Individual Differences in Resting-state Brain Rhythms
Uniquely Predict Second Language Learning Rate
and Willingness to Communicate in Adults
Chantel S. Prat, Brianna L. Yamasaki, and Erica R. Peterson
Abstrakt
■ The current study used quantitative electroencephalography
(qEEG) to characterize individual differences in neural rhythms
at rest and to relate them to fluid reasoning ability, to first lan-
guage proficiency, and to subsequent second language (L2)
learning ability, with the goal of obtaining a better understand-
ing of the neurocognitive bases of L2 aptitude. Mean spectral
power, laterality, and coherence metrics were extracted across
theta, alpha, beta, and gamma frequency bands obtained from
eyes-closed resting-state qEEG data from 41 adults aged 18–
34 Jahre. Participants then completed 8 weeks of French train-
ing using a virtual language and cultural immersion software.
Results replicate and extend previous studies showing that faster
learners have higher beta power recorded over right hemi-
Kugel (RH) electrode sites, greater laterality (RH − LH/RH +
LH) of alpha and beta bands, and greater coherence between
RH frontotemporal sites across all frequencies, although only
coherence measures survived multiple comparisons. Increased
coherence within and between RH networks was also asso-
ciated with greater posttest declarative memory scores and with
more accurate speech during learning. Total speech attempts, In
Kontrast, correlated with bilaterally distributed small-world net-
work configurations, as indexed by lower power and coherence
over high-frequency (beta and gamma) bands recorded over
frontotemporal networks in both hemispheres. Results from
partial correlations and regression analyses suggest that the
neural predictors of L2 learning rate, posttest proficiency, Und
total speech attempts varied in their degree of overlap with
qEEG correlates of first language proficiency and fluid reason-
ing abilities, but that neural predictors alone explained 26–60%
of the variance in L2 outcomes. ■
EINFÜHRUNG
The concept of a “talent” for language learning has cap-
tivated researchers, educators, and laypersons alike for
centuries (z.B., Handschin & Henmon, 1930; Russell,
1858). Perhaps this is because language is one of the
quintessential human abilities, or perhaps it is because
many of us have attempted to learn a second language
(L2) and failed. Differences in the ease with which lan-
guages can be acquired across the lifespan have been
the focus of extensive research on learning and neural
plasticity (z.B., Birdsong, 1999; Snow & Hoefnagel-Höhle,
1978). Despite the fact that acquiring an L2 in adulthood
is difficult, large individual differences, typically referred
to as L2 aptitude, existieren (z.B., Dörnyei, 2009), and re-
searchers from a variety of fields have attempted to mea-
sure and understand these differences with incremental
success (see Wen, Biedroń, & Skehan, 2017, für eine Rezension).
Theoretical perspectives of L2 aptitude have histori-
cally progressed iteratively with the development of be-
havioral batteries for assessing it (see Wen et al., 2017;
Dörnyei & Skehan, 2003, for reviews) and by extension
to theories of L2 learning. One central research question
Universität Washington
© 2018 Massachusetts Institute of Technology
that has been addressed through such psychometric
investigations is whether L2 aptitude consists of one or
many aptitudes. Zum Beispiel, the foundational work of
Carroll (z.B., Dörnyei, 2009; Carroll & Sapon, 1959)
resulted in a four-factor model of L2 aptitude, welche
included phonemic coding ability, grammatical sensitivity,
inductive language learning ability, and associative mem-
ory strength. A related theoretical question is understand-
ing the extent to which L2 aptitude is specific to language
learning or reflects a more general ability to learn complex
skills or acquire knowledge (z.B., Gardner & Lambert, 1965).
Relatively recently, neuroscientists have “thrown their
hat into the ring” in attempts to answer these questions.
The majority of this research has measured neural changes
associated with different levels of L2 proficiency (Chee,
Soon, Lee, & Pallier, 2004; Mechelli et al., 2004; Perani
et al., 1998), and a subset of studies have investigated
neural changes longitudinally as a function of increasing
exposure to an L2 (see Mamiya, Richards, Coe, Eichler, &
Kuhl, 2016; Li, Legault, & Litcofsky, 2014, für eine Rezension).
Jedoch, very few studies have related individual dif-
ferences in brain functioning to language aptitude or to
the ability to acquire an L2 in the future, adopting, in effect,
a neuropsychometric approach (Chai et al., 2016; Prat,
Yamasaki, Kluender, & Stocco, 2016; Qi, Han, Garel, San
Zeitschrift für kognitive Neurowissenschaften 31:1, S. 78–94
doi:10.1162/jocn_a_01337
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Chen, & Gabriela, 2015; Tan et al., 2011). Wichtig, zwei
of these existing studies (Chai et al., 2016; Prat et al., 2016)
have measured properties of brain functioning at “rest,”
and a third focused on static properties of white matter
Struktur (Qi et al., 2015). Such research is critical, as it allows
one to partially disentangle any intrinsic properties of brain
functioning associated with facile learning from differences
that result from the way an individual’s brain might process
or respond to linguistic stimuli.
To understand the neuropsychometric properties of
resting-state data obtained from quantitative electro-
encephalography (qEEG) Maßnahmen, the current study
employs an individual differences approach (Prat, 2011),
relating variability in network-level brain functioning at
rest and a battery of cognitive variables to the subsequent
ease (or difficulty) with which one acquires an L2 in adult-
hood. The goals of this approach are (1) to better under-
stand the neurocognitive basis of L2 acquisition and (2) Zu
characterize the patterns of brain functioning associated
with “good” L2 learning. Although these goals are inter-
related, their implications are largely separate. Der erste
goal is centered on the belief that comprehensive theo-
ries of L2 acquisition should be able to account for indi-
vidual differences in brain functioning and their behavioral
consequences (z.B., Prat, 2011). The second provides a
template for what optimal functioning looks like, welche
can be used to target brain-based interventions (sehen
Hanslmayr, Sauseng, Doppelmayr, Schabus, & Klimesch,
2005; Klimesch, Sauseng, & Gerloff, 2003, for examples of
this approach within intelligence research).
In the first exploration of resting-state qEEG predictors
of L2 aptitude, Prat and colleagues (2016) showed that
the power in beta and gamma frequency bands of neural
oscillations recorded primarily over right hemisphere
(RH) electrode sites was the strongest predictor of sub-
sequent L2 learning rates. Based on the correspondence
between these findings and what is known about the role
of the RH and first language (L1) proficiency, Prat et al.
(2016) proposed that these results may reflect the known
relation between L1 proficiency and L2 aptitude. Specifi-
cally, according to the RH dynamic spillover hypothesis,
monolingual individuals who are more proficient in their
native language have more specialized patterns of brain
Aktivierung, which are more highly left-lateralized, Ergebnis-
ing in recruitment of fewer RH resources for language
processes (Prat, 2011; Prat, Mason, & Nur, 2011). Es ist
worth noting, Jedoch, that laterality differences in qEEG
recorded over the scalp do not necessarily translate to
ipsilateral cortical sources.
Prat and colleagues (2016) also found that individual
differences in laterality of alpha power predicted L2 learn-
ing rate. Based on the correspondence between this re-
sult and previous research linking resting-state qEEG to
fluid intelligence (Thatcher, Norden, & Biver, 2005), Sie
proposed that this finding may highlight a more general
role of fluid cognitive abilities in L2 acquisition (Sasaki,
1996; Gardner & Lambert, 1965).
The goal of the current study was to extend these pre-
vious results and our knowledge of the neurocognitive
basis of L2 aptitude more broadly by exploring the de-
gree to which qEEG indicators of L2 aptitude reflect
individual differences in linguistic specific abilities (als
measured by L1 proficiency) versus more general cor-
relates of fluid reasoning (as measured by the Raven’s
Advanced Progressive Matrices [RAPM]). Zusätzlich,
we explored multiple facets of L2 learning to look for
convergence and divergence of neural predictors. In einem
recent review of the neural basis of language aptitude,
Biedron (2015) states that “One of the most important
obstacles in examining foreign language aptitude is the
heterogeneity of the construct” (P. 15). By leveraging
what is known about the neural basis of learning, Flüssigkeit
reasoning, and individual differences in L1 proficiency,
the current experiment investigated neural oscillations,
properties of intrinsic brain functioning that are known
to relate to subsequent cognitive performance (sehen
Klimesch, 1999, für eine Rezension). Speziell, we used met-
rics obtained through qEEG recorded while participants
sat with their eyes closed to predict subsequent L2 learn-
ing ability.
The current study employed the Operational Language
and Culture Training System (OLCTS), a virtual immer-
sion language training software that provides simulta-
neous training in language and sociocultural norms
( Johnson, Friedland, Watson, & Surface, 2012). This soft-
ware, which has been used to investigate L2 aptitude
(Prat et al., 2016), was designed to enable military per-
sonnel to function safely and effectively in foreign coun-
tries by attaining functional language proficiency in very
short time periods. The training is organized hierarchi-
cally around goal-based scenarios (z.B., obtaining transpor-
Station). It requires the learner to interact with intelligent
avatars by speaking and by choosing socially relevant be-
haviors such as eye contact (or lack thereof ), interruption
of ongoing speech, and formality of intonation.
The current investigation extends the previous work
by Prat and colleagues (2016) in several important ways.
Erste, qEEG data were analyzed using a network-level ap-
proach, which assessed not only mean power and later-
ality in functional networks known to correlate with
language learning (Kepinska, Pereda, Caspers, & Schiller,
2017) but also spectral coherence within and between
these networks. This allows us to make further infer-
ences about the mechanisms driving differences in power
and laterality. Zweite, total speech attempts and speak-
ing accuracy during L2 learning were added to L2 learn-
ing rate and posttest declarative memory as outcome
Maßnahmen. This allows us to assess whether procedural
(speaking accuracy) and declarative (translational
accuracy) outcome variables are explained by the same
neural metrics. Endlich, the current study involves a much
larger sample size than the original, allowing us to examine
which of the effects originally measured on 16 Teilnehmer
replicates in a group of 41 Teilnehmer.
Prat, Yamasaki, and Peterson
79
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
METHODEN
Teilnehmer
Forty-seven healthy adults aged 18–34 years (33 Frauen)1
were initially recruited for participation in this study. Von
diese, data were analyzed from the 41 participants who
completed the 8-week French training program. Partial
data from 16 of these participants linking qEEG power
to L2 learning rate was previously reported in a brief
communication article (Prat et al., 2016). All participants
were monolingual English speakers with no previous expo-
sure to French or to any L2 before the age of 10 and re-
ported low proficiency to any L2 exposed to at any point.
All participants gave informed consent according to the
standards set forth by the University of Washington insti-
tutional review board and were paid for their participation.
Materials
Behavioral Assessments of Cognitive Abilities
A battery of standardized or commonly used tests of fluid
information processing (Raven’s Advanced Progressive
Matrices, Arthur & Day, 1994; Reading Span and Operation
Span, Unsworth, Heitz, Schrock, & Engle, 2005), executive
functioning (Simon Task, Stocco et al., 2017; 3-back updat-
ing-Aufgabe, Kane, Conway, Miura, & Colflesh, 2007; color–
shape-switching task, Monsell, 2003; Continuous Performance
Test, Cohen, Barch, Fuhrmann, & Servan-Schreiber, 1999;
Attentional Blink task, Raymond, Shapiro, & Arnell, 1992;
Broadbent & Broadbent, 1987), probabilistic learning
(Probabilistic Stimulus Selection task, Frank, Seeberger,
& O’Reilly, 2004), and English reading ability (Nelson–
Denny Reading Test [NDRT]; Braun, Fishco, & Hanna,
1993) were used to assess cognitive abilities behaviorally.
These tasks are described in more detail in Prat et al.
(2016) and in the supplementary material.2
Language Training Software
The OLCTS ( Johnson et al., 2012), a virtual immersion
language training program, was used to provide partici-
pants with 8 weeks of French language training. Der
OLCTS provides immersive, goal-directed cultural and
language training, including comprehension and produc-
tion components (using speech recognition software), als
well as sociocultural practices and pragmatic instruction.
These skills are applied through goal-directed scenarios
involving interactions with avatars and then tested explic-
itly through quizzes that incorporate all aspects of linguistic
proficiency (written comprehension, verbal production,
and auditory comprehension). The OLCTS is organized
by “units” or “levels” that terminate with these quizzes.
Participants were not allowed to advance until they ob-
tained a minimum accuracy of 50% on each quiz. Bei der
end of each 30-min training session, the final level was
recorded. A regression line was fit to terminal-level data
at each session to calculate a rate of L2 learning. For each
participant, the OLCTS records the total number of speech
productions and number of “correct” speech attempts. A
speech attempt is considered correct when the utterance
is recognized, with the highest likelihood, from a classifica-
tion model that uses utterances of both native speakers
and of learners in the “correct” model and also includes a
“garbage” parameter. Speaking accuracy was determined
by taking the ratio of correct speech attempts to total
speech attempts as measured by the software.
The French–English Translation Posttest of
French Proficiency
Long-term declarative memory for items studied was
assessed using a French–English translation test, welche
was created individually for each participant based on
the materials they encountered over the 8-week session
of language training. The test required participants to
select the correct English translation for 50 French words
or phrases they learned during training. The number of
items drawn from each level was proportionate to the
amount of material covered on that level.
Procedures
Participant Training and Testing
All participants initially completed two 1.5-hr testing
sessions in which all behavioral measures were collected.
Zusätzlich, 5 min of eyes-closed resting-state EEG were
collected at 128 Hz using wireless, EPOC 16-channel
Emotiv headsets, with reference channels set at DMS
and CRL electrodes over the parietal lobe. The presen-
tation order of the behavioral tests varied across partici-
Hose, but EEG was always collected at the beginning of
one of the behavioral sessions. Following these two ses-
sionen, all participants received sixteen 30-min language train-
ing sessions using the OLCTS over the course of 8 weeks.
qEEG Analysis
Initial EEG data were analyzed using the procedures out-
lined in the preliminary report (Prat et al., 2016), welche
were slightly modified from Doppelmayr, Klimesch, Stadler,
Pöllhuber, and Heine (2002). Zusamenfassend, Die 5 min of resting-
state data were segmented into 2-sec epochs with 50%
overlap and cleaned for blinks, Bewegung, and impedance
artifacts, and the artifact-free data were analyzed using
fast Fourier transform to measure average power in theta
(4–7.5 Hz), alpha (8–12.5 Hz), beta (13–29.5 Hz), Und
low-gamma (30–40 Hz) frequency bands for each indi-
viduell. Any channel that did not have at least 75 artifact-free
epochs was not included in the data analysis, welche re-
sulted in the removal of 13 Kanäle (2.2% of the data),
which were treated as missing in subsequent analyses.
Spectral coherence was calculated for all pairwise combi-
nations of electrodes. As in other publications (z.B.,
Sauseng, Klimesch, Schabus, & Doppelmayr, 2005),
80
Zeitschrift für kognitive Neurowissenschaften
Volumen 31, Nummer 1
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
coherence Cx,j( F ) between two channels x and y at a
frequency f was computed according to the formula:
Cx;y fð Þ ¼
Px;y fð Þ2
Px fð Þ (cid:2) Py fð Þ
where the Px,j( F ) is the cross-power density and the
Px( F ) and Py( F ) are the power densities of x and y,
jeweils. The calculations were carried out using the
default spectral analysis function in the R statistical soft-
ware (R Core Team, 2013).
To reduce the dimensionality of this qEEG data in a the-
oretically driven way, we then collapsed the 14 qEEG
channels into networks based on recent data obtained
from phase synchronization during an artificial grammar
learning task (Kepinska et al., 2017). The resulting five
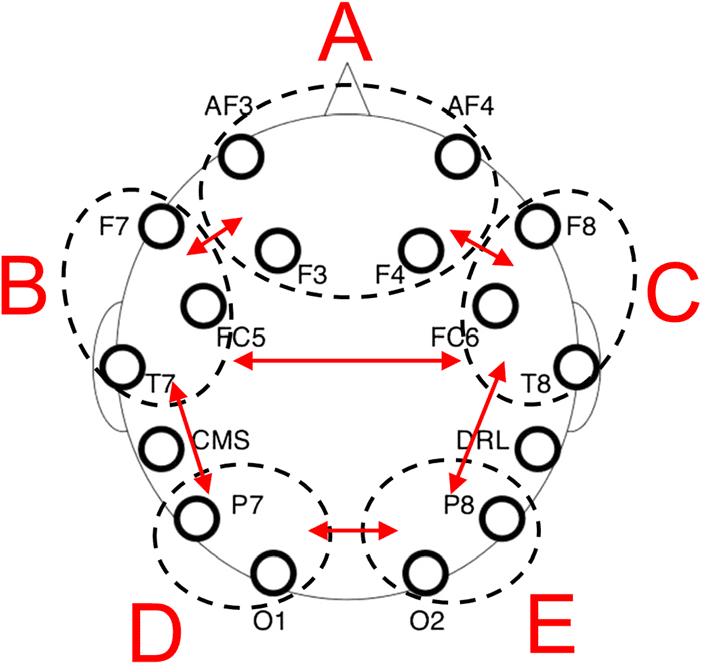
Netzwerke, as depicted in Figure 1, war (A) medial frontal
consisting of AF3, AF4, F3, and F4 electrodes; (B) LH
frontotemporal consisting of F7, FC5, and T7 electrodes;
(C) RH frontotemporal consisting of F8, FC6, and T8
Elektroden; (D) LH posterior consisting of P7 and O1 elec-
trodes; Und (E) RH posterior consisting of P8 and O2 elec-
trodes. Mean power across each of the four frequency
bands was computed over each network by averaging
the results of the fast Fourier transform analysis on indi-
vidual channels within each network. From these data,
two laterality indices were then calculated for each of the
four frequency bands by subtracting mean power in each
frequency band in the LH network from the mean power
in the corresponding RH network and dividing by the
average of the two values (z.B., C − B/Average(C, B)).
Mean within- and between-network spectral coherence
analyses were then computed by averaging coherence
within networks and between network pairs, as indicated
by red arrows in Figure 1.
Figur 1. Schematic depiction of network-level qEEG data analysis
conducted based on Kepinska et al. (2017). Letters correspond to
networks labeled herein: (A) medial-frontal, (B) LH frontotemporal,
(C) RH frontotemporal, (D) LH posterior, Und (E) RH posterior. Red
arrows indicate between-network coherence analyses that were
conducted as reported in Table 1.
Group-level network coherence analysis supported our
choice of networks, as all within network coherence values
were significantly greater than between network values
(siehe Tabelle 1). False discovery rates (FDRs) were also calcu-
lated and reported for each of the analyses (Benjamini &
Hochberg, 1995).
ERGEBNISSE
Individual Differences in Indicators of
L2 Learning
Individual differences in the ability to acquire an L2 in
adulthood were apparent in each of the four metrics of
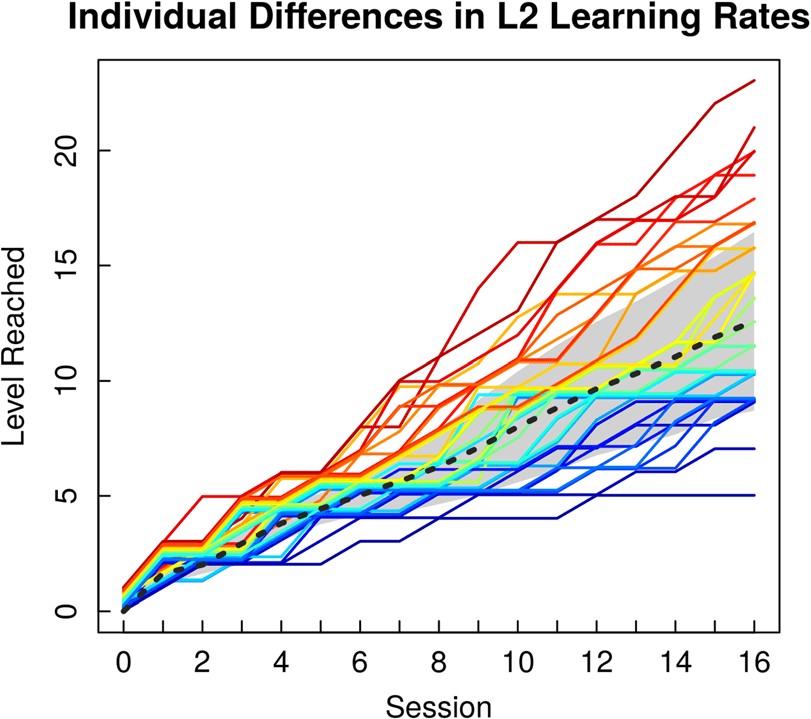
L2 learning acquired. With respect to L2 learning rate,
the fastest learner progressed three and a half times as
quickly through the training program as the slowest
learner did (mean = 0.80, range = 0.38–1.32, SD =
0.22). Individual learning rates are depicted in Figure 2.
A similar amount of variability was observed for propor-
tion of correct speech attempts (mean = 0.24, range =
0.13–0.33, SD = .05), with even greater variability ob-
served for total speech attempts (mean = 2025, range =
280–3586, SD = 825) across the 8-week session. In con-
trast, performance on the declarative memory posttest
was much less variable (mean = 91.7%, range = 70–100%,
SD = 7.86%).
Correlations between the four indicators of L2 learn-
ing suggest that they were not completely independent.
Speziell, accuracy on the declarative memory post-
test was positively correlated with rate of L2 learning,
rs(41) = .54, p = .001, demonstrating that the fastest
learners also retained the most information following
Ausbildung. A positive correlation between total speech
attempts and proportion of correct speech attempts,
rs(41) = .44, p = .004, was also observed, showing that
willingness to communicate, as indexed by total speech
attempts, was positively correlated with accuracy of
Rede, as indexed by the percentage of these utterances
recognized as correct by the software.
Cognitive Predictors of L2 Learning
L2 Learning Rate
As in our previous experiment, none of the cognitive tests
correlated significantly with L2 learning rate ( ps > .05).
Declarative Memory Posttest
Performance on the declarative memory posttest was posi-
tively correlated with L1 proficiency, as measured by the
NDRT, rs(41) = .45, p = .001. None of the other cognitive
measures reliably correlated with declarative memory post-
test scores.
Total Speech Attempts
Total speech attempts were negatively correlated with
the attentional blink size, rs(41) = −.40, p = .010, solch
Prat, Yamasaki, and Peterson
81
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Tisch 1. Mean Group-level Within and Between-network Coherence Values (SEM in Parentheses) with Paired Sample t Test
Comparison Values Demonstrating that All Within-network Coherence Values Were Significantly Greater than All Between-network
Coherence Values
Within-network Coherence
Between-network Coherence
t Test
A) Medial frontal
A) Medial frontal
B) LH frontotemporal
B) LH frontotemporal
B) LH frontotemporal
C) RH frontotemporal
C) RH frontotemporal
C) RH frontotemporal
D) LH posterior
D) LH posterior
E) RH posterior
E) RH posterior
.755 (.006)
.755 (.006)
.625 (.008)
.625 (.008)
.625 (.008)
.710 (.012)
.710 (.012)
.710 (.012)
.607 (.008)
.607 (.008)
.742 (.007)
.742 (.007)
A-B
A-C
B-A
B-D
B-C
C-A
C-E
C-D
D-B
D-E
E-C
E-D
.613 (.005)
.651 (.006)
.613 (.005)
.525 (.002)
.565 (.005)
.651 (.006)
.619 (.006)
.565 (.005)
.525 (.002)
.545 (.004)
.619 (.006)
.545 (.004)
T(40) = 26.8, P < .001
t(40) = 16.5, p < .001
t(40) = 2.7, p = .009
t(40) = 12.5, p < .001
t(40) = 10.3, p < .001
t(40) = 7.5, p < .001
t(40) = 15.6, p < .001
t(40) = 9.6, p < .001
t(37) = 9.2, p < .001
t(37) = 7.5, p < .001
t(36) = 18.0, p < .001
t(36) = 26.6, p < .001
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
that people who spoke aloud more frequently had reliably
smaller attentional blinks. Total speech attempts were
also positively correlated with working memory updating
accuracy as measured by total accuracy, rs(41) = .34,
p = .031, and distractor accuracy, rs(41) = .40, p = .010,
on the 3-back updating task.
Relating Individual Differences in Resting-state
qEEG Power to L2 Aptitude Variables
Fluid Reasoning Ability
Performance on the abbreviated RAPM did not correlate
with mean power averaged across any of the five
networks.
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
Proportion of Accurate Speech
None of the cognitive tests correlated significantly with
proportion of accurate speech ( ps > .05), although verbal
working memory as measured by the reading span task
approached significance, rs(41) = .29, p = .067.
Figur 2. L2 learning rates for individual participants plotted as a
function of level reached at the end of each 30-min training session (mean
level depicted by dashed line, standard deviation of the mean in gray).
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
L1 Proficiency
Performance on the NDRT was correlated with alpha
power in the RH in both frontotemporal, rs(37) = .33,
p = .049, and posterior, rs(37) = .34, p = .039, Netzwerke
as well as in the LH posterior network, rs(38) = .33,
p = .046. L1 proficiency was also correlated with mean
theta power averaged over bilateral frontal, rs(40) =
.36, p = .023, and RH posterior, rs(37 ) = .36, p =
.031, Netzwerke.
Relating Individual Differences in Resting-state
qEEG Power to L2 Learning
L2 Learning Rate
Consistent with previous research (Prat et al., 2016), mean
power in the beta frequency range recorded over the RH
predicted subsequent L2 learning rate. Speziell, mean
beta power over the RH posterior cluster was significantly
positively correlated with L2 learning rate, rs(37) = .39,
p = .019, and the correlation between mean beta power
over the RH frontotemporal electrode cluster and L2
learning rate approached significance, rs(37) = .31, p =
.063. Both correlations retained or reached significance
when either L1 proficiency or fluid intelligence were
partialed out ( ps < .025).
82
Journal of Cognitive Neuroscience
Volume 31, Number 1
Declarative Memory Posttest
Performance on the declarative memory posttest did
not correlate with qEEG power at rest ( ps > .10). Das
finding also replicates previous research (Prat et al.,
2016).
Total Speech Attempts
Total number of speech attempts was negatively correlated
with mean beta power recorded over both left fronto-
zeitlich, rs(40) = −.38, p = .016, and right fronto-
zeitlich, rs(37) = −.41, p = .013, sites and approached
significance over the medial frontal network, rs(40) =
−.31, p = .054. Number of speech attempts was also
negatively correlated with mean gamma power recorded
over RH frontotemporal, rs(37) = −.41, p = .012, Und
hintere, rs(37) = −.48, p = .003, Netzwerke. Diese
negative correlations retained or reached significance
when either L1 proficiency or fluid intelligence were
partialed out ( ps < .05).
Proportion of Accurate Speech
Proportion of accurate speech attempts did not correlate
with qEEG power at rest, although a negative correlation
between mean beta power in the left frontotemporal
network and accurate speech approached significance,
rs(40) = −.28, p = .077.
None of the reported correlations between L2 variables
and resting-state qEEG power remained significant when
FDR corrections for multiple comparisons were applied.
All bivariate correlations between the four OLCTS out-
come measures and power in the five resting-state net-
works are listed in Table 2.
Relating Individual Differences in Resting-state
qEEG Laterality to L2 Aptitude Variables
Fluid Reasoning Ability
Performance on the RAPM did not correlate with fronto-
temporal or posterior laterality in any power band.
L1 Proficiency
NDRT scores were positively correlated with fronto-
temporal laterality across theta, rs(37) = .36, p = .030,
and alpha, rs(37) = .34, p = .042, frequency bands.
rate was positively correlated with alpha laterality in both
frontotemporal, rs(37) = .44, p = .006, and posterior,
rs(35) = .40, p = .016, networks, whereas learning rate
only correlated with laterality of beta power in posterior
regions, rs(35) = .34, p = .046. When controlling for
L1 proficiency, only frontotemporal alpha laterality
remained significant ( p = .017), although the resulting
correlation with posterior alpha laterality, rs(32) = .30,
p = .084, was not significantly different than the original
correlation ( p = .65). Correlations with posterior beta
laterality and L2 learning rate also decreased when L1
proficiency was partialed out, rs(32) = .16, p = .381,
although the correlations were still not significantly dif-
ferent (correlation difference: p = .48). Similarly, when
controlling for fluid intelligence, only frontotemporal al-
pha laterality remained significant, rs(34) = .40, p = .017,
although the resulting correlation with posterior alpha
laterality, rs(32) = .33, p = .055, was not significantly
different than the original correlation ( p = .76). Cor-
relations with posterior beta laterality and L2 learning
rate also decreased when fluid intelligence was partialed
out, rs(32) = .17, p = .327, although the correlations
were still not significantly different ( p = .48).
Declarative Memory Posttest
Declarative memory posttest scores were not correlated
with resting-state qEEG laterality ( ps > .10).
Total Speech Attempts
Total speech attempts did not correlate with any index of
power laterality at rest ( ps > .17).
Proportion of Accurate Speech
Proportion of accurate speech did not correlate with
resting-state laterality; Jedoch, the correlation with laterality
in frontotemporal beta power approached significance,
rs(37) = .31, p = .067.
None of the reported correlations between L2 variables
and resting-state qEEG laterality remained significant
when FDR corrections for multiple comparisons were
applied. All bivariate correlations between the four
OLCTS outcome measures and laterality indices are listed
in Table 3.
Relating Individual Differences in Resting-state
qEEG Laterality to L2 Learning
Relating Individual Differences in Resting-state
qEEG Coherence to L2 Aptitude Variables
L2 Learning Rate
Consistent with previous research (Prat et al., 2016), L2
learning rate was positively correlated with laterality in
alpha and beta frequency bands. Speziell, L2 learning
Fluid Reasoning Ability
Performance on the RAPM was positively correlated with
coherence within the RH posterior network across theta
frequencies, rs(37) = .40, p = .014.
Prat, Yamasaki, and Peterson
83
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Tisch 2. All Bivariate Correlations (Spearman’s Rho) between Mean Power Obtained from Resting-state qEEG Networks and L2
Outcome Variables
Netzwerk
L2 Learning Rate
Posttest Scores
Speech Attempts
Correct Speech
.158 (40)
.049 (40)
.252 (40)
.179 (40)
.014 (40)
.018 (40)
.021 (40)
−.024 (40)
.286 (37)
.227 (37)
.308 (37)
.161 (37)
.235 (38)
.062 (38)
.132 (38)
.029 (38)
.286 (37)
.154 (37)
.385 (37)*
.129 (37)
A. Medial Frontal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
B. Left Frontotemporal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
C. Right Frontotemporal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
D. Left Posterior
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
E. Right Posterior
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
ns are in parentheses.
*P < .05. **p < .01. L1 Proficiency .180 (40) −.005 (40) .066 (40) −.023 (40) −.090 (40) −.033 (40) .013 (40) −.005 (40) .154 (37) .166 (37) .088 (37) −.116 (37) .170 (38) .089 (38) .086 (38) .064 (38) .173 (37) .100 (37) .179 (37) .093 (37) .107 (40) −.047 (40) −.307 (40) −.304 (40) −.259 (40) −.213 (40) −.379 (40)* −.250 (40) −.115 (37) −.104 (37) −.406 (37)* −.410 (37)* .121 (38) .116 (38) −.260 (38) −.205 (38) .028 (37) .028 (37) −.288 (37) −.481 (37)** .259 (40) .084 (40) −.027 (40) .040 (40) −.152 (40) −.092 (40) −.283 (40) −.124 (40) .138 (37) .110 (37) −.010 (37) −.122 (37) .280 (38) .275 (38) .061 (38) .017 (38) .216 (37) .226 (37) −.011 (37) −.216 (37) l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / / 3 1 1 7 8 1 7 8 8 1 2 6 / j o c n _ a _ 0 1 3 3 7 p d . f b y g u e s t t o n 0 8 S e p e m b e r 2 0 2 3 Performance on the NDRT was positively correlated with coherence within medial frontal network over theta, rs(41) = .32, p = .045, and alpha, rs(41) = .31, p = .048, frequencies and negatively correlated with coherence between left and right posterior networks over the theta frequency band, rs(41) = −.33, p = .035. Relating Individual Differences in Resting-state qEEG Coherence to L2 Learning L2 Learning Rate Rate of L2 learning was positively correlated with mean coherence within the right frontotemporal network across all power bands, rs(41) = .35, p = .026. This was driven by 84 Journal of Cognitive Neuroscience Volume 31, Number 1 Table 3. All Bivariate Correlations (Spearman’s Rho) Between Laterality Indices Obtained from Mean Power Averaged across Resting-state qEEG Networks and L2 Outcome Variables L2 Learning Rate Posttest Scores Speech Attempts Correct Speech Frontal theta (4–7.5 Hz) alpha (8–12.5 Hz) beta (13–29.5 Hz) gamma (30–40 Hz) Posterior theta (4–7.5 Hz) alpha (8–12.5 Hz) beta (13–29.5 Hz) gamma (30–40 Hz) ns are in parentheses. *p < .05. **p < .01. .242 (37) .440 (37)** .119 (37) .053 (37) .208 (35) .404 (35)* .339 (35)* .014 (35) .219 (37) .245 (37) −.029 (37) −.250 (37) −.044 (35) .139 (35) .107 (35) −.042 (35) .227 (37) .184 (37) .126 (37) −.082 (37) −.058 (35) −.215 (35) −.107 (35) −.204 (35) .243 (37) .211 (37) .305 (37) .077 (37) −.051 (35) −.061 (35) −.012 (35) −.129 (35) l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / / 3 1 1 7 8 1 7 8 8 1 2 6 / j o c n _ a _ 0 1 3 3 7 p d . f b y g u e s t t o n 0 8 S e p e m b e r 2 0 2 3 strong positive correlations between learning rate and right frontotemporal coherence in theta, rs(41) = .46, p = .003, and alpha, rs(41) = .38, p = .014, frequency ranges. Conversely, rate of L2 learning was negatively correlated with coherence within the left posterior network across all power bands, rs(38) = −.39, p = .016. In particular, coherence across alpha, rs(38) = −.35, p = .029, and beta, rs(38) = −.36, p = .025, frequency bands was significantly negatively related to L2 learn- ing rate. These correlations remained significant when controlling for L1 proficiency and fluid intelligence ( ps < .05). Declarative Memory Posttest Similar to rate of L2 learning, accuracy on the declarative memory posttest was positively correlated with mean coherence within the right frontotemporal network across all power bands, rs(41) = .48, p = .001. This was reflected by strong positive correlations between post- test score and right frontotemporal coherence in theta, rs(41) = .54, p < .001; alpha, rs(41) = .50, p = .001; and beta, rs(41) = .40, p = .010, frequency ranges. In addition, mean coherence between right frontotemporal and right posterior regions across all power bands significantly pre- dicted declarative memory posttest scores, rs(41) = .37, p = .016. This was driven by correlations over theta, rs(41) = .38, p = .013, and alpha, rs(41) = .39, p = .013, frequency ranges. Coherence over the theta frequency band between right frontotemporal and medial–frontal networks was also positively correlated with posttest performance, rs(41) = .35, p = .027. When controlling for L1 proficiency or fluid reasoning abilities, the cor- relations remained significant ( ps < .03). Total Speech Attempts Total speech attempts were reliably negatively correlated with gamma coherence within the left frontotemporal network, rs(41) = −.49, p = .001. On the contrary, mean coherence across frequencies in the left posterior net- work was positively correlated with total speech attempts, rs(38) = .33, p = .045, driven primarily by strong positive correlations with alpha coherence, rs(38) = .46, p = .004, and more moderate correlations with beta coherence, rs(38) = .34, p = .035. When controlling for L1 proficiency, these correlations remained marginally significant ( ps < .053). When controlling for fluid intelligence, the correla- tion with left frontotemporal gamma coherence remained highly significant ( p = .001), whereas the correlation with left posterior coherence across all channels dropped below significance, rs(35) = .31, p = .060, but was not significantly lower than the original ( p = .93). Proportion of Accurate Speech Proportion of accurate speech was also correlated with higher coherence in RH networks, averaged across all power bands between RH frontotemporal and posterior networks, rs(41) = .32, p = .041. Although this trend was observed for all frequency bands below gamma ( ps < .10), it was significant only over the beta frequency band, Prat, Yamasaki, and Peterson 85 rs(41) = .34, p = .029, when ranges were considered sep- arately. These correlations dropped below significance when either L1 proficiency or fluid reasoning abilities were factored out ( ps < .10), but the resulting correla- tions were not significantly different than the originals ( ps > .70).
Positive correlations between declarative memory and
right frontotemporal coherence and negative correlations
between total speech attempts and gamma coherence
between left frontotemporal networks remained signifi-
cant after FDR corrections for multiple comparisons were
applied. All correlations between the four OLCTS out-
come measures and coherence metrics are listed in
Tisch 4.
Regression Analyses
L2 Learning Rate
When the six predictors of L2 learning rate (right poste-
rior mean beta power, frontotemporal and posterior
alpha laterality, posterior beta laterality, mean within
right frontotemporal coherence across all bands, Und
mean within left posterior coherence across all bands)
were entered into a simultaneous regression analysis,
the model was significant, F(6, 25) = 3.89, p = .007,
and explained 48% of the variance in L2 learning rate,
with a Bayesian Information Criteria (BIC) von 98.35.
Only right posterior beta power (b = .33, t = 2.17,
p = .04) and frontotemporal alpha laterality (b = .36,
t = 2.12, p = .044) were significant predictors. By com-
parison, when L1 language proficiency and fluid reason-
ing were entered along with the qEEG variables into
the model, the significance was approximately the same,
F(8, 23) = 3.21, p = .013, and explained 53% of the
variance in L2 learning rate, with a BIC of 102.37. Rechts
posterior beta power remained the strongest predictor,
although it was not significant in the full model (b =
.29, t = 1.83, p = .08). Both models showed very strong
evidence of fitting the data better than the null model
(BIC = 122.77; model probabilities = 99.99% for both
the EEG-only and EEG plus L1 proficiency and fluid rea-
soning model). The probability that the EEG-only model
explained more variance in L2 learning rate than the EEG
plus behavioral model was 88.20%. Jedoch, given that
this represents only “positive” evidence for a difference,
the data suggest that the two models fit the data similarly
well ( Wagenmakers, 2007; Raftery, 1995).
Declarative Memory Posttest
model was significant, F(3, 37) = 4.39, p = .010, Und
explained 26% of the variance, with a BIC of 121.42.
Jedoch, only RH frontotemporal to posterior coher-
ence approached significance (b = .335, t = 1.83, p =
.076) as a predictor. By comparison, when L1 profi-
ciency and fluid reasoning were entered along with the
three qEEG variables into the model, the significance
was increased, F(5, 35) = 6.37, P < .001, and explained
48% of the variance in L2 learning rate, with a BIC
of 114.80. Only L1 proficiency explained significant vari-
ance in this model (b = .45, t = 3.59, p = .001). When
compared with a null model (BIC = 122.77), the only
model to demonstrate strong evidence for a better fit to
the data than the null model was the EEG plus L1 pro-
ficiency and fluid reasoning model (model probability =
98.17%).
Total Speech Attempts
When the six predictors of total speech attempts (mean
left and right frontotemporal beta power, mean right
frontotemporal and posterior gamma power, mean within
left frontotemporal gamma coherence, and mean within
left posterior coherence across all bands) were entered
into a simultaneous regression analysis, the model was
highly significant, F(6, 25) = 6.34, p < .001, and ex-
plained 60% of the variance, with a BIC of 520.73.
Mean within left posterior coherence across all fre-
quency bands (b = .41, t = 2.88, p = .008) and mean
right posterior gamma power (b = −.53, t = −2.52, p =
.018) were significant predictors. By comparison, when
L1 proficiency and fluid intelligence were entered along
with the six qEEG variables into the model, the model
remained highly significant, F(8, 23) = 5.35, p = .001,
and explained 65% of the variance, with a BIC of
523.62. Again, however, only mean within left posterior
coherence across all frequencies (b = .36, t = 2.55, p =
.018) and mean right posterior gamma power (b =
−.47, t = −2.22, p = .036) remained significant when
the variables competed for variance. There was very
strong evidence that both the EEG-only and the EEG
plus L1 proficiency and fluid reasoning models ex-
plained significantly more variance in total speech at-
tempts than the null model (BIC = 673.45; model
probabilities = 100.00% for both the EEG-only and the
EEG, L1 proficiency, and fluid reasoning model). Again,
there was only positive evidence to suggest that the
EEG-only model fit the data better than the EEG plus
behavioral model (model probability = 80.97%), which
suggests that both models fit the data similarly well.
When the three predictors of declarative posttest scores
(mean within right frontotemporal network and between
right frontotemporal and posterior network coherence
across all frequency ranges, and mean between right
frontotemporal and medial frontal theta coherence) were
entered into a simultaneous regression analysis, the
Proportion of Correct Speech
Because there was only one significant correlation be-
tween proportion of correct speech and resting-state
qEEG, we did not run regression analyses on this out-
come variable.
86
Journal of Cognitive Neuroscience
Volume 31, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
j
o
c
n
_
a
_
0
1
3
3
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 4. All Bivariate Correlations (Spearman’s Rho) between Network Coherence Indices and L2 Outcome Variables
Network Coherence
L2 Learning Rate
Posttest Scores
Speech Attempts
Correct Speech
Within Medial Frontal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
Within Left Frontotemporal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
Within Left Posterior
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
Within Right Frontotemporal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
Within Right Posterior
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
−.026 (41)
.032 (41)
−.107 (41)
−.108 (41)
−.035 (41)
.213 (41)
.078 (41)
.046 (41)
−.033 (41)
.066 (41)
−.238 (38)
−.354 (38)*
−.363 (38)*
−.188 (38)
−.388 (38)*
.458 (41)**
.380 (41)*
.261 (41)
.248 (41)
.348 (41)*
.034 (37)
.155 (37)
.132 (37)
.131 (37)
.153 (37)
Left Frontotemporal to Right Frontotemporal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
.063 (41)
−.029 (41)
−.005 (41)
−.046 (41)
.026 (41)
gamma (30–40 Hz)
All
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
j
o
c
n
_
a
_
0
1
3
3
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
−.019 (41)
.001 (41)
.010 (41)
−.005 (41)
.021 (41)
.242 (41)
.091 (41)
.003 (41)
−.154 (41)
.094 (41)
.008 (38)
−.070 (38)
−.061 (38)
.103 (38)
−.107 (38)
.542 (41)**
.504 (41)**
.398 (41)**
.261 (41)
.484 (41)**
.123 (37)
.272 (37)
.287 (37)
.261 (37)
.298 (37)
.117 (41)
.081 (41)
.139 (41)
.144 (41)
.182 (41)
−.064 (41)
−.100 (41)
−.044 (41)
−.145 (41)
−.083 (41)
−.122 (41)
−.180 (41)
−.243 (41)
−.492 (41)**
−.230 (41)
.229 (38)
.455 (38)**
.342 (38)*
−.066 (38)
.327 (38)*
−.003 (41)
−.092 (41)
−.049 (41)
−.240 (41)
−.066 (41)
.019 (37)
.269 (37)
−.062 (37)
−.314 (37)
−.087 (37)
−.018 (41)
−.119 (41)
−.083 (41)
−.302 (41)
−.102 (41)
−.271 (41)
−.103 (41)
−.166 (41)
−.236 (41)
−.236 (41)
−.196 (41)
−.076 (41)
−.145 (41)
−.284 (41)
−.142 (41)
.159 (38)
.251 (38)
.090 (38)
.001 (38)
.120 (38)
.098 (41)
.113 (41)
.125 (41)
−.137 (41)
.114 (41)
.206 (37)
.097 (37)
.031 (37)
−.046 (37)
.037 (37)
.064 (41)
.089 (41)
−.034 (41)
−.088 (41)
.048 (41)
Prat, Yamasaki, and Peterson
87
Table 4. (continued )
Network Coherence
L2 Learning Rate
Posttest Scores
Speech Attempts
Correct Speech
Left Posterior to Right Posterior
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
−.091 (41)
−.154 (41)
−.128 (41)
−.129 (41)
−.108 (41)
Left Frontotemporal to Medial Frontal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
.046 (41)
−.074 (41)
−.130 (41)
−.150 (41)
−.050 (41)
Right Frontotemporal to Medial Frontal
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
.231 (41)
.057 (41)
.090 (41)
.084 (41)
All
.073 (41)
Left Frontotemporal to Left Posterior
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
All
−.040 (41)
−.183 (41)
−.230 (41)
−.195 (41)
−.220 (41)
Right Frontotemporal to Right Posterior
theta (4–7.5 Hz)
alpha (8–12.5 Hz)
beta (13–29.5 Hz)
gamma (30–40 Hz)
.263 (41)
.292 (41)
.077 (41)
.051 (41)
All
.187 (41)
ns are in parentheses.
*p < .05.
**p < .01.
−.082 (41)
−.074 (41)
−.033 (41)
.023 (41)
−.022 (41)
.076 (41)
.010 (41)
.011 (41)
−.014 (41)
.017 (41)
.345 (41)*
.264 (41)
.253 (41)
.286 (41)
.262 (41)
−.072 (41)
−.016 (41)
−.104 (41)
−.155 (41)
−.096 (41)
.384 (41)*
.386 (41)*
.278 (41)
.170 (41)
.374 (41)*
−.090 (41)
.162 (41)
.035 (41)
−.116 (41)
−.008 (41)
−.011 (41)
−.140 (41)
−.167 (41)
−.271 (41)
−.181 (41)
−.149 (41)
−.172 (41)
−.230 (41)
−.163 (41)
−.219 (41)
.041 (41)
.086 (41)
.037 (41)
−.128 (41)
.076 (41)
.069 (41)
−.059 (41)
.074 (41)
−.117 (41)
.013 (41)
−.078 (41)
.064 (41)
−.185 (41)
−.202 (41)
−.126 (41)
−.041 (41)
.077 (41)
−.107 (41)
−.237 (41)
−.095 (41)
−.061 (41)
.079 (41)
−.088 (41)
.073 (41)
−.077 (41)
.122 (41)
.124 (41)
−.016 (41)
−.098 (41)
.089 (41)
.290 (41)
.273 (41)
.342 (41)*
.133 (41)
.320 (41)*
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
j
o
c
n
_
a
_
0
1
3
3
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
88
Journal of Cognitive Neuroscience
Volume 31, Number 1
DISCUSSION
The results reported herein provide extended evidence
that characteristics of intrinsic brain functioning, as
measured by resting-state qEEG, can be used to predict
individual differences in subsequent L2 acquisition in
adulthood. Results from the current experiment repli-
cated and extended our original report (Prat et al.,
2016) by demonstrating again that mean beta power re-
corded over RH regions and laterality of frontotemporal
alpha and posterior beta power (as indexed by RH −
LH/average of the two) were associated with faster L2
learning. The current experiment provided further evi-
dence about the mechanisms underpinning these differ-
ences by showing that learning rate, declarative knowledge,
and speaking accuracy were also correlated with greater
coherence within and between RH networks. With respect
to learning rate and memory performance, this increased
coherence was most prevalent in theta and alpha fre-
quency ranges, which have been widely demonstrated to
reflect memory and general cognitive processes (Klimesch,
1999). Using a neuropsychometric approach, the results
of the current study also showed that combined neural
metrics explained 48% of the variance in subsequent L2
learning rate, which is comparable to that explained by
standardized behavioral measures such as the MLAT
(Carroll, 1958). In addition, the current results provide
neural evidence supporting the hypothesis that L2 apti-
tude is a multifaceted construct, involving a combination
of general and language-specific abilities. These results
are linked to their theoretical implications in greater
detail in subsequent sections.
Relating qEEG Predictors of L2 Learning to L1
Proficiency and Fluid Reasoning Abilities
One of the motivations of the current experiment was to
explore the overlap between neural predictors of L2 learn-
ing and neural correlates of L1 proficiency. Some overlap
between these two variables was observed, particularly
with respect to alpha and theta laterality recorded over
the frontotemporal networks, which correlated both with
L1 proficiency and with L2 learning rate. However, when
L1 proficiency was partialed out of the correlation
between L2 learning rate and laterality, the resulting
correlations did not significantly change. Thus, al-
though individual differences in L1 proficiency and L2
outcome measures shared some resting-state qEEG fea-
tures, the relation between L2 learning rate and qEEG
laterality was not explained by L1 proficiency.
Another hypothesis explored herein is that the relation
between intrinsic brain functioning and L2 aptitude is ex-
plained, in part, by domain general cognitive mechanisms
such as fluid reasoning. Interestingly, the correlations be-
tween our abbreviated RAPM and resting-state qEEG were
confined to increased theta coherence within the RH
posterior network, which did not overlap with any of the
other L2 aptitude predictors. This may be because shorten-
ing the RAPM reduced our power to detect individual dif-
ferences in fluid reasoning abilities. It is worth noting again
that alpha laterality has been correlated with fluid intelli-
gence (Thatcher et al., 2005), so this remains an area of
interest for future investigation. Along these lines, when
fluid intelligence was partialed out of the relation between
L2 learning rate and alpha and beta laterality, the correla-
tion between posterior alpha and beta laterality decreased,
although not significantly so. Taken together, these results
suggest that the relation between L2 learning and qEEG
characterizations of intrinsic brain functioning at rest is
not significantly explained by fluid reasoning abilities.
Perhaps the strongest piece of evidence about the
roles of L1 proficiency and fluid reasoning abilities in L2
aptitude can be gleaned from the regression analysis
data. Specifically, when NDRT and RAPM scores were
added to the qEEG predictors, there was no evidence that
the resulting models predicted L2 learning rate or total
speaking attempts better than the qEEG models alone. In
fact, positive evidence suggested that the qEEG alone
models explained the data better. This pattern of results,
when combined with the lack of behavioral correlations
and lack of significant changes in partial correlations, sug-
gests that neither L1 proficiency (as measured by NDRT)
nor fluid reasoning abilities (as measured by abbreviated
RAPM) explain the association between resting-state brain
functioning and individual differences in L2 learning
outcomes.
In contrast, regression analyses conducted on the de-
clarative memory posttest suggested that only the model
that included L1 proficiency, fluid reasoning, and qEEG
indicators explained the data significantly. In addition,
when behavioral and brain metrics were entered simul-
taneously, L1 proficiency was the only significant pre-
dictor of posttest performance. These analyses provide
specific evidence that L1 proficiency and increased RH
coherence contribute uniquely to ultimate posttest
proficiency obtained and that L1 proficiency explains
more of the variance than RH functioning. Taken to-
gether, results from these analyses provide preliminary
evidence about the relation between fluid reasoning
abilities, L1 proficiency, and resting-state qEEG as pre-
dictors of various facets of L2 aptitude. As a second
step, the cognitive implications of the current set of re-
sults are discussed in light of what is known about the
neurocognitive bases of neural oscillations.
Leveraging Neural Oscillations to Understand the
Cognitive Bases of L2 Learning
In the current study, we operationalized individual differ-
ences in L2 language learning in four ways: (1) rate of L2
learning, which corresponds to the most basic definition
of aptitude (e.g., Carroll, 1958); (2) long-term declarative
memory for vocabulary learned, which corresponds to
the amount of crystallized information retained across
Prat, Yamasaki, and Peterson
89
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
j
o
c
n
_
a
_
0
1
3
3
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
learning, (3) total speech attempts produced, which re-
flects a willingness to engage with the software aloud,
and (4) the percentage of speech utterances correctly rec-
ognized by the software, which reflects combined decla-
rative (knowing what to say) and procedural (knowing
how to pronounce it) skills.
Relating Beta Power and Laterality to L2 Learning Rate
Consistent with our previous study, increased beta power
in RH posterior regions and increased laterality over
these regions correlated with faster L2 learning, although
only the later survived multiple corrections. Beta fre-
quency bands have become increasingly implicated in
online linguistic processes (see Weiss & Mueller, 2012,
for a review) and seem to be particularly important for
maintaining or binding information that unfolds over
time in memory. For example, experiments measuring
online memory encoding have shown that increased
power in beta (12–20 Hz) was associated with the encod-
ing of novel auditory stimuli (Haenschel, Baldeweg, Croft,
Whittington, & Gruzelier, 2000). In addition, qEEG record-
ings obtained intracranially from the anterior parahippo-
campal regions have shown evidence for increased beta
(19–25 Hz) power and synchronization as a function of in-
creasing working memory task demands (Axmacher,
Schmitz, Wagner, Elger, & Fell, 2008). In a recent review
of cortical plasticity changes associated with short-term
L2 learning, Li and colleagues (2014) summarized evidence
suggesting that increases in RH hippocampal volumes
accompany L2 learning in the laboratory setting.
Despite the increasing amount of evidence implicating
beta frequencies to online language processing, it can be
difficult to understand how characteristics of beta at rest
might predict subsequent language learning. One critical
link comes from a study that used independent compo-
nent analysis to define multiple resting-state networks
and to correlate them with resting-state qEEG character-
istics (Mantini, Perrucci, Del Gratta, Romani, & Corbetta,
2007). They identified six resting-state networks, and the
strongest correlation between beta power and these net-
works was observed for Resting-State Network 4, which
consisted primarily of bilateral temporal networks pro-
posed to be involved in auditory and phonological sys-
tems. Taken together, the existing research suggests
that power and synchronization recorded over the RH
posterior network in beta frequencies may relate to sys-
tems involved in either memory encoding and mainte-
nance broadly construed or more specifically to auditory/
phonological encoding processes. We see this as an
important area for future exploration.
Relating Beta Power to Total Speech Attempts
It is worth noting that the relation between beta power
and the willingness to engage verbally with the software,
as indexed by total speech attempts, was nonoverlapping
with the predictors of L2 learning rate. Specifically, lower
beta power in bilateral frontotemporal regions (with no
laterality effects) was correlated with an increased num-
ber of speech attempts. Taken together, this may reflect a
difference in the importance of frontotemporal versus
posterior beta networks, a difference in the nature of the
two outcome measures (which were not correlated with
one another), or some combination of the two. Below,
we discuss a third option, centered on the interaction be-
tween high- and low-frequency qEEG bands, hemispheres,
and network configuration.
Individual Differences in LH and RH Network
Configuration Differentially Predict L2
Learning Outcomes
The network coherence analyses discussed for the first
time herein contribute critically to our understanding of
the nature of the individual differences in resting-state
qEEG reported previously (Prat et al., 2016). With one ex-
ception discussed subsequently, increased coherence
within and between RH networks was associated with
better L2 learning outcomes whereas decreased coher-
ence within LH was associated with better L2 learning
outcomes (see Figure 3). Specifically, increased coher-
ence across all power bands within the RH frontotemporal
network was predictive both of rate of L2 learning and of
ultimate declarative memory proficiency level achieved,
and increased coherence of all power bands between RH
frontotemporal and posterior networks was positively
correlated with both posttest proficiency and total speak-
ing accuracy measures. Although none of these measures
reflects a “pure” index of L2 learning aptitude, their shared
reliance on declarative memory performance, combined
with the fact that these effects were often largest in alpha
and theta frequency bands, suggests that the availability of
large-scale, undifferentiated networks may promote more
facile initial L2 learning.
The fact that these results were significant when aver-
aging across power bands is also consistent with previous
research showing that increased white matter organiza-
tion in the RH corresponds to better L2 learning (e.g.,
Mamiya et al., 2016; Qi et al., 2015). Qi and colleagues
(2015) suggested that their findings, linking the quality
of the superior and inferior longitudinal fasciculi in the
RH to subsequent Mandarin learning success, may be re-
lated to the particular auditory and visuospatial demands
of Mandarin, which are both tonal and logographic.
However, Mamiya and colleagues (2016) found similar re-
sults in native Chinese learners of English. The fact that the
current study involved native English learners of French
provides further evidence that the relevance of the RH
for L2 learning may not be specific to the perceptual de-
mands of the language. In addition, despite the conver-
gence between our findings and structural brain research,
it is important to note that, without sufficient scalp cover-
age to assess source localization, it is unclear whether
90
Journal of Cognitive Neuroscience
Volume 31, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
j
o
c
n
_
a
_
0
1
3
3
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
j
o
c
n
_
a
_
0
1
3
3
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
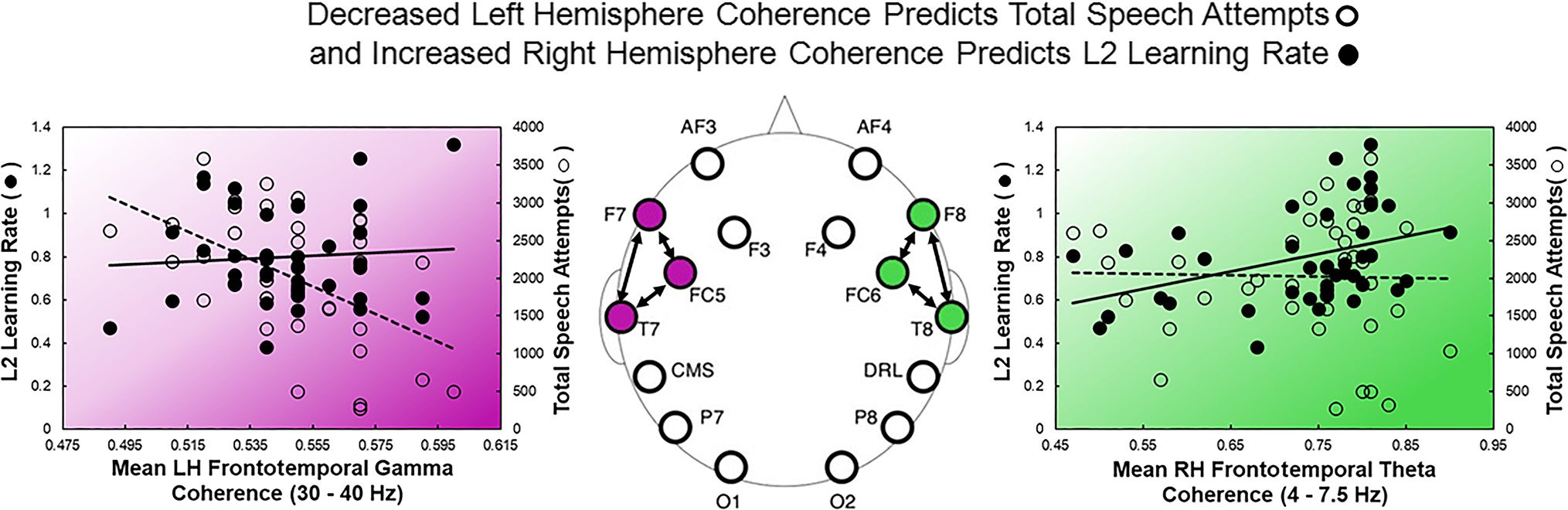
Figure 3. Scatterplots depicting individual differences in rate of L2 learning (filled circles) and total speech attempts (open circles) as a function of
LH coherence over gamma bands (in purple) and RH coherence over theta bands (in green). Electrode locations are displayed in center.
the qEEG coherence recorded over the RH reflects large-
scale networks that are generated in the RH. We see this
as an important next step for this research.
In contrast, lower coherence recorded over LH fronto-
temporal networks in the gamma frequency band (which
is known to reflect local cortical firing patterns) was
strongly associated with a higher willingness to com-
municate. In addition, lower coherence across all fre-
quency bands within the LH posterior network was
correlated with faster rates of L2 learning. Only one L2
learning outcome (L2 learning rate) was associated with
both higher RH coherence and lower LH coherence.
Thus, these differences may reflect fundamental distinc-
tions in the neural computations reflected by the various
L2 outcome tasks.
Relating LH Posterior Coherence to L2 Aptitude
Perhaps the most puzzling findings in the current study
show that decreased coherence in the LH posterior net-
work across all frequency bands was predictive of an in-
creased rate of L2 learning but a decreased number of
total speech attempts (the one exception to the LH/RH
coherence distinction). One might suppose that this re-
flects the fact that speech acts take time and that people
who voluntarily engage in more speech acts move more
slowly through the software than those who don’t.
Although the correlation between the two variables does
trend in that direction, rs(41) = −.26, p = .38, it is not
significant. These results are even more perplexing when
viewed in light of a recent study investigating resting-
state connectivity using fMRI and L2 aptitude (Chai et al.,
2016). Specifically, this study, which also investigated
native English speakers learning French, showed that
increased connectivity between the left visual word form
area (a posterior region) and the superior temporal gyrus
correlated with faster subsequent L2 reading rate. Thus,
our results may suggest that increased lexical access
through reading pathways is associated with a greater
willingness to engage verbally with the software; however,
it seems as though facility with L2 lexical access should
also enable individuals to move through the software
more rapidly. As discussed in the next section, a better
understanding of the nature of individual differences
in total speech attempts is necessary to gain traction on
this puzzle.
Individual Differences in Network-level
Functioning and Speaking Behaviors
Among the most novel findings of the current experi-
ment are those that address the neurocognitive corre-
lates of learners’ willingness to engage verbally with the
software (total speech attempts). Such individual differ-
ences have been characterized in the literature as “will-
ingness to communicate” (Burgoon, 1976; MacIntyre,
2007) and are difficult to measure in canonical classroom
or immersion environments. It is unclear whether exist-
ing research, which has focused on individuals’ willing-
ness to engage verbally with other live speakers, can be
applied to our language training paradigm, which in-
volves interacting with computers. One possible link is
that all language learners were in the same room with
another person (the experimenter) seated at a different
table. It is worth noting that there are several portions of
the learning experience where speaking aloud is optional
and that doing so required learners to produce foreign
sounds in the presence of the experimenter. Thus, it is
plausible that some of the key factors that have been
related to willingness to communicate (e.g., positive self-
confidence, low communication apprehension, high moti-
vation) may drive differences in total speech attempts,
whereas it is less likely that other variables (international
attitudes) drive these differences (Öz, Demirezen, &
Pourfeiz, 2015; Fallah, 2014; Yashima, Zenuk-Nishide, &
Shimizu, 2004; Yashima, 2002).
Our data suggest that this willingness to speak was
largely correlated with the presence of small-world
Prat, Yamasaki, and Peterson
91
networks (or focal representations) as indexed by lower
power in higher-frequency ranges (gamma and beta),
thought to reflect more focal network coupling, over
bilateral frontotemporal and RH posterior regions and
by lower coherence between LH frontotemporal regions
over gamma frequencies. These predictors of total
speech attempts were completely nonoverlapping with
the other L2 outcome variables, suggesting that if total
speech attempts index social cognition in some way,
the neural indicators of these skills are not overlapping
with those associated with L2 learning rate, posttest de-
clarative memory, or even with speaking accuracy, which
was positively correlated with total speech attempts. In
fact, each of these outcome measures was associated
with greater coherence in RH networks. Taken together,
these findings limit our ability to consider the role of
social motivation in RH contributions to L2 learning.
However, our data do provide novel characterizations
of individual differences in willingness to communicate
aloud in experimental settings.
It is critical to note here that total speech attempts
were predicted by two cognitive variables, attentional
blink size and working memory updating accuracy.
Although these two variables are considerably different
in nature, they both deal with the ability to encode seri-
ally presented information in memory. With respect to
the attentional blink in particular, recent work in our
lab has shown that bilingual language experience cor-
relates with the size of attentional blink observed, such
that bilingual individuals who exhibit more balanced
use of both of their languages on a daily basis (or more
frequent use of their less dominant language) were also
shown to have smaller attentional blinks ( Yamasaki,
Stocco, & Prat, 2018). The data reported herein suggests
that this relation may be driven, in part, by individual
differences in willingness to communicate, which may
be underpinned by differences in executive attention.
We see this as an important area for future investigation,
as willingness to communicate has been largely viewed
as a social/personality variable rather than a cognitive
one. Because willingness to communicate relates to
speaking proficiency in our current data and more
broadly to L2 proficiency in the literature (Knell &
Chi, 2012), understanding its neurocognitive under-
pinnings is critical to understanding individual differ-
ences in L2 learning.
Summary
vocabulary knowledge, and 60% of the variance in total
speech attempts. We also provided evidence that
patterns of network connectivity recorded over the left
hemisphere (LH) and RH differentially predict L2 learn-
ing outcomes and share little variance with correlates
of L1 proficiency or fluid reasoning abilities.
Interpretation of these results and their generaliz-
ability are subject to several limitations, however. The first
is to clarify that we are talking about laterality patterns as
recorded over the scalp, which may or may not directly
reflect differences in the functioning of the two hemi-
spheres. Although this is an important limitation, links
between our research and patterns of structural connec-
tivity in the RH (e.g., Mamiya et al., 2016; Qi et al., 2015)
and changes in RH gray matter following short periods of
L2 learning (as discussed in Li et al., 2014) provide some
evidence that the effects we are measuring may be tied
to differences in brain structure. A second limitation is
that our results come from a brief, 8-week exposure to
an L2. Although similar, and sometimes shorter, training
periods are common in L2 aptitude research (see Li
et al., 2014), they do limit the likelihood that learners will
reach advanced proficiency. A third limitation is that the
language training protocol used herein involves a com-
puter software that includes both language and culture
training modules. Thus, it is unclear whether these results
will generalize to classroom learning, to natural immersion
environments, or to longer learning periods that result in
greater proficiency. Nonetheless, the use of the OLCTS
allowed us to ensure that all learners were exposed to
the same training environment and enabled us to collect
data on voluntary speech acts and the accuracy of these
acts, which were uniquely correlated with qEEG indicators
at rest. In addition, it is unclear whether the predictors we
have identified are specific to language learning or repre-
sent some more general facility with learning and memory
that could be applied to multiple complex tasks. In fact,
existing work on alpha and intelligence and beta and
memory suggests the latter or at least some combination
of the two. Finally, although a number of recent experi-
ments have investigated changes in qEEG signals during
artificial language learning (Kepinska et al., 2017; de
Diego-Balaguer, Fuentemilla, & Rodriguez-Fornells,
2011), further research is needed to link the patterns
of brain activity at rest described herein with patterns
of brain activity associated with language aptitude dur-
ing active learning. We see this as an exciting avenue for
future research.
The results reported herein replicate and extend pre-
vious research using qEEG (Prat et al., 2016), resting-state
fMRI (Chai et al., 2016), and structural connectivity
(Mamiya et al., 2016; Qi et al., 2015) metrics of intrinsic
network-level brain functioning to predict variance in the
ability to acquire an L2 in adulthood. Taken together, our
qEEG indicators explained 48% of the variance in rate
of L2 learning, 26% of the variance in declarative posttest
Acknowledgments
This research was funded by a grant from the Office of Naval
Research (ONRBAA13-003) entitled “Training the Mind and
Brain: Investigating Individual Differences in the Ability to
Learn and Benefit Cognitively from Language Training.” We
would like to thank Andrea Stocco, Jose Ceballos, Malayka
Mottarella, and Justin Abernethy for their help with earlier
drafts of the manuscript.
92
Journal of Cognitive Neuroscience
Volume 31, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
j
o
c
n
_
a
_
0
1
3
3
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Reprint requests should be sent to Chantel S. Prat, Department
of Psychology, University of Washington, Box 351525, Seattle,
WA 98195-1525, or via e-mail: csprat@uw.edu.
Notes
1. Although there were unequal numbers of men and women
included in this study, the groups did not differ in any of the L2
learning variables or qEEG indices (all ps > .24).
2.
from https://figshare.com/s/b99fc1dc3cd3c4634695.
Supplementary material for this paper can be retrieved
VERWEISE
Arthur, W., & Day, D. (1994). Development of a short form for
the Raven’s Advanced Progressive Matrices Test. Educational
and Psychological Measurement, 54, 394–403.
Axmacher, N., Schmitz, D., Wagner, T., Elch, C., & Fell, J. (2008).
Interactions between medial temporal lobe, prefrontal
Kortex, and inferior temporal regions during visual working
Erinnerung: A combined intracranial EEG and functional
magnetic resonance imaging study. Zeitschrift für Neurowissenschaften,
28, 7304–7312.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false
discovery rate: A practical and powerful approach to multiple
testing. Journal of the Royal Statistical Society. Serie B,
Methodological, 57, 289–300.
Biedron, A. (2015). Neurology of foreign language aptitude.
Studies in Second Language Learning and Teaching, 5, 13–40.
Birdsong, D. (Ed.) (1999). Second language acquisition and
the critical period hypothesis. Mahwah, NJ: Erlbaum.
Broadbent, D., & Broadbent, M. (1987). From detection to
identification: Response to multiple targets in rapid serial
visual presentation. Wahrnehmung & Psychophysics, 42, 105–113.
Braun, J., Fishco, V., & Hanna, G. (1993). Nelson–Denny
Reading Test: Manual for scoring and interpretation forms
G & H. Rolling Meadows, IL: Riverside Publishing.
Burgoon, J. K. (1976). The unwillingness-to-communicate scale:
Development and validation. Communication Monographs,
43, 60–69.
Carroll, J. (1958). A factor analysis of two foreign language
aptitude batteries. Journal of General Psychology, 59, 3–19.
Carroll, J., & Sapon, S. (1959). Modern Language Aptitude Test.
San Antonio, TX: Psychological Corporation.
Chai, X., Berken, J., Barbeau, E., Soles, J., Callahan, M., Chen, J.,
et al. (2016). Intrinsic functional connectivity in the adult
brain and success in second-language learning. Zeitschrift für
Neurowissenschaften, 36, 755–761.
Chee, M., Soon, C., Lee, H., & Pallier, C. (2004). Left insula
Aktivierung: A marker for language attainment in bilinguals.
Verfahren der Nationalen Akademie der Wissenschaften, USA., 101,
15265–15270.
Cohen, J., Barch, D., Fuhrmann, C., & Servan-Schreiber, D. (1999).
Context-processing deficits in schizophrenia: Converging
evidence from three theoretically motivated cognitive tasks.
Journal of Abnormal Psychology, 108, 120–133.
de Diego-Balaguer, R., Fuentemilla, L., & Rodriguez-Fornells, A.
(2011). Brain dynamics sustaining rapid rule extraction
from speech. Zeitschrift für kognitive Neurowissenschaften, 23,
3105–3120.
Doppelmayr, M., Klimesch, W., Stadler, W., Pöllhuber, D., &
Heine, C. (2002). EEG alpha power and intelligence.
Intelligence, 30, 289–302.
Dörnyei, Z. (2009). The psychology of second language
Erwerb. Oxford, Vereinigtes Königreich: Oxford University Press.
Dörnyei, Z., & Skehan, P. (2003). Individual differences in
second language learning. In C. J. Doughty & M. H. Long
(Hrsg.), The handbook of second language acquisition
(S. 589–630). Oxford: Blackwell Publishing.
Fallah, N. (2014). Willingness to communicate in English,
communication self-confidence, motivation, shyness
and teacher immediacy among Iranian English-major
undergraduates: A structural equation modeling approach.
Learning and Individual Differences, 30, 140–147.
Frank, M., Seeberger, L., & O’Reilly, R. (2004). By carrot or by
stick: Cognitive reinforcement learning in parkinsonism.
Wissenschaft, 306, 1940–1943.
Gardner, R., & Lambert, W. (1965). Language aptitude, intelligence,
and second-language achievement. Journal of Educational
Psychologie, 56, 191–199.
Haenschel, C., Baldeweg, T., Croft, R. J., Whittington, M., &
Gruzelier, J. (2000). Gamma and beta frequency oscillations
in response to novel auditory stimuli: A comparison of
human electroencephalogram (EEG) data with in vitro
Modelle. Verfahren der Nationalen Akademie der Wissenschaften,
USA., 97, 7645–7650.
Handschin, C. H., & Henmon, V. (1930). Prognosis tests in
the modern foreign languages. New York: Macmillan.
Hanslmayr, S., Sauseng, P., Doppelmayr, M., Schabus, M., &
Klimesch, W. (2005). Increasing individual upper alpha
power by neurofeedback improves cognitive performance in
human subjects. Applied Psychophysiology and Biofeedback,
30, 1–10.
Johnson, W., Friedland, L., Watson, A., & Surface, E. (2012).
The art and science of developing intercultural competence.
In P. Durlach & A. Lesgold (Hrsg.), Adaptive technologies
for training and education (S. 261–286). New York:
Cambridge University Press.
Kane, M., Conway, A., Miura, T., & Colflesh, G. (2007). Arbeiten
Erinnerung, attention control, and the N-back task: A question
of construct validity. Journal of Experimental Psychology.
Learning, Memory, and Cognition, 33, 615.
Kepinska, O., Pereda, E., Caspers, J., & Schiller, N. (2017).
Neural oscillatory mechanisms during novel grammar
learning underlie language analytical abilities. Brain and
Language, 175, 99–110.
Klimesch, W. (1999). EEG alpha and theta oscillations reflect
cognitive and memory performance: A review and analysis.
Brain Research Reviews, 29, 169–195.
Klimesch, W., Sauseng, P., & Gerloff, C. (2003). Enhancing
cognitive performance with repetitive transcranial magnetic
stimulation at human individual alpha frequency. Der
Europäisches Journal für Neurowissenschaften, 17, 1129–1133.
Knell, E., & Chi, Y. (2012). The roles of motivation, affective
attitudes, and willingness to communicate among Chinese
students in early English immersion programs. International
Education, 41, 66.
Li, P., Legault, J., & Litcofsky, K. (2014). Neuroplasticity as a
function of second language learning: Anatomical changes
in the human brain. Kortex, 58, 301–324.
MacIntyre, P. D. (2007). Willingness to communicate in the
second language: Understanding the decision to speak as a
volitional process. The Modern Language Journal, 91,
564–576.
Mamiya, P., Richards, T., Coe, B., Eichler, E., & Kuhl, P. (2016).
Brain white matter structure and COMT gene are linked to
second-language learning in adults. Verfahren der
Nationale Akademie der Wissenschaften, USA., 113, 7249–7254.
Mantini, D., Perrucci, M., Del Gratta, C., Roma, G., & Corbetta,
M. (2007). Electrophysiological signatures of resting state
networks in the human brain. Verfahren des Nationalen
Akademie der Wissenschaften, USA., 104, 13170–13175.
Mechelli, A., Crinion, J., Noppeney, U., O’Doherty, J., Aschenbrenner,
J., Frackowiak, R., et al. (2004). Neurolinguistics: Structural
plasticity in the bilingual brain. Natur, 431, 757.
Prat, Yamasaki, and Peterson
93
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Monsell, S. (2003). Task switching. Trends in den Kognitionswissenschaften,
7, 134–140.
Öz, H., Demirezen, M., & Pourfeiz, J. (2015). Willingness to
communicate of EFL learners in Turkish context. Learning
and Individual Differences, 37, 269–275.
Perani, D., Paulesu, E., Galles, N., Dupoux, E., Dehaene, S.,
Bettinard, V., et al. (1998). The bilingual brain: Proficiency
and age of acquisition of the second language. Gehirn, 121,
1841–1852.
Prat, C. (2011). The brain basis of individual differences in
language comprehension abilities. Language and Linguistics
Compass, 5, 635–649.
Prat, C., Mason, R., & Nur, M. (2011). Individual differences in
the neural basis of causal inferencing. Brain and Language,
116, 1–13.
Prat, C., Yamasaki, B., Kluender, R., & Stocco, A. (2016). Resting
state qEEG predicts rate of second language learning in
Erwachsene. Brain and Language, 157–158, 44–50.
Qi, Z., Han, M., Garel, K., San Chen, E., & Gabriela, J. (2015).
White-matter structure in the right hemisphere predicts
Mandarin Chinese learning success. Zeitschrift für
Neurolinguistics, 33, 14–28.
R Core Team. (2013). R: A language and environment for
statistical computing. Vienna: R Foundation for Statistical
Computing.
Raftery, A. (1995). Bayesian model selection in social research.
Soziologische Methodologie, 25, 111–163.
Raymond, J., Shapiro, K., & Arnell, K. (1992). Temporary
suppression of visual processing in an RSVP task: Ein
attentional blink? Journal of Experimental Psychology.
Human Perception and Performance, 18, 849–860.
Russell, C. W. (1858). The life of Cardinal Mezzofanti. London:
Longman, Braun, and Co.
Sasaki, M. (1996). Second language proficiency, foreign language
aptitude, and intelligence: Quantitative and qualitative
Analysen (Bd. 6). New York: Peter Lang Pub Incorporated.
Sauseng, P., Klimesch, W., Schabus, M., & Doppelmayr, M. (2005).
Fronto-parietal EEG coherence in theta and upper alpha
reflect central executive functions of working memory.
International Journal of Psychophysiology, 57, 97–103.
Snow, C., & Hoefnagel-Höhle, M. (1978). The critical period
for language acquisition: Evidence from second language
learning. Kinderentwicklung, 49, 1114–1128.
Stocco, A., Murray, N. L., Yamasaki, B. L., Renno, T. J.,
Nguyen, J., & Prat, C. S. (2017). Individual differences in
the Simon effect are underpinned by differences in the
competitive dynamics in the basal ganglia: An experimental
verification and a computational model. Cognition, 164,
31–45.
Bräunen, L., Chen, L., Yip, V., Chan, A., Yang, J., Gao, J., et al. (2011).
Activity levels in the left hemisphere caudate–fusiform
circuit predict how well a second language will be learned.
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
108, 2540–2544.
Thatcher, R., Norden, D., & Biver, C. (2005). EEG and intelligence:
Relations between EEG coherence, EEG phase delay and
power. Clinical Neurophysiology, 116, 2129–2141.
Unsworth, N., Heitz, R., Schrock, J., & Engle, R. (2005). Ein
automated version of the operation span task. Behavior
Research Methods, 37, 498–505.
Wagenmakers, E. (2007). A practical solution to the pervasive
problems of p-values. Psychonomic Bulletin and Review,
14, 779–804.
Weiss, S., & Mueller, H. (2012). “Too many betas do not spoil
the broth”: The role of beta brain oscillations in language
Verarbeitung. Grenzen in der Psychologie, 3, 1-15.
Wen, Z., Biedroń, A., & Skehan, P. (2017). Foreign language
aptitude theory: Yesterday, today and tomorrow. Language
Teaching, 50, 1–31.
Yamasaki, B. L., Stocco, A., & Prat, C. S. (2018). Relating
individual differences in bilingual language experiences to
executive attention. Language, Cognition, and Neuroscience,
33, 1128–1151.
Yashima, T. (2002). Willingness to communicate in a second
Sprache: The Japanese EFL context. The Modern Language
Zeitschrift, 86, 54–66.
Yashima, T., Zenuk-Nishide, L., & Shimizu, K. (2004). Der
influence of attitudes and affect on willingness to
communicate and second language communication.
Language Learning, 54, 119–152.
94
Zeitschrift für kognitive Neurowissenschaften
Volumen 31, Nummer 1
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
1
1
7
8
1
7
8
8
1
2
6
/
J
Ö
C
N
_
A
_
0
1
3
3
7
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3