Neural Responses to Ambiguity Involve Domain-general
and Domain-specific Emotion Processing Systems
Maital Neta1, William M. Kelley2, and Paul J. Whalen2
Abstrakt
■ Extant research has examined the process of decision making
under uncertainty, specifically in situations of ambiguity. Wie-
immer, much of this work has been conducted in the context of
semantic and low-level visual processing. An open question is
whether ambiguity in social signals (z.B., emotional facial expres-
sionen) is processed similarly or whether a unique set of processors
come on-line to resolve ambiguity in a social context. Our work
has examined ambiguity using surprised facial expressions, als
they have predicted both positive and negative outcomes in the
Vergangenheit. Speziell, whereas some people tended to interpret sur-
prise as negatively valenced, others tended toward a more posi-
tive interpretation. Hier, we examined neural responses to social
ambiguity using faces (surprise) and nonface emotional scenes
(International Affective Picture System). Darüber hinaus, we examined
whether these effects are specific to ambiguity resolution (d.h.,
judgments about the ambiguity) or whether similar effects would
be demonstrated for incidental judgments (z.B., nonvalence judg-
ments about ambiguously valenced stimuli). We found that a dis-
tinct task control (d.h., cingulo-opercular) network was more
active when resolving ambiguity. We also found that activity in
the ventral amygdala was greater to faces and scenes that were
rated explicitly along the dimension of valence, consistent with
findings that the ventral amygdala tracks valence. Taken together,
there is a complex neural architecture that supports decision
making in the presence of ambiguity: (A) a core set of cortical
structures engaged for explicit ambiguity processing across stim-
ulus boundaries and (B) other dedicated circuits for biologically
relevant learning situations involving faces. ■
EINFÜHRUNG
Uncertainty is a prominent feature of our decision pro-
Prozess (Kahneman, Slovic, & Tversky, 1982), as it is ubiqui-
tous in our realistic settings, and poses a major obstacle
to effective decision making (Brunsson, 1985; Corbin,
1980). Although such decision-making processes have
been studied for several decades (Tversky & Kahneman,
1974), we have only recently begun to understand the
neural architecture that allows for performing such a
complex task. A clear understanding of these cognitive
functions requires, Dann, also a clear definition of uncer-
tainty. Whereas some describe this concept as a psycholog-
ical state in which an individual lacks sufficient knowledge
about the outcome(S) of a given choice (see Platt &
Huettel, 2008, für eine Rezension), a more specific kind of uncer-
tainty is derived from ambiguity. In large part, research on
ambiguity has been studied in the realm of linguistic prop-
erties, as ambiguity resolution is a central problem in lan-
guage comprehension (Rodd, Davis, & Johnsrude, 2005;
see MacDonald, Pearlmutter, & Seidenberg, 1994, for a
Rezension). More recently, work in neuroeconomics has com-
pared the effects of risk (levels of probability) and ambigu-
ität (uncertain probabilities) in the context of choice (Erheben,
Snell, Nelson, Rustichini, & Glimcher, 2010; Hsu, Bhatt,
Adolphs, Tranel, & Camerer, 2005).
1Washington University School of Medicine, 2Dartmouth College
Although this work has examined decision-making pro-
cesses in the presence of ambiguity, it remains unclear how
this may differ from the process of resolving ambiguity (d.h.,
making a discrete decision about an ambiguous stimulus,
specifically along the dimension of the ambiguity). In other
Wörter, an incidental judgment in the presence of ambigu-
ity might require participants to judge the gender of faces
that have been morphed along the dimension of race,
whereas an explicit resolution judgment would require par-
ticipants to judge the races of those same faces. Some stud-
ies have revealed that a set of cortical regions collectively
referred to as the cingulo-opercular network (dorsal ACC
[dACC]/medial superior frontal cortex and the bilateral
frontal operculum [FO]/anterior insula [AI]; see Dosenbach
et al., 2006) has been linked to decision making in the pres-
ence of semantic ambiguity (d.h., the selection between
competing alternatives; Thompson-Schill, DʼEsposito,
Aguirre, & Farah, 1997) and when detecting ambiguity in
visual motion (Sterzer, Russ, Preibisch, & Kleinschmidt,
2002). With respect to face processing, one study found
that these regions were more active only when participants
were asked to resolve the ambiguity (d.h., when making
race judgments about faces that are racially ambiguous
[morphed Asian and white faces] and gender judgments
about faces that are gender-ambiguous [morphed male
and female faces; Demos, Wig, Moran, & Kelley, 2004]).
One goal of the current work was to determine whether
certain aspects of the task demands specifically recruit this
© 2013 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 25:4, S. 547–557
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
Ö
9
C
0
N
7
_
9
A
/
_
J
0
Ö
0
C
3
N
6
3
_
A
P
_
D
0
0
B
3
j
6
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
F
.
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Netzwerk, das ist, when it is recruited by the mere presence
of ambiguity or when specifically making decisions that re-
solve the ambiguity.
To date, much of the work examining ambiguity has
been conducted in the context of semantic and low-level
visual processing. An open question is whether ambiguity
in social signals (z.B., emotional facial expressions) Sind
processed in a similar manner or whether a unique set
of processors come on-line to resolve ambiguity in an
emotional context. Zum Beispiel, surprised facial expres-
sions are ambiguous in that they have predicted both
positive and negative outcomes in the past. Tatsächlich, Das
ambiguity can be exploited to demonstrate that people
show individual differences in their propensity to inter-
pret surprised faces as either positive or negative. Im
absence of contextual information that can be used to
disambiguate the valence of this expression, some people
interpret surprised faces negatively, whereas others inter-
pret them positively (Neta & Whalen, 2010; Neta, Norris, &
Whalen, 2009; Kim, Somerville, Johnstone, Alexander, &
Whalen, 2003). The only known reports of neural re-
sponses to ambiguity of surprised faces has shown that
the amygdala is modulated by ambiguity but only in the
absence of a task that requires ambiguity resolution per
se (Kim et al., 2003, 2004). Taken together, the main goal
of this study was to examine ambiguity as it relates to social
Signale, both in face and other nonface emotional stimuli
(d.h., International Affective Picture System [IAPS] scenes)
and to determine how task demands might modulate
responses to ambiguity (explicit valence ratings of ambig-
uously valenced stimuli, as compared with nonvalence
evaluations).
Zu diesem Zweck, we examined behavioral and neural re-
sponses to ambiguous stimuli across two stimulus and
task categories. Speziell, we presented face stimuli
(surprised, angry, and happy expressions) sowie
scenes (ambiguous, clearly negative, and clearly positive)
and asked participants to make either a valence or non-
valence judgment about each one. In both stimulus cate-
gories, the clearly valenced stimuli were included purely
to serve as anchors for behavioral valence ratings of the
ambiguous stimuli. We predicted that there might be a
set of cortical regions that come on-line when effortful
processing of ambiguity is required (d.h., explicit valence
judgments of ambiguously valenced stimuli), whereas the
amygdala may play a dual role, in accordance with ventral
and dorsal subregions tracking valence and arousal, Re-
spectively, that may be critically dependent on the task
demands.
METHODEN
Teilnehmer
Thirty-five healthy participants (right-handed, without
neurological disease, and with normal/corrected vision;
21 Frauen) volunteered. None were aware of the purpose
of the experiment, and they were all compensated for their
participation through monetary payment or course credit.
Written informed consent was obtained from each partici-
pant before the session, and all procedures were approved
by the Dartmouth College Committee for the Protection of
Human Subjects. Three participants were removed because
of technical complications with the stimulus-presenting
computer, and an additional participant was removed be-
cause of nonnormative ratings (z.B., happy expressions were
rated as negative on greater than 40% of trials). Infolge,
the final sample contained 31 Teilnehmer (19 Frauen). Alle
included participants tested within normal limits for de-
pression (Beck Depression Inventory [BDI]; Beck, Station,
& Mendelson, 1961; M = 3.74, SE = .66) and anxiety
(Spielberger, Gorsuch, & Lushene, 1988; State-Trait
Anxiety Inventory: STAIs = 31.06 ± 1.21, STAIt = 34.32 ±
1.43). One additional participant was removed from the
fMRI data analysis because of scanner-related artifact, Also
the final sample for those analyses contained 30 Teilnehmer
(18 Frauen).
Stimuli
For the face task, we used 24 face identities (12 men) Pos-
ing angry, happy, and surprised expressions. We selected
images of 14 Identitäten (7 Frauen, 7 men) from the NimStim
standardized facial expression stimulus set (Tottenham
et al., 2009) Und 20 Identitäten (10 Frauen, 10 men) aus
the averaged Karolinska Directed Emotional Faces data-
base (Lundqvist, Flykt, & Öhman, 1998). Of the 34 indi-
viduals whose images were included in the experiment,
some posed all three expressions, and some posed only
one or two of the expressions, providing us with 48 dis-
crete stimuli. The facial expressions in this stimulus set
have been validated by a separate set of participants who
labeled each expression; only faces correctly labeled more
als 60% of the time were included.
Wichtig, not all of the identities are represented
in each expression condition. Jedoch, the focus of this
study was to assess the ambiguous conditions specifically
(faces and scenes), and the clearly valenced conditions
(z.B., happy and angry expressions) were included only
to serve as anchors that would ground participantsʼ re-
sponses to the ambiguous (z.B., surprised) Reize. Als
solch, we used twice as many face identities in the surprise
condition compared with the angry and happy face con-
ditions, and so, the conditions were never intended to be
matched for identity. Eher, we selected identities within
each expression condition that were labeled the most
accurately in terms of the normative data provided with
the stimulus set, so to ensure that our clearly valenced
faces were the ideal anchors (d.h., the most clearly valenced
stimuli we could present), and the same goal was applied
for the surprised faces.
For the scenes task, we used images from IAPS (Lang,
Bradley, & Cuthbert, 2008) that were previously rated as
either positive, negative, or ambiguous in valence. Ambiguity
548
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 4
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
Ö
9
C
0
N
7
_
9
A
/
_
J
0
Ö
0
C
3
N
6
3
_
A
P
_
D
0
0
B
3
j
6
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
T
.
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
was based on discordant valence ratings across participants
(some rated images as positive and others as negative; d.h.,
high standard deviation in valence ratings; Tisch 1). Der
mean (SEM) valence ratings on the normative 9-point scale
(1 = very unpleasant, 9 = very pleasant) war 2.39 (0.08)
for negative pictures, 7.93 (0.07) for positive pictures, Und
5.19 (0.26) for ambiguous pictures (F(2, 10) = 1469.44,
P < .001), such that positive pictures were rated as more
positive than ambiguous and negative pictures, and negative
pictures were rated as more negative than ambiguous pic-
tures (all ps < .001). Importantly, arousal was equated across
these three conditions: the mean (SEM) arousal ratings on
the normative 9-point scale (1 = low arousal, 9 = high
arousal) were 5.36 (0.20) for negative pictures, 4.96 (0.18)
for positive pictures, and 4.99 (0.18) for ambiguous pictures
(F(2, 10) = 1.56, p = .26; Table 1).
In a behavioral pilot, we recruited 20 new participants
(11 women). None were aware of the purpose of the ex-
periment, and they were all compensated for their par-
ticipation through monetary payment or course credit.
Written informed consent was obtained from each par-
ticipant before the session, and all procedures were
approved by Dartmouth College Committee for the Pro-
tection of Human Subjects. Each participant viewed 114
scenes and rated each image as positive or negative
(consistent with the methods in Experiment 1). Of the
114 scenes, 24 were defined as clearly negative, 24 as
clearly positive, and 66 as ambiguous, based on the valence
and arousal ratings provided with the stimuli (Table 1).
Each image was presented for 1500 msec on a black back-
ground, with an intertrial interval of 500 msec, during
which a white fixation cross appeared on the screen. As
with the face stimuli, 24 images were used, and each par-
ticipant viewed 50% ambiguous images and 50% clear
valence images (25% clear positive and 25% clear nega-
tive). Thus, from the data collected in the pilot, we se-
lected 12 clearly negative and 12 clearly positive images
and 24 with an ambiguous valence (i.e., chosen because
they had the highest standard deviation in valence ratings
across participants). Indeed, the standard deviation in rat-
ings of ambiguous scenes was significantly greater than
the standard deviation in ratings of clearly valenced scenes
(t(23) = 19.2, p < .001).
Finally, to directly compare the conditions across stimu-
lus sets, we ran a Stimulus (IAPS, faces) × Valence (ambig-
uous, negative, positive) repeated-measures ANOVA on
the normative ratings associated with each stimulus set.
For valence ratings, we found a significant main effect of
Valence (as expected) but no main effect of Stimulus
( p > .2). For arousal ratings, Jedoch, there was a main
effect of Stimulus (F(1, 11) = 5.66, p = .04), where arousal
ratings were significantly higher for IAPS than faces, welche
is somewhat expected given the nature of the stimuli.
Experiment Design and Parameters
The fMRI paradigm consisted of eight functional runs of
four experimental blocks each: two blocks each of facial
expressions and IAPS stimuli interleaved. Each block began
with a brief (2 Sek) task instruction statement: “valence”
or “gender” (nonvalence) for the face blocks and “valence”
or “social/nonsocial” (d.h., “Does the picture contain a
person?”; nonvalence) for the IAPS blocks. Each block con-
sisted of 24 Bilder, 12 from the ambiguous conditions and
six from each clear valence condition (positive, negative).
All images were presented sequentially for 500 ms, mit
an ISI of 1500 ms, in a randomized fashion for all condi-
tionen. An additional 24 trials were presented randomly
within each block to provide jitter, during which a white
fixation cross appeared on the screen, Und 24 fixation trials
were presented between each block (Figur 1). The order
of the blocks was counterbalanced across participants.
During imaging, participants responded by pressing one
of two buttons with their dominant hand. Darüber hinaus, Die
ratings of surprised faces on the valence task allowed for
the determination of positivity–negativity bias and RT.
Following each scanning session, participants also com-
pleted the following behavioral scales: BDI (Beck et al.,
1961) and the STAIs/STAIt (Spielberger et al., 1988).
Imaging Parameters
Images were acquired on a Philips Achieva 3.0-T scanner
(Philips Medical Systems, Bothell, WA), equipped with a
SENSE birdcage head coil. Anatomical T1-weighted images
were collected using a high-resolution 3-D magnetization-
prepared rapid gradient-echo sequence, mit 160 contig-
uous 1-mm thick sagittal slices (Echozeit = 4.6 ms,
repetition time = 9.8 ms, Sichtfeld = 240 mm, umdrehen
Winkel = 8°, Voxelgröße = 1 × 0.94 × 0.94 mm). Funktional
Tisch 1. Results of Stimulus-matching Procedure for IAPS Scenes
Negative Pictures
Ambiguous Pictures
Positive Pictures
Mean (SEM ) valence
Mean (SEM ) arousal
2.39 (0.08)
5.36 (0.20)
5.19 (0.26)
4.99 (0.18)
7.93 (0.07)
4.96 (0.18)
Valence scores are based on the normative 9-point scale (1 = very unpleasant, 9 = very pleasant). There was a significant difference between
each condition of pictures (F(2, 10) = 1469.44, P < .001, η2 = .95), such that positive pictures were rated as more positive than ambiguous and
negative pictures, and negative pictures were rated as more negative than ambiguous pictures (all ps < .001).
Arousal was also based on the normative 9-point scale (1 = low arousal, 9 = high arousal ). Arousal was equated across these three conditions
(F(2, 10) = 1.56, p = .26, η2 = .10).
Neta, Kelley, and Whalen
549
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
o
9
c
0
n
7
_
9
a
/
_
j
0
o
0
c
3
n
6
3
_
a
p
_
d
0
0
b
3
y
6
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
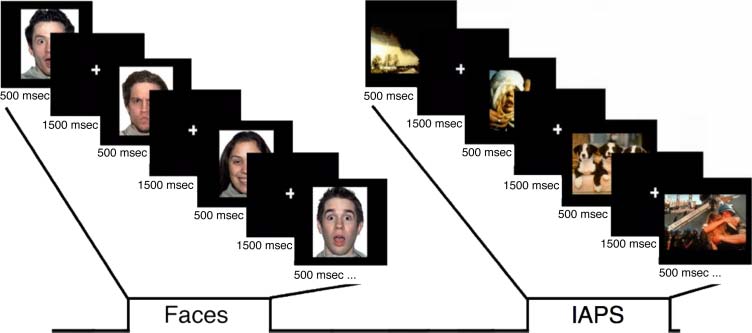
Figure 1. A depiction of the
experimental design. The
paradigm consisted of eight
runs of four experimental
blocks each: two blocks each
of facial expressions and IAPS
stimuli interleaved. Each block
began with a brief (2000 msec)
instruction statement: “valence”
or “gender” (nonvalence) for
the face blocks and “valence” or
“social/nonsocial” (nonvalence;
i.e., “Does the picture contain
a person?”) for the IAPS
blocks. Each block consisted
of 24 images, 12 from the
ambiguous conditions (e.g.,
surprised faces) and six from
each clear valence condition
(positive, negative; e.g., happy and angry faces, respectively). All images were presented sequentially for 500 msec, with an ISI of 1500 msec, in a
randomized fashion for all conditions. An additional 24 trials were presented randomly within each block to provide jitter, during which a white fixation
cross appeared on the screen, and 24 fixation trials were presented between each block. The order of the blocks was counterbalanced across participants.
images were acquired using an echo-planar T2*-weighted
imaging sequence. Each volume consisted of 36 interleaved
3-mm thick slices, AC–PC aligned, with 0.5-mm interslice
gap (echo time = 35 msec, repetition time = 2000 msec,
field of view = 240 mm, flip angle = 90°, voxel size = 3 ×
3 × 3.5 mm). Despite our focus on the amygdala, we used
slices aligned with the AC–PC plane so that we could effec-
tively interrogate whole-brain effects of processing am-
biguity across stimulus and task categories.
fMRI Analysis
Processing of fMRI data took place in SPM2 ( Wellcome
Department of Cognitive Neurology, London, United
Kingdom). First, several preprocessing steps were em-
ployed to increase the signal-to-noise ratio before for-
mation of statistical images. Slice time correction was
employed to correct for acquisition delays within func-
tional volumes. Next, realignment corrected for participant
head motion within and across runs using rigid body trans-
formation. Functional data then underwent unwarping,
which corrects for image distortions caused by move-
ment by susceptibility interactions (Andersson, Hutton,
Ashburner, Turner, & Friston, 2001). Functional data were
normalized to the EPI.mnc template, which warped data
into Montreal Neurological Institute space. Normalized
functional data were then spatially smoothed (6-mm
FWHM) using a Gaussian kernel.
Next, the general linear model was performed to examine
separate task contributions to the fMRI signal. Surprised,
angry, and happy expressions as well as ambiguous, neg-
ative, and positive IAPS were modeled as separate task
regressors, which were convolved with a canonical hemo-
dynamic response function. These task regressors were
included in a general linear model along with covariates
of noninterest (session mean, run regressor, linear trend,
and six movement parameters derived from realignment
corrections) to compute parameter estimates (images con-
taining weighted parameter estimates) for each comparison
at each voxel and for each participant.
Contrast maps were then entered into a random effects
model, which accounts for intersubject variability and
allows population-based inferences to be drawn. Param-
eter estimates for the ambiguous stimuli (i.e., surprised
faces and ambiguously valenced IAPS) were then sub-
mitted to a 2 × 2 ANOVA examining the effects of material
type (faces vs. IAPS) and task instruction (explicit valence
judgments vs. incidental non judgments). Brain regions
showing significant main effects in these analyses were ex-
tracted as ROIs (threshold: p < .001, as determined by
Monte Carlo simulations implemented in AlphaSim within
AFNI software [Cox, 1996] correcting for the whole brain).
Given the current studyʼs focus on the amygdala, we im-
posed a significance threshold of p < .05, corrected for mul-
tiple comparisons over the amygdala volume (∼4500 mm3,
defined using the Automated Anatomical Labeling atlas;
Maldjian, Laurienti, Kraft, & Burdette, 2003) and also deter-
mined by Monte Carlo simulations, a strategy we have im-
plemented in previous studies (Davis, Johnstone, Mazzulla,
Oler, & Whalen, 2010; Kim et al., 2003, 2010; Johnstone,
Somerville, Alexander, et al., 2005).
ROI
ROI analyses were conducted using the MarsBaR tool within
SPM2 ( Wellcome Department of Imaging Neuroscience,
London, United Kingdom). Spherical regions (6-mm ra-
dius) were defined around each of these peak activations,
and all significant voxels ( p < .001) were included. Signal
intensities for each ROI were then calculated separately for
each task comparison (i.e., ambiguously valenced faces and
IAPS, for the valence and nonvalence tasks separately) and
examined statistically using repeated-measures ANOVA.
Then, the BOLD signal (beta weights) from the significantly
550
Journal of Cognitive Neuroscience
Volume 25, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
o
9
c
0
n
7
_
9
a
/
_
j
0
o
0
c
3
n
6
3
_
a
p
_
d
0
0
b
3
y
6
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
activated voxels was extracted from each ROI for each
participant and submitted for off-line testing.
RESULTS
Behavioral Results
Valence Ratings
Angry and happy expressions were rated as consistently
negative (95.2% of trials) and positive (93.1% of trials), re-
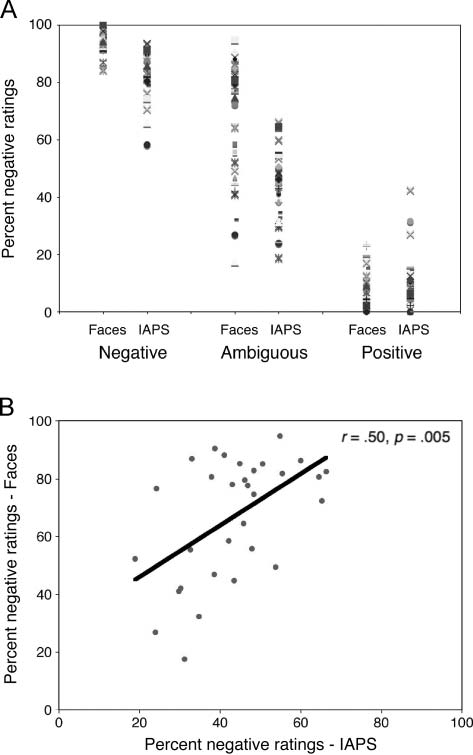
spectively. Figure 2A shows that these same participants
differed in their tendency to interpret surprised faces
negatively versus positively. As a manipulation check of
the valence conditions (ambiguously, as compared with
clearly valenced stimuli), a Stimulus (IAPS, faces) × Valence
(ambiguous, negative, positive) repeated-measures ANOVA
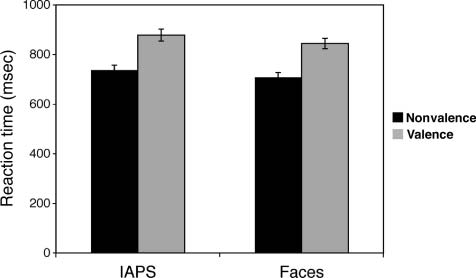
Figure 3. RTs (mean ± standard error) are greater when rating
the valence of ambiguous stimuli, as compared with an incidental
nonvalence rating, and they were also greater for the ambiguous
scenes than faces.
revealed a significant main effect of Valence (F(2, 29) =
1576.1, p < .001), and pairwise comparisons (LSD cor-
rected) revealed that negative stimuli were rated as more
negative than ambiguous stimuli, which were rated as
more negative than positive stimuli ( ps < .001). Impor-
tantly, there was a significant positive correlation in ratings
of ambiguous IAPS and faces (r = .50, p = .005; Figure 2B),
such that participants who tended to interpret surprised
faces as positive also tended to interpret ambiguous IAPS
as positive.
RT
A Stimulus (IAPS, faces) × Task (valence, nonvalence)
repeated-measures ANOVA revealed a significant main
effect of Stimulus (F(1, 30) = 7.3, p = .011), and pairwise
comparisons (LSD corrected) revealed that participants
took longer to rate the IAPS, as compared with the face
stimuli (Figure 3). A significant main effect of Task (F(1,
30) = 295.3, p < .001) revealed that RTs were longer dur-
ing the valence as compared with the nonvalence task.
There was no significant Stimulus × Task interaction
( p > .7).
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
Ö
9
C
0
N
7
_
9
A
/
_
J
0
Ö
0
C
3
N
6
3
_
A
P
_
D
0
0
B
3
j
6
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
/
T
.
F
Questionnaire Data
As noted in the Methods section, we administered per-
sonality and trait measure scales to document that these
participants fell within a healthy psychiatric range on cer-
tain measures (d.h., depression, anxiety) across all partici-
Hose. Results indicated that all scores for depression
(BDI) and anxiety (STAI) were within normal limits.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Figur 2. Individual differences in rating valence of surprised expressions
and ambiguous scenes. (A) Consistent with previous work, angry faces
are rated consistently negative, happy faces are consistently positive,
and surprised ratings vary from negative to positive, revealing individual
differences in how people interpret the valence of these expressions.
(B) There was a significant positive correlation between ratings for
ambiguous faces and scenes. Individuals who tended to rate surprised
faces as positive also tended to rate the ambiguous IAPS scenes as positive.
fMRI Results
A voxelwise whole-brain 2 × 2 repeated-measures ANOVA
on the ambiguous trials with the factors of Stimulus (IAPS,
faces) and Task (valence, nonvalence) revealed brain re-
gions showing a main effect of Stimulus, a main effect of
Neta, Kelley, and Whalen
551
38.92
Left medial superior frontal gyrus
Brain Regions Preferentially Sensitive to Task
Task, and an interaction between Stimulus and Task in re-
sponse to ambiguity (Tisch 2). To explore the directionality
of each effect, regions identified in the statistical F maps
were examined further using ROI analyses. It should be
noted that when RTs were regressed out, none of the re-
ported effects changed.
Tisch 2. Brain Regions Identified as Responding to Ambiguous
Stimuli
X
j
z
F
Region
Main Effect of Stimulus (Faces vs. Scenes)
−33 −39 −21
364.66
Left fusiform gyrus
33 −36 −21
201.71 Right fusiform gyrus
37.0
Right ACC
23.78
Left ACC
32.61
Left dACC
28.14 Right inferior frontal gyrus
3
−6
36
45
−3 −42
12
39
18
57
−6
−36
−51
6
15
39
24
54
57
27.70
Left superior frontal gyrus
18 −30
26.70
Left superior temporal gyrus
−57 −3 −21
34.26
Left middle temporal gyrus
54 −6 −27
38.48 Right inferior temporal gyrus
−27 −90
−39 −84
36 −84
33
97.35
Left superior occipital gyrus
9
170.36
Left middle occipital gyrus
18
124.58 Right middle occipital gyrus
Main Effect of Task ( Valence vs. Nonvalence)
0
6
42
−30
45
−54
−30
30
18
30
48
27
24 −9
21 −3
69.82 Medial superior frontal cortex
35.09 Right dACC
47.56 Right FO
46.84
Left insula
6
18
51
48
36
34.42 Right inferior frontal gyrus
6
9
43.33
Left inferior frontal gyrus
24.26
Left middle frontal gyrus
18
15.07 Right superior frontal gyrus
48 −42
−12 −78
12 −75
0
6
9
18.51 Right middle temporal gyrus
18.42
Left cuneus
18.76 Right cuneus
Stimulus × Task Interaction
−30 −63
−39 −54
−6
51
33
48
21
21.78
Left middle occipital gyrus
17.61
Left inferior parietal lobule
16.89
Left medial frontal gyrus
Brain Regions Preferentially Sensitive to Stimulus
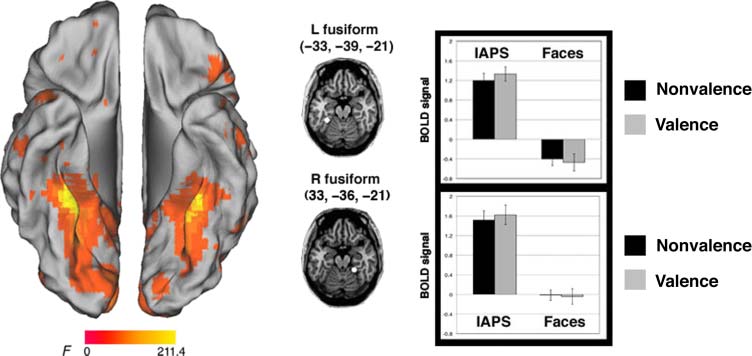
The main effect of Stimulus revealed activations in inferior
temporal cortex, with a peak on the bilateral medial fusiform
gyrus (X, j, z: left = −33, −39, −21; right = 33, −36, −21;
main effects of Stimulus, links: F(1, 29) = 191.0, P < .001;
right: F(1, 29) = 126.3, p < .001). ROI analyses revealed that
activity here was greater for the IAPS stimuli than the face
stimuli (Figure 4). Many of the other regions that showed
a main effect of Stimulus (Table 2) revealed a similar pattern
(i.e., greater activity for IAPS than faces), with the exception
of the right ACC (x, y, z = 3, 36, 6; main effects of stimulus:
F(1, 29) = 45.8, p < .001) and the right inferior fron-
tal gyrus (x, y, z = 57, 12, 24; main effects of Stimulus:
F(1, 29) = 14.7, p = .001), which showed greater activity
for the faces than the IAPS stimuli. Of these regions, only
the latter one also showed a main effect of Task (F(1, 29) =
11.6, p = .002), with a preferential response to the valence,
as compared with the nonvalence task. None of these
regions showed a significant interaction ( p > .1).
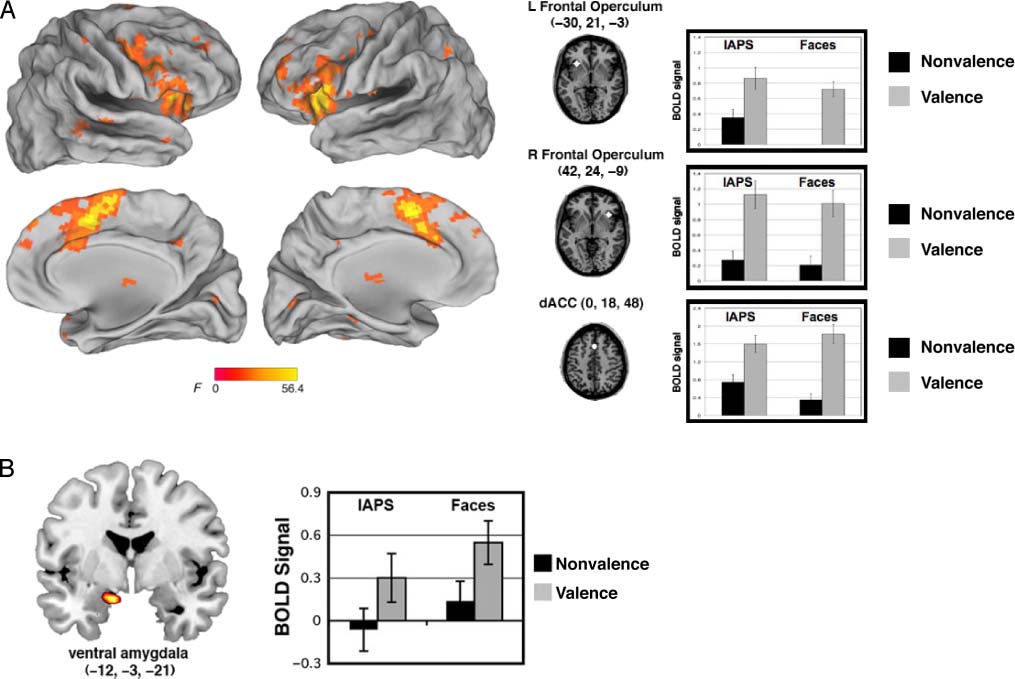
The main effect of Task revealed activity in the bilateral AI/FO
(left = −31, 21, −3; right = 36, 21, −3; main effects of task,
links: F(1, 29) = 38.9, P < .001; right: F(1, 29) = 49.1, p <
.001) as well as in the dACC (x, y, z = 0, 18, 48; main effect
of Task: F(1, 29) = 49.8, p < .001; Figure 5A). ROI analyses
in all of the regions that showed a main effect of Task (Table 2)
revealed that activity here was greater for the explicit valence
judgment task than the nonvalence (gender and social) task
for both categories (faces and scenes, respectively). More-
over, the left AI/FO and the left inferior frontal gyrus also
showed a main effect of Stimulus (F(1, 29) = 12.5, p =
.001; F(1, 29) = 15.6, p < .001; respectively), with a prefer-
ential response to the IAPS than face stimuli. Several of these
regions, including the dACC, showed a significant inter-
action of Stimulus and Task (F(1, 29) = 5.4, p = .03), where
there was a preferential response to the IAPS, but only
during the nonvalence task.
Finally, with a lower significance threshold (see fMRI
Analysis), we found a region of ventral amygdala (x, y,
z = −12, −3, −21) also showed a main effect of Task.
An ROI analysis was used to examine Stimulus (IAPS,
faces) × Task (valence, nonvalence) effects. We confirmed
this main effect of Task (F(1, 29) = 10.6, p = .003), but no
other main effects or interactions were significant ( ps >
.2). Like the other regions showing a main effect of Task,
there was greater activity here for the valence task as com-
pared with the nonvalence task (Figure 5B).1
Brain Regions Exhibiting an Interaction of
Stimulus and Task
Three distributed cortical regions demonstrated a cross-
over interaction, including the left middle occipital gyrus
552
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 4
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
Ö
9
C
0
N
7
_
9
A
/
_
J
0
Ö
0
C
3
N
6
3
_
A
P
_
D
0
0
B
3
j
6
G
3
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
.
T
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Figur 4. Brain regions active
for the main effect of stimulus.
Activity in the bilateral ventral
temporal cortex was greater for
the IAPS, as compared with the
face stimuli.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
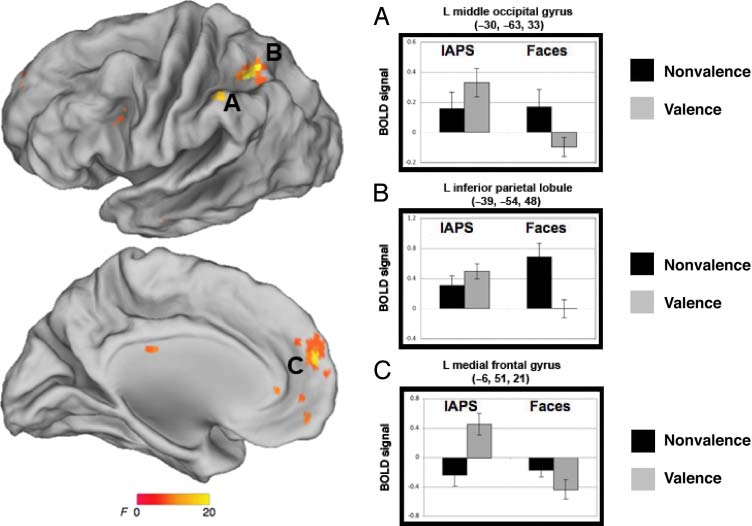
(X, j, z = −30, −63, 33; interaction: F(1, 29) = 16.0, p =
.001), the left inferior parietal lobule (X, j, z = −39, −54,
48; interaction: F(1, 29) = 17.8, P < .001), and the left
medial frontal gyrus (x, y, z = −6, 51, 21; F(1, 29) = 11.3,
p = .002; Figure 6).
DISCUSSION
The neural architecture for responding to ambiguity relies,
in part, on cognitive control processing that is recruited
when participants make a judgment that requires the res-
olution of ambiguity. We found that activity in regions
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
o
9
c
0
n
7
_
9
a
/
_
j
0
o
0
c
3
n
6
3
_
a
p
_
d
0
0
b
3
y
6
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Brain regions active for the main effect of task. (A) Three task control regions, comprising the cingulo-opercular network, showed
greater activity during the valence task, as compared with the nonvalence task (gender judgment for face stimuli, social judgment for IAPS scenes).
(B) A region of ventral amygdala showed greater activity to ambiguous stimuli during the valence task.
Neta, Kelley, and Whalen
553
Figure 6. Brain regions
exhibiting an interaction of
stimulus and task include (A)
left medial frontal gyrus, (B) left
inferior parietal lobule, and (C)
left middle occipital gyrus.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
o
9
c
0
n
7
_
9
a
/
_
j
0
o
0
c
3
n
6
3
_
a
p
_
d
0
0
b
3
y
6
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
comprising a task control network (i.e., cingulo-opercular
network) provides a central part of this architecture, as it
also has been shown to play a crucial role in processing
ambiguity in other domains (i.e., linguistic, low-level per-
ception). This domain-general processing is consistent
with our behavioral results that demonstrated that individ-
ual differences in the bias to resolve ambiguity positively or
negatively were correlated across the stimulus boundaries
(i.e., the same participants who showed a positive bias
when resolving ambiguity of facial expressions showed a
similar positive bias when resolving ambiguity in scenes).
Finally, we found that activity in the ventral amygdala was
greater to faces and scenes that were rated explicitly along
the dimension of valence, consistent with findings that the
ventral amygdala tracks valence (Whalen et al., 2009; Kim
et al., 2003). Here, we discuss the implications of these
findings, specifically in the context of task control and
specific emotion processing systems.
The Function of the Cingulo-opercular Network
A meta-analysis of 10 different tasks showed that three
cortical regions, comprising the cingulo-opercular net-
work (dACC and the bilateral AI/FO), showed three dis-
tinct types of task-control signals: (1) signals tied to the
start of a task, which are likely related to the instantia-
tion of task parameters; (2) activity sustained at a con-
stant level across the task period, which likely reflects
maintenance of those parameters; and (3) error-related
activity, which is likely related to performance feed-
back (Dosenbach et al., 2006). Thus, this network has
been referred to as a “core” task set system (Dosenbach
et al., 2006, 2007), which is thought to be involved in
controlling task performance through the stable main-
tenance of task parameters, as well as making and moni-
toring choices in accordance with those parameters (see
Dosenbach, Fair, Cohen, Schlaggar, & Petersen, 2008, for
a review).
Aside from these higher-order cognitive functions,
there are widely replicated findings implicating the dACC
in conflict monitoring (Botvinick, Braver, Barch, Carter, &
Cohen, 2001; Miller & Cohen, 2001). Moreover, the AI
shows transient responses related to electrodermal arousal
(Critchley, Elliott, Mathias, & Dolan, 2000), empathy
(Singer et al., 2004), the generation of subjective feelings
and integration of sensory inputs (see Medford & Critchley,
2010, for a review), and other socioemotional processing
(Chang, Smith, Dufwenberg, & Sanfey, 2011; Sanfey, Rilling,
Aronson, Nystrom, & Cohen, 2003; Lane, Fink, Chua, &
Dolan, 1997). This region is also shown, via resting state
functional connectivity (see Dosenbach et al., 2007; Fox,
Corbetta, Snyder, Vincent, & Raichle, 2006; Fox et al.,
2005) to coactivate with the dACC as part of a network
that is nonspecifically involved in general goal-directed
cognition (Chang, Yarkoni, Khaw, & Sanfey, 2012; Yarkoni,
Poldrack, Nichols, Van Essen, & Wager, 2011; Dosenbach
et al., 2006). Thus, task-related manipulations (e.g., con-
flict monitoring) that drive activity in one of these regions
(dACC) is likely to also affect activation in the others
(bilateral AI), providing further evidence that these regions
comprise a network or system that is implicated in many
instances requiring attentional resources or task control
(see also Nee, Wager, & Jonides, 2007; Duncan & Owen,
2000, for meta-analyses of cognitive control paradigms).
554
Journal of Cognitive Neuroscience
Volume 25, Number 4
As such, although the dACC and AI are independently
implicated in specific processes (e.g., conflict monitoring,
generating subjective feelings), the specific role of these
regions likely lies in the domain of general task control. In
other words, many different task demands are likely
to require an ability to sustain attention, monitor task
parameters, and modulate arousal (Nelson et al., 2010;
Dosenbach et al., 2006), but this does not indicate that
the specific role for these regions is to, for example, gen-
erate subjective feelings.
A Role for the Cingulo-opercular Network
in Ambiguity
We found increased activity in this network in response to
surprised faces and ambiguous IAPS, but only when mak-
ing judgments that require the resolution of ambiguity.
This network has been shown to respond to ambiguity
in semantic (Thompson-Schill et al., 1997), visual mo-
tion (Sterzer et al., 2002), and face processing (Demos
et al., 2004) paradigms. This is consistent with the notion
that these regions are recruited during tasks where a
stimulus–response mapping needs to be made concern-
ing the ambiguous stimulus. In other words, ambiguity is
defined here as something that can be interpreted in more
than one way and, in turn, requires a selection between
competing alternatives. Research in neuroeconomics has
demonstrated a link between the AI and ambiguity in deci-
sion making, particularly in situations that require behav-
ioral flexibility that is dependent on contextual analysis
(Huettel, Stowe, Gordon, Warner, & Platt, 2006), although
ambiguous decisions (multiple possible outcomes with
unknown probabilities) are made more quickly than risky
decisions (multiple possible outcomes with known prob-
abilities). In other words, as in our own data showing that
these regions were still active during explicit valence judg-
ments even when RT was regressed out, activity in this
network seems to be unrelated to time on task (see also Neta,
Schlaggar, & Petersen, under revision, but see Grinband
et al., 2011).
Recent work on perceptual recognition found that ac-
tivity in the AI/FO region remained near baseline until the
moment of recognition, suggesting a relation to the mo-
ment of the decision itself (Ploran et al., 2007). Similarly,
the dACC was shown to learn from experiences by mon-
itoring and integrating outcomes (Behrens, Woolrich,
Walton, & Rushworth, 2007), playing a crucial role in
the update (late) period of such decision making, not
during the initial computation of uncertainty (see Singer,
Critchley, & Preuschoff, 2009, for a review). These find-
ings suggest that these regions may not be involved in
the process of deciding how to resolve ambiguous stim-
uli, but rather, they are crucial at the point during which
ambiguity is resolved per se. Taken together, these data
support the notion that activity in the cingulo-opercular
network is not simply related to task difficulty, and we
propose that this activity is most readily observed in tasks
that (1) require a decision relevant to the ambiguity (i.e.,
valence) and (2) require resolution rather than simple
detection or perception of ambiguity. Future work can
investigate how these transient responses to ambiguity
relate to the transient error-related activity identified in
the same regions (Neta et al., under revision).
Limitations
We note several limitations in interpreting the present find-
ings. First, the IAPS stimuli were significantly more arous-
ing than the face stimuli. As noted in the Methods section,
this is somewhat expected given the nature of these stim-
ulus categories, where the IAPS scenes carry much more
information than individual facial expressions. Given this
effect, the brain regions that were found to show a main
effect of stimulus may be, at least in part, modulated by this
difference in arousal. However, we focus here predomi-
nantly on the main effects of task, which is not impacted
by these arousal effects.
Second, although we compared two stimulus categories,
one of individual facial expressions and another that rep-
resents nonface emotional stimuli (IAPS scenes), we note
that approximately half of our scenes contained (at least
part of ) faces and human forms. As such, the present data
showing similar neural responses to ambiguity between
these two stimulus sets might still require a social element,
and the present result can be mostly cautiously summa-
rized by saying that this network was similarly involved in
processing ambiguity to directly presented faces and other
faces within more complex social scenes.
Finally, based on previous work showing that a region of
the dorsal amygdala tracks surprised faces during passive
viewing (Kim et al., 2003), we predicted that this region
would show greater activity to stimuli with an ambiguous,
as compared with clear valence, and that this effect might
be modulated by task demands. Indeed, this would fit well
with nonhuman animal work implicating the amygdala in
associative orienting, meaning that it functions to alert
other brain areas in instances of uncertainty when there
is a greater amount to learn (Gallagher, Graham, &
Holland, 1990; Kapp, Frysinger, Gallagher, & Haselton,
1979). However, we did not find a region of dorsal amyg-
dala in the omnibus ANOVA in the current study. It could
be that our task manipulation did not allow for the sen-
sitivity to pick up on the amygdala response to ambiguity
in the implicit nonvalence task (as our previous work has
demonstrated such an effect only for passive viewing). In-
deed, meta-analyses have demonstrated that amygdala re-
sponses to emotional stimuli can be diminished during an
active task, as compared with passive viewing (Costafreda,
Brammer, David, & Fu, 2008). Future work might help to
disentangle these effects by directly comparing passive
viewing to responses during both explicit and implicit
evaluations of ambiguous stimuli.
Neta, Kelley, and Whalen
555
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
o
9
c
0
n
7
_
9
a
/
_
j
0
o
0
c
3
n
6
3
_
a
p
_
d
0
0
b
3
y
6
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Conclusions
The brain regions that were recruited while making a valence
judgment about ambiguously valenced stimuli (i.e., the
cingulo-opercular network, ventral amygdala) were active
for both the ambiguous faces (surprised expressions) and
scenes. Moreover, the bias with which people resolve
ambiguity is consistent across these categories of stimuli.
Thus, the neural and behavioral responses required when
resolving ambiguity transcends stimulus boundaries and
is better defined by an uncertainty in stimulus–response
mapping that is created when people are required to make
a selection between competing alternatives. At a weaker
threshold, a targeted analysis of the amygdala based on
previous data showed that the dorsal amygdala showed
the highest response to surprised faces, but only during
the nonvalence task. Together, these data suggest that,
although some of the domain-general processes that come
on-line in the “face” of ambiguity are similar across many
categories of ambiguity (faces and scenes in the present
work; semantic, visual motion, and economic decision-
making ambiguity in previous work), there may be other
dedicated circuits for some social and emotional ambiguity
and, specifically, for biologically relevant learning situations
involving faces.
Acknowledgments
We thank J. V. Haxby for advice on experimental design and com-
ments on the manuscript as well as S. E. Petersen and D. D.
Wagner for fruitful discussions about results. We also thank
G. Wolford for statistical advising and comments on the manu-
script, C. J. Norris for advice on experimental design, and A. L.
Palmer and S. V. Fogelson for discussion about methods. Finally,
we thank R. A. Loucks for assistance in recruitment of participants
and help with data collection and survey data entry. This work
was supported by NIMH 080716.
Reprint requests should be sent to Maital Neta, Department of
Neurology, Washington University School of Medicine, Room 2220,
4525 Scott Ave., St. Louis, MO 63110, or via e-mail: maital@npg.
wustl.edu, maital@dartmouth.edu.
Note
1. Given our previous demonstrations of amygdala responses
to ambiguous facial expressions during passive viewing (Davis
et al., 2010; Whalen et al., 2009; Kim et al., 2003), we predicted
that there would be a dorsal amygdala response to ambiguity
that was also modulated by task demands but may be limited
to face stimuli. Accordingly, a small volume correction (see
fMRI Analysis) revealed a region in the dorsal amygdala (x, y,
z = −24, −3, −9) that was more active for surprised as com-
pared with clearly valenced expressions, only for the nonva-
lence task. This effect was not observed for the IAPS at this
more liberal threshold.
REFERENCES
Andersson, J. L. R., Hutton, C., Ashburner, J., Turner, R., &
Friston, K. J. (2001). Image distortion correction in fMRI:
A quantitative evaluation. Neuroimage, 16, 217–240.
Beck, A. T., Ward, C., & Mendelson, M. (1961). Beck Depression
Inventory (BDI). Archives of General Psychiatry, 4,
561–571.
Behrens, T. E. J., Woolrich, M. W., Walton, M. E., & Rushworth,
M. F. S. (2007). Learning the value of information in an
uncertain world. Nature Neuroscience, 10, 1214–1221.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., &
Cohen, J. D. (2001). Conflict monitoring and cognitive
control. Psychological Review, 108, 624–652.
Brunsson, N. (1985). The irrational organization: Irrationality
as a basis for organizational action and change.
Chichester, UK: Wiley.
Chang, L. J., Smith, A., Dufwenberg, M., & Sanfey, A. G. (2011).
Triangulating the neural, psychological, and economic
bases of guilt aversion. Neuron, 70, 560–572.
Chang, L. J., Yarkoni, T., Khaw, M. W., & Sanfey, A. G. (2012).
Decoding the role of the insula in human cognition:
Functional parcellation and large-scale reverse inference.
Cerebral Cortex.
Corbin, R. M. (1980). Decisions that might not get made. In T.
Wallsten (Ed.), Cognitive processes in choice and decision
behavior (pp. 47–67). Hillsdale, NJ: Erlbaum.
Costafreda, S. G., Brammer, M. J., David, A. S., & Fu, C. H. Y.
(2008). Predictors of amygdala activation during the
processing of emotional stimuli: A meta-analysis of 385 PET
and fMRI studies. Brain Research Reviews, 58, 57–70.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research, 29, 162–173.
Critchley, H. D., Elliott, R., Mathias, C. J., & Dolan, R. J. (2000).
Neural activity relating to generation and representation of
galvanic skin conductance responses: A functional magnetic
resonance imaging study. Journal of Neuroscience, 20,
3033–3040.
Davis, F. C., Johnstone, T., Mazzulla, E. C., Oler, J. A., & Whalen,
P. J. (2010). Regional response differences across the human
amygdaloid complex during social conditioning. Cerebral
Cortex, 20, 612–621.
Demos, K. E., Wig, G. S., Moran, J. M., & Kelley, W. M. (2004).
A role for ambiguity resolution in left inferior prefrontal
cortex. Washington, DC: Society for Neuroscience.
Dosenbach, N. U. F., Fair, D. A., Cohen, A. L., Schlaggar, B. L., &
Petersen, S. E. (2008). A dual-networks architecture of top–
down control. Trends in Cognitive Sciences, 12, 99–105.
Dosenbach, N. U. F., Fair, D. A., Miezin, F. M., Cohen, A. L.,
Wenger, K. K., Dosenbach, R. A. T., et al. (2007). Distinct
brain networks for adaptive and stable task control in
humans. Proceedings of the National Academy of
Sciences, U.S.A., 104, 11073–11078.
Dosenbach, N. U. F., Visscher, K. M., Palmer, E. D., Miezin,
F. M., Wenger, K. K., Hyunseon, C. K., et al. (2006). A core
system for the implementation of task sets. Neuron, 50,
799–812.
Duncan, J., & Owen, A. M. (2000). Common regions of the
human frontal lobe recruited by diverse cognitive demands.
Trends in Neurosciences, 23, 475–483.
Fox, M. D., Corbetta, M., Snyder, A. Z., Vincent, J. L., & Raichle,
M. E. (2006). Spontaneous neuronal activity distinguishes
human dorsal and ventral attention systems. Proceedings
of the National Academy of Sciences, U.S.A., 103,
10046–10051.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences, U.S.A., 102,
9673–9678.
Gallagher, M., Graham, P. W., & Holland, P. C. (1990).
The amygdala central nucleus and appetitive Pavlovian
556
Journal of Cognitive Neuroscience
Volume 25, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
o
9
c
0
n
7
_
9
a
/
_
j
0
o
0
c
3
n
6
3
_
a
p
_
d
0
0
b
3
y
6
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
conditioning: Lesions impair one class of conditioned
behavior. Journal of Neuroscience, 10, 1906–1911.
Grinband, J., Savitskaya, J., Wager, T. D., Teichert, T., Ferrera,
V. P., & Hirsch, J. (2011). The dorsal medial frontal cortex is
sensitive to time on task, not response conflict or error
likelihood. Neuroimage, 57, 303–311.
Hsu, M., Bhatt, M., Adolphs, R., Tranel, D., & Camerer, C. F.
(2005). Neural systems responding to degrees of uncertainty
in human decision-making. Science, 310, 1680–1683.
Huettel, S. A., Stowe, C. J., Gordon, E. M., Warner, B. T., &
Platt, M. L. (2006). Neural signatures of economic
preferences for risk and ambiguity. Neuron, 49, 765–775.
Johnstone, T., Somerville, L. H., Alexander, A. L., Oakes, T. R.,
Davidson, R. J., Kalin, N. H., et al. (2005). Stability of
amygdala BOLD response to fearful faces over multiple
scan sessions. Neuroimage, 25, 1112–1123.
Kahneman, D., Slovic, P., & Tversky, A. (Eds.) (1982). Judgment
under uncertainty: Heuristics and biases. Cambridge:
Cambridge University Press.
Kapp, B. S., Frysinger, M., Gallagher, M., & Haselton, J. R.
(1979). Amygdala central nucleus lesions: Effect on heart
rate conditioning in the rabbit. Physiology Behavior, 23,
1009–1117.
Kim, H., Somerville, L. H., Johnstone, T., Alexander, A., &
Whalen, P. J. (2003). Inverse amygdala and medial prefrontal
cortex responses to surprised faces. NeuroReport, 14,
2317–2322.
Kim, H., Somerville, L. H., Johnstone, T., Polis, S., Alexander,
A. L., Shin, L. M., et al. (2004). Contextual modulation of
amygdala responsivity to surprised faces. Journal of
Cognitive Neuroscience, 16, 1730–1745.
Kim, M. J., Loucks, R. A., Neta, M., Davis, F. C., Oler, J. A.,
Mazzulla, E. C., et al. (2010). Behind the mask: The influence
of mask-type on amygdala responses to fearful faces. Social
Cognitive & Affective Neuroscience, 5, 363–368.
Lane, R. D., Fink, G. R., Chua, P., & Dolan, R. J. (1997). Neural
activation during selective attention to subjective emotional
responses. NeuroReport, 8, 3969–3972.
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (2008).
International affective picture system (IAPS): Affective
ratings of pictures and instruction manual. Gainesville,
FL: University of Florida.
Levy, I., Snell, J., Nelson, A. J., Rustichini, A., & Glimcher,
P. W. (2010). Neural representation of subjective value
under risk and ambiguity. Journal of Neurophysiology,
103, 1036–1047.
Lundqvist, D., Flykt, A., & Öhman, A. (1998). The Karolinska
Directed Emotional Faces–KDEF. CD ROM from Department
of Clinical Neuroscience Psychology section, Karolinska
Institutet, ISBN 91-630-7164-9.
MacDonald, M. C., Pearlmutter, N. J., & Seidenberg, M. S.
(1994). The lexical nature of syntactic ambiguity resolution.
Psychological Review, 101, 676–703.
Maldjian, J. A., Laurienti, P. J., Kraft, R. A., & Burdette, J. H.
(2003). An auto-mated method for neuroanatomic and
cytoarchitectonic atlas-based interrogation of fMRI data sets.
Neuroimage, 19, 1233–1239.
Medford, N., & Critchley, H. D. (2010). Conjoint activity of
anterior insular and anterior cingulate cortex: Awareness and
response. Brain Structure and Function, 214, 535–549.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202.
Nee, D. E., Wager, T. D., & Jonides, J. (2007). Interference
resolution: Insights from a meta-analysis of neuroimaging
tasks. Cognitive and Affective Behavior Neuroscience,
7, 1–17.
Nelson, S. M., Dosenbach, N. U. F., Cohen, A. L., Wheeler, M. E.,
Schlaggar, B. L., & Petersen, S. E. (2010). Role of the anterior
insula in task-level control and focal attention. Brain
Structure and Function, 214, 669–680.
Neta, M., Norris, C. J., & Whalen, P. J. (2009). Corrugator
muscle responses are associated with individual differences
in positivity–negativity bias. Emotion, 9, 640–648.
Neta, M., Schlaggar, B. L., & Petersen, S. E. (under revision).
Seperable responses to error, ambiguity, and reaction time in
cingulo-opercular task control regions.
Neta, M., & Whalen, P. J. (2010). The primacy of negative
interpretations when resolving the valence of ambiguous
facial expressions. Psychological Science, 21, 901–907.
Platt, M. L., & Huettel, S. A. (2008). Risky business: The
neuroeconomics of decision making under uncertainty.
Nature Neuroscience, 11, 398–403.
Ploran, E. J., Nelson, S. M., Velanova, K., Donaldson, D. I.,
Petersen, S. E., & Wheeler, M. E. (2007). Evidence
accumulation and the moment of recognition: Dissociating
perceptual recognition processes using fMRI. Journal of
Neuroscience, 27, 11912–11924.
Rodd, J. M., Davis, M. H., & Johnsrude, I. S. (2005). The neural
mechanisms of speech comprehension: fMRI studies of
semantic ambiguity. Cerebral Cortex, 15, 1261–1269.
Sanfey, A. G., Rilling, J. K., Aronson, J. A., Nystrom, L. E., &
Cohen, J. D. (2003). The neural basis of economic
decision-making in the Ultimatum Game. Science, 300,
1755–1758.
Singer, T., Critchley, H. D., & Preuschoff, K. (2009). A common
role of insula in feelings, empathy, and uncertainty. Trends
in Cognitive Sciences, 13, 334–340.
Singer, T., Seymour, B., OʼDoherty, J., Kaube, H., Dolan, R. J., &
Frith, C. D. (2004). Empathy for pain involves the affective
but not sensory components of pain. Science, 303,
1157–1162.
Spielberger, C. D., Gorsuch, R. L., & Lushene, R. E. (1988).
STAI-Manual for the State Trait Anxiety Inventory.
Palo Alto, CA: Consulting Psychologists Press.
Sterzer, P., Russ, M. O., Preibisch, C., & Kleinschmidt, A. (2002).
Neural correlates of spontaneous direction reversals in ambiguous
apparent visual motion. Neuroimage, 15, 908–916.
Thompson-Schill, S. L., DʼEsposito, M., Aguirre, G. K., & Farah,
M. J. (1997). Role of left inferior prefrontal cortex in retrieval
of semantic knowledge: A reevaluation. Proceedings of the
National Academy of Sciences, U.S.A., 94, 14792–14797.
Tottenham, N., Tanaka, J., Leon, A. C., McCarry, T., Nurse, M.,
Hare, T. A., et al. (2009). The NimStim set of facial expressions:
Judgments from untrained research participants. Psychiatry
Research, 168, 242–249.
Tversky, A., & Kahneman, D. (1974). Judgment under uncertainty:
Heuristics and biases. Science, 185, 1124–1131.
Whalen, P. J., Davis, F. C., Oler, J. A., Kim, H., Kim, M. J., & Neta, M.
(2009). Human amygdala responses to facial expressions of
emotion. In P. J. Whalen & E. A. Phelps (Eds.), The human
amygdala (pp. 265–288). New York: Guilford Press.
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of
human functional neuroimaging data. Nature Methods, 8,
665–670.
Neta, Kelley, and Whalen
557
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
4
2
5
5
/
4
4
7
/
1
5
9
4
4
7
5
/
0
1
8
7
3
7
o
9
c
0
n
7
_
9
a
/
_
j
0
o
0
c
3
n
6
3
_
a
p
_
d
0
0
b
3
y
6
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1