Patterns of Brain Activity Supporting Autobiographical
Memory, Prospection, and Theory of Mind, Und
Their Relationship to the Default Mode Network
R. Nathan Spreng1 and Cheryl L. Grady1,2
Abstrakt
■ The ability to rise above the present environment and reflect
upon the past, the future, and the minds of others is a fundamen-
tally defining human feature. It has been proposed that these
three self-referential processes involve a highly interconnected
core set of brain structures known as the default mode network
(DMN). The DMN appears to be active when individuals are en-
gaged in stimulus-independent thought. This network is a likely
candidate for supporting multiple processes, but this idea has
not been tested directly. We used fMRI to examine brain activity
during autobiographical remembering, prospection, and theory-
of-mind reasoning. Using multivariate analyses, we found a com-
mon pattern of neural activation underlying all three processes in
the DMN. Zusätzlich, autobiographical remembering and pros-
pection engaged midline DMN structures to a greater degree
and theory-of-mind reasoning engaged lateral DMN areas. A func-
tional connectivity analysis revealed that activity of a critical node
in the DMN, medial prefrontal cortex, was correlated with activity
in other regions in the DMN during all three tasks. We conclude
that the DMN supports common aspects of these cognitive behav-
iors involved in simulating an internalized experience. ■
EINFÜHRUNG
Remembering oneʼs past (autobiographical memory),
imagining oneʼs future (prospection), and imagining
the thoughts and feelings of others (theory of mind) Sind
similar in that they all involve simulating an experience
that is distinct from stimulus-driven behavior (Buckner &
Carroll, 2007). Two domains, autobiographical memory
and prospection, have shown substantial behavioral and
neural overlap. They are correlated in their phenomeno-
logical detail (DʼArgembeau & Van der Linden, 2004) Und
temporal distribution (Spreng & Levine, 2006). Weiter-
mehr, they appear to share a common set of active brain
Bereiche (Addis, Wong, & Schacter, 2007; Szpunar, Watson, &
McDermott, 2007; Okuda et al., 2003). Focal lesions to the
medial-temporal lobes have resulted in co-occurring defi-
cits in memory and imagining novel scenarios (Hassabis,
Kumaran, Vann, & Maguire, 2007; Tulving, 1985).
Autobiographical memory, prospection, and theory of
mind are also interrelated (Perner, 2000; Suddendorf &
Corballis, 1997). Simulation-based accounts of theory-
of-mind resemble the process of remembering oneʼs past
and imagining oneʼs future. Theory-of-mind reasoning
can involve imagining the mindset of others and simulat-
ing their experience in order to understand their internal
Staaten (Blakemore & Decety, 2001). In all three instances,
the individual must recreate an experience outside of
their personal present moment (Buckner & Carroll,
1Rotman Research Institute at Baycrest, Toronto, Ontario, Kanada,
2Universität von Toronto, Ontario, Kanada
2007). This trio of abilities emerges at the same time de-
velopmentally, at about 3.5 Jahre alt (Perner, Kloo, &
Gornik, 2007; Suddendorf, 1999). Autobiographical
memory and theory of mind are not necessarily mutually
dependent in adults, Jedoch, as shown in two patients
with retrograde amnesia who performed well on theory-
of-mind tasks (Rosenbaum, Stuss, Levine, & Tulving,
2007). Trotzdem, a quantitative summary of multiple
independent observations has found substantial neural
overlap between these three cognitive domains (Spreng,
Mar, & Kim, 2009). To date, no study has directly tested
the idea of common neural underpinnings for autobio-
graphical memory, prospection, and theory-of-mind rea-
soning within the same individuals.
There is evidence of overlap between the functional
neuroanatomy of these domains and that of the default
mode network (DMN; Spreng et al., 2009; Buckner &
Carroll, 2007). The DMN is an intrinsically organized core
set of brain regions observed to be more metabolically ac-
tive at “rest.” The network includes midline frontal and
parietal structures, medial and lateral temporal lobes, Und
lateral parietal cortex (Buckner, Andrews-Hanna, & Schacter,
2008; Mazoyer et al., 2001; Raichle et al., 2001; Shulman
et al., 1997). Behaviorally, the DMN may underlie stimulus-
independent thought (McGuire, Paulesu, Frackowiak, &
Frith, 1996). Self- and other-attribution judgments have
been associated with medial prefrontal cortex (MPFC) ac-
tivity (Mitchell, Banaji, & Macrae, 2005; Gusnard, Akbudak,
Shulman, & Rachel, 2001), a reliable node of the DMN.
daher, the process of projecting the self into the past,
© 2009 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 22:6, S. 1112–1123
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
.
F
T
/
.
.
Ö
N
1
8
M
A
j
2
0
2
1
the future, or into the minds of others may all rely on the
DMN (Buckner et al., 2008; Buckner & Carroll, 2007). A
separate network of regions that is “anticorrelated” to
the DMN appears to be related to stimulus-driven atten-
tion that is common with many cognitive tasks. This net-
work is typically referred to as the task-positive network
(Fox et al., 2005). These two networks account for large-
scale patterns of covariance across hundreds of neuro-
Bildgebungsstudien (Toro, Fuchs, & Paus, 2008).
In der vorliegenden Studie, Wir haben die Hypothese getestet, dass
adults who are engaged in autobiographical remember-
ing, prospection, and theory-of-mind reasoning would
activate the same set of brain regions across all three
Bedingungen. This hypothesis is consistent with the idea
that the DMN mediates these processes. We used a multi-
variate method of analysis known as partial least squares
(PLS) to identify where activity covaried across the three
experimental conditions in the entire brain, and to see
if this pattern was consistent with the DMN. To provide
additional evidence that the DMN, and not the task-
positive network, was engaged across the task conditions,
we conducted a functional connectivity analysis of brain
regions known to be part of these networks (Fox et al.,
2005). We did so by examining the correlations of the
DMN and task-positive seed regions across the three
experimental conditions and the rest of the brain.
METHODEN
Teilnehmer
fMRI scans of 16 healthy participants (Durchschnittsalter = 25.9 Jahre,
SD = 4.0; 5 men; 1 left-handed) were acquired while
they engaged in cued autobiographical remembering,
prospection, and theory-of-mind reasoning. Teilnehmer
had normal or corrected-to-normal vision and no history
of neurological or psychiatric illness. Years of education
averaged 18.0 Jahre (SD = 1.8). Participants gave written
informed consent before inclusion in the study, which was
approved by the ethics committee of Baycrest Centre.
Stimuli and Procedure
The study comprised three experimental conditions and
a sensorimotor control condition. In the experimental
Bedingungen, participants engaged in cued autobiographi-
cal remembering, prospection, and theory-of-mind rea-
soning. Four 10-min runs, each consisting of 8 trials of
each condition, for a total of 32 trials per condition, war
presented and counterbalanced across participants. Der
study paradigm was designed to match visual input but vary
the task demands.
In all experimental trials, a photograph and associated
word were used as cues and were presented on the
same screen for 4 Sek. Der 96 photographs were selected
from the International Affective Picture System (IAPS: Lang,
Bradley, & Cuthbert, 2005). Image valence ranged from
positive to neutral to negative while depicting people in
various situations. By IAPS standards, the range of images
was moderate (without graphic depictions of sex or muti-
lation): pleasure mean = 5.35 (SD = 1.8), arousal mean =
5.29 (SD = 1.5), and dominance mean = 5.36 (SD = 2.0).
Matched for valence, arousal, and dominance, the stimuli
were randomized within and unique to each run and were
counterbalanced across conditions.
Each photograph contained one or more persons, solch
as a family around a dinner table. Below the photograph, A
related cue word, such as “Family” was displayed. Nach dem
4-sec viewing period, participants were presented with the
instruction screen for 10 Sek. On the instruction screen,
participants were directed to use the photograph and the
word as a cue to remember an event, imagine a future
event or imagine the thoughts and feelings of someone
in the photograph. Zum Beispiel: (A) “Remember a time
when you went out with your family” (remembering), (B)
“Imagine a time you will go out with your family” (prospec-
tion), oder (C) “Imagine what the father in the picture was
thinking and feeling” (theory-of-mind reasoning). To en-
sure sufficient time for autobiographical memory retrieval,
a 10-sec interval was selected based on electrophysiolog-
ical evidence (c.f. Conway, Pleydell-Pearce, Whitecross, &
Sharpe, 2003). Nächste, participants rated how clearly they
engaged in the recollection or imagining (1 = Very clearly,
2 = Somewhat clearly, 3 = Not clearly at all; siehe Abbildung 1).
In the control condition, participants viewed 32 scram-
bled photographs that were meaningless but matched the
experimental photographs for perceptual input. The words
“Control Stimulus” were displayed underneath each image
(vgl. Burianova & Grady, 2007). This was followed by a but-
ton press instruction (z.B., “Press the key that corresponds
to the letter ‘P’.”). As in the experimental conditions, Re-
sponses were made by pressing 1, 2, oder 3 on a keypad
(z.B., “1 = N, 2 = B, 3 = P”). Responses were recorded
using a fiber-optic response pad placed in the participantsʼ
right hand. One participantʼs responses were not recorded.
All participants went through a three-step training ses-
sion prior to scanning that included instruction, üben,
und Diskussion. Training included an instructional conver-
sation about autobiographical events being unique to time
and place, envisaging future scenarios, and imagining the
thoughts and feelings of others. After completing five prac-
tice trials for each condition, participant compliance was
assessed. This included discussing the ability to engage
the task for 8 Zu 10 Sek, ease of engagement, and making
a clarity judgment at the end of the 10-sec interval. Six par-
ticipants requested to complete another practice run to
clarify what was expected of them. All participants found
the task to be easy and a majority spontaneously reported
it to be enjoyable after the scanning session.
Image Preprocessing and Analysis
Brain imaging data were acquired with a 3.0-T Siemens
scanner with a standard head coil. Anatomical scans
Spreng and Grady
1113
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
/
.
F
.
T
.
Ö
N
1
8
M
A
j
2
0
2
1
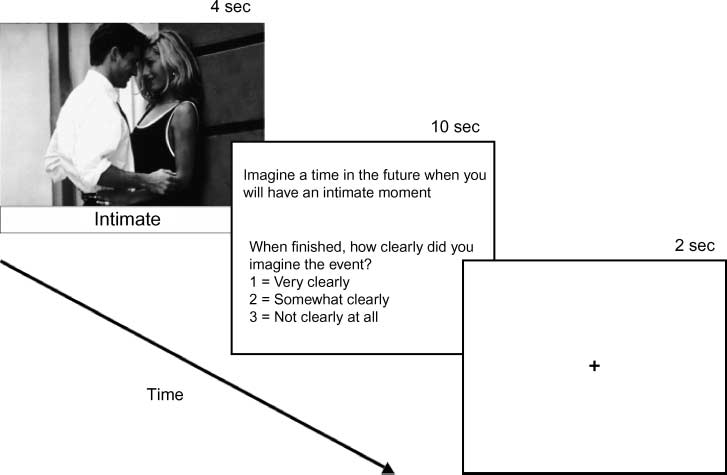
Figur 1. Example of an
experimental trial. Teilnehmer
viewed a photograph for 4 Sek.
Nächste, a condition-specific
instruction with three clarity
ratings was displayed for 10 Sek.
The intertrial interval (fixation)
War 2 Sek. In the example,
the question relates to the
future condition. Other
participants may have seen
the photograph paired in the
autobiographical memory
condition with the instruction,
“Remember a time you had
an intimate moment”; oder, Die
theory-of-mind condition with
the instruction, “Imagine what
the woman was thinking and
feeling.” Each photograph was
presented only once per
participant. Photographs were
presented in their original color.
were acquired using a T1-weighted volumetric MRI (TR =
2000 ms, TE = 2.63 ms, 160 axial slices, 1.0 mm thick,
FOV = 256 mm). Four runs of BOLD functional scans were
acquired with a T2*-weighted EPI pulse sequence (TR =
2000 ms, TE = 30 ms, flip angle = 70°, FOV = 200 mm,
64 × 64 acquisition matrix), consisting of 30 contiguous,
5-mm-thick axial slices.
Images were reconstructed in AFNI (Cox, 1996), Wo
they were first corrected for respiration and heart rate.
Slice-timing was corrected to the first slice; nächste, Bewegung
was corrected using a 3-D Fourier transform interpolation
using a functional volume that minimized the amount of
motion to less than 2 mm. Endlich, participantsʼ images
were spatially normalized to MNI space and smoothed
using a 6-mm FWHM Gaussian filter in SPM5. The result-
ing voxel size was 4 × 4 × 4 mm.
To analyze the fMRI data, we used spatio-temporal PLS,
a multivariate functional neuroimaging analysis tool de-
signed to identify whole-brain patterns of activity that
are correlated with task (McIntosh, Chau, & Protzner,
2004). PLS assessed the covariance between brain voxels
(BOLD signal) and the experimental design to identify a
limited number of components (latent variables, LVs)
that optimally relate the two data matrices. This data-
driven approach was similar to a principal component
analysis and determined whole-brain patterns of activity.
Unlike the standard univariate analysis that examines
the independent activity of any single voxel, PLS is ca-
pable of detecting brain-wide systems that covary with
the experimental design. With PLS, the hemodynamic re-
sponse is not modeled explicitly, in the sense of defining
a canonical response; stattdessen, the algorithm determines
the response that best characterizes the conditions of
interest.
When applying PLS analysis to event-related data, Die
results provide a set of brain regions that are reliably re-
lated to the task conditions at eight poststimulus TRs
(16 Sek) for each LV (McIntosh et al., 2004). Activity at
each time point for each event was normalized to activity
in the first TR of the trial, defined here as the onset of the
instructions. As we were primarily interested in activity
related to the conditions, and not in stimulus-related
activity to the cue photographs/words per se, we exam-
ined those neuroimages beginning with the instructions
to engage in autobiographical remembering, prospec-
tion, theory-of-mind reasoning, or the control task. Der
TRs sampling brain activity during the interval of time
spent viewing the photographs were not included in
the analysis (see Discussion for potential carryover effects
of picture viewing). In PLS, each brain voxel is given a
weight, known as a salience (akin to a component load-
ing). This salience is proportional to the covariance of
activity with the task contrast at each time point on each
LV. Multiplying the salience by the BOLD signal value in
that voxel and summing the product across all voxels
gives a brain score for each participant for each time
point on a given LV (like a component score). Diese
scores can be used to examine differences in brain activ-
ity across conditions, as greater activity in brain areas
with positive (or negative) saliences on an LV will yield
positive (or negative) mean scores for a given condition
over each time point. The plots of mean brain scores
show the pattern of activity across the brain, expressed
over the 16-sec period, and are analogous to hemody-
namic response functions typically plotted for individual
Gehirnregionen.
The significance of each LV was determined by per-
mutation testing (McIntosh, Bookstein, Haxby, & Grady,
1114
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 6
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
.
.
F
/
T
.
Ö
N
1
8
M
A
j
2
0
2
1
1996), verwenden 500 permutations. In a second, independent
step, the reliability of the saliences for the brain voxels,
characterizing each pattern identified by an LV, was de-
termined by bootstrap resampling to estimate the stan-
dard errors for each voxel (Efron & Tibshirani, 1985).
For each voxel at each TR, the salience/standard error
Verhältnis, or bootstrap ratio (BSR), was calculated and is
roughly equivalent to a Z-score. Peak voxels with a BSR
greater than four were considered reliable and approxi-
mates a p value of 10−4. Clusters containing at least 10
reliable voxels were extracted, and a local maximum for
each cluster was defined as the voxel with a BSR higher
than any other voxel in a 2-cm cube centered on that
voxel. Although most regions showed reliable activations
across multiple time points, results are reported from
the bootstrap ratios for the fourth and fifth TR (d.h., bei
8–12 sec after instruction onset) as a representative index
of brain activity in time. In focusing on these early and
middle time points, brain activity associated with clarity
judgments, made at approximately 9 sec after instruction
onset (see Results), would be minimized. Locations of
the maxima are reported in the stereotaxic coordinates
of MNI space.
In order to directly examine task covariance in neural
activity at the voxel level, changes in BOLD signal inten-
sity for each TR were extracted for peak voxels from the
task PLS analysis from the first LV in DMN regions (loca-
tions specified by Buckner et al., 2008, Tisch 1).
A common way of assessing activity in the DMN and in
the task-positive network is to measure resting state
functional connectivity using MPFC and inferior parietal
Sulkus (IPS) seed regions, jeweils (Fox et al.,
2005). In order to test the hypothesis that the three
experimental conditions are associated with the DMN,
and not the task-positive network, we sought to identify
functionally connected networks of brain regions asso-
ciated with the experimental conditions using “seed”
PLS (McIntosh, 1999). Seed PLS correlates activity in a
given region of interest with activity in all other brain
voxels to assess the functional connectivity of this region
and then examines how these correlation patterns differ
across experimental conditions (McIntosh, 1999). Wir
evaluated whether seeds in MPFC (0, 52, −16) and IPS
(24, −64, 48) could reinstate the intrinsically organized
default and task-positive networks in a task-related
functional connectivity analysis. The coordinates from
Fox et al. (2005) were transformed from Talairach to
MNI using the Lancaster transformation (Lancaster et al.,
2007). These seeds were selected because the regions
are reliably engaged across conditions (Spreng et al.,
2009; Toro et al., 2008) and functionally connected to the
respective networks at rest (Fox et al., 2005). In seed PLS,
correlation values represent the relationship between
activity in a voxel and the whole-brain pattern identified in
the analysis. We considered any region to be part of either
the DMN or the task-positive network if it was within 2 cm
of the average coordinate from a resting state functional
Tisch 1. Peak Regions Associated with LV1
Location
Hemis BA
X
j
z
Ratio
Past, Future, Other > Control
STS/MTG
Pre-SMA
ACC
RSC
MPFC
MFG
IFG (orbitalis)
Kleinhirn
PHC
Putamen
Putamen
IFG (triangularis)
PCC
MFG
MTG
ES G
IPL
Frontal pole
L
LR
L
L
L
L
L
R
R
R
L
L
L
L
L
L
L
21 −68 −36 −4
8
−4
32 −12
20
32
29
−8 −44
9 −12
6 −44
56
16
52
20
4
44
52
47 −44
36 −12
20 −88 −36
32 −60
NA
30
NA
20
NA −20
45 −48
8
4
28
8 −52
24
21 −64 −12 −16
20 −60 −12 −28
39 −40 −76
10 −12
64
40
24
4
12
8
4
24
36
9.4
8.6
8.0
7.8
7.8
7.7
7.4
7.1
7.1
6.9
6.7
6.6
6.4
6.3
6.2
6.1
5.9
5.9
LR
23
−4 −48
Control > Past, Future, Other
MT+
MT+
Insula
Thalamus
Paracentral lobule
Kleinhirn
Precentral gyrus
Postzentraler Gyrus
R
L
L
R
L
L
R
L
19
28 −60 −16 −15.3
19 −48 −80
8 −12.5
13 −48
0
−4 −11.2
NA
20 −28
31 −16 −40
0
48
−8 −72 −44
52
0
1 −32 −32
28
72
NA
4
−9.3
−6.5
−6.3
−6.0
−5.7
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
/
.
.
F
.
T
Clusters reported at TR5 with a minimum of 10 voxels with a BSR
greater than 4. Hemis = hemisphere; L = left; R = right; BA = Brodmannʼs
Bereich; ACC = anterior cingulate cortex; HC = hippocampus; IFG =
inferior frontal gyrus; IPL = inferior parietal lobule; ITG = inferior tem-
poral gyrus; MCC = middle cingulate cortex; MFG = middle frontal gyrus;
MOG = middle occipital gyrus; MPFC = medial prefrontal cortex; MT+ =
ventral occipital cortex; MTG = middle temporal gyrus; PCC = posterior
cingulate cortex; PCu = precuneus; PHC = parahippocampal gyrus;
RSC = retrosplenial cortex; SOG = superior occipital gyrus; SMA = sup-
plementary motor area; SMG = supramarginaler Gyrus; STS = superior
temporal sulcus. Coordinates are in MNI space.
Ö
N
1
8
M
A
j
2
0
2
1
Konnektivitätsstudie (Fox et al., 2005) and a large-scale
meta-analysis of coactivation patterns from 792 neuro-
imaging papers (Toro et al., 2008) and within the same neuro-
anatomical structure (z.B., posterior cingulate cortex).
Spreng and Grady
1115
ERGEBNISSE
There were no differences in the proportion of clarity
ratings between the experimental conditions (all ps >
.05). The majority of responses were rated as “very
clearly” (75–78%) followed by “somewhat clearly” (16–
22%). Participants rated a small proportion of responses
(2–5%) as “not clearly at all.” The pattern of brain activity
remained unchanged when the data from responses that
were rated as “not clearly at all” were excluded from the
Analyse. daher, we included all trials for each condi-
tion in the fMRI analysis. Response times were signifi-
cantly faster for the control condition (mean = 2.9 Sek,
SD = 3.0 Sek) than the autobiographical memory (mean =
9.0 Sek, SD = 1.4 Sek), prospection (mean = 9.4 Sek,
SD = 1.1 Sek), and theory-of-mind (mean = 8.4 Sek, SD =
1.5 Sek). The experimental conditions did not differ [F(3,
60) = 66.41, P < .001, post hoc Tukeyʼs HSD p < .05].
Autobiographical memory, prospection, and theory-
of-mind shared a common pattern of neural activity that
accounted for 81.42% of the covariance in the data and
was reliably differentiated from the control condition
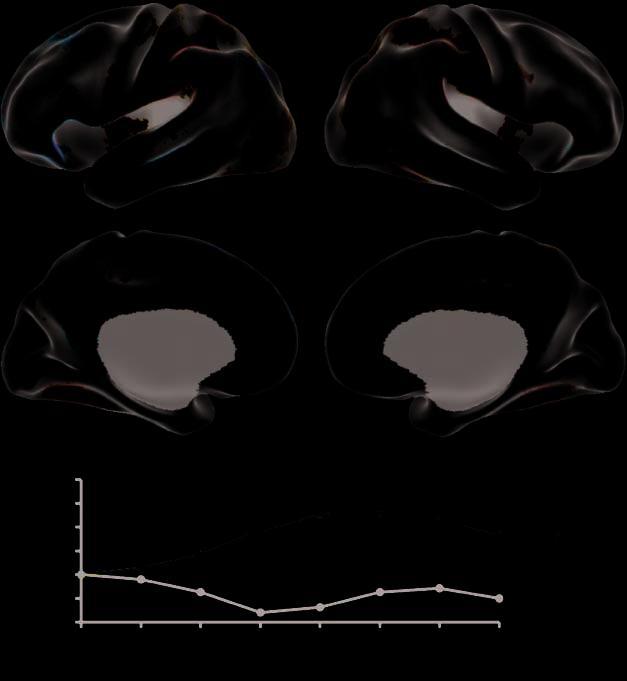
(LV1; p = .002; Figure 2). This common pattern included
activation of the midline structures in frontal and parietal
lobes; left-lateralized activation of inferior frontal gyrus,
temporal pole, middle temporal gyrus, superior tem-
poral sulcus and inferior parietal lobule; and activation
of right medial-temporal lobe (Table 1). In contrast, the
control task was associated with increased bilateral activ-
ity in lateral parietal (including IPS) and occipital cortex
(Table 1). Burianova and Grady (2007) observed a similar
pattern of activation with the same control task, which is
similar to the task-positive network revealed by resting
state functional connectivity (Fox et al., 2005), only with-
out dorsolateral prefrontal cortex. A second significant
pattern of brain activity dissociated autobiographical mem-
ory and prospection from theory-of-mind, accounting for
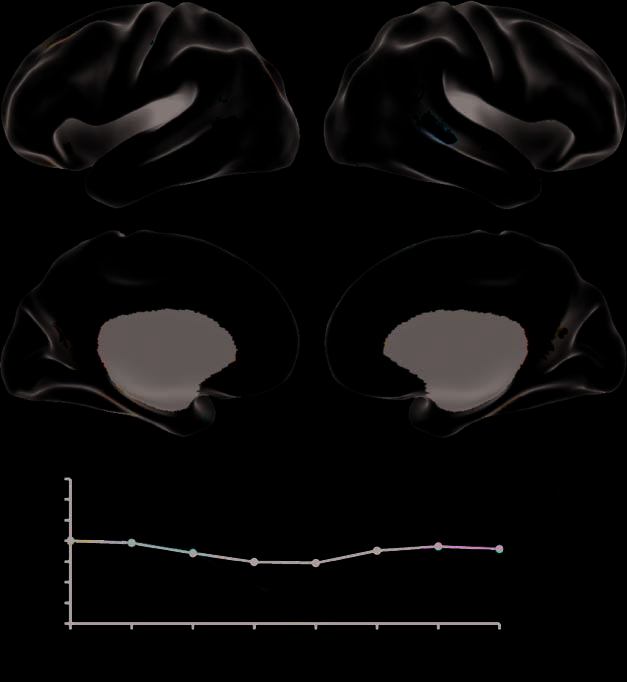
13.75% of the covariance in the data (LV2; p = .056; Fig-
ure 3). Autobiographical memory and prospection demon-
strated relatively greater frontal and parietal midline activity,
along with substantial involvement of the hippocampal
formation (Table 2). Theory-of-mind, in contrast, engaged
the lateral temporal regions and right temporo-parietal
junction (Table 2).
A critical question is whether these brain regions in-
volved in autobiographical memory, prospection, and
theory-of-mind are part of the DMN. The pattern of
activity associated with LV1 and autobiographical mem-
ory, prospection, and theory of mind closely resembles
the DMN, although the spatial extent of MPFC is smaller
and the overall pattern is less bilateral than previous char-
acterizations of the DMN (e.g., Buckner et al., 2008;
Figure 2). Indeed, many areas of activity associated with
LV1 and LV2 were located within 2 cm of published values
Figure 2. LV1 shows a
functional differentiation
between autobiographical
memory (Past), prospection
(Future), and theory-of-mind
(Other) conditions
(represented as warm colors)
and control (cool colors) at
TR4. Left medial and lateral
views are on the left. The
temporal brain score chart
(bottom) conveys changes in
brain activity related to task
over time. For each LV, mean
brain scores (summed scores
of activity across the entire
brain of each participants and
averaged across participants)
show the divergence between
experimental and control
conditions over time (eight
2-sec TRs), and are analogous
to hemodynamic response
functions typically plotted for
individual brain regions.
1116
Journal of Cognitive Neuroscience
Volume 22, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
6
e
-
1
p
1
d
1
f
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
o
1
c
2
n
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
2
8
0
2
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
t
.
/
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 3. LV2 differentiates
autobiographical memory (Past)
and prospection (Future)
(represented as cool colors)
from theory-of-mind (Other)
(warm colors), where control
is roughly at zero at TR5.
Temporal brain scores (bottom)
demonstrated maximal
differentiation at the fifth TR.
Left medial and lateral views
are on the left.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
6
e
-
1
p
1
d
1
f
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
o
1
c
2
n
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
2
8
0
2
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
.
f
.
t
o
n
1
8
M
a
y
2
0
2
1
for nodes within the DMN (Toro et al., 2008; Fox et al.,
2005). The conditions differentially activated DMN for LV2
(Figure 3). Remembering the past or thinking about the
future preferentially activated midline and medial temporal
lobe structures, whereas theory-of-mind reasoning prefer-
entially activated lateral temporal and parietal regions.
Thus, brain activity across the conditions showed overlap,
as well as some functional specialization, within the DMN.
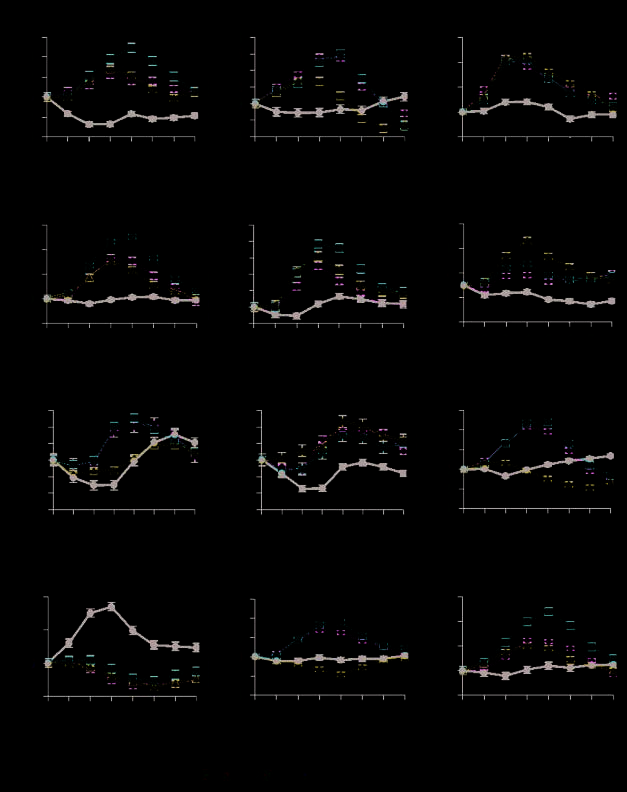
Hemodynamic response function plots of peak voxels
in DMN regions from LV1 (Figure 4A–I) demonstrated a
nearly uniform pattern of covariance, dissociating auto-
biographical memory, prospection, and theory of mind
from control in dorsal and ventral MPFC, bilateral lateral
temporal cortex, bilateral inferior parietal lobule, and right
parahippocampus. The theory-of-mind condition was not
dissociated from control in left parahippocampus (Fig-
ure 4G). The contribution of theory of mind to the pattern
of activity in posterior cingulate also was less robust (Fig-
ure 4I). A recently published formal graph-analytic network
analysis of resting state functional connectivity correlations
revealed five distinct yet highly interconnected hubs: MPFC,
bilateral IPL, and two in posterior cingulate, one rostral (−2,
−38, 36) and one caudal (−1, −53, 32) (Buckner et al., 2009;
coordinates in MNI space). Response function plots of these
two hubs within posterior cingulate cortex (Figure 4K, L)
demonstrated theory of mind covarying with autobiograph-
ical memory and prospection in only the caudal region of
posterior cingulate. Both regions reliably contributed to the
pattern of activity observed in LV1. However, only the ros-
tral portion was reliably involved in the pattern identified by
LV2 (BSR = −10.3) and was associated with autobiograph-
ical memory and prospection. The caudal region was not
(BSR = −2.4).
Both seed PLS analyses revealed a significant pattern
of functional connectivity ( p = .002) that was reliable
for MPFC (a key node in the DMN) and IPS (a key node
in the task-positive network) based on a 99% confidence
interval, for all three experimental conditions. In each
experimental condition, measures of overall brain activity
were significantly correlated with activity in the seed
voxel across subjects: in the MPFC seed analysis, r =
.87 for autobiographical memory, r = .88 for prospec-
tion, and r = .74 for theory-of-mind; in the IPS seed anal-
ysis, r = .81 for autobiographical memory, r = .95 for
prospection, and r = .85 for theory of mind. The hemo-
dynamic response function plots for the two seed voxels
are shown in Figure 4B and J.
If thinking about the past, future, or the thoughts and
feelings of others was associated with the DMN then the
MPFC seed should be correlated with core regions of the
DMN (Buckner et al., 2008) in these three conditions.
This was indeed the case—the MPFC seed was correlated
with activity in the left frontal pole, left inferior parietal
lobule, medial parietal cortex, bilateral medial temporal
Spreng and Grady
1117
Table 2. Peak Regions Associated with LV2
DISCUSSION
Location
Hemis BA
x
y
z
Ratio
Other > Past, Future
STS/MTG
Temporo-parietal junction
STS
Past, Future > Other
PCC
PCC
RSC
PHC
Fusiform gyrus
PHC
HC
ACC
Frontal pole
Kleinhirn
MFG
Thalamus
Kleinhirn
IFG (orbitalis)
PHC
Head of caudate
Brainstem
R
R
L
LR
LR
L
L
L
R
L
LR
L
R
L
R
R
L
R
L
L
21/22
48 −36 −4
39
48 −48
22 −52 −48
20
12
8.0
7.2
6.6
31 −4 −36
40 −14.5
31 −8 −60
24 −12.0
29 −8 −44
8 −11.5
36 −24 −40 −20 −10.7
19 −32 −80
44 −10.1
30
8 −40
4 −9.6
NA −20 −20 −24 −8.1
32 −4
10 −16
40
64
4 −8.1
12 −7.8
NA
16 −44 −52 −7.3
6 −24
16
52 −7.0
NA
NA
4 −8
12 −6.8
44 −72 −44 −6.8
47 −32
24 −8 −6.6
36
28 −40 −12 −6.4
NA −12
12 −4 −6.0
NA −4 −20 −28 −5.4
lobes, and bilateral inferior temporal gyri (Tisch 3). Der
only DMN regions not included in this pattern of connec-
tivity were the right lateral parietal cortex and right frontal
pole. Alternativ, if thinking about the past, future, oder der
thoughts and feelings of others was associated with the
task-positive network, functional connectivity would be
expected within the core regions of this network, wie zum Beispiel
anterior insula, frontal eye fields, supplementary motor
Bereich, dorsolateral prefrontal and ventral occipital cortex.
Jedoch, within the task-positive network, activity in the
IPS voxel was only correlated with contralateral IPS region
and left ventral occipital cortex (Tisch 3). No other core re-
gions of this network were significantly correlated. Stattdessen,
the IPS seed was functionally associated with some DMN
Regionen, including medial parietal cortex, as well as medial
and lateral temporal cortex (Tisch 3). Both seeds demon-
strated functional correlations with regions outside of the
networks as well, such as cerebellum, cuneus, and occipital
Kortex (Tisch 3). Gesamt, the seed PLS analyses indicated
more engagement of the DMN than the task-positive net-
work during the experimental conditions.
In this study, we investigated the neural mechanisms
underlying autobiographical memory, prospection, Und
theory of mind, and determined whether these domains
rely upon a pattern of brain activity operating within the
DMN. We used PLS analysis of fMRI data to examine
whole-brain patterns of activity associated with thinking
about the past, the future, and the thoughts and feelings
of others. We then used seed PLS to examine the func-
tional connectivity of brain regions that have previously
been shown to be functionally associated with the DMN
and the task-positive network while participants are at
ausruhen (Fox et al., 2005).
Autobiographical memory, prospection, and theory of
mind shared a common pattern of brain activity, inkl-
ing midline structures in the frontal and parietal lobes.
This is consistent with brain activity related to self- Und
other-referential thought (DʼArgembeau et al., 2008;
Johnson et al., 2006; Northoff & Bermpohl, 2004). Addi-
tionally, activation was observed in right medial temporal
lobe and left-lateralized activation in inferior frontal
gyrus, middle frontal gyrus, temporal pole, lateral tem-
poral lobe, and inferior parietal lobule. Vor allem, we found
activation of these DMN regions in the experimental
Bedingungen, relative to the control condition, despite the
fact that our participants may have reverted back to the
default mode after completing the easy and rapidly per-
formed sensorimotor control task. Daher, autobiographical
Erinnerung, prospection, and theory-of-mind tasks may acti-
vate DMN regions over and above spontaneous default
Aktivität. The observed regions may support the genera-
tive processing related to each of the experimental condi-
tionen, including semantic retrieval (Binder, Desai, Graves,
& Conant, in press; Martin & Chao, 2001); the recollection
of generic scripts that frame the remembered or imagined
scene; and the integration of sensorimotor modalities com-
prising these scenes. Gesamt, the functional coordination
of these heteromodal regions may be sufficient for pro-
ducing self-relevant stimulus-independent thought.
Autobiographical memory and prospection demon-
strated greater frontal and parietal midline activity,
relative to theory-of-mind. Vor allem, there was also sub-
stantial involvement of the hippocampal formation for
autobiographical memory and prospection: This may
be due to the temporal extension of the self and greater
recombination of contextualized event details involved
in remembering and prospection (Hassabis, Kumaran, &
Maguire, 2007; Schacter & Addis, 2007; Moscovitch, Nadel,
Winocur, Gilboa, & Rosenbaum, 2006). Im Gegensatz, theory-
of-mind reasoning uniquely activated superior temporal
sulcus and right temporo-parietal junction, which may be
preferentially engaged in mental and biological inferen-
cing more specific to this domain (Gallagher & Frith, 2003;
Sachsen & Kanwisher, 2003). Alternately, the medial/ lateral
dissociation may be due to the allocation of attention to
internal representations versus external stimuli (Buckner
1118
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 6
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
.
T
.
F
.
/
Ö
N
1
8
M
A
j
2
0
2
1
Figur 4. BOLD signal
response function plots.
The magnitude of
hemodynamic response
function change is on the
y-axis; TRs are on the x-axis.
Bootstrap ratios (BSR) Sind
from LV1 at TR4. A and C–I
are peak LV1 voxels in default
mode regions. B is the DMN
seed region, contiguous to
the peak ventral MPFC voxel
(−4, 48, −16), not pictured.
As indicated by the BSR, alle
voxels were highly reliable and
remain so over multiple time
points; Jedoch, some of these
Regionen (B, E, F, G) did not
meet the 10-voxel cluster size
cutoff for reporting in Table 1.
J is the task-positive seed
region. K and L are discrete
cortical network hubs in
rostral posterior cingulate
Kortex (rosPCC) and caudal
posterior cingulate cortex
(cauPCC) identified by graph
Analyse (Buckner et al., 2009).
Please refer to Table 1 Notiz
for abbreviations.
et al., 2008). Das ist, participants engaged in memory and
prospection may have used the cue to access internal rep-
resentations, whereas in the theory-of-mind condition, Par-
ticipants may have needed to retrieve more information
about the photographs in order to imagine the mental
states of the depicted people.
Although autobiographical memory, prospection, Und
theory-of-mind are types of stimulus-independent
thought, and appear to engage the DMN, there is little
consensus on whether the DMN is actually responsible
for these kinds of processing. To further complicate the
issue, monkeys show DMN activity (Vincent et al., 2007)
but may have little concept of time (Roberts, 2002); In
Kontrast, children with rudimentary autobiographical
Erinnerung, prospection, and theory-of-mind skills do not
have a fully developed DMN (Fair et al., 2008). Studying
evolutionary changes to the frontal poles and lateral parietal
Kortex, and investigating developmental maturation of
brain characteristics such as frontal white matter myelina-
tion, could resolve these issues eventually. Trotzdem,
current functional neuroimaging methods are sufficient
for examining the hypothesis that stimulus-independent
thought, such as autobiographical memory, prospection,
and theory-of-mind, share a common neural basis in
humans within the DMN. Our task-related functional con-
nectivity analysis was capable of recapitulating most of the
DMN, thereby supporting this hypothesis. Absent was right
lateral parietal cortex, which is hypothesized to be involved
in heteromodal integration between external salience and
internal representations (Corbetta, Patel, & Shulman, 2008),
a task demand predominant in only the theory-of-mind
condition (LV2). The right frontal pole was also absent from
the connectivity pattern, which is surprising considering
that damage to this region can result in impaired perspec-
tive taking (Stuss, Gallup, & Alexander, 2001) and episodic
Erinnerung (Wheeler, Stuss, & Tulving, 1997). The task-positive
Netzwerk, Jedoch, was not associated with autobiograph-
ical memory, prospection, or theory-of-mind. Stattdessen, Die
results from the seed PLS analyses suggest that IPS, a key
node of the task-positive network, can in fact be correlated
with areas of the DMN when engaged in structured in-
trospective behaviors, even though the two networks are
“anticorrelated” when functional connectivity is assessed
during the resting state (Fox et al., 2005). This disparity
Spreng and Grady
1119
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
.
/
.
.
T
F
Ö
N
1
8
M
A
j
2
0
2
1
Tisch 3. Functional Connectivity of the MPFC and rIPS Associated with Autobiographical Memory, Prospection, and Theory-of-Mind
MPFC Seed
IPS Seed
fc-Region
Hemis
BA
X
j
z
Ratio
fc-Region
Hemis
BA
X
j
z
Ratio
Default Mode Regions
LR
7
0 −68
7.8
PCu
LR
7
0 −60
44
7.2
PCu
PCC
HC
HC
ITG/MTG
ES G
Frontal pole
IPL
36
28
31/23
−8 −40
NA
NA
−36 −28 −12
36 −20 −16
20/21 −64 −16 −20
20
10
39
48 −16 −28
−8
56
−48 −60
8
24
L
L
R
L
R
L
L
12.1
6.6
8.3
6.3
4.9
6.4
5.3
PHC
PHC
MTG
IPS
MT+
Task-positive Regions
Other Regions
Kleinhirn
SOG
Cuneus
Precentral gyrus
Precentral gyrus
MCC
L
LR
LR
R
R
R
NA
−16 −60 −16
6.2
Kleinhirn
17
18
6
6
24
4 −88
−4 −84
52
0
52 −12
8 −16
0
12
28
28
40
Kleinhirn
7.1 MOG
5.5
Cuneus
LR
Cuneus
IFG
Brainstem
SMG
8.1
6.9
7.8
Lingual gyrus
R
R
L
L
L
L
R
R
L
L
L
R
L
28 −20 −28 −12
34
32
0 −20
6.9
5.8
21
60 −28 −12
9.4
7 −20 −64
37 −48 −60
48
4
14.7
6.6
NA −28 −48 −24
NA
36 −48 −28
6.6
8.3
19 −36 −76
20
12.0
17
18
44
−4 −72
24 −60
36
4
8
8
28
NA −12 −28 −28
40 −60 −24
18 −28 −76
36
−8
8.7
6.5
7.8
6.4
5.1
5.1
fc-Regions are functionally connected regions in the network. Regions labeled as DMN or task-positive are within 2 cm of published coordinates for
this network and within the same brain structure.
suggests that regions comprising the default mode and
task-positive networks may not be mutually inhibitory,
or may be a consequence of the functional connectivity
methodology (such as regressing out global mean signal;
Murphy, Birn, Handwerker, Jones, & Bandettini, 2009).
Jedoch, the absence of a positive association between
IPS and the experimental conditions in the task PLS analysis
limits strong conclusions.
No previous experiment has directly examined the
relations among autobiographical memory, prospection,
and theory-of-mind. One study compared cued autobio-
graphical recollection from personal photos against a
theory-of-mind type control task where participants
imagined scenarios from strangerʼs photographs, inkl-
ing the thoughts and feelings of people in the photos
(Gilboa, Winocur, Grady, Hevenor, & Moscovitch, 2004).
The results from their univariate analysis of autobio-
graphical memory closely resembled the midline and
medial-temporal regions found here in LV2.1 Univariate
Analyse, Jedoch, does not detect the optimal relationship
between brain activity and conditions unlike the multivari-
ate analysis that was used in this study. Consistent with stud-
ies comparing autobiographical memory and prospection,
a common pattern of neural activity is observed (Schacter
& Addis, 2007), although some subsystems may differ
(Addis, Pan, Vu, Laiser, & Schacter, 2009). Vor allem, Die
present work demonstrated common and differential re-
cruitment between autobiographical, prospection, Und
theory-of-mind. Multivariate methods are not only more
sensitive than the standard General Linear Model in detect-
ing distributed patterns of brain activity (McIntosh et al.,
2004), but are capable of quantifying the amount of covar-
iance associated with neural events that can be explained
by the tasks. We found that the amount of variance in brain
1120
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 6
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
.
.
/
T
.
F
Ö
N
1
8
M
A
j
2
0
2
1
activity shared by the experimental conditions actually was
greater than that which differentiated them, relative to the
sensorimotor control condition. The shared pattern of
neural activity in LV1 and the MPFC seed PLS analyses sup-
port the hypothesis that the same network is responsible
for the core processing of the experimental tasks. Weiter,
LV2, which accounts for less variance in neural activity, may
reveal functional specialization of regions within DMN
subsystems. Resting state functional connectivity analysis
supports this view. Seed activity in posterior cingulate,
medial prefrontal, or lateral parietal cortex are correlated
with the entire default mode network (Fox et al., 2005;
Buckner et al., 2008). Autobiographical memory and pros-
pection, Jedoch, are associated with the hippocampal
formation subsystem (Vincent et al., 2006). Weiter, con-
nectivity with a dorsal MPFC seed region is associated with
the theory-of-mind regions (Buckner et al., 2008). Core re-
gions common to autobiographical memory, prospection,
and theory-of-mind may support the construction and
integration of personally salient information. Subsystems
may provide specialized information to the core DMN.
This may include information from prior experiences
via input from the medial-temporal lobes, as well as elabo-
rated representations of other peopleʼs actions and inten-
tions with input from posterior STS and temporo-parietal
junction.
The common brain activity across the experimental
conditions might simply reflect neural processes driven
by the carryover effect of viewing complex photographs
and/or making a manual rating in response to a mental
Ereignis; Jedoch, this is unlikely for several reasons. Erste,
we limited the time points included in the analysis to
those where self-projection was taking place and not
when photos were being viewed. Zusätzlich, Aktivität
was normalized to the first TR after the photographs
were viewed, reducing the impact of the BOLD signal
related to prior perception. With spatio-temporal PLS,
the progression of BOLD signal change over time is ana-
lyzed. In focusing our interpretation on the fourth and
fifth TR, the impact of BOLD signal associated with photo-
graph viewing was further reduced, although may still
account for some covariance. Zweite, the shared pattern
of brain activation is also unlikely to be related to the clarity
ratings to any great extent. Most studies of autobiograph-
ical memory include a behavioral rating, such as amount
of detail, immediately following the recollective interval.
Participant awareness of an impending rating is unlikely
to confound the phenomenological experience and asso-
ciated neural activity of these processes prior to ratings
actually being made, although these processes undoubt-
edly influence the rating itself. The absence of ratings,
Jedoch, would eliminate a means to assess compliance.
Außerdem, in this study, ratings occurred, on average,
9 sec after the participants were instructed to begin re-
collecting, planning, or thinking about the thoughts and
feelings of others. Infolge, BOLD signals related to
clarity ratings fell outside the interpreted analysis.
Dritte, Burianova and Grady (2007) used a similar para-
digm and found the first LV dissociated experimental
Bedingungen (autobiographical, episodic, and semantic
memory retrieval) from a sensorimotor control condition
identical to the one used here. In their experiment, com-
plex photographs were also viewed and a behavioral
rating was made in the experimental conditions. Im
control condition, scrambled images were also presented
and a manual response was made. Wichtig, the re-
gions identified by Burianova and Grady as common
across retrieval conditions showed little overlap with
the regions identified here as common to autobiograph-
ical memory, prospection, and theory of mind. Jedoch,
both experiments found increased activity across ex-
perimental conditions in left lateral inferior and middle
frontal gyri (Brodmannʼs areas 47, 8). These regions are
involved in cognitive control (Vincent, Kahn, Snyder,
Rachel, & Buckner, 2008; Brass, Derrfuss, Forstmann,
& von Cramon, 2005), and thus, are likely to be involved
across multiple cognitive domains. Taken together, Die
evidence from both studies, as well as the high rates of
participant compliance (as indicated by the clarity ratings),
makes it likely that the shared pattern of covariance (seen
after the instructions are presented but before the clarity
ratings are made) is related to the shared processes of the
experimental conditions themselves.
The current set of analyses provides converging evidence
that DMN regions are engaged with specific cognitive behav-
iors. These behaviors may not be limited to those observed
Hier, but may also include related processes such as moral
decision-making (Greene, Sommerville, Nystrom, Darley, &
Cohen, 2001), fictional narrative simulation (Mar & Oatley,
2008; Mar, 2004), or mind-wandering (Christoff, Gordon,
Smallwood, Schmied, & Schooler, 2009; Mason et al., 2007). In
order to determine the neural (In)dependence of these
processes, Jedoch, neuropsychological investigations into
co-occurring autobiographical and interpersonal deficits with
compromised functional integrity of the midline, and lateral
parietal and lateral temporal cortex are also necessary.
Our analyses were designed to investigate the neural
mechanisms underlying autobiographical memory, pros-
pection, and theory-of-mind, and to determine if these
domains rely upon a pattern of brain activity operating
within the DMN. The results provide the first direct evi-
dence to support previous independent observations
that projection of the self into the past, the future, oder
the minds of others engages a common set of brain
regions largely overlapping with the DMN (Spreng et al.,
2009; Buckner & Carroll, 2007). Task-related functional
connectivity analysis revealed reliable correlations within
the core regions of the DMN, and only two nodes in the
task-positive network, suggesting that the DMN was
engaged during our experimental conditions, but not the
task-positive network. The present study does not directly
compare the experimental tasks with “rest.” Although pre-
and postscan questionnaires have helped clarify resting
state cognition (z.B., Andrews-Hanna, Huang, Reidler, &
Spreng and Grady
1121
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
.
T
F
.
.
/
Ö
N
1
8
M
A
j
2
0
2
1
Buckner, 2008; Mason et al., 2007), we remain unsure what
people do while resting. In order to circumvent this issue,
novel techniques and analyses are needed to assess default
cognition and its neural associations. Zum Beispiel, eins
study assessed the overlap between episodic memory
and theory-of-mind tasks, and then determined peak ac-
tivityʼs relationship with the DMN using resting state func-
tional connectivity analysis (Andrews-Hanna, Sachsen, Poulin,
& Buckner, 2007). The present set of analyses provides
converging evidence that the neuroanatomical correlates
of autobiographical memory, prospection, and theory-of-
mind are shared and that the DMN is involved in the pro-
cessing necessary to carry out these specific cognitive
behaviors. This sharing of neural resources may allow
remembering the past and imagining the future to provide
a “long view” in decision-making, over and above stimulus–
reward driven behavior, thereby facilitating theory-of-mind
and promoting social cooperation (Boyer, 2008).
Danksagungen
We thank the MRI technologists at Baycrest Centre, Magda
Wojtowicz, Dale Stevens, and Karen Spreng for their assistance
with this project. This work was supported by a grant from the
Canadian Institutes of Health Research (MOP14036 to C. L. G.),
by infrastructure support from the Ontario Research Fund and
the Canadian Foundation for Innovation. DR. Grady also is
supported by the Canada Research Chairs program.
Reprint requests should be sent to R. Nathan Spreng, Department
of Psychology, Harvard Universität, 33 Kirkland Street, Cambridge,
MA 02138, oder per E-Mail: nathan.spreng@gmail.com.
Notiz
1. During the preparation of this article, we learned of a highly
relevant paper by Rabin, Gilboa, Stuss, Mar, and Rosenbaum
(2010) who used a very similar paradigm as Gilboa et al., (2004),
but included a common baseline to assess conjunction as well as
differences between autobiographical memory and theory of
Geist. Despite different methods, the authors report a striking
consistency with the present findings.
VERWEISE
Addis, D. R., Pan, L., Vu, M.-A., Laiser, N., & Schacter, D. L. (2009).
Constructive episodic simulation of the future and the past:
Distinct subsystems of a core brain network mediate imagining
and remembering. Neuropsychologie, 47, 2222–2238.
Addis, D. R., Wong, A. T., & Schacter, D. L. (2007).
Remembering the past and imagining the future: Common
and distinct neural substrates during event construction
and elaboration. Neuropsychologie, 45, 1363–1377.
Andrews-Hanna, J. R., Huang, C., Reidler, J., & Buckner, R. L.
(2008). Functional connectivity within the default network
linked to spontaneous internal mentation. Neurowissenschaften
Meeting Planner. Washington, Gleichstrom: Society for Neuroscience,
Program No. 483.23.
Andrews-Hanna, J. R., Sachsen, R., Poulin, R., & Buckner, R. L.
(2007). The default system overlaps activation during
theory of mind and episodic memory retrieval tasks.
Neuroscience Meeting Planner. San Diego, CA: Society for
Neurowissenschaften, Program No. 421.17.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L.
(in press). Where is the semantic system? A critical review
and meta-analysis of 120 functional neuroimaging studies.
Hirnrinde, doi:10.1093/cercor/bhp055.
Blakemore, S. J., & Decety, J. (2001). From the perception of
action to the understanding of intention. Nature Reviews
Neurowissenschaften, 2, 561–567.
Boyer, P. (2008). Evolutionary economics of mental time travel?
Trends in den Kognitionswissenschaften, 12, 219–224.
Brass, M., Derrfuss, J., Forstmann, B., & von Cramon, D. Y.
(2005). The role of the inferior frontal junction area in
cognitive control. Trends in den Kognitionswissenschaften, 9, 314–316.
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L. (2008). Der
brainʼs default network: Anatomy, Funktion, and relevance to
Krankheit. Annalen der New York Academy of Sciences, 1124, 1–38.
Buckner, R. L., & Carroll, D. C. (2007). Self-projection and
das Gehirn. Trends in den Kognitionswissenschaften, 11, 49–57.
Buckner, R. L., Sepulcre, J., Talukdar, T., Krienen, F. M., Liu, H.,
Hedden, T., et al. (2009). Cortical hubs revealed by
intrinsic functional connectivity: Mapping, assessment of
Stabilität, and relation to Alzheimerʼs disease. Zeitschrift für
Neurowissenschaften, 29, 1860–1873.
Burianova, H., & Grady, C. L. (2007). Common and unique
neural activations in autobiographical, episodic, and semantic
retrieval. Zeitschrift für kognitive Neurowissenschaften, 19, 1520–1534.
Conway, M. A., Pleydell-Pearce, C. W., Whitecross, S. E., & Sharpe, H.
(2003). Neurophysiological correlates of memory for experienced
and imagined events. Neuropsychologie, 41, 334–340.
Corbetta, M., Patel, G., & Shulman, G. L. (2008). Der
reorienting system of the human brain: From environment
to theory of mind. Neuron, 58, 306–324.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research, 29, 162–173.
Christoff, K., Gordon, A. M., Smallwood, J., Schmied, R., &
Schooler, J. W. (2009). Experience sampling during fMRI
reveals default network and executive system contributions
to mind wandering. Proceedings of the National Academy
of Sciences, USA., 106, 8719–8724.
DʼArgembeau, A., Feyers, D., Majerus, S., Collette, F., Van der
Linden, M., Maquet, P., et al. (2008). Self-reflection across
Zeit: Cortical midline structures differentiate between
present and past selves. Social Cognitive and Affective
Neurowissenschaften, 3, 244–252.
DʼArgembeau, A., & Van der Linden, M. (2004). Phenomenal
characteristics associated with projecting oneself back into the
past and forward into the future: Influence of valence and
temporal distance. Consciousness & Cognition, 13, 844–858.
Efron, B., & Tibshirani, R. (1985). The bootstrap method for
assessing statistical accuracy. Behaviormetrika, 17, 1–35.
Fair, D. A., Cohen, A. L., Dosenbach, N. U. F., Church, J. A.,
Miezin, F. M., Barch, D. M., et al. (2008). The maturing
architecture of the brainʼs default network. Proceedings of
the National Academy of Sciences, USA., 105, 4028–4032.
Fuchs, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Rachel, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
102, 9673–9678.
Gallagher, H. L., & Frith, C. D. (2003). Functional imaging of
“theory of mind”. Trends in den Kognitionswissenschaften, 7, 77–83.
Gilboa, A., Winocur, G., Grady, C. L., Hevenor, S. J., &
Moscovitch, M. (2004). Remembering our past: Funktional
neuroanatomy of recollection of recent and very remote
personal events. Hirnrinde, 14, 1214–1225.
Greene, J. D., Sommerville, R. B., Nystrom, L. E., Darley, J. M.,
& Cohen, J. D. (2001). An fMRI investigation of emotional
engagement in moral judgment. Wissenschaft, 293, 2105–2108.
1122
Zeitschrift für kognitive Neurowissenschaften
Volumen 22, Nummer 6
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
F
.
.
.
/
T
Ö
N
1
8
M
A
j
2
0
2
1
Gusnard, D. A., Akbudak, E., Shulman, G. L., & Rachel, M. E.
(2001). Medial prefrontal cortex and self-referential mental
Aktivität: Relation to a default mode of brain function.
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
98, 4259–4264.
Hassabis, D., Kumaran, D., & Maguire, E. A. (2007). Using
imagination to understand the neural basis of episodic
Erinnerung. Zeitschrift für Neurowissenschaften, 26, 14365–14374.
Hassabis, D., Kumaran, D., Vann, S. D., & Maguire, E. A. (2007).
Patients with hippocampal amnesia cannot imagine new
Erfahrungen. Proceedings of the National Academy of
Wissenschaften, USA., 104, 1726–1731.
Johnson, M. K., Raye, C. L., Mitchell, K. J., Touryan, S. R., Greene, E.
J., & Nolen-Hoeksema, S. (2006). Dissociating medial frontal
and posterior cingulate activity during self-reflection. Sozial
Cognitive and Affective Neuroscience, 1, 56–64.
Lancaster, J. L., Tordesillas-Gutierrez, D., Martinez, M., Salinas,
F., Evans, A., Zilles, K., et al. (2007). Bias between MNI and
Talairach coordinates analyzed using the ICBM-152 brain
Vorlage. Kartierung des menschlichen Gehirns, 28, 1194–1205.
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (2005).
International affective picture system (IAPS): Affektiv
ratings of pictures and instruction manual. Technisch
Report A-6. University of Florida, Gainesville, FL.
Mar, R. A. (2004). The neuropsychology of narrative: Story
comprehension, story production and their interrelation.
Neuropsychologie, 42, 1414–1434.
Mar, R. A., & Oatley, K. (2008). The function of fiction is the
abstraction and simulation of social experience. Perspectives
on Psychological Science, 3, 173–192.
Martin, A., & Chao, L. L. (2001). Semantic memory and the
Gehirn: Structure and processes. Aktuelle Meinung in
Neurobiology, 11, 194–201.
Mason, M. F., Norton, M. ICH., Van Horn, J. D., Wegner, D. M.,
Grafton, S. T., & Macrae, C. N. (2007). Wandering minds: Der
default network and stimulus-independent thought. Wissenschaft,
315, 393–395.
Mazoyer, B., Zago, L., Mellet, E., Bricogne, S., Etard, O., Houdé,
O., et al. (2001). Cortical networks for working memory and
executive functions sustain the conscious resting state in
man. Brain Research Bulletin, 54, 287–298.
McGuire, P. K., Paulesu, E., Frackowiak, R. S. J., & Frith, C. D.
(1996). Brain activity during stimulus independent thought.
NeuroReport, 7, 2095–2099.
McIntosh, A. R. (1999). Mapping cognition to the brain through
neural interactions. Memory, 7, 523–548.
McIntosh, A. R., Bookstein, F. L., Haxby, J. V., & Grady, C. L.
(1996). Spatial pattern analysis of functional brain images
using partial least squares. Neurobild, 3, 143–157.
McIntosh, A. R., Chau, W., & Protzner, A. B. (2004).
Spatiotemporal analysis of event-related fMRI data using
partial least squares. Neurobild, 23, 764–775.
Mitchell, J. P., Banaji, M. R., & Macrae, C. N. (2005). The link
between social cognition and self-referential thought in the
medial prefrontal cortex. Zeitschrift für kognitive Neurowissenschaften,
17, 1306–1315.
Moscovitch, M., Nadel, L., Winocur, G., Gilboa, A., &
Rosenbaum, R. S. (2006). The cognitive neuroscience of
remote episodic, semantic and spatial memory. Current
Opinion in Neurobiology, 16, 179–190.
Murphy, K., Birn, R. M., Handwerker, D. A., Jones, T. B., &
Bandettini, P. A. (2009). The impact of global signal
regression on resting state correlations: Are anti-correlated
networks introduced? Neurobild, 44, 893–905.
Northoff, G., & Bermpohl, F. (2004). Cortical midline structures
and the self. Trends in den Kognitionswissenschaften, 8, 102–107.
Okuda, J., Fujii, T., Ohtake, H., Tsukiura, T., Tanji, K., Suzuki,
K., et al. (2003). Thinking of the future and past: The roles of
the frontal pole and the medial temporal lobes. Neurobild,
19, 1369–1380.
Perner, J. (2000). Memory and theory of mind. In F. ICH. M. Craik
& E. Tulving (Hrsg.), Oxford handbook of memory
(S. 297–312). New York: Oxford University Press.
Perner, J., Kloo, D., & Gornik, E. (2007). Episodic memory
Entwicklung: Theory of mind is part of re-experiencing
experienced events. Infant and Child Development, 16, 471–490.
Rabin, J. S., Gilboa, A., Stuss, D. T., Mar, R. A., & Rosenbaum,
R. S. (2010). Common and unique neural correlates of
autobiographical memory and theory of mind. Zeitschrift
of Cognitive Neuroscience, 22, 1095–1111.
Rachel, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode
of brain function. Proceedings of the National Academy
of Sciences, USA., 98, 676–682.
Roberts, W. A. (2002). Are animals stuck in time? Psychological
Bulletin, 128, 473–489.
Rosenbaum, R. S., Stuss, D. T., Levine, B., & Tulving, E. (2007).
Theory of mind is independent of episodic memory. Wissenschaft,
318, 1257.
Sachsen, R., & Kanwisher, N. (2003). People thinking about
thinking people—The role of the temporo-parietal junction
in “theory of mind”. Neurobild, 19, 1835–1842.
Schacter, D. L., & Addis, D. R. (2007). The cognitive neuroscience
of constructive memory: Remembering the past and imagining
the future. Philosophical Transactions of the Royal Society
of London, Serie B, Biological Sciences, 362, 773–786.
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L., Miezin,
F. M., Rachel, M. E., et al. (1997). Common blood flow
changes across visual tasks: II. Decreases in cerebral cortex.
Zeitschrift für kognitive Neurowissenschaften, 9, 648–663.
Spreng, R. N., & Levine, B. (2006). The temporal distribution of
past and future autobiographical events across the lifespan.
Memory & Cognition, 34, 1644–1651.
Spreng, R. N., Mar, R. A., & Kim, A. S. (2009). Das Gemeinsame
neural basis of autobiographical memory, prospection,
navigation, theory of mind and the default mode: A quantitative
meta-analysis. Zeitschrift für kognitive Neurowissenschaften, 21, 489–510.
Stuss, D. T., Gallup, G. G., & Alexander, M. P. (2001). The frontal
lobes are necessary for “theory of mind”. Gehirn, 124, 279–286.
Suddendorf, T. (1999). The rise of the metamind. In M. C. Corballis
& S. Lea (Hrsg.), The descent of mind: Psychological perspectives
on hominid evolution. Oxford: Oxford University Press.
Suddendorf, T., & Corballis, M. C. (1997). Mental time travel
and the evolution of the human mind. Genetic Social and
General Psychology Monographs, 123, 133–167.
Szpunar, K. K., Watson, J. M., & McDermott, K. B. (2007).
Neural substrates of envisioning the future. Proceedings of
the National Academy of Sciences, USA., 104, 642–647.
Toro, R., Fuchs, P. T., & Paus, T. (2008). Functional coactivation
map of the human brain. Hirnrinde, 18, 2553–2559.
Tulving, E. (1985). Memory and consciousness. Canadian
Psychologie, 26, 1–12.
Vincent, J. L., Kahn, ICH., Snyder, A. Z., Rachel, M. E., & Buckner,
R. L. (2008). Evidence for a frontoparietal control system
revealed by intrinsic functional connectivity. Zeitschrift für
Neurophysiologie, 100, 3328–3342.
Vincent, J. L., Patel, G. H., Fuchs, M. D., Snyder, A. Z., Bäcker, J. T.,
Van Essen, D. C., et al. (2007). Intrinsic functional architecture
in the anaesthetized monkey brain. Natur, 447, 83–86.
Vincent, J. L., Snyder, A. Z., Fuchs, M. D., Shannon, B. J., Andrews,
J. R., Rachel, M. E., et al. (2006). Coherent spontaneous
activity identifies a hippocampal–parietal memory network.
Journal of Neurophysiology, 96, 3517–3531.
Wheeler, M. A., Stuss, D. T., & Tulving, E. (1997). Toward a
theory of episodic memory: The frontal lobes and autonoetic
Bewusstsein. Psychological Bulletin, 121, 331–354.
Spreng and Grady
1123
D
Ö
w
N
l
Ö
A
D
e
D
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
F
R
Ö
M
D
H
Ö
T
w
T
N
P
Ö
:
A
/
D
/
e
M
D
ich
F
T
R
Ö
P
M
R
C
H
.
S
P
ich
l
D
v
ich
e
R
e
R
C
C
T
.
H
M
A
ich
R
e
.
D
u
C
Ö
Ö
M
C
/
N
J
A
Ö
R
C
T
ich
N
C
/
e
A
–
P
R
D
T
ich
2
C
2
l
6
e
–
1
P
1
D
1
F
2
/
1
2
9
2
3
/
9
6
5
/
8
1
4
1
Ö
1
C
2
N
/
1
2
0
7
0
6
9
9
5
2
6
1
4
2
8
/
2
J
Ö
P
C
D
N
.
B
j
2
0
G
0
u
9
e
.
S
T
2
Ö
1
N
2
8
0
2
8
.
S
P
e
D
P
F
e
M
B
j
B
e
G
R
u
2
0
e
2
S
3
T
/
J
T
.
.
.
F
/
Ö
N
1
8
M
A
j
2
0
2
1