Individual Differences in Face Identity Processing
with Fast Periodic Visual Stimulation
Buyun Xu1, Joan Liu-Shuang2, Bruno Rossion2,3, and James Tanaka1
Abstrakt
■ A growing body of literature suggests that human individuals
differ in their ability to process face identity. These findings
mainly stem from explicit behavioral tasks, such as the Cambridge
Face Memory Test (CFMT). Jedoch, it remains an open ques-
tion whether such individual differences can be found in the ab-
sence of an explicit face identity task and when faces have to be
individualized at a single glance. In the current study, we tested
49 participants with a recently developed fast periodic visual stim-
ulation (FPVS) paradigm [Liu-Shuang, J., Norcia, A. M., & Rossion,
B. An objective index of individual face discrimination in the right
occipitotemporal cortex by means of fast periodic oddball stimu-
lation. Neuropsychologie, 52, 57–72, 2014] in EEG to rapidly, ob-
jectively, and implicitly quantify face identity processing. Im
FPVS paradigm, one face identity (A) was presented at the
frequency of 6 Hz, allowing only one gaze fixation, with different
face identities (B, C, D) presented every fifth face (1.2 Hz; d.h.,
AAAABAAAACAAAAD…). Results showed a face individuation re-
sponse at 1.2 Hz and its harmonics, peaking over occipitotem-
poral locations. The magnitude of this response showed high
reliability across different recording sequences and was significant
in all but two participants, with the magnitude and lateralization
differing widely across participants. There was a modest but sig-
nificant correlation between the individuation response ampli-
tude and the performance of the behavioral CFMT task, despite
the fact that CFMT and FPVS measured different aspects of face
identity processing. Taken together, the current study highlights
the FPVS approach as a promising means for studying individual
differences in face identity processing. ■
EINFÜHRUNG
It is generally understood that humans are face experts
who can recognize a familiar face in a blink of an eye
without conscious effort or forethought (Tanaka, 2001;
Diamond & Carey, 1986). Jedoch, the past decade
has seen a number of behavioral studies suggesting that,
like most human traits and capacities, the ability to
recognize face identities lies on a continuum (z.B.,
Rhodes, Jeffery, Taylor, Hayward, & Ewing, 2014;
DeGutis, Wilmer, Mercado, & Cohan, 2013; Dennett,
McKone, Edwards, & Susilo, 2012; Wilmer et al., 2012;
Germine, Duchaine, & Nakayama, 2011; Wilhelm et al.,
2010; Bowles et al., 2009; Russell, Duchaine, & Nakayama,
2009). On one end of the continuum are the “super-face
recognizers” who can identify a vast catalog of faces after
a single presentation under limited viewing conditions
(Bobak, Bennetts, Parris, Jansari, & Bate, 2016; Russell
et al., 2009). On the other end are people who have
experienced no brain damage or trauma and yet have a life-
long impairment recognizing the faces of close friends and
relatives (“developmental prosopagnosia” or “congenital
prosopagnosia”; Duchaine & Nakayama, 2004, 2006A;
Behrmann & Avidan, 2005). Within the extremes, Gesicht
1University of Victoria, 2University of Louvain, 3Centre Hospitalier
Regional Universitaire de Nancy
© 2017 Massachusetts Institute of Technology
recognition performance as measured behaviorally is a nor-
mally distributed skill that is mediated by factors such as age
(Germine et al., 2011; Hildebrandt, Sommer, Herzmann, &
Wilhelm, 2010), Sex (Sommer, Hildebrandt, Kunina-
Habenicht, Schacht, & Wilhelm, 2013), or sociability
(Halliday, MacDonald, Scherf, & Tanaka, 2014; Bate, Parris,
Haslam, & Kay, 2010).
Individual differences in face identity processing are
typically measured explicitly using behavioral methods.
The most widely used behavior test is the Cambridge
Face Memory Test (CFMT; Duchaine & Nakayama,
2006B), which requires participants to first memorize
faces and then to recognize each of those faces among
other foil faces under different viewing conditions (z.B.,
viewpoint changes, with added noise). In standard ERPs
extracted from EEG recordings, the most prominent
signature of neural activities related to face processing
is the N170 component. Faces elicit a robust negative
response in the brain wave approximately 170 msec after
the onset of the face stimuli (Bentin, Allison, Chip, Perez,
& McCarthy, 1996), and this N170 response is larger to
faces than nonface objects with the largest difference ob-
served at right occipital-temporal locations on the scalp
(see Rossion & Jacques, 2011, für eine Rezension). Kaltwasser,
Hildebrandt, Recio, Wilhelm, and Sommer (2014) Und
Herzmann, Kunina, Sommer, and Wilhelm (2010) gebraucht
structural equation modeling techniques and found a
Zeitschrift für kognitive Neurowissenschaften 29:8, S. 1368–1377
doi:10.1162/jocn_a_01126
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
Ö
6
C
4
N
3
_
6
A
/
_
J
0
Ö
1
C
1
N
2
6
_
A
P
_
D
0
1
B
1
j
2
G
6
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
F
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
modest relation across individuals between the N170
latency (but not amplitude) and behavioral performance
of an explicit face recognition task using an old/new
recognition paradigm.
Jedoch, whether individual differences in face iden-
tity processing can be captured and quantified without an
explicit behavioral task remains an open question. Das ist
an important issue because certain individuals may have
difficulties understanding task instructions (young chil-
dren, neurological or psychiatric populations, usw.), Und
the individual differences in performance at the CFMT
or other explicit behavioral tests can be greatly influ-
enced by factors such as memory, motivation, attention,
decision processes, und so weiter. Darüber hinaus, although it is
often stated that faces can be recognized in a blink of an
eye (d.h., a “single glance”; Hsiao & Cottrell, 2008; Willis &
Todorov, 2006), faces are shown for a long (or an un-
limited) period in explicit behavioral tests such as the
CFMT or the widely used Benton Face Recognition Test
(Benton & Van Allen, 1968). Somit, whether individual
differences in face identity processing can be captured
in a task where only one gaze fixation is allowed on a
face remains unknown.
To shed light on these questions, the current study em-
ployed a fast periodic visual stimulation (FPVS) paradigm
coupled with EEG recording to measure individual differ-
ences in face identity processing (Rossion & Boremanse,
2011). In the specific FPVS oddball paradigm used here
(Liu-Shuang, Norcia, & Rossion, 2014), same identity face
stimuli are presented at a fast fixed rate (6 Hz, oder 6 images/
Sek, general stimulation frequency), allowing only one
gaze fixation per face. Such periodic stimulation elicits a
periodic response at the same driving frequency and its
harmonics (often called “steady-state visual evoked po-
tentials”), which can be captured using scalp EEG record-
ings (Regan, 1966, 1989; see Norcia, Appelbaum, Ales,
Cottereau, & Rossion, 2015, für eine Rezension). Wichtig,
every fifth image in this stimulation sequence shows a dif-
ferent face identity, so that face identity changes occur at
a rate of 1.2 Hz (Figur 1). In these conditions, the pres-
ence of a periodic response at 1.2 Hz and its harmonics
(d.h., individuation response) directly reflects the detec-
tion of face identity changes (d.h., individuation response).
Zusätzlich, the face individuation response captured in
this way reflects high-level face-specific processing. Für
Beispiel, the individuation response resists large changes
of stimulus size, but its magnitude is largely and signifi-
cantly reduced with inversion and contrast reversal (Liu-
Shuang et al., 2014), two image manipulations known to
disproportionately affect perceptual processing of faces
relative to other object categories ( Yin, 1969; Galper,
1970, jeweils). Darüber hinaus, a recent study showed that
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
Ö
6
C
4
N
3
_
6
A
/
_
J
0
Ö
1
C
1
N
2
6
_
A
P
_
D
0
1
B
1
j
2
G
6
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
.
/
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
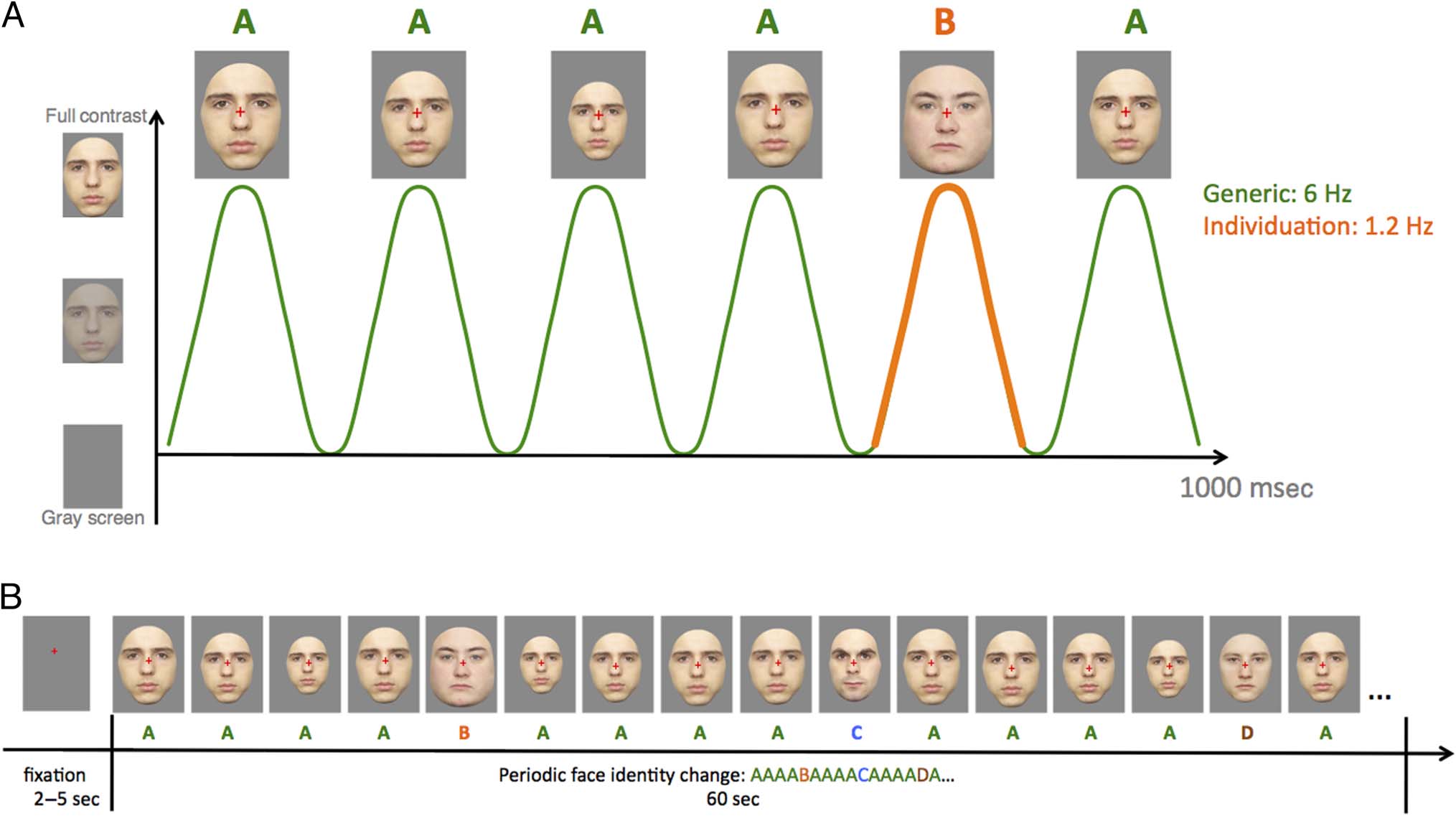
Figur 1. Schematic illustration of the experimental paradigm (adapted from Liu-Shuang et al., 2014). (A) Faces were presented by sinusoidal
contrast modulation at a rate of 6 Hz. A base face was presented four times successively, with a different oddball face presented at every fifth cycle
of presentation. To avoid pixel-wise processing, face size was randomly varied between 80% minimum and 120% maximum at every stimulation
Zyklus. (B) The stimulation sequence started with a fixation cross appearing on the screen for 2–5 sec, after which faces were shown at a rate of 6 Hz
für 60 Sek. Participants were asked to fixate on the cross positioned on the face stimuli and to respond to nonperiodic fixation-cross color changes.
Xu et al.
1369
a specific impairment in face identity processing could
be evidenced through this approach. More precisely,
although all typically developed participants showed sig-
nificant face individuation responses, a well-documented
patient with acquired prosopagnosia (Rossion et al., 2003)
showed no face individuation responses while her general
visual response remained intact relative to these age-
matched participants (Liu-Shuang, Torfs, & Rossion,
2016).
FPVS offers several additional advantages for measur-
ing individual differences in face identity processing.
Erste, FPVS measures face identity processing directly,
without having to subtract or regress out any control
condition responses. Most studies measuring face indi-
viduation in electrophysiology have used adaptation
paradigms in which the response to a series of repeated
face identities is compared with responses to different
face identities (z.B., Jacques, d’Arripe, & Rossion, 2007;
Heisz, Watter, & Shedden, 2006). Im Gegensatz, our FPVS
paradigm contains an inherent subtraction such that re-
sponses at 1.2 Hz and harmonics are directly driven by
the perceived change of face identity. Zweite, the data
analysis is objective because only responses at the prede-
fined frequency of stimulation and its harmonics need to
be considered. This is one of the advantages of FPVS-EEG
and frequency domain analysis over standard ERP mea-
sures because no arbitrary definition of the component
time window is needed, which can be quite subjective
and different across studies. Dritte, the periodic FPVS sig-
nal can easily be segregated from artifacts and spontane-
ous brain activity (Srinivasan, Bibi, & Nunez, 2006; Regan,
1989). More precisely, providing that a high-frequency
resolution is obtained by recording and analyzing data
from a sufficiently long time window, the signal in the
EEG spectrum is found in a few specific and very narrow
frequency bins corresponding to the stimulation frequen-
cies (d.h., if stimulated at 6 Hz, responses will be found in
frequency bins of 6 Hz and its harmonics only). In con-
trast, the background EEG noise and artifacts (alpha
waves, blinks, muscle potentials, usw) are not periodic
and will be distributed across many frequencies (broad-
band), thus having a reduced impact on the frequencies
of interest (Rossion, 2014; Regan, 1989). Infolge, FPVS
signals are more resilient to noise as compared with ERP
and require less strenuous preprocessing and artifact re-
jection steps, providing a measure of perceptual process-
ing that is minimally transformed from the original data
(Rossion, 2014). daher, only a small number of se-
quences are necessary to obtain a significant response,
an advantage that is valuable in studies of individual dif-
ferences in some specific populations (Kinder, infants,
people with cognitive disabilities, usw.). Vierte, the short
presentation time of each face (z.B., 167 msec per face
with a 6-Hz presentation rate) prevents extraneous sac-
cades and cognitive processes from interfering with face
identity processing, which only requires a very brief pre-
sentation time (Alonso-Prieto, Van Belle, Liu-Shuang,
Norcia, & Rossion, 2013). Last but not least, the FPVS re-
sponse can be obtained without contamination from de-
cisional or motor processes, because the observers do
not have to perform any explicit face identity processing
tasks (z.B., Liu-Shuang et al., 2014). daher, based on
these advantages, we hypothesize that FPVS will allow for
sensitive and reliable measures of individual face identity
processing abilities.
Zusammenfassend, the purposes of the current study were to
(1) investigate whether the FPVS approach is sensitive to
individual differences in face identity processing and (2)
study the relationship between the individual differences
in this task-independent neurophysiological measure
captured with FPVS and an explicit behavioral measure
with CFMT.
METHODEN
Teilnehmer
We tested 49 Teilnehmer (24 men, all right-handed) WHO
were recruited from the University of Victoria with com-
pensation of bonus course credit. All of the participants
were white and reported normal or corrected-to-normal
vision and no history of brain injury. The average age of
the participants was 21.93 years with a standard deviation
von 3.69 Jahre. The experiment and consenting procedures
were approved by the human research ethics committee
of University of Victoria.
Cambridge Face Memory Test
Participants first performed the CFMT before they went
to the EEG laboratory. In the CFMT, participants first
completed an introductory phase in which they memo-
rized six target faces. A target face was first presented
from three different views (front, right profile, left pro-
file) für 3 sec per view. Participants then performed
3 three-alternative forced-choice trials, with one trial for
each of the three views. The process was repeated for the
remaining five faces, ergebend 18 Versuche. Nächste, partici-
pants studied frontal views of the same six target faces
für 20 sec and then were presented with 30 forced-choice
test displays in the no-noise phase. Each display con-
tained one target face and two distractor faces. Partici-
pants were told to select the face that matched one of
the original six target faces. The matching faces varied
from their original presentation in terms of lighting con-
dition, pose, oder beides. Nächste, participants completed the
noise phase of the task where they were presented with
the six target faces to study, followed by 24 test displays
presented in Gaussian noise. The data from the trials in
the no-noise phase and noise phase were used to calcu-
late the behavioral face recognition performance scores.
On average, participants took 10–15 min to perform the
CFMT.
1370
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 8
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
Ö
6
C
4
N
3
_
6
A
/
_
J
0
Ö
1
C
1
N
2
6
_
A
P
_
D
0
1
B
1
j
2
G
6
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Stimuli
Stimuli used in the FPVS experiment consisted of full-
front colored photographs of 25 male and 25 female faces
with neutral facial expression (see Liu-Shuang et al.,
2014). Each face picture was taken under the same con-
ditions of lighting and background and with the same
face-to-camera distance. External features such as hair
and ears were cropped out using Adobe Photoshop,
and the isolated faces were put against a neutral gray
background. Final images were resized to a height of
250 pixels and width from 175 Zu 197 pixels. The average
visual angle of the stimuli was 7.1° in height by 5.2° in
width at a 58-cm viewing distance and 1280 × 1024
display resolution.
Verfahren
The experiment was composed of four 60-sec sequences,
with two sequences presenting all female faces and two
sequences presenting all male faces. Half of the partici-
pants were presented the sequences with the order of
female-male-male-female, and the other half were
presented with the order of male-female-female-male.
Within each sequence (Figure 1A), face pictures were
presented six times per second (frequency = 6 Hz)
through sinusoidal contrast modulation using a custom
script running with Psychtoolbox in MATLAB 2010 (Der
Mathworks, Inc., Natick, MA) (z.B., Liu-Shuang et al.,
2014; Rossion & Boremanse, 2011). Each cycle lasted
166.7 msec and began with a uniform gray background
from which a face appeared as its contrast increased. Full
contrast was reached at 83.3 msec and then decreased at
the same rate. The size of each face varied randomly
zwischen 80% Und 120% In 2% steps at every cycle of pre-
sentation to minimize low-level face feature overlap. In
every sequence, a fixation cross first appeared on the
screen for 2–5 sec (randomly jittered across sequences),
after which faces were shown at a rate of 6 Hz for 60 Sek.
Within the 60 sec of stimulation, one face was randomly
selected as the “base” face (d.h., Face A) wiederholen
throughout the sequence. Every fifth face was replaced
by different identity “oddball” faces, randomly selected
from the remaining 24 faces of the same sex (d.h., Faces
B, C, D…; Figure 1B), forming a stimulation pattern of
A A A A B A A A A C A A A A D A A A A E A A … . A s a r e s u l t ,
participants were presented with a face six times every
zweite (general base frequency rate, frequency =
6 Hz) and a face identity change five times every 6 Sek
(individuation frequency, frequency/5 = 1.2 Hz). Re-
sponses at the general base frequency rate and its
harmonics reflect general visual processing, whereas re-
sponses at the individuation frequency harmonics reflect
face identity processing. Participants performed an
orthogonal task in which they were asked to detect the
brief (200 ms) color change (from red to blue) of a
fixation cross situated in the center of the face stimuli
below the eyes, to ensure that they maintained a constant
level of attention. Color changes occurred randomly in
each sequence for eight times. No explicit instructions
were given to the participants to pay attention to the face
Identitäten, and although all of the participants mentioned
that they noticed the face identity change, none of them
reported that the identity change was periodic. Partici-
pants performed at ceiling for this task, with an accuracy
von 98.8% (95% CI [98.3%, 99.2%] and response time of
523.5 ms (95% CI [492.7, 554.4]).
EEG Acquisition
The EEG was recorded using a montage of 36 electrode
sites in accordance to the extended International 10–
20 System ( Jasper, 1958). Signals were acquired using
Ag–AgCl ring electrodes mounted in a nylon electrode
cap with an abrasive, conductive gel (EASYCAP GmbH,
Herrsching-Breitbrunn, Deutschland). Signals were ampli-
fied by low-noise electrode differential amplifiers with a
frequency response of DC 0.017–67.5 Hz (90-dB octave
roll-off ) and digitized at a rate of 250 Hz. Digitized signals
were recorded to disk using Brain Vision Recorder soft-
ware (Brain Products GmbH, München, Deutschland). Der
impedances were maintained below 10 kΩ. The EEG
was recorded using the average reference.
EEG Analysis
EEG signal was processed using Letswave 5 (Mouraux &
Iannetti, 2008) and MATLAB 2012. EEG data were 0.10- Zu
100-Hz band-pass filtered using a Butterworth filter with a
slope of 24 dB/octet. All channels were rereferenced to
an average reference. The continuous EEG recording of
each participant was cropped into four 60-sec segments
time-locked to the onset of the first cycle of presentation
and the offset of the last cycle of presentation in each
stimulation sequence. For the calculation of responses
at the level of individual participants, data of the four se-
quences were first averaged in the time domain to
reduce EEG noise (d.h., activity unrelated to the stimula-
tion). A fast Fourier transform was then applied, and fre-
quency amplitude was extracted at a resolution of 1/60 =
0.017 Hz. Jedoch, for the purposes of reliability calcu-
lation (see Reliability Analysis section), fast Fourier trans-
form was also applied separately to each of the four
sequences in the time domain. Baseline- corrected ampli-
tudes were calculated following the method used in the
previous studies (z.B., Liu-Shuang et al., 2016; Retter &
Rossion, 2016; Dzhelyova & Rossion, 2014A). More
precisely, at each frequency bin, the mean amplitude of
Die 20 surrounding frequency bins (10 on each side,
excluding the immediately adjacent bin) was subtracted
from the frequency bin of interest.
Group level z scores were used to determine the range
of frequency harmonics to take into account and were
Xu et al.
1371
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
Ö
6
C
4
N
3
_
6
A
/
_
J
0
Ö
1
C
1
N
2
6
_
A
P
_
D
0
1
B
1
j
2
G
6
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
.
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
calculated using grand-averaged frequency amplitudes
pooled across all channels. The mean and standard devi-
ation of the 20 frequency bins (10 on each side, excluding
the immediately adjacent bin) surrounding the frequency
of interest were used to calculate the z score. A conser-
vative threshold of z > 3.1 ( P < .001, one-tailed; i.e.,
signal > noise) was used, in line with previous research
(Liu-Shuang et al., 2016). After the selection of the rele-
vant harmonic frequencies, the face individuation re-
sponse and the general visual response were quantified
as the sum of these harmonics (Retter & Rossion, 2016).
Reliability Analysis
The internal reliability of behavioral and EEG measures
was calculated with Cronbach’s alpha, using the individ-
ual items of the CFMT and individual sequences of the
FPVS experiment.
ERGEBNISSE
EEG Data
Frequency Domain Analysis
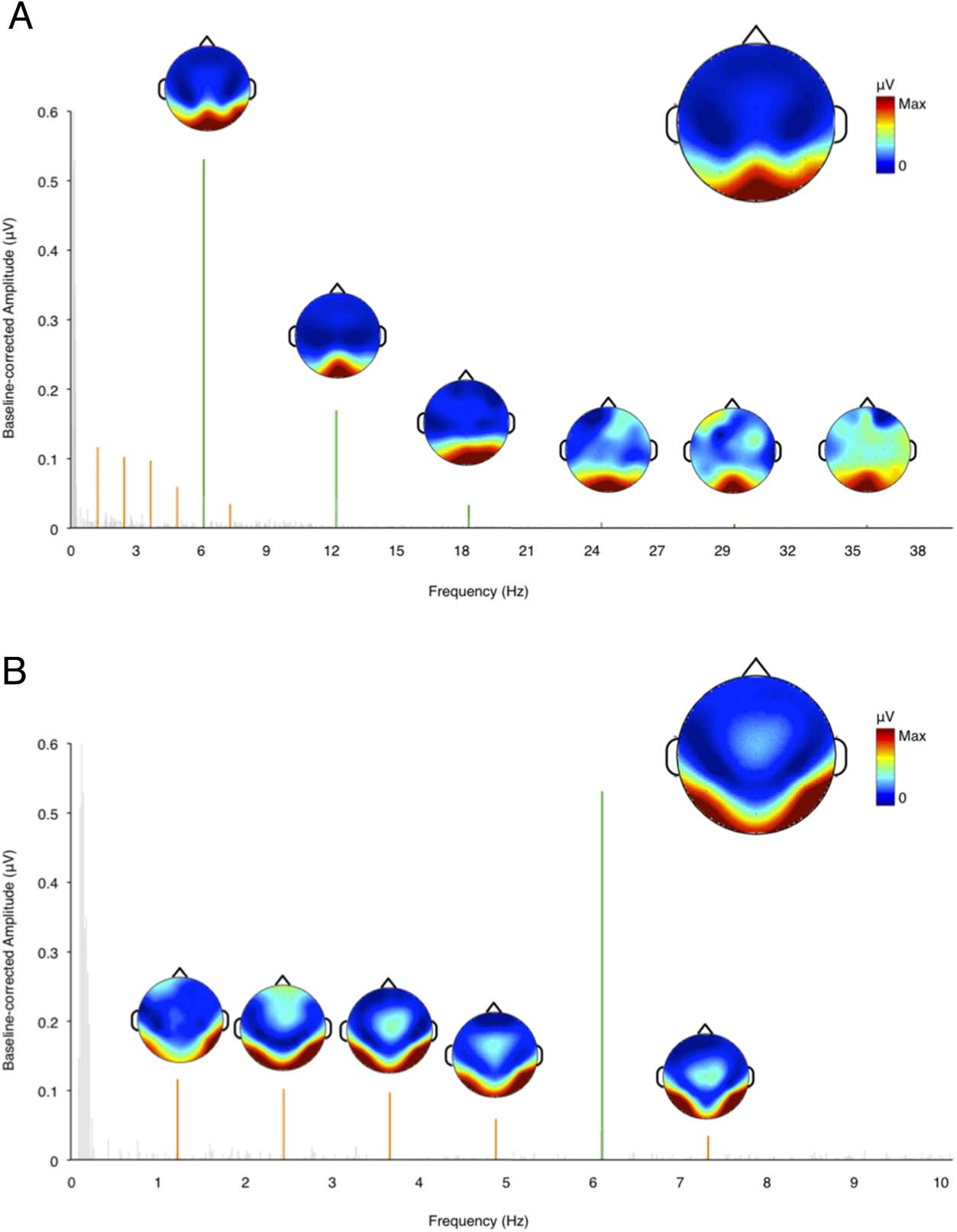
General visual responses were found at 6 Hz and its har-
monics (12 Hz, 18 Hz, …) in all participants. At the group
Ebene, responses were significant until the sixth harmonic
(36 Hz; see Table 1A). We therefore considered the first
six harmonics for analysis. As shown in Figure 2A, allgemein
visual responses had a medial occipital scalp distribution.
For each participant, the aggregated general response
was quantified as the sum of baseline-corrected ampli-
tudes of the first six harmonics on channels Oz and
POz. The internal consistency reliability (Cronbach’s al-
pha) of this general visual response was .98 across the
four individual sequences.
For the face individuation response, significant peaks
were found at 1.2 Hz and its harmonics (2.4 Hz, 3.6 Hz,
Tisch 1. Group Level z Scores for the Fundamental Frequency
and Harmonics of the (A) General and (B) Individuation
Response Based on the Average of All Channels
(A) General Response
(B) Individuation Response
Frequency (Hz)
z Scores
Frequency (Hz)
z Scores
F = 6
2F = 12
3F = 18
4F = 24
5F = 30
6F = 36
7F = 42
79.03
42.03
21.53
8.72
5.59
6.58
0.98
F/5 = 1.2
2F/5 = 2.4
3F/5 = 3.6
4F/5 = 4.8
6F/5 = 7.2
7F/5 = 8.4
8F/5 = 9.6
7.47
10.17
17.81
14.88
8.72
2.45
0.67
Numbers in bold indicate significant responses (z > 3.1).
…) and remained significant at the group level until the
sixth harmonic (7.2 Hz). On the basis of previous studies
as well as the topography of individual harmonics in the
current study, data were grouped into two occipitotem-
poral regions (right hemisphere: P8, PO8, TP10; links
hemisphere: P7, PO7, TP9; Figure 2B). Despite the short
recording time (d.h., 4 min), significant individuation re-
sponses were found at these regions in all participants
but two. We quantified the face individuation response
in each individual participant by summing the baseline-
corrected amplitudes of the first six harmonics, excluding
the fifth harmonic as it was confounded with the 6-Hz
general frequency. At the group level, a t test showed that
the aggregated individuation responses were significantly
larger (T(48) = 2.43, p = .01, one-tailed) over the right
occipital–temporal channels (M = 1.15, 95% CI [0.93,
1.37]) compared with the left occipital–temporal chan-
nels (M = 0.89, 95% CI [0.70, 1.09]). The internal consis-
tency reliability (Cronbach’s alpha) for the individuation
response score across four sequences was .87 when the
responses were aggregated from both the left and right
occipital–temporal locations.
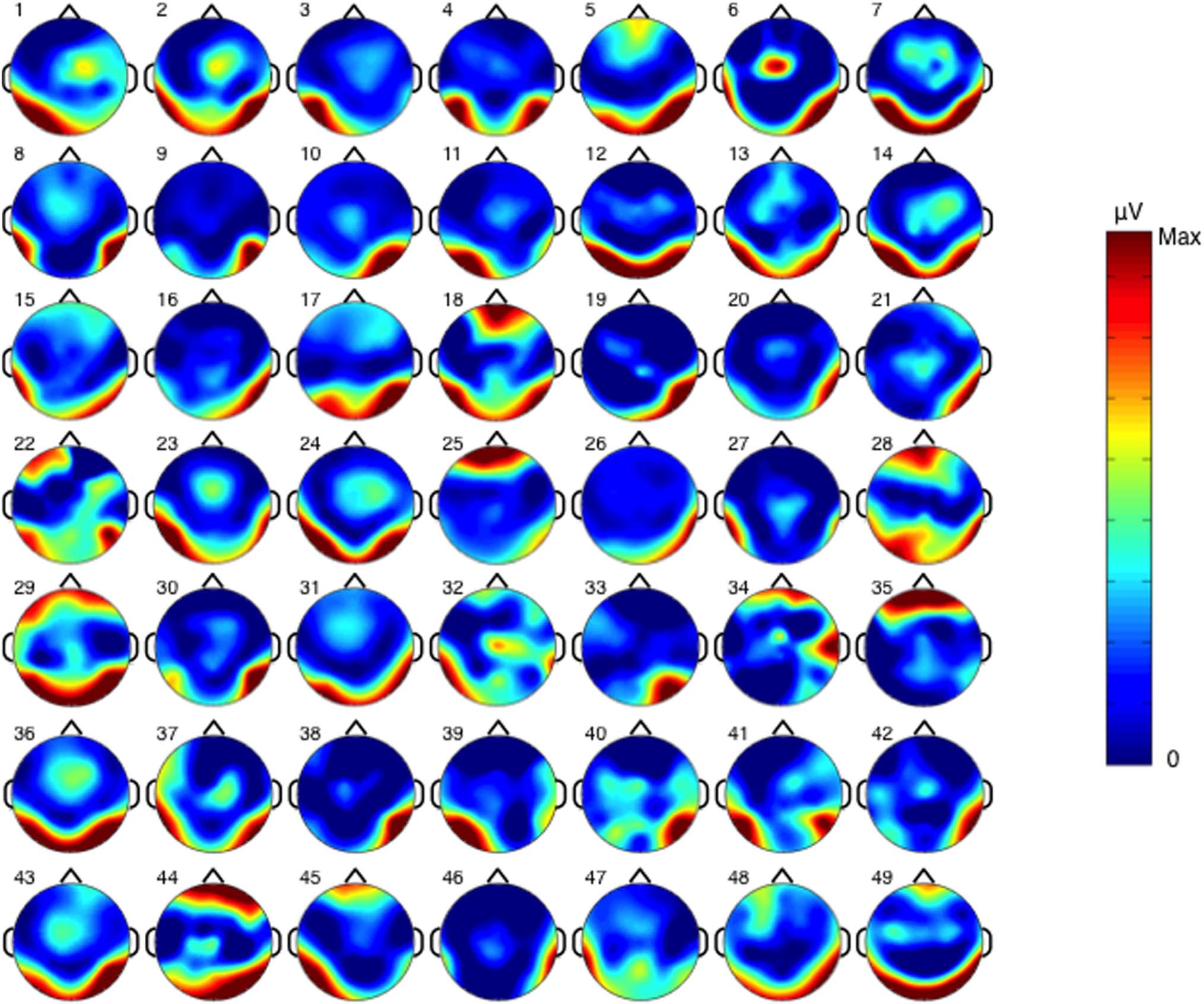
From visual inspection (Figur 3), individual differ-
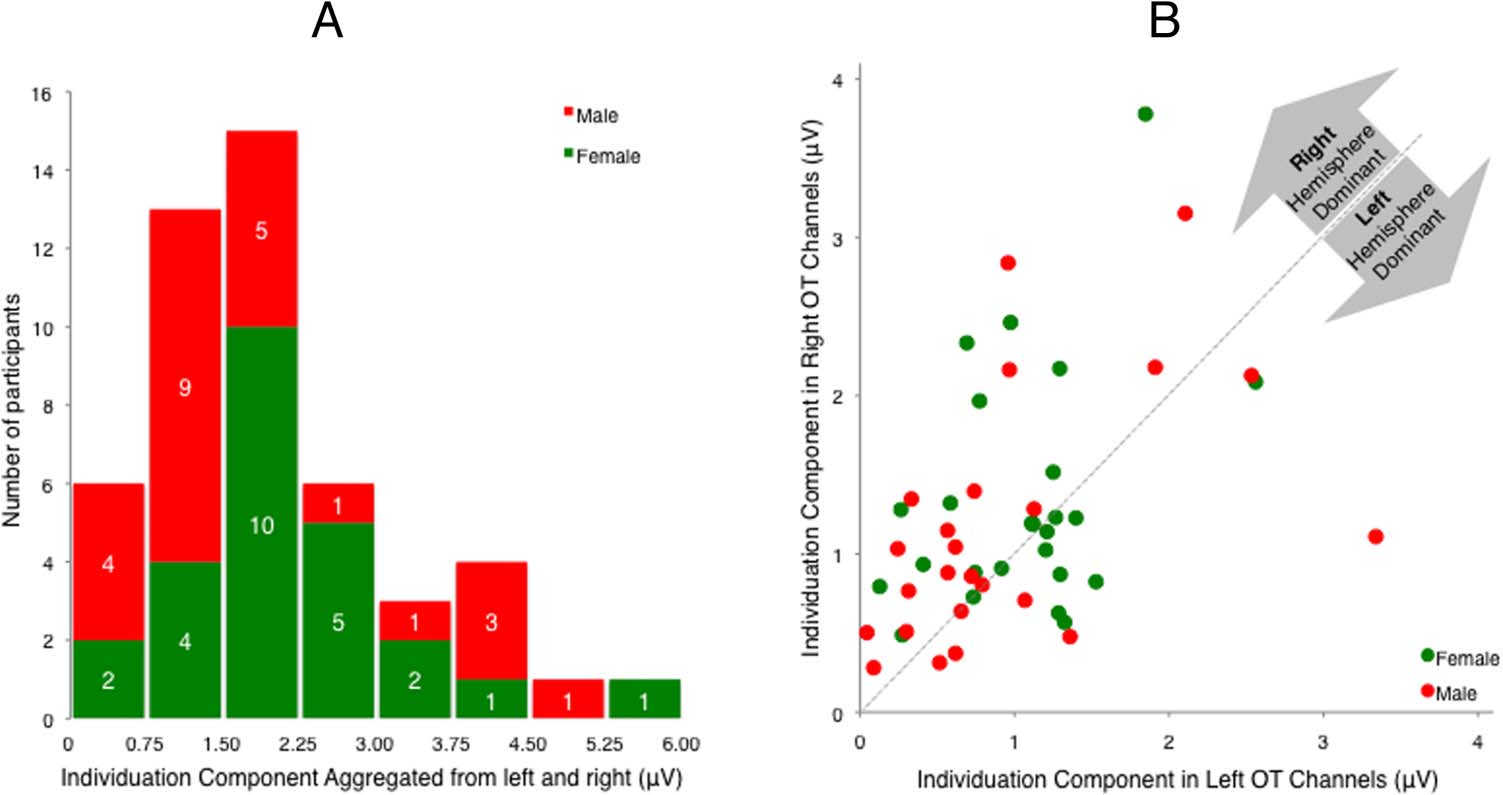
ences are evident in the individuation response. Der
magnitude of the individuation response varied from
0.17 Zu 5.43 μV across participants, as can be seen from
the histogram in Figure 4A. Regarding hemispheric later-
alization, 18 von 49 Teilnehmer (37%) showed left lateral-
ization (below the diagonal line on Figure 4B). Upon
closer inspection, the proportion of left-lateralized re-
sponses was larger for female (11/25 = 44%) verglichen
with male (7/24 = 29%) Teilnehmer, although this differ-
ence was not significant (χ2 = 1.16, p = .28). To take this
individual difference of hemispheric lateralization into
account for further analyses, we aggregated the face indi-
viduation responses over the left and right occipitotem-
poral channels.
Behavioral Data
The averaged accuracy in the CFMT task was 74.1% (95%
CI [69.7%, 78.5%]), with an accuracy of 77.6% (95% CI
[73.2%, 82.0%]) in the no-noise phase and 69.6% (95%
CI [64.2%, 75.0%]) in the noise phase. The internal reli-
ability (Cronbach’s alpha) for the CFMT based on the
sample of the current study was .89 (54 Artikel), mit
the reliabilities for the no-noise and noise phases of .86
(30 Artikel) Und .78 (24 Artikel), jeweils.
Correlations between EEG Amplitudes and
CFMT Performance
Pearson correlation coefficients between the general re-
sponse, the individuation response, and the CFMT per-
formance were calculated. The general response
measured from the medial occipital channels was not
correlated with the individuation response measured
1372
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 8
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
Ö
6
C
4
N
3
_
6
A
/
_
J
0
Ö
1
C
1
N
2
6
_
A
P
_
D
0
1
B
1
j
2
G
6
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
/
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Figur 2. Baseline-corrected
amplitude spectra showing
responses at the (A) allgemein
visual response (Grün) Und
(B) face individuation response
(orange) averaged across all
Kanäle. The horizontal axis
refers to response frequency
in Hertz, and the vertical axis
refers to the baseline-corrected
response amplitude in
microvolts. The scalp plot on
the top right side of each
graph shows that the scalp
distribution of the sum of the
baseline-corrected amplitude
at the harmonics contains
responses significantly larger
than the noise (z > 3.1).
from occipitotemporal channels (r = .15, p = .29, 95% CI
[−0.13, 0.41]). Darüber hinaus, there was no significant corre-
lation between the general response and the CFMT per-
Form (r = −.07, p = .65, 95% CI [−0.34, 0.25]).
Jedoch, the CFMT performance significantly correlated
with the aggregated individuation response from the left
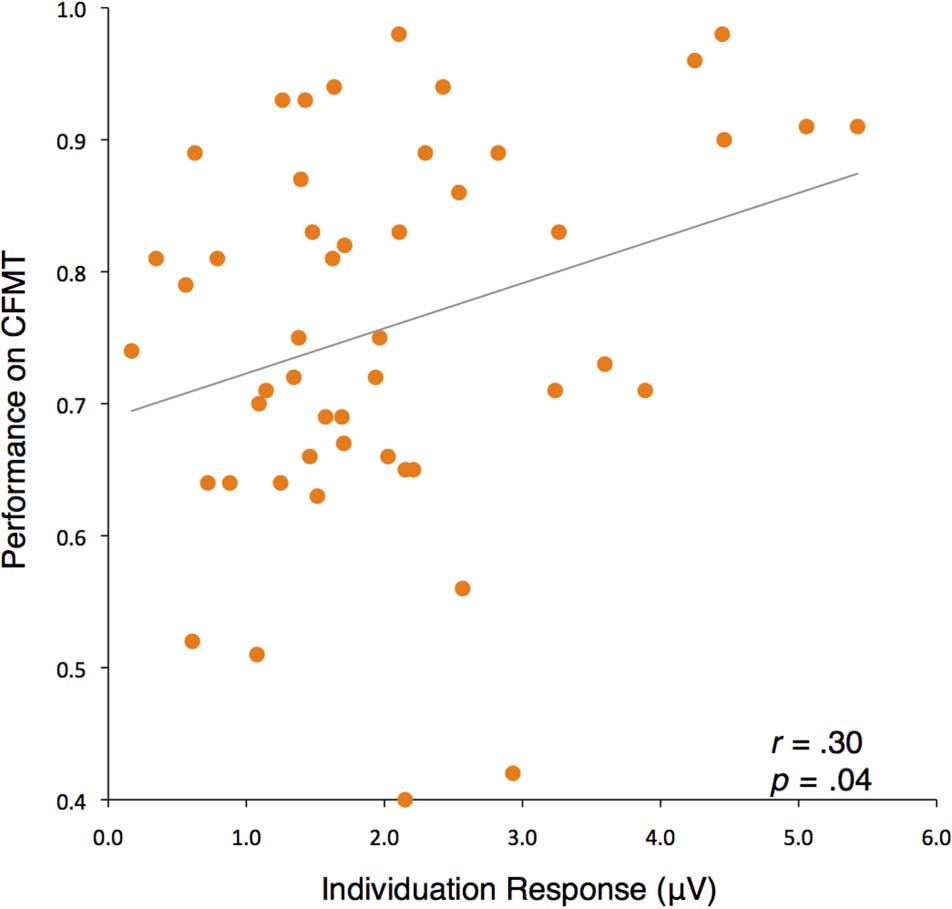
and right occipitotemporal channels (r = .30, p = .04,
95% CI [0.02, 0.54]; Figur 5).
DISKUSSION
The current study used an FPVS-EEG approach (Liu-
Shuang et al., 2014; Rossion & Boremanse, 2011) Zu
measure individual differences in face identity process-
ing. We presented participants with face stimuli at a fast
rate of 6 Hz (general base frequency rate), with face iden-
tity changes occurring every fifth face (1.2 Hz, individua-
tion frequency). Within only 4 min of EEG recording, Wir
found general responses at 6 Hz and harmonics and
lrobust face individuation responses at 1.2 Hz and har-
monics, thus replicating findings from previous studies
(Liu-Shuang et al., 2014, 2016; Dzhelyova & Rossion,
2014A, 2014B). The magnitude of the face individuation
response, but not of the general response, significantly
correlated with behavioral performance in the CFMT.
Although the FPVS-EEG approach has been used in a
number of studies to measure individualization of faces
(see Rossion, 2014, für eine Rezension; Liu-Shuang et al.,
2016, for the most recent study), it remained unknown
whether it was sensitive enough to elicit reliable brain re-
sponse from most individuals in a few minutes and
whether it was sensitive to the individual differences in
face identity processing in the normal population. Al-
though most of the previous studies used a relatively
small sample (z.B., 10–15 participants), the current study
recruited a larger sample of individuals (n = 49) Und
showed that we were able to obtain significant face indi-
viduation response in all but two participants using a con-
servative threshold (d.h., the signal in the frequency bin of
interest is significantly larger than the surrounding noise,
at the level of z > 3.1). Zusätzlich, this response was
consistently localized over the occipitotemporal channels
Xu et al.
1373
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
Ö
6
C
4
N
3
_
6
A
/
_
J
0
Ö
1
C
1
N
2
6
_
A
P
_
D
0
1
B
1
j
2
G
6
u
.
e
P
S
T
D
Ö
F
N
B
0
j
7
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
/
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
in most of the participants. Noch, our measures showed
significant individual differences in amplitude and lateral-
ization pattern.
Interessant, there was shared variance (9%) zwischen
individual EEG measures and behavioral performance on
the CFMT, although a number of factors contribute to
these measures. Trotzdem, the relatively weak corre-
lation coefficient suggests that the two measures reflect
different aspects of the face identity processing. It should
be emphasized that the weak correlation cannot be
attributed to low internal reliability of the measures.
The fidelity of the measures is crucial because it con-
strains the overall magnitude of their correlation ( Wilmer
et al., 2012; Schmidt & Hunter, 1996). In previous stud-
ies, it has been shown that the CFMT has high internal
consistency reliability (.86 < α < .90) in measuring face
recognition ability ( Wilmer et al., 2010), and this was rep-
licated in the current study (α = .89). The current study
also provided evidence, for the first time, that the FPVS-
EEG measure has high reliability (α = .87 for the individ-
uation response), despite the fact that it was obtained by
only four 60-sec sequences of recording. Most impres-
sively, different base faces were used in each stimulation
sequence, and none of the sequences were identical.
This high reliability may be attributed to the large num-
ber of variable individual face discrimination measures
(i.e., 72 in a 60-sec stimulation sequence) contributing
to the overall face individualization response in each
sequence.
Because both the CFMT and the FPVS measures had
high internal consistency reliability, the obtained correla-
tion between the EEG marker of facial identity and the
CMFT performance can be assumed to be robust and rel-
atively noise free. Thus, the modest size of the correlation
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
o
6
c
4
n
3
_
6
a
/
_
j
0
o
1
c
1
n
2
6
_
a
p
_
d
0
1
b
1
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 3. The individual scalp plots of the baseline-corrected amplitude of the individuation response summed from the significant harmonics
at the frequency bins of 1.2, 2.4, 3.6, 4.8, and 7.2 Hz. Each scalp plot is colored using 0 μV as the minimum and the largest magnitude of the
individuation response of that individual participant as the maximum, and hotter color stands for a larger individuation response.
1374
Journal of Cognitive Neuroscience
Volume 29, Number 8
Figure 4. Individual differences
in the lateralization and
amplitude of the EEG
individuation response. In
both graphs, data from male
participants are colored in
red, and data from female
participants are colored in
green. (A) Distribution of the
amplitude of the individuation
response of all participants.
The individuation response
was calculated by aggregating
the responses in both the
right and left occipitotemporal
channels. The horizontal axis
refers to the individuation
response bins, and the vertical
axis refers to the number of
participants falling into the
bins. (B) Lateralization of
face individuation responses.
The horizontal axis refers to the aggregated individuation response from channels at the left occipital temporal channel locations of P7, PO7,
and TP9. The vertical axis refers to the aggregated individuation response from channels at the right occipital temporal channel locations of P8, PO8,
and TP10. The diagonal line refers to the when the response is equal in the left and right occipital temporal locations.
between these two measures can rather be explained by
them targeting different aspects of face identity pro-
cessing. On the one hand, the CFMT is an explicit and
cognitively complex memory task that tests memory for
face identity. Performance on the CFMT requires atten-
tion, memory (encoding, storage, and retrieval), and
decision-making that go beyond the mere perceptual dif-
ferentiation of face identity. Moreover, individual differ-
Figure 5. Scatter plots showing the relationship between the
performance on CFMT and the individuation response aggregated from
the left and right occipitotemporal regions.
ences in the speed of face identity recognition are not
considered in the CFMT measure, despite the fact that
it is an important aspect of individual differences in this
function ( Wilhelm et al., 2010). On the other hand, the
EEG individuation response captures face identity dis-
crimination processes, without using an explicit behav-
ioral task, involving posterior regions of the visual
extrastriate cortex, and is thus closer to isolating percep-
tual processes. Moreover, in FPVS, the visual system is put
under greater constraint such that face identity needs to
be extracted at a single glance. Thus, we speculate that
the shared variance between FPVS and CFMT indicates
their common operation of processing face identity,
whereas the unaccounted variance is likely due not only
to the additional cognitive operations that are necessary
for the CFMT task (decision-making strategies, memory,
attention, etc.) but also to potential irrelevant factors
contributing to the magnitude of the EEG responses
obtained with FPVS (skull thickness, orientation of the
sources due to cortical folding, etc.).
The current study is the first to study the correlation
between the neural responses obtained using FPVS and
behavioral measures in face processing. Future studies
should correlate the FPVS responses and the perfor-
mance of a behavior face perception task that requires
participants to process faces in a briefer time duration.
In addition, showing that the FPVS response correlates
more with behavioral performance with faces than non-
face object processing tasks can be another important
evidence for the validity of the FPVS paradigm. Moreover,
future studies could focus on isolating a “real” face iden-
tity processing response of the behavioral tasks, for in-
stance, by using the shared variance of multiple face
Xu et al.
1375
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
o
6
c
4
n
3
_
6
a
/
_
j
0
o
1
c
1
n
2
6
_
a
p
_
d
0
1
b
1
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
recognition tasks requiring different components of the
face processing system (Bruce & Young, 1986) and con-
sidering both speed and accuracy (e.g., Sommer et al.,
2013; Hildebrandt, Wilhelm, Schmiedek, Herzmann, &
Sommer, 2011; Hildebrandt et al., 2010; Wilhelm et al.,
2010). Aggregation of the responses from several mea-
surements can average out the noise in each measure-
ment and thus provide a clear measure of the cognitive
processes of interest (Rushton, Brainerd, & Pressley,
1983). For example, the latent variable approach em-
ployed in the studies by Herzmann et al. (2010) and
Kaltwasser et al. (2014) using structural equation model-
ing can be used to abstract from task-specific variance
and measurement error. Those techniques are likely to
provide a more valid estimation of brain–behavior rela-
tionships in face processing. Last but not least, in the
EEG measure, future study can further improve the valid-
ity of the face individuation response by regressing out
a weak individuation response to control stimuli such
as inverted faces (Liu-Shuang et al., 2014) in the FPVS.
The current findings have critical practical implica-
tions. FPVS is a promising new technique that reveals
individual differences in rapid individual face discrimi-
nation as indicated by a reliable, valid, objective, task-
independent, and direct measure of brain activity (EEG).
It provides opportunities to easily test large samples of
participants from various backgrounds without confound-
ing perceptual abilities with general cognitive abilities.
Moreover, responses to a same experiment can be easily
compared across development and between various neu-
ro-typical and neuro-atypical populations.
Acknowledgments
This research was supported by the Chinese Scholarship Coun-
cil fellowship awarded to Buyun Xu, a postdoctoral grant from
the Belgian National Foundation for Scientific Research (no.
1230016F) awarded to Joan Liu-Shuang; PDR T.0207.16 FNRS
and ERC facessvep 284025 grants awarded to Bruno Rossion;
and the Temporal Dynamics of Learning Center (NSF grant
no. SBE-0542013) and the National Sciences and Engineering
Research Council of Canada grants awarded to James Tanaka.
We thank Adriano Boremanse and Benvenuto Jacob for tech-
nique supports for setting up the toolboxes and computer for
the FPVS testing and Bonnie Heptonstall and Alison Campbell
for data collection. We also thank the action editor and two
anonymous reviewers for their helpful comments.
Reprint requests should be sent to Buyun Xu, Department of
Psychology, University of Victoria, Cornett A191, 3800 Finnerty Rd.,
Victoria, BC, Canada, or via e-mail: xubuyun@uvic.ca.
REFERENCES
Alonso-Prieto, E., Van Belle, G., Liu-Shuang, J., Norcia, A. M.,
& Rossion, B. (2013). The 6 Hz fundamental stimulation
frequency rate for individual face discrimination in the
right occipito-temporal cortex. Neuropsychologia, 51,
2863–2875.
Bate, S., Parris, B., Haslam, C., & Kay, J. (2010). Socio-emotional
functioning and face recognition ability in the normal
population. Personality and Individual Differences, 48,
239–242.
Behrmann, M., & Avidan, G. (2005). Congenital prosopagnosia:
Face-blind from birth. Trends in Cognitive Sciences, 9, 180–187.
Bentin, S., Allison, T., Puce, A., Perez, E., & McCarthy, G. (1996).
Electrophysiological studies of face perception in humans.
Journal of Cognitive Neuroscience, 8, 551–565.
Benton, A. L., & Van Allen, M. W. (1968). Impairment in facial
recognition in patients with cerebral disease. Cortex, 4, 344.
Bobak, A. K., Bennetts, R. J., Parris, B. A., Jansari, A., & Bate, S.
(2016). An in-depth cognitive examination of individuals with
superior face recognition skills. Cortex, 82, 48–62.
Bowles, D. C., McKone, E., Dawel, A., Duchaine, B., Palermo, R.,
Schmalzl, L., et al. (2009). Diagnosing prosopagnosia: Effects
of ageing, sex, and participant–stimulus ethnic match on
the Cambridge Face Memory Test and Cambridge Face
Perception Test. Cognitive Neuropsychology, 26, 423–455.
Bruce, V., & Young, A. (1986). Understanding face recognition.
British Journal of Psychology, 77, 305–327.
DeGutis, J., Wilmer, J., Mercado, R. J., & Cohan, S. (2013). Using
regression to measure holistic face processing reveals a
strong link with face recognition ability. Cognition, 126, 87–100.
Dennett, H. W., McKone, E., Edwards, M., & Susilo, T. (2012).
Face aftereffects predict individual differences in face
recognition ability. Psychological Science, 23, 1279–1287.
Diamond, R., & Carey, S. (1986). Why faces are and are not
special: An effect of expertise. Journal of Experimental
Psychology: General, 115, 107.
Duchaine, B. C., & Nakayama, K. (2004). Developmental
prosopagnosia and the Benton Facial Recognition Test.
Neurology, 62, 1219–1220.
Duchaine, B. C., & Nakayama, K. (2006a). Developmental
prosopagnosia: A window to content-specific face processing.
Current Opinion in Neurobiology, 16, 166–173.
Duchaine, B., & Nakayama, K. (2006b). The Cambridge Face
Memory Test: Results for neurologically intact individuals and
an investigation of its validity using inverted face stimuli and
prosopagnosic participants. Neuropsychologia, 44, 576–585.
Dzhelyova, M., & Rossion, B. (2014a). The effect of parametric
stimulus size variation on individual face discrimination
indexed by fast periodic visual stimulation. BMC
Neuroscience, 15, 87.
Dzhelyova, M., & Rossion, B. (2014b). Supra-additive
contribution of shape and surface information to individual
face discrimination as revealed by fast periodic visual
stimulation. Journal of Vision, 14, 15.
Galper, R. E. (1970). Recognition of faces in photographic
negative. Psychonomic Science, 19, 207–208.
Germine, L. T., Duchaine, B., & Nakayama, K. (2011). Where
cognitive development and aging meet: Face learning ability
peaks after age 30. Cognition, 118, 201–210.
Halliday, D. W., MacDonald, S. W., Scherf, S. K., & Tanaka, J. W.
(2014). A reciprocal model of face recognition and autistic
traits: Evidence from an individual differences perspective.
PLoS One, 10, e94013.
Heisz, J. J., Watter, S., & Shedden, J. M. (2006). Automatic face
identity encoding at the N170. Vision Research, 46, 4604–4614.
Herzmann, G., Kunina, O., Sommer, W., & Wilhelm, O. (2010).
Individual differences in face cognition: Brain–behavior
relationships. Journal of Cognitive Neuroscience, 22,
571–589.
Hildebrandt, A., Sommer, W., Herzmann, G., & Wilhelm, O.
(2010). Structural invariance and age-related performance
differences in face cognition. Psychology and Aging, 25, 794.
Hildebrandt, A., Wilhelm, O., Schmiedek, F., Herzmann, G., &
Sommer, W. (2011). On the specificity of face cognition
compared with general cognitive functioning across adult
age. Psychology and Aging, 26, 701.
1376
Journal of Cognitive Neuroscience
Volume 29, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
o
6
c
4
n
3
_
6
a
/
_
j
0
o
1
c
1
n
2
6
_
a
p
_
d
0
1
b
1
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Hsiao, J. H.-W., & Cottrell, G. (2008). Two fixations suffice in
face recognition. Psychological Science, 19, 998–1006.
Jacques, C., d’Arripe, O., & Rossion, B. (2007). The time course
of the inversion effect during individual face discrimination.
Journal of Vision, 7, 3.
Jasper, H. H. (1958). The ten twenty electrode system of the
international federation. Eletroencephalography and
Clinical Neurophysiology, 10, 371–375.
Kaltwasser, L., Hildebrandt, A., Recio, G., Wilhelm, O., &
Sommer, W. (2014). Neurocognitive mechanisms of individual
differences in face cognition: A replication and extension.
Cognitive, Affective & Behavioral Neuroscience, 14, 861–878.
Liu-Shuang, J., Norcia, A. M., & Rossion, B. (2014). An objective
index of individual face discrimination in the right occipito-
temporal cortex by means of fast periodic oddball
stimulation. Neuropsychologia, 52, 57–72.
Liu-Shuang, J., Torfs, K., & Rossion, B. (2016). An objective
electrophysiological marker of face individualisation
impairment in acquired prosopagnosia with fast periodic
visual stimulation. Neuropsychologia, 83, 100–113.
Mouraux, A., & Iannetti, G. D. (2008). Across-trial averaging
of event-related EEG responses and beyond. Magnetic
Resonance Imaging, 26, 1041–1054.
Norcia, A. M., Appelbaum, L. G., Ales, J. M., Cottereau, B. R., &
Rossion, B. (2015). The steady-state visual evoked potential
in vision research: A review. Journal of Vision, 15, 4.
Regan, D. (1966). Some characteristics of average steady-state
and transient responses evoked by modulated light.
Electroencephalography and Clinical Neurophysiology,
20, 238–248.
Regan, D. (1989). Human brain electrophysiology: Evoked
potentials and evoked magnetic fields in science and
medicine. New York: Elsevier.
Retter, T. L., & Rossion, B. (2016). Uncovering the neural
magnitude and spatio-temporal dynamics of natural image
categorization in a fast visual stream. Neuropsychologia,
91, 9–28.
Rhodes, G., Jeffery, L., Taylor, L., Hayward, W. G., & Ewing, L.
(2014). Individual differences in adaptive coding of face
identity are linked to individual differences in face
recognition ability. Journal of Experimental Psychology:
Human Perception and Performance, 40, 897.
Rossion, B. (2014). Understanding individual face
discrimination by means of fast periodic visual stimulation.
Experimental Brain Research, 232, 1599–1621.
Rossion, B., & Boremanse, A. (2011). Robust sensitivity to facial
revealed by steady-state visual-evoked potentials. Journal of
Vision, 11, 16.
Rossion, B., Caldara, R., Seghier, M., Schuller, A. M., Lazeyras, F.,
& Mayer, E. (2003). A network of occipito temporal
face sensitive areas besides the right middle fusiform gyrus
is necessary for normal face processing. Brain, 126,
2381–2395.
Rossion, B., & Jacques, C. (2011). The N170: Understanding the
time-course of face perception in the human brain. In S. Luck
& E. Kappenman (Eds.), The Oxford handbook of ERP
components (pp. 115–142). Oxford: Oxford University Press.
Rushton, J. P., Brainerd, C. J., & Pressley, M. (1983). Behavioral
development and construct validity: The principle of
aggregation. Psychological Bulletin, 94, 18.
Russell, R., Duchaine, B., & Nakayama, K. (2009). Super-
recognizers: People with extraordinary face recognition
ability. Psychonomic Bulletin & Review, 16, 252–257.
Schmidt, F. L., & Hunter, J. E. (1996). Measurement error in
psychological research: Lessons from 26 research scenarios.
Psychological Methods, 1, 199.
Sommer, W., Hildebrandt, A., Kunina-Habenicht, O., Schacht,
A., & Wilhelm, O. (2013). Sex differences in face cognition.
Acta Psychologica, 142, 62–73.
Srinivasan, R., Bibi, F. A., & Nunez, P. L. (2006). Steady-state
visual evoked potentials: Distributed local sources and
wave-like dynamics are sensitive to flicker frequency. Brain
Topography, 18, 167–187.
Tanaka, J. W. (2001). The entry point of face recognition:
Evidence for face expertise. Journal of Experimental
Psychology: General, 130, 534.
Wilhelm, O., Herzmann, G., Kunina, O., Danthiir, V., Schacht,
A., & Sommer, W. (2010). Individual differences in perceiving
and recognizing faces—One element of social cognition.
Journal of Personality and Social Psychology, 99, 530.
Willis, J., & Todorov, A. (2006). First impressions making up
your mind after a 100-ms exposure to a face. Psychological
Science, 17, 592–598.
Wilmer, J. B., Germine, L., Chabris, C. F., Chatterjee, G.,
Gerbasi, M., & Nakayama, K. (2012). Capturing specific
abilities as a window into human individuality: The example
of face recognition. Cognitive Neuropsychology, 29, 360–392.
Wilmer, J. B., Germine, L., Chabris, C. F., Chatterjee, G.,
Williams, M., Loken, E., et al. (2010). Human face recognition
ability is specific and highly heritable. Proceedings of the
National Academy of Sciences, U.S.A., 107, 5238–5241.
Yin, R. K. (1969). Looking at upside-down faces. Journal of
identity in the right human occipito-temporal cortex as
Experimental Psychology, 81, 141.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
8
2
9
1
/
3
8
6
/
8
1
1
3
9
6
5
8
3
/
3
1
2
7
6
8
o
6
c
4
n
3
_
6
a
/
_
j
0
o
1
c
1
n
2
6
_
a
p
_
d
0
1
b
1
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Xu et al.
1377