PERSPECTIVE
Cognitive genomics: Linking genes to
behavior in the human brain
Genevieve Konopka
Department of Neuroscience, UT Southwestern Medical Center, Dallas, TX 75390-9111, USA
ABSTRAKT
Correlations of genetic variation in DNA with functional brain activity have already provided
a starting point for delving into human cognitive mechanisms. Jedoch, these analyses do
not provide the specific genes driving the associations, which are complicated by intergenic
localization as well as tissue-specific epigenetics and expression. The use of brain-derived
expression datasets could build upon the foundation of these initial genetic insights and yield
genes and molecular pathways for testing new hypotheses regarding the molecular bases of
human brain development, Erkenntnis, und Krankheit. Daher, coupling these human brain gene
expression data with measurements of brain activity may provide genes with critical roles in
brain function. Jedoch, these brain gene expression datasets have their own set of caveats,
most notably a reliance on postmortem tissue. In this perspective, I summarize and examine
the progress that has been made in this realm to date, and discuss the various frontiers
remaining, such as the inclusion of cell-type-specific information, additional physiological
measurements, and genomic data from patient cohorts.
Progress in understanding the inner workings of the brain has come a long way from the preneu-
roscience era of phrenology, when we were limited to conjectures about human behavior
based on the shape of the skull. Over the past quarter century, technological breakthroughs
have given us the ability to noninvasively peer into the operations of the human brain during
behavior, by means of a host of imaging and physiological techniques. Funktionelle Bildgebung
has provided elegant maps of human activity at rest, as well as during any number of cogni-
tive tasks. By coupling these results with neuroanatomical and structural imaging, function and
structure can be married to identify brain regions that work in concert to execute specific func-
tionen. Außerdem, when such approaches are carried out in patients with neuropsychiatric
disorders, the regional brain activity relevant to cognitive phenotypes can be uncovered.
Genetic Contributions to Cognition
Determining the relative contribution of genes to cognition has been a longstanding interest
in the field of genetic research. Recent inquiries have focused on unlocking the genetic and
molecular mechanisms underlying human brain activity (see the discussion and references
in Medland, Jahanshad, Neale, & Thompson, 2014, and Thompson, Ge, Glahn, Jahanshad,
& Nichols, 2013). Key insights have been made, such as the heritability of functional brain
Netzwerke (Fornito et al., 2011; Fu et al., 2015; Glahn et al., 2010; Yang et al., 2016) und das
correlation of genetic variation in altered functional connectivity in specific diseases
or phenotypes (see the references in Gaiteri, Mostafavi, Honey, De Jager, & Bennett, 2016;
Hernandez, Rudie, Grün, Bookheimer, & Dapretto, 2015). Als solche, these advances could
have profound implications for how we diagnose and treat such disorders (see the discus-
sion and references in Matthews & Hampshire, 2016). Außerdem, genome-wide association
Keine offenen Zugänge
Tagebuch
Zitat: Konopka G. (2017). Kognitiv
genomics: Linking genes to behavior
in the human brain. Netzwerk
Neurowissenschaften, 1(1), 3–13.
doi:10.1162/netn_a_00003
DOI:
http://doi.org/10.1162/netn_a_00003
zusätzliche Informationen:
Erhalten: 16 September 2016
Akzeptiert: 22 November 2016
Konkurrierende Interessen: The author has
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Genevieve Konopka
Genevieve.Konopka@utsouthwestern.edu
Handling-Editor:
Olaf Sporns
Urheberrechte ©: © 2017
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Microarrays:
Technology that uses preselected
oligonucleotides to quantify RNA
amounts on a genome-wide basis.
(See Box 1.)
In situ hybridization:
Probe-based technology used to
assess the spatial expression of
specific RNA molecules (See Box 1.)
RNA sequencing (RNA-seq):
Unbiased quantification of RNA
molecules on a genome-wide basis.
(See Box 1.)
studies have identified specific genomic loci that are significantly associated with subcorti-
cal brain structures (Hibar et al., 2015); with educational attainment as a proxy for cognition
in general
(Okbay, Beauchamp, et al., 2016); with personality traits such as subjective
well-being, depressive symptoms, and neuroticism (Okbay, Baselmans, et al., 2016); and with
cognitive disorders such as schizophrenia (Schizophrenia Working Group of the Psychiatric
Genomics Consortium, 2014). These molecular and genetic insights provide a baseline for
ultimately pinpointing drug targets in a number of cognitive disorders, as well as deepening
our understanding of both the developmental and evolutionary origins of human cognition.
Daher, further investigations into the molecular mechanisms underlying human brain activity
are needed to bridge the gap between genes and behavior.
Quantifying Gene Expression in the Human Brain
The genome revolution, followed rapidly by implementation of the high-throughput techno-
logies of microarrays and next-generation sequencing, has permitted investigations of human
brain gene expression in a spatiotemporal manner, by quantifying RNA amounts at a genome-
wide level (z.B., Kang et al., 2011). The analysis of gene transcription across the entire human
brain allows for distinguishing the genes expressed in specific brain regions during a given
developmental time period, and thus results in a quantitative measurement of gene expres-
sion levels. These datasets are different from the genetic associations mentioned above, In
which changes at the DNA level are identified. Such genetic variation might be within re-
gions of DNA of unknown functional significance (z.B., do the variants affect gene expres-
sion?) and might also interact with unknown epigenetic markers in a tissue-specific manner,
leading to further ambiguity about the resultant gene expression. Surveying the vast transcrip-
tional landscape of the developing and adult human brain has been facilitated by the work
of the Allen Institute for Brain Science in collaboration with a number of academic groups,
to develop several reference gene expression atlases of the human brain, by using a combi-
nation of in situ hybridization, microarrays, and RNA sequencing (RNA-seq) throughout the
human lifespan (Hawrylycz et al., 2015; Hawrylycz et al., 2012; Miller et al., 2014; Zeng et al.,
2012; see Box 1). One of the caveats to these assessments of human brain gene ex-
pression is that they are naturally limited to postmortem tissue. Although careful statistical
analyses take into consideration experimental covariates such as postmortem interval and RNA
Qualität,
there is always the possibility that patterns of gene expression in behaving
individuals cannot be fully recapitulated in postmortem tissue. Trotzdem, these assess-
ments provide critical insights into human brain gene expression patterns, based on develop-
mental stage (Kang et al., 2011; Miller et al., 2014), Geschlecht (Kang et al., 2011), hemispheric
lateralization (or lack thereof; Hawrylycz et al., 2012; Johnson et al., 2009; Pletikos et al.,
2014), and human-specific evolution (Bakken et al., 2016; Bernard et al., 2012). These obser-
vations can then be compared with disease-relevant datasets. Zum Beispiel, genetic data from
autism spectrum disorder (ASD) patients were integrated with the BrainSpan gene expression
dataset (www.brainspan.org) to identify ASD-relevant coexpression networks (Parikshak et al.,
2013; Willsey et al., 2013). Außerdem, genomic profiling of disease tissue itself can be
insightful, as has been the case for ASD, where such profiling has identified differentially ex-
pressed networks of mRNAs and microRNAs in ASD brains as compared to matched con-
trols (Voineagu et al., 2011; Y. E. Wu, Parikshak, Belgard, & Geschwind, 2016). Zusammen, diese
studies of human brain gene expression have facilitated the prioritization of specific genes and
molecular pathways for further in-depth analyses. Jedoch, such follow-up studies are largely
limited to animal models, and there appears to be a large divide between what can be observed
at the gene expression level in brain tissue and at the behavioral level in humans.
Netzwerkneurowissenschaften
4
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Kasten 1. Descriptions of Gene Expression Detection Methods
In situ hybridization is carried out by hybridizing gene-specific RNA probes to tissue samples.
This method provides spatial resolution of the mRNA expression of individual genes. Hoch-
quality tissue specimens and highly specific probes to each gene are required for accurate
detection. A major advantage of in situ hybridization is the ability to couple it with immuno-
histochemistry to make mRNA and protein correlations. Jedoch, the quantification of in
situ hybridization is challenging, depending on the method used. The greatest advantages of
in situ hybridization over the other technologies discussed below are its spatial resolution and
ability to detect expression from small amounts of tissue.
Microarrays for gene expression rely on hybridization of an RNA sample to a chip spotted
with predetermined oligonucleotide probes. Although recent microarrays can provide full
genome coverage and can detect, Zum Beispiel, small RNAs, the technology is still not com-
pletely unbiased. For one thing, novel gene transcripts and unannotated regions of a genome
will not be detected or quantified using microarrays. Darüber hinaus, microarrays are less useful
for querying expression in species for which specific microarrays are not available, nor will
microarrays provide cellular-resolution expression information. Jedoch, well-established
algorithms exist for analyzing microarray data, and their results are highly consistent when
comparing many samples on the same type of microarray. The processing and analysis of
microarray data is also less costly in terms of time, money, and computational power needed.
RNA sequencing, or RNA-seq, uses next-generation sequencing technology to quantify the
expression of all expressed genes in an RNA sample in an unbiased manner, without any
a priori information about the sample. Expression information is limited by how the RNA sam-
ple is processed—that is, is all of the RNA being processed, or are polyadenylated transcripts
selected? Expression information can also be limited by the length of the sequencing read
(z.B., 50 vs. 150 base pairs, or single- vs. paired-end reads) and the amount of sequencing
depth carried out. Such parameters can limit the ability to make informed analysis of features
such as the expression of noncoding RNAs or RNA splicing. Zusätzlich, there is no agreed-
upon method for analyzing RNA-seq data. Jedoch, all of these analyses and more can be
carried out, given the appropriate sample preparation, sequencing method, and analytical
pipeline. RNA-seq alone will not provide cellular-level resolution unless it is coupled with
other technologies, such as microfluidics or flow cytometry (z.B., single-cell RNA-seq). Der
advantages of RNA-seq include the unbiased detection of expression, which makes the tech-
nology compatible with any species of interest and any type of RNA, and a larger dynamic
range of detection than with microarrays (d.h., it is easier to detect low-abundance expression).
Correlations of Human Brain Gene Expression With Functional Imaging Data
Bridging the divide between genes and behavior in human brains has entered a new chap-
ter with the first studies to compare brain gene expression and functional imaging datasets from
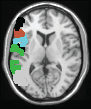
humans (Feige. 1)
(Hawrylycz et al., 2015; Krienen, Yeo, Ge, Buckner, & Sherwood, 2016;
Richiardi et al., 2015; G. Z. Wang et al., 2015). These studies utilized functional magnetic
resonance imaging (fMRT) data obtained during the resting state. It remains to be determined
whether gene expression data from postmortem tissues are more related to brain activity dur-
ing the resting state or better reflect an “acute” active state resulting from a specific function
or task. Since detailed behavioral data from brain donors are unavailable, resting-state cor-
relations are a reasonable starting point. Zusätzlich, transcriptional responses to stimuli are
on the order of minutes, whereas postmortem times are typically on the order of hours, wieder
supporting relevance to the resting state. Endlich, all of these studies focused their comparisons
on the neocortex; one reason for this is the relative similarity in gene expression across cortical
Functional magnetic resonance
Bildgebung (fMRT):
Noninvasive method for measuring
brain activity, based on blood
oxygenation levels that provide
spatial and temporal resolution on
the order of millimeters and seconds,
jeweils.
Netzwerkneurowissenschaften
5
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Correlated Genes
Brain Activity
Fractional amplitude of low frequency fluctuations (fALFF) aus 282
individuals from 2 separate sources (Wang et al. 2015)
Functional connectivity from 274 individuals from 2 separate sources
(Richiardi et al. 2015)
Functional connectivity from 447 individuals (Hawrylycz et al. 2015)
Functional connectivity from 1000 individuals (Krienen et al. 2015)
Brain Gene Expression
Genome-wide RNA-seq & microarray data from
11 cortical areas in 40 individuals from 3 separate sources
(Wang et al. 2015)
Genome-wide microarray data from
88 (Richiardi et al. 2015) oder
52 cortical areas (Hawrylycz et al. 2015) oder
microarray expression of 19 genes in the cortex (Krienen et al. 2016)
In 6 individuals from the Allen Institute for Brain Science database
Figur 1.
Integrating functional brain activity and gene expression in the human brain has re-
sulted in the identification of a small group of genes that are likely important for resting-state
functional networks.
Bereiche, relative to the large gene expression differences when comparing cortical to noncortical
Bereiche (Hawrylycz et al., 2015), and the other reason is the availability of independent cortical
expression datasets.
In the study by Richiardi et al. (2015), four well-characterized functional networks from
15 subjects were compared to microarray data from high-resolution gene expression profiling
of six postmortem brains that was carried out by the Allen Brain Institute. This comparison
identified ~78 genes significantly correlated with the functional networks. Common polymor-
phisms in these genes were further found to correlate with functional networks in the larger
(~259 subjects) IMAGEN cohort, for whom both single-nucleotide polymorphism and fMRI
data were available (Schumann et al., 2010). Endlich, Richiardi et al. further validated the gene
list in mouse orthologs, showing that the expression of these genes correlated with mouse
brain connectivity. The correlated gene list was enriched for neuron-specific genes and for
genes encoding ion channels, as well as for genes associated with Alzheimer’s disease and
schizophrenia. These results were consistent with a study by Hawrylycz et al. (2015) that used
the same gene expression dataset, along with independent functional connectivity MRI data
aus 447 subjects that are part of the Human Connectome Project (Van Essen et al., 2013).
In the Hawrylycz et al. (2015) Studie, the authors set out to determine the genes with the most
consistent pattern of gene expression across human brains. Using the genes with the most
consistent differential expression across the cortex, they found that the same genes identified
in Richiardi et al. (2015) were among the genes with the greatest correlations of expression
with functional connectivity.
In G. Z. Wang et al. (2015), RNA-seq and microarray data from three independent datasets
wurden benutzt, containing samples from approximately 40 individuals. Each of these three gene
expression studies was compared to two independent fMRI datasets, mit 84 Und 198 Fächer,
and the consensus of the correlations within the default-mode network was reported. In diesem
Studie, a limited number of cortical areas were included (five to ten), due to sample availabil-
ität. Trotzdem, 38 genes were identified that correlated significantly with the default-mode
Netzwerkneurowissenschaften
6
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Netzwerk. These genes were also enriched for neuron-specific genes, genes encoding ion chan-
nels, and genes implicated in ASD. Insbesondere, genes relevant to interneuron identity were
suggested to play a potentially key role in orchestrating the brain activity assessed by fMRI.
One of the challenges of interpreting the results from these first studies to correlate human-
brain gene expression and fMRI signals is that the correlations across regions might be biased
toward differences in cell-type proportions in these regions. Somit, the identified enrichment
of neuron-specific genes could be an artifact of the relative variation in neuronal proportions in
these cortical regions. This is further exemplified by the finding that human brain gene expres-
sion is most conserved with respect to mouse brain gene expression among neuron-relevant
genes (Hawrylycz et al., 2015), and the correlations between gene expression and functional
connectivity are conserved between human and mouse brains (Richiardi et al., 2015). The dis-
tinctions in areal and cell-type-specific expression patterns driving the correlations with activity
should be addressed with improved technology for single-cell expression profiling and higher-
resolution imaging, as will be discussed below. One additional challenge in interpreting these
findings is that there is no manner in which to assign causation to the correlations. In other
Wörter, there is no way to determine whether the gene expression patterns are a result of brain
Aktivität, or whether the brain activity is somehow driven by the gene expression patterns. Eins
way to assess this would be to measure gene expression in vivo before, während, and after a task,
something that is clearly not possible in humans. Jedoch, there has been remarkable progress
in following the translation of single mRNAs in live cells, including neurons, as well as in liv-
ing animals such as Drosophila (Halstead et al., 2015; C. Wang, Han, Zhou, & Zhuang, 2016;
B. Wu, Eliscovich, Yoon, & Singer, 2016; Yan, Hoek, Vale, & Tanenbaum, 2016). daher,
such technology might one day be available for the study of humans.
Because the gene expression data discussed here derive from postmortem sources, diese
studies require the use of separate cohorts from MRI studies for comparison. Mit anderen Worten,
the gene expression data were not derived from tissue from the subjects who underwent fMRI.
Zusätzlich, there was no overlap between the imaging and gene expression cohorts across
the Richiardi et al. (2015) and the G. Z. Wang et al. (2015) Studien, nor were the methods or
networks analyzed in the same way. Zum Beispiel, Richiardi et al. examined correlations across
brain regions in four networks (dorsal default-mode, salience, sensorimotor, and visuospatial),
whereas G. Z. Wang et al. focused on the default-mode network and an indicator of activ-
ität, fractional amplitude of low-frequency fluctuations (fALFF), from within each individual
region, followed by correlations across the regions. These differences make it all the more
remarkable that the results displayed statistically significant overlap in the genes identified
(G. Z. Wang et al., 2015). Zum Beispiel, NECAB2, NEFH, SCN1B, and SYT2 were identified in
both the Richiardi et al. and G. Z. Wang et al. Studien. Such overlaps suggest that even when
using different cohorts and different methods, the expression of genes underlying brain activity
at rest can be identified. Perhaps even more remarkable were the results of a subsequent study
by Krienen et al. (2016) that focused on 19 genes enriched in human supragranular layers and
that identified specific corticocortical connectivity networks correlated with these genes using
functional connectivity MRI measurements. Both NEFH and SYT2 are among these 19 genes.
Although the human gene expression data in the Krienen et al. study are the same as in the
Richiardi et al. and Hawrylycz et al. Studien, the imaging dataset was derived from an inde-
pendent cohort of 1,000 subjects from the Brain Genomics Superstruct Project (Holmes et al.,
2015). Daher, these overlaps in genes suggest intriguing hypotheses to test regarding the roles of
these specific genes in directing human-specific corticocortical connections. Zum Beispiel, Sind
there genetic polymorphisms or differential expression patterns of these two genes in patients
with either abnormal connectivity patterns, as assessed by imaging, or abnormal cognitive
Fractional amplitude of
low-frequency fluctuations (fALFF):
Measurement of brain activity from
fMRI datasets that compares signals
derived from low frequencies
(0.01–0.08 Hz) to all frequency
Signale.
Netzwerkneurowissenschaften
7
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
phenotypes or disorders? There are already examples of altered NEFH levels in brain tis-
sue from individuals with schizophrenia (Pinacho et al., 2016) or alcoholism (Iwamoto et al.,
2004); altered levels of SYT2 mRNA in schizophrenia (McMeekin et al., 2016); and associa-
tion of SYT2 with both attention-deficit/hyperactivity disorder (Sánchez-Mora et al., 2013) Und
cocaine dependence (Fernàndez-Castillo et al., 2012). Whether these changes and variants
can be linked to neuroanatomical alterations remains to be determined.
Zusammen, these studies build upon previous work that has supported a genetic basis for
functional connectivity and brain activity, showing evidence for mRNA expression correla-
tions with these measurements. The genes identified are synaptic genes enriched in neurons;
Jedoch, it is likely that this list of genes is not exhaustive, and as further detailed genomic
profiling is carried out, additional refined lists will be uncovered. The differences among the
genes identified raise important considerations. G. Z. Wang et al. (2015) only used data from
male subjects, due to sample availability, in one of the gene expression studies. It is unlikely
that gene list differences arose from such sample constraints, but as greater numbers of corti-
cal RNA-seq datasets become available, those differences can be tested. Zusätzlich, as was
previously mentioned, G. Z. Wang et al. used a region-of-interest approach to calculate ac-
tivity, while the other studies employed functional connectivity. Cognition can be studied at a
network level, and these networks span the gamut from those quantitated with fMRI, down to
those assessed by RNA sequencing (Petersen & Spurns, 2015). Jedoch, before these initial
studies using human datasets were carried out, it was not known whether gene expression pat-
terns would fall in line with the networks observed using functional connectivity approaches.
The gene expression data are processed from each region independently of the others, Und
the data can be collated from different individuals within the same brain regions with mini-
mal variation (Hawrylycz et al., 2012). Zusätzlich, the inability to directly manipulate gene
expression or brain regional activity in humans makes testing the contribution of the observed
gene list to cognitive function challenging. Zum Beispiel, as was already mentioned, there is
no way to test causation in humans: das ist, whether the level of expression of a particular
gene might result in altered brain activity, or whether altered brain activity results in a change
in gene expression. Jedoch, such experiments could be carried out in animal models if the
same correlations between expression and activity were valid.
Comparisons With Nonhuman Brain Expression and Activity
Wie, Dann, do these results in humans compare to what was already known in nonhu-
man brains? Such comparison would open up the possibility of translating the observations
in humans into model systems.
Initial work in worms provided direct evidence for gene
expression signatures indicating neuronal connectivity (Kaufman, Dror, Meilijson, & Ruppin,
2006). Subsequent studies in rodents have supported the idea of brain gene expression
correlating with structural connectivity (French & Pavlidis, 2011; Ji, Fakhry, & Deng, 2014;
Wolf, Goldberg, Manor, Sharan, & Ruppin, 2011).
Interessant, the gene ontology of these
correlated genes suggests a role for neuronal projection and guidance molecules in connec-
tivity, as might be predicted. In another study, the genes correlated with network hub connec-
tivity were identified as those involved in energy metabolism (z.B., oxidative synthesis and ATP
metabolism; Fulcher & Fornito, 2016), highlighting genes of interest for studies of brain disor-
ders featuring both metabolic deficiencies and abnormal connectivity, such as Alzheimer’s
Krankheit. Zusätzlich, a significant number of ASD-relevant genes were among the structurally
correlated genes (French & Pavlidis, 2011) as in the findings of G. Z. Wang et al. (2015). More-
über, follow-up work using human samples suggested that not only are there correlations be-
tween gene expression and structural connectivity, but that gene expression might be one of
Netzwerkneurowissenschaften
8
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Transcriptome:
The entire set of transcripts or
messenger RNAs from a particular
tissue and/or organism.
the driving forces behind the observed connectivity (Goel, Kuceyeski, LoCastro, & Raj, 2014),
and therefore a more causative correlation between these features might be discernible.
From a global gene expression perspective, the recent detailed profiling of the human brain
transcriptome has allowed for extensive comparisons with both the mouse and rhesus macaque
brain transcriptomes (Bakken et al., 2016; Hawrylycz et al., 2012). These comparisons have
uncovered highly correlated expression in similar regions of the human and mouse brains, Par-
ticularly when the genes were more related to neuronal expression patterns (Hawrylycz et al.,
2015). This is in line with data from Richiardi et al. (2015) that demonstrated similar correla-
tions of mouse brain gene expression and mouse brain connectivity when the same gene list
identified in the human brain was applied to the mouse. Of note, some genes vary in their
brain expression patterns between species (Hawrylycz et al., 2015), and it would be interesting
to examine whether these particular genes demonstrate differential correlations with human
brain activity measurements in patient populations with cognitive disorders. Such human-
relevant expression differences become even more notable when rhesus macaque is included
in the comparisons, since this allows for comparison with a more closely related primate
Spezies, rather than the mouse. The study by Bakken et al. (2016) compared human, rhesus
macaque, rat, and mouse brain expression, including developmental expression data as well
as some cortical-layer data. While the authors confirmed and further delineated genes with
conserved patterns of expression in mammalian brain, they also identified a subset of genes
that demonstrate human-specific expression patterns across cortical development. Wieder, ex-
amining whether the expression pattern of these genes over development show differential
correlations with developmental functional brain activity could be relevant to understanding
cognitive disorders. Zusammen, these studies suggest that conserved gene expression across
brain regions underlies many features of mammalian brain structure and activity, and they also
prioritize specific genes for detailed study in animal disease models.
Future Directions
In the last few years, there has been significant progress into elucidating the molecular mech-
anisms of human brain functional networks. We know that there is a genetic, heritable
component to these brain networks, as well as patterns of gene expression that may direct
ihnen. We have also learned that these genes and their expression may be tied to the human
brain’s evolution and cognitive disorders. Was, Dann, are the next frontiers for delving deeper
into these mechanisms?
In den vergangenen Jahren, efforts have been made to standardize experimental approaches in order
to permit the collation and comparison of datasets across studies. This has been particu-
larly noteworthy among both imaging and genetic consortia. Kürzlich, these collaborative
efforts on the imaging front have expanded to include imaging of specific cognitive disorders.
Zum Beispiel, the ABIDE consortium has provided novel insights into ASD (Di Martino et al.,
2014), and the IMAGEN Consortium (Schumann et al., 2010) has combined genetics and
imaging to study a variety of phenotypes relevant to psychiatric disorders. The Human Connec-
tome Project (Glasser, Coalson, et al., 2016; Glasser, Schmied, et al., 2016) has given these and
all other imaging studies a new baseline from which to derive deviations from the norm. In
the field of neurogenetics, this collaborative approach has also been critical for tackling as-
sessments of genetic associations with disease. Recent large-scale efforts to examine schizo-
phrenia (Schizophrenia Working Group of the Psychiatric Genomics, 2014) and depression
(CONVERGE Consortium, 2015) have required the inclusion of
thousands of
samples just to scratch the surface of the common genetic features of these disorders. Der
tens of
Netzwerkneurowissenschaften
9
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Endophenotype:
Phenotype(S) related to a specific
disorder that are quantifiable and
have a genetic component.
Electroencephalogram (EEG):
Noninvasive method for measuring
brain activity, on the basis of
electrical activity assessed with
electrodes placed on the scalp.
deposition of raw gene expression data from human brain tissue into repositories such
as NCBI GEO, alongside the efforts of
the Allen Brain Institute (http://human.brain-
map.org (Hawrylycz et al., 2015; Hawrylycz et al., 2012) and the BrainSpan Consortium
(www.brainspan.org), has further permitted detailed analyses and hypothesis testing by many
research groups. The fruits of all of these efforts can now be combined in a number of ways to
ask whether the correlations between gene expression and brain activity differ across multiple
Parameter, such as endophenotypes, medication use and response, or other comorbidities
when queried in patient populations. The abundance of genetic information from the increas-
ing number of genome-wide association studies and large-scale whole-genome sequencing
efforts can also be layered into these analyses, when expression data are lacking from a partic-
ular patient group, to make predictive correlations regarding whether a particular gene might
be expected to have altered expression in a particular disorder.
Natürlich, an ideal scenario would take a longitudinal approach in which populations
of patients and unaffected controls could be genotyped, phenotyped, imaged, und folgte
throughout the lifespan, culminating in tissue donation (sehen, z.B., the MyConnectome project:
Poldrack et al., 2015). Such longitudinal within-subjects analyses could last longer than the
average research career of individuals, but ultimately they would be quite informative for our
understanding of cognitive disorders. Although transcriptional data would necessarily be col-
lected at only one time point, the aggregation of data across many individuals might permit
inferences about how gene expression is relevant to specific phenotypes. Delimited within-
subjects approaches could also potentially be used with surgical patients. Zum Beispiel, surgi-
cal patients for drug-resistant epilepsy not only have the epileptic foci removed during surgery,
but they might also require removal of the adjacent unaffected tissue to obtain access to the
foci (Spencer, Spencer, Mattson, Williamson, & Novelly, 1984). Such patients who undergo
preoperative imaging studies could be included in a within-subjects study to correlate brain
activity or structure from the imaging study with gene expression profiling from the surgical
resections. Zusätzlich, other measurements of brain activity in these patients, such as intracra-
nial electroencephalography or single-unit recordings, could be coupled with measurements
of gene expression obtained from the surgically resected tissue.
Endlich, one of the most exciting advances on the gene expression front has been the adapta-
tion of fluidics and improved next-generation sequencing library preparations to permit the as-
sessment of genome-wide gene expression in single cells or single nuclei, with minimal signal
amplification biases. These approaches have been applied to both surgical and postmortem
brain tissue from humans (Darmanis et al., 2015; Johnson et al., 2015; Krishnaswami et al.,
2016; Lake et al., 2016; Pollen et al., 2015). Such studies are revising the definitions of cell
types in the brain, and their data have important downstream implications for how researchers
can interpret the phenotypic outcomes of genetic variants in patients or animal models. Während
it is unlikely that genomic profiling of every cell in a human brain will be carried out in the
near future, discrete assessment of particular brain regions relevant to a particular disorder,
or samples derived from surgical patients who had undergone functional or physiological
assessment, should be particularly revealing. Zum Beispiel, the study by Lake et al. (2016),
in which they profiled over 3,000 nuclei across six cortical areas from a postmortem brain,
provided evidence to support the idea of brain region and local heterogeneity among neu-
ronal subtypes, thus further emphasizing the need to explore these relationships at the cellular
Ebene. The incorporation of improved parcellation maps of the human brain (Ding et al., 2016;
Glasser, Coalson, et al., 2016) should also further facilitate integrating measurements of gene
expression that take into account regional heterogeneity with functional measurements. Für
Beispiel, these new maps could direct refinements of postmortem tissue dissections for gene
Netzwerkneurowissenschaften
10
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
expression studies. It is not yet clear how to ideally integrate cellular-resolution gene expres-
sion data with similar resolutions of brain activity, should such levels be achievable in humans.
Zum Beispiel, network approaches with cellular resolution that work in genetically modified
animals or with viral injections (z.B., calcium sensors such as GCaMP) await future advances
in imaging for a less-invasive approach in humans. The advent of noninvasive approaches, Also-
called “molecular fMRI,” in nonhuman systems may eventually translate into human studies
(Bartelle, Barandov, & Jasanoff, 2016). This approach would utilize a probe (z.B., a reporter
with a neuron-specific driver introduced virally via injection) that can provide a quantitative
readout to detect changes in gene expression at a cellular level when it is coupled with MRI.
Jedoch, in the meantime, one could imagine that less high-throughput comparisons using
electrophysiological methods in cultured human neurons or brain organoids could still be
illuminating. Because strong correlations with direct physiological stimulations and blood-
oxygen-level dependent signals, and between single-cell transcriptomics and electrophysiol-
Ogy, have been achieved in rodents (Lee et al., 2016; Tasic et al., 2016), it is possible that a
connection between fMRI, molecular or physiological stimulation in vivo, and gene expression
could be made in humans.
While connecting genes to behavior is an ultimate goal for many neuroscientists, the many
layers of regulation intervening between DNA, the ensemble of cellular activities, and the mul-
titude of circuit and network combinations make deciphering these connections challenging.
Darüber hinaus, these relationships are further complicated by cell-specific epigenetics and gene ex-
pression, and by a greater appreciation of somatic mutations during human brain development
(Lodato et al., 2015). Trotzdem, as has been evidenced by the few studies discussed here
in detail, major inroads are being made by linking mRNA levels in the brain to fMRI studies
beim Menschen. Clearly, more has yet to be accomplished. Jedoch, these first steps have opened
up the possibility of combining diverse human brain datasets to achieve new insights into the
molecular mechanisms of cognitive functions.
ACKNOWLEDGMENTS
I thank Maria Chahrour and the members of the Konopka lab for comments on the manuscript.
The research in the Konopka lab is supported by grants from NIMH, NIDCD, the NSF, the U.T.
BRAIN initiative, the Simons Foundation, and the James S. McDonnell Foundation.
VERWEISE
Bakken, T. E., Müller, J. A., Ding, S. L., Sunkin, S. M., Schmied, K. A.,
Ng, L., . . . Lein, E. S. (2016). A comprehensive transcriptional
map of primate brain development. Natur, 535, 367–375. doi:
10.1038/nature18637
Bartelle, B. B., Barandov, A., & Jasanoff, A. (2016). Molecular fMRI.
Zeitschrift für Neurowissenschaften, 36, 4139–4148.
Bernard, A., Lubbers, L. S., Tanis, K. Q., Luo, R., Podtelezhnikov,
A. A., Finney, E. M., . . . Lein, E. S. (2012). Transcriptional
architecture of the primate neocortex. Neuron, 73, 1083–1099.
doi:10.1016/j.neuron.2012.03.002
CONVERGE Consortium. (2015). Sparse whole-genome sequenc-
ing identifies two loci for major depressive disorder. Natur,
523, 588–591.
Darmanis, S., Sloan, S. A., Zhang, Y., Enge, M., Caneda, C., Shuer,
L. M., . . . Quake, S. R. (2015). A survey of human brain transcrip-
tome diversity at the single cell level. Verfahren des Nationalen
Akademie der Wissenschaften, 112, 7285–7290.
Di Martino, A., Yan, C. G., Li, Q., Denio, E., Castellanos, F. X.,
Alaerts, K., . . . Milham, M. P. (2014). The autism brain imaging
data exchange: Towards a large-scale evaluation of the intrinsic
brain architecture in autism. Molekulare Psychiatrie, 19, 659–667.
doi:10.1038/mp.2013.78
Ding, S. L., Royall, J. J., Sunkin, S. M., Ng, L., Facer, B. A., Lesnar,
P., . . . Lein, E. S. (2016). Comprehensive cellular-resolution atlas
of the adult human brain. Zeitschrift für Vergleichende Neurologie,
524, 3127–3481. doi:10.1002/cne.24080
Fernàndez-Castillo, N., Cormand, B., Roncero, C., Sánchez-Mora,
C., Grau-Lopez, L., Gonzalvo, B., . . . Ribasés, M. (2012). Can-
didate pathway association study in cocaine dependence: Der
control of neurotransmitter release. World Journal of Biological
Psychiatrie, 13, 126–134. doi:10.3109/15622975.2010.551406
Netzwerkneurowissenschaften
11
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Fornito, A., Zalesky, A., Bassett, D. S., Meunier, D., Ellison-Wright,
ICH., Yücel, M., . . . Bullmore, E. T. (2011). Genetic influences
on cost-efficient organization of human cortical functional net-
funktioniert. Zeitschrift für Neurowissenschaften, 31, 3261–3270. doi:10.1523/
JNEUROSCI.4858-10.2011
French, L., & Pavlidis, P. (2011). Relationships between gene expres-
sion and brain wiring in the adult rodent brain. PLoS Computa-
tional Biology, 7, e1001049.
Fu, Y., Ma, Z., Hamilton, C., Liang, Z., Hou, X., Ma, X., . . . Zhang,
N. (2015). Genetic influences on resting-state functional net-
funktioniert: A twin study. Kartierung des menschlichen Gehirns, 36, 3959–3972.
doi:10.1002/hbm.22890
Fulcher, B. D., & Fornito, A. (2016). A transcriptional signature of
hub connectivity in the mouse connectome. Verfahren der
Nationale Akademie der Wissenschaften, 113, 1435–1440.
Gaiteri, C., Mostafavi, S., Honey, C. J., De Jager, P. L., & Bennett,
D. A. (2016). Genetic variants in Alzheimer disease: Molekular
and brain network approaches. Nature Reviews Neurology, 12,
413–427.
Glahn, D. C., Winkler, A. M., Kochunov, P., Almasy, L., Duggirala,
R., Carless, M. A., . . . Blangero, J. (2010). Genetic control over
the resting brain. Proceedings of the National Academy of Sci-
zen, 107, 1223–1228. doi:10.1073/pnas.0909969107
Glasser, M. F., Coalson, T. S., Robinson, E. C., Hacker, C. D.,
Harwell, J., Yacoub, E., . . . Van Essen, D. C. (2016). A multi-
modal parcellation of human cerebral cortex. Natur, 536,
171–178.
Glasser, M. F., Schmied, S. M., Marcus, D. S., Andersson,
J. L.,
Auerbach, E. J., Behrens, T. E., . . . Van Essen, D. C. (2016). Der
Human Connectome Project’s neuroimaging approach. Natur
Neurowissenschaften, 19, 1175–1187. doi:10.1038/nn.4361
Goel, P., Kuceyeski, A., LoCastro, E., & Raj, A. (2014). Spatial pat-
terns of genome-wide expression profiles reflect anatomic and
fiber connectivity architecture of healthy human brain? Human
Brain Mapping, 35, 4204–4218.
Halstead, J. M., Lionnet, T., Wilbertz, J. H., Wippich, F., Ephrussi,
A., Singer, R. H., . . . Chao, J. A. (2015). Translation: An RNA
biosensor for imaging the first round of translation from single
cells to living animals. Wissenschaft, 347, 1367–1671.
Hawrylycz, M., Müller, J. A., Menon, V., Feng, D., Dolbeare, T.,
Guillozet-Bongaarts, A. L., . . . Lein, E. (2015). Canonical ge-
netic signatures of the adult human brain. Naturneurowissenschaften,
18, 1832–1844. doi:10.1038/nn.4171
Hawrylycz, M. J., Lein, E. S., Guillozet-Bongaarts, A. L., Shen, E. H.,
Ng, L., Müller, J. A., . . . Jones, A. R. (2012). An anatomically com-
prehensive atlas of the adult human brain transcriptome. Natur,
489, 391–399. doi:10.1038/nature11405
Hernandez, L. M., Rudie, J. D., Grün, S. A., Bookheimer, S., &
Dapretto, M. (2015). Neural signatures of autism spectrum dis-
orders: Insights into brain network dynamics. Neuropsychophar-
macology, 40, 171–189.
Hibar, D. P., Stein,
J. L., Renteria, M. E., Arias-Vasquez, A.,
Desrivieres, S., Jahanshad, N., . . . Medland, S. E. (2015). Com-
mon genetic variants influence human subcortical brain struc-
tures. Natur, 520, 224–229. doi:10.1038/nature14101
Holmes, A. J., Hollinshead, M. O., O’Keefe, T. M., Petrov, V. ICH.,
Fariello, G. R., Wald, L. L., . . . Buckner, R. L. (2015). Gehirn
Genomics Superstruct Project initial data release with structural,
functional, and behavioral measures. Scientific Data, 2, 150031.
doi:10.1038/sdata.2015.31
Iwamoto, K., Bundo, M., Yamamoto, M., Ozawa, H., Saito, T., &
Kato, T. (2004). Decreased expression of NEFH and PCP4/PEP19
in the prefrontal cortex of alcoholics. Neuroscience Research,
49, 379–385.
Ji, S., Fakhry, A., & Deng, H. (2014). Integrative analysis of the
connectivity and gene expression atlases in the mouse brain.
NeuroImage, 84, 245–253.
Johnson, M. B., Kawasawa, Y. ICH., Mason, C. E., Krsnik, Z., Coppola,
G., Bogdanovic, D., . . . Sestan, N. (2009). Functional and evo-
lutionary insights into human brain development through global
transcriptome analysis. Neuron, 62, 494–509.
Johnson, M. B., Wang, P. P., Atabay, K. D., Murphy, E. A., Doan,
R. N., Hecht, J. L., . . . Walsh, C. A. (2015). Single-cell analy-
sis reveals transcriptional heterogeneity of neural progenitors in
human cortex. Naturneurowissenschaften, 18, 637–646.
Kang, H. J., Kawasawa, Y. ICH., Cheng, F., Zhu, Y., Xu, X., Li, M., . . .
Sestan, N. (2011). Spatio-temporal transcriptome of the human
Gehirn. Natur, 478, 483–489. doi:10.1038/nature10523
Kaufman, A., Dror, G., Meilijson, ICH., & Ruppin, E. (2006). Gene ex-
pression of Caenorhabditis elegans neurons carries information
on their synaptic connectivity. PLoS Computational Biology, 2,
e167. doi:10.1371/journal.pcbi.0020167
Krienen, F. M., Yeo, B. T., Ge, T., Buckner, R. L., & Sherwood, C. C.
(2016). Transcriptional profiles of supragranular-enriched genes
associate with corticocortical network architecture in the human
Gehirn. Verfahren der Nationalen Akademie der Wissenschaften, 113,
E469–E478.
Krishnaswami, S. R., Grindberg, R. V., Novotny, M., Venepally, P.,
Lacar, B., Bhutani, K., . . . Lasken, R. S. (2016). Using single nuclei
for RNA-seq to capture the transcriptome of postmortem neurons.
Nature Protocols, 11, 499–524. doi:10.1038/nprot.2016.015
Lake, B. B., Ai, R., Kaeser, G. E., Salathia, N. S., Yung, Y. C., Liu,
R., . . . Zhang, K. (2016). Neuronal subtypes and diversity re-
vealed by single-nucleus RNA sequencing of the human brain.
Wissenschaft, 352, 1586–1590. doi:10.1126/science.aaf1204
Lee, H. J., Weitz, A. J., Bernal-Casas, D., Duffy, B. A., Choy, M.,
Kravitz, A. V., . . . Lee, J. H. (2016). Activation of direct and in-
direct pathway medium spiny neurons drives distinct brain-wide
responses. Neuron, 91, 412–424.
Lodato, M. A., Woodworth, M. B., Lee, S., Evrony, G. D., Mehta,
B. K., Karger, A., . . . Walsh, C. A. (2015). Somatic mutation in
single human neurons tracks developmental and transcriptional
Geschichte. Wissenschaft, 350, 94–98. doi:10.1126/science.aab1785
Matthews, P. M., & Hampshire, A. (2016). Clinical concepts emerg-
ing from fMRI functional connectomics. Neuron, 91, 511–528.
J. H.,
McCullumsmith, R. E., Hendrickson, R. C., Gamble, K. L., . . .
Cowell, R. M. (2016). Cortical PGC-1α-dependent transcripts
are reduced in postmortem tissue from patients with schizophre-
nia. Schizophrenia Bulletin, 42, 1009–1017. doi:10.1093/
schbul/sbv184
E. K., Meador-Woodruff,
McMeekin,
Lucas,
J.,
L.
Netzwerkneurowissenschaften
12
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Cognitive genomics: Linking genes to behavior in the human brain
Medland, S. E., Jahanshad, N., Neale, B. M., & Thompson, P. M.
(2014). Whole-genome analyses of whole-brain data: Arbeiten
within an expanded search space. Naturneurowissenschaften, 17,
791–800.
Müller, J. A., Ding, S. L., Sunkin, S. M., Schmied, K. A., Ng, L., Szafer,
A., . . . Lein, E. S. (2014). Transcriptional landscape of the prenatal
human brain. Natur, 508, 199–206. doi:10.1038/nature13185
Okbay, A., Baselmans, B. M., De Neve, J. E., Turley, P., Nivard,
M. G., Fontana, M. A., . . . Cesarini, D. (2016). Genetic vari-
ants associated with subjective well-being, depressive symp-
toms, and neuroticism identified through genome-wide analyses.
Nature Genetics, 48, 624–633.
Okbay, A., Beauchamp, J. P., Fontana, M. A., Lee, J. J., Pers, T. H.,
Rietveld, C. A., . . . Benjamin, D. J. (2016). Genome-wide associ-
ation study identifies 74 loci associated with educational attain-
ment. Natur, 533, 539–542. doi:10.1038/nature17671
Parikshak, N. N., Luo, R., Zhang, A., Won, H., Lowe,
J. K.,
Chandran, V., . . . Geschwind, D. H. (2013). Integrative functional
genomic analyses implicate specific molecular pathways and
circuits in autism. Cell, 155, 1008–1021.
Petersen, S. E., & Spurns, Ö. (2015). Brain networks and cognitive
architectures. Neuron, 88, 207–219.
Pinacho, R., Villalmanzo, N., Meana, J. J., Ferrer, ICH., Berengueras,
A., Haro, J. M., . . . Ramos, B. (2016). Altered CSNK1E, FABP4
and NEFH protein levels in the dorsolateral prefrontal cortex in
schizophrenia. Schizophrenia Research, 177, 88–97.
Pletikos, M., Sousa, A. M., Sedmak, G., Meyer, K. A., Zhu, Y., Cheng,
F., . . . Sestan, N. (2014). Temporal specification and bilaterality
of human neocortical topographic gene expression. Neuron, 81,
321–332.
Poldrack, R. A., Laumann, T. O., Koyejo, O., Gregory, B., Hover, A.,
Chen, M. Y., . . . Mumford, J. A. (2015). Long-term neural and
physiological phenotyping of a single human. Nature Communi-
Kationen, 6, 8885. doi:10.1038/ncomms9885
Pollen, A. A., Nowakowski, T. J., Chen, J., Retallack, H., Sandoval-
Espinosa, C., Nicholas, C. R., . . . Kriegstein, A. R. (2015). Mo-
lecular identity of human outer radial glia during cortical
Entwicklung. Cell, 163, 55–67. doi:10.1016/j.cell.2015.09.004
Richiardi, J., Altmann, A., Milazzo, A. C., Chang, C., Chakravarty,
M. M., Banaschewski, T., . . . IMAGEN Consortium. (2015).
Correlated gene expression supports synchronous activity in
brain networks. Wissenschaft, 348, 1241–1244. doi:10.1126/Wissenschaft.
1255905
Sánchez-Mora, C., Cormand, B., Ramos-Quiroga, J. A., Hervás, A.,
Bosch, R., Palomar, G., . . . Ribasés, M. (2013). Evaluation of
common variants in 16 genes involved in the regulation of neu-
rotransmitter release in ADHD. European Neuropsychopharma-
cology, 23, 426–435. doi:doi:10.1016/j.euroneuro.2012.07.014
Schizophrenia Working Group of the Psychiatric Genomics Con-
sortium (2014). Biologisch
insights from 108 schizophrenia-
associated genetic loci. Natur, 511, 421–427. doi:10.1038/
nature13595
Schumann, G., Loth, E., Banaschewski, T., Barbot, A., Barker, G.,
Büchel, C.,
. . . IMAGEN Consortium (2010). The IMAGEN
Studie: Reinforcement-related behaviour in normal brain function
and psychopathology. Molekulare Psychiatrie, 15, 1128–1139.
doi:10.1038/mp.2010.4
Spencer, D. D., Spencer, S. S., Mattson, R. H., Williamson, P. D.,
& Novelly, R. A. (1984). Access to the posterior medial tem-
poral lobe structures in the surgical treatment of temporal lobe
epilepsy. Neurochirurgie, 15, 667–671.
Tasic, B., Menon, V., Nguyen, T. N., Kim, T. K., Jarsky, T., Yao,
Z., . . . Zeng, H. (2016). Adult mouse cortical cell taxonomy
revealed by single cell transcriptomics. Naturneurowissenschaften, 19,
335–346.
Thompson, P. M., Ge, T., Glahn, D. C., Jahanshad, N., & Nichols,
T. E. (2013). Genetics of the connectome. NeuroImage, 80,
475–488.
Van Essen, D. C., Schmied, S. M., Barch, D. M., Behrens, T. E., Yacoub,
E., Ugur-Auto, K., & the WU–Minn HCP Consortium. (2013). Der
WU–Minn Human Connectome Project: An overview. Neuro-
Image, 80, 62–79.
Voineagu, ICH., Wang, X., Johnston, P., Lowe, J. K., Tian, Y., Horvath,
S., . . . Geschwind, D. H. (2011). Transcriptomic analysis of autis-
tic brain reveals convergent molecular pathology. Natur, 474,
380–384.
Wang, C., Han, B., Zhou, R., & Zhuang, X. (2016). Real-time imag-
ing of translation on single mRNA transcripts in live cells. Cell,
165, 990–1001.
Wang, G. Z., Belgard, T. G., Mao, D., Chen, L., Berto, S., Preuss,
T. M., . . . Konopka, G. (2015). Correspondence between resting-
state activity and brain gene expression. Neuron, 88, 659–666.
Willsey, A. J., Sanders, S. J., Li, M., Dong, S., Tebbenkamp, A. T.,
Muhle, R. A., . . . Zustand, M. W. (2013). Coexpression networks
implicate human midfetal deep cortical projection neurons in
the pathogenesis of autism. Cell, 155, 997–1007. doi:10.1016/
j.cell.2013.10.020
Wolf, L., Goldberg, C., Manor, N., Sharan, R., & Ruppin, E. (2011).
Gene expression in the rodent brain is associated with its regional
Konnektivität. PLoS Computational Biology, 7, e1002040.
Wu, B., Eliscovich, C., Yoon, Y. J., & Singer, R. H. (2016). Transla-
tion dynamics of single mRNAs in live cells and neurons. Wissenschaft,
352, 1430–1435.
Wu, Y. E., Parikshak, N. N., Belgard, T. G., & Geschwind, D. H.
(2016). Genome-wide, integrative analysis implicates microRNA
dysregulation in autism spectrum disorder. Naturneurowissenschaften,
19, 1463–1476. doi:10.1038/nn.4373
Yan, X., Hoek, T. A., Vale, R. D., & Tanenbaum, M. E. (2016). Dy-
namics of translation of single mrna molecules in vivo. Cell, 165,
976–989.
Yang, Z., Zuo, X. N., McMahon, K. L., Craddock, R. C., Kelly, C., von
Zubicaray, G. ICH., . . . Wright, M. J. (2016). Genetic and environ-
mental contributions to functional connectivity architecture of
das menschliche Gehirn. Hirnrinde, 26, 2341–2352. doi:10.1093/
cercor/bhw027
Zeng, H., Shen, E. H., Hohmann,
J. G., Oh, S. W., Bernard,
A., Royall, J. J., . . . Jones, A. R. (2012). Large-scale cellular-
resolution gene profiling in human neocortex reveals species-
specific molecular signatures. Cell, 149, 483–496. doi:10.1016/
j.cell.2012.02.052
Netzwerkneurowissenschaften
13
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
1
1
3
1
0
9
1
8
4
3
N
e
N
_
A
_
0
0
0
0
3
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3