Objects and Categories: Feature Statistics and
Object Processing in the Ventral Stream
Lorraine K. Tyler1, Shannon Chiu1, Jie Zhuang1, Billi Randall1,
Barry J. Devereux1, Paul Wright1, Alex Clarke1,
and Kirsten I. Taylor1,2,3
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
Abstrakt
■ Recognizing an object involves more than just visual anal-
yses; its meaning must also be decoded. Extensive research
has shown that processing the visual properties of objects relies
on a hierarchically organized stream in ventral occipitotemporal
Kortex, with increasingly more complex visual features being
coded from posterior to anterior sites culminating in the peri-
rhinal cortex (VR China) in the anteromedial temporal lobe (aMTL).
The neurobiological principles of the conceptual analysis of
objects remain more controversial. Much research has focused
on two neural regions—the fusiform gyrus and aMTL, both of
which show semantic category differences, but of different
types. fMRI studies show category differentiation in the fusiform
gyrus, based on clusters of semantically similar objects, wohingegen
category-specific deficits, specifically for living things, are asso-
ciated with damage to the aMTL. These category-specific defi-
cits for living things have been attributed to problems in
differentiating between highly similar objects, ein Prozess, der
involves the PRC. To determine whether the PRC and the fusi-
form gyri contribute to different aspects of an objectʼs meaning,
with differentiation between confusable objects in the PRC and
categorization based on object similarity in the fusiform, Wir
carried out an fMRI study of object processing based on a feature-
based model that characterizes the degree of semantic simi-
larity and difference between objects and object categories.
Participants saw 388 objects for which feature statistic infor-
mation was available and named the objects at the basic level
while undergoing fMRI scanning. After controlling for the ef-
fects of visual information, we found that feature statistics that
capture similarity between objects formed category clusters
in fusiform gyri, such that objects with many shared features
(typical of living things) were associated with activity in the
lateral fusiform gyri whereas objects with fewer shared features
(typical of nonliving things) were associated with activity in
the medial fusiform gyri. Significantly, a feature statistic reflect-
ing differentiation between highly similar objects, enabling
object-specific representations, was associated with bilateral
PRC activity. These results confirm that the statistical charac-
teristics of conceptual object features are coded in the ventral
stream, supporting a conceptual feature-based hierarchy, Und
integrating disparate findings of category responses in fusi-
form gyri and category deficits in aMTL into a unifying neuro-
cognitive framework. ■
EINFÜHRUNG
Recognizing an object involves not only an analysis of its
visual properties but also the computation of its mean-
ing. The neural system supporting visual analysis has been
characterized as a hierarchical neurobiological system of
increasing feature complexity in occipitotemporal cortex.
Simple visual features are integrated into more complex
feature combinations from visual cortex to anterior
temporal regions along ventral occipitotemporal cortex
(Tanaka, 1996; Ungerleider & Mishkin, 1982). At the apex
of this stream, perirhinal cortex (VR China) is claimed to per-
form the most complex visual feature integrations required
to discriminate between highly similar objects (Murray,
Bussey, & Saksida, 2007; Bussey, Saksida, & Murray, 2002;
Murray & Bussey, 1999).
1University of Cambridge, 2University Hospital Basel, 3Univer-
sity Center for Medicine of Aging, Basel, Schweiz
© 2013 Massachusetts Institute of Technology
The neurobiological principles of the conceptual anal-
ysis of objects remain more controversial, with studies
primarily focusing on category structure (Mahon &
Caramazza, 2009; Martin, 2007; Tyler & Moss, 2001; Chao,
Haxby, & Martin, 1999; McRae, de Sa, & Seidenberg, 1997;
Warrington & Shallice, 1984; Warrington & McCarthy,
1983) and its organizing principles. These are claimed to
include domain or category membership or different
property types that are shared among members of a cate-
gory (z.B., visuell, functional, and motor properties). Object
categories have been associated with two neural regions
in the ventral stream: the fusiform gyrus and the antero-
medial temporal cortex (Martin, 2007; Moss, Rodd,
Stamatakis, Bright, & Tyler, 2005; Tyler et al., 2004;
Humphreys & Forde, 2001; Chao et al., 1999; Warrington
& Shallice, 1984). Evidence for category differentiation in
the fusiform gyrus comes from fMRI studies with healthy
volunteers in which different parts of the fusiform prefer-
entially respond to different object categories such as tools
Zeitschrift für kognitive Neurowissenschaften 25:10, S. 1723–1735
doi:10.1162/jocn_a_00419
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
Ö
9
C
7
N
0
_
0
A
/
_
J
0
Ö
0
C
4
N
1
9
_
A
P
_
D
0
0
B
4
j
1
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
F
/
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
and animals (Chao et al., 1999) whereas neighboring
regions of the lateral occipital complex show little category
selectivity (Op de Beeck, Torfs, & Wagemans, 2008).
Category effects in the ventral stream have also been
observed in studies of neuropsychological patients who
show category-selective deficits. The most frequently re-
ported findings are for category-specific deficits for living
things in response to damage in anteromedial temporal
lobe (aMTL; Moss et al., 2005; Tyler et al., 2004; Humphreys
& Forde, 2001; Warrington & Shallice, 1984). Im Gegensatz,
patients with anterolateral temporal lobe damage have a
generalized semantic impairment and no category-specific
Beeinträchtigung (Noppeney et al., 2007; Rogers et al., 2006;
Moss et al., 2005). This distinction between anteromedial
and anterolateral involvement has been further supported
by neuroimaging studies with healthy volunteers, welche
show that living things preferentially engage the aMTL
(Taylor, Moss, Stamatakis, & Tyler, 2006; Moss et al.,
2005; Tyler et al., 2004).
Category-specific deficits for living things following
damage to the anteromedial temporal cortex have been
attributed to patientsʼ difficulties in differentiating be-
tween highly similar objects (Taylor et al., 2006; Moss
et al., 2005; Tyler et al., 2004). Although patients with
aMTL damage have no difficulty in determining the cate-
gory of an object, they are exceptionally poor at differen-
tiating between similar objects, and this pattern is most
marked for living things, especially animals (Moss et al.,
2005; Tyler et al., 2004; Moss, Tyler, & Jennings, 1997), welche
are among the most highly confusable objects according
to property norm data (Randall, Moss, Rodd, Greer, &
Tyler, 2004; McRae et al., 1997; Keil, 1986; Malt & Schmied,
1984). In patients with category-specific deficits, aMTL
damage tends to be extensive, but one region within it—
the PRC—may be the primary contributor to the deficit
(Kivisaari, Tyler, Monsch, & Taylor, 2012; Tyler et al.,
2004) because this region provides the neural infra-
structure for complex feature integration that enables the
fine-grained differentiation required for distinguishing
between highly similar objects (Barense, Henson, & Graham,
2011; Moss et al., 2005; Tyler et al., 2004). Other findings
support this suggestion: PRC lesions in nonhuman pri-
mates are associated with deficits in the ability to differen-
tiate between highly ambiguous objects (Saksida, Bussey,
Buckmaster, & Murray, 2007; Bussey et al., 2002) Und
patients with aMTL damage including the PRC have dif-
ficulty in complex feature ambiguity tasks (Barense, Gaffan,
& Graham, 2007; Barense et al., 2005).
To determine whether the PRC and the fusiform gyri
contribute to different aspects of an objectʼs meaning
with differentiation between confusable objects in the
PRC and category differentiation in the fusiform, we car-
ried out an fMRI study of object processing based on a
feature-based model that characterizes the degree of
semantic similarity and difference between objects and
object categories. Feature-based models assume that
conceptual representations are componential in nature:
that they are made up of smaller elements of meaning,
referred to as features, properties or attributes. Sie
account for categorization on the assumption that seman-
tic categories are based on feature similarity, although
models differ with respect to the nature of the attributes
considered and the similarity computations they hypothe-
Größe (Schmied & Medin, 1981). Componentiality, although
not universally accepted, is now widely assumed in cog-
nitive psychology (Mirman & Magnuson, 2008; Gotts &
Plaut, 2004; Randall, Moss, Rodd, Greer, & Tyler, 2004;
Cree & McRae, 2003; McRae et al., 1997) and accounts
for behavioral aspects of processing the semantics of
Objekte (Taylor, Devereux, Acres, Randall, & Tyler, 2012;
Randall et al., 2004; Pexman, Holyk, & Monfils, 2003;
McRae et al., 1997). This type of model also has the poten-
tial to capture the characteristics that distinguish objects
from each other and thus enable individuation between
similar objects. Although features that are shared by many
objects provide the basis for categorization, those that are
distinctive of a specific object enable similar objects to be
differentiated from each other (Taylor et al., 2012; Taylor,
Salamoura, Randall, Moss, & Tyler, 2008; Cree & McRae,
2003; McRae & Cree, 2002; Tyler & Moss, 2001).
The model used in this study was based on 2526 fea-
tures derived from a large-scale norming study of 541
concepts (Taylor et al., 2012; McRae, Cree, Seidenberg,
& McNorgan, 2005). In this study, participants generated
verbal feature lists for each concept. Although the listed
Merkmale (z.B., has stripes) are not intended to literally re-
flect all real features of a particular object, the statistical
regularities of these features do reflect systematic statis-
tical regularities we experience in the world that capture
the content and structure of conceptual representations,
provide a basis for categorization, and predict responses
to semantic tasks using both words (Grondin, Lupker, &
McRae, 2009; Randall et al., 2004; McRae et al., 1997) Und
pictures (Clarke, Taylor, Devereux, Randall, & Tyler,
2013; Taylor et al., 2012, Experimente 1 Und 2). Two key
aspects of conceptual representation have been tested
and validated in these cognitive studies and form the basis
of the current study. One important variable is the extent
to which an objectʼs features are shared by many (z.B.,
many animals have fur) or few concepts (z.B., few animals
have stripes). The property norm statistics show that living
Dinge (z.B., Tiere) have many shared and few distinctive
Merkmale, whereas nonliving things (z.B., Werkzeuge) have fewer
shared and relatively more distinctive features (Cree &
McRae, 2003; Tyler & Moss, 2001). The issue is whether
sharedness will be associated with activity in the fusiform,
and if so, whether these property statistics will be asso-
ciated with differentiation within the fusiform. Speziell,
will the differential effects of sharedness overlap with the
medial-lateral distinction in the fusiform such that the
effects of greater sharedness will overlap with the lateral
fusiform, known to be associated with animals (Chao
et al., 1999), and effects of fewer shared features will over-
lap with tool-associated regions of the medial fusiform
1724
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
Ö
9
C
7
N
0
_
0
A
/
_
J
0
Ö
0
C
4
N
1
9
_
A
P
_
D
0
0
B
4
j
1
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
T
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
(Chao et al., 1999)? Das ist, does the feature statistic vari-
able of Sharedness explain fusiform gyrus activity as well
as the living/nonliving variable?
A second feature statistic variable is that of correla-
tional strength or feature co-occurrence, where highly
correlated features (z.B., has eyes and has ears) co-occur
frequently and mutually coactivate, facilitating feature
Integration (McRae et al., 1997; Rosch, Mervis, Gray,
Johnson, & Boyes-Braem, 1976). Property norm statistics
show that living things have more weakly correlated dis-
tinctive features compared with nonliving things (Taylor
et al., 2008; Randall et al., 2004), making them more dif-
ficult to differentiate from other category members. As a
consequence, living things are disadvantaged relative to
nonliving things on those tasks that require differentia-
tion between similar objects. One such task is basic level
identification, which requires differentiating between
similar objects by integrating a conceptʼs distinctive fea-
tures with its shared features. Zum Beispiel, a basic level
naming response cannot be made on the basis of indi-
vidual features such as the shared features has legs
(Hund?) or lives in zoos (elephant?), or the distinctive fea-
ture has stripes (shirt?). Stattdessen, the individual shared
and distinctive features must be integrated together
(has legs + lives in zoos + has stripes) to know that
the concept is, Zum Beispiel, a tiger. This process is facili-
tated by the correlational strength of a conceptʼs distinc-
tive features: Concepts with weakly correlated distinctive
Merkmale, which are more difficult to integrate with the
other object features, place greater demands on the
complex feature integration computations required for
basic level identification. Im Gegensatz, concepts with rela-
tively highly correlated distinctive features are identified
at the basic level more quickly than concepts with weakly
correlated distinctive features (Taylor et al., 2012; Randall
et al., 2004). The issue is whether the PRC, welches ist
claimed to integrate the most complex feature conjunc-
tionen, will be preferentially engaged by these processes.
In the present fMRI study, we used a basic level nam-
ing task because it engages the entire ventral stream
(Tyler et al., 2004) and therefore allows us to determine
whether regions within the stream are involved in pro-
cessing different aspects of an objectʼs meaning. To
determine whether feature statistic variables account
for activity within different regions of the stream, we cor-
related activity with the two conceptual structure vari-
ables Sharedness and Correlation × Distinctiveness
(see Task Design and Materials) after visual effects had
been accounted for. We predicted that these two vari-
ables would differentiate between neural regions where
activity is driven by (A) similarity of conceptual structure
reflecting category structure (d.h., the relative amount of
shared features within a concept) Und (B) differentiation
between similar concepts (Correlation × Distinctiveness,
d.h., the relative extent to which the distinctive features
critical to basic level differentiation are correlated with
other features in the concept).
METHODEN
Teilnehmer
Fifteen healthy, right-handed, native British English
speakers participated in the fMRI study (nine men; mean
age = 24 Jahre, SD = 5 Jahre). The major exclusion
criteria were bilingualism; left-handedness; MR contra-
indications; neurological, psychiatric, or hormonal dis-
orders; dyslexia; and color blindness. All participants
had normal or corrected-to-normal vision, gave informed
consent, and were paid for their participation. The study
was approved by the East of England–Cambridge Central
Research Ethics Committee.
Task Design and Materials
This fMRI study measured the influence of feature statis-
tic variables on BOLD activity associated with basic level
picture naming while controlling for visual variables. Wir
selected all picturable concepts (n = 388) from an angli-
cized version (Taylor et al., 2012) of the McRae et al.
(2005) property norm set. The pictures had high exem-
plarity (d.h., ratings on a 7-point Likert scale, which reflect
the goodness with which the picture represented the
written concept word, with seven reflecting a perfect rep-
resentation). An independent group of 17 healthy indi-
viduals gave a mean rating (±SD) von 5.11 (±0.88),
ensuring that the object pictures are representative of
the concept. The mean naming and concept agreement
for the picture set was 76% Und 82% based on a further
independent sample of 20 healthy participants. The fea-
ture statistic variables were based on standard measures
of “feature distinctiveness” (d.h., 1/[number of concepts
the feature occurs in]) and the correlational strength of
Merkmale (Taylor et al., 2012; Randall et al., 2004; Vigliocco,
Vinson, Lewis, & Garrett, 2004; Tyler & Moss, 2001; McRae
et al., 1997; Rosch et al., 1976) calculated based on the
entire set of 517 anglicized feature norm concepts (Taylor
et al., 2012; McRae et al., 2005). “Feature distinctiveness”
has higher values for more distinctive features such as
has stripes, but lower values for shared features such as
has fur. We are interested in these shared features, seit
they provide the basis for categorization.
From these two standardly used measures, we calcu-
lated two feature statistic indices. The first measure,
Sharedness, is a measure of the degree of sharedness
of the features in a concept (d.h., how often a conceptʼs
features occur in other concepts). For each concept,
Sharedness is defined as 1 minus the square root of the
mean distinctiveness of the conceptsʼ features (Die
square root transformation was applied to reduce the skew
of the distribution). Sharedness has high values for con-
cepts with proportionately more shared features (z.B.,
Tiere) and low values for concepts with proportionately
more distinctive features (z.B., Werkzeuge; siehe Tabelle 1).
Zweite, we calculated Correlation × Distinctiveness
as the slope of the regression of correlational strength
Tyler et al.
1725
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
Ö
9
C
7
N
0
_
0
A
/
_
J
0
Ö
0
C
4
N
1
9
_
A
P
_
D
0
0
B
4
j
1
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
.
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Tisch 1. Characteristics of All Stimuli and Separately for Living and Nonliving Objects: Key Feature Statistic Variables (C×D =
Correlation × Distinctiveness; NOF = Number of Features; NODF = Number of Distinctive Features), Objective Visual Complexity
(Size of JPEG Files), Naming and Concept Agreement, and Naming Latency for Correct Trials (RT)
Alle
Living
Nonliving
Sharedness
C×D
Correlational strengtha
NOF
NODF
Visual complexity ( JPEG file size)
Naming agreement (%)
Concept agreement (%)
RT (correct name, ms)
Mean
0.45
0.79
0.51
12.6
3.9
10.5
76
82
965
SD
0.15
0.84
0.07
3.3
2.7
0.6
28
25
77
Mean
0.53
0.57
0.52
13.1
2.9
10.7
71
75
990
SD
0.12
0.80
0.07
3.5
2.3
0.5
32
30
82
Mean
0.40
0.94
0.50
12.2
4.5
10.4
79
87
951
SD
0.14
0.84
0.07
3.2
2.7
0.6
25
20
75
aMean correlation of shared features within concept.
on distinctiveness values over all features in the concept,
excluding highly distinctive features that occur in only
one or two concepts because their correlational strength
values may be spurious (Taylor et al., 2008, 2012; Cree,
McNorgan, & McRae, 2006). This measure represents
the relative correlational strength of shared versus dis-
tinctive features within a concept. Daher, concepts with
low or negative Correlation × Distinctiveness values have
relatively weakly correlated distinctive compared with
shared features, generating greater demands on complex
conceptual integration processes that bind distinctive
with shared features to enable basic level identification
(siehe Tabelle 1).
We also constructed variables to represent the visual
information present in the pictures using Gabor-filtered
images to capture the spatial position and orientation
of the objects. Related Gabor filter models have been
used to model perceptual processing in the visual system
(Nishimoto et al., 2011; Naselaris, Prenger, Kay, Oliver, &
Gallant, 2009; Kay, Naselaris, Prenger, & Gallant, 2008).
Gray-scale versions of the images were reduced to 153 ×
153 pixels before applying Gabor filters with four orien-
tations (0°, 45°, 90°, 130°) and five spatial frequencies (1,
2, 4, 8, 16 Hz [cycles/image]). Nächste, the set of 388 Gabor-
filtered pictures were vectorized, und das 23,409 pixel ×
388 picture matrix was entered into a PCA using the Matlab
function “princomp” (Mathworks, Natick, MA). Each result-
ing component described part of the variance in pixel
intensity across the set of pictures, with the first compo-
nent describing the strongest common influence on
variance (d.h., overall intensity) and subsequent com-
ponents describing progressively more subtle components
of the variance (difference in intensity between the top
and bottom of the image, center and surround, usw.). Der
first eight components were selected, according to the
Scree test, and together explained 60% of the variance in
image intensity. The loadings of these eight components
on the 388 images were used to model the visual proper-
ties of the pictures.
All pictures, on a white background, were resized to fit
comfortably on a computer screen with the longest axis
spanning maximally 750 pixels horizontally or 550 pixels
vertically (maximum visual angle 12.2° horizontally or 9.0°
vertically) and were saved as JPEG images using identical
compression settings. zusätzlich zu diesen 388 Objekte,
the stimuli in the fMRI study included fixation crosses
(n = 50) and phase-scrambled images of target stimuli
(n = 54; scrambled using the Fourier method in Matlab)
as low level visual baselines.
The items were presented in the same pseudoran-
domized order for each participant. The pseudorandom-
ization ensured that no more than two items from the
same semantic category (z.B., Tier, furniture, vege-
table) or beginning with the same phoneme followed
one another. The pseudorandomization of pictures with
fixation and scrambled images ensured a jittered, geo-
metric distribution of stimulus onset asynchronies for
the picture stimuli, which optimizes detection of BOLD
Aktivität.
fMRI Procedure
Pictures and baseline stimuli were pseudorandomized
and presented in two blocks of approximately equal
Länge. Each stimulus was displayed in the center of a
projection screen in the scanner for 2000 msec followed
by an intertrial interval of 1100 ms. Participants were
instructed to name aloud each picture as quickly and
accurately as possible, to respond to phase-scrambled
images by saying “blur” aloud, and to fixate on a fixation
1726
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 10
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
Ö
9
C
7
N
0
_
0
A
/
_
J
0
Ö
0
C
4
N
1
9
_
A
P
_
D
0
0
B
4
j
1
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
F
T
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
cross without responding. E-Prime software (Psychologie
Software Tools, Sharpsburg, PA) controlled presentation
and timing of stimuli. Participantsʼ spoken responses
were collected during scanning using an OptiMRI noise-
canceling microphone system (Optoacoustics Ltd.,
Moshav Mazoe, Israel). Stutters and no responses were
scored as incorrect. Darüber hinaus, only object names that ex-
actly corresponded to the concept name in the feature
norm study (McRae et al., 2005) were scored as correct
and thus included in the fMRI analyses to ensure that
the corresponding feature statistic data were valid mea-
sures of the processed concept (z.B., although both
“bread” and “loaf” could describe a given object, Sie
may be associated with different nonperceptual and per-
ceptual features).
Participantsʼ response latencies were calculated using
in-house software on the voice recordings made during
the scan. Continuous scanner recordings were filtered
to suppress frequencies over 700 Hz using a Chebyshev
type I filter and split into segments containing naming re-
sponses to individual items. Naming onsets were deter-
mined relative to picture onset using custom software
by finding the first time point where both (A) the root
mean square power exceeded 5 standard deviations
above a pre-object baseline period and (B) this root mean
square power level was exceeded for at least 40 ms. Any
naming latencies less than 500 msec were manually veri-
fied and corrected if necessary. To reduce the influence of
outlying response latencies, we inverse-transformed
the individual RTs (Ratcliff, 1993) then retransformed them
after averaging for each participant to give the harmonic
mean (ms).
Image Acquisition
Scanning was conducted on a 3T Siemens Tim Trio sys-
tem at the MRC Cognition and Brain Sciences Unit,
Cambridge, England. Continuous functional scans were
collected using gradient-echo EPI with 32 Scheiben, 3 mm iso-
tropic voxel dimensions, repetition time = 2 Sek, echo
time = 30 ms, Sichtfeld = 192 × 192 mm, matrix =
64 × 64, flip angle = 78°. T1-weighted anatomical MPRAGE
scans were acquired with repetition time = 2250 ms,
Echozeit = 3 ms, inversion time = 900 ms, field
of view = 256 mm × 240 mm × 160 mm, matrix size =
256 × 240 × 160.
Imaging Analyses
fMRI data were preprocessed and analyzed with SPM5
Software ( Wellcome Trust Centre for Neuroimaging,
www.fil.ion.ucl.ac.uk/spm/software/spm5/) implemented
in Matlab (Mathworks, Natick, MA). Preprocessing com-
prised slice time correction, within-subject realignment
(Bewegungskorrektur), unified spatial normalization, Und
spatial smoothing with an 8-mm FWHM Gaussian
smoothing kernel. Low-frequency noise was removed
using a high-pass filter with a period of 128 sec in the
SPM general linear model (GLM).
Each participantʼs data were analyzed with the GLM
using the canonical haemodynamic response function.
The correctly named pictures, incorrectly named pictures,
and two baseline conditions were modeled as separate
regressors. We also modeled the visual and feature statis-
tic variables as parametric modulators of the regressor for
correctly named pictures: Erste, the eight PCA compo-
nents characterizing the Gabor-filtered images, Dann
Sharedness and finally Correlation × Distinctiveness.
Each modulator was orthogonalized with respect to pre-
vious modulators, ensuring that the effects of the feature
statistic variables were not confounded with the visual
Variablen. We confirmed that the Sharedness and C×D
variables were not correlated with each other (r = .06,
p > .05) and that there were no significant correlations
between any of the eight Gabor PCA variables and either
Sharedness or C×D (max r < .14, all family-wise error
[FEW] p > .05). The model also included the six move-
ment parameters produced by realignment (über) als
nuisance variables.
The GLM in SPM includes implicit masking, which by
default excludes voxels with signal below 80% of the
mean signal over all voxels in the brain. This heuristic
is used to avoid including brain regions with low BOLD
signal because of variations in magnetic susceptibility,
such as the anterior temporal regions under investiga-
tion. Because this heuristic approach may exclude voxels
with low but reliable BOLD signal, we lowered the im-
plicit masking threshold to 10% and then defined reli-
able voxels using a more specific measure of temporal
signal-to-noise ratio (TSNR). We calculated TSNR maps
for each participant by dividing the mean functional im-
age intensity over time at each voxel by its standard de-
viation. We then calculated a group-averaged TSNR map
and defined reliable voxels as those with mean TSNR >
40 (Murphy, Bodurka, & Bandettini, 2007). The group-
averaged TSNR map indicated adequate reliability of sig-
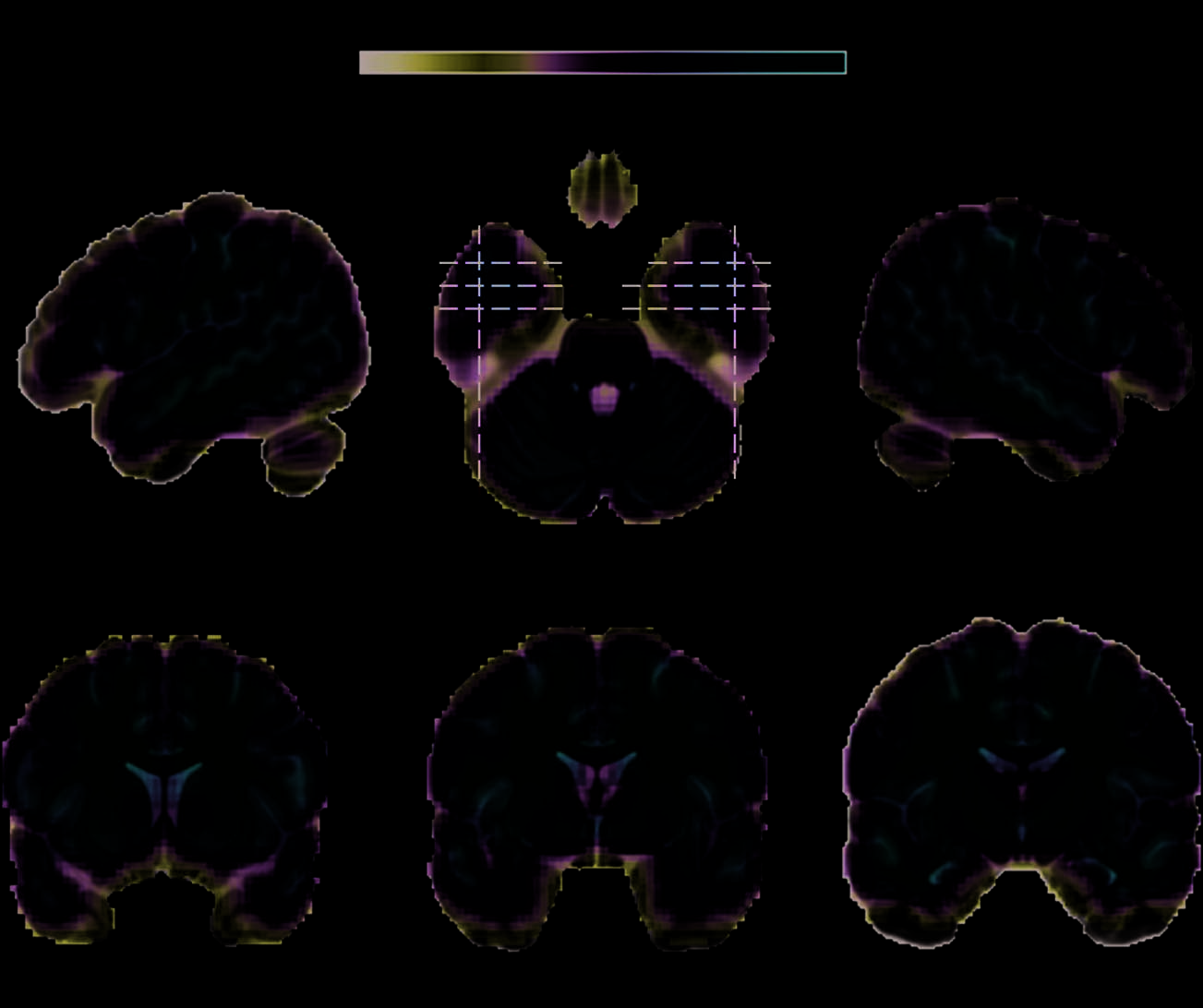
nal in the aMTL region including the PRC (Figur 1).
Subsequent group-level analyses included only voxels with
group mean TSNR > 40.
Group level random effects analyses were run by enter-
ing parameter estimate images from each participantʼs
GLM into one-sample t tests or F contrasts. Results were
thresholded at voxel level p < .01 uncorrected and clus-
ter level p < .05 with FWE correction for multiple com-
parisons. To explore more completely the a priori
predictions that Sharedness would modulate activity in
the fusiform gyri and C×D activity in the PRC, we report
additional results using a lower cluster size threshold.
This is especially critical with respect to the predicted
effects of Correlation × Distinctiveness in the PRC,
since this region is known to show small changes in
BOLD signal (Cohen & Bookheimer, 1994). For this rea-
son, results for Correlation × Distinctiveness are shown
both at the standard threshold noted above and at a
Tyler et al.
1727
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. TSNR around PRC
is sufficient for detection of
BOLD activity. Color bar shows
group mean TSNR, where a
minimum of 40 is needed to
detect BOLD activity. Slice
positions are reported in
MNI coordinates and shown
as dotted lines on the axial
section.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
reduced threshold of cluster-level p < .05 uncorrected,
including only those voxels with low but reliable signal
(i.e., intermediate TSNR between 40 and 100). Given
our a priori hypothesis that Sharedness would modu-
late activity within the fusiform gyrus, we examined the
effect of Sharedness within the fusiform gyrus without
cluster-level thresholding. We defined the extent of the
fusiform ROI using the Harvard-Oxford atlas (fsl.fmrib.ox.
ac.uk/fsl/fslwiki/Atlases). Voxels were included if the
atlas labeled them as “temporal fusiform cortex, posterior
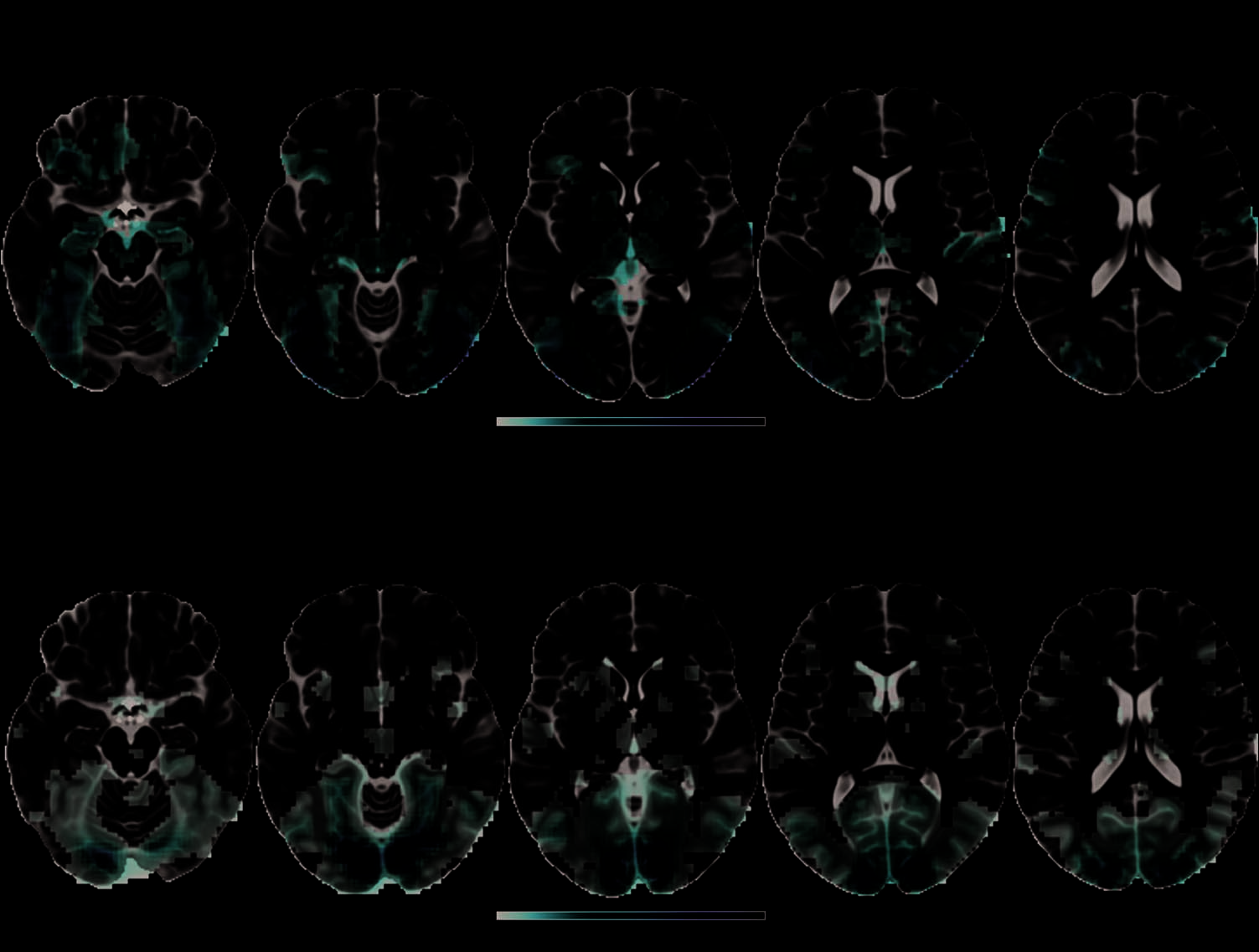
Figure 2. Brain activity
associated with picture
naming. (top) Contrast of
basic level naming versus
scrambled images overlaid
on the ventral cerebral
surface. Activation specific to
naming meaningful objects
(controlling for verbal output)
was found along the anterior
to posterior extent of the
ventral stream. (bottom)
Activity explained by the
visual model, where effects
were focused in the bilateral
occipital poles, with weaker
effects extending to the
posterior parts of the fusiform
and inferior temporal gyri
(see text for details). Color
bars represent voxel t and
F values (degrees of freedom).
1728
Journal of Cognitive Neuroscience
Volume 25, Number 10
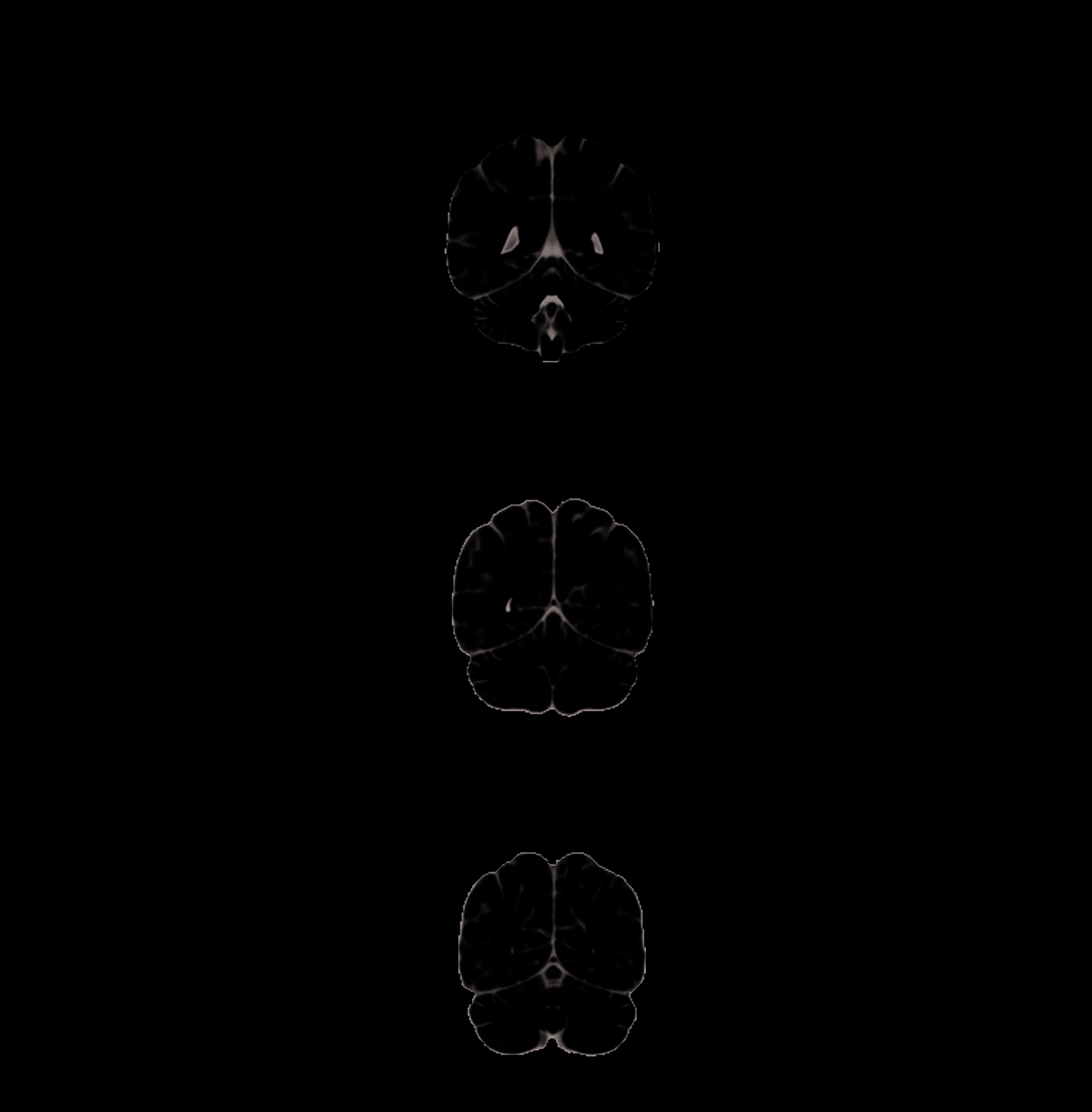
Figure 3. Sharedness of
object features modulates
BOLD activity within the
fusiform gyrus. (top) Objects
with relatively more shared
features were associated with
greater BOLD activity in the
bilateral lateral fusiform gyri,
regions previously associated
with activity for animals
(Martin, 2007; Chao et al.,
1999), consistent with the
greater number of shared
features in animals than tools
(voxel level threshold p < .01,
cluster level threshold p < .05
FWE). (bottom) The a priori
prediction that Sharedness
would differentially modulate
the medial and lateral fusiform
gyri was tested within an
anatomically defined fusiform
ROI without cluster level
thresholding. Objects with
more shared features (orange)
produce activity in lateral
fusiform and those with
fewer shared features (blue;
corresponding to tools)
produce activity in the
bilateral medial fusiform gyri.
Slice positions are given as
MNI coordinates, and color
bars represent voxel t values
(degrees of freedom).
division” or “temporal occipital fusiform cortex” with a
probability of >10%.
In follow-up analyses, we determined whether Shared-
ness generated differential activity in medial and lateral
fusiform gyri and whether this tracked category effects
in the fusiform (Chao et al., 1999). We defined linear
ROIs in each hemisphere as lines of voxels between
medial and lateral points in the fusiform gyri in three
coronal planes at Montreal Neurological Institute
(MNI) y = −48, −57, and −66 mm (Figur 4, center col-
umn). These planes crossed the anterior, Mitte, Und
posterior parts of the category effects reported by Chao
et al. (1999). We avoided examining activity in succes-
sive sagittal planes (d.h., from medial to lateral along
the x axis in MNI space), as this is relatively imprecise
and may dilute effects by including nonresponsive vox-
els. Stattdessen, we defined linear ROIs following the actual
anatomy of the fusiform, which is slightly oblique to the
x axis in MNI space. Parameter estimates were extracted
from successive voxels along the linear ROIs from two
additional GLMs, the first based on explicit categories
(living and nonliving) and second on the Sharedness
Variable. As in the main model described above, modu-
lators were orthogonalized serially, with the eight visual
parametric modulators entered first. The ninth para-
metric modulator was either category membership
(+1 for living, −1 for nonliving) or the Sharedness value
for each concept. In this way, we examined the differ-
ential sensitivity of medial and lateral fusiform gyri to
category level information defined either explicitly or
using feature statistics in two separate but comparable
Modelle.
ERGEBNISSE
Picture Naming Performance
Seventy-three percent of responses were scored as
correct according to the criteria described in the fMRI
Verfahren. A further 8% identified the correct concept,
but using a verbal label that did not correspond to that
in the property norm study (z.B., “loaf ” instead of
“bread”). Of the remaining 19% of responses scored as
incorrect, 4% were no responses, 2% were stutters, Und
13% were the incorrect concept (z.B., “lion” instead of
“tiger”). This accuracy rate is comparable to those
obtained in other studies using large sets of pictures
and similar criteria for coding errors (z.B., Taylor et al.,
2012; Graves, Grabowski, Mehta, & Gordon, 2007; Alario
et al., 2004; Barry, Morrison, & Ellis, 1997; Levelt, Schriefers,
Tyler et al.
1729
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
Ö
9
C
7
N
0
_
0
A
/
_
J
0
Ö
0
C
4
N
1
9
_
A
P
_
D
0
0
B
4
j
1
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Meyer, Pechman, & Vorberg, 1991). The mean (±SD) overt
basic level naming latency over all correct items was
965 ms (±77; siehe Tabelle 1), comparable with previous
fMRI studies (z.B., Graves et al., 2007).
Object-related Activity in the Ventral Stream
To identify the neural regions associated with object pro-
Abschließen, BOLD activity associated with the correctly
named objects was contrasted with the phase-scrambled
Bilder (voxel level p < .001, cluster level p < .05 FWE).
We found activity throughout the occipital lobes and
bilateral ventral streams through to posterior PRC, ex-
tending to right anterior PRC and hippocampus and left
amygdala (Figure 2, left), and also in bilateral ventral pre-
central cortices and left OFC, replicating previous find-
ings (Tyler et al., 2004).
The following analyses focus on the visual and feature
statistics variables (see Task Design and Materials). We
first correlated BOLD responses with visual features, rep-
resented by eight PCA components derived from the
Gabor-filtered images. These eight regressors were
entered into a one-way ANOVA and tested using an
“effects of interest” F contrast. Because this contrast
tests for voxels showing a response to any one of the
eight visual regressors, we report results using a more
conservative voxel level threshold of p < .05 FWE with
a minimal cluster size threshold of 10 voxels, because
correction was applied at the voxel level. Significant
main effects were observed in the bilateral occipital
poles (Figure 2, right; peak voxels in each hemisphere:
MNI 12, −90, −3 mm, F(8, 98) = 88.3, and MNI 15, −90,
0 mm, F(8, 98) = 88.1), similar to previous results (e.g.,
Kwong et al., 1992). Outside the occipital lobe, we found
weaker activation extending anteriorly along the fusi-
form and lingual gyri (peak voxels outside occipital lobe:
MNI 27, −51, −9 mm, F(8, 98) = 23.8 and MNI −24, −48,
−9 mm, F(8, 98) = 21.2) to the posterior end of the
inferior temporal gyrus.
Objects with higher values on the Sharedness variable
(i.e., greater degree of feature sharedness) produced
Figure 4. Activity patterns
within the fusiform gyrus for
the contrast of living versus
nonliving objects closely track
the correlation with Sharedness,
but not the correlation with
C×D. Linear ROIs (center
column; see Imaging Analyses)
traversing the fusiform gyri at
y = −48 mm (A), −57 mm (B),
and −66 mm (C) were used to
extract activity values (t values)
in successive voxels from left
to right. The resulting plots
(left and right columns) confirm
region-specific similarities in
activation between the living
versus nonliving contrast
(black) and the correlation
with Sharedness (solid gray)
but not the correlation with
C×D (dashed gray). Regions
responding preferentially to
living things (relative to
nonliving things) also respond
to concepts with relatively more
shared features, and regions
responding preferentially to
nonliving things (relative to
living things) also respond
to concepts with relatively fewer
shared features. In contrast,
these regions are not
modulated by the requirement
for feature integration (C×D).
The fine and course dashed
horizontal reference lines
indicate t values corresponding
to p < .01 and .001,
respectively.
1730
Journal of Cognitive Neuroscience
Volume 25, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
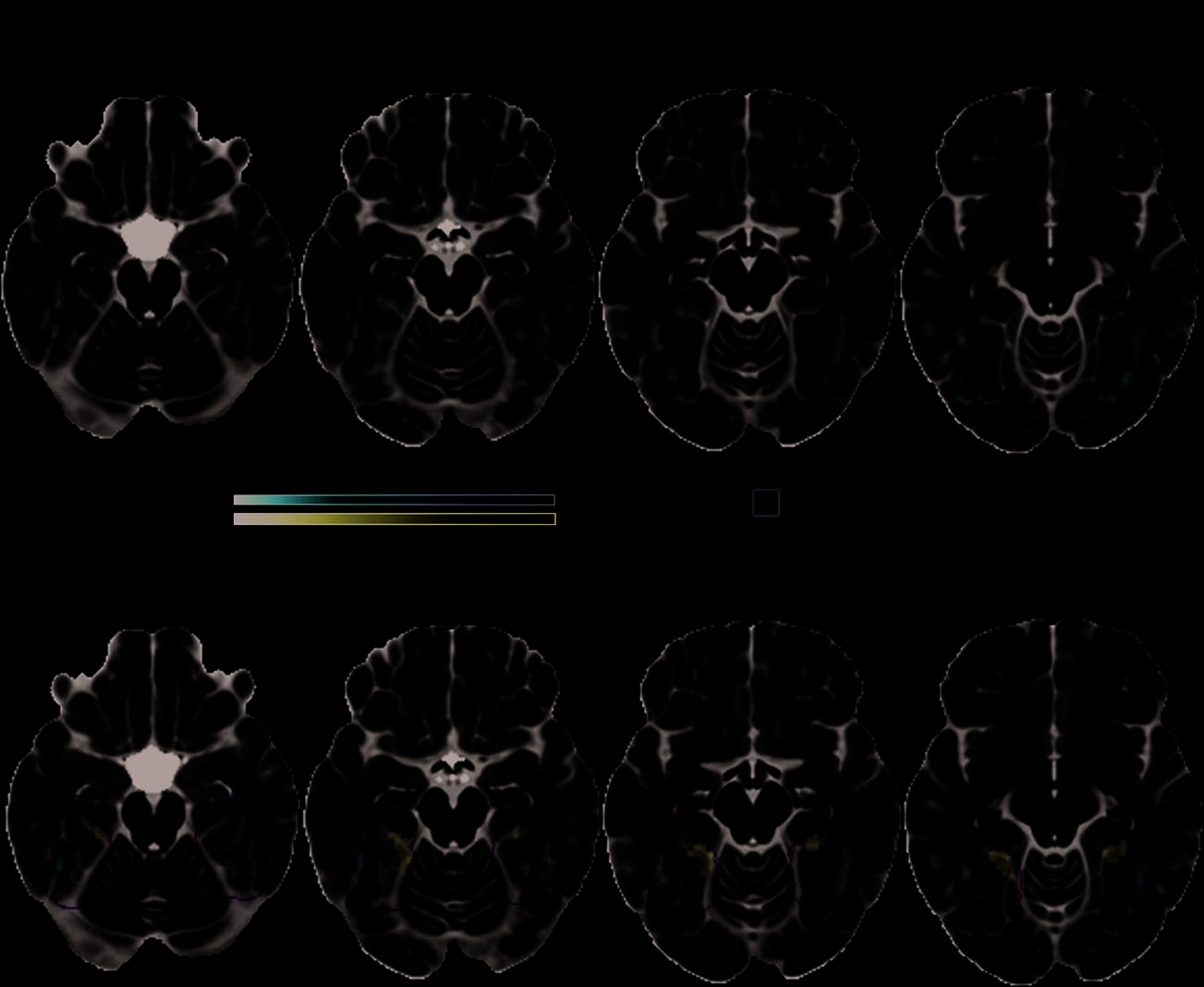
Figure 5. The feature statistic
Correlation × Distinctiveness
modulates BOLD activity in
anteromedial temporal cortex.
(top) Objects with lower
Correlation × Distinctiveness
values—indicating relatively
weakly correlated distinctive
features requiring more
complex feature integration
processes for their unique
identification—were associated
with greater activity in the
anteromedial temporal cortex
including the left PRC at voxel
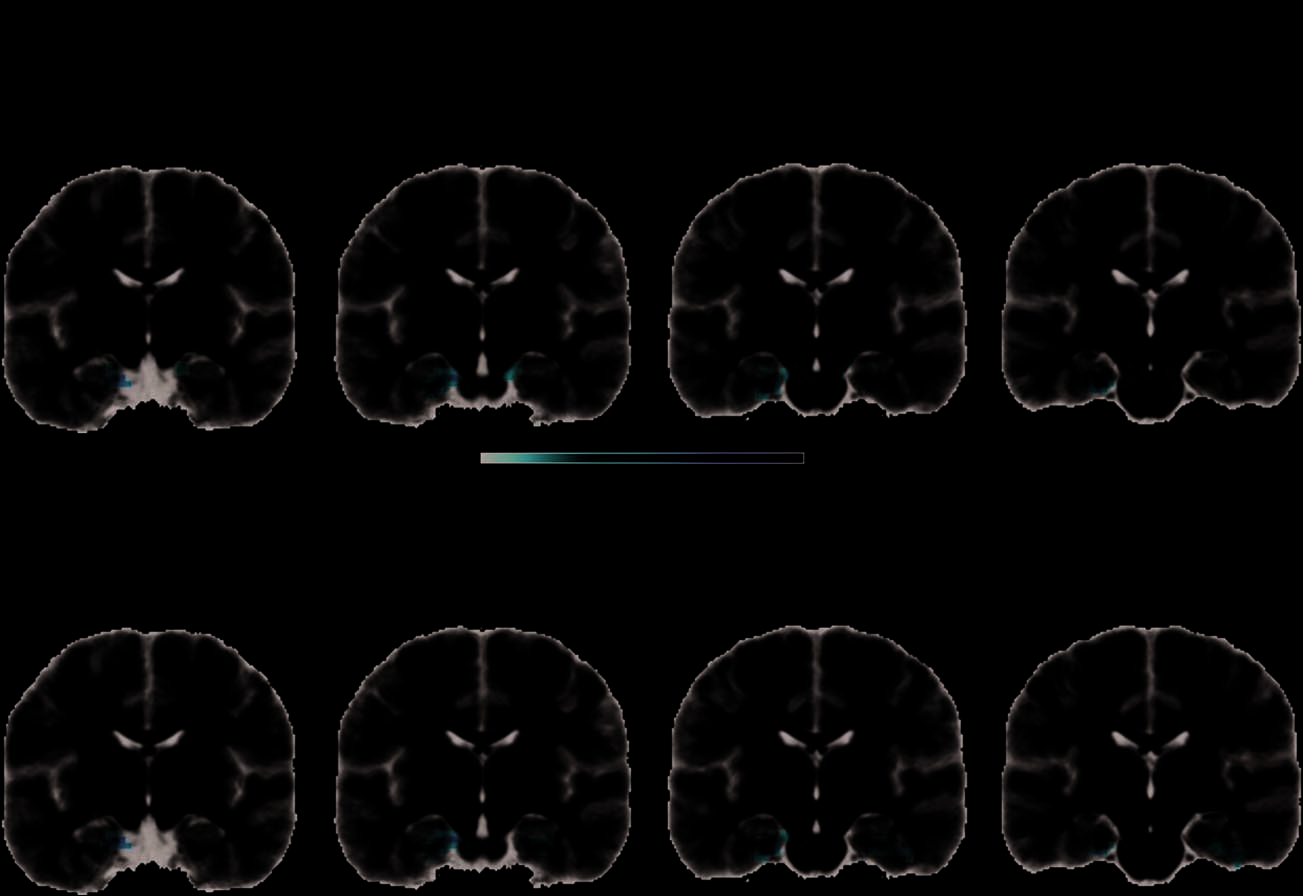
level p < .01, cluster level p <
.05 FWE. (bottom) At voxel level
p < .01, uncorrected cluster
level p < .05 in voxels with
intermediate TSNR of 40–100,
bilateral PRC activation was
seen. To maximize anatomic
localizability of the clusters with
respect to the PRC (Pruessner
et al., 2002), clusters are shown
on the average participant brain.
Slice positions are reported as
MNI coordinates and the
color bar represents voxel
t values (degrees of freedom).
greater activity in bilateral lateral fusiform gyri (Figure 3,
top; peak voxels MNI 42, −39, −27 mm, t(14) = 4.9 and
MNI −39, −48, −18 mm, t(14) = 4.2) and posterior
occipital and ventral occipitotemporal regions (peak
voxel MNI 12, −90, 3 mm, t(14) = 6.5). These regions
corresponded to those previously reported as showing
greater activity to animals than tools (Chao et al., 1999).
To further explore these effects, in particular in medial
fusiform regions, we examined activity within the fusi-
form gyrus without cluster-level FWE correction (Fig-
ure 3, bottom). At this threshold, we saw the same
positive effect of Sharedness in the lateral fusiform,
and in addition a negative effect of Sharedness, cor-
responding to increased activity for objects with lower
values of the Sharedness variable (i.e., fewer shared
features) in bilateral medial fusiform gyri (peak voxels
MNI 30, −45, −9 mm, t(14) = 3.8 and MNI −24, −45,
−15 mm, t(14) = 3.6). This result is consistent with
previous reports of greater activity in medial fusiform
gyri to tools than animals (Chao et al., 1999).
We then examined anatomical variability of activity
within the fusiform gyri along lines of voxels traversing
medial to lateral sites (see Imaging Analyses; Figure 4,
center column). Medial and lateral fusiform regions
showed opposite effects, with lateral regions more active
for items with more shared features and medial regions
more active for objects with less shared features (Figure 4,
gray lines on plots). Lateral voxels at MNI X = ±36–39 mm
typically showed the highest positive correlation with
Sharedness, whereas medial voxels at MNI X = ±24–
27 mm typically showed the lowest negative correlation.
Moreover, the medial to lateral pattern of activity for the
Sharedness variable tracked closely with the pattern pro-
duced by contrasting the explicit categories “living” and
“nonliving” (Figure 4, black lines on plots): The lateral
fusiform gyri responded more to living things (which
typically have more shared features), whereas the medial
fusiform gyri showed the reverse pattern, responding
more to nonliving things (which have fewer shared and
more distinctive features). To confirm that the living
and nonliving objects used in this study differed in their
degree of sharedness, we compared the Sharedness
values for the living and nonliving objects. Consistent with
previous studies, living things had significantly greater
feature sharedness (mean Sharedness = .53, SD = .12)
than nonliving things (mean Sharedness = .40, SD = .14;
difference between mean living and nonliving; t(386) =
9.27, p < .0001).
The effects of the Correlation × Distinctiveness vari-
able showed that objects whose distinctive features were
relatively more weakly correlated than their shared fea-
tures (typical of living things) elicited stronger activity
in the anteromedial temporal cortex, primarily left peri-
rhinal and entorhinal cortices (Figure 5, left; peak voxel:
MNI −24, −21, −27 mm, t(14) = −6.9). There were no
significant activations associated with relatively more
strongly correlated distinctive compared with shared fea-
tures. We further explored these effects at a reduced
threshold (see Imaging Analyses). Whereas the positive
contrast remained nonsignificant, the negative contrast
Tyler et al.
1731
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
now revealed bilateral PRC activation associated with
naming concepts with relatively more weakly correlated
distinctive than shared features (see Figure 5, right; peak
voxels: −24, −21, −27, t(14) = −6.9 and 39, −24, −24,
t(14) = −6.0). Two small clusters were also found in the
left medial superior and orbital frontal lobe (peak voxels:
MNI −6, 63, 37 mm, t(14) = −5.1 and MNI −3, 57,
36 mm, t(14) = −4.29). Because distinctive features must
be integrated with a conceptʼs shared features for basic
level identification, confusable concepts with relatively
weakly correlated distinctive features require more com-
plex feature integration processes supported by the bilat-
eral PRC for their unique identification (Taylor, Devereux,
& Tyler, 2011; Tyler & Moss, 2001).
DISCUSSION
In this study, we used a feature-based model of semantics
to determine how two key aspects of object semantics—
object-specific and category information—are neurally
represented and processed. This feature-based approach
to object representations in the brain has previously been
validated in cognitive studies, which show that feature
statistics affect conceptual processing (Taylor et al.,
2012; Randall et al., 2004; Pexman et al., 2003; Cree,
McRae, & McNorgan, 1999; McRae et al., 1997). The sta-
tistics used in these experiments are based on features
obtained from large-scale property norming studies
(e.g., McRae et al., 2005). Participants in property norm
studies are biased to report salient, verbalizable, and dis-
tinguishing features (McRae et al., 1997, 2005; Tyler et al.,
2000). Although it is assumed that these biases do not
interact with concept type or category, this assumption
requires experimental validation. Thus, although feature
statistics derived from feature norm studies are generally
regarded as the “gold standard” for characterizing the se-
mantics of concepts, surpassing semantic feature data ob-
tained using automatic extraction algorithms on large-scale
corpora (Devereux, Pilkington, Poibeau, & Korhonen,
2010), the inherent biases in feature norming data repre-
sent a potential limitation in any study using these data.
Behavioral studies have shown that a variety of feature
statistics—including those used here—affect behavioral
responses (Taylor et al., 2012; Randall et al., 2004; McRae
et al., 1997). For example, Taylor et al. (2012) showed
that shared features facilitated category decisions on ob-
jects whereas the ease with which distinctive features
could be integrated into a concept (as measured by the
Correlation × Distinctiveness variable) facilitated object-
specific identification, reflecting the functional relevance
of different types of information carried by the relationship
between different features within a concept.
Consistent with the behavioral studies, in the present
experiment, we found that feature statistics, reflecting dif-
ferent aspects of an objectʼs meaning, activated different
neural regions. Sharedness—which captures the degree
to which a conceptʼs features are shared with other con-
cepts and thus forms the basis of category organization—
modulated activity within the fusiform gyri. Moreover,
we found greater activation in lateral fusiform gyri for
objects with higher Sharedness and greater activation in
medial fusiform gyri for objects with lower Sharedness.
This profile of lateral to medial activity within each hemi-
sphere closely tracked activity for living and nonliving
things, with greater Sharedness showing similar effects as
living things and less Sharedness showing similar effects
as nonliving things. This correspondence between the
effects of Sharedness and living things is consistent with
the claim that living things have higher proportions of
shared properties than nonliving objects (Randall et al.,
2004; Cree & McRae, 2003; Rosch et al., 1976), a pattern
replicated in the present set of objects where living things
had more shared properties than nonliving things. Feature-
based models of semantics claim that category structure
(e.g., living, nonliving things) is an emergent property of
feature statistics ( Vigliocco et al., 2004; Cree & McRae,
2003; Garrard, Lambon Ralph, Hodges, & Patterson, 2001;
Tyler & Moss, 2001; Durrant-Peatfield, Tyler, Moss, & Levy,
1997), raising the possibility that features may provide
an organizing principle for category structure in the
brain. However, this is a difficult hypothesis to test empiri-
cally because of the inherent interdependence between
Sharedness and category.
In contrast to the effects in the fusiform, feature statis-
tics that differentiate between similar objects and enable
object-specific representations were associated with
aMTL activity, including in the bilateral PRCs. These results
further support the view that the meaning of concrete
objects is neurally coded in terms of feature-based repre-
sentations. Taken together, the present findings suggest a
hierarchy of semantic processing in the ventral stream
with similar computational properties as has been pro-
posed for the hierarchical model of perceptual object pro-
cessing developed in nonhuman primates (Taylor et al.,
2011; Taylor, Moss, & Tyler, 2007; Tyler et al., 2004). This
perceptual model claims that simple visual features are
coded in posterior ventral occipital sites, with increas-
ingly more complex feature combinations computed from
posterior to anterior regions in ventral temporal cortex
(Tanaka, 1996; Ungerleider & Mishkin, 1982). Nonhuman
primate IT neurons code for moderately complex fea-
tures (Tanaka, 1996), whereas PRC, at the endpoint of this
hierarchical system, generates feedback signals to bind
the relevant information in IT cortex together (pair-coding
properties; Higuchi & Miyashita, 1996), thereby coding
for the most complex feature combinations necessary to
disambiguate highly confusable objects (Murray et al.,
2007; Bussey et al., 2002). Critically, nonhuman primate
research also demonstrates that the ventral stream codes
not only perceptual object properties, but also the mean-
ing of these properties (Hoffman & Logothetis, 2009;
Sigala & Logothetis, 2002). Such findings demonstrate
that, in contrast to the less flexible, retinotopic coding
at the posterior end of the ventral stream, information
1732
Journal of Cognitive Neuroscience
Volume 25, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
represented in the anterior region becomes tuned to
meaningful, task-relevant features with experience. More-
over, just as the visual hierarchy is achieved by recurrent
activity between posterior and anterior sites within the
ventral stream (Hegdé, 2008), so communication between
the fusiform and anterior temporal lobe during object
processing is underpinned by recurrent activity between
these two regions (Clarke, Taylor, & Tyler, 2011).
The finding that the two anatomically distinct regions
associated with category selectivity in previous studies—
aMTL and fusiform—are sensitive to different aspects of
an objectʼs semantic features supports the suggestion
that posterior and anterior sites differ in their ability to
integrate less or more complex semantic feature conjunc-
tions, respectively. In this study, we found that feature
statistics that capture the similarity between objects
(shared features) were associated with activity in fusiform
regions previously linked to category-specific responses
(Martin, 2007). Category responses (e.g., viewing, match-
ing) require simpler feature conjunctions than those
needed for fine-grained differentiation between similar
objects within a category; these latter involve the most
complex feature conjunctions (Moss et al., 2005; Tyler
et al., 2004) that are reflected in our second measure
of Correlation × Distinctiveness. This measure captures
the relative strength of the correlation for distinctive
and shared features within an object, and thus the ease
with which features can be integrated, thus enabling
similar objects to be successfully differentiated from
each other. According to property norm data, these
are typically living things, especially animals (Taylor
et al., 2008, 2012; see also Table 1). The present re-
sults show that objects with this statistical profile en-
gage the aMTL, especially the PRC, more than objects
whose distinctive features are strongly correlated with its
other features and are thus easier to differentiate one
from another.
These results are consistent with neuropsychological
studies demonstrating that patients with aMTL damage
that includes the PRC have particular problems differ-
entiating between highly similar objects, especially ani-
mals (Tyler & Moss, 2001). These patients exhibit a very
specific kind of category-specific deficit; they have con-
siderable difficulties with the distinctive properties of
objects and thus are unable to differentiate similar objects
from each other. Thus, they have little difficulty in iden-
tifying an objectʼs category but cannot differentiate be-
tween members within the same category. Moreover, this
problem is much more pronounced for living than non-
living things (Moss, Tyler, & Devlin, 2002; Tyler & Moss,
2001; Moss, Tyler, Durrant-Peatfield, & Bunn, 1998). The
feature-based model described here can account for this
behavioral effect in patients and its association with a
specific neural region.
Supporting evidence for a posterior to anterior shift in
neural activity in terms of the types of integration compu-
tations required comes from studies that manipulate the
subjectʼs task and consequently the kind of conceptual
representation required (Clarke et al., 2011; Tyler et al.,
2004). These task manipulations require participants to
process an object at different levels of specificity—as a
member of a category or as a specific object (animal,
camel ). Naming an object as a member of a category
requires simpler feature conjunctions and activity is
confined to posterior sites. Making an object-specific re-
sponse to the same object, which requires the computa-
tion of more complex feature conjunctions, also engages
aMTL (Barense, Henson, Lee, & Graham, 2010; Moss
et al., 2005; Tyler et al., 2004). This is consistent with the
notion that both perceptual and conceptual object pro-
cessing progresses from a coarse- to fine-grained analysis
along the ventral stream (Clarke et al., 2013; Taylor et al.,
2012; Tyler et al., 2004). The use of a basic level naming
task in this study ensured functional activity along the
entire ventral stream (Tyler et al., 2004) as well as the
integration of distinctive with shared object features re-
quired for unique object identification, thus enabling the
measurement of the effects of both the relative proportion
of shared features and the relative correlational strength
of shared and distinctive features.
In conclusion, this study suggests that a conceptual
hierarchy, analogous to the perceptual hierarchy, and
based on semantic feature statistics which capture statis-
tical regularities of concepts experienced in the world,
underpins the recognition of meaningful objects in the
ventral temporal cortex. By combining a cognitive model
of semantic representations with a neurobiological model
of hierarchical processing in the ventral stream, it accounts
for variation in neural activity as a function of the semantic
structure of individual objects and the relationship be-
tween objects, and provides a unifying framework for
heretofore unconnected findings of category responses
in the fusiform (e.g., Chao et al., 1999) and category effects
in anteromedial temporal cortex (Moss et al., 2005).
Acknowledgments
This work was supported by a grant from the Medical Research
Council (G0500842) to L. K. T., a British Academy (grant LRG-
45583) grant to L. K. T. and K. I. T., a Newton Trust grant to
L. K. T., the European Research Council (ERC) under the
European Communityʼs Seventh Framework Programme (FP7/
2007–2013)/ERC (grant 249640) to L. K. T., and a Swiss National
Science Foundation Ambizione Fellowship (grant PZ00P1_
126493) to K. I. T. We are very grateful for the training data
for FIRST, used to create the Harvard-Oxford atlas, particularly
to David Kennedy at the CMA, and also to Christian Haselgrove,
Centre for Morphometric Analysis, Harvard; Bruce Fischl,
Martinos Center for Biomedical Imaging, MGH; Janis Breeze
and Jean Frazier, Child and Adolescent Neuropsychiatric Research
Program, Cambridge Health Alliance; Larry Seidman and Jill
Goldstein, Department of Psychiatry of Harvard Medical School;
and Barry Kosofsky, Weill Cornell Medical Center.
Reprint requests should be sent to Professor Lorraine K. Tyler,
Centre for Speech, Language and the Brain, Department of
Psychology, University of Cambridge, Downing Street, Cambridge
CB2 3EB, UK, or via e-mail: lktyler@csl.psychol.cam.ac.uk.
Tyler et al.
1733
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
REFERENCES
Alario, F. X., Ferrand, L., Laganaro, M., New, B., Frauenfelder,
U. H., & Segui, J. (2004). Predictors of picture naming speed.
Behavior Research Methods, Instruments, & Computers: A
Journal of the Psychonomic Society, Inc., 36, 140–155.
Barense, M. D., Bussey, T. J., Lee, A. C. H., Rogers, T. T., Davies,
R. R., Saksida, L. M., et al. (2005). Functional specialization
in the human medial temporal lobe. The Journal of
Neuroscience, 25, 10239–10246.
Barense, M. D., Gaffan, D., & Graham, K. (2007). The human
medial temporal lobe processes online representations of
complex objects. Neuropsychologia, 45, 2963–2974.
Barense, M. D., Henson, R. N. A., & Graham, K. S. (2011).
Perception and conception: Temporal lobe activity
during complex discriminations of familiar and novel
faces and objects. Journal of Cognitive Neuroscience, 23,
3052–3067.
Barense, M. D., Henson, R. N. A., Lee, A. C. H., & Graham, K. S.
(2010). Medial temporal lobe activity during complex
discrimination of faces, objects, and scenes: Effects of
viewpoint. Hippocampus, 20, 389–401.
Barry, C., Morrison, C., & Ellis, A. (1997). Naming the Snodgrass
and Vanderwart pictures: Effects of age of acquisition,
frequency, and name agreement. The Quarterly Journal of
Experimental Psychology, 50A, 560–585.
Bussey, T. J., Saksida, L. M., & Murray, E. A. (2002). Perirhinal
cortex resolves feature ambiguity in complex visual
discriminations. European Journal of Neuroscience, 15,
365–374.
Chao, L. L., Haxby, J. V., & Martin, A. (1999). Attribute-based
neural substrates in temporal cortex for perceiving and
knowing about objects. Nature Neuroscience, 2, 913–919.
Clarke, A., Taylor, K. I., Devereux, B., Randall, B., & Tyler, L. K.
(2013). From perception to conception: How meaningful
objects are processed over time. Cerebral Cortex, 23,
187–197.
Clarke, A., Taylor, K. I., & Tyler, L. K. (2011). The evolution of
meaning: Spatiotemporal dynamics of visual object recognition.
Journal of Cognitive Neuroscience, 23, 1887–1899.
Cohen, M. S., & Bookheimer, S. Y. (1994). Localization of brain
function using magnetic resonance imaging. Trends in
Neurosciences, 17, 268–277.
Cree, G. S., McNorgan, C., & McRae, K. (2006). Distinctive
features hold a privileged status in the computation of word
meaning: Implications for theories of semantic memory.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 32, 643–658.
Cree, G. S., & McRae, K. (2003). Analyzing the factors
underlying the structure and computation of the meaning
of chipmunk, cherry, chisel, cheese, and cello (and many
other such concrete nouns). Journal of Experimental
Psychology: General, 132, 163–201.
Cree, G. S., McRae, K., & McNorgan, C. (1999). An attractor
model of lexical conceptual processing: Simulating semantic
priming. Cognitive Science, 23, 371–414.
Devereux, B. J., Pilkington, N., Poibeau, T., & Korhonen, A.
(2010). Towards unrestricted, large-scale acquisition of
feature-based conceptual representations from corpus data.
Research on Language and Computation, 7, 137–170.
Durrant-Peatfield, M. R., Tyler, L. K., Moss, H. E., & Levy, J. P.
(1997). The distinctiveness of form and function in category
structure: A connectionist model. In M. G. Shafto & P. Langley
(Eds.), Proceedings of the 19th Annual Conference of the
Cognitive Science Society (pp. 193–198). Mahwah, NJ:
Erlbaum.
Garrard, P., Lambon Ralph, M. A., Hodges, J. R., & Patterson, K.
(2001). Prototypicality, distinctiveness, and intercorrelation:
Analyses of the semantic attributes of living and nonliving
concepts. Cognitive Neuropsychology, 18, 125–174.
Gotts, S. J., & Plaut, D. C. (2004). Connectionist approaches to
understanding aphasic perseveration. Seminars in Speech
and Language, 25, 323–334.
Graves, W., Grabowski, T., Mehta, S., & Gordon, J. (2007). A
neural signature of phonological access: Distinguishing the
effects of word frequency from familiarity and length in overt
picture naming. Journal of Cognitive Neuroscience, 19,
617–631.
Grondin, R., Lupker, S. J., & McRae, K. (2009). Shared features
dominate semantic richness effects for concrete concepts.
Journal of Memory and Language, 60, 1–19.
Hegdé, J. (2008). Time course of visual perception: Coarse-to-
fine processing and beyond. Progress in Neurobiology, 84,
405–439.
Higuchi, S., & Miyashita, Y. (1996). Formation of mnemonic
neuronal responses to visual paired associates in
inferotemporal cortex is impaired by perirhinal and
entorhinal lesions. Proceedings of the National Academy
of Sciences, U.S.A., 93, 739–743.
Hoffman, K. L., & Logothetis, N. K. (2009). Cortical mechanisms
of sensory learning and object recognition. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 364, 321–329.
Humphreys, G. W., & Forde, E. M. E. (2001). Hierarchies,
similarity, and interactivity in object recognition: Category-
specific neuropsychological deficits. Behavioral and Brain
Sciences, 24, 453–509.
Kay, K., Naselaris, T., Prenger, R., & Gallant, J. (2008).
Identifying natural images from human brain activity. Nature,
452, 352–356.
Keil, F. C. (1986). The acquisition of natural kind and artifact
terms. In W. Domopoulous & A. Marras (Eds.), Language
learning and concept acquisition (pp. 133–153). Norwood,
NJ: Ablex.
Kivisaari, S. L., Tyler, L. K., Monsch, A. U., & Taylor, K. I. (2012).
Medial perirhinal cortex disambiguates confusable objects.
Brain, 135, 3757–3769.
Kwong, K. K., Belliveau, J. W., Chesler, D. A., Goldberg, I. E.,
Weisskoff, R. M., Poncelet, B. P., et al. (1992). Dynamic
magnetic resonance imaging of human brain activity during
primary sensory stimulation. Proceedings of the National
Academy of Sciences, U.S.A., 89, 5675–5679.
Levelt, W. J. M., Schriefers, H., Meyer, A. S., Pechman, T., &
Vorberg, D. (1991). The time course of lexical access in
speech production: A study of picture naming. Psychological
Review, 98, 122–142.
Mahon, B. Z., & Caramazza, A. (2009). Concepts and categories:
A cognitive neuropsychological perspective. Annual Review
of Psychology, 60, 27–51.
Malt, B. C., & Smith, E. (1984). Correlated properties in natural
categories. Journal of Verbal Learning and Verbal Behavior,
23, 250–269.
Martin, A. (2007). The representation of object concepts in the
brain. Annual Review of Psychology, 58, 25–45.
McRae, K., & Cree, G. S. (2002). Factors underlying category-
specific semantic deficits. In E. M. E. Forde & G. W.
Humphreys (Eds.), Category-specificity in brain and mind
(pp. 211–249). Hove, UK: Psychology Press.
McRae, K., Cree, G. S., Seidenberg, M. S., & McNorgan, C.
(2005). Semantic feature production norms for a large set of
living and nonliving things. Behavior Research Methods, 37,
547–559.
McRae, K., de Sa, V. R., & Seidenberg, M. S. (1997). On the
nature and scope of featural representations of word
meaning. Journal of Experimental Psychology: General, 126,
99–130.
1734
Journal of Cognitive Neuroscience
Volume 25, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Mirman, D., & Magnuson, J. S. (2008). Attractor dynamics and
semantic neighborhood density: Processing is slowed by near
neighbors and speeded by distant neighbors. Journal of
Experimental Psychology: Learning, Memory, and
Cognition, 34, 65–79.
Moss, H. E., Rodd, J. M., Stamatakis, E. A., Bright, P., & Tyler, L. K.
(2005). Anteromedial temporal cortex supports fine-grained
differentiation among objects. Cerebral Cortex, 15, 616–627.
Moss, H. E., Tyler, L. K., & Devlin, J. T. (2002). The emergence
of category-specific deficits in a distributed semantic system.
In E. M. E. Forde & G. W. Humphreys (Eds.), Category
specificity in brain and mind (pp. 115–145). Hove, UK:
Psychology Press.
Moss, H. E., Tyler, L. K., Durrant-Peatfield, M., & Bunn, E. M.
(1998). “Two eyes of a see-through”: Impaired and intact
semantic knowledge in a case of selective deficit for living
things. Neurocase, 4, 291–310.
Moss, H. E., Tyler, L. K., & Jennings, F. (1997). When leopards
lose their spots: Knowledge of visual properties in category-
specific deficits for living things. Cognitive Neuropsychology,
14, 901–950.
Murphy, K., Bodurka, J., & Bandettini, P. A. (2007). How long to
scan? The relationship between fMRI temporal signal to noise
and necessary scan duration. Neuroimage, 34, 565–574.
Murray, E. A., & Bussey, T. J. (1999). Perceptual-mnemonic
functions of the perirhinal cortex. Trends in Cognitive
Sciences, 3, 142–151.
Murray, E. A., Bussey, T. J., & Saksida, L. M. (2007). Visual
perception and memory: A new view of medial temporal
lobe function in primates and rodents. Annual Review of
Neuroscience, 30, 99–122.
Naselaris, T., Prenger, R., Kay, K., Oliver, M., & Gallant, J. (2009).
Bayesian reconstruction of natural images from human brain
activity. Neuron, 63, 902–915.
Nishimoto, S., Vu, A. T., Naselaris, T., Benjamini, Y., Yu, B., &
Gallant, J. (2011). Reconstructing visual experiences from
brain activity evoked by natural movies. Current Biology, 21,
1641–1646.
Noppeney, U., Patterson, K., Tyler, L. K., Moss, H., Stamatakis,
E. A., Bright, P., et al. (2007). Temporal lobe lesions and
semantic impairment: A comparison of herpes simplex virus
encephalitis and semantic dementia. Brain, 130, 1138–1147.
Op de Beeck, H. P., Torfs, K., & Wagemans, J. (2008). Perceived
shape similarity among unfamiliar objects and the organization
of the human object vision pathway. The Journal of
Neuroscience, 28, 10111–10123.
Pexman, P. M., Holyk, G. G., & Monfils, M. H. (2003). Number-
of-features effects and semantic processing. Memory &
Cognition, 31, 842–855.
Pruessner, J. C., Kohler, S., Crane, J., Pruessner, M., Lord, C.,
Byrne, A., et al. (2002). Volumetry of temporopolar, perirhinal,
entorhinal and parahippocampal cortex from high-resolution
MR images: Considering the variability of the collateral sulcus.
Cerebral Cortex, 12, 1342–1353.
Randall, B., Moss, H. E., Rodd, J. M., Greer, M., & Tyler, L. K.
(2004). Distinctiveness and correlation in conceptual structure:
Behavioral and computational studies. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 30, 393–406.
Ratcliff, R. (1993). Methods for dealing with reaction time
outliers. Psychological Bulletin, 114, 510–532.
cortex and semantic memory: Reconciling findings from
neuropsychology and functional imaging. Cognitive,
Affective, and Behavioral Neuroscience, 6, 201–213.
Rosch, E., Mervis, C. B., Gray, W. D., Johnson, D. M., &
Boyes-Braem, P. (1976). Basic objects in natural categories.
Cognitive Psychology, 8, 382–439.
Saksida, L. M., Bussey, T. J., Buckmaster, C. A., & Murray, E. A.
(2007). Impairment and facilitation of transverse patterning
after lesions of the perirhinal cortex and hippocampus,
respectively. Cerebral Cortex, 17, 108–115.
Sigala, N., & Logothetis, N. K. (2002). Visual categorization
shapes feature selectivity in the primate temporal cortex.
Nature, 415, 318–320.
Smith, E. E., & Medin, D. L. (1981). Categories and concepts.
Cambridge, MA: Harvard University Press.
Tanaka, K. (1996). Inferotemporal cortex and object vision.
Annual Review of Neuroscience, 19, 109–140.
Taylor, K. I., Devereux, B. J., Acres, K., Randall, B., & Tyler, L. K.
(2012). Contrasting effects of feature-based statistics on
the categorisation and identification of visual objects.
Cognition, 122, 363–374.
Taylor, K. I., Devereux, B. J., & Tyler, L. K. (2011). Conceptual
structure: Towards an integrated neurocognitive account.
Language and Cognitive Processes (Cognitive Neuroscience
of Language), 26, 1368–1401.
Taylor, K. I., Moss, H. E., Stamatakis, E. A., & Tyler, L. K. (2006).
Binding crossmodal object features in perirhinal cortex.
Proceedings of the National Academy of Sciences, U.S.A.,
103, 8239–8244.
Taylor, K. I., Moss, H. E., & Tyler, L. K. (2007). The conceptual
structure account: A cognitive model of semantic memory
and its neural instantiation. In J. Hart & M. Kraut (Eds.),
The neural basis of semantic memory (pp. 265–301).
Cambridge: Cambridge University Press.
Taylor, K. I., Salamoura, A., Randall, B., Moss, H., & Tyler, L. K.
(2008). Clarifying the nature of the distinctiveness by domain
interaction in conceptual structure: Comment on Cree,
McNorgan, and McRae (2006). Journal of Experimental
Psychology: Learning, Memory, and Cognition, 34, 719–725.
Tyler, L. K., & Moss, H. E. (2001). Towards a distributed account
of conceptual knowledge. Trends in Cognitive Sciences, 5,
244–252.
Tyler, L. K., Moss, H. E., Durrant-Peatfield, M. R., & Levy, J.
(2000). Conceptual structure and the structure of concepts: A
distributed account of category-specific deficits. Brain and
Language, 75, 195–231.
Tyler, L. K., Stamatakis, E. A., Bright, P., Acres, K., Abdallah, S.,
Rodd, J. M., et al. (2004). Processing objects at different levels
of specificity. Journal of Cognitive Neuroscience, 16,
351–362.
Ungerleider, L., & Mishkin, M. (1982). Two cortical visual
systems. In D. J. Ingle, M. A. Goodale, & R. J. W. Mansfiled
(Eds.), Analysis of visual behavior (pp. 549–586).
Cambridge, MA: MIT Press.
Vigliocco, G., Vinson, D. P., Lewis, W., & Garrett, M. F. (2004).
Representing the meanings of object and action words: The
featural and unitary semantic space hypothesis. Cognitive
Psychology, 48, 422–488.
Warrington, E. K., & McCarthy, R. (1983). Category-specific
access dysphasia. Brain, 106, 859–878.
Rogers, T. T., Hocking, J., Noppeney, U., Mechelli, A., Gorno-
Tempini, M., Patterson, K., et al. (2006). Anterior temporal
Warrington, E. K., & Shallice, T. (1984). Category specific
semantic impairments. Brain, 107, 829–854.
Tyler et al.
1735
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
7
0
2
/
3
1
1
7
9
2
4
3
5
/
9
1
2
1
7
7
o
9
c
7
n
0
_
0
a
/
_
j
0
o
0
c
4
n
1
9
_
a
p
_
d
0
0
b
4
y
1
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1