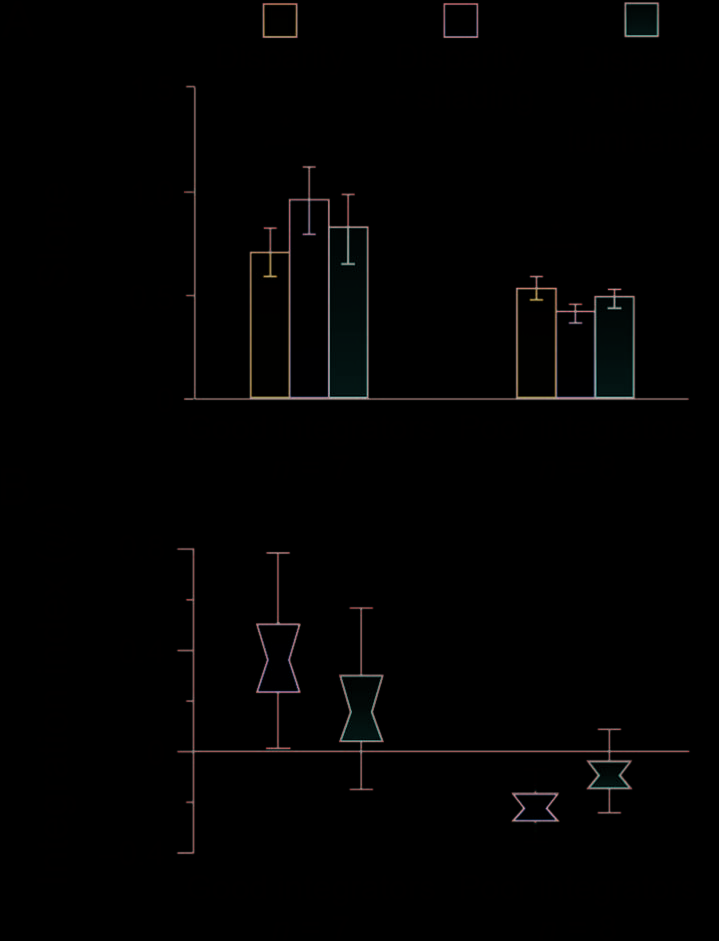Perceptual Integration for Qualitatively Different 3-D
Cues in the Human Brain
Dicle Dövencioğlu1*, Hiroshi Ban1,2*, Andrew J. Schofield1,
and Andrew E. Welchman1
Abstrakt
■ The visual systemʼs flexibility in estimating depth is remark-
able: We readily perceive 3-D structure under diverse condi-
tions from the seemingly random dots of a “magic eye”
stereogram to the aesthetically beautiful, but obviously flat,
canvasses of the Old Masters. Noch, 3-D perception is often en-
hanced when different cues specify the same depth. This per-
ceptual process is understood as Bayesian inference that
improves sensory estimates. Despite considerable behavioral
support for this theory, insights into the cortical circuits in-
volved are limited. Darüber hinaus, extant work tested quantitatively
similar cues, reducing some of the challenges associated with
integrating computationally and qualitatively different signals.
Here we address this challenge by measuring fMRI responses
to depth structures defined by shading, binocular disparity,
and their combination. We quantified information about depth
configurations (convex “bumps” vs. concave “dimples”) in dif-
ferent visual cortical areas using pattern classification analysis.
We found that fMRI responses in dorsal visual area V3B/KO
were more discriminable when disparity and shading concur-
rently signaled depth, in line with the predictions of cue inte-
gration. Wichtig, by relating fMRI and psychophysical tests
of integration, we observed a close association between depth
judgments and activity in this area. Endlich, using a cross-cue
transfer test, we found that fMRI responses evoked by one
cue afford classification of responses evoked by the other. Das
reveals a generalized depth representation in dorsal visual cor-
tex that combines qualitatively different information in line with
3-D perception. ■
EINFÜHRUNG
Many everyday tasks rely on depth estimates provided by
the visual system. To facilitate these outputs, das Gehirn
exploits a range of inputs: from cues related to distance
in a mathematically simple way (z.B., binocular disparity,
motion parallax) to those requiring complex assumptions
and prior knowledge (z.B., shading, occlusion; Burge,
Fowlkes, & Banks, 2010; Kersten, Mamassian, & Yuille,
2004; Mamassian & Goutcher, 2001). These diverse
signals each evoke an impression of depth in their own
Rechts; Jedoch, the brain aggregates cues (Landy, Maloney,
Johnston, & Jung, 1995; Buelthoff & Mallot, 1988;
Dosher, Sperling, & Wurst, 1986) to improve perceptual
judgments (Knill & Saunders, 2003).
Here we probe the neural basis of integration, testing
binocular disparity and shading depth cues that are com-
putationally quite different. Auf den ersten Blick, these cues
may appear so divergent that their combination would
be prohibitively difficult. Jedoch, perceptual judgments
show evidence for the combination of disparity and shad-
ing (Lovell, Bloj, & Harris, 2012; Lee & Saunders, 2011;
Schiller, Slocum, Jao, & Wiener Würstchen, 2011; Vuong, Domini,
1University of Birmingham, 2Japan Society for the Promotion
of Science
*These authors contributed equally to this work.
& Caudek, 2006; Doorschot, Kappers, & Koenderink,
2001; Buelthoff & Mallot, 1988), and the solution to
this challenge is conceptually understood as a two-stage
Verfahren (Landy et al., 1995) in which cues are first ana-
lyzed quasi-independently followed by the integration
of cue information that has been “promoted” into com-
mon units (such as distance). Darüber hinaus, observers can
make reliable comparisons between the perceived depth
from shading and stereoscopic, as well as haptic, compar-
ison stimuli (Schofield, Rock, Sun, Jiang, & Georgeson,
2010; Königreich, 2003), suggesting some form of com-
parable information.
To gain insight into the neural circuits involved in pro-
cessing 3-D information from disparity and shading, pre-
vious brain imaging studies have tested for overlapping
fMRI responses to depth structures defined by the two
Hinweise, yielding locations in which information from dis-
parity and shading converge (Nelissen et al., 2009; Georgieva,
Todd, Peeters, & Orban, 2008; Sereno, Trinath, Augath, &
Logothetis, 2002). Although this is a useful first step, Das
previous work has not established integration: Zum Beispiel,
representations of the two cues might be collocated within
the same cortical area, but represented independently. Von
Kontrast, recent work testing the integration of disparity
and motion depth cues, indicates that integration occurs in
higher dorsal visual cortex (area V3B/kinetic occipital [KO];
Ban, Preston, Meeson, & Welchman, 2012). This suggests a
© 2013 Massachusetts Institute of Technology Published under a
Creative Commons Attribution 3.0 Unportiert (CC–BY 3.0) Lizenz
Zeitschrift für kognitive Neurowissenschaften 25:9, S. 1527–1541
doi:10.1162/jocn_a_00417
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
Ö
9
C
5
N
6
_
4
A
/
_
J
0
Ö
0
C
4
N
1
7
_
A
P
_
D
0
0
B
4
j
1
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
candidate cortical locus in which other types of 3-D infor-
mation may be integrated; Jedoch, it is not clear whether
integration would generalize to (ich) more complex depth
structures and/or (ii) different cue pairings.
Erste, Ban and colleagues (2012) used simple fronto-
parallel planes that can suboptimally stimulate neurons
selective to disparity-defined structures in higher portions
of the ventral ( Janssen, Vogels, & Orban, 2000) and dorsal
streams (Srivastava, Orban, De Maziere, & Janssen, 2009)
compared with more complex curved stimuli. It is there-
fore possible that other cortical areas (especially those in
the ventral stream) would emerge as important for cue
integration if more “shape-like” stimuli were presented.
Zweite, it is possible that information from disparity
and motion are a special case of cue conjunctions, Und
daher, integration effects may not generalize to other
depth signal combinations. Insbesondere, depth from dis-
parity and from motion have computational similarities
(Richards, 1985) and joint neuronal encoding (DeAngelis
& Uka, 2003; Anzai, Ohzawa, & Freeman, 2001; Bradley,
Qian, & Andersen, 1995) and can, grundsätzlich, support
metric (absolute) judgments of depth. Im Gegensatz, Die
3-D pictorial information provided by shading relies on
a quite different generative process that is subject to dif-
ferent constraints and prior assumptions (Thompson,
Fleming, Creem-Regehr, & Stefanucci, 2011; Fleming,
Dror, & Adelson, 2003; Koenderink & van Doorn, 2003;
Mamassian & Goutcher, 2001; Sun & Perona, 1998; Horn,
1975).
To test for cortical responses related to the integration
of disparity and shading, we assessed how fMRI re-
sponses change when stimuli are defined by different
Hinweise (Figure 1A). We used multivoxel pattern analysis
(MVPA) to assess the information contained in fMRI re-
sponses evoked by stimuli depicting different depth con-
figurations (convex vs. concave hemispheres to the left
vs. right of the fixation point). We were particularly inter-
ested in how information about the stimulus contained in
the fMRI signals changed depending on the cues used to
depict depth in the display. Intuitiv, we would expect
that discriminating fMRI responses should be easier
when differences in the depicted depth configuration
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
Ö
9
C
5
N
6
_
4
A
/
_
J
0
Ö
0
C
4
N
1
7
_
A
P
_
D
0
0
B
4
j
1
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
F
T
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Figur 1. Stimulus illustration and experimental procedures. (A) Links: Cartoon of the disparity and/or shading defined depth structure.
One of the two configurations is presented: bumps to the left, dimples to the right. Rechts: Stimulus examples rendered as red–cyan anaglyphs.
(B) Illustration of the psychophysical testing procedure. (C) Illustration of the fMRI block design. (D) Illustration of the vernier task performed by
participants during the fMRI experiment. Participants compared the horizontal position of a vertical line flashed (250 ms) to one eye against
the upper vertical nonius element of the crosshair presented to the other eye.
1528
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 9
were defined by two cues rather than just one (d.h., diff-
ferences defined by disparity and shading together
should be easier to discriminate than differences defined
by only disparity). The theoretical basis for this intuition
can be demonstrated based on statistically optimal dis-
crimination (Ban et al., 2012), with the extent of the
improvement in the two-cue case providing insight into
whether the underlying computations depend on the
integration of two cues or rather having colocated but
independent depth signals.
To appreciate the theoretical predictions for a cortical
area that responds to integrated cues versus colocated
but independent signals, first consider a hypothetical
area that is only sensitive to a single cue (z.B., shading).
If shading information differed between two presented
Reize, we would expect neuronal responses to change,
providing a signal that could be decoded. Im Gegensatz,
manipulating a nonencoded stimulus feature (z.B., dis-
Parität) would have no effect on neuronal responses,
meaning that our ability to decode the stimulus from
the fMRI response would be unaffected. Such a compu-
tationally isolated processing module is biologically
rather unlikely, so next, we consider a more plausible
scenario where an area contains different subpopulations
of neurons, some of which are sensitive to disparity and
others to shading. In this case, we would expect to be
able to decode stimulus differences based on changes
in either cue. Darüber hinaus, if the stimuli contained differ-
ences defined by both cues, we would expect decoding
performance to improve, where this improvement is pre-
dicted by the quadratic sum of the discriminabilities for
changes in each cue. This expectation can be understood
graphically by conceiving of discriminability based on
shading and disparity cues as two sides of a right-angled
triangle, where better discriminability equates to longer
side lengths; the discriminability of both cues together
equals the triangleʼs hypotenuse whose length is deter-
mined based on a quadratic sum (d.h., the Pythagorean
equation) and is always at least as good as the discrimina-
bility of one of the cues.
The alternative possibility is a cortical region that inte-
grates the depth cues. Under this scenario, we also
expect better discrimination performance when two cues
define differences between the stimuli. Wichtig,
Jedoch, unlike the independence scenario, when stimu-
lus differences are defined by only one cue, a fusion
mechanism is adversely affected. Zum Beispiel, if contrast-
ing stimulus configurations differ in the depth indicated
by shading but disparity indicates no difference, Die
fusion mechanism combines the signals from each cue
with the result that it is less sensitive to the combined
estimate than the shading component alone. Durch con-
sequence, if we calculate a quadratic summation predic-
tion based on MVPA performance for depth differences
defined by single cues (d.h., disparity; shading) we will
find that empirical performance in the combined cue
Fall (d.h., disparity + shading) exceeds the prediction
(Ban et al., 2012). Here we exploit this expectation to
identify cortical responses to integrated depth signals,
seeking to identify discrimination performance that is
“greater than the sum of its parts” due to the detrimental
effects of presenting stimuli in which depth differences
are defined in terms of a single cue.
Zu diesem Zweck, we generated random dot patterns (Feige-
ure 1A) that evoked an impression of four hemispheres,
two concave (“dimples”) and two convex (“bumps”). Wir
formulated two different types of display that differed
in their configuration: (1) bumps left–dimples right (de-
picted in Figure 1A) versus (2) dimples left–bumps right.
We depicted depth variations from (ich) binocular dis-
Parität, (ii) shading gradients, Und (iii) the combination
of disparity and shading. Zusätzlich, we employed a con-
trol stimulus (iv) in which the overall luminance of the
top and bottom portions of each hemisphere differed
(Ramachandran, 1988; disparity + binary luminance).
Perceived depth for these (deliberately) crude approxi-
mations of the shading gradients relied on disparity.
We tested for integration using both psychophysical
and fMRI discrimination performance for the component
Hinweise (ich, ii) with that for stimuli containing two cues (iii,
iv). We reasoned that a response based on integrating
cues would be specific to concurrent cue stimulus (iii)
and not be observed for the control stimulus (iv).
METHODEN
Teilnehmer
Twenty observers from the University of Birmingham
participated in the fMRI experiments. Of these, five were
excluded due to excessive head movement during scan-
ning, meaning that the correspondence between voxels
required by the MVPA technique was lost. Excessive
movement was defined as ≥4 mm over an 8-min run,
and we excluded participants if they had fewer than five
runs below this cut-off as there was insufficient data for
the MVPA. Generally, participants were able to keep still:
The average absolute maximum head deviation relative
to the start of the first run for included participants was
1.2 mm versus 4.5 mm for excluded participants. More-
over only one included participant had an average head
motion of >2 mm per run, and the mode of the head
movement distribution across participants was <1 mm.
Six women and nine men were included; 12 right-
handed. Mean age was 26 ± 1.2 (SEM ) years. Authors
A.E.W. H.B. participated; all other participants were
naive to the purpose of study. Four participants
had taken part in Ban et al.ʼs (2012) Participants
had normal or corrected-to-normal vision pre-
screened for stereo deficits. Experiments approved
by University Birmingham Science Engineer-
ing Ethics Committee; observers gave written informed
consent.
Dövencioğlu al.
1529
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
> 0) and selected to the top 300 voxels as
data for the classification algorithm (Preston, Li, Kourtzi,
& Welchman, 2008). To minimize baseline differences
between runs we z-scored the response time course of
each voxel and each experimental run. To account for
the hemodynamic response lag, the fMRI time series
were shifted by two repetition times (4 Sek). Thereafter,
we averaged the fMRI response of each voxel across the
16 sec stimulus presentation block, obtaining a single test
pattern for the multivariate analysis per block. To remove
potential univariate differences (that can be introduced
after z-score normalization due to averaging across time
points in a block and grouping the data into train vs. test
data sets), we normalized by subtracting the mean of all
voxels for a given volume (Serences & Boynton, 2007),
with the result that each volume had the same mean
value across voxels and differed only in the pattern of
Aktivität. We performed multivoxel pattern analysis using
a linear support vector machine (SVMlight toolbox) clas-
sification algorithm. We trained the algorithm to dis-
tinguish between fMRI responses evoked by different
stimulus configurations (z.B., convex to the left vs. Zu
the right of fixation) for a given stimulus type (z.B., dis-
Parität). Participants typically took part in eight runs, jede
of which had three repetitions of a given spatial config-
uration and stimulus type, creating a total of 24 patterns.
We used a leave-one-run-out cross-validation procedure:
We trained the classifier using seven runs (d.h., 21 pat-
Seeschwalben) and then evaluated the prediction performance
of the classifier using the remaining, nontrained data (d.h.,
three patterns). We repeated this, leaving a single run out
im Gegenzug, and calculated the mean prediction accuracy across
cross-validation folds. Accuracies were represented in units
of discriminability (D
0) using the formula:
D 0 ¼ 2erfinvð2p−1Þ
ð1Þ
where erfinv is the inverse error function and p is the
proportion of correct predictions.
For tests of transfer between disparity and shading
Hinweise, we used a Recursive Feature Elimination method
(De Martino et al., 2008) to detect sparse discriminative
patterns and define the number of voxels for the SVM
classification analysis. In each feature elimination step,
five voxels were discarded until there remained a core
set of voxels with the highest discriminative power. To
avoid circular analysis, the Recursive Feature Elimination
method was applied independently to the training pat-
terns of each cross-validation fold, resulting in eight sets
of voxels (d.h., one set for each test pattern of the leave-
one-run-out procedure). This was done separately for
each experimental condition, with final voxels for the
SVM analysis chosen based on the intersection of voxels
from corresponding cross-validation folds. A standard
SVM was then used to compute within- and between-
cue prediction accuracies. This feature selection method
was required for transfer, in line with evidence that it
improves generalization (De Martino et al., 2008).
We conducted repeated-measures GLM in SPSS (IBM,
Inc., Armonk, New York) applying Greenhouse-Geisser correc-
tion when appropriate. Regression analyses were also
conducted in SPSS. Für diese Analyse, we considered the
use of repeated-measures MANCOVA (and found results
consistent with the regression results); Jedoch, Die
integration indices (defined below) we use are partially
correlated between conditions because their calculation
depends on the same denominator, violating the GLMʼs
assumption of independence. We therefore limited our
analysis to the relationship between psychophysical and
fMRI indices for the same condition, for which the psycho-
physical and fMRI indices are independent of one another.
Quadratic Summation and Integration Indices
We formulate predictions for the combined cue condi-
tion (d.h., disparity + shading) based on the quadratic
summation of performance in the component cue condi-
tionen (d.h., disparity, shading). As outlined in the Intro-
duktion, this prediction is based on the performance of
1532
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 9
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
Ö
9
C
5
N
6
_
4
A
/
_
J
0
Ö
0
C
4
N
1
7
_
A
P
_
D
0
0
B
4
j
1
G
7
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
introduced during fMRI measurement (scanner noise,
observer movement), colocated but independent re-
sponses can yield a positive index (see the fMRI simula-
tions by Ban et al., 2012 in their Supplementary Figure 3).
Daher, the integration index alone cannot be taken as
definite evidence of cue integration and therefore needs
to be considered in conjunction with the other tests. To
assess statistical significance of the integration indices,
we used bootstrapped resampling as our use of a ratio
makes distributions non-Normal, and thus a nonpara-
metric procedure more appropriate.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
Figur 2. Psychophysical results. (A) Behavioral tests of integration.
Bar graphs represent the between-subject mean slope of the
psychometric function. *P < .05. (B) Psychophysical results as an
integration index. Distribution plots show bootstrapped values: The
center of the “bowtie” represents the median, the colored area depicts
68% confidence values, and the upper and lower error bars represent
95% confidence intervals.
an ideal observer model that discriminates pairs of inputs
(visual stimuli or fMRI response patterns) based on the
optimal discrimination boundary. Psychophysical tests
indicate that this theoretical model matches human per-
formance in combining cues (Knill & Saunders, 2003;
Hillis, Ernst, Banks, & Landy, 2002).
To compare measured empirical performance in dis-
parity + shading condition with the prediction derived
from the component cue conditions, we calculate a ratio
index (Ban et al., 2012; Nandy & Tjan, 2008) whose gen-
eral form is
Index ¼ CDþS
p
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
þ C2
C2
D
S
−1
ð2Þ
where CD, CS, and CD+S are sensitivities for disparity,
shading, and the combined cue conditions, respectively.
If the responses of the detection mechanism to the dis-
parity and shading conditions (CD, CS) are independent
of each other, performance when both cues are available
(CD+S) should match the quadratic summation pre-
diction, yielding a ratio of 1 and thus an index of zero.
A value of less than zero suggests suboptimal detection
performance, and a value above zero suggests that the
component sources of information are not independent
(Ban et al., 2012; Nandy & Tjan, 2008). However, a value
above zero does not preclude the response of indepen-
dent mechanisms: Depending on the amount of noise
RESULTS
Psychophysics
To assess cue integration psychophysically, we measured
observersʼ sensitivity to slight differences in the depth
profile of the stimuli (Figure 1B). Participants viewed
two shapes sequentially, and decided which had the
greater depth (i.e., which bumps were taller or which
dimples were deeper). By comparing a given standard
stimulus against a range of test stimuli, we obtained
psychometric functions. We used the slope of these
functions to quantify observersʼ sensitivity to stimulus
differences (where a steeper slope indicates higher sen-
sitivity). To determine whether there was a perceptual
benefit associated with adding shading information to
the stimuli, we compared performance in the disparity
condition with that in the disparity and shading condi-
tion. Surprisingly, we found no evidence for enhanced
performance in the disparity and shading condition at
the group level, F(1, 14) < 1, p = .38. In light of previous
empirical work on cue integration, this was unexpected
(e.g., Lovell et al., 2012; Schiller et al., 2011; Vuong
et al., 2006; Doorschot et al., 2001; Buelthoff & Mallot,
1988) and prompted us to consider the significant var-
iability between observers, F(1, 14) = 62.23, p < .001,
in their relative performance in the two conditions. In
particular, we found that some participants clearly bene-
fited from the presence of two cues; however, others
showed no benefit and some actually performed worse
relative to the disparity only condition. Poorer perfor-
mance might relate to individual differences in the as-
sumed direction of the illuminant (Schofield, Rock, &
Georgeson, 2011); ambiguity or bistability in the inter-
pretation of shading patterns (Wagemans, van Doorn,
& Koenderink, 2010; Liu & Todd, 2004); and/or differ-
ences in cue weights (Lovell et al., 2012; Schiller et al.,
2011; Knill & Saunders, 2003; we return to this issue in
the Discussion). To quantify variations between partici-
pants in the relative performance in two conditions, we
calculated a psychophysical integration index (ψ):
ψ ¼ SDþS
SD
−1
ð3Þ
Dövencioğlu et al.
1533
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
fMRI Measures
Before taking part in the main experiment, each partici-
pant underwent a separate fMRI session to identify ROIs
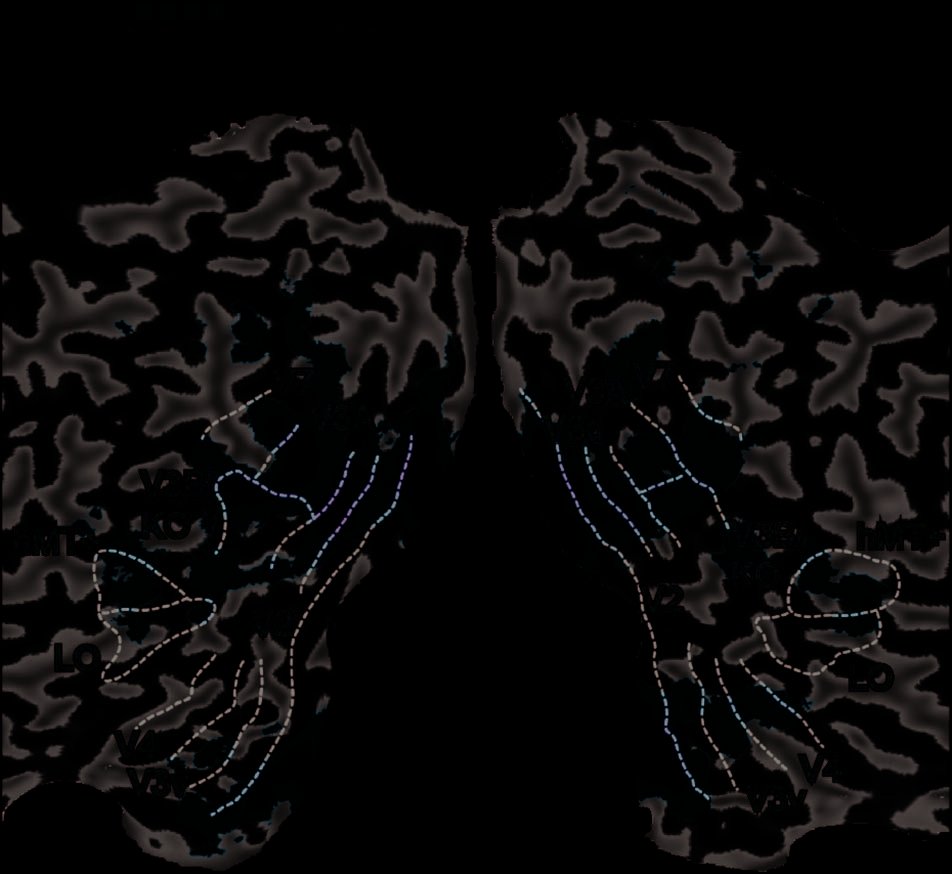
within the visual cortex (Figure 3). We identified retino-
topically organized cortical areas based on polar and
eccentricity mapping techniques (Tyler et al., 2005;
Tootell & Hadjikhani, 2001; DeYoe et al., 1996; Sereno
et al., 1995). In addition, we identified area LO involved
in object processing (Kourtzi & Kanwisher, 2001), the
human motion complex (hMT+/ V5; Zeki et al., 1991),
and the KO region, which is localized by contrasting
motion-defined contours with transparent motion (Zeki
et al., 2003; Dupont et al., 1997). Responses to the KO
localizer overlapped with the retinotopically localized
area V3B and were not consistently separable across
participants and/or hemispheres (see also Ban et al.,
2012) so we denote this region as V3B/KO. A representa-
tive flatmap of the ROIs is shown in Figure 3, and Table 1
provides mean coordinates for V3B/KO.
We then measured fMRI responses in each of the ROIs
and were a priori particularly interested in the V3B/KO
region (Ban et al., 2012; Tyler, Likova, Kontsevich, & Wade,
2006). We presented stimuli from four experimental condi-
tions (Figure 1) under two configurations: (a) bumps to the
left of fixation, dimples to the right or (b) bumps to the
right, dimples to the left, thereby allowing us to contrast
fMRI responses to convex versus concave stimuli.
To analyze our data, we trained a machine learning
classifier (SVM) to associate patterns of fMRI voxel activity
and the stimulus configuration (convex vs. concave) that
gave rise to that activity. We used the performance of the
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
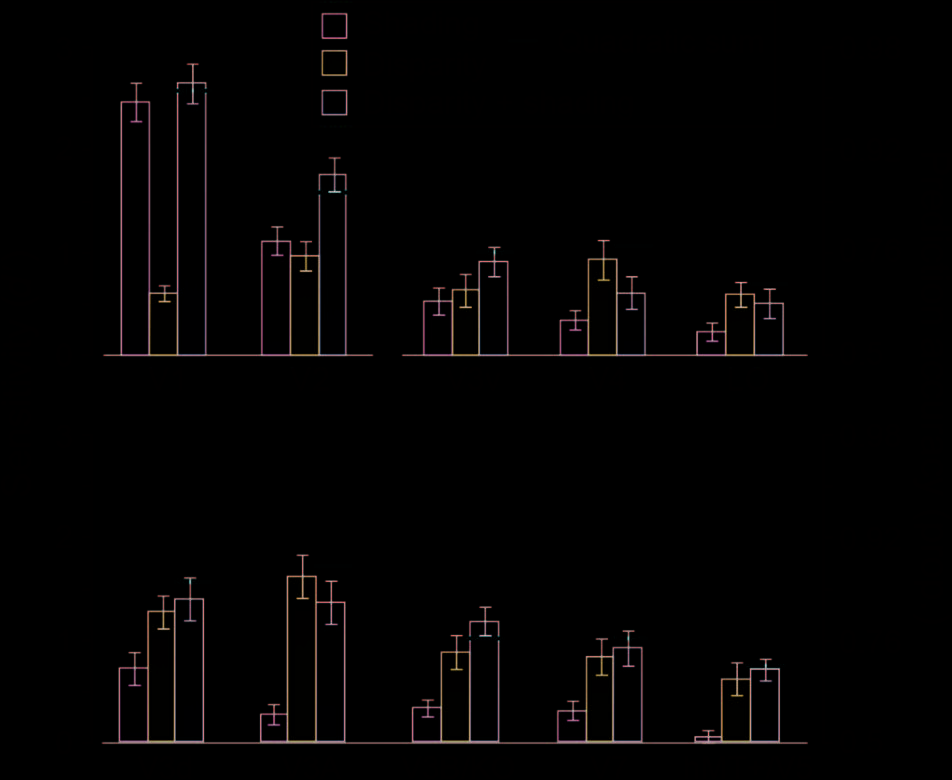
Figure 4. Performance in predicting the convex versus concave
configuration of the stimuli based on the fMRI data measured in
different ROIs. The bar graphs show the results from the “single cue”
experimental conditions, the “disparity + shading” condition, and the
quadratic summation prediction (horizontal red line). Error bars
indicate SEM.
Figure 3. Representative flat maps from one participant showing the
left and right ROIs. The sulci are depicted in darker gray than the gyri.
Shown on the maps are retinotopic areas, V3B/KO, the human motion
complex (hMT+/ V5), and LO area. The activation on the maps shows
the results of a searchlight classifier analysis that moved iteratively
throughout the measured cortical volume, discriminating between
stimulus configurations. The color code represents the t value of the
classification accuracies obtained. This procedure confirmed that we had
not missed any important areas outside those localized independently.
p
where SD+S is sensitivity in the combined condition and
SD is sensitivity in the disparity condition. This index is
based on the quadratic summation test (Ban et al.,
2012; Nandy & Tjan, 2008; see Methods) where a value
above zero suggests that participants integrate the depth
information provided by the disparity and shading cues
when making perceptual judgments. In this instance we
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
assumed that SD ≈
Þ
þ S 2
ðS 2
because our attempts to
S
D
measure sensitivity to differences in depth amplitude
defined by shading alone in pilot testing resulted in such
poor performance that we could not fit a reliable psycho-
metric function. Specifically, discriminability for the max-
0 = 0.3 ± 0.25 for
imum presented depth difference was d
0 = 3.9 ± 0.3 for disparity,
shading alone, in contrast to d
that is, SD
2 ≫ SS
2.
We used a clustering algorithm on ψ to determine
whether our participants formed different subgroups.
In particular, SPSSʼs two-step clustering algorithm (apply-
ing Schwarzʼs Bayesian Criterion for cluster identifica-
tion) indicated two subgroups: Participants with ψ >
0.1 were associated with cluster 1 and participants with
ψ < −0.1 with cluster 2; hereafter, we refer to these
groups as good integrators (n = 7) and poor integrators
(n = 8). By definition, these post hoc groups differed
in the relative sensitivity to disparity and disparity +
shading conditions (Figure 2). Our purpose in forming
these groups, however, was to test the link between dif-
ferences in perception and fMRI responses.
1534
Journal of Cognitive Neuroscience
Volume 25, Number 9
classifier in decoding the stimulus from independent fMRI
data (i.e., leave-one-run-out cross validation) as a measure
of the information about the presented stimulus within a
particular region of cortex.
We could reliably decode the stimulus configuration in
the four conditions in almost every ROI (Figure 4), and
there was a clear interaction between conditions and
ROIs, F(8.0, 104.2) = 8.92, p < .001. This widespread
sensitivity to differences between convex versus concave
stimuli is not surprising in that a range of features might
modify the fMRI response (e.g., distribution of image
intensities, contrast edges, mean disparity, etc.). The
machine learning classifier may thus decode low-level
image features, rather than “depth” per se. We were
therefore interested not in overall prediction accuracies
between areas (which are influenced by our ability to
measure fMRI activity in different anatomical locations).
Rather, we were interested in the relative performance
between conditions, and whether this related to between-
observer differences in perceptual integration. We there-
fore considered our fMRI data subdivided based on the
behavioral results (significant interaction between condi-
tion and group [good vs. poor integrators]: F(2.0, 26.6) =
4.52, p = .02).
First, we wished to determine whether fMRI decoding
performance improved when both depth cues indicated
depth differences. Prediction accuracies for the concurrent
stimulus (disparity + shading) were statistically higher
than the component cues in areas V2, F(3, 39) = 7.47,
p < .001, and V3B/KO, F(1.6, 21.7) = 14.88, p < .001.
To assess integration, we compared the extent of improve-
ment in the concurrent stimulus relative to a minimum
bound prediction (Figures 4 and 5, red lines) based on
the quadratic summation of decoding accuracies for
“single cue” presentations (Ban et al., 2012). This cor-
responds to the level of performance expected if dis-
parity signals and shading signals are collocated in a
cortical area, but represented independently. If perfor-
mance exceeds this bound, it suggests that cue repre-
sentations are not independent, as performance in the
“single” cue case was attenuated by the conflicts that
result from “isolating” the cue. We found that perfor-
mance was higher (outside the SEM) than the quadratic
summation prediction in areas V2 and V3B/KO (Figure 5).
However, this result was only statistically reliable in
V3B/KO. Specifically in V3B/KO, there was a significant
interaction between the behavioral group and experi-
mental condition, F(2, 26) = 5.52, p = .01, with decod-
ing performance in the concurrent (disparity +
shading) condition exceeding the quadratic summation
prediction for good integrators, F(1, 6) = 9.27, p = .011,
but not for the poor integrators, F(1, 7) < 1, p = .35
(Figure 5). In V2, there was no significant difference be-
tween the quadratic summation prediction and the mea-
sured data in the combined cue conditions, F(2, 26) < 1,
p = .62, nor an interaction, F(2, 26) = 2.63, p = .091. We
quantified the extent of integration using a boot-
strapped index (ϕ) that contrasted decoding perfor-
mance in the concurrent condition (d
D+S) with the
quadratic summation of performance with “single” cues
(d
D and d
S):
0
0
0
ϕ ¼
p
d0
DþS
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
D2 þ d0
d0
S2
−1
ð4Þ
Using this index, a value of zero corresponds to the
performance expected if information from disparity
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
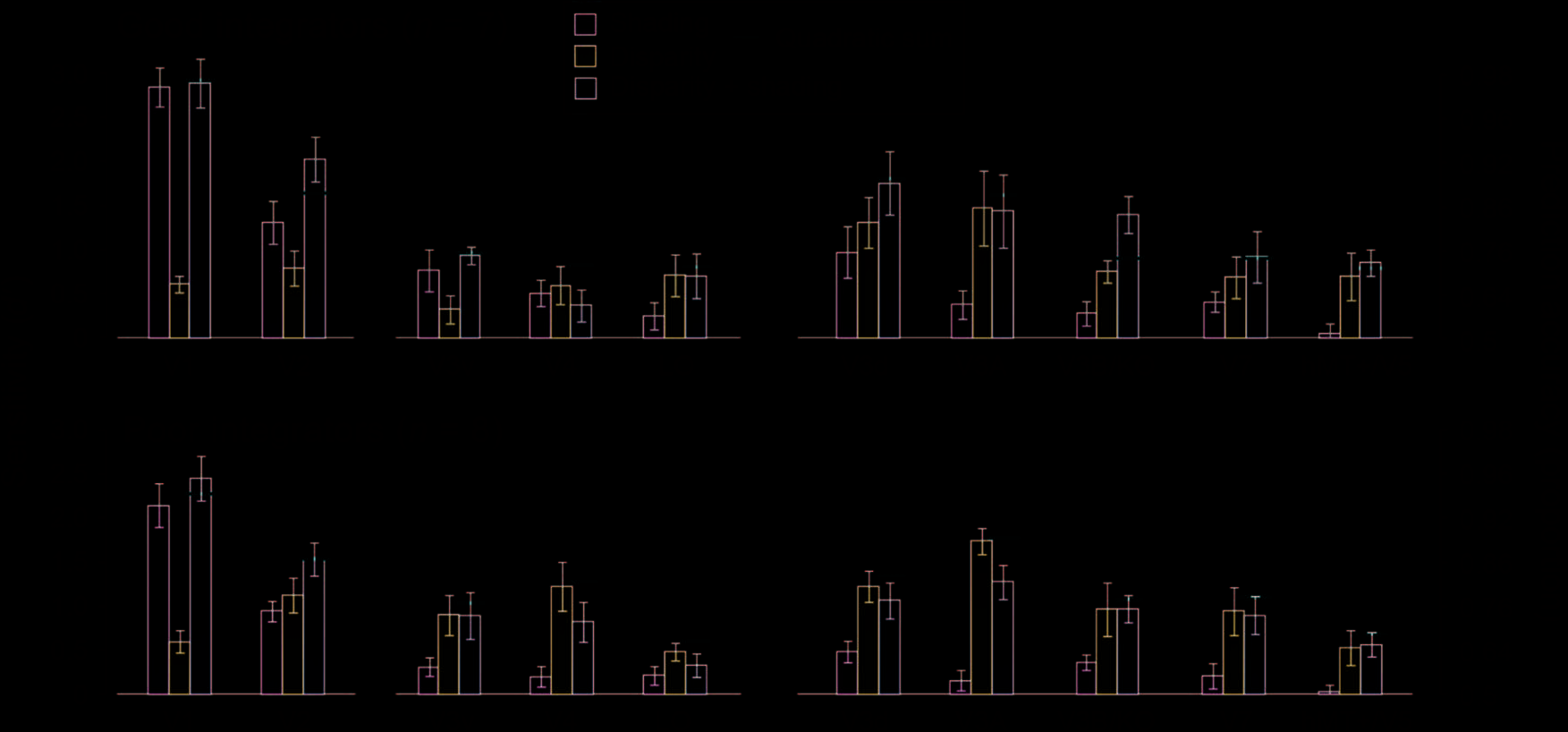
Figure 5. Prediction performance for fMRI data separated into the two groups based on the psychophysical results (“good” vs. “poor”
integrators). The bar graphs show the results from the “single cue” experimental conditions, the “disparity + shading” condition, and the
quadratic summation prediction (horizontal red line). Error bars indicate SEM.
Dövencioğlu et al.
1535
that approximated luminance differences in the shaded
stimuli but did not, per se, evoke an impression of
depth. As the fMRI response in a given area may reflect
low-level stimulus differences (rather than depth from
shading), we wanted to rule out the possibility that
improved decoding performance in the concurrent dis-
parity + shading condition could be explained on the
basis that two separate stimulus dimensions (disparity
and luminance) drive the fMRI response. The quadratic
summation test should theoretically rule this out; never-
theless, we contrasted decoding performance in the
concurrent condition versus the binary control (disparity
+ binary luminance) condition. We reasoned that if
enhanced decoding is related to the representation of
depth, superquadratic summation effects would be lim-
ited to the concurrent condition. On the basis of a
significant interaction between subject group and condi-
tion, F(2, 26) = 5.52, p = .01, we found that this was true
for the good integrator subjects in area V3B/KO: sensi-
tivity in the concurrent condition was above that in the
binary control condition, F(1, 6) = 14.69, p = .004. By
contrast, sensitivity for the binary condition in the poor
integrator subjects matched that of the concurrent
group, F(1, 7) < 1, p = .31, and was in line with quadratic
summation. Results from other ROIs (Table 2) did not
suggest the clear (or significant) differences that were
apparent in V3B/KO.
As a further line of evidence, we used regression ana-
lyses to test the relationship between psychophysical and
fMRI measures of integration. Although we would not
anticipate a one-to-one mapping between them (the
fMRI data were obtained for differences between concave
vs. convex shapes, whereas the psychophysical tests
measured sensitivity to slight differences in the depth
profile), our group-based analysis suggested a corre-
spondence. We found a significant relationship between
the fMRI and psychophysical integration indices in V3B/
KO (Figure 6B) for the concurrent (R = 0.57, p = .026)
but not the binary luminance (R = 0.10, p = .731) con-
dition. This result was specific to area V3B/KO (Table 3)
and, in line with the preceding analyses, suggests a rela-
tionship between activity in area V3B/KO and the percep-
tual integration of disparity and shading cues to depth.
As a final assessment of whether fMRI responses related
to depth structure from different cues, we tested whether
training the classifier on depth configurations from one cue
(e.g., shading) afforded predictions for depth configura-
tions specified by the other (e.g., disparity). To compare
the prediction accuracies on this cross-cue transfer with
baseline performance (i.e., training and testing on the
same cue), we used a bootstrapped transfer index:
T ¼ 2d0
T
þ d0
d0
D
S
ð5Þ
0
where d
½ (d
0
D + d
T is between-cue transfer performance and
S) is the average within-cue performance. A
0
Figure 6. (A) fMRI based prediction performance as an integration
index for the two groups of participants in area V3B/KO. A value of
zero indicates the minimum bound for fusion as predicted by quadratic
summation. The index is calculated for the “disparity + shading” and
“disparity + binary shading” conditions. Data are presented as notched
distribution plots. The center of the “bowtie” represents the median, the
colored area depicts 68% confidence values, and the upper and lower
error bars represent 95% confidence intervals. (B) Correlation between
behavioral and fMRI integration indices in area V3B/KO. Psychophysics
and fMRI integration indices are plotted for each participant for disparity
+ shading and disparity + binary luminance conditions. The Pearson
correlation coefficient (R) and p value are shown. (C) The transfer index
values for V3B/KO for the good and poor integrator groups. Using this
index, a value of 1 indicates equivalent prediction accuracies when
training and testing on the same cue versus training and testing on
different cues. Distribution plots show the median, 68% and 95%
confidence intervals. Dotted horizontal lines depict a bootstrapped
chance baseline based on the upper 95th centile for transfer analysis
obtained with randomly permuted data.
and shading are collocated, but independent. We found
that only in areas V2 and V3B/ KO was the integration
index for the concurrent condition reliably above zero
for the good integrators (Figure 6A; Table 2).
To provide additional evidence for neuronal responses
related to depth estimation, we used the binary lumi-
nance stimuli as a control. We constructed these stimuli
such that they contained a very obvious low-level feature
1536
Journal of Cognitive Neuroscience
Volume 25, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 2. Probabilities Associated with Obtaining a Value of Zero for the fMRI Integration Index in the (i) Disparity + Shading
Condition and (ii) Luminance Control Condition
Disparity + Shading
Disparity + Binary Luminance
Cortical Area
Good Integrators
Poor Integrators
Good Integrators
Poor Integrators
V1
V2
V3v
V4
LO
V3d
V3A
V3B/KO
V7
hMT+/ V5
0.538
0.004
0.294
0.916
0.656
0.253
0.609
<0.001
0.298
0.315
0.157
0.419
0.579
0.942
0.944
0.890
1.000
0.629
0.595
0.421
0.999
0.607
0.726
0.987
0.984
0.909
0.999
0.327
0.844
0.978
0.543
0.102
1.000
0.628
0.143
0.234
0.961
0.271
0.620
0.575
Values are from a bootstrapped resampling of the individual participantsʼ data using 10,000 samples. Bold formatting indicates Bonferroni-corrected
significance.
value of one using this index indicates that prediction
accuracy between cues equals that within cues. To provide
a baseline for the transfer that might occur by chance, we
calculated the transfer index on data sets for which we ran-
domly shuffled the condition labels, such that we broke the
relationship between fMRI response and the stimulus that
evoked the response. We calculated shuffled transfer per-
formance 1000 times for each ROI and used the 95th cen-
tile of the resulting distribution of transfer indices as the
Table 3. Results for the Regression Analyses Relating the
Psychophysical and fMRI Integration Indices in Each ROI
Cortical
Area
V1
V2
V3v
V4
LO
V3d
V3A
V3B/KO
V7
hMT+/ V5
Disparity +
Shading
Disparity +
Binary Luminance
R
−0.418
0.105
−0.078
0.089
0.245
0.194
0.232
0.571
0.019
0.411
p
.121
.709
.782
.754
.379
.487
.405
.026
.946
.128
R
−0.265
−0.394
0.421
−0.154
−0.281
−0.157
−0.157
0.097
−0.055
−0.367
p
.340
.146
.118
.584
.311
.577
.577
.731
.847
.178
The table shows the Pearson correlation coefficient (R) and the signifi-
cance of the fit as a p value for the “disparity + shading” and “disparity +
binary luminance” conditions.
cut-off for significance. We found reliable evidence for
transfer between cues in area V3B/KO (Figure 6C) for the
good, but not poor, integrator groups. Furthermore, this
effect was specific to V3B/KO and was not observed in
other areas (Table 4). Together with the previous ana-
lyses, this result suggests a degree of equivalence between
representations of depth from different cues in V3B/KO
that is related to an individualʼs perceptual interpretation
of cues.
To ensure we had not missed any important loci of
activity outside the areas we sampled using our ROI local-
izers, we conducted a searchlight classification analysis
(Kriegeskorte, Goebel, & Bandettini, 2006) in which we
moved a small spherical aperture (diameter = 9 mm)
through the sampled cortical volume performing MVPA
on the difference between stimulus configurations for
the concurrent cue condition (Figure 3). This analysis
indicated that discriminative signals about stimulus dif-
ferences were well captured by our ROI definitions.
Our main analyses considered MVPA of the fMRI
responses partitioned into two groups based on psycho-
physical performance. To ensure that differences in
MVPA prediction performance between groups related
to the pattern of voxel responses for depth process-
ing, rather than the overall responsiveness of different
ROIs, we calculated the average fMRI activations (percent
signal change) in each ROI for the two groups of partici-
pants. Reassuringly, we found no evidence for statistically
reliable differences between groups across conditions
and ROIs (i.e., no ROI × Group interaction: F(3.3,
43.4) < 1, p = .637; no Condition × Group interaction:
F(3.5, 45.4) < 1, p = .902; and no ROI × Condition × Group
interaction: F(8.6, 112.2) = 1.06, p = .397). Moreover,
limiting this analysis to V3B/KO provided no evidence
Dövencioğlu et al.
1537
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 4. Probabilities Associated with the Transfer between
Disparity and Shading Producing a Transfer Index above the
Random (Shuffled) Baseline
Cortical Area
Good Integrators
Poor Integrators
V1
V2
V3v
V4
LO
V3d
V3A
V3B/KO
V7
hMT+/ V5
0.247
0.788
0.121
0.478
0.254
0.098
0.295
<0.001
0.145
0.124
0.748
0.709
0.908
0.062
0.033
0.227
0.275
0.212
0.538
0.302
These p values are calculated using bootstrapped resampling with
10,000 samples. Bold formatting indicates Bonferroni-corrected
significance.
for a difference in the percent signal change between
groups (i.e., no Condition × Group interaction: F(3.1,
40.3) < 1, p = .586). Furthermore, we ensured that we
had sampled from the same cortical location in both
groups by calculating the mean Talairach location of
V3B/KO subdivided by groups (Table 1). This confirmed
that we had localized the same cortical region in both
groups of participants.
To guard against artifacts complicating the interpreta-
tion of our results, we took specific precautions during
scanning to control attentional allocation and eye move-
ments. First, participants performed a demanding vernier
judgment task at fixation. This ensured equivalent atten-
tional allocation across conditions, and, as the task was
unrelated to the depth stimuli, psychophysical judgments
and fMRI responses were not confounded and could not
thereby explain between-subject differences. Second, the
attentional task served to provide a subjective measure of
eye vergence (Popple et al., 1998). In particular, partici-
pants judged the relative location of a small target flashed
(250 msec) to one eye, relative to the upper vertical
nonius line (presented to the other eye; Figure 1D).
We fit the proportion of “target is to the right” responses
as a function of the targetʼs horizontal displacement. Bias
(i.e., deviation from the desired vergence position) in this
judgment was around zero suggesting that participants
were able to maintain fixation with the required vergence
angle. Using a repeated-measures ANOVA, we found that
there were no significant differences in bias between Stim-
ulus Conditions, F(1.5, 21.4) = 2.59, p = .109, Sign of
Curvature, F(1, 14) = 1.43, p = .25, and no interaction,
F(2.2, 30.7) = 1.95, p = .157. Furthermore, there were
no differences in the slope of the psychometric functions:
no effect of Condition, F(3, 42) < 1, p = .82, or Curvature,
F(1, 14) < 1, p = .80, and no interaction, F(3, 42) < 1,
p = .85.
Third, our stimuli were constructed to reduce the
potential for vergence differences: Disparities to the left
and right of the fixation point were equal and opposite, a
constant low spatial frequency pattern surrounded the
stimuli, and participants used horizontal and vertical
nonius lines to monitor their eye vergence.
DISCUSSION
Here we provide three lines of evidence that activity in
dorsal visual area V3B/KO reflects the integration of dis-
parity and shading depth cues in a perceptually relevant
manner. First, we used a quadratic summation test to
show that performance in concurrent cue settings im-
proves beyond that expected if depth from disparity
and shading are collocated but represented indepen-
dently. Second, we showed that this result was specific
to stimuli that are compatible with a 3-D interpretation
of shading patterns. Third, we found evidence for
cross-cue transfer. Importantly, the strength of these re-
sults in V3B/KO varied between individuals in a manner
that was compatible with their perceptual use of inte-
grated depth signals.
These findings complement evidence for the integra-
tion of disparity and relative motion in area V3B/KO
(Ban et al., 2012) and suggest both a strong link with
perceptual judgments and a more generalized represen-
tation of depth structure. Such generalization is far from
trivial: Binocular disparity is a function of an objectʼs 3-D
structure, its distance from the viewer and the separation
between the viewerʼs eyes; by contrast, shading cues (i.e.,
intensity distributions in the image) depend on the type
of illumination, the orientation of the light source with
respect to the 3-D object, and the reflective properties of
the objectʼs surface (i.e., the degree of Lambertian and
Specular reflectance). As such disparity and shading pro-
vide complementary shape information: They have quite
different generative processes, and their interpreta-
tion depends on different constraints and assumptions
(Doorschot et al., 2001; Blake, Zisserman, & Knowles,
1985). Taken together, these results indicate that the 3-D
representations in the V3B/KO region are not specific to
specific cue pairs (i.e., disparity–motion) and generalize
to more complex forms of 3-D structural information
(i.e., local curvature). This points to an important role
for higher portions of the dorsal visual cortex in comput-
ing information about the 3-D structure of the surround-
ing environment.
Individual Differences in Disparity and
Shading Integration
One striking, and unexpected feature of our findings was
that we observed significant between-subject variability in
the extent to which shading enhanced performance, with
1538
Journal of Cognitive Neuroscience
Volume 25, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
some participants benefitting, and others actually per-
forming worse. What might be responsible for this varia-
tion in performance? Although shading cues support
reliable judgments of ordinal structure (Ramachandran,
1988), shape is often underestimated (Mingolla & Todd,
1986) and subject to systematic biases related to the
estimated light source position (Mamassian & Goutcher,
2001; Sun & Perona, 1998; Curran & Johnston, 1996;
Pentland, 1982) and light source composition (Schofield
et al., 2011). Moreover, assumptions about the position
of the light source in the scene are often esoteric: Most
observers assume overhead lighting, but the strength of
this assumption varies considerably (Thomas, Nardini, &
Mareschal, 2010; Wagemans et al., 2010; Liu & Todd,
2004), and some observers assume lighting from below
(e.g., 3 of 15 participants in Schofield et al., 2011). Our
disparity + shading stimuli were designed such that the
cues indicated the same depth structure to an observer
who assumed lighting from above. Therefore, it is quite
possible that observers experienced conflict between the
shape information specified by disparity and that deter-
mined by their interpretation of the shading pattern.
Such participants would be “poor integrators” only inas-
much as they failed to share the assumptions typically
made by observers (i.e., lighting direction, lighting com-
position, and Lambertian surface reflectance) when inter-
preting shading patterns. In addition, participants may
have experienced alternation in their interpretation of
the shading cue across trials (i.e., a weak light-from-above
assumption that has been observed quite frequently;
Thomas et al., 2010; Wagemans et al., 2010); aggregating
such bimodal responses to characterize the psychometric
function would result in more variable responses in the
concurrent condition than in the “disparity“ alone condi-
tion, which was not subject to perceptual bistability. Such
variations could also result in fMRI responses that vary
between trials; in particular, fMRI responses in V3B/KO
change in line with different perceptual interpretations
of the same (ambiguous) 3-D structure indicated by
shading cues (Preston, Kourtzi, & Welchman, 2009). This
variation in fMRI responses could thereby account for
reduced decoding performance for these participants.
An alternative possibility is that some of our observers
did not integrate information from disparity and shading
because they are inherently poor integrators. Although
cue integration both within and between sensory modal-
ities has been widely reported in adults, it has a develop-
mental trajectory and young children do not integrate
signals (Gori, Del Viva, Sandini, & Burr, 2008; Nardini,
Jones, Bedford, & Braddick, 2008; Nardini, Bedford, &
Mareschal, 2010). This suggests that cue integration may
be learnt via exposure to correlated cues (Atkins, Fiser, &
Jacobs, 2001) where the effectiveness of learning can
differ between observers (Ernst, 2007). Furthermore,
although cue integration may be mandatory for many
cues where such correlations are prevalent (Hillis et al.,
2002), interindividual variability in the prior assumptions
that are used to interpret shading patterns may cause
some participants to lack experience of integrating shad-
ing and disparity cues (at least in terms of how these are
studied in the laboratory).
These different possibilities are difficult to distinguish
from previous work that has looked at the integration of
disparity and shading signals and reported individual re-
sults. This work indicated that perceptual judgments are
enhanced by the combination of disparity and shading
cues (Lovell et al., 2012; Schiller et al., 2011; Vuong
et al., 2006; Doorschot et al., 2001; Buelthoff & Mallot,
1988). However, between-participant variation in such
enhancement is difficult to assess given that low numbers
of participants were used (mean per study = 3.6, max =
5), a sizeable proportion of whom were not naive to the
purposes of the study. Here we find evidence for inte-
gration in both authors H.B. and A.W., but considerable
variability among the naive participants. In common with
Wagemans et al. (2010), this suggests that interobserver
variability may be significant in the interpretation of
shading patterns in particular and integration more gen-
erally, providing a stimulus for future work to explain the
basis for such differences.
Responses in Other ROIs
When presenting the results for all the participants, we
noted that performance in the disparity + shading con-
dition was statistically higher than for the component
cues in area V2 as well as in V3B/KO (Figure 3). Our sub-
sequent analyses did not provide evidence that V2 is a
likely substrate for the integration of disparity and shad-
ing cues. However, it is possible that the increased de-
coding performance—around the level expected by
quadratic summation—is due to parallel representations
of disparity and shading information. It is unlikely that
either signal is fully elaborated, but V2ʼs more spatially
extensive receptive fields may provide important infor-
mation about luminance and contrast variations across
the scene that provide signals important when interpret-
ing shape from shading (Schofield et al., 2010).
Previous work (Georgieva et al., 2008) suggested that
the processing of 3-D structure from shading is primarily
restricted in its representation to a ventral locus near the
area we localize as LO (although Gerardin, Kourtzi, &
Mamassian, 2010 suggested V3B/ KO is also involved
and Taira, Nose, Inoue, & Tsutsui, 2001 reported wide-
spread responses). Our fMRI data supported only weak
decoding of depth configurations defined by shading in
LO, and more generally across higher portions of both
the dorsal and ventral visual streams (Figures 3 and 4).
Indeed, the highest prediction performance of the MVPA
classifier for shading (relative to overall decoding accura-
cies in each ROI) was observed in V1 and V2, which is
likely to reflect low-level image differences between stim-
ulus configurations rather than an estimate of shape from
shading per se. Nevertheless, our findings from V3B/KO
Dövencioğlu et al.
1539
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
make it clear that information provided by shading
contributes to fMRI responses in higher portions of the
dorsal stream. Why then is performance in the “shading”
condition so low? Our experimental stimuli purposefully
provoked conflicts between the disparity and shading
information in the “single cue” conditions. Therefore,
the conflicting information from disparity that the viewed
surface was flat is likely to have attenuated fMRI re-
sponses to the “shading alone” stimulus. Indeed, given
that sensitivity to disparity differences was so much great-
er than for shading, it might appear surprising that we
could decode shading information at all. Previously, we
used mathematical simulations to suggest that area
V3B/KO contains a mixed population of responses, with
some units responding to individual cues and others fus-
ing cues into a single representation (Ban et al., 2012).
Thus, residual fMRI decoding performance for the shad-
ing condition may reflect responses to nonintegrated
processing of the shading aspects of the stimuli. This
mixed population could help support a robust perceptual
interpretation of stimuli that contain significant cue con-
flicts: for example, the reader should still be able to gain
an impression of the 3-D structure of the shaded stimuli
in Figure 1, despite conflicts with disparity).
In summary, previous fMRI studies suggest a number
of locations in which 3-D shape information might be
processed (Nelissen et al., 2009; Sereno et al., 2002).
Here we provide evidence that area V3B/KO plays an
important role in integrating disparity and shading cues,
compatible with the notion that it represents 3-D struc-
ture from different signals (Tyler et al., 2006) that are sub-
ject to different prior constraints (Preston et al., 2009). Our
results suggest that V3B/KO is involved in 3-D estimation
from qualitatively different depth cues, and its activity
may underlie perceptual judgments of depth.
Acknowledgments
The work was supported by the Japan Society for the Promotion
of Science (H22,290), the Wellcome Trust (095183/Z/10/Z), the
EPSRC (EP/F026269/1), and the Birmingham University Imaging
Centre.
Reprint requests should be sent to Andrew E. Welchman,
School of Psychology, University of Birmingham, Edgbaston,
Birmingham, B15 2TT, UK, or via e-mail: A.E.Welchman@
bham.ac.uk.
REFERENCES
Anzai, A., Ohzawa, I., & Freeman, R. D. (2001). Joint-encoding of
motion and depth by visual cortical neurons: Neural basis
of the Pulfrich effect. Nature Neuroscience, 4, 513–518.
Atkins, J. E., Fiser, J., & Jacobs, R. A. (2001). Experience-
dependent visual cue integration based on consistencies
between visual and haptic percepts. Vision Research, 41,
449–461.
Ban, H., Preston, T. J., Meeson, A., & Welchman, A. E. (2012).
The integration of motion and disparity cues to depth in
dorsal visual cortex. Nature Neuroscience, 15, 636–643.
Belhumeur, P. N., Kriegman, D. J., & Yuille, A. L. (1999).
The bas-relief ambiguity. International Journal of Computer
Vision, 35, 33–44.
Blake, A., Zisserman, A., & Knowles, G. (1985). Surface
descriptions from stereo and shading. Image and Vision
Computing, 3, 183–191.
Bradley, D. C., Qian, N., & Andersen, R. A. (1995). Integration of
motion and stereopsis in middle temporal cortical area of
macaques. Nature, 373, 609–611.
Buelthoff, H. H., & Mallot, H. A. (1988). Integration of depth
modules - Stereo and shading. Journal of the Optical Society
of America: A, 5, 1749–1758.
Burge, J., Fowlkes, C. C., & Banks, M. S. (2010). Natural-scene
statistics predict how the figure-ground cue of convexity
affects human depth perception. Journal of Neuroscience,
30, 7269–7280.
Curran, W., & Johnston, A. (1996). The effect of illuminant
position on perceived curvature. Vision Research, 36,
1399–1410.
De Martino, F., Valente, G., Staeren, N., Ashburner, J.,
Goebel, R., & Formisano, E. (2008). Combining multivariate
voxel selection and support vector machines for mapping
and classification of fMRI spatial patterns. Neuroimage, 43,
44–58.
DeAngelis, G. C., & Uka, T. (2003). Coding of horizontal
disparity and velocity by MT neurons in the alert macaque.
Journal of Neurophysiology, 89, 1094–1111.
DeYoe, E. A., Carman, G. J., Bandettini, P., Glickman, S., Wieser, J.,
Cox, R., et al. (1996). Mapping striate and extrastriate visual
areas in human cerebral cortex. Proceedings of the National
Academy of Sciences, U.S.A., 93, 2382–2386.
Doorschot, P. C. A., Kappers, A. M. L., & Koenderink, J. J.
(2001). The combined influence of binocular disparity and
shading on pictorial shape. Perception & Psychophysics,
63, 1038–1047.
Dosher, B. A., Sperling, G., & Wurst, S. A. (1986). Tradeoffs
between stereopsis and proximity luminance covariance as
determinants of perceived 3-D structure. Vision Research,
26, 973–990.
Dupont, P., De Bruyn, B., Vandenberghe, R., Rosier, A. M.,
Michiels, J., Marchal, G., et al. (1997). The kinetic occipital
region in human visual cortex. Cerebral Cortex, 7,
283–292.
Ernst, M. O. (2007). Learning to integrate arbitrary signals from
vision and touch. Journal of Vision, 7, 1–14.
Fleming, R. W., Dror, R. O., & Adelson, E. H. (2003). Real-world
illumination and the perception of surface reflectance
properties. Journal of Vision, 3, 347–368.
Georgieva, S. S., Todd, J. T., Peeters, R., & Orban, G. A. (2008).
The extraction of 3-D shape from texture and shading in the
human brain. Cerebral Cortex, 18, 2416–2438.
Gerardin, P., Kourtzi, Z., & Mamassian, P. (2010). Prior
knowledge of illumination for 3-D perception in the human
brain. Proceedings of the National Academy of Sciences,
U.S.A., 107, 16309–16314.
Gori, M., Del Viva, M., Sandini, G., & Burr, D. C. (2008). Young
children do not integrate visual and haptic form information.
Current Biology, 18, 694–698.
Grill-Spector, K., Kushnir, T., Hendler, T., & Malach, R. (2000).
The dynamics of object-selective activation correlate with
recognition performance in humans. Nature Neuroscience,
3, 837–843.
Hillis, J. M., Ernst, M. O., Banks, M. S., & Landy, M. S. (2002).
Combining sensory information: Mandatory fusion within,
but not between, senses. Science, 298, 1627–1630.
Horn, B. K. P. (1975). Obtaining shape from shading
information. In P. H. Winston (Ed.), The psychology of
computer vision (pp. 115–155). New York: McGraw Hill.
1540
Journal of Cognitive Neuroscience
Volume 25, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Janssen, P., Vogels, R., & Orban, G. A. (2000). Three-
dimensional shape coding in inferior temporal cortex.
Neuron, 27, 385–397.
Kersten, D., Mamassian, P., & Yuille, A. (2004). Object perception
as Bayesian inference. Annual Review of Psychology, 55,
271–304.
Kingdom, F. A. A. (2003). Color brings relief to human vision.
Nature Neuroscience, 6, 641–644.
Knill, D. C., & Saunders, J. A. (2003). Do humans optimally
integrate stereo and texture information for judgments of
surface slant? Vision Research, 43, 2539–2558.
Koenderink, J. J., & van Doorn, A. J. (2003). Shape and shading.
In L. M. Chalupa & J. S. Werner (Eds.), The visual
neurosciences (pp. 1090–1105). Cambridge, MA: MIT Press.
Kourtzi, Z., & Kanwisher, N. (2001). Representation of
perceived object shape by the human lateral occipital
complex. Science, 293, 1506–1509.
Kriegeskorte, N., Goebel, R., & Bandettini, P. (2006).
Information-based functional brain mapping. Proceedings of
the National Academy of Sciences, U.S.A., 103, 3863–3868.
Landy, M. S., Maloney, L. T., Johnston, E. B., & Young, M. (1995).
Measurement and modeling of depth cue combination - In
defense of weak fusion. Vision Research, 35, 389–412.
Larsson, J., Heeger, D. J., & Landy, M. S. (2010). Orientation
selectivity of motion-boundary responses in human visual
cortex. Journal of Neurophysiology, 104, 2940–2950.
Lee, Y. L., & Saunders, J. A. (2011). Stereo improves 3-D shape
discrimination even when rich monocular shape cues are
available. Journal of Vision, 11, Article 2.
Liu, B., & Todd, J. T. (2004). Perceptual biases in the
interpretation of 3-D shape from shading. Vision Research,
44, 2135–2145.
Lovell, P. G., Bloj, M., & Harris, J. M. (2012). Optimal integration
of shading and binocular disparity for depth perception.
Journal of Vision, 12, 1–18.
Mamassian, P., & Goutcher, R. (2001). Prior knowledge on the
illumination position. Cognition, 81, B1–B9.
Mingolla, E., & Todd, J. T. (1986). Perception of solid shape
from shading. Biological Cybernetics, 53, 137–151.
Nandy, A. S., & Tjan, B. S. (2008). Efficient integration across
spatial frequencies for letter identification in foveal and
peripheral vision. Journal of Vision, 8, 1–20.
Nardini, M., Bedford, R., & Mareschal, D. (2010). Fusion of
visual cues is not mandatory in children. Proceedings of the
National Academy of Sciences, U.S.A., 107, 17041–17046.
Nardini, M., Jones, P., Bedford, R., & Braddick, O. (2008).
Development of cue integration in human navigation.
Current Biology, 18, 689–693.
Nelissen, K., Joly, O., Durand, J. B., Todd, J. T., Vanduffel, W.,
& Orban, G. A. (2009). The extraction of depth structure
from shading and texture in the macaque brain. Plos One,
4, e8306.
Pentland, A. P. (1982). Finding the illuminant direction. Journal
of the Optical Society of America, 72, 448–455.
Popple, A. V., Smallman, H. S., & Findlay, J. M. (1998). The area
of spatial integration for initial horizontal disparity vergence.
Vision Research, 38, 319–326.
Preston, T. J., Kourtzi, Z., & Welchman, A. E. (2009). Adaptive
estimation of three-dimensional structure in the human
brain. Journal of Neuroscience, 29, 1688–1698.
Preston, T. J., Li, S., Kourtzi, Z., & Welchman, A. E. (2008).
Multivoxel pattern selectivity for perceptually relevant
binocular disparities in the human brain. Journal of
Neuroscience, 28, 11315–11327.
Ramachandran, V. S. (1988). Perception of shape from shading.
Nature, 331, 163–166.
Richards, W. (1985). Structure from stereo and motion. Journal
of the Optical Society of America: A, 2, 343–349.
Schiller, P. H., Slocum, W. M., Jao, B., & Weiner, V. S. (2011).
The integration of disparity, shading and motion parallax
cues for depth perception in humans and monkeys. Brain
Research, 1377, 67–77.
Schofield, A. J., Rock, P. B., & Georgeson, M. A. (2011). Sun and
sky: Does human vision assume a mixture of point and
diffuse illumination when interpreting shape-from-shading?
Vision Research, 51, 2317–2330.
Schofield, A. J., Rock, P. B., Sun, P., Jiang, X. Y., & Georgeson,
M. A. (2010). What is second-order vision for? Discriminating
illumination versus material changes. Journal of Vision, 10.
Serences, J. T., & Boynton, G. M. (2007). The representation of
behavioral choice for motion in human visual cortex. Journal
of Neuroscience, 27, 12893–12899.
Sereno, M. E., Trinath, T., Augath, M., & Logothetis, N. K.
(2002). Three-dimensional shape representation in monkey
cortex. Neuron, 33, 635–652.
Sereno, M. I., Dale, A. M., Reppas, J. B., Kwong, K. K.,
Belliveau, J. W., Brady, T. J., et al. (1995). Borders of
multiple visual areas in humans revealed by functional
magnetic resonance imaging. Science, 268, 889–893.
Srivastava, S., Orban, G. A., De Maziere, P. A., & Janssen, P.
(2009). A distinct representation of three-dimensional shape
in macaque anterior intraparietal area: Fast, metric, and
coarse. Journal of Neuroscience, 29, 10613–10626.
Sun, J., & Perona, P. (1998). Where is the sun? Nature
Neuroscience, 1, 183–184.
Taira, M., Nose, I., Inoue, K., & Tsutsui, K. (2001). Cortical areas
related to attention to 3-D surface structures based on
shading: An fMRI study. Neuroimage, 14, 959–966.
Thomas, R., Nardini, M., & Mareschal, D. (2010). Interactions
between “light-from-above” and convexity priors in visual
development. Journal of Vision, 10, 6.
Thompson, W. B., Fleming, R. W., Creem-Regehr, S. H., &
Stefanucci, J. K. (2011). Illumination, shading and shadows.
Chapter 9 of Visual perception from a computer graphics
perspective. Boca Raton, FL: Taylor and Francis.
Tootell, R. B. H., & Hadjikhani, N. (2001). Where is “dorsal V4”
in human visual cortex? Retinotopic, topographic and
functional evidence. Cerebral Cortex, 11, 298–311.
Tootell, R. B., Hadjikhani, N., Hall, E. K., Marrett, S.,
Vanduffel, W., Vaughan, J. T., et al. (1998). The retinotopy
of visual spatial attention. Neuron, 21, 1409–1422.
Tyler, C. W., Likova, L. T., Chen, C.-C., Kontsevich, L. L., &
Wade, A. R. (2005). Extended concepts of occipital
retinotopy. Current Medical Imaging Reviews, 1, 319–329.
Tyler, C. W., Likova, L. T., Kontsevich, L. L., & Wade, A. R.
(2006). The specificity of cortical region KO to depth
structure. Neuroimage, 30, 228–238.
Vuong, Q. C., Domini, F., & Caudek, C. (2006). Disparity and
shading cues cooperate for surface interpolation. Perception,
35, 145–155.
Wagemans, J., van Doorn, A. J., & Koenderink, J. J. (2010). The
shading cue in context. i-Perception, 1, 159–177.
Zeki, S., Perry, R. J., & Bartels, A. (2003). The processing of
kinetic contours in the brain. Cerebral Cortex, 13, 189–202.
Zeki, S., Watson, J. D., Lueck, C. J., Friston, K. J., Kennard, C., &
Frackowiak, R. S. (1991). A direct demonstration of functional
specialization in human visual cortex. Journal of
Neuroscience, 11, 641–649.
Dövencioğlu et al.
1541
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
9
2
5
1
/
5
9
2
/
7
1
1
5
9
2
4
7
5
/
6
1
3
7
8
7
o
9
c
5
n
6
_
4
a
/
_
j
0
o
0
c
4
n
1
7
_
a
p
_
d
0
0
b
4
y
1
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1