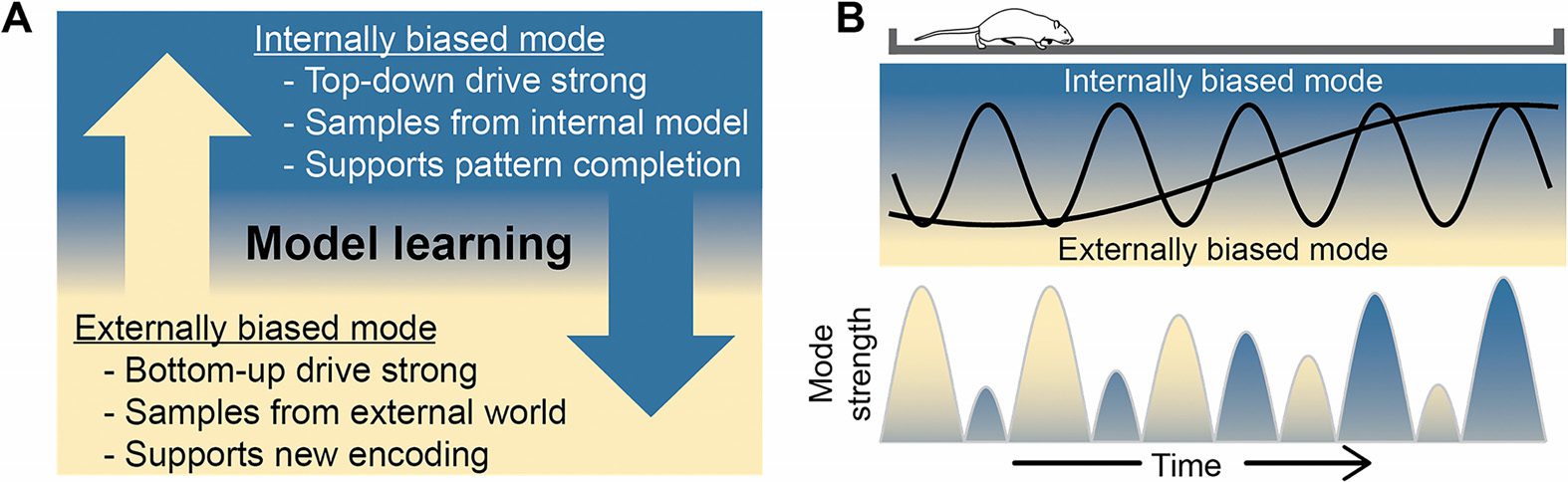
Figur 1. Switching between internally and externally biased modes. (A) Schematic of internally
and externally biased modes of processing. (B) Illustration of switching between external and in-
ternal drivers of activity at multiple timescales simultaneously. In the time it takes for an animal to
navigate a linear track, areas like the hippocampus switch between internally and externally biased
modes at fast and slow timescales simultaneously, as caricatured by the two black sine waves. Der
net result, as shown by the bottom mode strength chart, is that each mode is sampled often but the
dominance of one mode over the other changes gradually.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
What are the modes between which neural systems vary? Building on ideas developed by
Hasselmo (1995), we refer to them as “internally biased” and “externally biased” modes. Der
basic properties of these modes are summarized in Figure 1. The “internal” mode is biased
toward recurrent or top-down drivers. In this mode, prior learning shapes the neural dynam-
ics, effectively driving the current activation state toward the learned patterns that comprise
an internal model. Funktional, this supports perceptual processing by enabling pattern com-
pletion and prediction. The “external” mode, in contrast, enhances the relative strength of
feedforward drive. In this mode, incoming signals from the world or afferent regions shape the
neural dynamics, effectively driving the current activation state to reflect the structure of the
Eingang.
Which anatomical pathways correspond to “internal” and “external” processing? The an-
swer depends on how information flows from the world to arrive at each neural system. In
the case of V1, “external” input is provided by the lateral geniculate nucleus, while “inter-
nal” input involves local pyramidal neurons and top-down visual projections. In the case of
the hippocampus, “external” input is provided via superficial layers of the entorhinal cortex,
while “internal” input depends on synapses within and between CA3 and CA1. When consid-
ering the cerebrum as a whole, most “external” input is associated with thalamic projections to
sensory cortices, while “internal” input occurs most strongly from higher-order cortices and in
medial temporal and limbic systems. As a rule of thumb, more “internal” circuits are located
a larger number of synapses away from the sensory periphery. Quantitative metrics can also
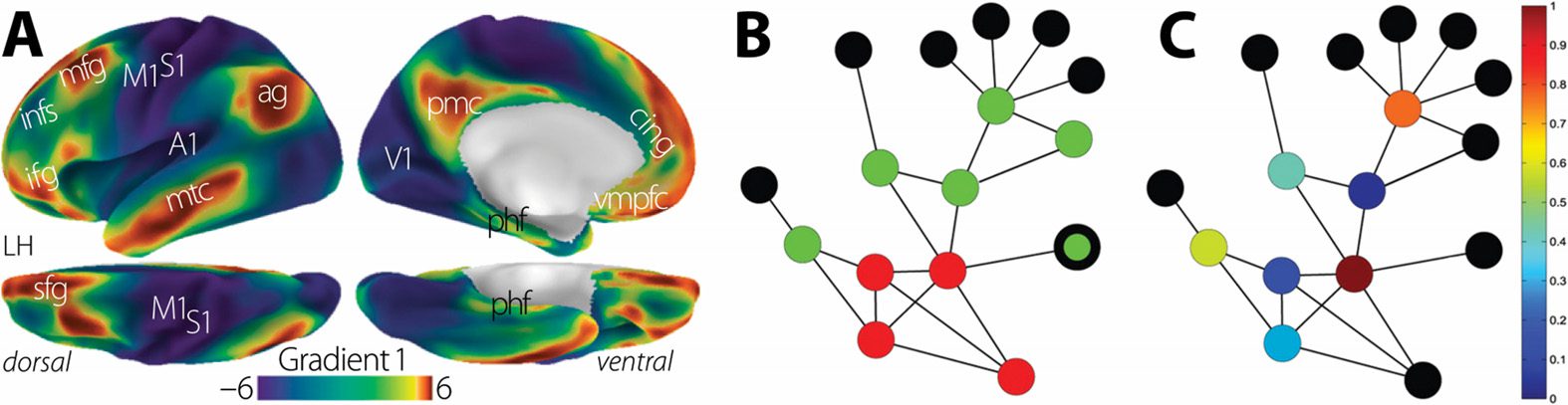
be defined: Figur 2 illustrates a large-scale gradient of external-internal processing derived
by Margulies et al. (2016) and network-theoretic tools that can quantify internal-external gra-
dients (Della Rossa, Dercole, & Piccardi, 2013). Most fundamentally, external pathways are
those that convey information from current or recent states of the world; internal pathways
are those that convey information from a model of the world shaped by days and years of
Erfahrung.
How might mode-switching aid learning? Rechnerisch, neural systems are believed
to learn by changing their representations according to the difference between expected and
observed input. Switching between modes that are biased toward new or old information
Netzwerkneurowissenschaften
340
Learning by switching internal and external modes
Figur 2. Defining internally and externally biased circuits. (A) Macroscopic gradients from inter-
nal to external processing can be defined based on covariation in functional connectivity patterns.
Network-theoretic tools such as (B) k-shell decomposition and (C) core-periphery profiling can also
be used to define a node-specific measure of distance from the network core. Panel A is adapted
from Margulies et al. (2016). Panels B and C are adapted from Della Rossa et al. (2013).
allows neural circuits to identify the contrast between these two sources of information, Und
thus to iteratively reduce the mismatch between them.
The sections below elaborate the evidence and reasoning behind this proposal. Erste, Wir
outline the long-standing computational motivation for mode-switching. Then we review
the empirical support for mode-switching in both the allocortex and the neocortex on faster
timescales (milliseconds and seconds) and slower timescales (minutes to hours). Empirical sup-
port for the existence of mode-switching is substantial, while evidence for its role in learning is
still in early stages. The theoretical utility of mode-switching leads us to the strong prediction
that where there is evidence for switching between internally and externally biased modes,
this may be used by the brain to benefit learning. Endlich, we consider why the fluctuations
occur on such a range of timescales, as well as the broader implications and predictions of
this framework.
COMPUTATIONAL ROLE OF SWITCHING MODES
To think and act adaptively, animals build internal models that reflect relevant properties of
die Umgebung. Models of the environment enable diverse behaviors (including navigation,
planning, tool use) and mental functions (search in noisy environments, Bilder, prediction).
Many algorithms are known to shape accurate internal models in neural networks. The sim-
plest algorithms are Hebbian: connections are strengthened between neurons that represent
stimuli that occur together in space and time (Hebb, 1949; Hopfield, 1982). These algorithms
become more powerful, Jedoch, when the learning process is further driven by a mismatch, oder
Fehler, signal (Rosenblatt, 1958; Widrow & Hoff, 1960). The mismatch signal can be generated
by switching between two modes. In the first mode, the network uses internal representations
to generate a “guess” about the present environment (auto-encoder) or near-future environ-
ment (predictor). In the second mode, network activity is primarily driven by external input.
To generate the mismatch signal, algorithms compare the patterns of activity resulting from
these two modes.
The mismatch can be computed at an output layer and then propagated back to earlier lay-
ers (z.B., backpropagation; Lillicrap, Cownden, Tweed, & Akerman, 2016; Rumelhart, Hinton
& Williams, 1986), or individual units within each layer of a network can compute mismatch
based on more local
Information (z.B., Bengio, Lee, Bornschein, Mesnard, & Lin, 2015;
Hinton & McClelland, 1988; O’Reilly, 1996; Smolensky, 1986). Synaptic weights are then
341
Backpropagation:
Learning by propagating an
output-layer error backward through
layers of a network, proportional to
how much each connection
contributed to the error.
Netzwerkneurowissenschaften
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
Trisynaptic pathway:
A three-synapse pathway for
information flow in the
hippocampus, from the entorhinal
cortex to the dentate gyrus, then on
to area CA3, and finally to area CA1.
iteratively adjusted away from the internal model and toward the external input, such that the
next time a similar input is encountered, the network is incrementally better able to anticipate
and process it.
We propose that different brain systems, operating on multiple different timescales, switch
between internal and external modes as a way of implementing a comparison computation of
this kind. Using the mismatch between external measurement and the expectation of the inter-
nal model, each neural circuit iteratively improves an internal model of its own “environment,”
which is the set of inputs it receives.
EMPIRICAL EVIDENCE FOR INTERNAL/EXTERNAL MODE SWITCHES
Below we consider a few examples of mode-switching systems (Tisch 1). This list is represen-
tative rather than exhaustive, with the aim of illustrating (A) the range of systems that exhibit
functionally relevant switching; (B) the range of timescales over which switching occurs; Und
(C) the possible neurophysiological commonalities across the examples.
Hippocampus
The hippocampus provides multiple examples of internal/external mode-switching with demon-
strable functional implications.
In hippocampal area CA1, “bottom-up” input arrives via a
monosynaptic pathway directly from the entorhinal cortex, while “top-down” input arrives
from hippocampal area CA3 after traveling the trisynaptic pathway from entorhinal cortex
through dentate gyrus to CA1 (van Strien, Cappaert, & Witter, 2009). The bottom-up path-
way is more directly influenced by the external world than the trisynaptic pathway, mit
fewer synapses separation from the environment. Electrophysiological analysis reveals that
when animals navigate, CA1 receives input from these two pathways alternately across the
phases of the theta rhythm in the local field potential (Brankack, Stewart, & Fuchs, 1993; Buzsáki,
Czopf, Kondákor, & Kellényi, 1986). The alternating phases of CA1 input are functionally dif-
ferent: CA1 firing represents information immediately in front of the animal during entorhinal
bottom-up drive as opposed to retrospective information during CA3 top-down drive
(Fernández-Ruiz et al., 2017; Itskov, Pastalkova, Mizuseki, Buzsáki, & Harris, 2008; O’Keefe
& Recce, 1993). At a physiological level, these alternating phases also differ with regard to
neural plasticity: long-term potentiation (LTP) versus long-term depression (LTD) are induced
by activity during the bottom-up and top-down phases, jeweils (Huerta & Lisman, 1995;
Hyman, Wyble, Goyal, Rossi, & Hasselmo, 2003). Such differences have led these phases to
be ascribed distinct functional roles with regard to memory formation (Hasselmo et al., 2002;
Norman, Newman, Detre, & Polyn, 2006). The mechanism supporting the rapid switching
is a combination of systems-level and cellular-level interactions that maintain the antiphase
relationship between these inputs (Buzsáki, 2002).
The internal and external drives to CA1 also switch on a timescale that is an order of magni-
tude slower than the intra-theta cycle described above (z.B., Figure 1B, with each phase lasting
∼500 ms instead of ∼50 ms). The key observation is that the strength of the bottom-up and top-
down inputs varies, not only within each theta cycle, but also across theta cycles (Colgin et al.,
2009; Fernández-Ruiz et al., 2017; Schomburg et al., 2014). Once one pathway becomes
activated, it remains activated for two to nine consecutive theta cycles (Colgin et al., 2009),
and the strength of the two pathways are inversely related such that it is rare to observe both
strong on the same theta cycle (Colgin et al., 2009).
Netzwerkneurowissenschaften
342
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
Tisch 1. Summary of internal versus external mode examples.
Timescale
Spatial scale
Internal mode
External mode
Hippocampal encoding
vs. retrieval
10s ms
Hippocampal encoding
vs. retrieval
100s ms
Single circuit:
hippocampal trisynaptic
loop
Trough of
CA1pyramidal theta:
CA3 input to CA1
stronger than
entorhinal input
Peak of
CA1pyramidal theta:
entorhinal input to
CA1 stronger than
CA3 input
Multiple circuits:
hippocampus and septal
circuits
Retrieval: lower
cholinergic tone;
CA3 drives CA1
Aktivität
Encoding: higher
cholinergic tone;
entorhinal cortex
drives CA1 activity
Neocortical
feedforward vs.
Rückmeldung
100s ms to 10s s
Changes often coherent
over ∼5–50 mm of
neocortex
Active vs. resting
wake periods
seconds–minutes
Most of brain
Non-REM vs. REM
10s minutes
Entire brain
Wake vs. schlafen
Hours
Entire brain
Field potential
filtered in 4–35 Hz
range is high and
high variance;
lower; cholinergic
tone; inhibition of
core thalamic input
and feedforward
corticocortical drive
Field potential
filtered in 4-35 Hz
range is low and low
variance; higher
cholinergic tone;
elevated
asynchronous firing,
detectable as
increase in
broadband power
Lower cholinergic
tone; higher overall
activity in default
mode network
Higher cholinergic
tone; lower overall
activity in default
mode network
REM: exploration of
cortical networks
containing
long-term memories
Non-REM:
dominance of
hippocampus,
containing more
recent memories
Sleep: minimal
influence of
Umfeld;
relatively lower
cholinergic tone in
cortex on average
Wake: (potential for)
strong influence of
Umfeld;
relatively higher
cholinergic tone in
cortex on average
Evidence that these slower fluctuations serve functional roles comes from behavioral stud-
ies in rodents showing that the bottom-up pathway remains strong during encoding of a novel
object or an unfamiliar maze and that the strength of the top-down pathway grows as animals
become able to use mnemonic recollection to drive behavior (z.B., Bieri, Bobbitt, & Colgin,
Netzwerkneurowissenschaften
343
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
Hippocampal theta oscillations:
Rhythmic fluctuations in the local
field potential of the hippocampus,
repeating cyclically at 6–10 Hz.
2014; Fernández-Ruiz et al., 2017). Manipulations that interfere with the bottom-up path-
Weg, such as administration of a cholinergic antagonist, shift the balance toward the top-
down pathway, and interfere with encoding (Douchamps et al., 2013; Newman, Gillet, Climer
in conjunction with evidence from in vitro studies
& Hasselmo, 2013). Such evidence,
(Hasselmo & Schnell, 1994), suggests that cholinergic modulation plays a central role in me-
diating the switching between bottom-up and top-down processing in the hippocampus.
Rechnerisch, both fast and slow forms of hippocampal switching reflect an alternation
between epochs of “encoding,” where bottom-up activity drives CA1, and epochs of “retrieval,”
where CA3 drives CA1 (Hasselmo, 1995; Hasselmo & Schnell, 1994; Meeter, Murre, &
Talamini, 2004). Switching between these two inputs may allow CA1 to compare them, sup-
porting alignment of environmental events and internal models (Bittner et al., 2015;
Hasselmo et al., 2002). Consistent with the idea that the regular switching of modes is im-
portant for learning, blocking theta oscillations via septal inactivation blocks hippocampal
dependent learning in rats (Mizumori, Perez, Alvarado, Barnes, & McNaughton, 1990).
Neocortex
The neocortex provides multiple examples of mode-switching, and these are most often stud-
ied in the context of perception rather than learning. The most well-known switch between
externally and internally biased modes is the “alpha” oscillation in the occipital neocortex.
Electrical potentials recorded from the scalp above the visual cortex oscillate on a timescale
of ∼100 ms (Adrian & Matthews, 1934). Decreases in the amplitude of these oscillations
are associated with greater engagement with the external visual world. Sensitivity to ex-
ternal visual stimuli appears to be particularly enhanced at the troughs of individual cycles
(Busch, Dubois, & VanRullen, 2009; Jensen & Mazaheri, 2010).
More generally across the mammalian neocortex, band-limited oscillations in the elec-
trical potential are observed with timescales typically ranging from ∼30 to 400 MS (Buzáki &
Draguhn, 2004; Groppe et al., 2013; Jasper & Penfield, 1949; Keitel & Brutto, 2016). Both the
amplitude and phase of these oscillations have been linked to switches between external and
internal processing modes. The amplitude of oscillations, especially in the range of 6–18 Hz,
decreases when local feedforward processing (external mode) is engaged in a particular
Bereich (Müller, Honey, Hermes, Rao, & Ojemann, 2014; Pfurtscheller & Da Silva, 1999), while
increases in amplitude (and peaks of individual oscillatory cycles) appear to be associated with
a more prior-driven, top-down (internal) mode of processing (Bastos et al., 2015; Halgren et al.,
2015; Sherman, Kanai, Seth, & VanRullen, 2016; Van Kerkoerle et al., 2014). Such a phasic
alternation between stimulus-driven and prior-driven activity has been posited as the basis of
perceptual learning in a recent computational model of early visual cortex (O’Reilly, Wyatte,
& Rohrlich, 2014).
The variation in feedforward strength need not occur with a fixed rhythmic timescale.
Much of the variation in field potentials, and in the amplitude of rhythmic processes, oc-
curs more slowly than 2 Hz, and may not have a tightly constrained timescale (Honey et al.,
2012; Leopold, Murayama, & Logothetis, 2003). Even in early visual cortices, slower vari-
ations in field potential (Schlosser, Karmos, Mehta, Ulbert, & Schroeder, 2008) and firing rate
(Engel et al., 2016) reflect phasic changes in attention and track perceptual sensitivity.
The strength of neocortical feedback and feedforward drive are continuously switching, Und
this will impact what is perceived and learned. Jedoch, experiments that manipulate the
strength and frequency of mode switches (keeping other factors constant) would be needed
Netzwerkneurowissenschaften
344
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
Acetylcholine:
An organic chemical that modulates
and drives activity in diverse regions
of central and peripheral nervous
Systeme.
to establish that the alternation of modes is itself driving learning.
Zusätzlich, we caution
against a one-to-one mapping between field potential amplitude and perceptual sensitivity:
Field potentials are only an aggregate measure of a system of interwoven lamina, and in some
tasks the most accurate inferences about the external world may occur at intermediate levels
of feedforward excitability (Linkenkaer-Hansen et al., 2004; McGinley, David, & McCormick,
2015A).
Common neuromodulatory and oscillatory patterns in hippocampus and neocortex.
Cholinergic tone is associated with an “external” mode of processing in the hippocampus
and in the cerebral cortex (Newman et al., 2013; Newman, Gupta, Climer, Monaghan, &
Hasselmo, 2012A). Hippocampally, cholinergic agonists in area CA1 induce a form of pre-
synaptic inhibition on the “top-down” inputs from area CA3, biasing processing toward the
bottom-up inputs from entorhinal cortex (Hasselmo & Schnell, 1994). In the piriform cortex,
acetylcholine acts to increase the relative strength of bottom-up input from the olfactory bulb
versus top-down input from cortical sources (Hasselmo & Bower, 1992; Linster & Cleland,
2016). In the neocortex, cholinergic modulation facilitates bottom-up processing by potentiat-
ing thalamocortical synapses (Disney, Aoki, & Hawken, 2007; Gil, Connors, & Amitai, 1997;
Hsieh, Cruikshank & Metherate, 2000). When acetylcholine drives sensory cortex to an ex-
ternal mode, this is reflected in decreased slow (∼2–10 Hz) field oscillations and increased
fidelity of sensory coding (Goard & Und, 2009; Pinto et al., 2013; see also Warburton, Wesnes,
Edwards, & Larrad, 1985).
Fluctuations in cholinergic and noradrenergic tone are associated with fluctuations in
arousal, as reflected in spontaneous variation in perceptual acuity and bias, and in physiolog-
ical parameters such as pupil diameter (de Gee et al., 2017; McGinley et al., 2015B). A study
of auditory sensory discrimination (McGinley et al., 2015A) demonstrated that false alarm
rates were highest in an auditory discrimination task when pupil diameter was largest and slow
auditory cortical oscillations were smallest. Daher, the “external mode” for neocortical circuits
appears to be associated with increased cholinergic tone (and perhaps noradrenergic tone) als
well as with decreases in local oscillatory activity below 30 Hz. Although the highest levels
of arousal are not always optimal for perceptual judgment and decision making (de Gee et al.,
2017; McGinley et al., 2015A), it seems clear that fluctuations in cholinergic and noradren-
ergic neuromodulation can bias the relative influence of internal and external sources of
Information.
Whole Brain
Awake rest states and the default mode network. Switching between internal and external
modes also occurs on longer timescales. We spend as much as half of our awake time in
disengaged states characterized by spans of seconds or minutes of reduced attention to our
immediate environment (Killingsworth & Gilbert, 2010; Monto, Verehrung, Voipio, & Verehrung, 2008;
Sadaghiani, Hesselmann, Friston, & Kleinschmidt, 2010). Why do we continually interleave
internally oriented processing in daily function? Our framework suggests this interleaving may
help to update internal models to reflect recent experience. In der Tat, brain activity occurring
after stimulus offset is associated with enhanced subsequent memory (Ben-Yakov & Dudai,
2011; Tambini, Ketz, & Davachi, 2010); disengaged time benefits creative problem solving
(Baird et al., 2012); and hundreds of experiments have demonstrated that information is bet-
ter retained when exposure is spaced across time rather than massed (Cepeda, Pasher, Vul,
Wixted, & Rohrer, 2006), suggesting that time away from information is important.
Netzwerkneurowissenschaften
345
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
Sharp wave ripple:
Local field potential deflection in the
hippocampus, accompanied by
140–250 Hz oscillations.
Awake rodents also spend substantial time in disengaged states. They replay past and possi-
ble future events during hippocampal sharp wave ripples in quiet rest periods (Diba & Buzsáki,
2007; Johnson & Redish, 2007), when sensory neocortex is in a synchronized “internal mode”
(McGinley et al., 2015B). Disrupting these ripples disrupts learning (Jadhav, Kemere, Deutsch,
& Frank 2012). Periods of quiet rest are associated with lower levels of acetylcholine relative
to active rest (Marrosu et al., 1995), and hippocampal sharp wave ripples are blocked by stim-
ulation of cholinergic inputs (Vandecasteele et al., 2014). Daher, this internal mode is again
associated with lower cholinergic tone.
The so-called default mode network is a set of interconnected regions far from the sensory
periphery, including the posterior parietal cortex, anterior medial cortex, and hippocampus
(Figure 2A). Although these individual areas, as discussed in the Hippocampus section above,
can oscillate between relatively internal and external modes of processing, their mean activa-
tion is associated with an overall internal mode of processing for the brain. The default mode
network is more active when participants are not engaged in a demanding externally oriented
Aufgabe (Mason et al., 2007; Raichle et al., 2001), and is therefore sometimes called a “task neg-
ative” network. Jedoch, regions of the default mode network can also be reliably recruited
by external input that demands understanding of semantics (Binder et al., 1999), constructing
scenarios (Hassabis & Maguire, 2007), comprehension of narratives or the viewpoint of others
(Mars et al., 2012; Simony et al., 2016), or imagining the past and future (Buckner & Carroll,
2007; Spreng, Mar, & Kim, 2009). These are all tasks that require use of high-level internal
Modelle. Daher, internal processing can be expressed as a persistent and brain-wide state, als
during mind-wandering, but it can also be more transiently and locally interwoven in ongoing
behavior and cognition in the external world.
Sleep and wake. At an even longer timescale, the transition between sleep and wake states
is perhaps the most dramatic example of switching between internal and external process-
ing modes. During sleep, there is minimal processing of immediate external input—neural
dynamics are governed almost exclusively by internal interactions.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
Wake-sleep algorithm:
An unsupervised learning algorithm
that iteratively samples bottom-up
Information (recognition
connections, “wake” phase) Und
top-down information (generative
connections, “sleep” phase).
Sleep benefits recently formed memories (Stickgold, 2013) and aids the integration of new
information into existing memory stores (Tamminen, Lambon Ralph, & Lewis, 2017). Weil
of the virtual absence of real-time environmental input, sleep may be an ideal time to update
internal models to better reflect recent information. The “wake-sleep” neural network learning
Algorithmus, which makes use of switches between internally and externally driven modes to
improve internally generated representations of the environment, was so named because of
this apparent correspondence (Hinton, Dayan, Frey, & Neal, 1995).
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Rapid eye movement (REM) und Non-REM-Schlaf. Within a night of sleep, there are several
alternations between non-REM and REM stages. This switching may also correspond to a rel-
atively internal versus external orientation, even within the sleeping brain. The hippocampus
stores traces of the details of recent experiences and replays these experiences during non-
REM sleep, often within sharp wave ripples (Nádasdy, Hirase, Czurkó, Csicsvari, & Buzsáki,
1999), which are temporally correlated with spindle events in cortex (Staresina et al., 2015).
This replay is thought to help “teach” cortex about these recent experiences, promoting sys-
tems consolidation—the transfer of information from hippocampus to cortex (McClelland,
In this sense, non-REM sleep approximates an externally
McNaughton, & O’Reilly, 1995).
driven mode for the neocortex. Although not as externally driven as when those events were
actually experienced, it is an opportunity to recap the details of the day’s events, providing
additional exposure to information that was recently acquired from the world.
Netzwerkneurowissenschaften
346
Learning by switching internal and external modes
During REM, in contrast, cortical dynamics are less influenced by the hippocampus and
driven more by long-established cortical representations (Diekelmann & Born 2010; Hasselmo,
1999). The interleaving of new information during non-REM with old information during REM
over the course of the night may then facilitate the integration of new memories into existing
cortical networks (Ficca & Salzarulo, 2004; Sara, 2017).
Daher, from the perspective of the cortex, non-REM sleep is an externally oriented mode,
using hippocampal retrieval as a proxy for the external world, whereas REM is an internally
oriented mode. Jedoch, the opposite is true from the viewpoint of the hippocampus. Dort,
low acetylcholine, as during wake, leads to a retrieval-like (internal) mode during non-REM
schlafen, and high acetylcholine leads to an encoding-like (extern) mode during REM sleep
(Hasselmo, 1999; Marrosu et al., 1995). These modes promote learning within the sleeping
hippocampus as well (Poe, Walsh, & Bjorness, 2010).
In sum, the non-REM/REM cycle may act as a shorter-timescale microcosm of the wake-
sleep cycle. In both the transition between sleep and wake and the transition between non-REM
und REM, there is a switch between relatively external and internal processing, happening at
opposing times for the sleeping hippocampus and cortex, which may drive learning to improve
internal models within these circuits.
CLARIFICATIONS, PREDICTIONS, AND OPEN QUESTIONS
Could the Brain Achieve Similar Learning Functions Without Switching Between Modes?
Why is it necessary to switch over time between external and internal modes? Could a cir-
cuit rather process feedforward and feedback signals simultaneously? Although feedforward
and feedback signals almost always commingle, there are two primary advantages to switch-
ing their relative efficacy. zuerst, if the same neural circuits simultaneously process external
and internal information, then it is difficult for those circuits to separate what is in the envi-
ronment from what is generated based on priors, memories, and expectations. When newly
encoded information is ubiquitously bound to retrieved memories it becomes difficult to dis-
tinguish related memories (Hasselmo & Bower, 1993), and reencoding of recently retrieved in-
formation can result in runaway synaptic strengthening (Newman, Shay, & Hasselmo, 2012B).
Zweitens, switching is an important ingredient in algorithms that converge toward accurate
high-dimensional internal models (Bengio et al., 2015; Heeger, 2017; O’Reilly, 1996). Als
noted by Marblestone, Wayne, and Kording (2016), although the specific implementations of
the relevant algorithms (recirculation, contrastive Hebbian learning, wake-sleep) may vary,
they all invoke “feedback connections that carry error phasically” (P. 6). The general principle
is that one should iteratively (A) use one’s best current model of the world to interpret incoming
data and (B) use new data to update one’s model of the world. Without switching, it is unclear
what data is coming from the environment, and so it is difficult to adjust the internal model
to better match that environment. Without switching, it is also difficult to fully instantiate an
internal representation when receiving strong environmental input.
What if different neural circuits were specialized for representing internal and external in-
Formation, thus avoiding the need for alternating modes over time? Although the brain does
separate externally biased and internally biased information sources in space (sensory versus
higher-order cortical regions, wie in Abbildung 2; granular versus supragranular layers of cortex;
possibly even dendrites versus soma, Guergiuev, Lillicrap, & Richards, 2016), das tut nicht
obviate the benefits of switching. Because switching provides individual neurons with infor-
mation from prior and subsequent layers in a neural network, it provides a higher-dimensional
347
Contrastive Hebbian learning:
Learning by adjusting connection
weights between neuron i and j
based on the difference in i − j
coactivity across bottom-up and
top-down phases.
Granular neocortex:
Intermediate layers of the cerebral
Kortex, which typically contain many
granule cells and are often a major
recipient of thalamic drive.
Netzwerkneurowissenschaften
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
learning signal, which is important for learning higher-dimensional models. It is possible for
neurons to update their weights using a common scalar error (such as a reward signal) das ist
generated in a separate neural system, but this form of learning does not solve the credit as-
signment problem in multilayer systems and is inefficient for high-dimensional representations
(Marblestone et al., 2016; Werfel, Xie, & Seung, 2005). One could generate an appropriately
high-dimensional learning signal in a separate error circuit if that circuit was of comparable
complexity to the circuit being trained, but this would be a very costly approach for wiring, Und
would require continuous coordination between the trainer and trainee. Stattdessen, a relatively
simple and robust approach is to allow circuits to generate error signals locally by switching
between consecutive bottom-up and top-down biased activity modes. This switching approach
has been adopted not only in machine learning (Kasten 1) but also in neuronal circuit models of
learning (z.B., O’Reilly et al., 2014).
Kasten 1. Sufficient ingredients for switch-based learning
Which of the many fluctuating processes in the nervous system will drive learning? Obwohl
it is difficult to say what ingredients are necessary, two general characteristics appear to be
sufficient: (A) The fluctuating modes of a circuit should be consecutively biased toward inter-
nal and external sources of information; (B) Processes for updating synaptic weights must be
sensitive to coactivity of neurons as well as to whether activity occurs in an internally biased
or externally biased phase.
(A) Internal and external switching: One phase of a fluctuation needs to be more influenced
by feedforward information, while the other phase needs to be more influenced by long-term
expectations. The external information is associated with volleys of spiking arriving from the
sensory periphery. The internal information is associated with volleys of spiking arriving from
higher-order cortical systems, and with patterns of synaptic connections that are stable over
days or longer.
(B) Plasticity dependent on activity across switches: A number of algorithms have been pro-
posed for training high-dimensional multilayer models by comparing local activity across
and within switches. Contrastive Hebbian learning is a supervised learning approach that
can approximate backpropagation in multilayer settings (O’Reilly, 1996; Xie & Seung, 2003;
see also Scellier & Bengio, 2017).
In contrastive Hebbian learning, the weight update can
be thought of as either (A) two separate weight updates with opposite signs during internal
and external phases; oder (B) a single weight update based on the difference in coactivity dur-
ing presentation of a stimulus and during presentation of the corresponding target. A more
neurobiologically grounded learning process is provided by the XCAL rule (O’Reilly et al.,
2014), which updates weights based on the difference in coactivity when the network is near
to and far from attractors; switching the gain of feedforward drive can push the system toward
and away from such attractors. Endlich, Hinton (2007) and Bengio et al. (2015) have argued
that spike-timing-dependent plasticity can approximate a weight update similar to backprop-
agation. Although weight updates are applied in the same way during internal or external
phases, switching of the network between feedforward- and feedback-dominated dynamics
is still required under these proposals. As Bengio et al. (2015) notiert, one of the missing in-
gredients for a biologically plausible approximation to backpropagation is for computations
to be “clocked to alternate between feedforward and backpropagation phases (since the latter
needs the formers results)” (P. 1).
Two general observations apply to switch-based learning. Erste, explicit supervision signals
are not a necessary component for switch-based learning in multilayer systems, because each
layer can treat its subsequent layer as an activation target. Zweite, switch-based learning need
not be restricted to neurons and synapses, and could also operate at the level of neuronal
assemblies that are adjusting their mutual efficacies.
Supervised learning:
A learning process in which input
data are paired with explicit labels or
Rückmeldung.
Netzwerkneurowissenschaften
348
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
How Can an Experimentalist Decide Whether a Region is in an Internal or External Mode?
It is important to clarify that internal and external modes are not determined by anatomy but by
information flow (Figure 1A). A circuit is pushed toward an external mode when it is influenced
by information arriving from the world. A circuit is pushed toward an internal mode when it
is influenced by information arriving from an internal model (or set of priors) regarding the
Welt. At any given moment, each circuit is subject to both internal and external influences
in some mixture, and their relative strengths will vary (z.B., Figure 1B). We have suggested,
as a rule of thumb, that pathways closer to the sensory periphery tend to convey external
Information, and so when these inputs are dominant then a circuit is in an external mode.
Jedoch, the flow of information can vary along fixed anatomy (as in the case of cortico-
hippocampal interactions during wake and sleep). Zusätzlich, fixed anatomy may not have
an easily interpretable “bottom up” or “top down” status: Projections to the hippocampus from
the nucleus reunions appear to be “bottom up,” as they originate in the thalamus, but because
they may convey information from the prefrontal cortex, they could be considered “top down.”
Although the large-scale anatomical architecture is strongly correlated with internal and ex-
ternal information flow (Figure 2A), more difficult cases can be decided via real (or imagined)
interventional tests. Insbesondere, the balance between internal and external information can
be determined by measuring the effects of (A) changing the world state and (B) changing the
content of the internal model. To measure external influence, one might ask the following:
If I make a small, transient change in the world (z.B., change the shape of a perceived face),
how much would that affect the current activity of the circuit I am measuring? To measure the
influence of internal information, one might ask the following: If I make a long-lasting change
to the state of the world (z.B., change the shape of all faces that are perceived and learned
over years), then how much would that affect the present state of the circuit? The balance of
internal and external information can be derived based on which kind of manipulation has the
greater effect on the circuit at a particular moment.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Predictions
(A) The strongest prediction of our framework, for which there is already evidence in some
of the discussed examples, is that switching between internally and externally biased modes
supports error-driven learning. daher, in the case of non-REM/REM cycles, Zum Beispiel, A
disruption of the switching should produce a deficit in integrating new hippocampal memories
with old information stored in neocortex. Ähnlich, if the relative contribution of feedforward
and feedback influences is altered, this should also impair learning of internal models. Für
Beispiel, if the time in external modes is relatively lengthened, our framework predicts that
new information would be encoded but would be less well linked to relevant aspects of the
existing model; if time in external modes is shortened, then new experience would not be
sufficient to correct errors in the existing model.
(B) If fluctuations between internally and externally biased modes reflect a learning process,
then the timescale of fluctuation is a timescale over which error signals are computed. Daher,
the error signal that drives learning on subsecond scales (z.B., across hippocampal theta cycles
or visual cortical alpha cycles) is computed on subsecond properties of the world, and models
subsecond relationships of this input. Im Gegensatz, the error signal computed across waking
and sleeping states can be influenced not only by transient features of the world, but also
by external features that vary over many hours. Daher, sleep may be especially necessary for
learning associations between events separated by many minutes or hours.
Netzwerkneurowissenschaften
349
Learning by switching internal and external modes
Within the neocortex, the timescale of fluctuation slows as one moves up the processing
hierarchy (Hasson, Chen, & Honey, 2015; Honey et al., 2012). If this fluctuation represents a
timescale for comparison that drives learning, then earlier cortical regions should be biased
to learn mostly about transient features of their input, while higher-order cortical regions may
learn in response to environmental features that change over many seconds or even minutes.
This could be tested by presenting a stimulus sequence that contains structure on both short
timescales (z.B., phonemes in an artificial language) and long timescales (z.B., probabilistic
syntax in an artificial language). The internal/external model can then be tested by manip-
ulating (via pharmacological or electromagnetic bias) the rate of switching between internal
and external modes. More rapid switching should lead participants to behaviorally exhibit
improved learning of the short timescale structure, relative to the long timescale structure.
Allgemeiner, if the extraction of slow features leads to the learning of invariant representa-
tionen (Wiskott & Sejnowski, 2002), increasingly slow mode fluctuations may bias consecutive
stages of processing to learn increasingly invariant representations. Computational modeling
indicates that, when learning multiscale structure that is nested in time, a learning system en-
dowed with a hierarchy of slowness can be more powerful (Mozer, 1992) and more efficient
(Chung, Ahn, & Bengio, 2016; Schmidhuber, 1992), with higher-order structure being learned
by the more slowly varying components of the model.
(C) The learning rate of an individual region might vary inversely according to the speed of its
internal/external switching. The learning rate can be thought of as the magnitude of synaptic
weight change at each learning opportunity. Learning rates likely vary as one descends the
hierarchy from the hippocampus (high learning rate) to medial and inferior temporal regions
(medium learning rate) to early visual cortex (slow learning rate; Ahissar & Hochstein, 2004;
Norman & O’Reilly, 2003; Yang & Maunsell, 2004; see also Lee & Sachsen, 2014).
(D) A final prediction of our model is that in order to drive learning, there should exist a
local mechanism in each neural system for comparing predicted states with observed states. Wenn
mode-switching occurs on the timescale of seconds, then information about the prior state can
be stored in circuit activation (z.B., a slowly changing distributed pattern; Baldassano et al.,
2016) and differences may be computed continuously in all phases (Hasselmo et al., 2002;
Norman et al., 2006). Jedoch, in systems where the internal-external switching occurs more
slowly (z.B., over minutes), implementing an update rule such as contrastive Hebbian learning
(see Box 1) would pose a greater challenge. There would need to be either (A) a trace that can
persist over minutes or longer, to allow for the computation of mismatch between the internal
and external phases, oder (B) a way to switch the sign of the weight update rule between the
internal and external phases. There are many biological and circuit mechanisms that can store
single-exposure information across minutes and hours (Barak & Tsodyks, 2014; Benna & Fusi,
2016; Reinartz, Biro, Gal, Giugliano, & Marom, 2014), but it is an important question to con-
sider which of these mechanisms could support computation of the mismatch between past
and present.
Open Questions
Once a mismatch is detected, when are internal models updated to reflect this mismatch? Is
there a bias to perform model updating during the internal or external mode? Increased cholin-
ergic and noradrenergic tone is associated with increased stimulus-dependent plasticity in the
neocortex and the hippocampus (Gu, 2002). At first glance, this suggests that mismatch-based
updates to the internal model may occur preferentially during the “external” mode. Jedoch,
Netzwerkneurowissenschaften
350
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
plasticity is rarely studied in settings where synapses are exposed to interleaved bottom-up
and top-down drive or in which neuromodulatory tone and stimulus features are varied dy-
namically. Daher, it remains an important open question whether the biological instantiation
of model updating occurs during one mode, or whether it may depend in a more complex
manner on the timing of the switching process.
How does the ongoing switch-based learning we have described relate to reinforcement?
Traditionell, reinforcement learning is considered a special case of error-driven learning in
which values are updated based on the difference between expected and received reward—
the reward prediction error (Sutton & Barto, 1998). These differences between expectation
and outcome can also be used to build models of the environment, thought to be used by the
basal ganglia and medial prefrontal cortex to predict outcomes and select actions (Alexander
& Braun, 2011; Daw, Gershman, Seymour, Dayan, & Dolan, 2011). There are clear connec-
tions between the principles of adaptive task-related neuromodulation (Yu & Dayan, 2005)
and the internal and external switching that we describe here. We have focused on the learn-
ing of high-dimensional multilevel internal models, a setting in which scalar reinforcement
is considered impractically inefficient on its own (Werfel et al., 2005). dennoch, Die
“internally generated” mode-switching processes we have described will be strongly mod-
ulated by goal state and reinforcement in real behavior (Kasten 2). It has even been suggested
that basal forebrain cholinergic projections, which modulate both cortical circuit state and
plasticity (Sugihara, Chen, & Sur, 2016), may provide a supervisory signal that modulates
local sensory learning (Hangya, Ranade, Lorenc, & Kepecs, 2015; see also Poort et al., 2015).
Diffuse supervision signals, including those from reward, could greatly accelerate the switch-
based learning we have described, and so are an important topic for future developments of
our framework.
Kasten 2. What drives switches between internal and external modes?
The transition between internal and external modes can be driven by both exogenous and
endogenous factors. Exogenous factors, which come from the sensorium, include reward-
ing, unexpected, or otherwise salient stimuli. Endogenous factors, which can operate inde-
pendently of environmental input, include receptor recovery from inactivation, endogenous
rhythmicity in neuronal population activity, as well as circadian and homeostatic metabolic
processes.
The exogenous and endogenous causes of switching can proceed independently, but in most
settings they will be coupled. Zum Beispiel, a salient stimulus can shift or reset the phase of an
otherwise endogenous oscillation, and circadian clocks become aligned to the presentation
of light and food. Daher, organisms will switch to an external mode when more information is
needed from the environment for the current task, and in parallel with this purposive switch-
ing, there are also multiple timescales of switching that arise from entrainment of endogenous
fluctuations to exogenous cues.
CONCLUSION
Motivated by the hypothesis that internal-external mode-switching can generically support
learning, we have reviewed neural circuits and systems that exhibit such fluctuations. Func-
tionally relevant switching unfolds in the hippocampus and neocortical regions on the scale of
milliseconds and seconds, and diffusely across the cerebrum, over minutes and hours within
Netzwerkneurowissenschaften
351
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
and across wake and sleep. The functional influence of cholinergic tone is similar across these
settings and timescales.
The fluctuating processes we reviewed cannot be present only because they serve learning:
They arise within a broader context of metabolic dynamics and developmental and evolution-
ary constraints. Jedoch, learning is a fundamental function of nervous systems, and internal
models are ubiquitously useful in the brain, because they allow for more efficient representa-
tions and robustness to noisy input. Switch-based learning allows internal models to be shaped
in a natural and general manner, using only local information. The ingredients of such learn-
ing appear to be available in many instances where biological systems oscillate around a set
Punkt (Kasten 1). In this way, each neural circuit, while swimming in the idiosyncratic inputs that
compose its world, moves gradually toward a more accurate model of that world.
ACKNOWLEDGMENTS
The authors thank Aaron Bornstein, Roy Cox, Michael Hasselmo, Kathrin Müsch, Kenneth
Norman, and Robert Stickgold for useful comments on earlier versions of this manuscript.
BEITRÄGE DES AUTORS
Christopher J. Honey: Konzeptualisierung; Writing – original draft. Ehren L. Newman: Concep-
tualization; Writing – original draft. Anna C. Schapiro: Konzeptualisierung; Writing – original
Entwurf; Christopher J. Honey, Ehren L. Newman, and Anna C. Schapiro: Writing – review &
Bearbeitung.
FUNDING INFORMATION
The authors gratefully acknowledge the support of the National Institutes of Health (F32-
NS093901 to ACS; MH111439-01 subaward CJH) and the Sloan Foundation (Forschung
Fellowship to CJH).
VERWEISE
Adrian, E. D., & Matthews, B. H. C.
(1934). The berger rhythm:
Potential changes from the occipital lobes in man. Gehirn, 57(4),
355–385.
Ahissar, M., & Hochstein, S. (2004). The reverse hierarchy theory of
visual perceptual learning. Trends in den Kognitionswissenschaften, 8(10),
457–464.
Alexander, W. H., & Braun, J. W. (2011). Medial prefrontal cortex
as an action-outcome predictor. Naturneurowissenschaften, 14(10),
1338–1344.
Baird, B., Smallwood, J., Mrazek, M. D., Kam, J. W., Franklin, M. S.,
& Schooler, J. W. (2012). Inspired by distraction mind wander-
ing facilitates creative incubation. Psychological Science, 23(10),
1117–1122.
Baldassano, C., Chen, J., Zadbood, A., Pillow, J. W., Hasson, U., &
Norman, K. A. (2016). Discovering event structure in continuous
narrative perception and memory. bioRxiv, 081018.
Barak, O., & Tsodyks, M.
(2014). Working models of working
Bastos, A. M., Vezoli,
Erinnerung. Aktuelle Meinung in der Neurobiologie, 25, 20–24.
J., Bosman, C. A., Schoffelen,
J. M.,
Oostenveld, R., Dowdall, J. R., . . . Fries, P.
(2015). Visual
areas exert feedforward and feedback influences through distinct
frequency channels. Neuron, 85(2), 390–401.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Bengio, Y., Lee, D. H., Bornschein, J., Mesnard, T., & Lin, Z. (2015).
Towards biologically plausible deep learning. arXiv:1502.04156
(2016). Computational principles of
synaptic memory consolidation. Naturneurowissenschaften, 19(12),
1697.
Benna, M. K., & Fusi, S.
Ben-Yakov, A., & Dudai, Y. (2011). Constructing realistic engrams:
Poststimulus activity of hippocampus and dorsal striatum predicts
subsequent episodic memory. Zeitschrift für Neurowissenschaften, 31(24),
9032–9042.
Bieri, K. W., Bobbitt, K. N., & Colgin, L. L. (2014). Slow and fast
gamma rhythms coordinate different spatial coding modes in
hippocampal place cells. Neuron, 82(3), 670–681.
Binder, Jeffrey R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S.,
(1999). Conceptual processing
Rao, S. M., & Cox, R. W.
during the conscious resting state: A functional MRI study.
Zeitschrift für kognitive Neurowissenschaften, 11(1), 80–93.
Bittner, K. C., Grienberger, C., Vaidya, S. P., Milstein, A. D.,
(2015). Conjunctive
Macklin, J. J., Suh, J., . . . Magee, J. C.
input processing drives feature selectivity in hippocampal CA1
Neuronen. Naturneurowissenschaften, 18(8), 1133–1142.
Brankack, J., Stewart, M., & Fuchs, S. E. (1993). Current source density
analysis of the hippocampal theta rhythm: Associated sustained
Netzwerkneurowissenschaften
352
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
.
T
Learning by switching internal and external modes
potentials and candidate synaptic generators. Gehirnforschung,
615(2), 310–327.
Buckner, R. L., & Carroll, D. C.
(2007). Self-projection and the
Gehirn. Trends in den Kognitionswissenschaften, 11(2), 49–57.
Busch, N. A., Dubois, J., & VanRullen, R.
ongoing EEG oscillations predicts visual perception.
Neurowissenschaften, 29(24), 7869–7876.
(2009). The phase of
Zeitschrift für
Buzsáki, G. (2002). Theta oscillations in the hippocampus. Neuron,
33(3), 325–340.
Buzsáki, G., Czopf, J., Kondákor, ICH., & Kellényi, L. (1986). Laminar
distribution of hippocampal rhythmic slow activity (RSA) im
behaving rat: Current-source density analysis, effects of urethane
and atropine. Gehirnforschung, 365(1), 125–137.
Buzsáki, G., & Draguhn, A. (2004). Neuronal oscillations in cortical
Netzwerke. Wissenschaft, 304(5679), 1926–1929.
Cepeda, N. J., Pashler, H., Vul, E., Wixted, J. T., & Rohrer, D. (2006).
Distributed practice in verbal recall tasks: A review and quan-
titative synthesis. Psychological Bulletin, 132(3), 354.
Chung, J., Ahn, S., & Bengio, Y.
(2016). Hierarchical multiscale
recurrent neural networks. arXiv:1609.01704
Colgin, L. L., Denninger, T., Fyhn, M., Hafting, T., Bonnevie, T.,
Jensen, O., . . . Moser, E. ICH. (2009). Frequency of gamma oscil-
lations routes flow of information in the hippocampus. Natur,
462(7271), 353–357.
Daw, N. D., Gershman, S. J., Seymour, B., Dayan, P., & Dolan, R. J.
(2011). Model-based influences on humans’ choices and striatal
prediction errors. Neuron, 69(6), 1204–1215.
de Gee,
J. W., Colizoli, O., Kloosterman, N. A., Knapen, T.,
Nieuwenhuis, S., & Donner, T. H.
(2017). Dynamic modula-
tion of decision biases by brainstem arousal systems. eLife, 6,
e23232.
Della Rossa, F., Dercole, F., & Piccardi, C.
(2013). Profiling
core-periphery network structure by random walkers. Scientific
Reports, 3, 1467.
Diba, K., & Buzsäki, G.
(2007). Forward and reverse hippocam-
pal place-cell sequences during ripples. Naturneurowissenschaften,
10(10), 1241–1242.
Diekelmann, S., & Born, J. (2010). The memory function of sleep.
Nature Reviews Neurowissenschaften, 11(2), 114–126.
Disney, A. A., Aoki, C., & Hawken, M. J. (2007). Gain modulation
by nicotine in macaque v1. Neuron, 56(4), 701–713.
Douchamps, V., Jeewajee, A., Blundell, P., Bürger, N., & Lever,
C. (2013). Evidence for encoding versus retrieval scheduling in
Zeitschrift für
the hippocampus by theta phase and acetylcholine.
Neurowissenschaften, 33(20), 8689–8704.
Duncan, K., Sadanand, A., & Davachi, L. (2012). Memory’s penum-
bra: Episodic memory decisions induce lingering mnemonic
biases. Wissenschaft, 337(6093), 485–487.
Engel, T. A., Steinmetz, N. A., Gieselmann, M. A., Thiele, A.,
Moore, T., & Boahen, K.
Selective modulation
of cortical state during spatial attention. Wissenschaft, 354(6316),
1140–1144.
(2016).
Fernández-Ruiz, A., Oliva, A., Groß, G. A., Maurer, A. P., Berényi,
A., & Buzsáki, G.
(2017). Entorhinal-CA3 dual-input control
of spike timing in the hippocampus by theta-gamma coupling.
Neuron, 93, 1213–1226.
Ficca, G., & Salzarulo, P.
(2004). What in sleep is for memory.
Sleep Medicine, 5(3), 225–230.
Gil, Z., Connors, B. W., & Amitai, Y.
(1997). Differential regu-
lation of neocortical synapses by neuromodulators and activity.
Neuron, 19(3), 679–686.
Goard, M., & Und, Y. (2009). Basal forebrain activation enhances
cortical coding of natural scenes. Naturneurowissenschaften, 12(11),
1444–1449.
Groppe, D. M., Bickel, S., Keller, C. J., Jain, S. K., Hwang, S. T.,
Harden, C., & Mehta, A. D. (2013). Dominant frequencies of rest-
ing human brain activity as measured by the electrocorticogram.
NeuroImage, 79, 223–233.
Gu, Q.
(2002). Neuromodulatory transmitter systems in the cor-
tex and their role in cortical plasticity. Neurowissenschaften, 111(4),
815–835.
Guergiuev, J., Lillicrap, T. P., & Richards, B. A. (2016). Deep learn-
ing with segregated dendrites. arXiv:1610.00161
Halgren, E., Kaestner, E., Marinkovic, K., Cash, S. S., Wang, C.,
Schomer, D. L., . . . Ulbert, ICH.
Laminar profile of
spontaneous and evoked theta: Rhythmic modulation of cortical
processing during word integration. Neuropsychologie, 76,
108–124.
(2015).
Hangya, B., Ranade, S. P., Lorenc, M., & Kepecs, A. (2015). Cen-
tral cholinergic neurons are rapidly recruited by reinforcement
Rückmeldung. Cell, 162(5), 1155–1168.
Hassabis, D., & Maguire, E. A.
(2007). Deconstructing episodic
memory with construction. Trends in den Kognitionswissenschaften, 11(7),
299–306.
Hasselmo, M. E. (1995). Neuromodulation and cortical function:
Modeling the physiological basis of behavior. Behavioural Brain
Forschung, 67(1), 1–27.
Hasselmo, M. E.
memory consolidation.
351–359.
(1999). Neuromodulation: Acetylcholine and
Trends in den Kognitionswissenschaften, 3(9),
Hasselmo, M. E. (2006). The role of acetylcholine in learning and
Erinnerung. Aktuelle Meinung in der Neurobiologie, 16(6), 710–715.
Hasselmo, M. E., Bodelón, C., & Wyble, B. P. (2002). A proposed
function for hippocampal theta rhythm: Separate phases of en-
coding and retrieval enhance reversal of prior learning. Neuronal
Computation, 14(4), 793–817.
Hasselmo, M. E., & Bower, J. M. (1992). Cholinergic suppression
specific to intrinsic not afferent fiber synapses in rat piriform (ol-
factory) Kortex. Journal of Neurophysiology, 67(5), 1222–1229.
Hasselmo, M. E., & Bower, J. M. (1993). Acetylcholine and memory.
Trends in Neuroscience, 16, 218–222.
Hasselmo, M. E., & Schnell, E.
(1994). Laminar selectivity of
the cholinergic suppression of synaptic transmission in rat hip-
pocampal region CA1: Computational modeling and brain slice
physiology. Zeitschrift für Neurowissenschaften, 14(6), 3898–3914.
Hasson, U., Chen, J., & Honey, C. J. (2015). Hierarchical process
Erinnerung: Memory as an integral component of information
Verarbeitung. Trends in den Kognitionswissenschaften, 19(6), 304–313.
Hebb, D. Ö. (1949). The organization of behavior: A neuropsycho-
logical theory. New York: Wiley.
Heeger, D. J.
(2017). Theory of cortical function. Proceedings of
the National Academy of Sciences, 114(8), 1773–1782.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Netzwerkneurowissenschaften
353
Learning by switching internal and external modes
Hinton, G. E. (2007). How to do backpropagation in a brain. Invited
talk at the NIPS 2007 Deep Learning Workshop.
Hinton, G. E., Dayan, P., Frey, B. J., & Neal, R. M.
(1995).
The “wake-sleep” algorithm for unsupervised neural networks.
Wissenschaft, 268(5214), 1158.
Hinton, G. E., & McClelland, J. L.
(1988). Learning representa-
tions by recirculation. In Neural information processing systems
(S. 358–366). New York: American Institute of Physics.
Honey, C. J., Thesen, T., Donner, T. H., Silbert, L. J., Carlson, C. E.,
Devinsky, Ö, . . . Hasson, U. (2012). Slow cortical dynamics and
the accumulation of information over long timescales. Neuron,
76(2), 423–434.
Hopfield, J. J. (1982). Neural networks and physical systems with
emergent collective computational abilities. Verfahren der
Nationale Akademie der Wissenschaften, 79(8), 2554–2558.
Hsieh, C. Y., Cruikshank, S. J., & Metherate, R. (2000). Differential
modulation of auditory thalamocortical and intracortical synap-
tic transmission by cholinergic agonist. Gehirnforschung, 880(1),
51–64.
Huerta, P. T., & Lisman, J. E. (1995). Bidirectional synaptic plastic-
ity induced by a single burst during cholinergic theta oscillation
in CA1 in vitro. Neuron, 15(5), 1053–1063.
Hyman, J. M., Wyble, B. P., Goyal, V., Rossi, C. A., & Hasselmo,
M. E. (2003). Stimulation in hippocampal region CA1 in behav-
ing rats yields long-term potentiation when delivered to the peak
of theta and long-term depression when delivered to the trough.
Zeitschrift für Neurowissenschaften, 23(37), 11725–11731.
Itskov, V., Pastalkova, E., Mizuseki, K., Buzsáki, G., & Harris, K. D.
information in
(2008).
hippocampus. Zeitschrift für Neurowissenschaften, 28(23), 5959–5964.
Theta-mediated dynamics of spatial
Jadhav, S. P., Kemere, C., Deutsch, P. W., & Frank, L. M.
(2012).
Awake hippocampal sharp-wave ripples support spatial memory.
Wissenschaft, 336(6087), 1454–1458.
Jasper, H., & Penfield, W.
(1949). Electrocorticograms in man:
Effect of voluntary movement upon the electrical activity of the
präzentraler Gyrus. Archiv für Psychiatrie und Nervenkrankheiten,
183(1–2), 163–174.
Jensen, O., & Mazaheri, A. (2010). Shaping functional architecture
by oscillatory alpha activity: Gating by inhibition. Grenzen in
Human Neuroscience, 4, 186.
Johnson, A., & Redish, A. D. (2007). Neural ensembles in CA3 tran-
siently encode paths forward of the animal at a decision point.
Zeitschrift für Neurowissenschaften, 27(45), 12176–12189.
(2016).
Individual human brain areas can
be identified from their characteristic spectral activation finger-
prints. PLoS Biology, 14(6), e1002498.
Keitel, A., & Brutto, J.
Killingsworth, M. A., & Gilbert, D. T. (2010). A wandering mind is
an unhappy mind. Wissenschaft, 330, 932.
Schlosser, P., Karmos, G., Mehta, A. D., Ulbert, ICH., & Schroeder, C. E.
(2008). Entrainment of neuronal oscillations as a mechanism of
attentional selection. Wissenschaft, 320(5872), 110–113.
Lee, R., & Sachsen, A. M. (2014). Modeling perceptual learning with
deep networks. 36th annual meeting of the Cognitive Science
Society, Quebec.
Leopold, D. A., Murayama, Y., & Logothetis, N. K.
(2003).
Very slow activity fluctuations in monkey visual cortex: Impli-
cations for functional brain imaging. Hirnrinde, 13(4),
422–433.
Lillicrap, T. P., Cownden, D., Tweed, D. B., & Akerman, C. J. (2016).
Random synaptic feedback weights support error backpropaga-
tion for deep learning. Nature Communications, 7.
Linkenkaer-Hansen, K., Nikulin, V. V., Verehrung, S., Ilmoniemi, R. J.,
(2004). Prestimulus oscillations enhance psy-
Zeitschrift für Neurowissenschaften,
& Verehrung, J. M.
chophysical performance in humans.
24(45), 10186–10190.
Linster, C., & Cleland, T. A.
(2016). Neuromodulation of olfac-
tory transformations. Aktuelle Meinung in der Neurobiologie, 40,
170–177.
Marblestone, A. H., Wayne, G., & Kording, K. P.
(2016). Toward
an integration of deep learning and neuroscience. Grenzen in
Computational Neuroscience, 10.
Margulies, D. S., Ghosh, S. S., Goulas, A., Falkiewicz, M.,
Huntenburg, J. M., Langs, G., . . . Jefferies, E. (2016). Situating the
default-mode network along a principal gradient of macroscale
cortical organization. Proceedings of the National Academy of
Wissenschaften, 113(44), 12574–12579.
Marrosu, F., Portas, C., Mascia, M. S., Casu, M. A., Fá, M.,
Giagheddu, M., . . . Gessa, G. L. (1995). Microdialysis measure-
ment of cortical and hippocampal acetylcholine release during
sleep-wake cycle in freely moving cats. Gehirnforschung, 671(2),
329–332.
Mars, R. B., Neubert, F. X., Noonan, M. P., Sallet, J., Toni, ICH., &
Rushworth, M. F. (2012). On the relationship between the “de-
fault mode network” and the “social brain.” Frontiers in Human
Neurowissenschaften, 6, 189.
Mason, M. F., Norton, M. ICH., Van Horn, J. D., Wegner, D. M.,
(2007). Wandering minds:
Grafton, S. T., & Macrae, C. N.
The default network and stimulus-independent thought. Wissenschaft,
315(5810), 393–395.
McClelland, J. L., McNaughton, B. L., & O’Reilly, R. C.
(1995).
Why there are complementary learning systems in the hippocam-
pus and neocortex: Insights from the successes and failures of
connectionist models of learning and memory. Psychological
Rezension, 102(3), 419.
McGinley, M. J., David, S. V., & McCormick, D. A. (2015A). Cor-
tical membrane potential signature of optimal states for sensory
signal detection. Neuron, 87(1), 179–192.
McGinley, M. J., Vinck, M., Reimer, J., Batista-Brito, R., Zagha,
E., Cadwell, C. R., . . . McCormick, D. A.
(2015B). Waking
state: Rapid variations modulate neural and behavioral re-
sponses. Neuron, 87(6), 1143–1161.
Meeter, M., Murre, J. M. J., & Talamini, L. M. (2004). Mode shifting
between storage and recall based on novelty detection in oscil-
lating hippocampal circuits. Hippocampus, 14(6), 722–741.
Müller, K. J., Honey, C. J., Hermes, D., Rao, R. P., & Ojemann, J. G.
(2014). Broadband changes in the cortical surface potential track
activation of functionally diverse neuronal populations. Neuro-
Image, 85, 711–720.
Mizumori, S. J. Y., Perez, G. M., Alvarado, M. C., Barnes, C. A., &
McNaughton, B. L. (1990). Reversible inactivation of the medial
septum differentially affects two forms of learning in rats. Gehirn
Forschung, 528(1), 12–20.
Netzwerkneurowissenschaften
354
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
Monto, S., Verehrung, S., Voipio, J., & Verehrung, J. M. (2008). Very slow EEG
fluctuations predict the dynamics of stimulus detection and os-
cillation amplitudes in humans. Zeitschrift für Neurowissenschaften, 28(33),
8268–8272.
Mozer, M. C. (1992). Induction of multiscale temporal structure. In
J. E. Moody, S. J. Hanson & R. P. Lippmann (Hrsg.), Advances
in Neural
Information Processing Systems 4 (S. 275–282).
Retrieved from https://papers.nips.cc
Nádasdy, Z., Hirase, H., Czurkó, A., Csicsvari, J., & Buzsáki, G.
(1999). Replay and time compression of recurring spike se-
quences in the hippocampus. Zeitschrift für Neurowissenschaften, 19(21),
9497–9507.
Newman, E. L., Gillet, S. N., Climer, J. R., & Hasselmo, M. E.
(2013).
Cholinergic blockade reduces theta-gamma phase
amplitude coupling and speed modulation of theta frequency
Zeitschrift für
consistent with behavioral effects on encoding.
Neurowissenschaften, 33(50), 19635–19646.
Newman, E. L., Gupta, K., Climer,
J. R., Monaghan, C. K., &
Hasselmo, M. E. (2012A). Cholinergic modulation of cognitive
Verarbeitung: Insights drawn from computational models. Frontiers
in Behavioral Neuroscience, 6, 24.
Newman, E. L., Shay, C. F., & Hasselmo, M. E. (2012B). Malignant
synaptic growth and Alzheimer’s disease. Future Neurology, 7(5),
557–571.
Norman, K. A., Newman, E., Detre, G., & Polyn, S. M. (2006). Wie
inhibitory oscillations can train neural networks and punish com-
petitors. Neural Computation, 18(7), 1577–1610.
Norman, K. A., & O’Reilly, R. C.
(2003). Modeling hip-
pocampal and neocortical contributions to recognition memory:
Psychological
A complementary-learning-systems approach.
Rezension, 110(4), 611.
O’Keefe, J., & Recce, M. L. (1993). Phase relationship between hip-
pocampal place units and the EEG theta rhythm. Hippocampus,
3(3), 317–330.
O’Reilly, R. C. (1996). Biologically plausible error-driven learning
using local activation differences: The generalized recirculation
Algorithmus. Neural Computation, 8(5), 895–938.
O’Reilly, R. C., Wyatte, D., & Rohrlich, J (2014). Learning through
time in the thalamocortical loops. arXiv:1407.3432
Verehrung, J. M., & Verehrung, S.
(2011). 22 roles of multiscale brain
activity fluctuations in shaping the variability and dynamics
of psychophysical performance. Progress in Brain Research,
193, 335.
Pfurtscheller, G., & Da Silva, F. L. (1999). Event-related EEG/MEG
synchronization and desynchronization: Basic principles. Clini-
cal Neurophysiology, 110(11), 1842–1857.
Pinto, L., Goard, M. J., Estandian, D., Xu, M., Kwan, A. C.,
Fast modulation of visual
(2013).
forebrain cholinergic neurons. Natur
Lee, S. H., . . . Und, Y.
perception by basal
Neurowissenschaften, 16(12), 1857–1863.
Poe, G. R., Walsh, C. M., & Bjorness, T. E.
(2010). Kognitiv
neuroscience of sleep. Progress in Brain Research, 185, 1.
Poort, J., Khan, A. G., Pachitariu, M., Nemri, A., Orsolic, ICH., Krupic,
J., . . . Hofer, S. B.
(2015). Learning enhances sensory and
multiple non-sensory representations in primary visual cortex.
Neuron, 86(6), 1478–1490.
Rachel, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L.
(2001). A default mode of
brain function. Verfahren der Nationalen Akademie der Wissenschaften,
98(2), 676–682.
Reinartz, S., Biro, ICH., Gal, A., Giugliano, M., & Marom, S. (2014).
Synaptic dynamics contribute to long-term single neuron
response fluctuations. Frontiers in Neural Circuits, 8, 71.
Rosenblatt, F.
(1958). The perceptron: A probabilistic model for
information storage and organization in the brain. Psychological
Rezension, 65(6), 386.
Rumelhart, D. E., Hinton, G. E., & Williams, R. J. (1986). Learning
representations by back-propagating errors. Natur, 323(6088),
533–536.
Sadaghiani, S., Hesselmann, G., Friston, K. J., & Kleinschmidt, A.
(2010). The relation of ongoing brain activity, evoked neural
responses, and cognition. Frontiers in Systems Neuroscience,
4, 20.
Sara, S. J.
(2017). Sleep to remember.
Zeitschrift für Neurowissenschaften,
37(3), 457–463.
Scellier, B., & Bengio, Y. (2017). Equilibrium propagation: Bridg-
ing the gap between energy-based models and backpropagation.
Frontiers in Computational Neuroscience, 11.
Schmidhuber, J.
(1992). Learning complex, extended sequences
using the principle of history compression. Neural Computation,
4(2), 234–242.
Schomburg, E. W., Fernández-Ruiz, A., Mizuseki, K., Berényi,
A., Anastassiou, C. A., Koch, C., & Buzsáki, G.
(2014).
Theta phase segregation of input-specific gamma patterns in
entorhinal-hippocampal networks. Neuron, 84(2), 470–485.
Sherman, M. T., Kanai, R., Seth, A. K., & VanRullen, R.
(2016).
Rhythmic influence of top-down perceptual priors in the phase
of prestimulus occipital alpha oscillations. Zeitschrift für Kognition
Neurowissenschaften, 28(9), 1318–1330.
Simony, E., Honey, C. J., Chen, J., Lositsky, O., Yeshurun, Y., Wiesel,
A., & Hasson, U. (2016). Dynamic reconfiguration of the default
mode network during narrative comprehension. Nature Commu-
nications, 7.
Smolensky, P.
Information processing in dynamical
Systeme: Foundations of harmony theory (No CU-CS-321-86).
Colorado University at Boulder, Department of Computer
Wissenschaft.
(1986).
Spreng, R. N., Mar, R. A., & Kim, A. S. (2009). The common neural
basis of autobiographical memory, prospection, navigation, Die-
ory of mind, and the default mode: A quantitative meta-analysis.
Zeitschrift für kognitive Neurowissenschaften, 21(3), 489–510.
Staresina, B. P., Bergmann, T. O., Bonnefond, M., van der Meij, R.,
Jensen, O., Deuker, L., . . . Fell J. (2015). Hierarchical nesting of
slow oscillations, spindles and ripples in the human hippocam-
pus during sleep. Naturneurowissenschaften, 18, 1679–1686.
Stickgold, Roberto.
(2013). Parsing the role of sleep in memory
Verarbeitung. Aktuelle Meinung in der Neurobiologie, 23(5), 847–853.
Sugihara, H., Chen, N., & Sur, M. (2016). Cell-specific modulation
of plasticity and cortical state by cholinergic inputs to the visual
Kortex. Journal of Physiology (Paris), 110(1), 37–43.
Sutton, R. S., & Barto, A. G.
(1998). Reinforcement learning: Ein
introduction (Bd. 1, NEIN. 1). Cambridge, MA: MIT Press.
Netzwerkneurowissenschaften
355
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Learning by switching internal and external modes
Tambini, A., Ketz, N., & Davachi, L. (2010). Enhanced brain corre-
lations during rest are related to memory for recent experiences.
Neuron, 65(2), 280–290.
Tamminen, J., Lambon Ralph, M. A., & Lewis, P. A.
(2017). Tar-
geted memory reactivation of newly learned words during sleep
triggers REM-mediated integration of new memories and existing
Wissen. Neurobiology of Learning and Memory, 137, 77–82.
Van Kerkoerle, T., Self, M. W., Dagnino, B., Gariel-Mathis, M. A.,
Poort, J., Van Der Togt, C., & Roelfsema, P. R.
(2014). Alpha
and gamma oscillations characterize feedback and feedforward
processing in monkey visual cortex. Verfahren des Nationalen
Akademie der Wissenschaften, 111(40), 14332–14341.
van Strien, N. M., Cappaert, N. L. M., & Witter, M. P.
(2009). The anatomy of memory: An interactive overview of
the parahippocampal-hippocampal network. Nature Reviews
Neurowissenschaften, 10(4), 272–282.
Vandecasteele, M., Varga, V., Berényi, A., Papp, E., Barthó, P.,
Venance, L., . . . Buzsáki, G.
(2014). Optogenetic activation
of septal cholinergic neurons suppresses sharp wave ripples and
enhances theta oscillations in the hippocampus. Proceedings of
the National Academy of Sciences, 111(37), 13535–13540.
VanRullen, R.
(2016). Perceptual cycles. Trends im kognitiven Bereich
Wissenschaften, 20(10), 723–735.
Warburton, D. M., Wesnes, K., Edwards, J., & Larrad, D.
(1985).
Scopolamine and the sensory conditioning of hallucinations.
Neuropsychobiology, 14(4), 198–202.
Werfel,
J., Xie, X., & Seung, H. S.
Learning curves
for stochastic gradient descent in linear feedforward networks.
Neural Computation, 17(12), 2699–2718.
(2005).
Widrow, B., & Hoff, M. E.
(1960). Adaptive switching circuits.
In IRE WESCON Convention Record (Bd. 4, NEIN. 1, S. 96–104).
Wiskott, L., & Sejnowski, T. J. (2002). Slow feature analysis: Un-
supervised learning of invariances. Neural Computation, 14(4),
715–770.
Xie, X., & Seung, H. S.
(2003). Equivalence of backpropagation
and contrastive Hebbian learning in a layered network. Neuronal
Computation, 15(2), 441–454.
Yang, T., & Maunsell, J. H. (2004). The effect of perceptual learn-
ing on neuronal responses in monkey visual area V4. Zeitschrift für
Neurowissenschaften, 24(7), 1617–1626.
Yu, A. J., & Dayan, P. (2005). Uncertainty, neuromodulation, Und
attention. Neuron, 46(4), 681–692.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
4
3
3
9
1
0
9
1
8
9
9
N
e
N
_
A
_
0
0
0
2
4
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Netzwerkneurowissenschaften
356