Independent Distractor Frequency and Age-of-Acquisition
Effects in Picture–Word Interference: fMRI Evidence
for Post-lexical and Lexical Accounts according
to Distractor Type
Greig I. de Zubicaray1, Michele Miozzo2, Kori Johnson1,
Niels O. Schiller3, and Katie L. McMahon1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
Abstrakt
■ In two fMRI experiments, participants named pictures with
superimposed distractors that were high or low in frequency or
varied in terms of age of acquisition. Pictures superimposed
with low-frequency words were named more slowly than those
superimposed with high-frequency words, and late-acquired
words interfered with picture naming to a greater extent than
early-acquired words. The distractor frequency effect (Experi-
ment 1) was associated with increased activity in left premotor
and posterior superior temporal cortices, consistent with the
operation of an articulatory response buffer and verbal self-
monitoring system. Umgekehrt, the distractor age-of-acquisition
Wirkung (Experiment 2) was associated with increased activity in
the left middle and posterior middle temporal cortex, konsis-
tent with the operation of lexical level processes such as lemma
and phonological word form retrieval. The spatially dissociated
patterns of activity across the two experiments indicate that dis-
tractor effects in picture–word interference may occur at lexical
or postlexical levels of processing in speech production. ■
EINFÜHRUNG
Forty years of psycholinguistic research have demon-
strated that saying a word, the most fundamental task in
Apropos, requires selecting from among a set of activated
word candidates (see Goldrick, 2007; Dell & Sullivan,
2004). Daher, if a speaker wants to say dog, other words
are activated in addition to the target word dog. To the
extent that multiple lexical candidates are activated, theo-
ries of word production need to identify the nature of
these candidates as well as the degree to which they inter-
fere with target word production.
To meet this challenge, researchers have developed
paradigms that permit introducing word competitors while
varying their characteristics (z.B., semantic, syntactic, oder
phonological). A widely used paradigm has been picture–
word interference (PWI; Rosinski, Golinkoff, & Kukish,
1975), in which participants named a picture of a target
object in the context of a superimposed distractor word
that they are instructed to ignore. An important empirical
observation in PWI is semantic interference (SI), the rela-
tive slowing of naming latencies to target pictures in the
context of categorically related compared with unrelated
distractors, Zum Beispiel, picture–word pairs like dog–fox.
1University of Queensland, 2University of Cambridge, 3Leiden
Universität
Informative results about the locus of distractor interfer-
ence were obtained by manipulating distractor frequency.
Results from several experiments converged in demon-
strating greater interference for low-frequency (LF) dis-
tractors than high-frequency (HF) distractors (Catling,
Dent, Johnston, & Balding, 2010; Dhooge & Hartsuiker,
2010; Miozzo & Caramazza, 2003). The distractor fre-
quency effect was obtained even with semantically un-
related picture–distractor pairs (z.B., superimposing the
HF distractor book and the LF distractor stool to the picture
Hund). Außerdem, when effects of distractor frequency
and SI were induced in the same experiments (Miozzo &
Caramazza, 2003), they did not interact and exhibited dif-
ferent time courses, indicating the two do not share the
same processing locus.
A general framework to explain the greater interference
of LF distractors presupposes limited processing capacity,
whereby word distractors are processed slightly ahead
of target pictures, causing a delay in target naming pro-
portional to the time needed to process the distractors
(Miozzo & Caramazza, 2003). Because processing time is
longer for LF than HF words, LF distractors would generate
greater interference, as indeed observed in the distractor
frequency effect. Jedoch, the locus of the processing
delay resulting in the distractor frequency effect has proven
more problematic to define. Two accounts have been
examined, hereafter referred to as “input” and “output”
© 2011 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 24:2, S. 482–495
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
Ö
7
C
4
N
5
_
8
A
/
_
J
0
Ö
0
C
1
N
4
1
_
A
P
_
D
0
0
B
1
j
4
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
.
F
T
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
accounts, jeweils (Mahon, Costa, Peterson, Vargas, &
Caramazza, 2007; Miozzo & Caramazza, 2003).
The “input” account locates the distractor frequency
effect at the level of the (spoken or orthographic) recog-
nition mechanisms leading to accessing word meaning.
Because LF words have lower resting activation levels than
HF words as is typically assumed in recognition models
(z.B., McClelland & Rumelhart, 1981; Morton, 1969; Aber
see Norris, 2006, for a critical discussion of this assump-
tion), then LF words should be recognized more slowly
and interfere comparatively more. The assumption that
HF words might be processed more quickly also receives
support from network modeling studies. Zum Beispiel, In
Steyvers and Tenenbaumʼs (2005) model of developing
semantic networks, word frequency influences the prob-
ability of connecting new nodes to existing nodes, resulting
in HF words having more central, highly connected nodes
that are more likely to be accessed first.
Im Gegensatz, the “output” account locates the distractor
frequency effect at the level of postlexical mechanisms lead-
ing to the assembly and execution of articulatory programs
(z.B., Mahon et al., 2007; Miozzo & Caramazza, 2003). Word
distractors are assumed to have a privileged relationship
with the articulators and enter an output buffer as phono-
logically well-formed responses. Außerdem, the speed at
which a response enters the output buffer is assumed to
be related to frequency and to influence the speed at which
the distractor can be excluded according to task-relevant
Kriterien. By entering the buffer faster, HF distractors are
excluded earlier, thus incurring shorter delays in picture
naming relative to LF distractors.
Discriminating between these alternative accounts has
implications for explanations of PWI and word production
theories alike. Zum Beispiel, if the “output” account were
correct, distractors would potentially interact with mul-
tiple levels of processing of word production, einschließlich
the level where phonological/articulatory information is
computed. Zum Beispiel, the SI effect could result from
categorically related distractors entering the buffer faster
because of semantic priming, with a decision mechanism
taking longer to exclude the distractor as it also satis-
fies some response relevant criteria (Mahon et al., 2007;
Miozzo & Caramazza, 2003). Somit, according to this ac-
zählen, distractors potentially influence prelexical semantic
processes as well as postlexical ones. Andererseits,
the distractor frequency effect appears problematic for
the lexical selection by competition (LSC) account that
proposes the time taken to select the target word depends
on the activation levels of the competing lexical nodes.
Entsprechend, the higher the activation level of compet-
ing words, the longer name selection takes (z.B., Levelt,
Roelofs, & Meyer, 1999; Starreveld & La Heij, 1996; Harley,
1993; Roelofs, 1992; La Heij, 1988). Because HF words
should be more strongly activated compared with LF
Wörter, this view incorrectly anticipates larger interference
for the former than the latter, unless additional assump-
tions are made (z.B., distractor blocking; Roelofs, 2005).
There are results seemingly favoring the “output” ac-
zählen. Zum Beispiel, the distractor frequency effect varied
as a function of distractor phonology, suggesting that this
is an effect occurring at the level where phonological/
articulatory information are processed in naming (Miozzo
& Caramazza, 2003). A further result was accrued by
Dhooge and Hartsuiker (2010; Experiment 2) by masking
written word distractors. They reasoned that if the out-
put account were correct, masking the distractor should
preclude the formation of a response in the articulatory
output buffer, thus eliminating the distractor frequency
Wirkung. Masking did eliminate the effect. Darüber hinaus, a di-
rect testing of the “input” account conducted by Miozzo
and Caramazza (2003; Experimente 2 Und 3) did not yield
results expected within this account. Althought distractor
interference should reduce when word recognition is facili-
tated (z.B., by repetition), it should increase if word recog-
nition is made more difficult (z.B., by cAsE aLtErNaTiOn).
Neither prediction was confirmed.
Each of the tests employed to examine the “input” and
“output” hypotheses relies on a complex series of assump-
tionen. Let us exemplify this point referring to case alter-
nation. This manipulation represents a valid test of the
“input” account if one assumes that (A) case alternation
slows ignored/unattended distractor processing and mod-
ulates the interference effect and (B) frequency effects in
word recognition and picture naming have the same locus
and should therefore interact. With respect to the first
assumption, case-alternated primes have no differential
impact on masked repetition priming (Forster, 1998) oder
on conventional priming at short SOAs (Lee, Honig, &
Lee, 2002), indicating a case alternation effect may only
appear for attended stimuli. With respect to the second
assumption, there is evidence suggesting that frequency
effects in word comprehension (reading) and spoken
word production are different, with frequency effects in
the former modality being relatively independent of se-
mantic processing while depending on the presence or
absence of semantic constraint in the latter (Gollan et al.,
2011). In brief, the complexity of assumptions underlying
the tests of “input” and “output” accounts would make
it desirable to acquire additional evidence to adjudicate
between the alternative proposals. We addressed this
issue from a novel perspective in the present investigation
by characterizing the neural correlates of the distractor
frequency effect.
A number of studies have adopted neuroimaging tech-
niques to provide converging evidence for the level(S) bei
which distractor effects occur in speech production (z.B.,
Righi, Blumstein, Mertus, & Worden, 2010; Bles & Jansma,
2008; Heim, Friederici, Schiller, Rüschemeyer, & Amunts,
2008; de Zubicaray, McMahon, Eastburn, & Pringle, 2006;
de Zubicaray, Wilson, McMahon, & Muthiah, 2001). Das
is because of the increasing realization that brain imag-
ing data represents an additional dependent variable of
relevance to spreading activation models (z.B., Goldrick,
2007; Dell & Sullivan, 2004). There is now a large literature
de Zubicaray et al.
483
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
Ö
7
C
4
N
5
_
8
A
/
_
J
0
Ö
0
C
1
N
4
1
_
A
P
_
D
0
0
B
1
j
4
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
F
.
/
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
relating brain activation data to stages of processing in
models of spoken word production (z.B., Acheson, Hamidi,
Binder, & Postle, 2011; Peeva et al., 2010; Schuhmann,
Schiller, Goebel, & Sack, 2009; Alario, Chainay, Lehericy, &
Cohen, 2006; Indefrey & Levelt, 2004). These studies have
identified roles for the midsection of the left middle tem-
poral gyrus in lexical semantic processing and the poste-
rior section of the middle and superior temporal gyri
( Wernickeʼs area) in phonological word form retrieval,
jeweils, within a predominantly left hemisphere cere-
bral network. During naming of depicted objects, die Zeit
course of activation in these two regions typically occurs
zwischen 150 Und 300 msec following initial visual object
recognition, with postlexical processes of syllabification
and phonetic encoding followed by articulation occurring
zwischen 300 Und 600 msec attributed to the posterior left
inferior pFC (Brocaʼs area) and premotor cortical areas,
jeweils (z.B., Acheson et al., 2011; Schuhmann et al.,
2009; Indefrey & Levelt, 2004). Folglich, these stud-
ies provide candidate brain regions for testing the “input”
and “output” accounts. Crucially, the two accounts make
contrasting predictions concerning the brain regions as-
sociated with the distractor frequency effect. Although the
“output” account anticipates activation related to distractor
frequency in brain regions supporting phonological and
articulatory processing, no such activation is anticipated by
the “input” account.
A second goal of the present investigation relates to the
nature of the distractor frequency effect. There was a con-
founding, in PWI studies, of distractor frequency with age
of acquisition (AoA; see Brysbaert & Neu, 2009; Brysbaert
& Ghyselinck, 2006). Frequency and AoA are highly in-
tercorrelated, although have also been demonstrated to
exert independent effects in reading, Apropos, and speech
comprehension (Brysbaert & Cortese, 2011; Cortese &
Khanna, 2007; Brysbaert & Ghyselinck, 2006). Zusätzlich,
a number of studies have provided evidence indicating
that frequency and AoA effects might have different loci
across a range of tasks (z.B., Catling & Johnston, 2009;
Dent, Johnston, & Humphreys, 2008). Further arguments
for the distinctiveness of AoA come from accounts attribut-
ing a processing advantage for early AoA words to network
plasticity—As the network develops, plasticity is reduced
resulting in less accessible representations for later ac-
quired words (z.B., Menenti & Burani, 2007; Lambon
Ralph & Ehsan, 2006; Ellis & Lambon Ralph, 2000). Consis-
tent with the growing body of evidence suggesting the
autonomy of AoA from frequency, Catling et al. (2010;
Experiment 2) were able to demonstrate a frequency-
independent distractor AoA effect in PWI, with late-acquired
words producing greater interference.
Parallel to differential effects demonstrated behaviorally,
partially distinct patterns of brain activation would possibly
emerge when investigating the fMRI correlates of fre-
quency and AoA. Behavioral data could thus help us to
identify possible candidate brain regions sensitive to AoA.
Belke, Brysbaert, Meyer, and Ghyselinck (2005) found an
interaction between the SI effect and AoA in blocked
cyclic naming, in which pictures are presented in categori-
cally homogeneous versus mixed contexts. They inter-
preted this result as indicating that the AoA effect occurs
at a lexical semantic (Lemma) level of processing (see also
Brysbaert & Ghyselinck, 2006). Of note, no interaction
between SI and frequency was found in the same task
(Santesteban, Costa, Pontin, & Navarrete, 2006), zu mit-
trasting pattern further suggesting the distinctiveness of
frequency and AoA effects. A number of studies have iden-
tified roles for the middle and posterior portions of the
left middle temporal gyrus (pMTG) in lexical semantic
Verarbeitung (z.B., Acheson et al., 2011; Peeva et al., 2010;
Indefrey & Levelt, 2004). These regions have demonstrated
increased activity in categorically related compared with
unrelated distractor conditions in fMRI studies of picture
Wir (z.B., de Zubicaray et al., 2001, 2006). In line with
the hypothesis that AoA is linked to lexical semantic pro-
Abschließen (Belke et al., 2005), it seems reasonable to anti-
cipate responsiveness of these regions to AoA.
The effect of distractor frequency and AoA in PWI were
tested in Experiments 1 Und 2, jeweils. We employed
sparse temporal sampling designs within two functional
MRI experiments, permitting overt naming responses to
be accurately recorded during scanning (z.B., Heim et al.,
2008; de Zubicaray et al., 2001, 2006).
EXPERIMENT 1
The goal of the first experiment was to determine the
locus of the distractor frequency effect, contrasting predic-
tions from input and output accounts. If the latter account
is correct, then we would expect to observe increased
activity for LF words in brain regions associated with artic-
ulation and control mechanisms. Reliable cerebral cor-
relates of postlexical stages of processing (syllabification,
phonetic encoding, and motor articulation) encompass
both left inferior frontal gyrus (IFG) and premotor cortices
(z.B., Peeva et al., 2010; Eickhoff, Heim, Zilles, & Amunts,
2009; Schuhmann et al., 2009; Tremblay & Gracco, 2009;
Alario et al., 2006; Indefrey & Levelt, 2004). Within the
premotor cortex, Alario et al. (2006) identified a rostro-
caudal gradient in the SMA corresponding to postlexical
Auswahl, phonetic encoding, and articulatory processes
during spoken word production (see also Peeva et al.,
2010). Tremblay and Gracco (2009) likewise recently
identified pre-SMA as playing a central role in response
selection of spoken words. Jedoch, as Dhooge and
Hartsuiker (2010) notiert, the output account does not
specify the nature of the control mechanism hypothesized
to operate on the articulatory buffer. They tentatively
proposed that the verbal self-monitoring system might be
responsible for this function, documenting a range of
experimental findings in support. In their meta-analysis,
Indefrey and Levelt (2004) ascribed the monitoring of
both internal and external speech to bilateral posterior
484
Zeitschrift für kognitive Neurowissenschaften
Volumen 24, Nummer 2
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
Ö
7
C
4
N
5
_
8
A
/
_
J
0
Ö
0
C
1
N
4
1
_
A
P
_
D
0
0
B
1
j
4
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
F
/
T
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
superior temporal gyri (pSTG), a finding supported by
more recent studies (see Price, 2010; z.B., Zheng, Munhall,
& Johnsrude, 2010). Daher, it seems reasonable to assume
that the premotor cortex (especially SMA), IFG, and pSTG
would show increased activity if the output account were
correct.
As noted in Introduction, the input account predicts
increased activity in the middle and posterior portions of
the left middle temporal gyrus and left posterior middle
and superior temporal gyri (pMTG/pSTG) as they have
been implicated in lexical processing generally (Lemma
selection and phonological word form retrieval, bzw-
aktiv; z.B., Acheson et al., 2011; Peeva et al., 2010; Indefrey
& Levelt, 2004) and during SI in PWI specifically (z.B.,
de Zubicaray et al., 2001, 2006).
in the center of each picture. Stimuli were back-projected
using a BenQ SL705X projector onto a screen that partici-
pants viewed through a mirror mounted on the head coil,
and subtended approximately 10° of visual angle when each
participant was positioned for imaging. A 30 db attenuating
headset was used to reduce gradient noise. Naming re-
sponses were recorded on digital audio files using a custom
positioned fiber-optic dual-channel noise-cancelling micro-
phone attached to the head coil (FOMRI-III, Optoacoustics
Ltd., Or-Yehuda, Israel; www.optoacoustics.com). Naming lat-
encies were determined automatically with a voice key soft-
ware custom written in Microsoft VisualBasic and verified
manually using Audacity software (audacity.sourceforge.
net) in case nonvocal noise triggered the voice key.
Methoden
Teilnehmer
Seventeen healthy volunteers (10 Frauen) with a mean
age of 22 Jahre (SD = 3.5 Jahre) performed the experi-
ment. All were undergraduate students of the University
of Queensland. All were right-handed and native English
speakers, with no history of neurological or psychiatric dis-
Befehl, substance dependence, or known hearing deficits.
All had normal or corrected-to-normal vision and gave in-
formed consent in accordance with the protocol approved
by the Medical Research Ethics Committee of the Univer-
sity of Queensland. They were reimbursed AUD$30 for
participating.
Materials
The materials were identical to those used by Catling et al.
(2010; Experiment 1). Forty-eight black-and-white line
drawings were selected from Snodgrass and Vanderwart
(1980). These were split evenly into early and late-acquired
picture sets that were matched on a range of linguistic vari-
ables following Barry, Hirsh, Johnston, and Williams (2001).
HF and LF distractors were matched on a range of lin-
guistic variables including AoA (for information about the
matching variables, see the appendix in Catling et al.,
2010). Each target picture was paired with an HF and an
LF word that did not share a semantic or phonological
relationship with it. Target pictures were also presented
without distractor words in a neutral condition to examine
a potentially independent effect of target picture AoA and
to determine the direction of distractor related activity in
the fMRI experiment.
A laptop PC running Microsoft VisualBasic and ExacTicks
(Ryle Design, Mt. Pleasant, Michigan) software was used to
show the picture and word stimuli and record vocal re-
sponses on digital audio files (sampling rate, 11 kHz). Line
drawings were presented in black on a luminous white
background, and the visual distractor words were shown
in black lowercase Times New Roman 18-point bold font
Verfahren
A PWI paradigm was employed. Participants were first
familiarized with the set of experimental pictures with the
appropriate label printed below. The size of the pictures,
including background, was approximately 10 cm wide by
10 cm high. Over two consecutive practice blocks they
were instructed to name the pictures as fast and as accu-
rately as possible. Erroneous naming responses were cor-
rected. In a final block, they viewed the pictures without
labels and were instructed to name the pictures per the
instructions above.
Two experimental blocks each comprising 72 Versuche
presented in pseudorandom order were then conducted
(48 target pictures presented in three conditions: neutral/
no distractor, HF distractor, and LF distractor). A short
break was permitted between the two blocks while a
structural image was acquired (see fMRI Acquisition be-
niedrig). Trial presentation was pseudorandomized across
participants using Mix software (van Casteren & Davis,
2006), such that two presentations of the same picture
were always interceded by at least five different pictures,
and trials from a given condition were presented no more
than twice in succession. Participants were instructed
to name the pictures as quickly and accurately as possible
while ignoring the distractor word. They were also in-
structed not to speak or move during image acquisition
Und, in the event of a naming error, not to correct their
response. Trial presentation involved the following se-
quence: A fixation point (+) was shown for 500 ms,
followed by the presentation of the superimposed target
and distractor for 750 ms. Intertrial interval was 15 Sek.
fMRI Acquisition
Scanning was performed using a Bruker Medspec (Erlangen,
Deutschland) 4T system equipped with a transverse electromag-
netic head coil for radiofrequency transmission and recep-
tion ( Vaughan et al., 2002). A point-spread function
mapping sequence was first acquired to correct geometric
distortions in the functional images (Zaitsev, Hennig, &
de Zubicaray et al.
485
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
Ö
7
C
4
N
5
_
8
A
/
_
J
0
Ö
0
C
1
N
4
1
_
A
P
_
D
0
0
B
1
j
4
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
T
/
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Speck, 2003). Functional images depicting BOLD contrast
were then acquired with a gradient-echo EPI sequence
optimized for both image quality and noise reduction
(64 × 64 Matrix, 36 axial slices, 3.5 mm in plane resolution,
Scheibendicke = 3.5 mm, effective repetition time [TR] =
15 Sek; Echozeit = 30 ms; flip angle = 90°; McMahon,
Pringle, Eastburn, & Maillet, 2004). Two blocks of 73 Bild
volumes were acquired using a sparse temporal sampling
sequence to capture the estimated peak BOLD signal re-
sponse to task-related neural activity (Eden, Joseph, Braun,
Braun, & Zeffiro, 1999; Elliott, Bowtell, & Morris, 1999).
For each trial, no field gradients were applied for a 4-sec
period of relative silence, allowing for stimulus presenta-
tion and the participantʼs overt verbal response. A single
image volume was then acquired within 3 Sek, approxi-
mately coincident with the trialʼs estimated peak BOLD
response. No field gradients were applied for an additional
8-sec period to allow the BOLD response to the gradient
noise to return to baseline (for a diagram of the imaging
protocol, siehe Abbildung 1 in de Zubicaray et al., 2001). Kopf
movement was limited by foam padding within the head
coil. A 3-D T1-weighted structural image was acquired be-
tween the two functional imaging runs using a magnetiza-
tion prepared rapid acquisition gradient-echo sequence
(MP-RAGE; 2563 Matrix; 0.9 mm3 voxels). Total imaging
time was approximately 50 min.
fMRI Data Preprocessing and Analysis
The fMRI data were preprocessed and analyzed using
statistical parametric mapping software (SPM8; Wellcome
Department of Imaging Neuroscience, Queen Square,
London, VEREINIGTES KÖNIGREICH.). The first volume in each fMRI block was
discarded, and the remaining images were realigned to
the first image of the first block using the INRIAlign tool-
box (Freire, Roche, & Mangin, 2002). A mean image was
generated from the realigned series and coregistered
to the T1-weighted image. The T1-weighted image was
next segmented using the “New Segment” procedure. Der
“DARTEL” toolbox (Aschenbrenner, 2007) was then employed
to create a custom group template from the segmented
gray and white matter images, and individual flow fields
were used to normalize the realigned fMRI volumes to
the Montreal Neurological Institute (MNI) atlas T1 tem-
plate. The images were resampled to 3 mm3 voxels and
smoothed with a 9-mm FWHM isotropic Gaussian kernel.
Global signal effects were then estimated and removed
using a voxel level linear model (Macey, Macey, Kumar, &
Harper, 2004).
We conducted a two-stage, mixed effects model statis-
tical analysis. Event types corresponding to distractor and
Fehler (see Behavioral Results below) conditions were
modeled as effects of interest with delta functions repre-
senting each onset and convolved with a basis function
consisting of a single finite impulse response with a win-
dow length corresponding to the TR. As the sparse image
sequence does not acquire BOLD time course informa-
tion, trials were not convolved with a conventional hemo-
dynamic response function (see Gracco, Tremblay, & Pike,
2005; Eden et al., 1999; Elliott et al., 1999). Hoch- and low-
pass filtering were not applied, because of the long TRs
involved and the use of detrending (Macey et al., 2004).
Linear contrasts were applied to each participantʼs param-
eter estimates at the fixed effects level, then entered in
a group level repeated measures ANOVA, in which co-
variance components were estimated using a restricted
maximum likelihood procedure to correct for nonspheric-
ität (Friston et al., 2002). Regions with significant main
effects and/or interactions were investigated with planned
t contrasts.
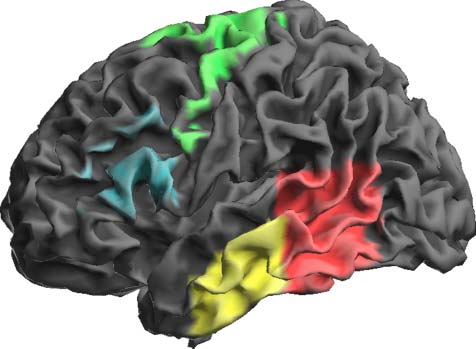
A priori ROIs (Figur 1) were defined using labeled
maximum likelihood gray matter maps from 3-D probabi-
listic atlases (Eickhoff et al., 2005; Hammers et al., 2003).
These were left mid-MTG and posterior temporal cortex
(Hammers et al., 2003) and left BA 44/45 (Brocaʼs area)
and premotor cortex (BA 6; Eickhoff et al., 2005). Letzteres
cytoarchitectonically defined region encompassed the
stereotactic MNI coordinates reported by both Tremblay
and Gracco (2009) and Alario et al. (2006) for their SMA
regions involved in postlexical selection, as well as the ven-
trolateral premotor area implicated in articulation (Peeva
et al., 2010; Indefrey & Levelt, 2004). A height threshold
of p < .005 was adopted in conjunction with a clus-
ter threshold of p < .05 estimated for the whole brain
(54 contiguous voxels) and for each ROI volume using
a Monte Carlo estimation procedure with 10,000 simula-
tions (alphasim, implemented in Analysis of Functional
NeuroImages toolkit, AFNI; National Institute of Mental
Health, Bethesda, MD). The height threshold of p < .005
threshold was adopted instead of the more commonly
used p < .001 to aid identification of possible overlap in
activated regions across experiments.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. Cerebral ROIs derived from maximum probability maps
(Eickhoff et al., 2005; Hammers et al. 2003) used in Experiments 1
and 2, rendered on a left hemisphere surface of an individual brain.
Middle temporal cortex (yellow), posterior temporal cortex (red),
premotor cortex (green), and Brocaʼs area (blue).
486
Journal of Cognitive Neuroscience
Volume 24, Number 2
Results
Behavioral Data
Trials scored as errors included incorrect or omitted
naming responses and dysfluencies (e.g., stuttering).
These were excluded from analysis (5.2%). Additional trials
were excluded in which naming onset RTs were <300
or>3000 ms (1.2%). Mean naming RTs as a function
of distractor condition are given in Table 1. Repeated mea-
sures ANOVAs were conducted for RTs with distractor
frequency and target picture AoA conditions with F1 treat-
ing participants as a random factor and F2 treating items
as a random factor. Because of the low error rates, diese
were not subjected to analysis.
There were significant main effects of target picture
AoA [F1(1, 16) = 6.92, MSE = 2283.15, P < .05, η2 =
0.30, and F2(1, 23) = 10.80, MSE = 5612.87, p < .005,
η2 = 0.32] and distractor frequency [F1(2, 32) = 28.87,
MSE = 2727.18, p < .001, η2 = 0.64, and F2(2, 46) =
37.04, MSE = 3215.37, p < .001, η2 = 0.62] and no sig-
nificant interaction (both Fs < 3). Paired samples t tests
showed a significant effect of distractor frequency [t1(16) =
2.8, p < .05 and t2(23) = 2.4, p < .05]. In summary, the re-
sults replicated those of Catling et al. (2010; Experiment 1)
with comparable RTs. Pictures with late-acquired names
were named more slowly than those with early-acquired
names, and the distractor frequency effect was confirmed;
pictures with LF distractors were named more slowly than
those with HF words.
fMRI Data
Data from a single participant were excluded from the
fMRI analyses because of excessive head movement during
image acquisition, defined as exceeding one voxel (3 mm)
within an imaging run. Group-averaged motion and rota-
tion parameters from the remaining 16 participants were
less than 1 mm and 1°, respectively, consistent with data re-
ported for sparse fMRI acquisitions in the literature (e.g.,
Gracco et al., 2005). A repeated measures ANOVA failed to
reveal any brain region demonstrating a significant main
effect of Target picture AoA or significant interaction be-
tween Target picture AoA and Distractor frequency, either
at the whole-brain level or in any of the a priori defined
ROIs, although it did reveal a significant main effect of
Distractor frequency. Therefore, the data were collapsed
across Target picture AoA for subsequent analyses. A
repeated measures ANOVA on the collapsed data (HF, LF
and, neutral conditions) revealed clusters showing a signifi-
cant main effect. These were subjected to post hoc planned
t contrasts.
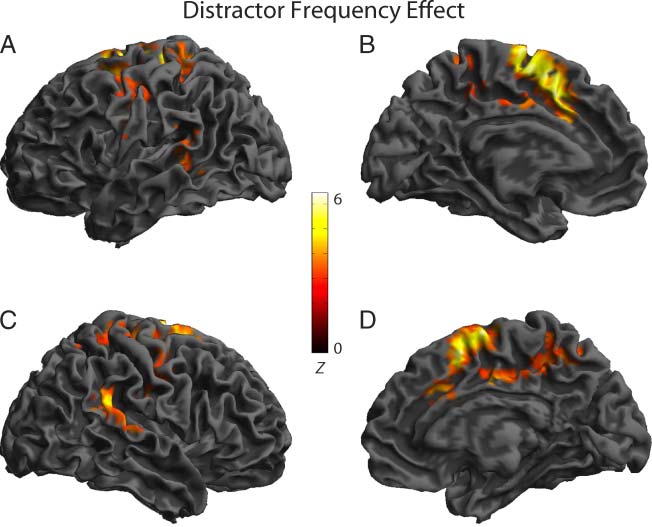
A planned contrast of LF > HF distractors revealed sig-
nificant activity at the whole-brain level in three large clus-
ters encompassing medial and lateral premotor, primary
sensorimotor and caudal anterior cingulate cortices bilater-
ally, left pSTG/pMTG and right supramarginal gyrus/pSTG.
ROI analyses revealed significant activity in the left me-
dial and lateral premotor cortex and the left pSTG/pMTG
(Tisch 2 and Figure 2). No other ROIs showed significant
Aktivität. Zusätzlich, no significant activity was observed for
the reverse contrast (HF > LF distractors) at the whole-
brain level or in the ROIs nor did contrasts of LF and HF
distractor conditions versus the neutral condition reveal
any significant activity.
Diskussion
The distractor frequency effect was replicated (Catling et al.,
2010, Experiment 1; Dhooge & Hartsuiker, 2010, Experi-
ment 1), confirming that the effect is robust. The fMRI
results may be interpreted as being consistent with an out-
put account (Mahon et al., 2007). Significantly increased
activity was observed in the medial and lateral premotor
cortex for LF versus HF distractor conditions, with a peak
corresponding to the boundary between the pre-SMA and
SMA. These regions have been associated with postlexical
selection and articulation by various researchers (Peeva
et al., 2010; Tremblay & Gracco, 2009; Alario et al., 2006;
Indefrey & Levelt, 2004). The absence of activity in the
IFG may be consistent with the notion that responses that
are already syllabified and phonetically encoded enter the
articulatory buffer (Mahon et al., 2007), as activity in this re-
gion precedes that in premotor and motor cortices during
speech production (Eickhoff et al., 2009). The increase in
activity in the left pSTG supports Dhooge and Hartsuikerʼs
(2010) proposal that the verbal self-monitoring system
may be responsible for checking the contents of the out-
put buffer and initiating the removal of inappropriate
responses. The absence of activity in the left mid-MTG indi-
cates that, contrary to the predictions of the input account,
Tisch 1. Naming Latencies for Experiment 1 (Distractor Frequency Effect)
Distractor Condition
Bilder
Low Frequency
High Frequency
Neutral (No Word Distractors)
Early
Late
Mean
1202 (356)
1215 (336)
1209 (345)
1175 (333)
1181 (357)
1178 (343)
Standard deviations are in parentheses.
1087 (323)
1142 (336)
1114 (328)
de Zubicaray et al.
487
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
Ö
7
C
4
N
5
_
8
A
/
_
J
0
Ö
0
C
1
N
4
1
_
A
P
_
D
0
0
B
1
j
4
G
1
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
ich
2
3
e
S
/
J
/
T
F
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Tisch 2. Cerebral Regions Showing Significant Activity in the Two Experiments
Peak MNI (x y z)
Z Score
Cluster Size ( Voxels)
LF > HF Words (Experiment 1)
Bilateral premotor, primary sensorimotor and
caudal anterior cingulate corticesa
Left pSTG/pMTGa
Right supramarginal gyrus/pSTGa
Left medial preSMA/SMAb
Left lateral premotor cortexb
Left posterior temporal cortexb
Late AoA > Early AoA Words (Experiment 2)
Left mid-MTG and pSTGa
Left inferior parietal lobule/angular gyrusa
Bilateral anterior cingulatea
Left posterior temporal cortexb
Left middle temporal cortexb
Late AoA Words > Neutral (Experiment 2)
Right mid-STG
Inferior parietal lobule and pSTG
Left mid-MTG and pMTG/STG
Left posterior temporal cortexb
3 12 51
−48 −42 3
57 −42 24
−3 9 51
−45 −12 51
48 −42 3
−51 −24 3
−42 −57 21
3 33 30
−42 −33 12
−42 −57 18
−54 −27 −6
42 −9 −3
60 −18 18
−51 −30 12
−51 −33 12
Neutral > Late AoA Words (Experiment 2)
Left parahippocampal gyrus
−12 −39 −6
Height threshold p < .005 and p < .05, cluster corrected.
aWhole-brain corrected.
bROI corrected.
4.64
3.13
4.0
4.6
3.12
3.13
4.4
3.61
3.68
3.29
3.27
3.27
4.0
3.42
3.6
3.51
3.6
1909
88
182
317
28
55
107
150
74
33
20
16
56
61
67
52
86
the activation levels of nontarget lexical (lemma) nodes are
unlikely to be responsible for the effect (e.g., Acheson et al.,
2011; Indefrey & Levelt, 2004; see Miozzo & Caramazza,
2003).
EXPERIMENT 2
The second experiment aimed to investigate whether the
effects of distractor frequency and AoA involve common or
distinct brain regions. According to the output account, the
activation levels of nontarget lexical nodes do not affect
selection (Mahon et al., 2007; also Miozzo & Caramazza,
2003). As neither early nor late-acquired words unrelated
to the target picture are relevant to the response required
by the task, this means the speed at which they enter the
buffer will be primarily responsible for the effect. As early-
acquired words are accessed first, they enter the buffer first
and are first to be excluded (e.g., Lambon Ralph & Ehsan,
2006; Steyvers & Tenenbaum, 2005). Hence, it seems rea-
sonable to presume that the output account predicts in-
creased activity for the late AoA distractor condition in
the same articulatory motor and verbal self-monitoring
regions (following Dhooge & Hartsuiker, 2010) predicted
for the distractor frequency effect in Experiment 1, namely,
premotor cortex (especially SMA), IFG, and pSTG. In addi-
tion, given the prior evidence afforded by Experiment 1,
we would expect similar patterns of activity to be involved.
The candidate regions for the input account are essentially
the same as those proposed for the distractor frequency
effect in Experiment 1, namely, left middle and posterior
temporal cortex, if the effect has a lexical semantic locus
as proposed by Belke et al. (2005).
488
Journal of Cognitive Neuroscience
Volume 24, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Cerebral regions
showing significantly increased
activity in Experiment 1 for
the independent distractor
frequency effect (LF > HF
Wörter) in the whole-brain
analysis superimposed on
the surface rendering of
an individual brain (height
thresholded at p < .005 and
cluster thresholded at p < .05):
(A) left hemisphere lateral view,
(B) left hemisphere medial view,
(C) right hemisphere lateral
view, and (D) right hemisphere
medial view.
Methods
Participants
Results
Behavioral Data
Seventeen healthy volunteers (11 women) with a mean
age of 22 years (SD = 4.75 years) performed the ex-
periment. None had participated in Experiment 1 and
met the criteria described in the Participants section of
Experiment 1.
Materials
Materials were identical to those in Catling et al. (2010;
Experiment 2). The same pictures as Experiment 1 were
employed, although early and late AoA distractors (again
matched on a range of linguistic variables including fre-
quency estimates from various corpora; see Catling et al.,
2010; Appendix) were used. Each target picture was paired
with an early AoA and late AoA word that did not share a
semantic or phonological relationship with it. Target pic-
tures were again presented without distractor words in
a neutral condition to examine any effect of target picture
AoA and to determine the direction of distractor related
activity in the fMRI experiment.
Procedure
The procedure was identical to that in Experiment 1.
fMRI Acquisition, Data Preprocessing, and Analysis
Scoring criteria were identical to Experiment 1. Error trials
were excluded from analysis (1.8%), as were RT outliers
(0.1%). Mean naming RTs as a function of distractor con-
dition are given in Table 3. Repeated measures ANOVAs
were conducted for RTs with Distractor AoA and Target
picture AoA conditions with F1 treating participants as a
random factor and F2 treating items as a random factor.
Because of the low error rates, these were not subjected
to analysis.
These revealed significant main effects of Target pic-
ture AoA [F1(1, 16) = 11.22, MSE = 1937.54, p < .005,
η2 = 0.41, and F2(1, 23) = 9.65, MSE = 9837.25, p < .01,
η2 = 0.29] and Distractor AoA [F1(2, 32) = 58.64, MSE =
1964.64, p < .001, η2 = 0.86, and F2(2, 46) = 47.09,
MSE = 2853.46, p < .001, η2 = .67] and no significant
interaction (both Fs < 1). Paired samples t tests showed a
Table 3. Naming Latencies for Experiment 2 (Distractor
AoA Effect)
Distractor Condition
Pictures
Early AoA
Late AoA
Neutral
(No Word Distractors)
Early
Late
Mean
1166 (261)
1225 (279)
1093 (3248)
1196 (313)
1239 (238)
1138 (268)
1180 (251)
1231 (258)
1116 (256)
These were identical to those in Experiment 1.
Standard deviations in parentheses.
de Zubicaray et al.
489
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
significant effect of Distractor AoA [t1(16) = 5.0, p < .001
and t2(23) = 3.0, p < .01]. In summary, the results repli-
cated those of Catling et al. (2010; Experiment 2) with
comparable RTs: Pictures with late-acquired names were
named more slowly than those with early-acquired names,
and the independent distractor AoA effect was confirmed:
Pictures with late-acquired distractors were named more
slowly than those with early-acquired words.
fMRI Data
Data from two participants were excluded from the fMRI
analyses because of excessive head movement based on
identical criteria to Experiment 1. Group-averaged motion
and rotation parameters from the remaining 15 partici-
pants were again less than 1 mm and 1°, respectively. A
repeated measures ANOVA failed to reveal any brain region
demonstrating a significant main effect of Target picture
AoA or significant interaction between Target picture AoA
and Distractor AoA either at the whole-brain level or in
any of the a priori defined ROIs, although it did reveal a
significant main effect of Distractor AoA. Therefore, the
data were collapsed across target picture AoA for sub-
sequent analyses. A repeated measures ANOVA on the col-
lapsed data revealed clusters showing a significant main
effect of Distractor (early, late, and neutral). These were
subjected to post hoc planned t contrasts.
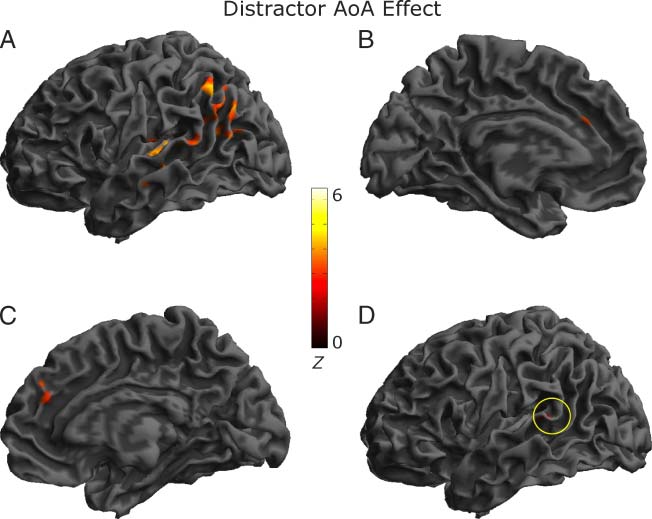
A planned contrast of Late > Early AoA distractors re-
vealed significant activity at the whole-brain level in three
clusters encompassing the left mid- MTG and pSTG, links
inferior parietal lobule/angular gyrus and bilateral ante-
rior cingulate (Figur 3). ROI analyses revealed significant
activity in two regions of left pSTG/MTG and left mid-
MTG (Tisch 2). No other ROIs showed significant activ-
ität. No significant activity was observed for the reverse
Kontrast (Early > Late AoA distractors) at the whole-brain
level or in the ROIs.
Zusätzlich, significant activity was observed at the whole-
brain level for the contrast of the Late AoA distractor >
Neutral conditions in the right mid-STG, inferior parietal
lobule and pSTG and left mid-MTG and pMTG/STG (Feige-
ure 2). ROI analyses also revealed significant activity in the
left pMTG/pSTG. No significant activity was detected in
the remaining ROIs. The reverse contrast (Neutral > Late
AoA) revealed a single cluster in the left parahippocampal
gyrus significant at the whole-brain level (Tisch 2). No sig-
nificant activity was observed in any of the ROI analyses
nor was any significant activity observed for the early AoA
distractor versus neutral contrasts either at the whole-brain
level or in ROI analyses.
Diskussion
The independent distractor AoA effect reported by Catling
et al. (2010; Experiment 2) was replicated. Significantly
increased activity was observed in the left mid-MTG and
pMTG/STG regions consistent with a role for these regions
in lexical selection and phonological word form retrieval
(z.B., Acheson et al., 2011; Peeva et al., 2010; Indefrey &
Levelt, 2004). The activity in the left pSTG might also be
considered consistent with the operation of the verbal
self-monitoring system, as Indefrey and Levelt (2004) Profi-
posed the region serves as a common store of lexical word
form representations for both production and perception.
We therefore calculated the overlap in activated voxels
between the distractor frequency (Experiment 1) Und
Figur 3. Cerebral regions
showing significantly increased
activity in Experiment 2 für
the independent distractor
AoA effect (Late > Early words)
in the whole-brain analysis
superimposed on the surface
rendering of an individual brain
(height thresholded at p < .005
and cluster thresholded at
p < .05): (A) left hemisphere
lateral view, (B) left hemisphere
medial view, (C) right
hemisphere medial view,
and (D) voxels overlapping
between the distractor
frequency (Experiment 1)
and AoA effects (circled).
490
Journal of Cognitive Neuroscience
Volume 24, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
AoA effects via inclusively masking the former with the
latter result. This revealed only 18 of 107 (or 17%) voxels
overlapped solely in the posterior portion of the left pSTG,
indicating that while the two effects share activity in pos-
terior pSTG that might be sensitive to the self monitoring
demands of the task, the distractor AoA effect is associ-
ated with additional activity in both middle and posterior
MTG consistent with the operation of lexical level pro-
cesses (Figure 3). More importantly, the absence of sig-
nificant activity in the premotor cortex or IFG precludes
an interpretation of a postlexical mechanism operating at
the level of an articulatory response buffer (Mahon et al.,
2007). This contrasts with the extensive activity observed
in the premotor cortex in Experiment 1, indicating that
the independent distractor frequency and AoA effects
involve different mechanisms.
GENERAL DISCUSSION
This article is the first to report fMRI data for the distractor
frequency and AoA effects in PWI. The results indicate
quite clearly that the two effects do not involve identical
processing mechanisms based on the observed spatial dis-
sociations in cortical activity across the two experiments.
Although hypotheses from two rival accounts of distrac-
tor interference in PWI were supported in each experi-
ment, neither account is capable of providing a complete
explanation. Rather, our data indicate that word distrac-
tors may affect each level of processing proposed by the
two accounts.
The Distractor Frequency Effect
Miozzo and Caramazza (2003) questioned the conventional
LSC account of word production based on a series of in-
vestigations of the distractor frequency effect. Subsequent
studies confirmed their basic findings by controlling for
the potentially confounding factor of AoA (Catling et al.,
2010, Experiment 1; Dhooge & Hartsuiker, 2010, Experi-
ment 1). The behavioral data from Experiment 1 replicated
Catling et al.ʼs results, confirming the robustness of the ef-
fect. In Experiment 1, the independent distractor frequency
effect in PWI was associated with significant increases in
activity in a series of cortical regions associated previously
with articulatory motor and verbal self-monitoring mecha-
nisms in spoken word production, including the medial
(SMA/pre-SMA) and lateral premotor cortex and left pSTG
(e.g., Peeva et al., 2010; Tremblay & Gracco, 2009; Alario
et al., 2006; Indefrey & Levelt, 2004). The premotor cortex
activity, in particular, may be considered consistent with
the output hypothesis, as it attributes the locus of the effect
to a postlexical articulatory response buffer (Mahon et al.,
2007). The activation observed in primary motor cortex
is also consistent with Peeva et al.ʼs (2010) recent pro-
posal that projections from medial and lateral premotor
cortices to primary motor cortex transform sublexical rep-
resentations into a set of motor commands to the speech
articulators.
It is difficult to reconcile the premotor cortex result
with an input account. Indefrey and Leveltʼs (2004) meta-
analysis of the time course of spoken word production
indicated premotor cortex activity occurs subsequent to
lexical (lemma and word form) processing in middle
and posterior temporal cortical areas. Nor do the results
appear to support Roelofsʼ (2005) modified LSC mech-
anism implemented in WEAVER++. In Roelofsʼ (2005)
WEAVER++ account, a selective attention mechanism re-
actively blocks distractor processing, favoring the produc-
tion of the target name. The speed of blocking depends
on the speed with which the distractor information be-
comes available. As HF words are accessed first (e.g.,
Steyvers & Tenenbaum, 2005), they are first to be blocked.
Roelofs, Piai, and Schriefers (2011) were able to demon-
strate that WEAVER++ accounts for the distractor fre-
quency effect and the results of Dhooge and Hartsuikerʼs
(2010) masking and SOA manipulations. However, the
WEAVER++ model still entails the assumption that the
activation levels of the lexical nodes of LF words are higher
once processing of HF words is reactively blocked, leading
to a prediction of increased activity in middle and poste-
rior MTG/STG. Roelofs (2008) associated the attentional
control mechanism in WEAVER++ with the anterior cingu-
late and IFG. The former region showed increased activity
as part of a larger cluster involving premotor and sensori-
motor regions, although this activity was located in the
cingulate motor area, indicating a more likely motor role.
The left IFG/Brocaʼs area did not show significant activity,
either at the whole-brain level or in an ROI analysis.
As Dhooge and Hartsuiker (2010) noted, the output ac-
count does not specify the nature of the control mecha-
nisms operating on the articulatory response buffer. They
proposed that this role is performed by the verbal self-
monitoring system. The finding of increased activity in
the pSTG is consistent with this proposal, given evidence
linking verbal self-monitoring to this region (Price, 2010;
Zheng et al., 2010; Indefrey & Levelt, 2004). Precisely
how the self-monitoring system might exclude responses
from the articulatory buffer remains to be specified (see
Roelofs et al., 2011). Another issue requiring specification
relates to the role in PWI played by SMA/pre-SMA, regions
sensitive to distractor frequency.
The Distractor AoA Effect
Experiment 2 replicated the independent distractor AoA
effect reported by Catling et al. (2010; Experiment 2), indi-
cating it is a robust effect. The direction of the AoA effect
(larger interference/activation for late vs. early words) is
in the opposite direction to that predicted by an LSC ac-
count operating under a typical assumption of lower rest-
ing activation for late-acquired words (e.g., Meschyan &
Hernandez, 2002). However, network modeling studies
indicate that early-acquired words are accessed relatively
de Zubicaray et al.
491
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
faster because they have more central, highly connected
nodes (Steyvers & Tenenbaum, 2005), possibly as a result
of network structuring through development (e.g., Menenti
& Burani, 2007; Lambon Ralph & Ehsan, 2006; Ellis &
Lambon Ralph, 2000). The AoA effect was associated in our
experiment with increases in activity in left mid-MTG/STG
and pMTG/STG, in addition to the anterior cingulate. The
former regions are related to lexical level processes, in-
cluding lemma and phonological word form retrieval
(e.g., Acheson et al., 2011; Peeva et al., 2010; Indefrey &
Levelt, 2004), supporting an input account in which repre-
sentations of early-acquired words are accessed first (e.g.,
Lambon Ralph & Ehsan, 2006; Steyvers & Tenenbaum,
2005). Of note, these regions have demonstrated increased
activity in categorically related compared with unrelated
distractor conditions in fMRI studies of SI effects (e.g., de
Zubicaray et al., 2001, 2006). Thus, these findings converge
with the interaction between AoA and SI observed by
Belke et al. (2005), a result interpreted by them as indi-
cating independent AoA effects occur at a lexical (lemma)
level of processing.
The ACC activity for the independent distractor AoA
effect is consistent with that observed previously for the
SI effect in PWI (e.g., de Zubicaray et al., 2001, 2006). This
activity was rostral to the cingulate motor area activity
observed for the distractor frequency effect. A prevailing
account of ACC activity during interference tasks is that
it reflects a general competition or conflict monitoring
mechanism (e.g., Botvinick, Cohen, & Carter, 2004). On
the basis of an assumption of competition among rival
lexical nodes for production, a “competition detector”
could monitor the total amount of activation of items at
a given level of processing and respond with an error
signal if a particular threshold is exceeded (Hartsuiker,
2006). This type of monitor was proposed by de Zubicaray
and colleagues (2001, 2006) based upon results from their
fMRI studies of SI effects, and appended to Harleyʼs (1993)
connectionist computational model in a successful simula-
tion of the SI effect (Hockey, Wiles, & de Zubicaray, 2005).
Other neuroimaging studies have reached similar conclu-
sions concerning the monitoring role of ACC in speech
production (e.g., Riès, Janssen, Dufau, Alario, & Burle,
2011; Christoffels, Formisano, & Schiller, 2007). Alterna-
tively, the activity might also be considered consistent with
Roelofsʼ (2008) LSC account in which ACC performs a role
in attentional control during speech production. The ad-
ditional activity observed in the left inferior parietal lobule
appears consistent with some role in distractor process-
ing, given the extensive research linking this region with
attention and reading (see Shaywitz & Shaywitz, 2008)
and its increased response in the distractor versus neutral
condition.
Implications for Input and Output Accounts
The contrasting patterns of activity observed for the inde-
pendent distractor frequency and AoA effects indicate that
neither input nor output accounts provide complete ex-
planations of these PWI effects. A number of recent studies
have challenged the reliability and generality of results
cited in support of the latter account (e.g., Janssen, Schirm,
Mahon, & Caramazza, 2008; cf. Mädebach, Oppermann,
Hantsch, Curda, & Jescheniak, 2011; Piai, Roelofs, &
Schriefers, 2011; also Mahon et al., 2007; cf. Lee &
de Zubicaray, 2010). However, an output account clearly
provides a better explanation of the premotor activity as-
sociated with the independent distractor frequency effect
than an input account, while the latter provides a better
account of the middle to posterior MTG/STG activity ob-
served in conjunction with the distractor AoA and SI ef-
fects. A way to reconcile our data to these accounts is to
propose that word distractors may affect the processes
implicated by each account.
The results of Experiment 1 converge with other find-
ings from neuroimaging studies indicating that distractor
effects may occur at multiple levels of information pro-
cessing in word production. The locus of the SI effect in
the postcue naming task has been variously attributed to
prelexical, lexical, and postlexical mechanisms (Mahon
et al., 2007; Dean, Bub, & Masson, 2001; Humphreys,
Lloyd-Jones, & Fias, 1995). In that task, participants view
two differently colored superimposed pictures, naming
the target according to a subsequently presented color
cue. When the distractor and target pictures are categori-
cally related, naming latencies are slower than when they
are unrelated. Dean et al. (2001) proposed that this in-
terference occurs when demands on the integration of
perceptual attributes (color) and conceptual representa-
tions are high because of feature overlap among category
exemplars. Recent fMRI evidence supports this proposal
(Hocking, McMahon, & de Zubicaray, 2010).
AoA Effects in Target Picture Names
Although main effects of target picture AoA were observed
in the naming latencies in both experiments, target pic-
ture AoA did not interact with the independent distractor
frequency or AoA effects, replicating the results of Catling
et al. (2010). Nor did target picture AoA produce signifi-
cant changes in fMRI activity. The absence of an interaction
is consistent with research indicating different processing
mechanisms for AoA effects according to task (Catling &
Johnston, 2009; Brysbaert & Ghyselinck, 2006). However,
in an fMRI study of covert picture naming, Ellis, Burani,
Izura, Bromiley, and Venneri (2006) reported increased
activation for pictures with late-acquired names in the
left middle occipital and fusiform gyri, a result they in-
terpreted as reflecting mapping of visual onto semantic
representations.
To investigate target picture AoA effects more directly
in our data, we performed a post hoc analysis involving
a direct t contrast of early and late target pictures in the
neutral (i.e., no distractor) condition, combining data
from participants across both experiments (n = 31). This
492
Journal of Cognitive Neuroscience
Volume 24, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
analysis likewise failed to reveal any significant activity.
As Ellis et al. (2006) employed a blocked experimental
design, it is possible that their findings represent the
strategic operation of a cognitive search mechanism like
that envisaged by Steyvers and Tenenbaum (2005). Alter-
natively, an absence of differential activity might also be
considered consistent with the architecture of neural net-
work models (e.g., Lambon Ralph & Ehsan, 2006; Zevin
& Seidenberg, 2002). According to these models, AoA
effects in picture naming are because of the arbitrary map-
pings between objects and their names (i.e., there is no
systematic relationship between semantic input and pho-
nological output representations). Although these effects
emerge in networks with distributed random representa-
tions, they do not emerge in networks with localist ones
(Zevin & Seidenberg, 2002). Hence, one would not neces-
sarily expect to observe localized activity attributable to a
specific level of processing in an fMRI experiment. This dif-
fers from explanations that attribute AoA effects with word
distractors to a lexical semantic level of processing (e.g.,
Belke et al., 2005; see also Brysbaert & Ghyselinck, 2006).
Summary and Conclusions
The two fMRI experiments reported here each provides
some support for input and output accounts of the dis-
tractor frequency and AoA effects, respectively. However,
given the different patterns of activity observed, neither
account is able to provide a complete explanation. The
fMRI data show the former effect engages regions involved
in postlexical processes, including articulation and verbal
self-monitoring, while the latter effect engages regions
linked with lexical level processes. These interpretations
should, nevertheless, be considered within the limitations
of inference afforded by neuroimaging, as fMRI data can-
not provide direct evidence of relationships between pro-
cesses and processors in speech production.
Acknowledgments
We thank Jonathan Catling for providing the distractor stimuli for
Experiments 1 and 2. This research was supported by Australian
Research Council (ARC) Discovery Project grant DP1092619.
Greig de Zubicaray was supported by ARC Future Fellowship
FT0991634. Niels O. Schiller was supported by the Netherlands
Institute for Advanced Study in the Humanities and Social Sci-
ences as a Fellow-in-Residence 2010/2011. Michele Miozzo was
supported by NIH grant DC006242.
Reprint requests should be sent to Greig I. de Zubicaray, School
of Psychology, University of Queensland, Brisbane, QLD 4072,
Australia, or via e-mail: greig.dezubicaray@uq.edu.au.
REFERENCES
Acheson, D. J., Hamidi, M., Binder, J. R., & Postle, B. R.
(2011). A common neural substrate for language
production and verbal working memory. Journal
of Cognitive Neuroscience, 23, 1358–1367.
Alario, F. X., Chainay, H., Lehericy, S., & Cohen, L. (2006).
The role of the supplementary motor area (SMA) in
word production. Brain Research, 1076, 129–143.
Ashburner, J. (2007). A fast diffeomorphic image registration
algorithm. Neuroimage, 38, 95–113.
Barry, C., Hirsh, K. W., Johnston, R. A., & Williams, C. L.
(2001). Age of acquisition, word frequency, and the
locus of repetition priming of picture naming. Journal
of Memory and Language, 44, 350–375.
Belke, E., Brysbaert, M., Meyer, A. S., & Ghyselinck, M.
(2005). Age of acquisition effects in picture naming:
Evidence for a lexical semantic competition hypothesis.
Cognition, 96, B45–B54.
Bles, M., & Jansma, B. M. (2008). Phonological processing
of ignored distractor pictures, an fMRI investigation.
BMC Neuroscience, 9, 20.
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004).
Conflict monitoring and anterior cingulate cortex:
An update. Trends in Cognitive Sciences, 8, 539–546.
Brysbaert, M., & Cortese, M. J. (2011). Do the effects of
subjective frequency and age of acquisition survive
better word frequency norms? Quarterly Journal of
Experimental Psychology, 64, 545–559.
Brysbaert, M., & Ghyselinck, M. (2006). The effect of age of
acquisition: Partly frequency related, partly independent.
Visual Cognition, 12, 992–1011.
Brysbaert, M., & New, B. (2009). Moving beyond Kucera
and Francis: A critical evaluation of current word frequency
norms and the introduction of a new and improved word
frequency measure for American English. Behavior Research
Methods, 41, 977–990.
Catling, J. C., Dent, K., Johnston, R. A., & Balding, R. (2010).
Age of acquisition, word frequency, and picture–word
interference. Quarterly Journal of Experimental
Psychology, 63, 1304–1317.
Catling, J. C., & Johnston, R. A. (2009). The varying effects
of age of acquisition. Quarterly Journal of Experimental
Psychology, 62, 50–62.
Christoffels, I. K., Formisano, E., & Schiller, N. O. (2007).
Neural correlates of verbal feedback processing: An fMRI
study employing overt speech. Human Brain Mapping,
28, 868–879.
Cortese, M. J., & Khanna, M. M. (2007). Age of acquisition
predicts naming and lexical-decision performance above
and beyond 22 other predictor variables: An analysis
of 2,342 words. Quarterly Journal of Experimental
Psychology, 60, 1072–1082.
de Zubicaray, G. I., McMahon, K. L., Eastburn, M. M., &
Pringle, A. (2006). Top–down influences on lexical
selection during spoken word production: A 4T fMRI
investigation of refractory effects in picture naming.
Human Brain Mapping, 27, 864–873.
de Zubicaray, G. I., Wilson, S. J., McMahon, K. L., & Muthiah, S.
(2001). The semantic interference effect in the picture–word
paradigm: An event-related fMRI study employing overt
responses. Human Brain Mapping, 14, 218–227.
Dean, M. P., Bub, D. N., & Masson, M. E. (2001). Interference
from related items in object identification. Journal of
Experimental Psychology: Learning, Memory, and
Cognition, 27, 733–743.
Dell, G. S., & Sullivan, J. M. (2004). Speech errors and
language production: Neuropsychological and connectionist
perspectives. In B. H. Ross (Ed.), The psychology of learning
and motivation (pp. 63–108). San Diego, CA: Elsevier.
Dent, K., Johnston, R. A., & Humphreys, G. W. (2008). Age of
acquisition and word frequency effects in picture naming:
A dual task investigation. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 34, 282–301.
de Zubicaray et al.
493
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Dhooge, E., & Hartsuiker, R. J. (2010). The distractor frequency
effect in picture–word interference: Evidence for response
exclusion. Journal of Experimental Psychology: Learning,
Memory, and Cognition, 36, 878–891.
Eden, G. F., Joseph, J. E., Brown, H. E., Brown, C. P., & Zeffiro,
T. A. (1999). Utilizing hemodynamic delay and dispersion
to detect fMRI signal change without auditory interference:
The behavioral interleaved gradients technique. Magnetic
Resonance in Medicine, 41, 13–20.
Eickhoff, S. B., Heim, S., Zilles, K., & Amunts, K. (2009).
A systems perspective on the effective connectivity of
overt speech production. Philosophical Transactions
of the Royal Society A, 367, 2399–2421.
Eickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C.,
Fink, G. R., Amunts, K., et al. (2005). A new SPM toolbox
for combining probabilistic cytoarchitectonic maps and
functional imaging data. Neuroimage, 25, 1325–1335.
Elliott, M. R., Bowtell, R. W., & Morris, P. G. (1999). The effect
of scanner sound in visual, motor, and auditory functional
MRI. Magnetic Resonance in Medicine, 41, 1230–1235.
Ellis, A., & Lambon Ralph, M. A. (2000). Age of acquisition
effects in adult lexical processing reflect loss of plasticity
in maturing systems: Insights from connectionist networks.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 26, 1103–1123.
Ellis, A. W., Burani, C., Izura, C., Bromiley, A., & Venneri, A.
(2006). Traces of vocabulary acquisition in the brain:
Evidence from covert object naming. Neuroimage, 33,
958–968.
Forster, K. I. (1998). The pros and cons of masked priming.
Journal of Psycholinguistic Research, 27, 203–233.
Freire, L., Roche, A., & Mangin, J. F. (2002). What is the best
similarity measure for motion correction in fMRI time series?
IEEE Transactions on Medical Imaging, 21, 470–484.
Friston, K. J., Glaser, D. E., Henson, R. N. A., Kiebel, S., Phillips, C.,
& Ashburner, J. (2002). Classical and Bayesian inference in
neuroimaging: Applications. Neuroimage, 16, 484–512.
Goldrick, M. (2007). Connectionist principles in theories of
speech production. In M. G. Gaskell (Ed.), The Oxford
handbook of psycholinguistics (pp. 515–530). Oxford,
U.K.: Oxford University Press.
Gollan, T. H., Slattery, T. J., Goldenberg, D., van Assche, E.,
Duyck, W., & Rayner, K. (2011). Frequency drives lexical
access in reading but not in speaking: The frequency-lag
hypothesis. Journal of Experimental Psychology: General,
140, 186–209.
Gracco, V. L., Tremblay, P., & Pike, G. B. (2005). Imaging
speech production using fMRI. Neuroimage, 26, 294–301.
Hammers, A., Allom, R., Koepp, M. J., Free, S. L., Myers, R.,
Lemieux, L., et al. (2003). Three-dimensional maximum
probability atlas of the human brain, with particular
reference to the temporal lobe. Human Brain Mapping,
19, 224–247.
Harley, T. A. (1993). Phonological activation of semantic
competitors during lexical access in speech production.
Language and Cognitive Processes, 8, 291–309.
Hartsuiker, R. J. (2006). Are speech error patterns affected
by a monitoring bias? Language and Cognitive Processes,
21, 856–891.
Heim, S., Friederici, A. D., Schiller, N. O., Rüschemeyer, S. A.,
& Amunts, K. (2008). The determiner congruency effect
in language production investigated with functional MRI.
Human Brain Mapping, 30, 928–940.
Hockey, A., Wiles, J., & de Zubicaray, G. (2005, November).
Interactivity, conflict and cognitive control: Computational
modelling of the language production system. Poster
presented at the Dynamical Neuroscience Conference,
Washington, DC.
Hocking, J., McMahon, K., & de Zubicaray, G. (2010).
Semantic interference in object naming: An fMRI study
of the postcue naming paradigm. Neuroimage, 50,
796–801.
Humphreys, G. W., Lloyd-Jones, T. J., & Fias, W. (1995).
Semantic interference effects on naming using a postcue
procedure: Tapping the links between semantics and
phonology with pictures and words. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 21, 961–980.
Indefrey, P., & Levelt, W. J. M. (2004). The spatial and temporal
signatures of word production components. Cognition,
92, 101–144.
Janssen, N., Schirm, W., Mahon, B. Z., & Caramazza, A.
(2008). Semantic interference in a delayed naming task:
Evidence for the response exclusion hypothesis. Journal
of Experimental Psychology: Learning, Memory, and
Cognition, 34, 249–256.
La Heij, W. (1988). Components of Stroop-like interference
in picture naming. Memory & Cognition, 16, 400–410.
Lambon Ralph, M. A., & Ehsan, S. (2006). Age of acquisition
effects depend on the mapping between representations and
the frequency of occurrence: Empirical and computational
evidence. Visual Cognition, 13, 928–948.
Lee, C. H., Honig, R., & Lee, Y. (2002). Phonological recoding of
mixed-case words in the priming task. Reading Psychology,
23, 199–216.
Lee, M. L., & de Zubicaray, G. I. (2010, September). Lexical
selection is by competition: A failure to replicate Mahon
et al.ʼs (2007) Experiment 7. Poster presented at the Sixth
International Workshop on Language Production, Edinburgh,
Scotland.
Levelt, W. J. M., Roelofs, A., & Meyer, A. S. (1999). A theory of
lexical access in speech production. Behavioral and Brain
Sciences, 22, 1–75.
Macey, P. M., Macey, K. E., Kumar, R., & Harper, R. M. (2004). A
method for removal of global effects from fMRI time series.
Neuroimage, 22, 360–366.
Mädebach, A., Oppermann, F., Hantsch, A., Curda, C., &
Jescheniak, J. D. (2011). Is there semantic interference in
delayed naming? Journal of Experimental Psychology:
Learning, Memory, and Cognition, 37, 522–538.
Mahon, B. Z., Costa, A., Peterson, R., Vargas, K. A., &
Caramazza, A. (2007). Lexical selection is not by competition.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 33, 503–533.
McClelland, J. L., & Rumelhart, D. E. (1981). An interactive
activation model of context effects in letter perception.
Psychological Review, 88, 375–407.
McMahon, K., Pringle, A., Eastburn, M., & Maillet, D. (2004).
Improving EPI imaging quality and sound levels with
bandwidth selection. Proceedings of the 12th Annual
Meeting of the International Society for Magnetic
Resonance in Medicine, Kyoto, Japan, 1033 (abstract).
Menenti, L., & Burani, C. (2007). What causes the effect of
age-of-acquisition in lexical processing? Quarterly Journal
of Experimental Psychology, 60, 652–661.
Meschyan, G., & Hernandez, A. (2002). Age of acquisition
and word frequency: Determinants of object-naming
speed and accuracy. Memory and Cognition, 30, 262–269.
Miozzo, M., & Caramazza, A. (2003). When more is less:
A counterintuitive effect of distractor frequency in the
picture–word interference paradigm. Journal of
Experimental Psychology: General, 132, 228–252.
Morton, J. (1969). The interaction of information in word
recognition. Psychological Review, 76, 165–178.
Norris, D. (2006). The Bayesian reader: Explaining word
recognition as an optimal Bayesian decision process.
Psychological Review, 113, 327–357.
494
Journal of Cognitive Neuroscience
Volume 24, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Peeva, M. G., Guenther, F. H., Tourville, J. A., Nieto-Castanon, A.,
Anton, J.-L., Nazarian, B., et al. (2010). Distinct representations
of phonemes, syllables, and supra-syllabic sequences in the
speech production network. Neuroimage, 50, 626–638.
Piai, V., Roelofs, A., & Schriefers, H. (2011). Semantic interference
in immediate and delayed naming and reading: Attention and
task decisions. Journal of Memory and Language, 64, 404–423.
Price, C. J. (2010). The anatomy of language: A review of
100 fMRI studies published in 2009. Annals of the New
York Academy of Sciences, 1191, 62–88.
Riès, S., Janssen, N., Dufau, S., Alario, F. X., & Burle, B. (2011).
General-purpose monitoring during speech production.
Journal of Cognitive Neuroscience, 23, 1419–1436.
Righi, G., Blumstein, S., Mertus, J., & Worden, M. (2010). Neural
systems underlying lexical competition: An eyetracking and
fMRI study. Journal of Cognitive Neuroscience, 22, 213–224.
Roelofs, A. (1992). A spreading-activation theory of lemma
retrieval in speaking. Cognition, 42, 107–142.
Roelofs, A. (2005). From Popper to Lakatos: A case for
cumulative computational modeling. In A. Cutler (Ed.),
Twenty-first century psycholinguistics: Four cornerstones
(pp. 313–330). Hillsdale, NJ: LEA.
Roelofs, A. (2008). Attention to spoken word planning:
Chronometric and neuroimaging evidence. Language
and Linguistics Compass, 2, 389–405.
Roelofs, A., Piai, V., & Schriefers, H. (2011). Selective attention
and distractor frequency in naming performance: Comment
on Dhooge and Hartsuiker (2010). Journal of Experimental
Psychology: Learning, Memory, and Cognition, 37, 1032–1038.
Rosinski, R. R., Golinkoff, R. M., & Kukish, K. S. (1975).
Automatic semantic processing in a picture–word
interference task. Child Development, 26, 247–253.
Santesteban, M., Costa, A., Pontin, S., & Navarrete, E. (2006).
The effect of word-frequency on lexical selection in speech
production: Evidence from semantic homogeneous naming
contexts. Cognitiva, 18, 75–84.
Schuhmann, T., Schiller, N. O., Goebel, R., & Sack, A. T.
(2009). The temporal characteristics of functional activation
in Brocaʼs area during overt picture naming. Cortex, 45,
1111–1116.
Shaywitz, S. E., & Shaywitz, B. A. (2008). Paying attention
to reading: The neurobiology of reading and dyslexia.
Development and Psychopathology, 20, 1329–1349.
Snodgrass, J. G., & Vanderwart, M. (1980). A standardized set of
260 pictures: Norms for name agreement, image agreement,
familiarity, and visual complexity. Journal of Experimental
Psychology: Human Learning and Memory, 6, 174–215.
Starreveld, P. A., & La Heij, W. (1996). Time-course analysis of
semantic and orthographic context effects in picture-naming.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 22, 896–918.
Steyvers, M., & Tenenbaum, J. B. (2005). The large scale
structure of semantic networks: Statistical analyses and a
model of semantic growth. Cognitive Science, 29, 41–78.
Tremblay, P., & Gracco, V. L. (2009). On the selection
of words and oral motor responses: Evidence of a
response-independent fronto-parietal network. Cortex,
46, 15–28.
van Casteren, M., & Davis, M. H. (2006). Mix, a program
for pseudorandomization. Behavior Research Methods,
38, 584–589.
Vaughan, J. T., Adriany, G., Garwood, M., Yacoub, E., Duong, T.,
DelaBarre, L., et al. (2002). Detunable transverse electromagnetic
(TEM) volume coil for high-field NMR. Magnetic Resonance
in Medicine, 47, 990–1000.
Zaitsev, M., Hennig, J., & Speck, O. (2003). Automated online
EPI distortion correction for fMRI applications. Proceedings
of the 11th Annual Meeting of the International Society
for Magnetic Resonance in Medicine, 1042 (abstract).
Zevin, J. D., & Seidenberg, M. S. (2002). Age of acquisition
effects in word reading and other tasks. Journal of
Memory and Language, 47, 1–29.
Zheng, Z., Munhall, K. G., & Johnsrude, I. S. (2010). Functional
overlap between regions involved in speech perception
and monitoring oneʼs own voice during speech production.
Journal of Cognitive Neuroscience, 22, 1770–1781.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
2
2
4
4
/
8
2
2
/
1
4
9
8
4
2
3
/
2
1
6
7
5
7
o
7
c
4
n
5
_
8
a
/
_
j
0
o
0
c
1
n
4
1
_
a
p
_
d
0
0
b
1
y
4
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
de Zubicaray et al.
495