Perceptual Grouping Reveals Distinct Roles for Sustained
Slow Wave Activity and Alpha Oscillations in
Working Memory
Gisella K. Diaz , Edward K. Vogel, and Edward Awh
Abstrakt
■ Multiple neural signals have been found to track the num-
ber of items stored in working memory ( WM). These signals
include oscillatory activity in the alpha band and slow-wave
components in human EEG, both of which vary with storage
loads and predict individual differences in WM capacity.
Jedoch, recent evidence suggests that these two signals play
distinct roles in spatial attention and item-based storage in WM.
Hier, we examine the hypothesis that sustained negative volt-
age deflections over parieto-occipital electrodes reflect the
number of individuated items in WM, whereas oscillatory activ-
ity in the alpha frequency band (8–12 Hz) within the same elec-
trodes tracks the attended positions in the visual display. Wir
measured EEG activity while participants stored the orientation
of visual elements that were either grouped by collinearity or
nicht. This grouping manipulation altered the number of individ-
uated items perceived while holding constant the number of
locations occupied by visual stimuli. The negative slow wave
tracked the number of items stored and was reduced in ampli-
tude in the grouped condition. Im Gegensatz, oscillatory activity
in the alpha frequency band tracked the number of positions
occupied by the memoranda and was unaffected by perceptual
grouping. Perceptual grouping, Dann, reduced the number of
individuated representations stored in WM as reflected by
the negative slow wave, whereas the location of each element
was actively maintained as indicated by alpha power. Diese
findings contribute to the emerging idea that distinct classes
of EEG signals work in concert to successfully maintain on-
line representations in WM. ■
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
J
Ö
C
N
_
A
_
0
1
7
1
9
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
EINFÜHRUNG
Visual working memory (VWM) is an on-line memory sys-
tem that enables the rapid access and updating of infor-
mation in the service of other cognitive tasks. Capacity
limits in working memory ( WM) exhibit robust correla-
tions with broad measures of intellectual ability ( Vogel
& Awh, 2008), suggesting that WM is integral to complex
Erkenntnis. Daher, there has been strong interest in delin-
eating the behavioral and neural processes that deter-
mine these capacity limits. Multiple neural correlates
have been found to monotonically scale with the number
of items in visual WM up to measured storage limits (ap-
proximately 3 Artikel), showing sustained activity over the
delay period (Fukuda, Mance, & Vogel, 2015; Todd &
Marois, 2004; Vogel & Machizawa, 2004). Darüber hinaus, diese
signals predict individual differences in visual WM capac-
ität (Luria, Balaban, Awh, & Vogel, 2016; Fukuda et al.,
2015; Vogel & Machizawa, 2004). Using scalp-EEG and
the ERP technique, past work has routinely found that
a contralateral and sustained negative voltage deflection,
or the contralateral delay activity (CDA), shows reliable
decreases in amplitude as items are added into WM
(Luria et al., 2016; Vogel & Machizawa, 2004). Ähnlich,
oscillatory activity in the alpha band has also been shown
The University of Chicago
© 2021 Massachusetts Institute of Technology
to track the number of items in WM (Sauseng et al., 2009;
Busch & Herrmann, 2003). Previous work often focused
on lateralized signals that required lateralized task de-
signs and distractors to balance visual stimulation for
both hemispheres. More recently, Fukuda et al. (2015)
used a whole-field design that did not require distractors
to examine whether whole-field signals were also sensi-
tive to the number of items in WM. They found that
parieto-occipital alpha power (7–9 Hz) and a sustained
negative voltage deflection over parieto-occipital elec-
trodes both tracked the number of items in WM.
In der Tat, it has been suggested that these two signals
are manifestations of the same neural process, such that
amplitude modulations of oscillatory activity can explain
the generation of slow evoked components, wie
CDA (van Dijk, van der Werf, Mazaheri, Medendorp, &
Jensen, 2010; Mazaheri & Jensen, 2008). Tatsächlich, latera-
lized sustained event-related fields in magnetoencepha-
lography and lateralized alpha power were found to be
strongly correlated, both spatially and temporally, in einem
WM task (van Dijk et al., 2010). Jedoch, the whole-field
signals in Fukuda et al. (2015) showed distinct time
courses and explained unique variance in predicting in-
dividual differences in WM capacity, suggesting that sus-
tained potentials and alpha-band oscillations reflect
distinct aspects of storage in visual WM ( Wang, Megla,
Zeitschrift für kognitive Neurowissenschaften 33:7, S. 1354–1364
https://doi.org/10.1162/jocn_a_01719
& Woodman, 2021; Hakim, Adam, Gunseli, Awh, &
Vogel, 2019; Wang, Rajsic, & Woodman, 2019; Bae &
Luck, 2018).
Hier, we examine the hypothesis that each signal pro-
vides unique information about the contents of visual
WM. Speziell, the negative slow wave can track the
number of individuated items in WM, whereas parieto-
occipital alpha power tracks the number of relevant
locations in WM. The distinction between items and
locations is a subtle distinction that has been examined
before using the CDA, which was found to be sensitive
to the number of items in WM regardless of the number
of locations (Ikkai, McCollough, & Vogel, 2010). Auf der
andererseits, oscillatory activity in the alpha frequency band
is known to track the deployment of spatial attention during
storage in visual WM (Foster, Sutterer, Serences, Vogel, &
Awh, 2016; Rihs, Michel, & Thut, 2007), but extant work
has not discriminated between item-based and location-
based explanations of this oscillatory activity. Nach
to our hypothesis, parieto-occipital alpha power will be
primarily sensitive to the number of attended locations
within the visual display rather than the number of individ-
uated items.
We used a perceptual grouping manipulation to dis-
criminate between item-based and location-based neural
Aktivität. Past work has shown that perceptual grouping
can yield strong improvements in WM performance such
that a larger number of elements can be remembered
when they are grouped. This effect has been observed
using a variety of grouping cues, including proximity
and connectedness cues (Xu, 2006; Jiang, Chun, &
Olson, 2004; Woodman, Vecera, & Luck, 2003), color
Ähnlichkeit (Morey, 2019; Gao, Gao, Tang, Shui, & Shen,
2016; Morey, Cong, Zheng, Price, & Morey, 2015; Brady
& Tenenbaum, 2013; Peterson & Berryhill, 2013; Shen,
Yu, Xu, & Gao, 2013; Quinlan & Cohen, 2012), shape
Ähnlichkeit (Mate & Baqués, 2009), amodal completion
( Walker & Davies, 2003), depth cues (Kristjánsson,
2006), and collinearity and closure cues (Gao et al.,
2016). Hier, we used collinearity cues to manipulate
the number of individuated items, while holding constant
the number of relevant locations. Daher, this manipulation
provided traction for distinguishing between item-based
and location-based neural signals.
In Experiment 1, we first establish that parieto-occipital
alpha power (8–12 Hz) and the negative slow wave track
the number of items stored with both color and spatial
memoranda, replicating the findings of Fukuda et al.
(2015) and also extending them to a spatial WM task.
Our primary motivation for Experiment 1 was to ensure
that both signals, especially parieto-occipital alpha power,
were not disrupted by a spatial task. This anticipatory
analysis was done to ensure a fair comparison between
the two signals in the second experiment, bei dem die
number of items was manipulated, but the number of spa-
tial locations was not. Speziell, we use collinearity cues
to perceptually group elements in Experiment 2, while
holding constant the number of elements in the display.
Pairs of stimuli were either aligned to create the percept
of a single item or misaligned to create the percept of two
Artikel. To anticipate the results, alpha power was sensitive
to the number of locations that were attended regardless
of whether the items were grouped or not, whereas the
negative slow wave tracked the number of individuated
items stored, revealing a smaller number of items with
grouping by collinearity. These findings provide further
evidence for the distinction between spatial and item-
based signals in visual WM.
METHODEN
Teilnehmer
Participants were recruited from the University of
Chicago and the surrounding community. Gesamt, 16
(six women, Durchschnittsalter = 22 Jahre) Und 23 (14 Frauen, mean
age = 22 Jahre) participants were run in Experiments 1
Und 2, jeweils. For Experiment 1, data from two
participants were excluded because of the participants’
voluntary withdrawal during the experiment session. In
addition, data from two participants in Experiment 1 Und
three participants in Experiment 2 were excluded because
of excessive EEG artifacts (< 150 trials remaining per
condition).
Experimental procedures were approved by the insti-
tutional review board at the University of Chicago. All
participants gave informed consent and were compen-
sated for their participation at a rate of $15 per hour.
Participants reported normal color vision and normal or
corrected-to-normal visual acuity.
For Experiment 1, our intended sample size was 16
participants given previous research showing that this is
a sufficient number of participants to observe the time
course of set size effects on both the negative slow wave
and parieto-occipital alpha power (Fukuda et al., 2015).
However, data from four of our 16 participants were ex-
cluded from analyses given insufficient number of trials
after artifact rejection or because of participants’ volun-
tary withdrawal from the experiment session. For
Experiment 2, our intended sample size was 20 partici-
pants given that our aim was to go one step further
and investigate whether our set size effects were sensitive
to perceptual grouping. Data from three of our initial 20
participants were excluded from our analyses after arti-
fact rejection. We replaced these three participants and
collected data from an additional three participants to
complete our intended sample size.
Apparatus
Participants were tested in a dimly lit, electrically shielded
chamber. Stimuli were generated using MATLAB (The
Mathworks) and the Psychophysics Toolbox (Brainard,
1997; Pelli, 1997). Stimuli were presented on a 24-in.
Diaz, Vogel, and Awh
1355
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
LCD monitor (refresh rate: 120 Hz, resolution: 1080 ×
1920 pixels) at a viewing distance of approximately 75 cm
and against a dark gray background.
EEG Acquisition
We recorded EEG activity using 30 active Ag/AgCl elec-
trodes mounted in an elastic cap (Brain Products
actiCHamp). We recorded from International 10–20 sites
Fp1, Fp2, F7, F3, Fz, F4, F8, FC5, FC1, FC2, FC6, C3, Cz,
C4, CP5, CP1, CP2, CP6, P7, P3, Pz, P4, P8, PO7, PO3,
PO4, PO8, O1, Oz, and O2. Two additional electrodes
were placed on the left and right mastoids, and a ground
electrode was placed at position Fpz. All sites were re-
corded with a right-mastoid reference and were rerefer-
enced off-line to the algebraic average of the left and
right mastoids. We recorded EOG using passive elec-
trodes with a ground electrode placed on the left cheek.
HEOG was recorded with a bipolar pair of electrodes
placed ∼1 cm from the external canthus of each eye,
and vertical EOG with a bipolar pair of electrodes placed
above and below the right eye. Data were filtered on-line
(low cutoff = 0.01 Hz, high cutoff = 80 Hz, slope from
low-to-high cutoff = 12 dB/octave) and were digitized at
500 Hz using BrainVision Recorder (Brain Products) run-
ning on a PC. During preparation, impedances were set
to be below 10 kΩ.
Eyetracking
We recorded gaze position using a desk-mounted infrared
eye-tracking system (EyeLink 1000 Plus, SR Research).
Gaze position was sampled at 1000 Hz. Stable head posi-
tion was maintained during the task using a chin rest.
The eye tracker was recalibrated as needed throughout
the session, including whenever participants removed
their chin from the chin rest.
Artifact Rejection
For artifact rejection, each trial was segmented into
−400 msec pretrial and 1750 msec poststimulus array
onset epochs. We used an automated procedure to flag
trials that were contaminated by ocular or EEG artifacts.
Next, we used this procedure as a guideline during manual
visual inspection where it was ultimately determined
which trials were to be rejected. Experimenters were blind
to condition when inspecting the data for artifacts. Trials
contaminated by artifacts were excluded from EEG analy-
ses but not from behavioral analyses. Participants were
excluded from the final sample if they had fewer than
150 artifact-free trials per condition.
An automated artifact detection procedure was used to
detect eye movements, blinks, and EEG artifacts. Trials
were flagged as containing a saccade if the Euclidean vec-
tor between the mean gaze positions in the first and sec-
ond halves of an 80-msec sliding window (advanced in
10-msec increments) was greater than 0.5° of visual an-
gle. When eye tracking data were not available, we used
HEOG to detect saccades. Trials were flagged as contain-
ing a saccade if the mean voltage during the first and sec-
ond halves of a 150-msec sliding window (advanced in
10-msec steps) exceeded 20 μV.
For blinks, trials were flagged as containing a blink if
the eye tracker could not detect the pupil at any point
during the trial. When eye tracking data were not avail-
able, we used vertical EOG to detect blinks. Trials were
flagged as containing a blink if the mean voltage during
the first and second halves of a 150-msec sliding window
(advanced in 10-msec steps) exceeded 30 μV.
For EEG artifacts, we flagged trials as containing voltage
drifts (e.g., skin potentials) if the absolute change in volt-
age from the first quarter of the trial to the last quarter of
the trial exceeded 100 μV. We flagged trials as including a
sudden step in voltage (which can occur when an elec-
trode is damaged) if the mean voltage during the first
and second halves of a 250-msec sliding window (ad-
vanced in 20-msec increments) differed by more than
100 μV. We marked trials as containing high-frequency
noise (e.g., muscle artifacts) if any electrode had a
peak-to-peak amplitude greater than 150 μV within a
15-msec sliding window (advanced in 50-msec incre-
ments). Finally, we flagged trials as containing amplifier
saturation if any electrode had 60 time points within a
200-msec sliding window (advanced in 50-msec incre-
ments) that were within 1 μV of each other.
Negative Slow Wave Analysis
EEG activity was calculated using a baseline from
−400 msec to 0 msec relative to the onset of the stimulus
array. The mean baseline amplitude was subtracted from
EEG amplitude at each time point in the trial. The base-
lined trials were then averaged for each condition to
create ERPs for each condition. We included 12 parieto-
occipital electrodes chosen a priori and based on previ-
ous findings: P7, P3, Pz, P4, P8, PO7, PO3, PO4, PO8, O1,
Oz, and O2. Statistical analyses were performed on data
that were not filtered beyond the .01- to 80-Hz on-line
data-acquisition filter. We low-pass filtered the data (30 Hz)
for illustrative purposes in the figures.
Parieto-Occipital Alpha Power Analysis
EEG signal processing was performed in MATLAB. We
band-pass filtered the raw EEG data using a filter from
the FieldTrip toolbox (ft_preproc_bandpassfilter.m;
Oostenveld, Fries, Maris, & Schoffelen, 2011) and then
extracted instantaneous power values for the alpha band
(8–12 Hz) by applying a Hilbert transform (hilbert.m) to
the filtered data. We calculated alpha power for the same
parieto-occipital electrodes as in the ERP analysis: P7, P3,
Pz, P4, P8, PO7, PO3, PO4, PO8, O1, Oz, and O2. For
illustrative purpose in the figures, we subtracted the mean
1356
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
baseline (−400 to 0 msec) at each time point in the trial
for each condition and converted to percent change from
baseline.
Stimuli
For both experiments, a black fixation dot (diameter =
0.20°) was presented at the center of a dark gray back-
ground and remained visible for the entire trial. The stimuli
were presented within a predetermined area (Experiment 1:
19.65° × 14.90°; exp. 2: 15.80° × 15.80°) and at least 0.75°
(exp. 1) or 3° (exp. 2) away from fixation. During the de-
lay interval, only the fixation dot remained on the screen.
After the delay, a single probe stimulus reappeared.
For Experiment 1, the stimuli array consisted of either
one or three circles (0.75°) placed randomly within a
quadrant, without replacement and at least 2.25° away
from each other if Set Size 3. For the color change detec-
tion task, the circles were each randomly rendered in one
color from seven possible colors (red, green, blue, yel-
low, magenta, white, and black) without replacement.
For the spatial change detection task, the circles were
rendered in black. After the delay interval, a single probe
stimulus reappeared that was either the “same” as one of
the originally presented stimuli or “different” from any of
the originally presented stimuli. For the color task, a
change could occur in the color of the stimulus, whereas
a change in spatial location could occur in the spatial
task. Specifically, the change in spatial location could oc-
cur in any angular direction (0°–360°) and at a randomly
determined distance (range: 3°–3.75°) relative to the
original stimulus. The probe stimulus was constrained
to its original quadrant and had to maintain the same
minimum distance requirement (2.25°) from the original
stimuli locations.
For Experiment 2, the stimuli array consisted of either
two or four black circles (3°) with rectangular gaps. The
locations of the stimuli were assigned in a serial manner.
The location of the first stimulus was randomly selected
to fall within the predetermined bounds. In relation to
the first stimulus, the location of the second stimulus
could occur in a randomly determined angular direction
(0°–360°) and 5.65° away from the first stimulus, while
maintaining the requirements of the first stimulus (i.e.,
remaining within the predetermined bounds), and effec-
tively forming a pair. For Set Size 4, the third and fourth
stimuli locations were determined in a similar manner
with the additional constraints that stimuli be at least 6°
away from stimuli belonging to a different pair and that
pairs of stimuli did not intersect. On half of the trials, the
gaps of each pair of stimuli were oriented toward each
other to form the percept of either one (Set Size 2) or
two (Set Size 4) items (grouped condition). On the re-
maining trials, the gaps were misaligned (ungrouped con-
dition). The same location configurations were used
across conditions (grouped and ungrouped) but were
unique for each participant. To avoid accidental pairs in
the ungrouped condition, the orientation of the second
stimulus in each pair had to be more than 5° clockwise
or counterclockwise from the orientation it would be in
the grouped condition.
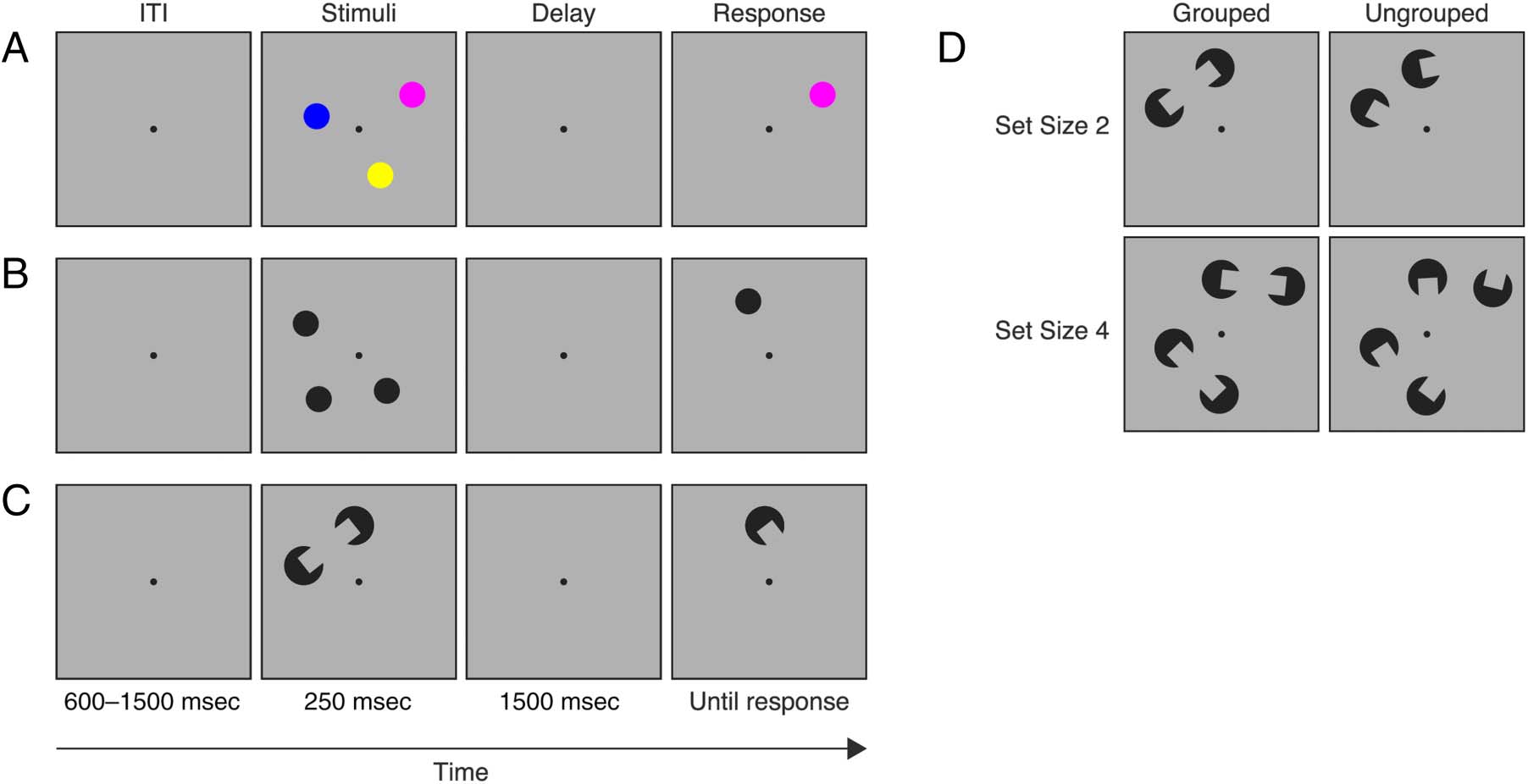
Experiment 1 Procedure
Participants performed a whole-field change detection
task (Figure 1A and 1B). The trial began with a fixation
dot presented at the center of the screen for a randomly
Figure 1. Color (A) and spatial (B) change detection tasks from Experiment 1. Perceptual grouping change detection task from Experiment 2 (C)
with example stimuli displays for each of the four conditions (D). ITI = intertrial interval.
Diaz, Vogel, and Awh
1357
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
determined duration between 600 and 1500 msec. A
stimuli array followed and was presented for 250 msec.
The stimuli array consisted of either one or three circles
that were either rendered in color (color change detection
task) or in black (spatial change detection task). Participants
were asked to remember as many of the stimuli as possible
over a 1500-msec blank delay interval. After the delay, a sin-
gle stimulus reappeared. Participants used a keyboard but-
ton press to indicate whether this probe stimulus was
presented at one of the previously occupied locations.
For the color task (Figure 1A), a change could occur in
the color of the stimulus, whereas a change in spatial loca-
tion could occur in the spatial task (Figure 1B). Participants
pressed the “z” or “/” key to indicate whether the probe
stimulus was the “same” or “different,” respectively, from
the stimuli display. There were no practice trials given be-
fore the formal experiment. Participants were given ver-
bal and written task instructions with the aid of an example
trial image similar to Figure 1A–C.
Within a block, half of the trials were “same” trials, and
the remaining half were “different” trials. Similarly, half of
the trials were Set Size 1, and the remaining half were Set
Size 3. Participants completed 20 blocks with each con-
taining 60 trials of either the color or spatial change detec-
tion task. There were an equal number of color and
spatial change detection blocks, which were completed
in an alternating order. The task for the first block was
counterbalanced across participants. Participants self-
initiated each block by pressing the spacebar key. The
experiment session was scheduled to take 3 hr, but the
actual duration of the session (∼2.5–3.5 hr) depended on
each participant’s pace because they initiated each block
and decided when (and if ) to take breaks between blocks.
Experiment 2 Procedure
The procedure was similar to Experiment 1 with the follow-
ing exceptions. The stimuli array consisted of either two or
four black circles with rectangular gaps (Figure 1C). On
half of the trials, the gaps of each pair of stimuli were ori-
ented toward each other to form the percept of either
one (Set Size 2) or two (Set Size 4) items. On the remain-
ing trials, the gaps were misaligned (Figure 1D). After the
delay, participants indicated whether the orientation of
the probe stimulus that reappeared was the “same” or
“different.” Half of the trials were Set Size 2, and the re-
maining half were Set Size 4. Participants completed 30
blocks of 40 trials each. The experiment session was
scheduled to take 3.5 hr (∼3–4 hr).
Experimental Design and Statistical Analysis
Both experiments used a 2 × 2 within-subject design. For
Experiment 1, the factors were set size (1 or 3) and task
type (color or spatial). The type of task alternated with
each block. For Experiment 2, the factors were set size
(2 or 4) and grouping condition (grouped or ungrouped).
Behavioral data were analyzed using a repeated-measures
ANOVA. Neural data were analyzed using repeated-
measures ANOVAs on averaged delay activity data and
cluster-based permutation tests (Sassenhagen & Draschkow,
2019; Maris & Oostenveld, 2007) on data averaged over
parieto-occipital electrodes. In Experiment 1, our focus was
on ANOVAs given that we were investigating robust set size
effects that had been observed before. In Experiment 2, we
focused on cluster-based permutation tests to search for
novel grouping effects in a data-driven way. For consis-
tency, we present results using both approaches for each
experiment. For the cluster-based permutation tests, we
first reduced our high-dimensional data (n time points ×
n participants; per condition) to a single value. Repeated-
measures t tests were calculated at each time point to as-
sess the difference between conditions (e.g., Set Size 1
vs. Set Size 3), which resulted in a map of t scores across
time. Time points were thresholded according to an a
priori defined criterion (which corresponded to a p value
of .05, two-sided), and adjacent time points with t scores
that exceeded this value were grouped together to form
a cluster. Clusters were summarized into a single number
by summing the t values, which produced a single value
for each cluster. It is important to note that the extent of
the cluster became fixed at the end of this first step and
individual time points were not visible to the next infer-
ence step. Specifically, the cluster structure was our only
test statistic and no statistical inference was made about
individual time points. Nevertheless, we next calculated
the probability that these values came from a null distri-
bution. Permutation tests were used to establish the
probability of our data under the null hypothesis given
that it is unclear what distribution of t value sums would
be expected under the null hypothesis. The number of
permutations was 10,000 or the maximum possible,
whichever was lowest. On each iteration and for each
time point, it was randomly determined if the first condi-
tion was subtracted from the second condition or vice
versa for each participant. Then for each iteration, the
cluster formation step was repeated. The cluster with
the highest sum of t values was identified, and the sum
of its t values was stored and became our surrogate-
null value for that permutation. After all iterations, the
cumulative density of these surrogate-null values was our
approximation of the values under the null hypothesis.
The p value then was calculated as the percentage of
surrogate-null values that the observed data exceeded.
RESULTS
Experiment 1
There was a main effect of Set Size, F(1, 11) = 17.99, p =
.001, η2 = .62, and Task, F(1, 11) = 8.18, p = .016, η2 =
.43, on accuracy, such that accuracy was higher for Set
Size 1 (M = 0.95, SD = 0.05) than Set Size 3 (M = 0.90,
1358
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
SD = 0.08) and for the color task (M = 0.94, SD = 0.05)
than the spatial task (M = 0.90, SD = 0.08). There was no
significant interaction between Set Size and Task on ac-
curacy, F(1, 11) = 0.05, p = .84, η2 = .004.
We used repeated-measures ANOVAs on averaged delay
activity from parieto-occipital electrodes to analyze the ef-
fects of Set Size and Task Type on both the negative slow
wave and parieto-occipital alpha power. Moreover, we
used nonparametric cluster-based permutation tests
(Maris & Oostenveld, 2007) to corroborate the effects of
Set Size and Task Type in a more nuanced manner.
Given previous research (Fukuda et al., 2015), we pre-
dicted that both signals would show characteristic set size
effects. Specifically, Fukuda et al. (2015) found a mono-
tonic increase in slow wave negativity and a monotonic
decrease in parieto-occipital alpha power with increases
in set size up to typical capacity (approximately 3 items).
The negative slow wave and parieto-occipital alpha
power were analyzed during the delay period (250–
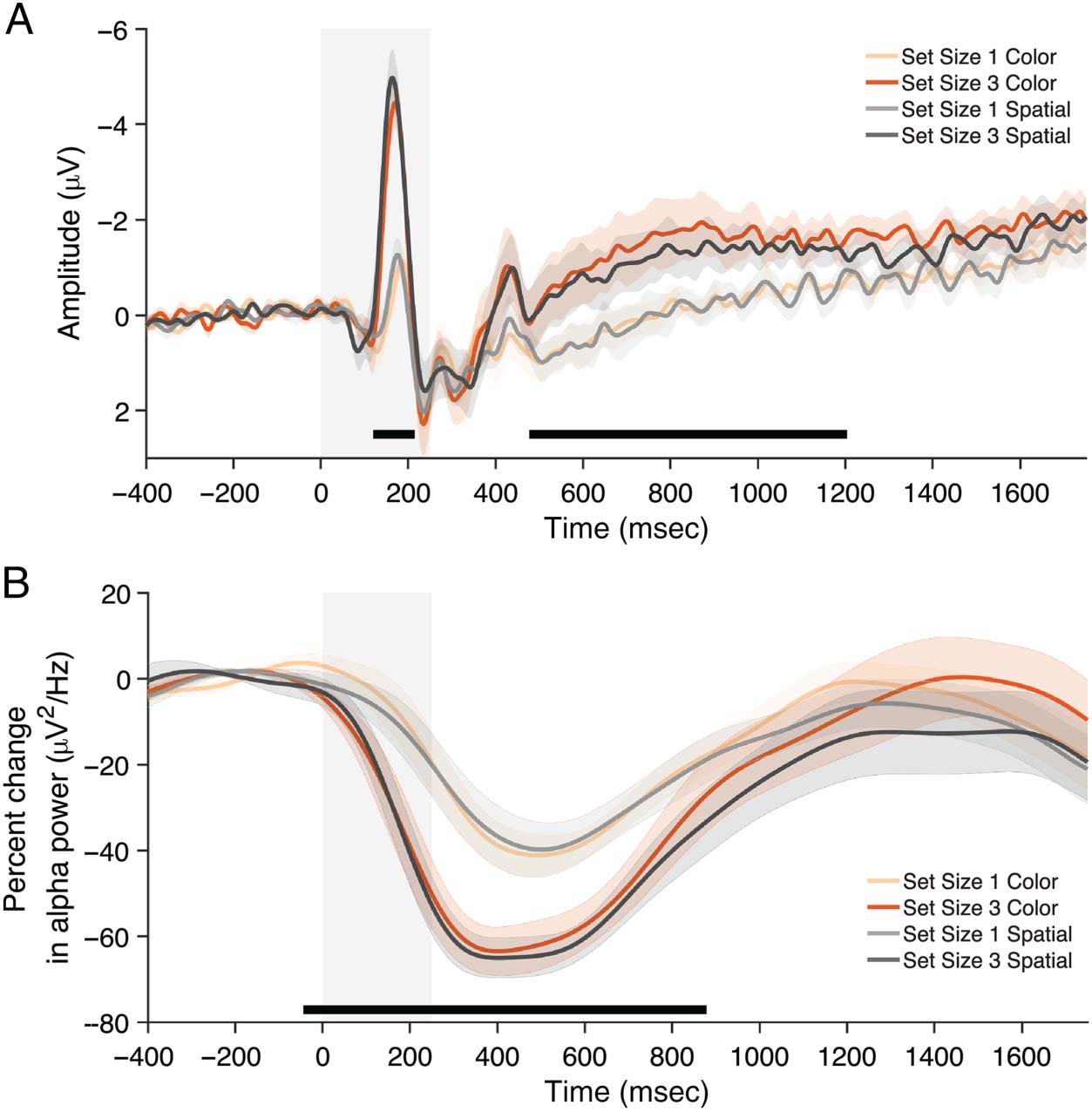
1750 msec). As predicted, there was a main effect of
Set Size on voltage at parieto-occipital electrodes
(Figure 2A; F(1, 11) = 16.99, p = .002, η2 = .61), such
that the amplitude was more negative for Set Size 3 than
Set Size 1. There was no main effect of Task, F(1, 11) =
0.47, p = .51, η2 = .04, and no significant interaction, F(1,
11) = 0.29, p = .60, η2 = .03. Cluster-based permutation
tests echoed these findings. Three clusters in the voltage
at parieto-occipital electrodes were found to be sensitive
to set size and extended from ∼130 to ∼210 msec, ∼490
to ∼1020 msec, ∼1020 to ∼1190 msec. The permutation
test indicated that the effects of Set Size were significant
( p = .048, p = .007, p = .044). There were no clusters
that were sensitive to Task Type. In short, we replicated
and extended previous findings that the negative slow
wave is sensitive to set size in a color change detection
task (Fukuda et al., 2015), as well as in a spatial change
detection task.
The main effect of Set Size on parieto-occipital alpha
power did not reach significance when averaging over
the entire delay interval (Figure 2B; F(1, 11) = 3.06, p =
.11, η2 = .22), although an early transient effect of Set Size
was evident with more alpha power suppression for Set
Size 3 than Set Size 1. There was a main effect of Task,
F(1, 11) = 6.83, p = .024, η2 = .38, which was driven by
the rapid return to baseline in the color task for Set Size 3
trials. Importantly, the interaction did not reach signifi-
cance, F(1, 11) = 1.03, p = .33, η2 = .09. Cluster-based per-
mutation tests revealed more nuanced results. There was a
cluster sensitive to set size that extended from ∼−30 to
∼870 msec. The permutation test indicated that there
was a significant effect of Set Size ( p = .006). There was
also a cluster that was sensitive to task, extending from
∼680 to ∼1560 msec. The permutation test indicated a sig-
nificant effect of task ( p = .003). Thus, we again replicated
Figure 2. Averaged negative
slow wave (A) and averaged
alpha power suppression (B)
observed at parieto-occipital
electrodes in Experiment 1.
Shaded regions indicate
duration of stimuli display.
Black bars indicate clusters
showing a significant
( p < .05) Set Size effect
from cluster-based
permutations tests.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Diaz, Vogel, and Awh
1359
previous findings from Fukuda et al. (2015) that parieto-
occipital alpha power shows a monotonic decrease with
an increase in set size for both a color and spatial change
detection task.
In summary, our findings from Experiment 1 replicate
those of Fukuda et al. (2015) and extend them to spatial
memoranda. For both color and spatial memoranda, both
the negative slow wave and parieto-occipital alpha power
are sensitive to set size. In Experiment 2, we investigate
whether these set size effects are further shaped by col-
linearity cues. Do grouping cues that compel the percep-
tion of fewer individuated objects also affect the
magnitude of these storage-related signals?
Experiment 2
There was a main effect of Set Size, F(1, 19) = 110.3, p <
.001, η2 = .85, and Grouping, F(1,19) = 146.7, p < .001,
η2 = .89, on accuracy, such that accuracy was higher for
Set Size 2 (M = 0.94, SD = 0.05) than Set Size 4 (M =
0.86, SD = 0.10) and for grouped trials (M = 0.95, SD =
0.05) compared to ungrouped trials (M = 0.85, SD = 0.10).
There was also a significant interaction between Set Size
and Grouping on accuracy, F(1,19) = 116.1, p < .001,
η2 = .86, such that there was a greater benefit of group-
ing for Set Size 4 than Set Size 2.
Nonparametric cluster-based permutation tests were
used to analyze the effects of set size and grouping on
both the negative slow wave and parieto-occipital alpha
power. In comparison to Experiment 1, the grouping ef-
fects of interest in Experiment 2 were more novel and the
timing and duration of any grouping effect was not
known. Accordingly, choosing an objective window of in-
terest for ANOVAs was not possible. Instead, we focused
on cluster-based permutation tests; however, we also re-
port the results of repeated-measures ANOVAs using
averaged delay activity from three equally sized windows
of 500 msec each (early, middle, and late delay).
Given previous research and our own data, we again
predicted that both signals would show characteristic
set size effects, with higher slow wave negativity and re-
duced alpha power for Set Size 4 compared to Set Size 2.
The central question of the study, however, was to inves-
tigate whether these signals were sensitive to grouping
when set size effects were present. To this end, we first
identified clusters that were sensitive to set size and then
examined whether those clusters were also sensitive to
grouping.
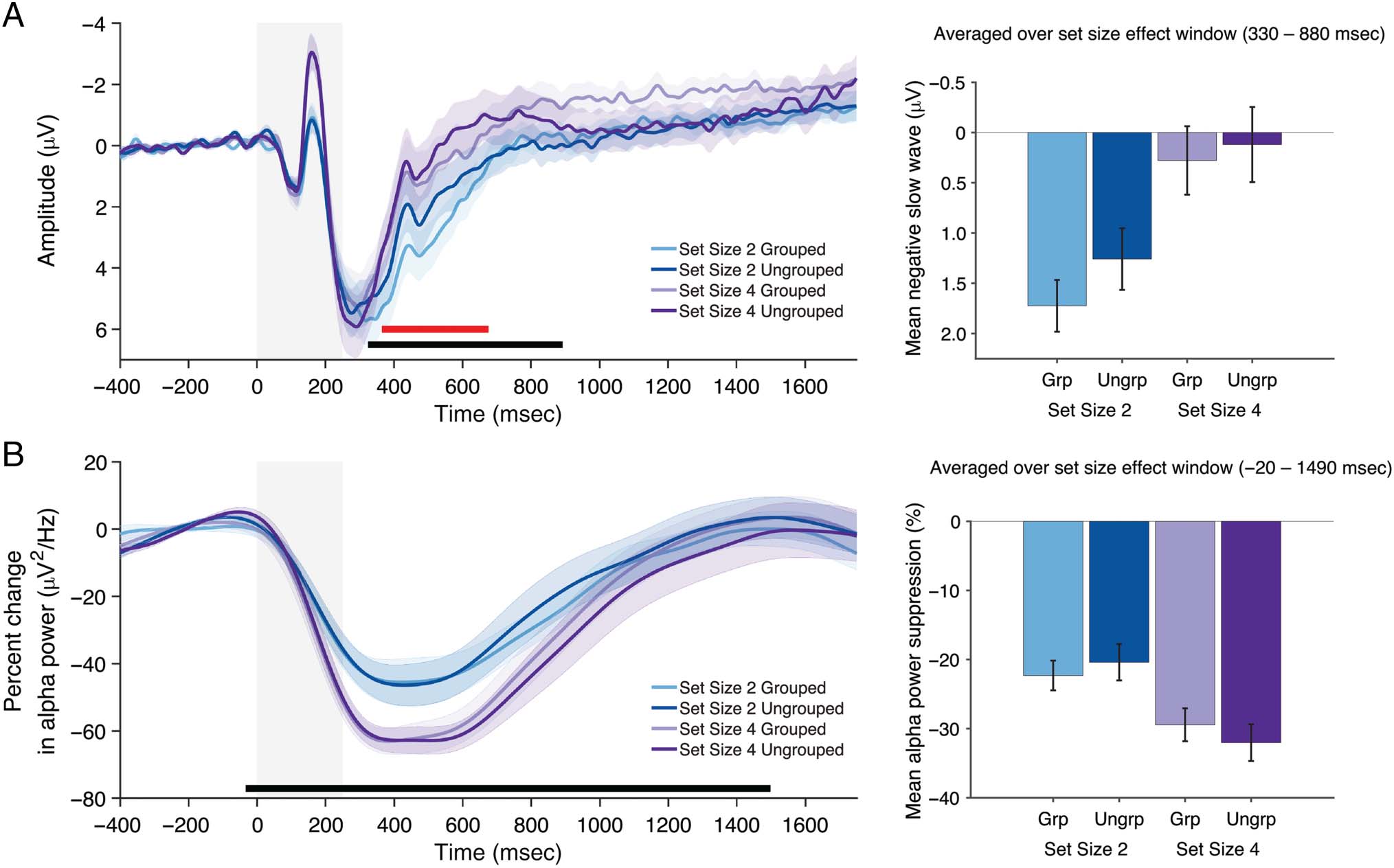
First, there was a main effect of Set Size and Grouping
on voltage at parieto-occipital electrodes during the early
time window (250–750 msec; Table 1). A cluster in the
voltage at parieto-occipital electrodes extended from
∼330 to ∼880 msec. The cluster-based permutation test
indicated that there was a significant effect of Set Size
(Figure 3A; p = .005), such that amplitude was more neg-
ative for Set Size 4 than Set Size 2. Within this cluster,
another cluster was identified that extended from ∼370
Table 1. Repeated-Measures ANOVAs for Neural Data in
Experiment 2
Terms
F-statistics df1 df2 p Value η2
19 <.001
.54
.32
.007
Negative Slow Wave
Early (250–750 msec)
Set Size (2, 4)
Condition (Ingrouped
to Ungrouped)
22.21
9.11
Set Size × Condition
3.07
Middle (750–1250 msec)
Set Size (2, 4)
Condition (Grouped,
Ungrouped)
Set Size × Condition
Late (1250–1750 msec)
Set Size (2, 4)
Condition (Grouped,
Ungrouped)
Set Size × Condition
Alpha Power
Early (250–750 msec)
Set Size (2, 4)
Condition (Grouped,
Ungrouped)
5.11
3.49
2.39
3.33
0.64
2.76
14.51
2.97
Set Size × Condition
0.08
Middle (750–1250 msec)
Set Size (2, 4)
Condition (Grouped,
Ungrouped)
10.32
3.01
Set Size × Condition
4.89
Late (1250–1750 msec)
Set Size (2, 4)
Condition (Grouped,
Ungrouped)
Set Size × Condition
2.61
0.58
3.84
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
19
19
19
19
19
19
19
19
19
19
19
19
19
19
19
19
19
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
.10
.14
.036
.077
.21
.16
.14
.11
.084
.43
.15
.03
.11
.13
.001
.10
.43
.14
.78
.00
.005
.10
.35
.14
.04
.21
.12
.46
.12
.03
.065
.17
We applied 2 (Set Size 2, 4) × 2 (Grouped, Ungrouped) repeated-
measures ANOVAs to voltage and alpha power at parieto-occipital elec-
trodes averaging over delay activity in three equally sized windows of
500 msec each. Significant ( p < .0167; Bonferroni-corrected for α =
.05) effects in bold.
1360
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Averaged negative slow wave (A) and averaged alpha power suppression (B) observed at parieto-occipital electrodes in Experiment 2.
Shaded regions indicate duration of stimuli display. Black and red bars indicate clusters showing a significant ( p < .05) set size or grouping effect,
respectively, from cluster-based permutation tests.
to ∼670 msec. The permutation test indicated that there
was a significant effect of Grouping ( p < .001), such that
amplitude was more negative when stimuli were un-
grouped relative to grouped. To assess the evidence for
grouping during this set size window, estimated Bayes
factors were calculated (BF01 = 0.89) and indicated that
the data were 1.13 times more likely to have occurred
under the alternative hypothesis.
Thus, we first replicated our finding from Experiment 1
that the negative slow wave was modulated by set size.
Moreover, we extended those findings by demonstrating
that the negative slow wave was also modulated by
grouping. The reduction in amplitude of the negative
slow wave for grouped stimuli is consistent with our hy-
pothesis that the negative slow wave tracks the number
of individuated items.
Halfway through the delay, we observed an unexpected
drop in amplitude in the negative slow wave for Set Size 4
Ungrouped, such that the set size effect was eliminated.
Although the mechanisms involved are unknown, it is pos-
sible that this pattern is because of the strategic reorient-
ing of attention to the remembered items within capacity.
In line with this speculation, we found that the drop in the
negative slow wave was restricted to participants with high
Set Size 4 Ungrouped performance, which is consistent
with the idea of strategic refocusing. Additional work is
needed to bolster the findings of this post hoc analysis.
In alpha power, there was a main effect of Set Size during
the early and middle time windows (250–750 msec, 750–
1250 msec; Table 1), but no main effect of Grouping. A
cluster extended from ∼−20 to ∼1490 msec with the per-
mutation test indicating that there was a significant effect
of Set Size (Figure 3B; p < .001), such that alpha power
was lower for Set Size 4 than Set Size 2. Despite this
robust effect of the number of positions, however, no
clusters showing this set size effect were sensitive to
the effects of perceptual grouping. Estimated Bayes fac-
tors during this set size window were calculated (BF01 =
1.51) and indicated that the data were 1.5 times more
likely to have occurred under the null hypothesis. This
suggests that participants maintained their attention on
the same number of positions, even though the number
of individuated items stored was reduced in the grouped
condition.
DISCUSSION
In line with previous research, perceptual grouping cues
enhanced VWM performance and also led to distinct
changes in EEG signals that track WM storage.
Diaz, Vogel, and Awh
1361
Specifically, the negative slow wave—a signal known to
track the number of items stored in visual WM (Fukuda
et al., 2015)—was reduced in amplitude for grouped stim-
uli, consistent with the idea that grouping reduces the num-
ber of individuated items. By contrast, parieto-occipital
alpha power reliably tracked the number of locations that
were attended, but this signal was completely unaffected
by perceptual grouping. This pattern of results suggests that
the negative slow wave is sensitive to the number of indi-
viduated items in WM, whereas parieto-occipital alpha
power tracks the number of visually attended locations.
These diverging effects of perceptual grouping dovetail
with recent evidence showing that the EEG signals that
track WM storage fall into at least two distinct classes.
Although both parieto-occipital slow waves and alpha
power track the number of relevant items in a WM task,
they have distinct temporal profiles, and explain distinct
between-subjects variance in WM capacity (Fukuda et al.,
2015). Moreover, these two EEG signals respond distinctly
to manipulations of attentional priority in WM (Günseli
et al., 2019) and to whether the task motivates the main-
tenance of item-based information (Hakim et al., 2019).
The perceptual grouping effect on the negative slow
wave is similar to the effects reported in studies that exam-
ined CDA, a lateralized ERP component that is observed at
similar electrode sites as the negative slow wave and is highly
sensitive to the number of items in VWM (Luria et al.,
2016; Vogel & Machizawa, 2004). Previous research charac-
terizing the negative slow wave was driven by the goal of
finding a whole-field alternative to the CDA that would
not require the filtering of distractors (Fukuda et al.,
2015). The findings, so far, are consistent with the hypoth-
esis that the CDA and negative slow wave are indexing the
same neural operation, although further work is needed
to determine how these signals are related. That said, pre-
vious research shows that the CDA tracks the number of
items regardless of the number of relevant locations and
even as the items undergo physical transformations. For
instance, the CDA reliably tracks additional items that are
added into VWM regardless of whether these additional
items are presented in the same locations as previously
encoded memoranda or not, which suggests that it ulti-
mately tracks the number of items rather than the num-
ber of attended positions (Ikkai et al., 2010). Moreover,
the CDA is sensitive to common fate cues, such that there
is a reduction in the CDA when common motion cues en-
courage the perception of multiple elements as a single
item (Luria & Vogel, 2014). There is also a reduction in
the CDA for identical colors relative to distinct colors,
which suggests that identical colors might be maintained
as a group in VWM (Peterson, Gözenman, Arciniega, &
Berryhill, 2015; Gao et al., 2011).
Importantly, the current work extends these findings
by providing a direct measure of information maintained
about each of the grouped elements, specifically spatial
information. Alpha oscillations provided a sensitive index
of the number of attended positions and revealed that
perceptual grouping yielded no reduction in the number
of attended positions. Specifically, parieto-occipital alpha
power continued to index the same number of spatial lo-
cations even when the stimuli were grouped, which sug-
gests that both constituents of the group were maintained.
This finding rules out the possibility that participants main-
tained only half of the group in grouped trials and used
that information to inform their decision at the time of re-
sponse. Instead, perceptual grouping boosted orientation
WM performance by reducing the number of individuated
representations stored without sacrificing spatial informa-
tion about each of the grouped elements. Our findings are
broadly consistent with past work that found a reduced
CDA for identical colors presumably maintained as a group
(Peterson et al., 2015; Gao et al., 2011). Future research
could extend the current work to include these similarity
cues or other grouping cues that are especially compel-
ling, such as common onset or common fate, or cues that
have been found to boost WM performance, such as prox-
imity (e.g., Woodman et al., 2003).
Finally, the distinct effects of perceptual grouping on
item-based and spatial EEG signals are consistent with a
broad class of models that distinguish between the num-
ber of individuated objects maintained in WM and the
specific featural information that is stored about each
of those objects (Fukuda, Vogel, Mayr, & Awh, 2010;
Awh, Barton, & Vogel, 2007; Xu & Chun, 2006). For ex-
ample, Xu and Chun (2009) described a neural object file
theory that distinguishes between an initial stage of ob-
ject individuation and a subsequent stage in which the
details of the individuated objects are apprehended.
This account is supported by their finding that grouped
objects elicited lower fMRI responses in inferior intrapar-
ietal sulcus (a brain region thought to track the number
of individuated items in WM) but higher responses in
superior intraparietal sulcus and the lateral occipital
complex, regions that are thought to be sensitive to the
information load or complexity of the stored items (Xu &
Chun, 2007). According to the neural object file account,
grouping reduced the number of individuated items in a
capacity-limited individuation stage, allowing more infor-
mation to be relayed to higher visual areas in the identi-
fication stage. Our results bolster this interpretation by
showing that distinct neural signals track the number of
individuated items in WM and the details of the specific
locations associated with each item. Thus, although neu-
ral signals tracking WM storage are sometimes viewed in
a monolithic fashion, there is growing evidence that dis-
tinct delay signals with distinct computational roles work
in parallel to maintain information in an on-line state
(Hakim, Awh, & Vogel, 2021).
Acknowledgments
Research was supported by National Institute of Mental Health
grant ROI MH087214 and Office of Naval Research grant
N00014-12-1-0972. We thank Ariana Gale and Clara Sava-Segal
for assistance with data collection.
1362
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Reprint requests should be sent to Gisella K. Diaz, Department
of Psychology, The University of Chicago, 940 East 57th Street,
Chicago, IL 60637, or via e-mail: gisella@uchicago.edu.
Funding Information
Edward K. Vogel, Office of Naval Research (https://dx.doi
.org/10.13039/100000006), grant number: N00014-12-1-
0972. Gisella K. Diaz and Edward K. Vogel, National
Institute of Mental Health (https://dx.doi.org/10.13039
/100000025), grant number: ROI MH087214.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/W = .108, and W/W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
Awh, E., Barton, B., & Vogel, E. K. (2007). Visual working
memory represents a fixed number of items regardless of
complexity. Psychological Science, 18, 622–628. https://doi.org
/10.1111/j.1467-9280.2007.01949.x, PubMed: 17614871
Bae, G.-Y., & Luck, S. J. (2018). Dissociable decoding of spatial
attention and working memory from EEG oscillations and
sustained potentials. Journal of Neuroscience, 38, 409–422.
https://doi.org/10.1523/JNEUROSCI.2860-17.2017, PubMed:
29167407
Brady, T. F., & Tenenbaum, J. B. (2013). A probabilistic model
of visual working memory: Incorporating higher order
regularities into working memory capacity estimates.
Psychological Review, 120, 85–109. https://doi.org/10.1037
/a0030779, PubMed: 23230888
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163/156856897X00357,
PubMed: 9176952
Busch, N. A., & Herrmann, C. S. (2003). Object-load and
feature-load modulate EEG in a short-term memory task.
NeuroReport, 14, 1721–1724. https://doi.org/10.1097
/00001756-200309150-00013, PubMed: 14512845
Foster, J. J., Sutterer, D. W., Serences, J. T., Vogel, E. K., & Awh,
E. (2016). The topography of alpha-band activity tracks
the content of spatial working memory. Journal of
Neurophysiology, 115, 168–177. https://doi.org/10.1152/jn
.00860.2015, PubMed: 26467522
Fukuda, K., Mance, I., & Vogel, E. K. (2015). α power
modulation and event-related slow wave provide dissociable
correlates of visual working memory. Journal of
Neuroscience, 35, 14009–14016. https://doi.org/10.1523
/JNEUROSCI.5003-14.2015, PubMed: 26468201
Fukuda, K., Vogel, E., Mayr, U., & Awh, E. (2010). Quantity, not
quality: The relationship between fluid intelligence and
working memory capacity. Psychonomic Bulletin & Review,
17, 673–679. https://doi.org/10.3758/17.5.673, PubMed:
21037165
Gao, Z., Gao, Q., Tang, N., Shui, R., & Shen, M. (2016).
Organization principles in visual working memory: Evidence
from sequential stimulus display. Cognition, 146, 277–288.
https://doi.org/10.1016/j.cognition.2015.10.005, PubMed:
26500190
Gao, Z., Xu, X., Chen, Z., Yin, J., Shen, M., & Shui, R. (2011).
Contralateral delay activity tracks object identity information
in visual short term memory. Brain Research, 1406, 30–42.
https://doi.org/10.1016/j.brainres.2011.06.049, PubMed:
21757188
Günseli, E., Fahrenfort, J. J., van Moorselaar, D., Daoultzis, K. C.,
Meeter, M., & Olivers, C. N. L. (2019). EEG dynamics reveal a
dissociation between storage and selective attention within
working memory. Scientific Reports, 9, 13499. https://doi.org
/10.1038/s41598-019-49577-0, PubMed: 31534150
Hakim, N., Adam, K. C. S., Gunseli, E., Awh, E., & Vogel, E. K.
(2019). Dissecting the neural focus of attention reveals
distinct processes for spatial attention and object-based
storage in visual working memory. Psychological Science,
30, 526–540. https://doi.org/10.1177/0956797619830384,
PubMed: 30817220
Hakim, N., Awh, E., & Vogel, E. K. (2021). Manifold visual
working memory. In R. H. Logie, V. Camos, & N. Cowan
(Eds.), Working memory: State of the science (pp. 44–84).
Oxford: Oxford University Press. https://doi.org/10.1093/oso
/9780198842286.003.0011
Ikkai, A., McCollough, A. W., & Vogel, E. K. (2010). Contralateral
delay activity provides a neural measure of the number of
representations in visual working memory. Journal of
Neurophysiology, 103, 1963–1968. https://doi.org/10.1152/jn
.00978.2009, PubMed: 20147415
Jiang, Y., Chun, M. M., & Olson, I. R. (2004). Perceptual
grouping in change detection. Perception & Psychophysics,
66, 446–453. https://doi.org/10.3758/BF03194892, PubMed:
15283069
Kristjánsson, Á. (2006). Surface assignment modulates object
formation for visual short-term memory. Perception, 35,
865–881. https://doi.org/10.1068/p5526, PubMed: 16970197
Luria, R., Balaban, H., Awh, E., & Vogel, E. K. (2016). The
contralateral delay activity as a neural measure of visual
working memory. Neuroscience and Biobehavioral Reviews,
62, 100–108. https://doi.org/10.1016/j.neubiorev.2016.01.003,
PubMed: 26802451
Luria, R., & Vogel, E. K. (2014). Come together, right now:
Dynamic overwriting of an object’s history through common
fate. Journal of Cognitive Neuroscience, 26, 1819–1828.
https://doi.org/10.1162/jocn_a_00584, PubMed: 24564468
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190. https://doi.org/10.1016/j.jneumeth
.2007.03.024, PubMed: 17517438
Mate, J., & Baqués, J. (2009). Visual similarity at encoding and
retrieval in an item recognition task. Quarterly Journal of
Experimental Psychology, 62, 1277–1284. https://doi.org/10
.1080/17470210802680769, PubMed: 19235099
Mazaheri, A., & Jensen, O. (2008). Asymmetric amplitude
modulations of brain oscillations generate slow evoked
responses. Journal of Neuroscience, 28, 7781–7787. https://doi
.org/10.1523/JNEUROSCI.1631-08.2008, PubMed: 18667610
Morey, C. C. (2019). Perceptual grouping boosts visual working
memory capacity and reduces effort during retention. British
Diaz, Vogel, and Awh
1363
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Journal of Psychology, 110, 306–327. https://doi.org/10.1111
/bjop.12355, PubMed: 30345501
Morey, C. C., Cong, Y., Zheng, Y., Price, M., & Morey, R. D.
(2015). The color-sharing bonus: Roles of perceptual
organization and attentive processes in visual working
memory. Archives of Scientific Psychology, 3, 18–29. https://
doi.org/10.1037/arc0000014
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis
of MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869. https://doi.org/10.1155/2011/156869, PubMed:
21253357
Pelli, D. G. (1997). The VideoToolbox software for visual
psychophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442. https://doi.org/10.1163/156856897X00366,
PubMed: 9176953
Peterson, D. J., & Berryhill, M. E. (2013). The Gestalt principle
of similarity benefits visual working memory. Psychonomic
Bulletin & Review, 20, 1282–1289. https://doi.org/10.3758
/s13423-013-0460-x, PubMed: 23702981
Peterson, D. J., Gözenman, F., Arciniega, H., & Berryhill, M. E.
(2015). Contralateral delay activity tracks the influence of
Gestalt grouping principles on active visual working memory
representations. Attention, Perception, & Psychophysics,
77, 2270–2283. https://doi.org/10.3758/s13414-015-0929-y,
PubMed: 26018644
Quinlan, P. T., & Cohen, D. J. (2012). Grouping and binding
in visual short-term memory. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 38, 1432–1438.
https://doi.org/10.1037/a0027866, PubMed: 22449133
Rihs, T. A., Michel, C. M., & Thut, G. (2007). Mechanisms of
selective inhibition in visual spatial attention are indexed
by alpha-band EEG synchronization. European Journal of
Neuroscience, 25, 603–610. https://doi.org/10.1111/j.1460
-9568.2007.05278.x, PubMed: 17284203
Sassenhagen, J., & Draschkow, D. (2019). Cluster-based
permutation tests of MEG/EEG data do not establish significance
of effect latency or location. Psychophysiology, 56, e13335.
https://doi.org/10.1111/psyp.13335, PubMed: 30657176
Sauseng, P., Klimesch, W., Heise, K. F., Gruber, W. R., Holz, E.,
Karim, A. A., et al. (2009). Brain oscillatory substrates of visual
short-term memory capacity. Current Biology, 19, 1846–1852.
https://doi.org/10.1016/j.cub.2009.08.062, PubMed: 19913428
Shen, M., Yu, W., Xu, X., & Gao, Z. (2013). Building blocks of
visual working memory: Objects or Boolean maps? Journal of
Cognitive Neuroscience, 25, 743–753. https://doi.org/10.1162
/jocn_a_00348, PubMed: 23249354
Todd, J. J., & Marois, R. (2004). Capacity limit of visual short-
term memory in human posterior parietal cortex. Nature,
428, 751–754. https://doi.org/10.1038/nature02466, PubMed:
15085133
van Dijk, H., van der Werf, J., Mazaheri, A., Medendorp, W. P., &
Jensen, O. (2010). Modulations in oscillatory activity with
amplitude asymmetry can produce cognitively relevant
event-related responses. Proceedings of the National
Academy of Sciences, U.S.A., 107, 900–905. https://doi.org
/10.1073/pnas.0908821107, PubMed: 20080773
Vogel, E. K., & Awh, E. (2008). How to exploit diversity for
scientific gain: Using individual differences to constrain
cognitive theory. Current Directions in Psychological
Science, 17, 171–176. https://doi.org/10.1111/j.1467-8721
.2008.00569.x
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity predicts
individual differences in visual working memory capacity.
Nature, 428, 748–751. https://doi.org/10.1038/nature02447,
PubMed: 15085132
Walker, P., & Davies, S. J. (2003). Perceptual completion and
object-based representations in short-term visual memory.
Memory & Cognition, 31, 746–760. https://doi.org/10.3758
/BF03196113, PubMed: 12956239
Wang, S., Megla, E. E., & Woodman, G. F. (2021). Stimulus-
induced alpha suppression tracks the difficulty of attentional
selection, not visual working memory storage. Journal of
Cognitive Neuroscience, 33, 536–562. https://doi.org/10.1162
/jocn_a_01637, PubMed: 33054550
Wang, S., Rajsic, J., & Woodman, G. F. (2019). The contralateral
delay activity tracks the sequential loading of objects into
visual working memory, unlike lateralized alpha oscillations.
Journal of Cognitive Neuroscience, 31, 1689–1698. https://
doi.org/10.1162/jocn_a_01446, PubMed: 31274391
Woodman, G. F., Vecera, S. P., & Luck, S. J. (2003). Perceptual
organization influences visual working memory.
Psychonomic Bulletin & Review, 10, 80–87. https://doi.org/10
.3758/BF03196470, PubMed: 12747493
Xu, Y. (2006). Understanding the object benefit in visual
short-term memory: The roles of feature proximity and
connectedness. Perception & Psychophysics, 68, 815–828.
https://doi.org/10.3758/BF03193704, PubMed: 17076349
Xu, Y., & Chun, M. M. (2006). Dissociable neural mechanisms
supporting visual short-term memory for objects. Nature,
440, 91–95. https://doi.org/10.1038/nature04262, PubMed:
16382240
Xu, Y., & Chun, M. M. (2007). Visual grouping in human parietal
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 104, 18766–18771. https://doi.org/10.1073/pnas
.0705618104, PubMed: 17998539
Xu, Y., & Chun, M. M. (2009). Selecting and perceiving multiple
visual objects. Trends in Cognitive Sciences, 13, 167–174.
https://doi.org/10.1016/j.tics.2009.01.008, PubMed: 19269882
1364
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
5
4
1
9
2
1
2
4
5
/
/
j
o
c
n
_
a
_
0
1
7
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3