METHODEN
More than the sum of its parts: Merging network
psychometrics and network neuroscience with
application in autism
Joe Bathelt1,2
, Hilde M. Geurts2
, and Denny Borsboom2
1Abteilung für Psychologie, Royal Holloway, University of London, Egham, Surrey, Vereinigtes Königreich
2Abteilung für Psychologie, University of Amsterdam, Amsterdam, die Niederlande
Schlüsselwörter: Netzwerk, Methoden, Psychometrics, Neuroimaging, Autism
Keine offenen Zugänge
Tagebuch
ABSTRAKT
Network approaches that investigate the interaction between symptoms and behaviours have
opened new ways of understanding psychological phenomena in health and disorder in recent
Jahre. In parallel, network approaches that characterise the interaction between brain
regions have become the dominant approach in neuroimaging research. In diesem Papier, Wir
introduce a methodology for combining network psychometrics and network neuroscience.
This approach utilises the information from the psychometric network to obtain neural
correlates that are associated with each node in the psychometric network (network-based
regression). Darüber hinaus, we combine the behavioural variables and their neural correlates in a
joint network to characterise their interactions. We illustrate the approach by highlighting the
interaction between the triad of autistic traits and their resting-state functional connectivity
associations. Zu diesem Zweck, we utilise data from 172 male autistic participants (10–21 years) aus
the autism brain data exchange (ABIDE, ABIDE-II) that completed resting-state fMRI and were
assessed using the autism diagnostic interview (ADI-R). Our results indicate that the network-
based regression approach can uncover both unique and shared neural correlates of
behavioural measures. Zum Beispiel, our example analysis indicates that the overlap between
communication and social difficulties is not reflected in the overlap between their functional
brain correlates.
ZUSAMMENFASSUNG DES AUTORS
The article introduces a method to combine common practices in network psychometrics and
network neuroimaging. Namely, we use the unique variance in behavioural measures as
regressors to identify unique neural correlates. This enables the description of brain-level and
behavioural-level data into a joint network while keeping the dimensionality of the results
manageable and interpretable. We illustrate this approach by showing the network of autistic
traits and their correlates in resting-state functional connectivity.
EINFÜHRUNG
The traditional view of psychiatric conditions conceptualizes mental disorders as latent con-
structs that are expressed in a set of manifest symptoms. Zum Beispiel, autism spectrum disor-
ders (ASDs) are typically described as a disorder rooted in an (as yet unknown) dysfunction
Zitat: Bathelt, J., Geurts, H. M., &
Borsboom, D. (2022). More than the
sum of its parts: Merging network
psychometrics and network
neuroscience with application in
autism. Netzwerkneurowissenschaften,
6(2), 445–466. https://doi.org/10.1162
/netn_a_00222
DOI:
https://doi.org/10.1162/netn_a_00222
Erhalten: 23 Juni 2021
Akzeptiert: 8 Dezember 2021
Konkurrierende Interessen: Die Autoren haben
erklärte, dass keine konkurrierenden Interessen bestehen
existieren.
Korrespondierender Autor:
Joe Bathelt
joe.bathelt@rhul.ac.uk
Handling-Editor:
Cornelis Jan Stam
Urheberrechte ©: © 2021
Massachusetts Institute of Technology
Veröffentlicht unter Creative Commons
Namensnennung 4.0 International
(CC BY 4.0) Lizenz
Die MIT-Presse
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
rooted in the biology of the human system, which manifests its effects in the domains of social
interaction, social communication, and restricted, repetitive patterns of behaviour, interests, oder
Aktivitäten (DSM-5). In this view, the manifested difficulties—typically interpreted as symptoms—
are viewed as indicators of a latent condition; in accordance, the information present in these
symptoms is commonly aggregated into a single score (z.B., by counting the number of symp-
toms or averaging subscales of questionnaires). This score is interpreted as a measure of the
severity of the mental disorder, and usually functions as a dependent variable in designs geared
to uncover the genetic background or neural correlates of the disorder.
An alternative view, which has emerged in recent years, interprets the relation between
symptoms and disorders differently, and emphasises the dynamic interaction between symp-
toms (Borsboom, 2017; Borsboom et al., 2018; Robinaugh et al., 2020). In this alternative net-
work approach to mental disorders, causal interaction between symptoms themselves is
brought to the forefront and disorders are seen as clustered states of symptoms that mutually
reinforce each other. Zum Beispiel, in the case of autism, restricted interests may limit the time
spent in social interactions, which may lead to reduced social communication skills, und das
limited success in social communication may in turn reinforce restricted interests. From this
Perspektive, an important part of the basis of mental disorders like autism would be expected
to lay in the disposition to develop individual symptoms and in the processes that govern inter-
actions between them. Infolge, the network approach does not focus on the level of aggre-
gate scores or functions thereof, but on the patterns of association that arise between symptom
Variablen. A host of statistical techniques has been developed to estimate and analyse such
symptom networks (Borsboom et al., 2021). This way of working has become a popular meth-
odology approach to the study of symptomatology and is beginning to find its way in clinical
üben (Kroeze et al., 2017).
By focusing attention on the interaction between symptoms, network approaches open up
new ways to study how mental disorders may be related to the brain. Insbesondere, von dem
point of view of network theory, neuroscientific approaches should focus on (A) mechanisms
or dysfunctions that lead symptoms to arise (z.B., mechanisms that promote repetitive behav-
iours), Und (B) processes that couple one symptom to another (z.B., mechanisms that link
repetitive behaviours to, sagen, problems in social interaction). To the extent that biological pro-
cesses are specific to individual symptoms or symptom-symptom interactions, aggregation of
scores is methodologically inadvisable because the shared variance between symptoms is
then likely to correspond to a biological amalgam of processes and mechanisms that will
be hard to tease apart.
Autism spectrum condition (ASC) is an ideal test case for applying the network perspective to
resolve ongoing debates about the associations between core behavioural features. ASC is char-
acterised by atypicalities in social interaction and communication, alongside restricted and
repetitive behaviours and narrow interests (RRBI). Jedoch, there is some debate regarding
the separation between the domains. Auf der einen Seite, it has been noted that atypicalities in all
three domains frequently co-occur (Wing & Gould, 1979), possibly reflecting a shared genetic
or neurocognitive mechanism. Andererseits, the ‘fractionable triad’ account suggests that
unique mechanisms contribute to difficulties in each domain (Happé & Ronald, 2008). Factor
analyses of scores from diagnostic interviews or behaviour ratings are inconclusive. Several
studies indicate that a large proportion of variance loads onto a single factor in principal com-
ponent analysis (Constantino et al., 2004; Szatmari et al., 2002), which is interpreted as support
for a single factor underlying the triad of autistic behaviours. Noch, other studies found support for
a factor structure with two, three, or more factors (Berument et al., 1999; DiLalla & Rogers,
1994; Kim et al., 2019; van Lang et al., 2006; Wadden et al., 1991). A meta-analysis indicated
RRBI:
Repetitive and restricted behaviours
or interests; potential domain of
autism spectrum condition
behaviours.
Netzwerkneurowissenschaften
446
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
that studies that identified more than one factor, typically divided the social aspects (sozial
interaction, communication) from the putatively less social aspects (RRBIs) (Mandy & Skuse,
2008). Jedoch, a more recent study that used a hierarchical cluster model indicated that com-
munication and RRBIs segregated together with social interaction as a separate factor (Kim
et al., 2018). Zusammenfassend, the behavioural evidence suggests that the domains of atypicality
in autism (social interaction, communication, RRBIs) are linked but can be fractionated. Das
provides an ideal test case for a network approach that aims to capture the relation between
associated variables.
In addition to investigations at the behavioural level, methodologies developed in the net-
work approach may be useful in disentangling neural correlates on symptoms and interactions
between them. In the case of ASC, neuroimaging research has associated the autistic trait triad
with partially distinct networks of brain regions. Social cognition associated with the social
interaction aspect of the autistic trait triad is thought to be supported by a distributed network
of brain regions, collectively referred to as the “social brain” (Müller & Fishman, 2018). Das
network comprises of the amygdala, the dorsal and ventral medial prefrontal cortices (dmPFC,
vmPFC), the anterior and posterior cingulate cortex (ACC, PCC), the posterior superior temporal
Sulkus (pSTS), the temporoparietal junction (TPJ), the inferior occipital gyrus (IOG), the fusiform
face area (FFA), and the insula. These regions showed increased responses in tasks that tap
social processing, including the decoding of facial expressions (Kadosh et al., 2010) and theory
of mind (Schurz et al., 2014). These social brain regions show reduced activation during social
processing tasks in autism (Di Martino et al., 2009; Patriquin et al., 2016). Außerdem, Die
connectivity between regions of the social regions is weaker in autism, yet the connectivity with
regions outside of the social brain is stronger compared to neurotypicals (Hagen et al., 2013;
Kana & Wadsworth, 2012; Supekar et al., 2013). Differences in social brain connectivity are
also apparent at rest. Assaf et al. (2010) reported reduced connectivity of the canonical default
mode (DMN) and salience network (SN) that contain social brain regions. Reduced connectiv-
ity of the DMN and SN was related to the severity of social symptoms. Communication atyp-
icalities in autism have been linked to differences in classic language areas, such as Broca’s and
Wernicke’s areas. Individuals with autism show reduced activation in the left inferior frontal
Kortex (Brocas Gegend) and increased activation in the superior temporal gyrus ( Wernicke’s area)
in lexical and semantic processing tasks (Harris et al., 2006; Just et al., 2004; Kana et al., 2006).
Differences in connectivity within the language network is also apparent at rest. The left inferior
frontal cortex (Brocas Gegend) has been found to show reduced connectivity in children and ado-
lescents with autism, while the superior temporal gyrus ( Wernicke’s area) shows reduced con-
nectivity in autistic adults (Y. Lee et al., 2017). Differences related to communication difficulties
in autism are also observed outside of the core language network. This includes increased acti-
vation of homologous areas in the right hemisphere during language processing (Just et al.,
2004; Kana et al., 2006) and activations in areas typically involved in visuospatial processing
(Kana et al., 2006; Samson et al., 2012), das ist, the lateral occipital cortex and inferior parietal
Sulkus. Ähnlich, studies that investigated functional connectivity at rest indicated an associa-
tion of higher communication difficulties with increased connectivity across the whole brain
(Uddin et al., 2013), reduced connectivity of the DMN and SN (Assaf et al., 2010; Maximo
& Kana, 2019), and increased connectivity of the lateral occipital cortex (Jung et al., 2017).
Repetitive and stereotyped behaviours and narrow interests (RRBIs) have been associated with
differences in systems involved in sensation, motor control, and reward-related processing.
Speziell, higher RRBIs scores are linked to greater connectivity between the striatum with
occipital and frontal areas (Dupong & Di Martino, 2020), alongside lower connectivity of the
striatum with cortical motor and sensory areas (Abbott et al., 2018; Maximo & Kana, 2019).
Netzwerkneurowissenschaften
447
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
Außerdem, higher RRBIs scores are associated with lower connectivity of fronto-parietal
Regionen (Abbott et al., 2018; J. M. Lee et al., 2016), broadly consistent with the DMN. Weiter-
mehr, SN hyperconnectivity has been found to predict RRBIs scores in autistic children (Uddin
et al., 2013).
As is apparent from the summary above, there is a rich literature on the brain correlates of
autistic traits. In manchen Fällen, the associations overlap. Zum Beispiel, the default mode and SN
are implicated across all domains of the autistic triad. Other associations appear to be unique
to each domain, Zum Beispiel, the involvement of the basal ganglia in RRBIs. The interplay
between brain systems that are associated with autistic behaviours may be key. Zum Beispiel,
the involvement of the same brain system may explain the overlap of characteristics at the
behavioural level. Jedoch, the interaction between autistic traits and their neural correlates
has so far not been investigated. A network approach that combines behavioural and brain
measures is primed to fill this gap. Jedoch, a clear link between the network psychometrics
approach and brain imaging research has so far been missing. Establishing this connection has
the potential to open a new line of research that investigates the relations between brain-level
and symptom-level networks and, thereby, holds the potential to uncover mechanisms of com-
pensation or vulnerability. Jedoch, the current approaches in psychiatric neuroimaging are
not well suited to this task. The dominant approaches are to either contrast groups of cases with
groups of controls, or to identify correlates of symptoms or putative cognitive endophenotypes.
Both approaches have limitations as has been discussed at length elsewhere (Borsboom, 2017;
Robinaugh et al., 2020). Most importantly for network psychometrics, these approaches implic-
itly assume that psychiatric conditions are latent constructs that determine, and therefore are
indicated by, symptoms or cognitive endophenotypes. In this manuscript, we provide a concep-
tual bridge that links network psychometrics with established methods in human neuroimaging.
Namely, we construct brain-level correlates by using the unique variance in behavioural mea-
sures as regressors. Subsequently, we explore the relationship between the symptom-level net-
work and its neural correlates.
To illustrate this approach, we focus on the behaviours associated with ASC. We use large
databases that collected resting-state fMRI (rsfMRI) and characterised autistics traits with the
same assessment instrument to study the neural correlates of symptom networks. Using these
Daten, we applied the connectome-based predictive modelling (CPM) method (Shen et al.,
2017) to identify the rsfMRI correlates of autistic traits. To characterise the relations between
behavioural and neuroimaging data, we applied network psychometric and causal inference
Methoden. Based on the psychometric literature on autistic traits summarized above, Wir
expected a closer association between social and communication difficulties than between
these traits with RRBIs. We did not have a strong expectation regarding the association
between the neural correlates.
MATERIALS AND METHODS
Network-Based Regression Method
The following section describes new methods for constructing networks based on brain cor-
relates of behavioural or cognitive measures.
Behaviour Network–Based Regressors
In neuroimaging, out-of-scanner task performance scores are often used to identify the neural
correlates of behaviours, particularly for analyses of resting-state data (fMRT, M/EEG) or struc-
tural data (diffusion-weighted imaging, morphological). Jedoch, the use of raw task scores
CPM:
Connectome-based predictive
modelling; approach to establish an
association between connectome
measures and behavioural measures.
Netzwerkneurowissenschaften
448
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
Tisch 1.
residuals after regression (4–6)
Correlation between ADI-R domain scores for either the original scores (1-3) oder der
1. Sozial
2. Communication
3. RRBI
4. Social residual
5. Communication residual
6. RRBI residual
1
1.00
0.66
0.38
0.73
0.56
0.38
2
0.66
1.00
0.36
0.00
0.93
0.36
3
0.38
0.36
1.00
0.00
0.00
1.00
4
0.73
0.00
0.00
1.00
0.00
0.00
5
0.56
0.93
0.00
0.00
1.00
0.00
6
0.38
0.36
1.00
0.00
0.00
1.00
has been criticised, because the task scores reflect many extraneous influences. One way to
create better regressors is to calculate the shared variance of several measures that are thought
to tap the same psychological construct (Poldrack & Yarkoni, 2016). This approach of creating
latent variables views the psychological or psychiatric constructs as latent entities indicated by
the scores. Im Gegensatz, in network psychometrics, one may be more interested in the unique
variance of each measure to create the nodes that make up the network. For neuroimaging,
obtaining correlates for measures while controlling for other measures is already common
üben, typically in the context of controlling for nuisance variables. This can be achieved
by simply regressing the effect of other variables from each variable and retaining the residual.
Zum Beispiel, in a network of three variables A, B, and C, the unique variance in A is given by
the residual term, epsilon, in the regression equation: yA = βBXB + βCXC + XIntercept + (cid:1). Als
expected, the association between behavioural scores was removed after regressing the effect
of the other behavioural scores (siehe Tabelle 1). The residual terms can be used as regressors to
obtain the neural correlates of the unique variance of each behavioural measure. In this way,
the nodes within a psychometric network can be used as regressors in established neuroim-
aging pipelines. This provides a methodological and conceptual bridge between network psy-
chometrics and neuroscience.
Associations Between Behaviour Scales and Functional Brain Connectivity
To identify the associations between functional connectivity and behavioural ratings, we used
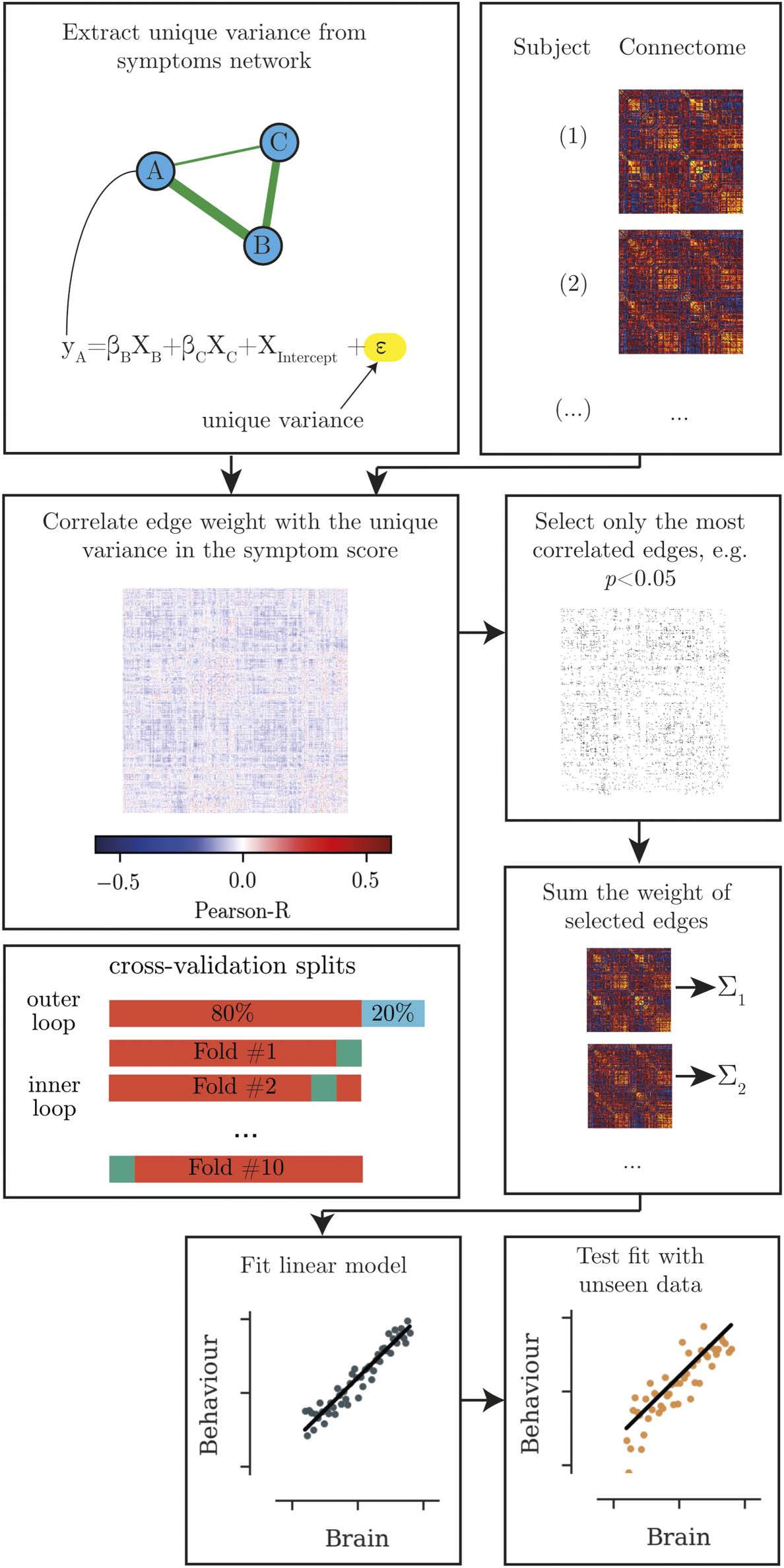
the CPM approach described by Shen et al. (2017) (siehe Abbildung 1 for an overview). Zusamenfassend, a set
of edges is identified that correlate with the behaviour ratings below a certain p value thresh-
alt. Dann, the edge weights are summed into a brain score and entered into a regression
model to estimate the association between the brain score and behaviour rating. For the cur-
rent analysis, we identified positively and negatively associated edges separately and entered
the summed edge weight for positively and negatively associated edges into a common mul-
tiple regression model:
ysymptom ¼ βBrainþ XBrainþ þ βBrain− XBrain− þ XIntercept þ (cid:1)
We were faced with multiple methodological choices that were difficult to determine a
priori, namely the number of regions of interest (ROIs) in the parcellation, the edge definition,
p value threshold, and global signal regression strategy. daher, we employed a shuffle split
cross-validation to find the combination of parameters that led to the best prediction of behav-
iour ratings in unseen data. For this parameter tuning, we randomly split the data into an 80%
training and 20% test set in an outer loop (siehe Abbildung 1 for an illustration of the cross-validation
Netzwerkneurowissenschaften
449
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 1. Overview of the analysis steps for identifying the brain correlates of behavioural measures. Erste, the unique variance in each
behavioural scale is calculated. The edges that shows the strongest correlation with the unique variance in the behavioural score are extracted.
The summed edge weight of the most highly associated edges is used to build a regression model, which is then tested in unseen data.
Netzwerkneurowissenschaften
450
Merging network psychometrics and network neuroscience
splits). We identified the associated edges and fitted the regression model in the training set
using 10-fold cross-validation. Dann, we compared the quality of prediction by using the
model with the held-out test data from the outer loop by calculating the correlation between
predicted and observed values (Shen et al., 2017). The parameter combination that pro-
duced the best prediction in unseen data used 300 ROIs, defined the edge weight through
Pearson correlation without global signal regression, and set the association p value thresh-
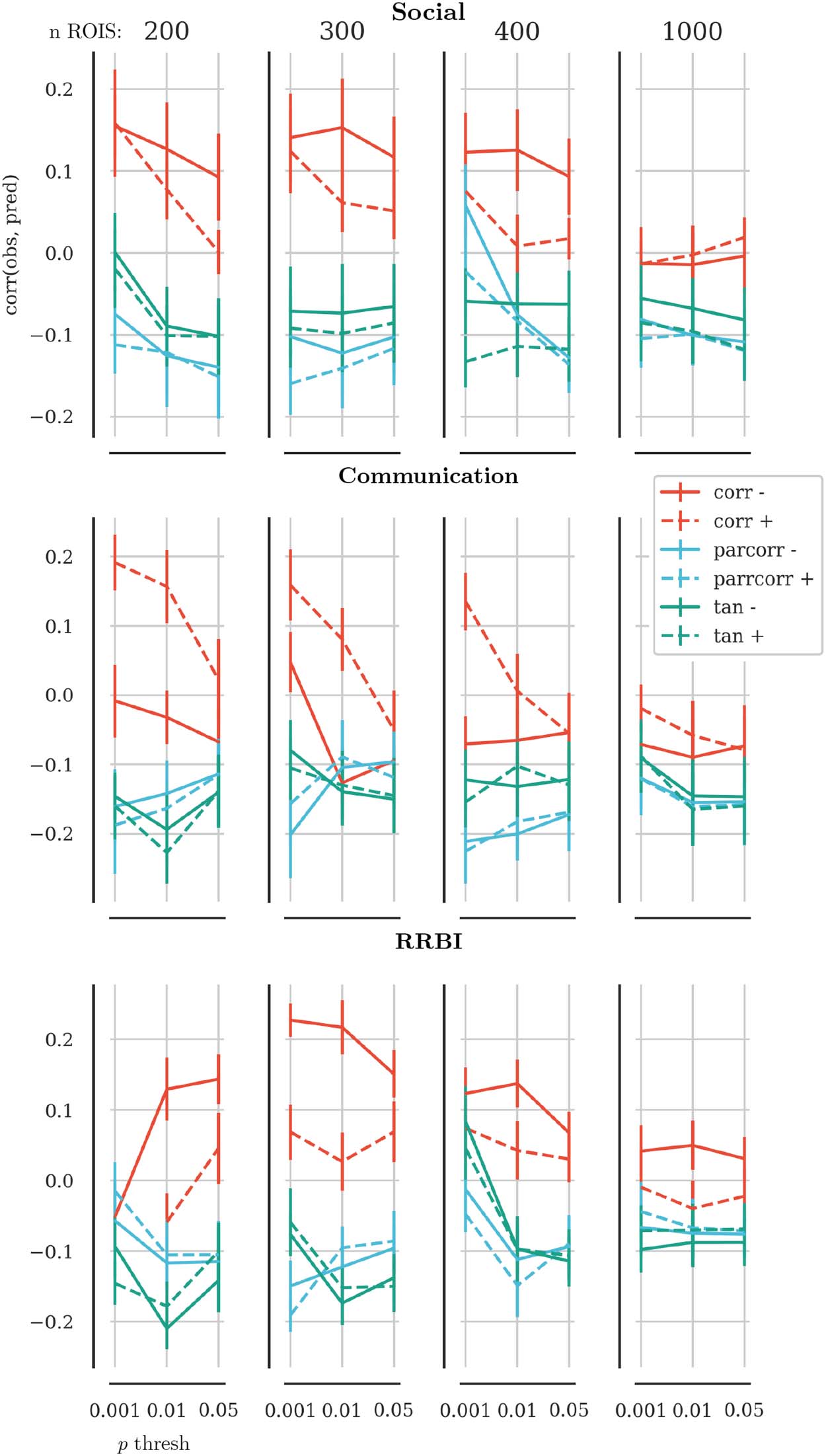
old to p < 0.001 (see Figure 2 for results of the parameter tuning). For the evaluation of the
final model, we only retained edges that were included across 7 out of 10 splits of the data.
We compared the performance of this model against 5,000 random permutations of the
behavioural data.
General Data Preparation
The following section describes the data and preprocessing steps that were used to illustrate
the network-based regression approach.
Participants
The analysis was based on data taken from the first and second Autism Brain Imaging Data
Exchange database (ABIDE; Di Martino et al., 2014; and ABIDE-II; Di Martino et al., 2017).
Both databases collated resting-state fMRI and phenotypic data from autistic participants from
19 international sites. There was no prior coordination between sites, which means that the
sites differed in their fMRI acquisition protocols and diagnostic procedures. Because of the
ensuing variability, we applied selection criteria to arrive at a more homogeneous sample.
Namely, we selected only male participants, because women were not well represented.
Furthermore, we selected only participants over 10 years of age, because of different scoring
criteria on the ADI-R for younger children. We further excluded participants older than 21 years,
because the relatively few participants older than 21 years were spread over a large age range.
In addition, we only selected participants with complete, research-reliable ADI-R assessments
and with complete structural and functional MRI that was rated as useable by expert human
assessors (note that the quality ratings are distributed with the phenotypic data). For further
quality control, we excluded 13 participants because their fMRI data fell outside the recom-
mended range on established quality metrics (framewise displacement > 0.5 mm; Power
et al., 2012; DVARS > 5%; Burgess et al., 2016). The final sample consisted of 172 autistic par-
ticipants (ABIDE: n = 127, ABIDE-II: n = 45; siehe Tabelle 2 for sample characteristics). Please note
that we were not aiming to obtain a representative sample. The purpose of the analysis was to
demonstrate the potential use of a network-based regression method. The selection criteria
were intended to create a more homogeneous sample with good quality imaging data.
Assessment of Autism Characteristics
The Autism Diagnostic Interview-Revised (ADI-R) is a standardised diagnostic interview for pri-
mary caregivers (Rutter et al., 2003). It focuses on a description of a child’s behaviour when
they were 4–5 years old and their current behaviour. An autism diagnosis is made with an
ADI-R algorithm that consists of 37 extracted items. In addition to a total score, subscores for
autistic traits (social interaction, communication, RRBI) can be obtained. The ADI-R shows a
high interrater agreement (0.94–0.96; Cicchetti et al., 2008) and high convergence with clin-
ical team assessments and another commonly used assessment protocol (75% Vereinbarung;
Mazefsky & Oswald, 2006), das ist, the autism observation schedule (ADOS). The ADI-R pro-
tocol is adjusted depending on the chronological and mental age of the participant. Fewer items
are included for children younger than 10 years or with an intellectual functioning outside of
ADI-R:
Autism Diagnostic Interview–
Überarbeitet; assessment for behaviours
associated with autism spectrum
condition.
Netzwerkneurowissenschaften
451
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 2. Results of the parameter tuning. The x-axis shows the p value threshold used to select edges that were associated with behaviour
ratings scores. The y-axis shows the correlation between the predicted scores and the observed scores in held-out data. Solid lines show results
based on connectomes that were constructed without global signal regression; dashed lines indicate results based on connectomes with global
signal regression. Red lines show results based on connectomes that used Pearson correlation as the edge definition, blue lines show the results
for partial correlations, and green lines for tangent-space embedding. The panel in each row show the results for brain parcellations with 200,
300, 400, Und 1,000 ROIs. The overall best results across behaviour scales were observed for 300 ROIs.
Netzwerkneurowissenschaften
452
Merging network psychometrics and network neuroscience
Tisch 2.
Sample characteristics
Alter
Mean ± SE
13.87 ± 0.204
Full-scale IQa
108.94 ± 1.015
Min-Max
10.04–20.09
86–149
N [%]
ADI-Rb
Sozial
Verbal
RRBI
19.59 ± 0.388
15.51 ± 0.316
5.94 ± 0.196
11,202
45,505
0–12
164 [95.35]
167 [97.09]
142 [82.56]
a Assessments: 86 WASI, 53 WISC-IV, 33 andere.
b Cutoff scores: Social > 10, Verbal > 8, RRBI > 3.
the typical range. Because the current analysis aimed to compare associations of scores, Wir
restricted our analysis to children above 10 years with a full-scale IQ in the typical range to
ensure that differences in ADI-R scores reflected the relative ranking of the severity of difficulties
in each domain. Please note that only summary scores for each domain (social interaction,
communication, RRBI) were part of the ABIDE data release, das ist, the individual items used
to calculate the summary scores were not available for analysis.
fMRI Processing
The data were processed using the standard configuration of the Configurable Pipeline for the
Analysis of Connectomes (C-PAC, v. 1.6.2; Craddock et al., 2013). C-PAC is an automated
state-of-the-art pipeline for reproducible processing of large-scale data. The current analysis
was run using the singularity image distributed via the C-PAC website to process the data
on a high-performance computing cluster. The configuration file for the pipeline is provided
via the Open Science Framework so that the results can be exactly reproduced (https://osf.io
/my8g6/?view_only=85c585b0f4ee4abb8e700afd2693b8c3). The full details of the process-
ing pipeline are available from the C-PAC website (https://fcp-indi.github.io/docs/user
/preprocessing). The following provides an overview of the preprocessing steps. For anatom-
ical preprocessing, a nonlinear transform between images and a 2-mm MNI brain-only tem-
plate were calculated using Advanced Normalisation Tools (ANTs) (Tustison et al., 2014). Der
images were then skull-stripped using AFNI’s 3dSkullStrip and subsequently segmented into
white matter ( WM), grey matter (GM), and cerebrospinal fluid (CSF) using FSL’s fast tool
(Schmied, 2002). The resulting WM mask was multiplied by a WM prior map that was trans-
formed into individual space using the inverse of the linear transforms calculated through
ANTs. A CSF mask was multiplied by a ventricle map derived from the Harvard–Oxford atlas
distributed with FSL (Smith et al., 2004). Skull-stripped images and GM tissue maps were trans-
formed into MNI space at 2-mm resolution.
For functional preprocessing, motion correction was performed using a two-stage approach
in which the images were first co-registered to the mean of the fMRI sequence, and then a new
mean was calculated and used as the target for a second registration (AFNI 3dvolreg; Cox &
Jesmanowicz, 1999). A 7-degree of freedom linear transform between the mean fMRI and the
structural image was calculated using FSL’s boundary-based registration (Greve & Fischl,
2009). Nuisance variable regression (NVR) was performed on the motion-corrected data using
a second-order polynomial, a 24-regressor model of motion (Friston et al., 1996), five nuisance
signals identified via principal components analysis of signals obtained from WM (CompCor;
Netzwerkneurowissenschaften
453
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
Behzadi et al., 2007), and the mean CSF signal. WM and CSF signals were extracted using the
previously described masks after transforming the fMRI data to match them in 2-mm space by
using the inverse of the linear fMRI-sMRI transform. The NVR procedure was performed twice,
with and without the inclusion of the global signal as a nuisance regressor. The results of the
processing strategies were both entered into the predictive model in the later stages of the
Analyse (siehe unten). The residuals of the NVR procedure were bandpass filtered (0.001 Hz <
f < 0.1 Hz), written into MNI space at 2-mm resolution and subsequently smoothed using a
6-mm full-width half-maximum kernel.
Then, the time series for the parcellation described by Schaefer et al. (2017) were extracted
with 200, 300, 400, and 1,000 parcels. We chose this parcellation because it provides a better
account of fMRI activations than previous data-driven parcellations and is based on a large
representative database (N = 1,489). We evaluated the optimal resolution for the purpose
of the current analysis in the predictive model. To ensure that all included regions were suf-
ficiently covered in all participants, we calculated the mean functional image, thresholded and
binarized it at 70% intensity, and calculated the overlap between the atlas ROIs and the result-
ing image in all participants. ROIs that had less than 50% overlap with the binarised intensity
image in any participant were excluded from the analysis. This resulted in the exclusion of
9 ROIs for the 200 ROI atlas, 16 for the 300 ROI atlas, 24 for the 400 ROI atlas, and 68 for the
1,000 ROI atlas. The excluded ROIs were located in the frontal and temporal pole.
Functional Connectome Construction
We calculated the functional connectome matrices using three approaches, namely correla-
tion, partial correlation, and tangent-space embedding. For the correlation approach, we cal-
culated the Pearson correlation between each pairwise combination of ROIs in the parcellation.
For the partial correlation approach, the Pearson correlation of each pairwise combination of
ROIs was calculated after regressing the effect of other ROIs from both time series. For the
tangent-space embedding approach, we used the method described by Varoquaux et al.
(2010) that models each participant as a deviation from the group average connectome. For
all approaches, the implementation of the method in Nilearn v. 0.6.2 was used (Abraham
et al., 2014).
To reduce the influence of extraneous variables, we applied a similar regression approach
to the Human Connectome Project Mega-Trawl analysis (https://db.humanconnectome.org
/megatrawl/HCP820_MegaTrawl_April2016.pdf). Namely, we regressed the effect of age,
age2, acquisition site, framewise displacement, spatial root-mean-square of the data after tem-
poral differencing (DVARS), intracranial volume (ICV), and total GM volume from each edge
in the functional connectome in an ordinary least-squares regression model. ICV and total GM
volume were estimated using FreeSurfer.
Quality Control
Functional MRI connectomics have been shown to be particularly sensitive to participant
movement (Power et al., 2012). In order to mitigate the influence of motion, we applied a
combination of steps. First, we only included participants with structural and functional data
that were rated as useable by expert human assessors. These assessments are included in the
phenotypic data file of the ABIDE and ABIDE-II database. For some sites, scans were assessed
by multiple assessors. We only included a participant if all assessors rated the data as useable.
Furthermore, we evaluated the functional data using metrics calculated using the MRIQC
pipeline (Esteban et al., 2017). We only included data that met conservative thresholds for
Network Neuroscience
454
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
n
e
n
_
a
_
0
0
2
2
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Merging network psychometrics and network neuroscience
framewise displacement (<0.5 mm; Power et al., 2012) and DVARS (<5%; Burgess et al.,
2016). In the remaining participants, the effect of participant motion was mitigated by control-
ling for nuisance signals, including noise components estimated in the WM (Behzadi et al.,
2007). Furthermore, we regressed mean framewise displacement and standardised DVARS
from each edge in the functional connectome. The quality metrics were not associated with
social and communication scores (all p > 0.16 unkorrigiert). RRBI scores were associated with
DVARS (p = 0.032 unkorrigiert), but not framewise displacement (p = 0.214 unkorrigiert). Wir
compared the effect of different de-confounding strategies as implemented in the confounds
package for Python (Raamana, 2020) using cross-validation. Regressing the effect of confound-
ing variables showed the numerically highest prediction but the differences between con-
founding strategies were negligible (augment: mean = 0.008, SE = 0.0011; dummy: mean =
0.008, SE = 0.0011; residualize: mean = 0.009, SE = 0.0011, F(2, 31638) = 0.62, p = 0.54;
all values indicate correlations between predicted and observed scores in unseen data).
Confound regression was employed for confound removal for the final analysis.
Interpretation of Imaging Results
To aid the interpretation of the neuroimaging associations, we obtained anatomical labels for
the centroid coordinates of the nodes that were implicated in the association between rsfMRI
connectivity and behavioural measures. For this purpose, we used FSL’s atlasquery tool with
the Harvard–Oxford Cortical Structural atlas. We report most likely labels for each centroid
coordinate.
Construction of the Behaviour and Brain Score Network
To assess the relations between behavioural scores and their neural correlates, we estimated
the network structure of a network based on the behavioural scores and the total brain
score for each behavioural measure. To construct the total brain score, the regression
weights were applied to the positive and negative brain scores. To determine the edges
of the network, we identified an unregularized Gaussian graphical model by minimizing
the extended Bayesian information criterion (BIC) using the glasso algorithm and stepwise
model selection. These analyses were performed in R (Ausführung 3.5.0) using the bootnet
v1.4.3 and qgraph v1.6.5 packages.
Subsequently, we identified the likely causal direction of the network edges (Pearl, 2014) In
the network of behavioural scores, rsfMRI correlates, and a combined network with behav-
ioural and rsfMRI correlate notes. For this purpose, we computed a Bayesian network,
described in a directed acyclic graph (DAG), using structure learning algorithms implemented
in the bnlearn v4.5 package (Scutari, 2009). The structure learned by these algorithms can
contain directed and undirected edges. We compared the structure learned through different
Algorithmen, namely the Grow-Shrink (Margaritis, 2003), Incremental Association (Tsamardinos
et al., 2006), Fast Incremental Association (Yaramakala & Margaritis, 2005), Interleaved Incre-
mental Association (Tsamardinos et al., 2003), and Max-Min Parents and Children (Tsamardinos
et al., 2006) Algorithmen. We also compared the structure learned when using different indepen-
dence test statistics, namely Pearson correlation, Fisher’s Z, Monte-Carlo permutation, Und
gegenseitige Information.
ERGEBNISSE
The CPM methods indicated a combination of edge weights that could predict behaviour
scores in held-out data (Sozial: mean = 0.14, SE = 0.054; Communication: mean = 0.05,
DAG:
Directed acyclic graph; gerichtet
graph that never forms a closed loop.
Netzwerkneurowissenschaften
455
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
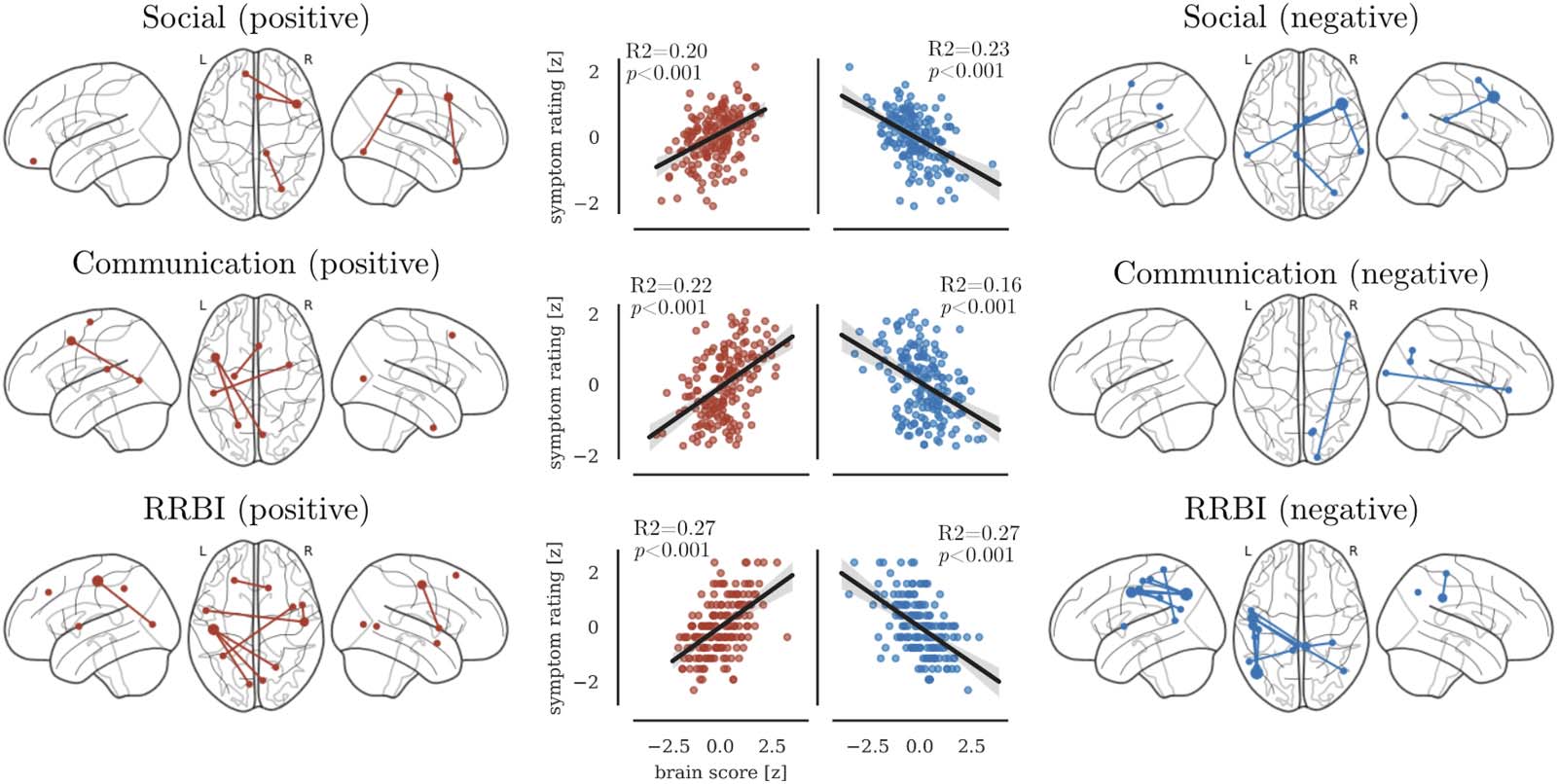
Figur 3. Association between unique variance in ADI-R domains with edges of the functional connectome identified through CPM. The left
panel shows edges that were positively associated with scores. The right panel shows negatively associated edges. The scatter plots show the
association between the summed brain score and the symptom ratings across the entire sample.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
SE = 0.043, RRBI: mean = 0.23, SE = 0.024; mean correlation between observed and pre-
dicted scores across 10 random 80/20 splits) and were significantly better in predicting behav-
iour scores compared to scrambled data (Sozial: p = 0.021; Communication: p = 0.014, RRBI:
p = 0.011; p value based on 5,000 permutations).
Social difficulties were positively associated with connections of the right prefrontal cortex,
and connections between the left fusiform gyrus and right postcentral gyrus (siehe Abbildung 3 Und
Tisch 3). Außerdem, social difficulties were negatively associated with connections between
the right middle frontal cortex with parietal areas in the left and right hemisphere (siehe Abbildung 3
und Tisch 3). Communication difficulties were positively associated with connections between
parietal areas in the left hemisphere with medial frontal and occipital areas (siehe Abbildung 3 Und
Tisch 3). Negative association between communication scores and rsfMRI connectivity were
found for connections between the right occipital areas with the inferior frontal gyrus and
precuneus (siehe Tabelle 3). RRBI scores showed the most extensive associations. RRBI scores
were positively associated with connections between the left postcentral gyrus with occipital
Bereiche, and between the right postcentral gyrus with the left insula. RRBI scores were negatively
associated with connections between left pre- and postcentral gyrus with occipital and poste-
rior cingulate areas (siehe Abbildung 3 und Tisch 3).
Combined Behaviour and Brain Network
We compared the network structure of a network based on behavioural measures and a net-
work based on the neural correlates of the behavioural measures (siehe Tabelle 4 for correlations
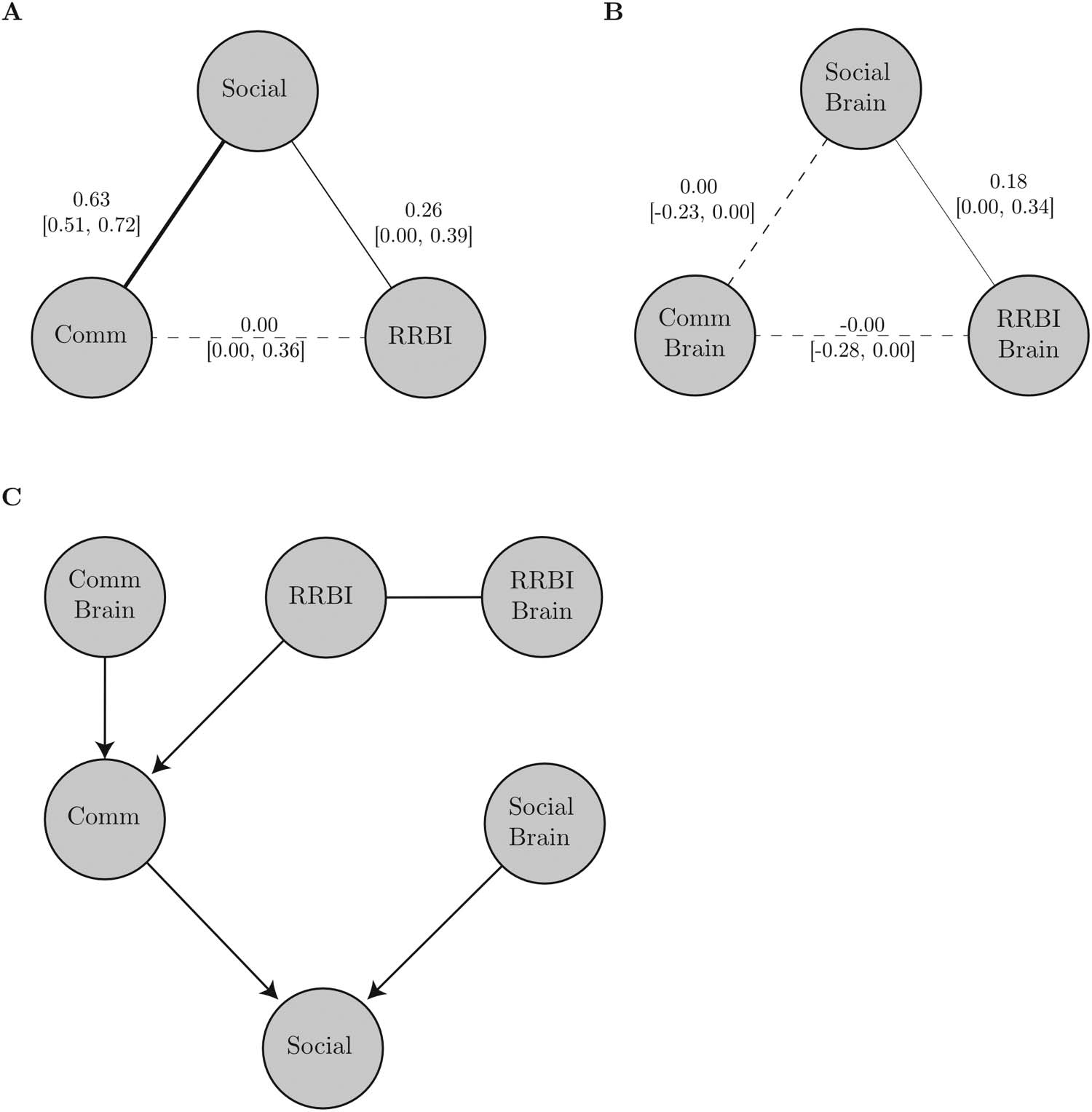
between all measures). As expected from previous studies, the behavioural network showed a
strong association between Communication and Social Scores, and a weaker association
between Social and RRBI scores (see Figure 4A). Im Gegensatz, the network of rsfMRI correlates
showed a strong association between the functional brain correlates of Social and RRBI scores,
but no association between the correlates of Social and Communication scores (see Figure 4B).
Nächste, we conducted a causal inference analysis using Bayesian networks. For the network
of behaviour scores, there were undirected interactions between all behaviour domains, Das
Netzwerkneurowissenschaften
456
Merging network psychometrics and network neuroscience
Tisch 3. Detailed description of edges that were associated with ADI-R scores
Sozial: positive
−10
MNI
48
Harvard-Oxford Label
−22
L Frontal Pole (45%)
28
4
−74
24
−12
−22
R Occipital Fusiform Gyrus (67%)
R Subcallosal Cortex (59%)
MNI
16
−36
16
44
12
44
Harvard-Oxford Label
46
R Middle Frontal Gyrus
(60%)
52
46
R Postcentral Gyrus (34%)
R Middle Frontal Gyrus
(60%)
Sozial: negative
−56
−38
16
L Juxtapositional Lobule Cortex (70%)
44
16
46
R Middle Frontal Gyrus
(60%)
−4
−8
60
L Planum Temporale (48%)
44
16
46
R Middle Frontal Gyrus
(60%)
−4
−38
36
L Cingulate Gyrus posterior division
36
−78
26
R Lateral Occipita (53%)
(81%)
64
−34
22
L Supramarginal Gyrus posterior
44
16
46
R Middle Frontal Gyrus
division (22%)
(60%)
6
0
64
R Juxtapositional Lobule Cortex (58%)
44
16
46
R Middle Frontal Gyrus
Communication:
−18
−64
6
L Intracalcarine Cortex (48%)
−42
positive
−44
−30
18
L Parietal Operculum Cortex (67%)
36
(60%)
8
0
48
L Middle Frontal Gyrus
(53%)
−44
R Temporal Fusiform Cortex
(44%)
−22
−12
68
L Precentral Gyrus (31%)
−42
8
48
L Middle Frontal Gyrus (53%)
4
8
20
54
R Superior Frontal Gyrus
(62%)
−74
8
R Intracalcarine Cortex
(62%)
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
Communication:
18
−98
14
R Occipital Pole (60%)
negative
RRBI: positive
12
−6
−44
−72
−78
−20
26
R Cuneal Cortex (43%)
8
L Intracalcarine Cortex (64%)
54
L Postcentral Gyrus (37%)
50
32
14
−44
8
−70
−20
−74
−44
−20
54
L Postcentral Gyrus (37%)
22
−60
−4
38
54
8
6
R Inferior Frontal Gyrus
(30%)
R Precuneous Cortex (35%)
L Postcentral Gyrus (37%)
R Intracalcarine Cortex
(62%)
R Intracalcarine Cortex
(35%)
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
−34
−52
−22
52
50
−48
0
32
−12
46
L Superior Parietal (38%)
6
L Central Opercular (63%)
42
50
L Superior Frontal (44%)
R Postcentral Gyrus (52%)
6
4
R Central Opercular Cortex (37%)
40
52
14
50
52
4
−12
R Insular Cortex (59%)
−12
24
6
−12
50
60
R Postcentral Gyrus (52%)
R Superior Frontal (61%)
4
R Central Opercular (37%)
50
R Postcentral Gyrus (52%)
Netzwerkneurowissenschaften
457
Merging network psychometrics and network neuroscience
RRBI: negative
MNI
−8
−8
−8
−16
−16
−50
−50
−50
−50
−50
Tisch 3.
(Fortsetzung )
Harvard-Oxford Label
42
L Precentral Gyrus (44%)
42
42
44
44
L Precentral Gyrus (44%)
L Precentral Gyrus (44%)
L Postcentral Gyrus (46%)
L Postcentral Gyrus (46%)
MNI
−66
−46
−4
46
−46
−46
−38
−64
−60
−66
40
36
42
24
40
Harvard-Oxford Label
L Lateral Occipital Cortex
(70%)
L Cingulate Gyrus (81%)
R Lateral Occipital Cortex
(62%)
L Angular Gyrus (50%)
L Lateral Occipital Cortex
(70%)
−48
−28
56
L Postcentral Gyrus (55%)
−46
−66
40
L Lateral Occipital Cortex
(70%)
−44
−20
54
L Postcentral Gyrus (37%)
−46
−66
40
L Lateral Occipital Cortex
(70%)
−8
−42
66
L Postcentral Gyrus (47%)
−54
−54
12
L Middle Temporal Gyrus
(33%)
−52
0
6
L Central Opercular Cortex (63%)
34
−34
62
R Postcentral Gyrus (44%)
6
6
−38
−38
36
36
R Cingulate Gyrus (70%)
R Cingulate Gyrus (70%)
Notiz. The MNI coordinates refer to the centroid of the ROI in the Schaefer parcellation with 300 ROIs at 2-mm resolution. The labels indicate the highest
probability labels in the Harvard-Oxford Cortical Structural atlas. L = left; R = right.
Ist, Sozial, Communication, and RRBI. This structure was indicated across all structure learning
algorithms and statistics for the independence tests. For the network of brain correlates, a struc-
ture with an undirected interaction between rsfMRI correlates of Social scores and RRBI scores
was indicated. There were no connections with the rsfMRI correlates of the Communication
Punktzahl. This structure was consistent across structure learning algorithms and independence test
Statistiken. When combining the behaviour and brain scores in one causal inference network, A
more complex network structure emerged (see Figure 4C). In this network, RRBI behavioural
scores predicted Communication behaviour scores, which in turn predicted Social behaviour
scores. Social and Communication behaviour scores were also predicted by their rsfMRI cor-
relate scores. This structure was consistent across algorithms and independence test statistics.
Tisch 4.
Pearson correlation between behaviour and brain nodes
1. Sozial
2. Comm
3. RRBI
4. Soc Brain
5. Comm Brain
6. RRBI Brain
1
1.00
0.66
0.38
0.49
0.21
0.24
2
3
4
5
6
1.00
0.36
0.09
0.47
0.18
1.00
0.12
−0.12
0.61
1.00
−0.05
0.18
1.00
−0.13
1.00
Notiz. Comm = communication; RRBI = repetitive and/or restricted behaviours and narrow interests; Soc =
sozial.
Netzwerkneurowissenschaften
458
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 4. Overview of the network analysis. Bootstrapped undirected network structure of the behavioural measures (A) and their resting-
state fMRI correlates (B). (A) Bootstrapped undirected network structure. Solid lines indicate significant edges; dashed lines indicate nonsig-
nificant ones. The thickness of the lines indicates the strength of the association. All edges are positive and solid lines indicate significant edges
(bootstrap p value < 0.05). The numbers indicate the bootstrapped median with the 2.5% and 98.5%ile in brackets. (C) Directed acyclic graph
determined through Bayesian network analysis with constrained-based learning algorithms for the combined brain–behaviour network.
Arrows indicated directed interactions. Lines indicate undirected interaction.
DISCUSSION
In this article, we present a novel approach to study the interaction between behavioural var-
iables and their neural correlates. We demonstrate this approach by investigating the relation-
ship between autistic traits and their resting-state functional connectivity correlates identified
through CPM (Shen et al., 2017). Our results indicate that the network of associations between
behavioural measures of autistic traits is not the same as the network of their neural correlates.
We further demonstrate that complex causal interactions become apparent when behavioural
traits and their neural correlates are considered together. This shows that the network-based
regression approach can provide new insights into otherwise inaccessible aspects of brain–
behaviour associations.
To determine if the results obtained through the CPM method with network-based regres-
sors were reasonable, we compared the findings to the results obtained using the traditional
Network Neuroscience
459
Merging network psychometrics and network neuroscience
approach to studying brain–behaviour relationships. Regarding the connections implicated in
each domain of the autistic traits, the current analysis indicated that lower connection strength
of the connections with the middle frontal cortex were associated with higher difficulties
across all autistic trait domains. This result is consistent with other reports that suggest a central
role of frontal networks in autism (Lin et al., 2019; Lynch et al., 2017), possibly due to a shared
influence of executive function difficulties mediated by the frontal cortex on all domains of
autistic traits (Yerys et al., 2019). Furthermore, weaker connections of lateral parietal areas
were associated with greater difficulties across domains. Hypoconnectivity of parietal areas
that form part of the default mode network are among the most consistent findings in autism
neuroimaging research (Padmanabhan et al., 2017). In conclusion, the findings using the CPM
method with network-based regressors are in line with previously implicated neural correlates
of autistic traits.
Critically, our analysis using network-based regressors also identified brain correlates that
were shared between only some of the behavioural traits. Communication and RRBIs were
both associated with greater connectivity of occipital areas. These connectivity differences
may be related to sensory oversensitivity (Green et al., 2016) or increased reliance on visuo-
spatial processing strategies (Kana et al., 2006; Samson et al., 2012). Furthermore, higher
connectivity of left postcentral gyrus was associated with both higher difficulties with social
interaction and RRBIs. Because the network-based regressors only contain the unique variance
in each measure, any remaining overlap hints at a shared mechanism at the brain level.
Our results also indicated unique associations for each domain of autistic traits. Because the
network-based regressors remove the overlap between the behavioural measures, these
findings suggest unique brain-level mechanisms. For instance, higher connectivity of the pre-
central gyrus was only associated with RRBIs but not the other autistic traits. This illustrates the
utility of the network-based regression approach to identify unique mechanisms of behav-
ioural traits.
The results obtained through network-based regression can be further employed to com-
pare the network structure at the behavioural and brain level. This provides further indication
about unique or shared mechanisms at each level of observation. In our autism example, there
was a closer association between social and communication scores than between these scores
and the RRBI score at the behavioural level. This result is consistent with the distinction
between “social” and “nonsocial” traits suggested by some psychometric and genetic studies
(Mandy & Skuse, 2008; Ronald et al., 2005) that is now incorporated in DSM-5. However, a
different structure was indicated for the network of the rsfMRI correlates of the autistic traits. In
the brain score network, correlates of social scores were associated with the correlates of RRBI
scores, but not with the correlates of communication scores. This suggests that the close asso-
ciation of social and communication scores at the behavioural level is not due to a shared
mechanism in brain function. In contrast, the association between the neural correlates of
social and RRBI scores indicates that there is at least some overlap in the brain functional sys-
tems underlying these behaviours. We further explored causal interaction within the networks.
This analysis indicated complex causal interactions between behavioural measures of autistic
traits and their neural correlates that could not be predicted from the network at either the
behavioural or functional brain level alone. This causal network indicated that Communica-
tion scores mediate the association between RRBI and Social scores when accounting for the
overlap in the associated functional brain systems. This shows that the network-based regres-
sion approach opens possibilities for analyses that contrast behavioural and brain-level asso-
ciations to identify potentially shared mechanisms by using the information at the behavioural
level to minimize any overlap in the regressors.
Network Neuroscience
460
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
n
e
n
_
a
_
0
0
2
2
2
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Merging network psychometrics and network neuroscience
To construct a network of neural correlates of behavioural traits, we had to address several
methodological challenges. Namely, in the first step of the analysis, we identified the functional
connectivity correlates of the unique variance in autistic traits in a large, openly accessible data-
base. The principal challenge here is the high dimensionality of the functional connectivity
data. Even with a relatively coarse parcellation of the brain, the number of features by far
exceeds the number of participants. For instance, a parcellation with 100 ROIs produces
4,950 features, assuming that the investigator excluded the diagonal and identical features in
a symmetric adjacency matrix. Several methods have been developed to tackle this problem.
These methods include complex machine learning pipelines, for example, HPC Netmats Meta-
trawls (https://db.humanconnectome.org/megatrawl/index.html), and dimensionality reduction
approaches, such as partial least-squares analysis (Krishnan et al., 2011) or canonical correla-
tion analysis (Zhuang et al., 2020). In the current analysis, we employed the CPM method (Shen
et al., 2017) for its relative simplicity and clear interpretability (Scheinost et al., 2019). This
method identifies functional connectome features that are most closely associated with the
behavioural variable of interest and mitigates overfitting through straightforward cross-
validation. Despite its simplicity, the method has been successfully employed to identify func-
tional connectome correlates in several studies (Beaty et al., 2018; Finn et al., 2015; Rosenberg
et al., 2016, 2018). Even though the modelling approach is relatively simple, the impact of sev-
eral methodological choices still needs to be evaluated. Our results show that the choice of the
functional connectivity metric, the resolution of the parcellation scheme, and the inclusion of
global signal regression strongly impact on the strength of association between the functional
connectivity features and the behavioural measures (see Figure 2). We recommend that
researchers who wish to apply the brain–behaviour network approach evaluate and report
the impact of these methodological choices on the strength of the brain–behaviour association.
The results of the current analysis show that CPM can identify functional connections that
are associated with each facet of the autistic trait triad. The summed brain scores significantly
predicted the behavioural ratings and explained between 20% and 27% of the variance. The
proportion of explained variance is lower than in studies that predicted cognitive task perfor-
mance (Rosenberg et al., 2016) but is similar to the explained variance in a large-scale study
with questionnaire measures (https://db.humanconnectome.org/megatrawl/index.html). Mea-
surement considerations aside, the explained variance in the current analysis may also be
lower due to the variability in the dataset. We utilised data from ABIDE and ABIDE-II, which
are retrospective collections that were acquired without prior harmonisation of the protocol
(Di Martino et al., 2014, 2017). Consequently, there is considerable variation between the
acquisition sites (He et al., 2020). We reduced this variability by applying stringent criteria
for participant inclusion and by regressing the effect of extraneous variables from the connec-
tome edges, but the remaining unaccounted variance is likely to have impacted on the amount
of variance that could be explained in the behavioural measures.
It has to be noted that the network-based regression method has some limitations. First,
relatively large samples are needed to obtain stable estimates of the networks. We used
summed scores in our example to reduce the dimensionality of the behavioural measures.
However, this involves a trade-off. Analyses with more detailed measures show that there is
considerable variation within each domain at the behavioural level. For instance, RRBIs are
dissociable into factors of repetitive behaviours and insistence on sameness (Szatmari et al.,
2006). Summing these dissociable domains may impact on the network structure at the behav-
ioural level and influence subsequent analyses. Researchers will need to decide on the appro-
priate degree of granularity in the behavioural measures when employing the network-based
regression approach for their research questions. Furthermore, the current analysis pipeline
Network Neuroscience
461
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
n
e
n
_
a
_
0
0
2
2
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Merging network psychometrics and network neuroscience
was constructed to maximise the correspondence between resting-state fMRI connectivity
and behavioural scores. Therefore, we selected the parameters that maximised the correla-
tion between the measures and used cross-validation to prevent overfitting. Future researcher
may wish to investigate the impact of the parameter choices, for example, through specifi-
cation curve analysis.
To conclude, we present a novel method to investigate the network structure of behavioural
measures and their neural correlates. This method uses the unique variance int the behavioural
variables to identify neural correlates. Subsequently, the behavioural and neural variables can
be treated as nodes to compare the network structure at the level of behaviour and brain func-
tion. We show how this approach can be used to investigate if similarities at the behavioural
level may be driven by similar mechanisms at the functional brain level. The approach pre-
sented here aligns closely with the shift towards complex system analysis in clinical psychol-
ogy (Borsboom et al., 2018) and expands the current toolkit to levels of analysis beyond
behavioural traits.
AUTHOR CONTRIBUTIONS
Joe Bathelt: Conceptualization; Formal analysis; Methodology; Software; Visualization; Writing –
original draft; Writing – review & editing. Hilde M. Geurts: Conceptualization; Supervision;
Writing – original draft; Writing – review & editing. Denny Borsboom: Conceptualization;
Methodology; Supervision; Writing – original draft; Writing – review & editing.
FUNDING INFORMATION
Joe Bathelt, Amsterdam Brain & Cognition Talent Grant, 2019. Hilde M. Geurts, VICI Grant
from the Netherlands Organisation for Scientific Research, Award ID: 453-16-006. Denny
Borsboom, Consolidator Grant from the European Research Council, Award ID: 647209.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
n
e
n
_
a
_
0
0
2
2
2
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Abbott, A. E., Linke, A. C., Nair, A., Jahedi, A., Alba, L. A., Keown,
C. L., Fishman, I., & Müller, R.-A. (2018). Repetitive behaviors in
autism are linked to imbalance of corticostriatal connectivity: A
functional connectivity MRI study. Social Cognitive and Affective
Neuroscience, 13(1), 32–42. https://doi.org/10.1093/scan
/nsx129, PubMed: 29177509
Abraham, A., Pedregosa, F., Eickenberg, M., Gervais, P., Mueller,
A., Kossaifi, J., Gramfort, A., Thirion, B., & Varoquaux, G.
(2014). Machine learning for neuroimaging with scikit-learn.
Frontiers in Neuroinformatics, 8, 14. https://doi.org/10.3389
/fninf.2014.00014, PubMed: 24600388
Assaf, M., Jagannathan, K., Calhoun, V. D., Miller, L., Stevens,
M. C., Sahl, R., O’Boyle, J. G., Schultz, R. T., & Pearlson,
G. D. (2010). Abnormal functional connectivity of default mode
sub-networks in autism spectrum disorder patients. NeuroImage,
53(1), 247–256. https://doi.org/10.1016/j.neuroimage.2010.05
.067, PubMed: 20621638
Beaty, R. E., Kenett, Y. N., Christensen, A. P., Rosenberg, M. D.,
Benedek, M., Chen, Q., Fink, A., Qiu, J., Kwapil, T. R., Kane,
M. J., & Silvia, P. J. (2018). Robust prediction of individual crea-
tive ability from brain functional connectivity. Proceedings of the
National Academy of Sciences, 115(5), 1087–1092. https://doi
.org/10.1073/pnas.1713532115, PubMed: 29339474
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. NeuroImage, 37(1), 90–101. https://doi.org/10
.1016/j.neuroimage.2007.04.042, PubMed: 17560126
Berument, S. K., Rutter, M., Lord, C., Pickles, A., & Bailey, A.
(1999). Autism screening questionnaire: Diagnostic validity.
British Journal of Psychiatry, 175(5), 444–451. https://doi.org/10
.1192/bjp.175.5.444, PubMed: 10789276
Borsboom, D. (2017). A network theory of mental disorders. World
Psychiatry, 16(1), 5–13. https://doi.org/10.1002/wps.20375,
PubMed: 28127906
Borsboom, D., Cramer, A. O. J., & Kalis, A. (2018). Brain disorders?
Not really: Why network structures block reductionism in psycho-
pathology research. The Behavioral and Brain Sciences, 42, e2.
https://doi.org/10.1017/S0140525X17002266, PubMed: 29361992
Network Neuroscience
462
Merging network psychometrics and network neuroscience
Borsboom, D., Deserno, M. K., Rhemtulla, M., Epskamp, S., Fried,
E. I., McNally, R. J., Robinaugh, D. J., Perugini, M., Dalege, J.,
Costantini, G., Isvoranu, A.-M., Wysocki, A. C., van Borkulo, C. D.,
van Bork, R., & Waldorp, L. J. (2021). Network analysis of multi-
variate data in psychological science. Nature Reviews Methods
Primers, 1(1), 1–18. https://doi.org/10.1038/s43586-021-00055-w
Burgess, G. C., Kandala, S., Nolan, D., Laumann, T. O., Power,
J. D., Adeyemo, B., Harms, M. P., Petersen, S. E., & Barch,
D. M. (2016). Evaluation of denoising strategies to address
motion-correlated artifacts in resting-state functional magnetic
resonance imaging data from the Human Connectome Project.
Brain Connectivity, 6(9), 669–680. https://doi.org/10.1089/brain
.2016.0435, PubMed: 27571276
Cicchetti, D. V., Lord, C., Koenig, K., Klin, A., & Volkmar, F. R.
(2008). Reliability of the ADI-R: Multiple examiners evaluate a
single case. Journal of Autism and Developmental Disorders,
38(4), 764–770. https://doi.org/10.1007/s10803-007-0448-3,
PubMed: 18058216
Constantino, J. N., Gruber, C. P., Davis, S., Hayes, S., Passanante,
N., & Przybeck, T. (2004). The factor structure of autistic traits.
Journal of Child Psychology and Psychiatry, and Allied Disci-
plines, 45(4), 719–726. https://doi.org/10.1111/j.1469-7610
.2004.00266.x, PubMed: 15056304
Cox, R. W., & Jesmanowicz, A. (1999). Real-time 3D image regis-
tration for functional MRI. Magnetic Resonance in Medicine,
42(6), 1014–1018. https://doi.org/10.1002/(SICI)1522-2594
(199912)42:6<1014::AID-MRM4>3.0.CO;2-F
Craddock, C., Sikka, S., Cheung, B., Khanuja, R., Ghosh, S. S., Yan,
C., … Milham, M. (2013). Towards automated analysis of con-
nectomes: The configurable pipeline for the analysis of connec-
tomes (C-PAC). Frontiers in Neuroinformatics, 42, 10–3389.
https://www.frontiersin.org/10.3389/conf.fninf.2013.09.00042
/event_abstract
Di Martino, A., O’Connor, D., Chen, B., Alaerts, K., Anderson, J. S.,
Assaf, M., Balsters, J. H., Baxter, L., Beggiato, A., Bernaerts, S.,
Blanken, L. M. E., Bookheimer, S. Y., Braden, B. B., Byrge, L.,
Castellanos, F. X., Dapretto, M., Delorme, R., Fair, D. A.,
Fishman, ICH., … Milham, M. P. (2017). Enhancing studies of the
connectome in autism using the autism brain imaging data
exchange II. Wissenschaftliche Daten, 4, 170010. https://doi.org/10.1038
/sdata.2017.10, PubMed: 28291247
Di Martino, A., Ross, K., Uddin, L. Q., Sklar, A. B., Castellanos,
F. X., & Milham, M. P. (2009). Functional brain correlates of
social and nonsocial processes in autism spectrum disorders:
An activation likelihood estimation meta-analysis. Biologisch
Psychiatrie, 65(1), 63–74. https://doi.org/10.1016/j.biopsych
.2008.09.022, PubMed: 18996505
Di Martino, A., Yan, C.-G., Li, Q., Denio, E., Castellanos, F. X.,
Alaerts, K., Anderson, J. S., Assaf, M., Bookheimer, S. Y.,
Dapretto, M., Deen, B., Delmonte, S., Dinstein, ICH., Ertl-Wagner,
B., Fair, D. A., Gallagher, L., Kennedy, D. P., Keown, C. L.,
Keyser, C., … Milham, M. P. (2014). The autism brain imaging
data exchange: Towards a large-scale evaluation of the intrinsic
brain architecture in autism. Molekulare Psychiatrie, 19(6), 659.
https://doi.org/10.1038/mp.2013.78, PubMed: 23774715
DiLalla, D. L., & Rogers, S. J. (1994). Domains of the childhood
autism rating scale: Relevance for diagnosis and treatment.
Journal of Autism and Developmental Disorders, 24(2), 115–128.
https://doi.org/10.1007/bf02172092, PubMed: 8040157
Dupong, ICH., & Di Martino, A. (2020). Hyper-connectivity of the stri-
atum related to restricted and repetitive behaviors’ severity in
children with ASD. BioRxiv, 2020.02.21.957993. https://doi.org
/10.1101/2020.02.21.957993
Esteban, O., Birman, D., Schaer, M., Koyejo, Ö. O., Poldrack, R. A.,
& Gorgolewski, K. J. (2017). MRIQC: Advancing the automatic
prediction of image quality in MRI from unseen sites. PLoS
Eins, 12(9), e0184661. https://doi.org/10.1371/journal.pone
.0184661, PubMed: 28945803
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang, J.,
Chun, M. M., Papademetris, X., & Polizist, R. T. (2015). Func-
tional connectome fingerprinting: Identifying individuals using
patterns of brain connectivity. Naturneurowissenschaften, 18(11),
1664–1671. https://doi.org/10.1038/nn.4135, PubMed: 26457551
Friston, K. J., Williams, S., Howard, R., Frackowiak, R. S. J., &
Turner, R. (1996). Movement-Related effects in fMRI time-series.
Magnetic Resonance in Medicine, 35(3), 346–355. https://doi.org
/10.1002/mrm.1910350312, PubMed: 8699946
Grün, S. A., Hernandez, L., Bookheimer, S. Y., & Dapretto, M.
(2016). Salience network connectivity in autism is related to
brain and behavioral markers of sensory overresponsivity. Zeitschrift
of the American Academy of Child and Adolescent Psychiatry,
55(7), 618–626. https://doi.org/10.1016/j.jaac.2016.04.013,
PubMed: 27343889
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain image
alignment using boundary-based registration. NeuroImage, 48,
63–72. https://doi.org/10.1016/j.neuroimage.2009.06.060,
PubMed: 19573611
Hagen, E. A. H. von D., Stoyanova, R. S., Baron-Cohen, S., &
Calder, A. J. (2013). Reduced functional connectivity within
and between ‘social’ resting state networks in autism spectrum
Bedingungen. Social Cognitive and Affective Neuroscience, 8(6),
694–701. https://doi.org/10.1093/scan/nss053, PubMed:
22563003
Happé, F., & Ronald, A. (2008). The “fractionable autism triad”: A
review of evidence from behavioural, genetic, cognitive and
neural research. Neuropsychology Review, 18(4), 287–304.
https://doi.org/10.1007/s11065-008-9076-8, PubMed: 18956240
Harris, G. J., Chabris, C. F., Clark, J., Urban, T., Aharon, ICH., Steele,
S., McGrath, L., Condouris, K., & Tager-Flusberg, H. (2006).
Brain activation during semantic processing in autism spectrum
disorders via functional magnetic resonance imaging. Brain and
Cognition, 61(1), 54–68. https://doi.org/10.1016/j.bandc.2005
.12.015, PubMed: 16473449
Er, Y., Byrge, L., & Kennedy, D. P. (2020). Nonreplication of func-
tional connectivity differences in autism spectrum disorder
across multiple sites and denoising strategies. Human Brain Map-
ping, 41(5), 1334–1350. https://doi.org/10.1002/ hbm.24879,
PubMed: 31916675
Jung, M., Tu, Y., Lang, C. A., Ortiz, A., Park, J., Jorgenson, K., Kong,
X.-J., & Kong, J. (2017). Decreased structural connectivity and
resting-state brain activity in the lateral occipital cortex is associ-
ated with social communication deficits in boys with autism
spectrum disorder. NeuroImage, 190, 205–212. https://doi.org
/10.1016/j.neuroimage.2017.09.031, PubMed: 28927730
Netzwerkneurowissenschaften
463
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
Nur, M. A., Cherkassky, V. L., Keller, T. A., & Minshew, N. J. (2004).
Cortical activation and synchronization during sentence compre-
hension in high-functioning autism: Evidence of underconnectiv-
ität. Gehirn: A Journal of Neurology, 127(8), 1811–1821. https://doi
.org/10.1093/brain/awh199, PubMed: 15215213
Kadosh, K. C., Kadosh, R. C., Dick, F., & Johnson, M. H. (2010).
Developmental changes in effective connectivity in the emerging
core face network. Hirnrinde, 21(6), 1389–1394. https://doi
.org/10.1093/cercor/bhq215, PubMed: 21045001
Kana, R. K., Keller, T. A., Cherkassky, V. L., Minshew, N. J., & Nur,
M. A. (2006). Sentence comprehension in autism: Thinking in
pictures with decreased functional connectivity. Gehirn: A Journal
of Neurology, 129(Pt 9), 2484–2493. https://doi.org/10.1093
/brain/awl164, PubMed: 16835247
Kana, R. K., & Wadsworth, H. M. (2012). “The archeologist’s career
ended in ruins”: Hemispheric differences in pun comprehension
in autism. NeuroImage, 62(1), 77–86. https://doi.org/10.1016/j
.neuroimage.2012.04.034, PubMed: 22548805
Kim, H., Keifer, C. M., Rodriguez-Seijas, C., Eaton, N. R., Lerner,
M. D., & Gadow, K. D. (2018). Structural hierarchy of autism
spectrum disorder symptoms: An integrative framework. Zeitschrift
of Child Psychology and Psychiatry, and Allied Disciplines,
59(1), 30–38. https://doi.org/10.1111/jcpp.12698, PubMed:
28195316
Kim, H., Keifer, C., Rodriguez-Seijas, C., Eaton, N., Lerner, M., &
Gadow, K. (2019). Quantifying the optimal structure of the
autism phenotype: A comprehensive comparison of dimensional,
categorical, and hybrid models. Journal of the American Acad-
emy of Child and Adolescent Psychiatry, 58(9), 876–886.
https://doi.org/10.1016/j.jaac.2018.09.431, PubMed: 30768420
Krishnan, A., Williams, L. J., McIntosh, A. R., & Abdi, H. (2011).
Partial least squares (PLS) methods for neuroimaging: A tutorial
and review. NeuroImage, 56(2), 455–475. https://doi.org/10
.1016/j.neuroimage.2010.07.034, PubMed: 20656037
Kroeze, R., van der Veen, D. C., Servaas, M. N., Bastiaansen, J. A.,
Oude Voshaar, R. C., Borsboom, D., Ruhe, H. G., Schoevers,
R. A., & Riese, H. (2017). Personalized feedback on symptom
dynamics of psychopathology: A proof-of-principle study. Zeitschrift
for Person-Oriented Research, 3(1), 1–10. https://doi.org/10
.17505/jpor.2017.01, PubMed: 33569120
Lee, J. M., Kyeong, S., Kim, E., & Cheon, K.-A. (2016). Abnormal-
ities of inter- and intra-hemispheric functional connectivity in
autism spectrum disorders: A study using the Autism Brain
Imaging Data Exchange database. Frontiers in Neuroscience,
10, 191. https://doi.org/10.3389/fnins.2016.00191, PubMed:
27199653
Lee, Y., Park, B.-Y., James, O., Kim, S.-G., & Park, H. (2017). Autism
spectrum disorder related functional connectivity changes in the
language network in children, adolescents and adults. Frontiers
in Human Neuroscience, 11, 418. https://doi.org/10.3389
/fnhum.2017.00418, PubMed: 28867997
Lin, H.-Y., Perry, A., Cocchi, L., Roberts, J. A., Tseng, W.-Y. ICH.,
Breakspear, M., & Gau, S. S.-F. (2019). Development of fronto-
parietal connectivity predicts longitudinal symptom changes in
young people with autism spectrum disorder. Translational Psy-
chiatry, 9(1), 86. https://doi.org/10.1038/s41398-019-0418-5,
PubMed: 30755585
Lynch, C. J., Breeden, A. L., You, X., Ludlum, R., Gaillard, W. D.,
Kenworthy, L., & Vaidya, C. J. (2017). Executive dysfunction in
autism spectrum disorder is associated with a failure to modulate
frontoparietal-insular hub architecture. Biologische Psychiatrie:
Cognitive Neuroscience and Neuroimaging, 2(6), 537–545.
https://doi.org/10.1016/j.bpsc.2017.03.008, PubMed: 29348041
Mandy, W. P. L., & Skuse, D. H. (2008). Research review: What is
the association between the social-communication element of
autism and repetitive interests, behaviours and activities? Zeitschrift
of Child Psychology and Psychiatry, and Allied Disciplines, 49(8),
795–808. https://doi.org/10.1111/j.1469-7610.2008.01911.x,
PubMed: 18564070
Margaritis, D. (2003). Learning Bayesian network model structure
aus Daten. Pittsburgh, PA: Carnegie-Mellon Univerity School
of Computer Science. https://apps.dtic.mil/sti/citations
/ADA461103
Maximo, J. O., & Kana, R. K. (2019). Aberrant “deep connectivity”
in autism: A cortico–subcortical functional connectivity magnetic
resonance imaging study. Autism Research, 12(3), 384–400.
https://doi.org/10.1002/aur.2058, PubMed: 30624021
Mazefsky, C. A., & Oswald, D. P. (2006). The discriminative ability
and diagnostic utility of the ADOS-G, ADI-R, and GARS for chil-
dren in a clinical setting. Autism: The International Journal of
Research and Practice, 10(6), 533–549. https://doi.org/10.1177
/1362361306068505, PubMed: 17088271
Müller, R.-A., & Fishman, ICH. (2018). Brain connectivity and neuro-
imaging of social networks in autism. Trends in Cognitive Sci-
zen, 22(12), 1103–1116. https://doi.org/10.1016/j.tics.2018
.09.008, PubMed: 30391214
Padmanabhan, A., Lynch, C. J., Schaer, M., & Menon, V. (2017).
The default mode network in autism. Biologische Psychiatrie: Cog-
nitive Neuroscience and Neuroimaging, 2(6), 476–486. https://
doi.org/10.1016/j.bpsc.2017.04.004, PubMed: 29034353
Patriquin, M. A., DeRamus, T., Libero, L. E., Laird, A., & Kana, R. K.
(2016). Neuroanatomical and neurofunctional markers of social
cognition in autism spectrum disorder. Kartierung des menschlichen Gehirns,
37(11), 3957–3978. https://doi.org/10.1002/ hbm.23288,
PubMed: 27329401
Pearl, J. (2014). Probabilistic reasoning in intelligent systems: Net-
works of plausible inference. Sonst.
Poldrack, R. A., & Yarkoni, T. (2016). From brain maps to cognitive
ontologies: Informatics and the search for mental structure.
Annual Review of Psychology, 67, 587–612. https://doi.org/10
.1146/annurev-psych-122414-033729, PubMed: 26393866
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., &
Petersen, S. E. (2012). Spurious but systematic correlations in
functional connectivity MRI networks arise from subject motion.
NeuroImage, 59(3), 2142–2154. https://doi.org/10.1016/j
.neuroimage.2011.10.018, PubMed: 22019881
Raamana, P. R. (2020). Conquering confounds and covariates in
machine learning with the python library confounds. Zenodo.
https://doi.org/10.5281/zenodo.3701528
Robinaugh, D. J., Hoekstra, R. H. A., Toner, E. R., & Borsboom, D.
(2020). The network approach to psychopathology: Eine Rezension von
the literature 2008–2018 and an agenda for future research. Psy-
chological Medicine, 50(3), 353–366. https://doi.org/10.1017
/S0033291719003404, PubMed: 31875792
Netzwerkneurowissenschaften
464
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
Ronald, A., Happé, F., & Plomin, R. (2005). The genetic relationship
between individual differences in social and nonsocial behav-
iours characteristic of autism. Developmental Science, 8(5),
444–458. https://doi.org/10.1111/j.1467-7687.2005.00433.x,
PubMed: 16048517
Rosenberg, M. D., Finn, E. S., Scheinost, D., Papademetris, X.,
Shen, X., Polizist, R. T., & Chun, M. M. (2016). A neuromarker
of sustained attention from whole-brain functional connectivity.
Naturneurowissenschaften, 19(1), 165–171. https://doi.org/10.1038/nn
.4179, PubMed: 26595653
Rosenberg, M. D., Hsu, W.-T., Scheinost, D., Polizist, R. T., &
Chun, M. M. (2018). Connectome-based models predict separa-
ble components of attention in novel individuals. Journal of Cog-
nitive Neuroscience, 30(2), 160–173. https://doi.org/10.1162
/jocn_a_01197, PubMed: 29040013
Rutter, M., Le Couteur, A., & Lord, C. (2003). Autism diagnostic
interview-revised. Western Psychological Services, 29(2003), 30.
Samson, F., Mottron, L., Soulières, ICH., & Zeffiro, T. A. (2012).
Enhanced visual functioning in autism: An ALE meta-analysis.
Kartierung des menschlichen Gehirns, 33(7), 1553–1581. https://doi.org/10
.1002/hbm.21307, PubMed: 21465627
Schäfer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, A. J., Eickhoff, S. B., & Yeo, B. T. T. (2017). Local-global
parcellation of the human cerebral cortex from intrinsic func-
tional connectivity MRI. BioRxiv, 135632. https://doi.org/10
.1101/135632
Scheinost, D., Noble, S., Horien, C., Greene, A. S., Lake, E. M. R.,
Salehi, M., Gao, S., Shen, X., O’Connor, D., Barron, D. S., Yip,
S. W., Rosenberg, M. D., & Polizist, R. T. (2019). Ten simple
rules for predictive modeling of individual differences in neuro-
Bildgebung. NeuroImage, 193, 35–45. https://doi.org/10.1016/j
.neuroimage.2019.02.057, PubMed: 30831310
Schurz, M., Radua, J., Aichhorn, M., Richlan, F., & Perner, J. (2014).
Fractionating theory of mind: A meta-analysis of functional brain
Bildgebungsstudien. Neuroscience and Biobehavioral Reviews, 42,
9–34. https://doi.org/10.1016/j.neubiorev.2014.01.009,
PubMed: 24486722
Scutari, M. (2009). Learning Bayesian networks with the bnlearn R
Package [Computer software]. 0908.3817.
Shen, X., Finn, E. S., Scheinost, D., Rosenberg, M. D., Chun, M. M.,
P a p a d e m e t r i s , X . , & C o n s t a b l e , R . T.
( 2 0 1 7 ) . U s i n g
connectome-based predictive modeling to predict individual behav-
ior from brain connectivity. Nature Protocols, 12(3), 506–518.
https://doi.org/10.1038/nprot.2016.178, PubMed: 28182017
Schmied, S. M. (2002). Fast robust automated brain extraction. Human
Brain Mapping, 17(3), 143–155. https://doi.org/10.1002/ hbm
.10062, PubMed: 12391568
Schmied, S. M., Jenkinson, M., Woolrich, M. W., Beckmann, C. F.,
Behrens, T. E. J., Johansen-Berg, H., Bannister, P. R., Luca,
M. D., Drobnjak, ICH., Flitney, D. E., Niazy, R. K., Saunders, J.,
Vickers, J., Zhang, Y., Stefano, N. D., Brady, J. M., & Matthews,
P. M. (2004). Advances in functional and structural MR image
analysis and implementation as FSL. NeuroImage, 23, S208–S219.
https://doi.org/10.1016/j.neuroimage.2004.07.051, PubMed:
15501092
Supekar, K., Uddin, L. Q., Khouzam, A., Phillips, J., Gaillard, W. D.,
Kenworthy, L. E., Yerys, B. E., Vaidya, C. J., & Menon, V. (2013).
Brain hyperconnectivity in children with autism and its links to
social deficits. Cell Reports, 5(3), 738–747. https://doi.org/10
.1016/j.celrep.2013.10.001, PubMed: 24210821
Szatmari, P., Georgiades, S., Bryson, S., Zwaigenbaum, L., Roberts,
W., Mahoney, W., Goldberg, J., & Tuff, L. (2006). Investigating
the structure of the restricted, repetitive behaviours and interests
domain of autism. Journal of Child Psychology and Psychiatry,
and Allied Disciplines, 47(6), 582–590. https://doi.org/10.1111/j
.1469-7610.2005.01537.X, PubMed: 16712635
Szatmari, P., Mérette, C., Bryson, S. E., Thivierge, J., Roy, M.-A.,
Cayer, M., & Maziade, M. (2002). Quantifying dimensions in
autism: A factor-analytic study. Journal of the American Academy
of Child and Adolescent Psychiatry, 41(4), 467–474. https://doi
.org/10.1 097 /000 04 58 3-20 02 04 000 -00 020 , Pub Med :
11931604
Tsamardinos, ICH., Aliferis, C. F., Statnikov, A. R., & Statnikov, E.
(2003). Algorithms for large scale Markov blanket discovery.
FLAIRS Conference, 2, 376–380. https://www.aaai.org/ Papers
/FLAIRS/2003/Flairs03-073.pdf
Tsamardinos, ICH., Braun, L. E., & Aliferis, C. F. (2006). The max-min
hill-climbing Bayesian network structure learning algorithm.
Machine Learning, 65(1), 31–78. https://doi.org/10.1007
/s10994-006-6889-7
Tustison, N. J., Cook, P. A., Klein, A., Song, G., Das, S. R., Duda,
J. T., Kandel, B. M., van Strien, N., Stein, J. R., Gee, J. C., &
Avants, B. B. (2014). Large-scale evaluation of ANTs and Free-
Surfer cortical thickness measurements. NeuroImage, 99, 166–179.
https://doi.org/10.1016/j.neuroimage.2014.05.044, PubMed:
24879923
Uddin, L. Q., Supekar, K., Lynch, C. J., Khouzam, A., Phillips, J.,
Feinstein, C., Ryali, S., & Menon, V. (2013). Salience network–
based classification and prediction of symptom severity in chil-
dren with autism. JAMA Psychiatry, 70(8), 869–879. https://doi
.org/10.1001/jamapsychiatry.2013.104, PubMed: 23803651
van Lang, N. D. J., Boomsma, A., Sytema, S., de Bildt, A. A., Kraijer,
D. W., Ketelaars, C., & Minderaa, R. B. (2006). Structural equa-
tion analysis of a hypothesised symptom model in the autism
spectrum. Journal of Child Psychology and Psychiatry, and Allied
Disciplines, 47(1), 37–44. https://doi.org/10.1111/j.1469-7610
.2005.01434.X, PubMed: 16405639
Varoquaux, G., Baronnet, F., Kleinschmidt, A., Fillard, P., & Thirion,
B. (2010). Detection of brain functional-connectivity difference
in post-stroke patients using group-level covariance modeling.
In T. Jiang, N. Navab, J. P. W. Pluim, & M. A. Viergever (Hrsg.),
Medical image computing and computer-assisted intervention –
MICCAI 2010. MICCAI 2010. Lecture Notes in Computer Science
(S. 200–208, Bd. 6361). Berlin, Deutschland: Springer. https://doi
.org/10.1007/978-3-642-15705-9_25, PubMed: 20879232
Wadden, N. P. K., Bryson, S. E., & Rodger, R. S. (1991). A closer look
at the Autism Behavior Checklist: Discriminant validity and factor
Struktur. Journal of Autism and Developmental Disorders, 21(4),
529–541. https://doi.org/10.1007/ bf02206875, PubMed:
1778965
Wing, L., & Gould, J. (1979). Severe impairments of social interac-
tion and associated abnormalities in children: Epidemiology and
classification. Journal of Autism and Developmental Disorders,
9(1), 11–29. https://doi.org/10.1007/bf01531288, PubMed: 155684
Netzwerkneurowissenschaften
465
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
T
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Merging network psychometrics and network neuroscience
Yaramakala, S., & Margaritis, D. (2005). Speculative Markov blan-
ket discovery for optimal feature selection. Fifth IEEE Interna-
tional Conference on Data Mining (ICDM’05), 4 S. https://doi
.org/10.1109/ICDM.2005.134
Yerys, B. E., Tunç, B., Satterthwaite, T. D., Antezana, L., Mosner,
M. G., Bertollo, J. R., Guy, L., Schultz, R. T., & Herrington,
J. D. (2019). Functional connectivity of fronto-parietal and
salience/ventral attention networks have independent
associations with co-occurring ADHD symptoms in children
with autism. Biologische Psychiatrie: Cognitive Neuroscience and
Neuroimaging, 4(4), 343–351. https://doi.org/10.1016/j.bpsc
.2018.12.012, PubMed: 30777604
Zhuang, X., Yang, Z., & Cordes, D. (2020). A technical review of
canonical correlation analysis for neuroscience applications.
Kartierung des menschlichen Gehirns, 41(13), 3807–3833. https://doi.org/10
.1002/hbm.25090, PubMed: 32592530
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
4
5
2
0
2
8
1
5
6
N
e
N
_
A
_
0
0
2
2
2
P
D
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Netzwerkneurowissenschaften
466