Perceptual Expectations Modulate Low-Frequency Activity:
A Statistical Learning Magnetoencephalography Study
Ying Joey Zhou1, Alexis Pérez-Bellido1,2, Saskia Haegens1,3, and Floris P. de Lange1
Abstrakt
■ Perceptual expectations can change how a visual stimulus
is perceived. Recent studies have shown mixed results in
terms of whether expectations modulate sensory representa-
tionen. Hier, we used a statistical learning paradigm to study
the temporal characteristics of perceptual expectations. Wir
presented participants with pairs of object images organized
in a predictive manner and then recorded their brain activity
with magnetoencephalography while they viewed expected
and unexpected image pairs on the subsequent day. Wir
observed stronger alpha-band (7–14 Hz) activity in response
to unexpected compared with expected object images.
Speziell, the alpha-band modulation occurred as early as
the onset of the stimuli and was most pronounced in left
occipito-temporal cortex. Given that the differential response
to expected versus unexpected stimuli occurred in sensory re-
gions early in time, our results suggest that expectations mod-
ulate perceptual decision-making by changing the sensory
response elicited by the stimuli. ■
EINFÜHRUNG
Perception can be understood as a process of probabilistic in-
Referenz, in which the brain actively generates predictions and
compares them with the sensory input it receives (de Lange,
Heilbron, & Kok, 2018; Friston, 2005). When sensory
input is inconsistent with the current predictions, Die
system generates a prediction error signal, which is fed
forward to update predictions. When sensory input is
consistent with current predictions, prediction errors are
minimal, resulting in an attenuated sensory response
(Summerfield & de Lange, 2014; Alink, Schwiedrzik, Kohler,
Singer, & Muckli, 2010; Murray, Kersten, Olshausen,
Schrater, & Wald, 2002).
On the implementation level of predictive perception,
it has been proposed that low- and high-frequency oscil-
lations may carry feedback predictions and feedforward
prediction errors, jeweils (Michalareas et al., 2016;
Bastos et al., 2012). This proposal is supported by neuro-
physiological evidence showing that oscillations in the
alpha (7–14 Hz) and gamma (40–90 Hz) range propagate
in a feedback and feedforward fashion, jeweils
(Bonaiuto et al., 2018; van Kerkoerle et al., 2014). In ad-
dition, many studies have shown that these oscillatory
signals correlate with behavior at different stages of per-
ceptual decision-making. Oscillatory alpha-band activity
has been shown to index the allocation of attentional re-
sources (Haegens, Händel, & Jensen, 2011; Thut, Nietzel,
1Radboud University, 2University of Barcelona, 3Columbia
Universität
© 2019 Massachusetts Institute of Technology
Brandt, & Pascual-Leone, 2006; Worden, Foxe, Wang, &
Simpson, 2000), the employment of sensory expectations
(Mayer, Schwiedrzik, Wibral, Singer, & Melloni, 2016;
Sherman, Kanai, Seth, & VanRullen, 2016), and other
top–down cognitive control functions (für eine Rezension, sehen
Sadaghiani & Kleinschmidt, 2016). Generally speaking,
alpha power decreases with task engagement and increases
when an area is disengaged ( Jensen, Bonnefond, &
VanRullen, 2012; Haegens, Nácher, Luna, Romo, & Jensen,
2011). Gamma band activity, andererseits, has been
shown to increase when sensory expectations are violated
(Bauer, Stenner, Friston, & Dolan, 2014; Arnal, Wyart, &
Giraud, 2011), consistent with the idea that prediction er-
rors may be fed forward to update the system’s predictions.
In addition to its implementation, recent studies have
also focused on how predictions (or perceptual expecta-
tionen) are employed by the brain. Some researchers have
suggested that expectations facilitate perception by en-
hancing the sensory representation per se. When a stimu-
lus is expected, the sensory representation is more precise
(Kok, Jehee, & de Lange, 2012) and participants’ sensitivity
to the signal is enhanced compared with an unexpected
stimulus (Cheadle, Egner, Wyart, Wu, & Summerfield,
2015; Wyart, Nobre, & Summerfield, 2012). Jedoch,
others have suggested that expectations facilitate percep-
tion by shifting the decision criterion without changing the
sensory representation. Zum Beispiel, Bang and Rahnev
(2017) compared conditions where the expectation cues
were provided either before or after the stimulus presen-
0) difference between
tation and found no sensitivity (D
these conditions. Another recent study used EEG to track
neural markers of sensory processing and found no
Zeitschrift für kognitive Neurowissenschaften 32:4, S. 691–702
https://doi.org/10.1162/jocn_a_01511
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
/
T
F
Ö
N
0
5
M
A
j
2
0
2
1
expectation modulations on sensory processing, al-
though expectations significantly modulated behavioral
RTs (Rungratsameetaweemana, Itthipuripat, Salazar, &
Serences, 2018). Taken together, it is still debated which
stage of the perceptual decision-making process is mod-
ulated by prior expectations.
Statistical learning is a commonly used paradigm to
study predictive perception in both human (Richter &
de Lange, 2019; Manahova, Mostert, Kok, Schoffelen,
& de Lange, 2018; Richter, Ekman, & de Lange, 2018;
Turk-Browne, Scholl, Johnson, & Chun, 2010) and non-
menschlich (Ramachandran, Meyer, & Olson, 2017; Meyer,
Ramachandran, & Olson, 2014; Meyer & Olson, 2011)
primates. Instead of providing explicit expectation cues
and instructing participants to expect upcoming stimuli,
expectations in statistical learning paradigm are learned
and employed in a more automatic and implicit manner
(Batterink, Reber, Neville, & Paller, 2015; Kim, Seitz,
Feenstra, & Shams, 2009). Modulatory effects of expec-
tations that are based on learned statistical regularities
are therefore less likely to be confounded by top–down
goal-directed attention, making it a useful paradigm to
study predictive perception. Studies using this paradigm
have consistently shown that participants respond faster
and more accurately when objects are presented in an
expected order (d.h., following the same arrangement
as during prior exposure), compared with when objects
are presented in an unexpected order. Darüber hinaus, Die
neural response in object-selective cortex is typically
stronger for unexpected, compared with expected,
object stimuli (Richter & de Lange, 2019; Kaposvari,
Kumar, & Vogels, 2018; Manahova et al., 2018; Richter
et al., 2018; Ramachandran et al., 2017; Meyer & Olson,
2011). This phenomenon—that unexpected stimuli eli-
cited a stronger neural response—is consistent with elec-
trophysiological studies showing that unexpected stimuli
lead to an enhanced response amplitude of ERP compo-
nents such as the N170 (Robinson, Breakspear, Jung, &
Johnston, 2018; Johnston et al., 2017) and the visual mis-
match negativity (see Stefanics, Kremláček, & Czigler,
2014, für eine Rezension).
The current study aimed to address whether expecta-
tions modulate sensory processing. We used a statistical
learning paradigm to characterize the temporal profile of
the brain’s response to expected and unexpected object
Bilder. Genauer, we characterized the strength
von (phase-locked) ERFs and (non-phase-locked) neural
oscillations to explore their modulation in predictive
perception. To preview, although we observed no differ-
ences in ERFs between the different expectation con-
ditions, we found stronger low-frequency oscillatory
activity over occipito-temporal cortex in response to an
image when it was unexpected than when it was ex-
pected. The temporal and spatial profiles of the observed
activity differences suggest that expectations derived
from statistical regularities modulate perception at an
early sensory processing stage.
METHODEN
Data Availability
All data and code used for stimulus presentation and
analysis are available online at the Donders Repository
at hdl.handle.net/11633/aacvnzlx.
Teilnehmer
Our target sample size was a priori set to 34, providing us
mit 80% power to detect two-sided experimental effects
that had at least a medium effect size (Cohen’s d > 0.5).
Power analysis was conducted with G*Power (Faul,
Erdfelder, Lang, & Buchner, 2007). Thirty-nine healthy
adult participants were recruited online via the SONA
System. All participants reported normal (or corrected-
to-normal) vision. Five participants were excluded from
analysis because of technical errors during data recording
or dropout after the first session, resulting in the planned
sample size of 34 Teilnehmer (21 Frauen; Durchschnittsalter =
23.4 Jahre, SD = 3.1 Jahre) in the reported analysis.
The study was approved by the local ethics committee
(CMO Arnhem-Nijmegen). All participants gave informed
consent before the experiment and received monetary
compensation for their participation.
Stimuli
A set of 80 object images of electronic and nonelectronic
items were chosen from an image database (cvcl.mit.edu/
MM/uniqueObjects.html; Brady, Konkle, Alvarez, & Oliva,
2008) as stimuli. For each participant, we randomly se-
lected nine images from the set as leading images and
six images as trailing images (three were of electronic
Artikel, and the other three were of nonelectronic items).
Color images were presented on a gray (RGB = [128,
128, 128]) background, spanning approximately 5° × 5°
(degrees) of visual angle (dva) on the screen. A bull’s
eye (outer black ring = 0.5° × 0.5° dva, innermost black
dot = 0.25° × 0.25° dva) was used as fixation and pre-
sented throughout the run. The stimuli were displayed
on an LCD screen during behavioral training and on a
semitranslucent screen (1920 × 1080 pixel resolution,
120-Hz refresh rate) back-projected by a PROpixx projector
( VPixx Technologies) during magnetoencephalography
(MEG) Aufnahmen. The experiment was programmed
with PsychtoolBox (Brainard, 1997) in MATLAB (Der
Mathworks, Inc.).
Verfahren
Participants reported to the laboratory on 2 consecutive
Tage, for a training session on Day 1 and MEG recording
session on Day 2. The training session on Day 1 served to
familiarize participants with the task and the predictive
relationship between the stimuli. Participants completed
eight blocks of 92 trials of the main task during training.
692
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 4
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
F
T
/
.
Ö
N
0
5
M
A
j
2
0
2
1
Participants were tested in the MEG on Day 2, während
which they performed six blocks of the main task and
six blocks of the functional localizer task while their
brain activity was recorded. They completed a 20-min
behavioral test after the MEG recording, during which
they performed the categorization task and the ques-
tionär. Depending on the availability of participants’
T1-weighted anatomical scans in our institute’s data-
base, some of the participants returned for a third
session, during which we obtained their T1-weighted
anatomical scans.
Experimental Paradigm
Main Task
Participants were presented with two object images in
quick succession. Each image was presented for 500 ms
with a 300-msec ISI in between. An intertrial interval of
900–1000 msec was used (Figure 1A shows schematic of
an example trial). Fifteen images were shown in different
image pairs throughout the experiment, of which nine al-
ways appeared first within any image pair (“leading im-
age”) and the remaining six always appeared second
(“trailing image”). The participants’ task was to press a
key as fast as possible when they saw an image presented
upside down, which occurred randomly in 88 (Ausbildung
session) Und 90 (MEG session) trials and equally likely
for either the leading or trailing image.
The predictive relationships of the 15 images were sub-
tly different in the training and MEG sessions. During train-
ing, six of the nine leading images were always followed by
the same trailing image (d.h., 100% predictive, referred to
as expected pairs), and three of them were followed
equally likely by any of the six trailing images (d.h., 16.7%
predictive, referred to as neutral pairs). Each expected pair
was presented 72 mal, and each neutral pair was pre-
gesendet 12 mal, ergebend 648 nonoddball trials during
Ausbildung. This information was provided to participants be-
fore they were shown any image; das ist, the fact that there
was a predictive structure was revealed but not the exact
pairwise relationships between images.
During the MEG session, unexpected trials were intro-
duced by manipulating the probabilistic relationship be-
tween the leading and trailing images (Figure 1D): Jede
leading image of the expected pairs was followed in
58.3% of the trials by the trailing image paired with it dur-
ing training and 41.7% by one of the other trailing images
(8.3% for each trailing image). Four hundred thirty-two
nonoddball trials were used during the MEG session.
The predictive relationship of the neutral pairs remained
the same. The neutral pairs were used as a baseline for
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
F
T
/
Ö
N
0
5
M
A
j
2
0
2
1
Figur 1. Experimental paradigm and behavioral results of the categorization task. (A) Schematic of the main task trial during MEG recording.
When an upside-down image was presented, participants made a button press as fast as possible before the end of the trial. When no upside-down
image was presented, participants made no motor response. The same trial presentation parameters were used for the categorization task, except
that participants had to respond within 1.2 sec after trailing image onset in each trial. (B) Schematic of the functional localizer trial. (C) Der
arrangement of the functional localizer (F) and the main task blocks (M) during MEG recording. (D) Occurrence of each image pair during the MEG
recording. “Lx” denotes the xth leading image, and “Ty” denotes the yth trailing image randomly selected for each participant. Image pairs of
the expected, neutral, and unexpected conditions were shaded in green, Blau, and orange, jeweils. (E) Accuracy and RTs (correct trials only) für
the categorization task. Error bars denote 95% confidence interval. Asterisk denotes significance ( P < .05) in repeated-measures ANOVA. ITI =
intertrial interval.
Zhou et al.
693
comparison, enabling us to ask whether the neural response
was suppressed when a stimulus was expected and/or en-
hanced when a stimulus was unexpected. Participants were
informed at the beginning of the MEG session that there
would be a small change in how images were paired,
yet the exact predictive structure was not explained.
Functional Localizer
To identify sensors most responsive to the experimental
stimuli, independent of perceptual expectations, func-
tional localizer trials were introduced during the MEG
session. Only the six trailing images were used as stimuli
in localizer trials. Each image was presented 110 times
(10 trials being upside down) in a pseudorandom order.
All trials started with a 500-msec fixation period, followed
by a 500-msec image presentation, and were separated by
an intertrial interval of 900–1100 msec (Figure 1B). Partic-
ipants were asked to respond as fast as possible (while
keeping fixation) when an image was presented upside
down.
Categorization Task
To assess participants’ knowledge about the predictive
relationships between images, we asked our participants
to perform a categorization task after their MEG record-
ing. We reasoned that, if participants learned the relation-
ship between images and could use this knowledge to
guide behavior, they would respond faster and more ac-
curately for expected trials. Participants were instructed
to report as fast as possible on every trial whether the
trailing image was of an electronic item or not. The same
trial structure and stimuli as those in the MEG main task
were used, with the only difference that the occurrence
of each image pair was halved to reduce participants’ fa-
tigue. A response window of 1.2 sec was used to encour-
age participants to prioritize response speed.
(CTF MEG Systems, VSM MedTech Ltd.) in a magnetically
shielded room. Three fiducial coils were placed at a par-
ticipant’s nasion and both ear canals, to provide online
monitoring of participant’s head position and offline an-
atomical landmarks for coregistration. Eye position was
recorded using an eye tracker (EyeLink, SR Research
Ltd.) during the MEG recordings.
Anatomical MRIs were obtained during a third session
or requested from the center’s database if available. To
improve coregistration of the MRIs and MEG data,
earplugs with a drop of Vitamin E were placed at par-
ticipants’ ear canals during MRI acquisition. These ana-
tomical scans were used for source reconstruction of
the MEG signals. Note that source analysis reported here
was based on 33 participants, as the anatomical MRIs of
one participant were not available because of dropout
after the MEG session.
Data Analysis
MEG Preprocessing
MEG data were preprocessed offline and analyzed using the
FieldTrip toolbox (Oostenveld, Fries, Maris, & Schoffelen,
2011) and custom-built MATLAB scripts. Trials of the main
task and localizer were segmented and processed sepa-
rately, given their different trial lengths. The data were
down-sampled to a sampling frequency of 400 Hz, after
applying a notch filter to remove line noise and har-
monics (at 50, 100, and 150 Hz). Bad channels and trials
were rejected via visual inspection before independent
component analysis. Independent component analysis
components were visually inspected, and those repre-
senting eye and heart artifacts were then projected out
of the data. For the resulting data, outlier trials of ex-
treme variance and trials within which participants
blinked during the presentation of images were further
removed from the data.
Questionnaire
ERFs
After the categorization task, participants were tested
on their explicit awareness of the predictive relation-
ships between images. Their task was to indicate using
a 4-point scale how frequently they had seen a given
image pair in the MEG main task (1 = most frequent
and 4 = less frequent). They were shown 24 image
pairs, consisting of the six expected pairs, six randomly
selected neutral pairs, six randomly selected unexpected
pairs, and six “swapped” pairs (i.e., image pairs whose
presentation order of leading and trailing images was
swapped).
Data Acquisition
Whole-head MEG data were acquired at 1200 Hz with a
275-channel CTF MEG system with axial gradiometers
Before calculating the ERFs, singe-trial data were baseline-
corrected using an interval of [−0.5, 0] sec for the main
task trials and [−0.2, 0] sec for the localizer trials. To avoid
the confounding influence of noise (in the planar transfor-
mation) because of unequal trial numbers across condi-
tions, trial numbers were equated via subsampling when
computing ERFs for different conditions. Specifically, we
subsampled an equal number of trials from each condition
before averaging over trials, such that the number of trials
per condition matched that in the condition of the fewest
trials. Planar gradients of the MEG field distribution were
then calculated, which makes interpretation of the sensor-
level data easier and enables comparing and averaging ERF
topographies across participants. We repeated the above-
mentioned procedure 10 times per condition, to ensure
every trial was used at least once, and then averaged over
694
Journal of Cognitive Neuroscience
Volume 32, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
4
e
6
-
9
p
1
d
f
2
0
/
1
3
3
2
2
/
5
4
5
/
6
o
9
c
1
n
/
_
a
1
_
8
0
6
1
1
5
3
1
3
1
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
1
0
1
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
all corresponding planar-combined averages to obtain
ERFs per condition.
Spectral Analysis
Time–frequency representations (TFRs) of each trial were
calculated by applying a fast Fourier transform to short
sliding time windows. For low frequencies (4–30 Hz), a
Hanning-tapered 300-msec sliding time window was used
in time steps of 50 msec and frequency steps of 1 Hz.
High-frequency (30–120 Hz) power was estimated using
a discrete prolate spheroidal sequences multitaper ap-
proach with a sliding time window of 200 msec in time
steps of 50 msec and frequency steps of 2 Hz, with ±10-
Hz smoothing (obtained by using three tapers). Singe-trial
TFRs were then averaged per condition. Power in the re-
sulting average TFRs was expressed relative to a baseline,
defined as [−0.6, −0.15] and [−0.4, −0.1] sec time-locked
to stimulus onset for low and high frequencies, respective-
ly. These time windows were used to prevent leakage of
poststimulus activity into the baseline window.
Source Reconstruction of Frequency-Domain Data
The partial canonical coherence beamformer approach
(Gross et al., 2001) was used to localize the sources of
the observed differences in the sensor-level TFR between
the expected and unexpected conditions. Volume con-
duction models were constructed based on a single-shell
model of individual participants’ anatomical MRIs (Nolte,
2003). They were then used to construct participant-
specific search grids (6-mm resolution), which were later
normalized to a template in Montreal Neurological
Institute space. For each grid point, lead fields were com-
puted with a reduced rank, which removes the sensitivity
to the direction perpendicular to the surface of the vol-
ume conduction model. We extracted 400-msec data seg-
ments ([0.8, 1.2] sec time-locked to stimulus onset) from
trials of both conditions and computed cross-spectral
density matrices using the multitaper method centered
at a frequency of 10 (±6) Hz. With the cross-spectral den-
sity matrices and the lead fields, a common spatial filter
(i.e., common for both conditions) was constructed for
each grid point for each participant. Using this common
spatial filter, the spatial distribution of power was then
estimated for the expected and unexpected conditions
separately. To visualize the difference between condi-
tions on source level, t statistic was computed for each
grid point as a proxy of the source’s contribution to the
difference.
Statistical Inference
Behavioral Data
Behavioral results of the post-MEG tasks allowed us to
evaluate whether participants had learned and used
(either implicitly or explicitly) the transitional relations
between images. Post-MEG behavioral data reported
here were based on 33 participants, as data from one par-
ticipant were not recorded because of a hardware error
during testing. For the categorization task, mean RTs and
accuracy of each condition were computed per partici-
pant. A within-participants repeated-measures ANOVA
was then applied separately to both measures. For the
questionnaire, the median rating score of each condition
was computed per participant and used as entries to the
repeated-measures ANOVA.
MEG Data
Statistical significance was evaluated using cluster-based
permutation tests (Maris & Oostenveld, 2007). The time
interval of interest was defined as 0.8–1.6 sec (i.e., the
800-msec window after the onset of the trailing image,
during which expectations could be either confirmed or
violated). For reference, we also considered the 0- to
0.8-sec time interval (i.e., the 800-msec window after
the onset of the leading image, during which expectations
about the trailing image may form). Pairwise permutation
tests between conditions were conducted separately for
the two intervals. For ERFs, data at each sensor and time
point were compared univariately between two condi-
tions and were used for clustering when the correspond-
ing two-tailed paired t test resulted in a p value smaller
than .05. A similar procedure was applied to TFRs, with
the only difference being that the clustering took place
in three dimensions including frequencies, sensors, and
time course. The sum of the t values within a cluster
was then computed as cluster-level statistic. The cluster
with the maximum sum was subsequently used as test
statistic. By randomizing the data across the two con-
ditions and recalculating the test statistic 5000 times, we
obtained a reference distribution of maximum cluster
t values to evaluate the statistic of the actual data. The
cluster of interest was considered significant when it fell
out of the 95% tail of the reference distribution.
RESULTS
Behavioral Results
During the MEG main task, participants had to press a
button for upside-down oddball images. They detected
the oddballs on 99.3% (range = 97.3–100%) of the occur-
rences with mean RTs of 518 msec (between-participants
SD = 44 msec), confirming their close engagement in the
task.
During the categorization task (after the MEG recording),
a significant difference in RT was observed for the three
types of image pairs, F(1.54, 49.13) = 6.619, p = .006
(Greenhouse–Geisser corrected). Specifically, expected im-
age pairs led to significantly shorter RT compared with unex-
pected pairs (Mexpected = 472 msec, Munexpected = 484 msec;
Zhou et al.
695
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
4
e
6
-
9
p
1
d
f
2
0
/
1
3
3
2
2
/
5
4
5
/
6
o
9
c
1
n
/
_
a
1
_
8
0
6
1
1
5
3
1
3
1
5
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
1
0
1
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
.
f
o
n
0
5
M
a
y
2
0
2
1
t(32) = −3.044, p = .005, uncorrected), with the RT of the
neutral pairs (Mneutral = 480 msec) in between (expected
vs. neutral: t(32) = −2.168, p = .037, uncorrected; invalid
vs. neutral: t(32) = −1.857, p = .073, uncorrected). Accu-
racy showed a similar yet insignificant pattern as RT, F(1.78,
57.05) = 2.983, p = .064 (Greenhouse–Geisser corrected).
This lack of statistical significance in accuracy may be be-
cause of a ceiling effect (average accuracy across all con-
ditions = 94.50%). Overall, participants’ performance in
the categorization task indicated that they learned the pre-
dictive relationship between images and benefited from
this knowledge when expectations were relevant.
Finally, we presented image pairs in expected, neutral,
unexpected, and swapped order and acquired frequency
ratings from participants. A significant difference in the
frequency ratings of the four trial types, F(2.75, 87.99) =
49.43, p < .001 (Greenhouse–Geisser corrected), was
found. Post hoc tests showed that participants reported
the expected pairs to have occurred most frequently
(Mexpected = 1.91, uncorrected p < .01 for expected vs.
neutral, expected vs. unexpected, and expected vs.
swapped) and the swapped pairs as the least frequent
(Mswapped = 3.45, uncorrected p < .001 for swapped vs.
expected, swapped vs. neutral, and swapped vs. unex-
pected), with the neutral and unexpected pairs (Mneutral =
2.31, Munexpected = 2.53) in between (neutral vs. un-
expected, p = .055, uncorrected). These results further
suggested that participants learned the predictive relation-
ship between images.
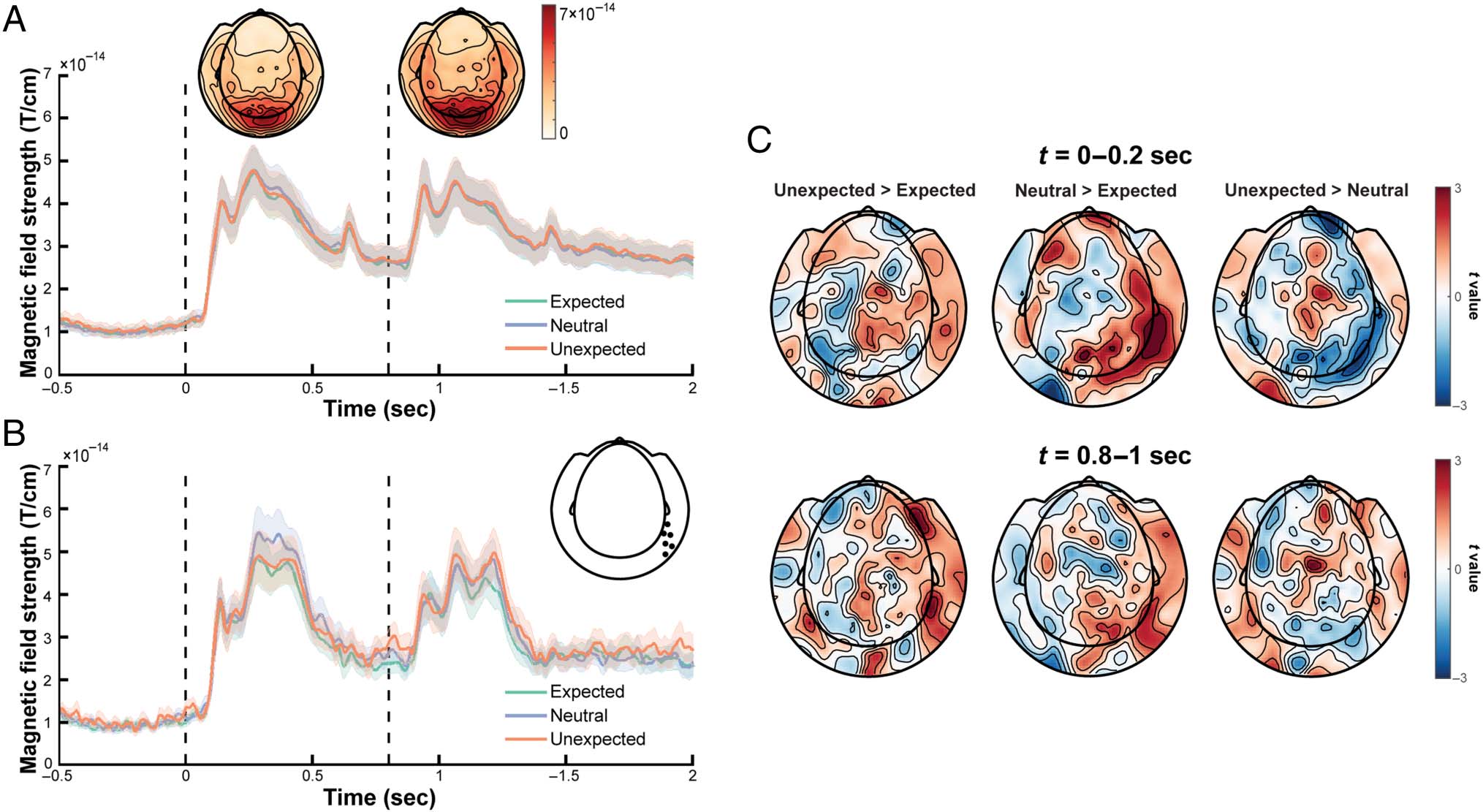
No ERF Amplitude Difference between Conditions
To investigate whether expectations modulated the re-
sponse to the trailing image, we contrasted the evoked
response to each of the three conditions (using three
pairwise permutation tests) in an 800-msec window start-
ing at the onset of the trailing image. We observed no sig-
nificant differences between any of the conditions ( p > .5
for all three pairwise permutation tests; Figure 2A and C,
bottom row). Wie erwartet, there were also no differences
between conditions for the evoked response to the lead-
ing image (d.h., pairwise permutation tests for an 800-msec
window aligned to the onset of leading image; Figure 2C,
top row). It could be argued that the statistical approach
that we used, a cluster-based permutation test that con-
trols the Type I error rate at the whole-brain level (involv-
ing 275 sensors), has reduced sensitivity compared with a
more focused analysis that tests for differences in a subset
of sensors of interest (z.B., occipital sensors). daher,
we repeated our analysis for a subset of sensors reported
to show different activations to expected and unexpected
stimuli in a previous study by Manahova et al. (2018).
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
/
.
F
T
Ö
N
0
5
M
A
j
2
0
2
1
Figur 2. ERFs. (A) Grand-averaged ERF of all sensors. Black dotted lines denote the onsets of the leading and trailing images, jeweils.
Topographic plots above show the ERF amplitude time-locked to the first 200 msec of the leading and trailing image presentations. Error bars denote
between-participants standard error. (B) Grand-averaged ERF of selected right occipito-temporal sensors (marked on the topography), based on
Manahova et al. (2018). Error bars denote between-participants standard error. (C) Topographies of the contrasts of interest (showing t values):
unexpected versus expected (links), neutral versus expected (center), and unexpected versus neutral (Rechts) for activity during the first 200 msec after
the leading image (top row) and the trailing image (bottom row).
696
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 4
Results for this ROI analysis showed no significant differ-
ences between the three conditions ( p = 1 for all three
pairwise permutation tests).
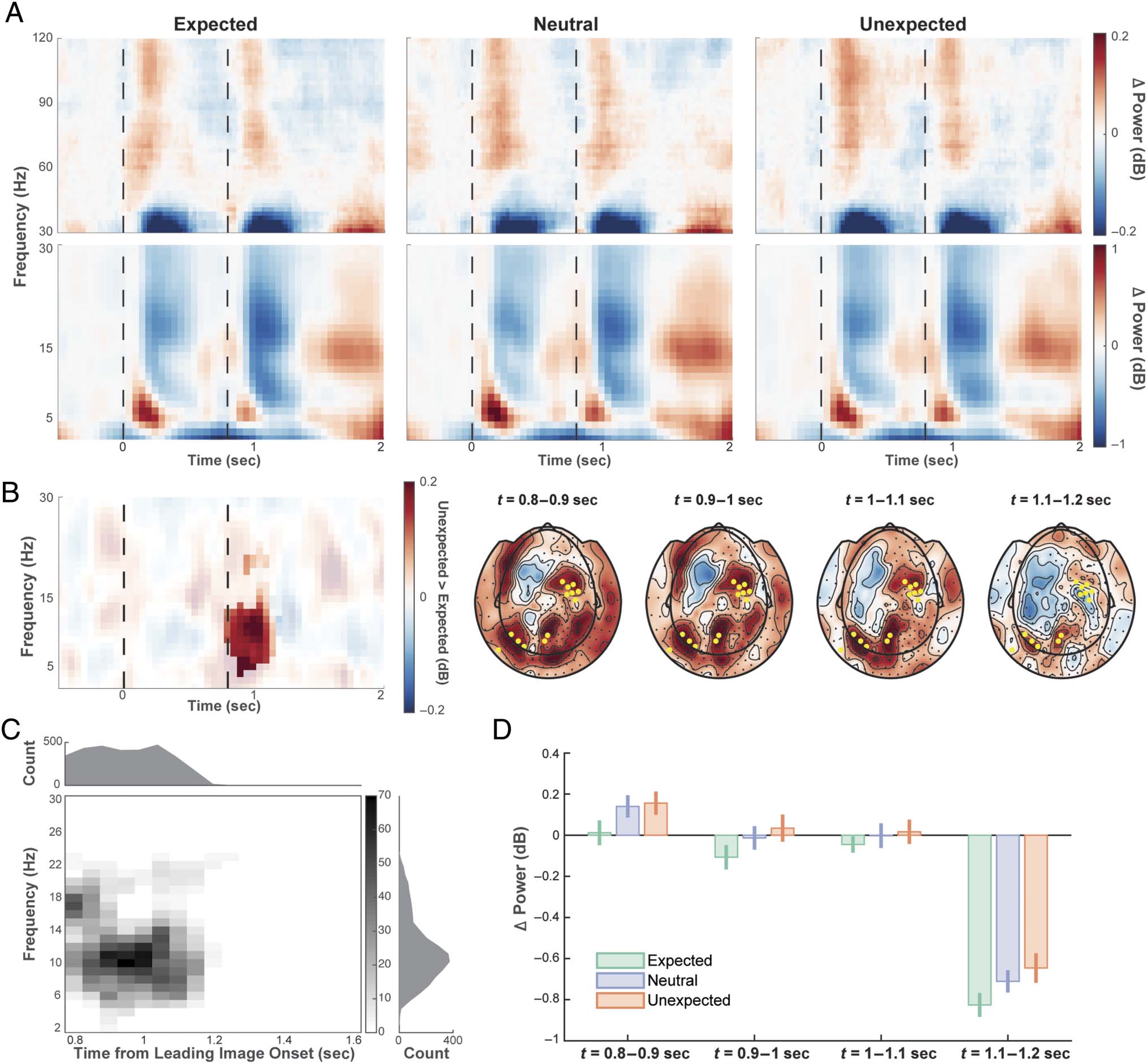
Unexpected Object Images Induce Stronger
Low-Frequency Power
Although we found no significant ERF differences be-
tween conditions, we did observe a significant difference
in low-frequency power between expected and unex-
pected conditions. When contrasting TFRs during the
processing of the trailing image, we found a significant
spectro-temporal cluster with stronger power for the
unexpected than the expected object stimuli, centered
around 10 Hz (time window of 0.8–1.2 sec after the onset
of the leading image, d.h., at onset of the trailing image, fre-
quency range = 6–26 Hz, p = 0.006, Bonferroni corrected;
Figure 3B and C).
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
/
F
.
T
Ö
N
0
5
M
A
j
2
0
2
1
Figur 3. TFRs. (A) TFRs of low and high frequencies of selected sensors (highlighted in B with yellow markers), showing power during task versus
baseline. Black dotted lines denote the onsets of the leading and trailing images. (B) TFRs showing the difference in power between expected and
unexpected conditions (of sensors highlighted with yellow markers), and the corresponding topographies over time for frequencies ranging from 4
Zu 16 Hz. Sensors contributed to the cluster were highlighted with black markers, with those contributed to the cluster at all time points tested highlighted
in yellow. (C) Overview of number of sensors that contributed to the significant contrast between expected and unexpected conditions, per time–
frequency sample. (D) Power change (relative to baseline) of frequencies ranging from 4 Zu 16 Hz of the three expectation conditions in different
time windows. Slightly different subsets of sensors contributed to the significant cluster (as in C) in different time windows; daher, the sensors used to
compute the bars for each time window differed. Error bars denote 95% confidence interval in within-subject comparison.
Zhou et al.
697
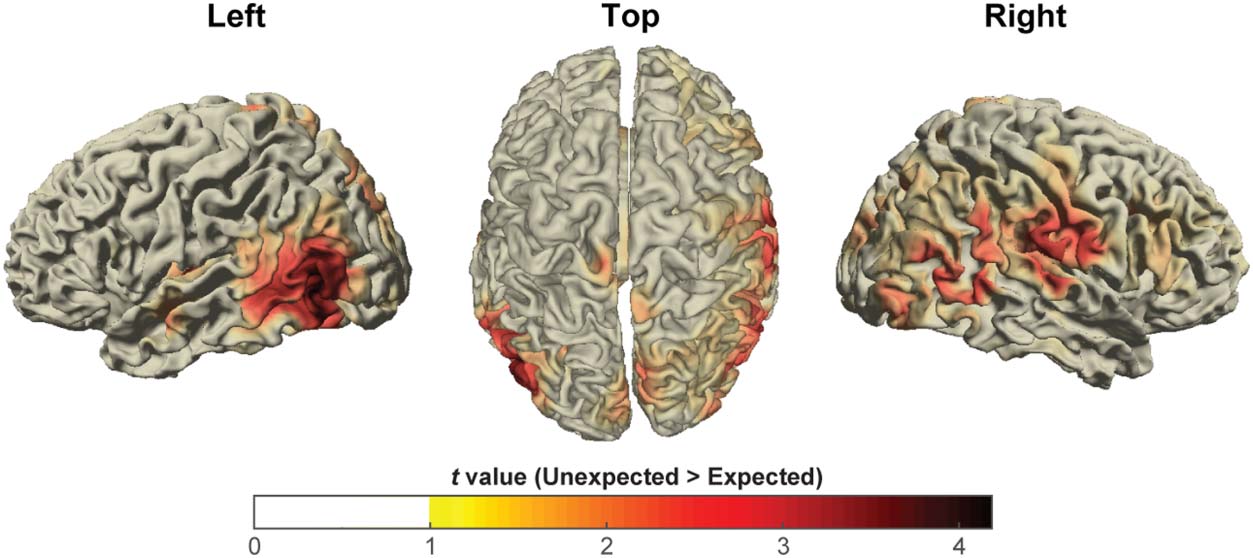
Figur 4. Quelle
reconstruction of the significant
effect for expected versus
unexpected conditions
(frequency range: 10 ± 6 Hz;
time window: [0.8, 1.2] Sek,
with respect to leading image
onset).
D
Ö
w
N
l
Ö
A
D
e
D
We used beamformer analysis to localize the source of
this power difference and found that it stemmed mainly
from the left occipito-temporal cortex (Figur 4). Pair-
wise comparisons between the neutral condition and
the other two conditions were not significant ( p > .3
for both permutation tests).
No Evidence for Phase-Locked Expectation
Modulation of Low-Frequency Activity
The diverging patterns of our ERF and TFR results—that
Ist, lack of differences in the evoked response versus sig-
nificantly different oscillatory patterns—suggest that the
low-frequency power differences between expected and un-
expected stimuli are unlikely to be phase locked. Beachten Sie, dass,
although our TFR analysis mainly reflected the induced
Aktivität, it might also have captured some evoked/phase-
locked responses. To address this further, we estimated
the phase-locked TFRs (Figure 5A) by computing the
time–frequency dynamics of the average ERFs for each con-
dition (see Swettenham, Muthukumaraswamy, & Singh,
2009, for a detailed description of the method). We then
repeated the comparisons between conditions, speziell
focusing on the contrast between the expected and unex-
pected conditions. There were no significant differences be-
tween conditions for the phase-locked TFRs ( p > .18;
Figure 5B). We additionally computed the phase-locked
TFRs using another method (Cohen, 2014) and compared
the resulting TFRs between conditions. This additional
analysis also did not indicate a difference between ex-
pected and unexpected conditions in the phase-locked
component ( p > .8). Zusammen, these analyses suggest
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
/
T
F
Ö
N
0
5
M
A
j
2
0
2
1
Figur 5. Phase-locked TFR. (A) Phase-locked TFRs of low frequencies. (B) TFR showing the difference in power between expected and
unexpected conditions (of selected sensors highlighted in yellow in Figure 3B), and the corresponding topographies over time for frequencies
ranging from 4 Zu 16 Hz.
698
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 4
that expectations modulated endogenous oscillations in a
non-phase-locked manner, rather than these oscillatory
patterns being evoked by the external stimulus.
No Difference between Conditions in
High-Frequency Power
Although we observed significant high-frequency activity in
response to both the leading and trailing images ( P < .01
for response to leading image vs. baseline and trailing
image vs. baseline; see Figure 3A, top row), our cluster-
based permutation tests revealed no significant differ-
ences between conditions for either time interval (for
all permutation tests, p > .3).
DISKUSSION
In the current study, we investigated the consequences
of prior expectation on the neural response to object
stimuli using MEG. Speziell, we aimed to address
whether expectations modulate perceptual processing
by changing early sensory representations and to charac-
terize the oscillatory activity induced by (violations of )
expectation. Using a statistical learning paradigm in
which participants implicitly acquired knowledge about
stimulus transitions, we found a modulatory effect of per-
ceptual expectations on low-frequency activity centered
at around 10 Hz, in the form of increased power in re-
sponse to unexpected relative to expected items.
No Effect of Expectation on ERFs
Somewhat surprisingly, we did not observe any expec-
tation modulations in the ERFs, in apparent contrast with
earlier electrophysiological studies in nonhuman primates
(Ramachandran et al., 2017; Meyer & Olson, 2011) Das
used similar stimuli and probabilistic transition rules. Von
Notiz, obwohl, primates were exposed to the object images
extensively for many days in these studies, which may
have caused object-selective neurons to alter their selec-
tivity profiles (Li & DiCarlo, 2008, 2010). Somit, es ist
plausible that, in these studies, expectations modulated
neuronal activity in a different manner than in our study,
where exposure to each image was limited. Zusätzlich,
attention was typically not controlled in these studies, als
primates were simply trained to maintain fixation during
stimulus presentation.
The lack of ERF amplitude differences that we observe
also appears at odds with earlier electrophysiological
studies in human participants, where multiple ERP
components have been shown to index prediction error
or surprise. Zum Beispiel, using predictable image se-
quences, Johnston et al. (2017) showed that violation
of expectations robustly modulated the N170 ERP com-
ponent across several stimulus categories. More gen-
erally, multiple studies have shown that surprising visual
events can elicit a so-called “visual mismatch negativity”
response (Kimura, Schröger, Czigler, & Ohira, 2010;
Kimura, Widmann, & Schröger, 2010; see Stefanics et al.,
2014, für eine Rezension), although the effects of perceptual
expectation are not always separable from stimulus adap-
tation in these designs. It is possible that these seemingly
inconsistent results are attributable to a difference in how
expectations were elicited in different studies and whether
the predictive relationship was task relevant (Richter &
de Lange, 2019; Larsson & Schmied, 2012).
Expectations Modulate Perception at Early Sensory
Processing Stage
It is still debated whether expectations modulate percep-
tion by enhancing early sensory signals (Cheadle et al.,
2015; Kok et al., 2012; Wyart et al., 2012) or whether this
modulation is restricted to later decision-related stages
(Rungratsameetaweemana et al., 2018; Bang & Rahnev,
2017). Our current findings support the notion that ex-
pectations modulate the perceptual decision-making
process at an early sensory stage, as substantiated by both
the temporal and spatial profiles of the expectation mod-
ulations we observed. Speziell, the observed modu-
latory effect of expectation coincided with the trailing
image onset and was short-lived (d.h., um 400 ms),
suggesting a modulation during the sensory processing
of the bottom–up input. Zusätzlich, source localization
showed that object-selective sensory regions (in particu-
lar, occipito-temporal cortex in both hemispheres) war
most strongly modulated, supporting the idea that top–
down expectations modulate activity of neuronal popula-
tions representing the sensory stimuli.
Beachten Sie, dass, in addition to modulating sensory pro-
Prozesse, expectations might also modulate later processes
such as setting the decision criterion. As the current
study used an oddball detection task where oddballs oc-
curred randomly throughout the experiment, we were un-
able to address whether expectations modulate decisional
stages beyond sensory processing. Außerdem, the use
of clearly visible object images instead of threshold-level
Reize (vgl. Bang & Rahnev, 2017) resulted in ceiling per-
formance for all participants, thus minimizing our op-
portunities to observe any modulation in decision criterion.
daher, future studies using threshold-level stimuli
may address whether expectations modulate decisional
stages in addition to modulation of sensory processes.
Violations of Sensory Expectations Result in
Stronger Endogenous Alpha-Band Oscillations
When an image stimulus was presented unexpectedly com-
pared with when it was presented as expected, a power in-
crease was observed in band-limited low-frequency activity
overlapping largely with the alpha band. It has recently
been proposed that alpha-band oscillations emerge from
recurrent interactions in a hierarchical network, in which
the higher-order region tries to predict the signal received
Zhou et al.
699
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
T
F
/
Ö
N
0
5
M
A
j
2
0
2
1
from the lower-order region (Alamia & VanRullen, 2019).
In der Tat, stimulus-induced alpha oscillations have been ob-
served when computing the visual impulse response func-
tion to randomly varying (unpredictable) visual input
(VanRullen & McDonald, 2012). In contrast with the “ca-
nonical microcircuits” model of predictive coding that em-
phasized the feedback prediction carrier role of alpha-band
oscillations (Bastos et al., 2012), Alamia and VanRullen
(2019) proposed that alpha-band traveling waves occur in
both the feedforward (during visual stimulation) and back-
ward (in the absence of visual input) directions. It is well
conceivable that the unexpected condition required a larger
amount of recurrent activity cycles as the inconsistency be-
tween sensory expectation and input requires an update of
sensory expectations, therefore leading to stronger power
in the alpha band. At present, this proposal is speculative,
Jedoch, and in need of further empirical evidence.
Alternativ, the power difference in alpha-band oscil-
lations might also be interpreted as a stronger alpha
power suppression for the expected stimuli. It has been
proposed that alpha oscillations gate information process-
ing by inhibiting task-irrelevant brain regions, and this in-
hibition is modulated by attention ( Jensen & Mazaheri,
2010). It has been reported that statistical regularities
can bias attention toward the regular input (Zhao, Al-
Aidroos, & Turk-Browne, 2013). daher, one might hy-
pothesize that the observed difference between expected
and unexpected trials may stem from stronger attentional
engagement for the expected stimuli. Although partici-
pants only paid limited attention to the stimuli, as they
were engaged in an oddball detection task that was de-
signed to minimize the task relevance of the nonoddball
Bilder, we cannot fully rule out the contribution of atten-
tion to the observed neural difference.
Alpha power in the neutral condition, in which all trail-
ing images were equally expected, was intermediate
between expected and unexpected images but did not
significantly differ from these conditions (Figure 3D).
We believe that this null result may be because of a lack
of statistical power, given that this difference is expected
to be more subtle than the difference between expected
and unexpected trials (Ramachandran et al., 2017).
Although we observed significant expectation modula-
tion of oscillatory activity in the alpha band, we did not
see any activity modulation in the high-frequency range.
This appears at odds with the proposal that low- Und
high-frequency oscillations reflect predictions and predic-
tion errors, jeweils (Bastos et al., 2012). Ein Faktor
that may have precluded our observing such an effect is
the stimulus set we used. Although we observed an in-
crease in high-frequency power time-locked to our stim-
uli, this reflected a broadband evoked response, eher
than the narrow-band gamma oscillatory activity that is
typically seen as the feedforward information carrier in
the predictive coding framework. Trotzdem, our observation
was consistent with previous reports that narrow-band
gamma oscillations are elicited by only certain types of
Reize (Hermes, Müller, Wandell, & Winawer, 2015).
Future research may be able to shed light on the stimulus
(In)dependence of high-frequency oscillatory activity for
expectation violations.
To summarize, our current study demonstrates that
perceptual expectations modulate perception at an early
stage during sensory processing. Wichtig, this modu-
latory effect of expectation is expressed by a power in-
crease of low-frequency oscillatory activity in response
to an unexpected stimulus. Our findings extend our un-
derstanding on how perceptual expectations are imple-
mented by the human brain.
Danksagungen
This work was supported by the European Union Horizon
2020 Programm (ERC Starting Grant 678286, “Contextvision”
awarded to F. P. D. L.), the Chinese Scholarship Council
(CSC20170800036 awarded to Y. J. Z.), die Niederlande
Organization for Scientific Research Vidi grant (NWO 016.
Vidi.185.137 awarded to S. H.), and the Beatriu de Pinòs 2017-
BP-00213 (AGAUR, awarded to A. P. B.). We thank Mats van Es,
Jan-Mathijs Schoffelen, Matthias Fritsche, and Eelke Spaak for
helpful discussions.
Reprint requests should be sent to Ying Joey Zhou, Donders
Institute for Brain Cognition and Behaviour, Radboud
Universität, 198328 Kapittelweg 29, Nijmegen, Gelderland 6525
EN, Die Niederlande, oder per E-Mail: ying.joey.zhou@gmail.com.
VERWEISE
Alamia, A., & VanRullen, R. (2019). Alpha oscillations and
traveling waves: Signatures of predictive coding? PLoS
Biology, 17, e3000487.
Alink, A., Schwiedrzik, C. M., Kohler, A., Singer, W., &
Muckli, L. (2010). Stimulus predictability reduces responses
in primary visual cortex. Zeitschrift für Neurowissenschaften, 30,
2960–2966.
Arnal, L. H., Wyart, V., & Giraud, A.-L. (2011). Transitions in
neural oscillations reflect prediction errors generated in
audiovisual speech. Naturneurowissenschaften, 14, 797–801.
Bang, J. W., & Rahnev, D. (2017). Stimulus expectation alters
decision criterion but not sensory signal in perceptual
decision making. Wissenschaftliche Berichte, 7, 17072.
Bastos, A. M., Usrey, W. M., Adams, R. A., Mangun, G. R., Fries,
P., & Friston, K. J. (2012). Canonical microcircuits for
predictive coding. Neuron, 76, 695–711.
Batterink, L. J., Reber, P. J., Neville, H. J., & Paller, K. A. (2015).
Implicit and explicit contributions to statistical learning.
Journal of Memory and Language, 83, 62–78.
Bauer, M., Stenner, M.-P., Friston, K. J., & Dolan, R. J. (2014).
Attentional modulation of alpha/beta and gamma oscillations
reflect functionally distinct processes. Zeitschrift für
Neurowissenschaften, 34, 16117–16125.
Bonaiuto, J. J., Meyer, S. S., Little, S., Rossiter, H., Callaghan, M. F.,
Dick, F., et al. (2018). Lamina-specific cortical dynamics in
human visual and sensorimotor cortices. eLife, 7, e33977.
Brady, T. F., Konkle, T., Alvarez, G. A., & Oliva, A. (2008). Visual
long-term memory has a massive storage capacity for object
Einzelheiten. Verfahren der Nationalen Akademie der Wissenschaften,
USA., 105, 14325–14329.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436.
700
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 4
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
F
T
.
/
Ö
N
0
5
M
A
j
2
0
2
1
Cheadle, S., Egner, T., Wyart, V., Wu, C., & Summerfield, C.
(2015). Feature expectation heightens visual sensitivity
during fine orientation discrimination. Journal of Vision,
15, 14.
Cohen, M. X. (2014). Analyzing neural time series data:
Theory and practice. Cambridge, MA: MIT Press.
de Lange, F. P., Heilbron, M., & Kok, P. (2018). How do
expectations shape perception? Trends im kognitiven Bereich
Wissenschaften, 22, 764–779.
Faul, F., Erdfelder, E., Lang, A. G., & Buchner, A. (2007).
G*Power 3: A flexible statistical power analysis program for
the social, behavioral, and biomedical sciences. Behavior
Research Methods, 39, 175–191.
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Serie B,
Biological Sciences, 360, 815–836.
Brutto, J., Kujala, J., Hämäläinen, M., Timmermann, L.,
Schnitzler, A., & Salmelin, R. (2001). Dynamic imaging of
coherent sources: Studying neural interactions in the human
Gehirn. Verfahren der Nationalen Akademie der Wissenschaften,
USA., 98, 694–699.
Haegens, S., Händel, B. F., & Jensen, Ö. (2011). Top–down
controlled alpha band activity in somatosensory areas
determines behavioral performance in a discrimination task.
Zeitschrift für Neurowissenschaften, 31, 5197–5204.
Haegens, S., Nácher, V., Luna, R., Romo, R., & Jensen, Ö.
(2011). α-Oscillations in the monkey sensorimotor network
influence discrimination performance by rhythmical
inhibition of neuronal spiking. Verfahren des Nationalen
Akademie der Wissenschaften, USA., 108, 19377–19382.
Hermes, D., Müller, K. J., Wandell, B. A., & Winawer, J. (2015).
Gamma oscillations in visual cortex: The stimulus matters.
Trends in den Kognitionswissenschaften, 19, 57–58.
Jensen, O., Bonnefond, M., & VanRullen, R. (2012). Ein
oscillatory mechanism for prioritizing salient unattended
Reize. Trends in den Kognitionswissenschaften, 16, 200–206.
Jensen, O., & Mazaheri, A. (2010). Shaping functional
architecture by oscillatory alpha activity: Gating by inhibition.
Grenzen der menschlichen Neurowissenschaften, 4, 186.
Johnston, P., Robinson, J., Kokkinakis, A., Ridgeway, S., Simpson,
M., Johnson, S., et al. (2017). Temporal and spatial localization
of prediction-error signals in the visual brain. Biologisch
Psychologie, 125, 45–57.
Kaposvari, P., Kumar, S., & Vogels, R. (2018). Statistical
learning signals in macaque inferior temporal cortex.
Hirnrinde, 28, 250–266.
Kim, R., Seitz, A., Feenstra, H., & Shams, L. (2009). Testen
assumptions of statistical learning: Is it long-term and
implicit? Neurowissenschaftliche Briefe, 461, 145–159.
Kimura, M., Schröger, E., Czigler, ICH., & Ohira, H. (2010). Human
visual system automatically encodes sequential regularities of
discrete events. Zeitschrift für kognitive Neurowissenschaften, 22,
1124–1139.
Manahova, M. E., Mostert, P., Kok, P., Schoffelen, J. M., &
de Lange, F. P. (2018). Stimulus familiarity and expectation
jointly modulate neural activity in the visual ventral stream.
Zeitschrift für kognitive Neurowissenschaften, 30, 1366–1377.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Zeitschrift für Neurowissenschaften
Methoden, 164, 177–190.
Mayer, A., Schwiedrzik, C. M., Wibral, M., Singer, W., & Melloni, L.
(2016). Expecting to see a letter: Alpha oscillations as
carriers of top-down sensory predictions. Hirnrinde,
26, 3146–3160.
Meyer, T., & Olson, C. R. (2011). Statistical learning of visual
transitions in monkey inferotemporal cortex. Verfahren
der Nationalen Akademie der Wissenschaften, USA., 108,
19401–19406.
Meyer, T., Ramachandran, S., & Olson, C. R. (2014). Statistical
learning of serial visual transitions by neurons in monkey
inferotemporal cortex. Zeitschrift für Neurowissenschaften, 34,
9332–9337.
Michalareas, G., Vezoli, J., van Pelt, S., Schoffelen, J. M.,
Kennedy, H., & Fries, P. (2016). Alpha–beta and
gamma rhythms subserve feedback and feedforward
influences among human visual cortical areas. Neuron,
89, 384–397.
Murray, S. O., Kersten, D., Olshausen, B. A., Schrater, P., &
Wald, D. L. (2002). Shape perception reduces activity in
human primary visual cortex. Verfahren des Nationalen
Akademie der Wissenschaften, USA., 99, 15164–15169.
Nolte, G. (2003). The magnetic lead field theorem in the quasi-
static approximation and its use for magnetoencephalography
forward calculation in realistic volume conductors. Physik
in Medicine and Biology, 48, 3637–3652.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis of
MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869.
Ramachandran, S., Meyer, T., & Olson, C. R. (2017). Prediction
suppression and surprise enhancement in monkey
inferotemporal cortex. Journal of Neurophysiology, 118,
374–382.
Richter, D., Ekman, M., & de Lange, F. P. (2018). Suppressed
sensory response to predictable object stimuli throughout
the ventral visual stream. Zeitschrift für Neurowissenschaften, 38,
7452–7461.
Richter, D., & de Lange, F. P. (2019). Statistical learning
attenuates visual activity only for attended stimuli. eLife, 8,
e47869.
Robinson, J. E., Breakspear, M., Jung, A. W., & Johnston, P. J.
(2018). Dose-dependent modulation of the visually evoked
N1/N170 by perceptual surprise: A clear demonstration of
prediction-error signalling. European Journal of
Neurowissenschaften. https://doi.org/10.1111/ejn.13920.
Kimura, M., Widmann, A., & Schröger, E. (2010). Human visual
Rungratsameetaweemana, N., Itthipuripat, S., Salazar, A., &
system automatically represents large-scale sequential
regularities. Gehirnforschung, 1317, 165–179.
Kok, P., Jehee, J. F. M., & de Lange, F. P. (2012). Less is more:
Expectation sharpens representations in the primary visual
Kortex. Neuron, 265–270.
Larsson, J., & Schmied, A. T. (2012). fMRI repetition suppression:
Neuronal adaptation or stimulus expectation? Zerebral
Kortex, 22, 567–576.
Li, N., & DiCarlo, J. J. (2008). Unsupervised natural experience
rapidly alters invariant object representation in visual cortex.
Wissenschaft, 321, 1502–1507.
Li, N., & DiCarlo, J. J. (2010). Unsupervised natural visual
experience rapidly reshapes size-invariant object representation
in inferior temporal cortex. Neuron, 67, 1062–1075.
Serences, J. T. (2018). Expectations do not alter early sensory
processing during perceptual decision-making. Zeitschrift für
Neurowissenschaften, 38, 5632–5648.
Sadaghiani, S., & Kleinschmidt, A. (2016). Brain networks
and α-oscillations: Structural and functional foundations of
cognitive control. Trends in den Kognitionswissenschaften, 20,
805–817.
Sherman, M. T., Kanai, R., Seth, A. K., & VanRullen, R. (2016).
Rhythmic influence of top–down perceptual priors in the
phase of prestimulus occipital alpha oscillations. Zeitschrift für
Cognitive Neuroscience, 28, 1318–1330.
Stefanics, G., Kremláček, J., & Czigler, ICH. (2014). Visual
mismatch negativity: A predictive coding view. Grenzen in
Human Neuroscience, 8, 666.
Zhou et al.
701
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
.
T
F
/
Ö
N
0
5
M
A
j
2
0
2
1
Summerfield, C., & de Lange, F. P. (2014). Expectation in
perceptual decision making: Neural and computational
mechanisms. Nature Reviews Neurowissenschaften, 15, 745–756.
Swettenham, J. B., Muthukumaraswamy, S. D., & Singh, K. D.
(2009). Spectral properties of induced and evoked gamma
oscillations in human early visual cortex to moving and
stationary stimuli. Journal of Neurophysiology, 102,
1241–1253.
Thut, G., Nietzel, A., Brandt, S. A., & Pascual-Leone, A. (2006).
Alpha-band electroencephalographic activity over occipital
cortex indexes visuospatial attention bias and predicts visual
target detection. Zeitschrift für Neurowissenschaften, 26, 9494–9502.
Turk-Browne, N. B., Scholl, B. J., Johnson, M. K., & Chun, M. M.
(2010). Implicit perceptual anticipation triggered by statistical
learning. Zeitschrift für Neurowissenschaften, 30, 11177–11187.
van Kerkoerle, T., Self, M. W., Dagnino, B., Gariel-Mathis, M. A.,
Poort, J., van der Togt, C., et al. (2014). Alpha and gamma
oscillations characterize feedback and feedforward
processing in monkey visual cortex. Verfahren der
Nationale Akademie der Wissenschaften, USA., 111, 14332–14341.
VanRullen, R., & McDonald, J. S. (2012). Perceptual echoes at
10 Hz in the human brain. Aktuelle Biologie, 22, 995–999.
Worden, M. S., Foxe, J. J., Wang, N., & Simpson, G. v. (2000).
Anticipatory biasing of visuospatial attention indexed by
retinotopically specific alpha-band electroencephalography
increases over occipital cortex. Zeitschrift für Neurowissenschaften,
20, RC63.
Wyart, V., Nobre, A. C., & Summerfield, C. (2012). Dissociable
prior influences of signal probability and relevance on visual
contrast sensitivity. Proceedings of the National Academy
of Sciences, USA., 109, 3593–3598.
Zhao, J., Al-Aidroos, N., & Turk-Browne, N. B. (2013). Attention
is spontaneously biased toward regularities. Psychological
Wissenschaft, 24, 667–677.
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
Ö
H
w
T
N
T
P
Ö
:
A
/
D
/
e
D
M
ich
F
R
T
Ö
P
M
R
C
H
.
P
S
ich
l
D
v
ich
R
e
e
R
C
T
C
.
M
H
A
ich
e
R
D
.
u
C
Ö
Ö
M
C
N
/
J
A
Ö
R
T
C
ich
C
N
e
/
–
A
P
R
D
T
ich
3
2
C
l
4
e
6
–
9
P
1
D
F
2
0
/
1
3
3
2
2
/
5
4
5
/
6
Ö
9
C
1
N
/
_
A
1
_
8
0
6
1
1
5
3
1
3
1
5
P
/
D
J
Ö
B
C
j
N
G
_
u
A
e
_
S
0
T
1
Ö
5
N
1
0
1
7
.
P
S
D
e
F
P
e
B
M
j
B
e
G
R
u
2
e
0
S
2
T
3
/
J
F
T
/
.
Ö
N
0
5
M
A
j
2
0
2
1
702
Zeitschrift für kognitive Neurowissenschaften
Volumen 32, Nummer 4