Imaging Informational Conflict: A Functional
Magnetic Resonance Imaging Study of
Numerical Stroop
J. Tang, H. D. Critchley, D. E. Glaser, R. J. Dolan, and B. Butterworth
Abstrakt
& We employed a parametric version of the comparison
Stroop paradigm to investigate the processing of numerical
magnitude and physical size under task-relevant and -irrelevant
conditions to investigate two theoretical issues: (1) What is the
fate of task-irrelevant information? (2) What is the
neural
neural basis of the resolution of the conflict between task-
relevant and -irrelevant information? We show in 18 healthy
adults that numerical magnitudes of numbers call for higher
processing requirements than physical sizes. The enhanced
activation elicited by numerical magnitudes is not modulated
by task relevance, indicating autonomous processing. More-
über, the normal behavioral distance effect when the numerical
dimension is task relevant and reversed distance effect when it
is not show that autonomous processing fully encodes
numerical magnitudes. Conflict trials elicited greater activation
in bilateral inferior frontal gyri, right middle frontal gyri, Und
right superior frontal gyri. We postulate two sources to the
conflict, nämlich, at cognitive and response levels. &
EINFÜHRUNG
Task-irrelevant information can interfere with perform-
ance. This was classically demonstrated by Stroop
(1935), using a task where naming the ink color of a word
is more error prone and slower when the word is a con-
flicting color word, Zum Beispiel, where the word GREEN
is printed in red. This is now known as the ‘‘Stroop
effect.’’ Nevertheless, the majority of responses in Stroop
task performance are correct, indicating appropriate res-
olution of the conflict between task-relevant and task-
irrelevant information (Derbyshire, Vogt, & Jones, 1998;
Fuhrmann, Mintun, & Cohen, 1995; George et al., 1994; Bench
et al., 1993; Pardo, Pardo, Janer, & Rachel, 1990).
Functional neuroimaging experiments of Stroop inter-
ference effects potentially provide insight into two im-
portant general theoretical questions: (1) What is the
neural fate of task-irrelevant information? More specifi-
cally, are there qualitative or quantitative differences in
the activation patterns that can be assumed to imply
different cognitive processing, or is the processing of the
stimulus dimensions autonomous so that task does not
modulate neural activity? (2) What is the neural basis of
the resolution of the conflict between task-relevant and
-irrelevant information?
We suggest that the classical color-word task is not
well suited to answering these questions because the
competing dimensions are unbalanced. Although words
University College London, Vereinigtes Königreich
interfere with color naming, color has little effect on
word naming: There is rarely a significant slowing when
naming the word ‘‘GREEN’’ printed in red, even when
color information has been manipulated to precede the
word by 400 ms (Glaser & Glaser, 1982). Außerdem,
it is impossible to order color names on a continuum, Also
even if one is able to parametrically vary the hue of a
color, it is impossible to have a parametric modulation
on the color name dimension.
The number Stroop task provides a more appro-
priate alternative, as described originally by Besner and
Coltheart (1979), and uses the dimensions numerical
magnitude and physical size. In a typical experiment,
both numerical magnitudes and physical sizes of the
numbers displayed vary; subjects have to select the
larger number in either the numerical or the physical
dimension. Trials may be congruent, where the nu-
merically larger number is physically larger (z.B., 3 5);
incongruent, where the numerically larger number is
physically smaller (z.B., 3 5); Und, in some experiments,
neutral where the numbers are displayed in the same
Größe (z.B., 3 5) for the numerical comparison task and
where the same numbers are displayed in different sizes
(z.B., 3 3) for the physical comparison task. The Stroop
effect manifests as interference (an increase in reaction
time and/or error rate in incongruent trials compared
with neutral trials) and/or facilitation (a decrease in
reaction time and/or error rate in congruent trials
compared with neutral trials) (z.B., Girelli, Lucangeli,
& Butterworth, 2000; Tzelgov, Meyer, & Henik, 1992;
D 2006 Massachusetts Institute of Technology
Zeitschrift für kognitive Neurowissenschaften 18:12, S. 2049–2062
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
F
.
.
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Foltz, Poltrock, & Potts, 1984; Henik & Tzelgov, 1982;
Besner & Coltheart, 1979). Facilitation is always sub-
stantially smaller than interference (für eine Rezension, sehen
MacLeod, 1991).
A further advantage of the number Stroop task is that
it provides a metric for processing ‘‘refinement’’: Die
numerical distance effect (Moyer & Landauer, 1967).
This effect,
like other symbolic and nonsymbolic dis-
tance effects, means that it is harder to discriminate
similar stimuli than dissimilar stimuli. In the case of
Zahlen, similarity is on the semantic dimension of
numerical magnitude. Comparison times are inversely
related to the numerical distance between two numbers,
so that, Zum Beispiel, 3 4 takes longer to compare than 3
5 (Moyer & Landauer, 1967), Und, generally, compari-
sons times are well captured by the Welford function
(Zorzi & Butterworth, 1999, and see below). The dis-
tance effect holds for written number words (Foltz et al.,
1984) and dot patterns (Buckley & Gillman, 1974). Daher,
the presence of a distance effect shows that processing
has gone beyond a mere large–small categorization and
that the numbers have been processed to a semantic
level that discriminates between the magnitudes of each
number (Tzelgov et al., 1992).
The effect can be observed in numerical Stroop
tasks (Pinel, Piazza, Le Bihan, & Dehaene, 2004; Fias,
Lammertyn, Reynvoet, Dupont, & Orban, 2003; Pinel,
Dehaene, Rivie`re, & Le Bihan, 2001; Foltz et al., 1984;
Hinrichs, Yurko & Hu, 1981; Duncan & MacFarland,
1980), but when physical size is manipulated as the
task-relevant dimension and numerical magnitude as
the task-irrelevant dimension, the numerical distance
effect can vanish (Rubinsten, Henik, Berger, & Shahar-
Shalev, 2002) or even reverse (Girelli et al., 2000; Henik
& Tzelgov, 1982). See below for an account of the
reversed distance effect.
In this study, we further examined the autonomous
aspect of numerical information processing. Autonomy,
by definition, is a property of automatic processes, Das
Ist, processes that are fast, effortless, and unconscious
(z.B., Logan, 1980; Shiffrin & Schneider, 1977; Posner &
Snyder, 1975). Here we use the concept autonomous to
refer to a process that takes place even when it is
irrelevant to the task at hand, so that it begins and runs
to completion without intention (Zbrodoff & Logan,
1986). The number Stroop paradigm is particularly well
suited to investigate the depth of processing because it
uses the distance effect and the reversed distance effect
as metrics.
A reversed distance effect in the task-irrelevant di-
mension (z.B., Girelli et al., 2000; Henik & Tzelgov,
1982) does not necessarily suggest that information is
processed differently in the task-irrelevant channel. Fol-
lowing Girelli et al. (2000),
if both dimensions are
processed autonomously, conflict will arise only when
competing outcomes are simultaneously active. It takes
less time to generate a candidate response when the
distance is greater. Daher, this candidate response in the
task-irrelevant dimension will be generated more quickly
to interfere with the generation of the task-relevant
response. Zum Beispiel, selecting the numerically larger
item will be slower for 3 5 als 3 5 because it will
be quicker to resolve the size difference in the first
Fall. In der Tat, the reversed distance effect, like the nor-
mal distance effect, can be construed as evidence that
the processing of numerical information has continued
autonomously to its normal completion.
There is already considerable evidence as to where
semantic processing of numbers is expressed in the
Gehirn. Pinel et al. (2001) has described two distinct
stages of numerical processing, nämlich,
identification
and semantic processing. The ventral occipitotemporal
areas are activated bilaterally by the visual shapes of
Arabic numerals (Dehaene & Cohen, 1995) during the
identification process. Word identification is thought to
be strictly left lateralized and to rely on the left ‘‘visual
word form area,’’ a region of the left fusiform gyrus that
is involved in the recognition of visual words (Shallice,
1988). Jedoch, Pinel and colleagues (Pinel et al., 2001;
Pinel et al., 1999) have provided evidence that the right
fusiform gyrus is implicated in the identification of
Arabic numerals.
Several studies implicate the parietal lobes in support-
ing a notation-independent semantic representation of
Mengen (see Dehaene & Cohen, 1995, für eine Rezension).
Genauer, the intraparietal sulcus and the pre-
cuneus have been implicated in the comparison process
of numbers, and the activity of these regions is modu-
lated by the numerical distance effect—smaller numer-
ical distances associated with high activation levels (Pinel
et al., 2001). Pinel et al. (2004) extended these findings
to other nonsymbolic continua, such as physical size and
luminance. By using a number Stroop paradigm, Sie
reported that during the numerical comparison task, Die
numerical distance effect was associated with enhanced
bilateral activity of the horizontal segment of the intra-
parietal sulci and the left precentral gyrus. Im Gegensatz,
during physical size comparisons with number stimuli,
correlates of the physical distance effect were found
predominantly in the right hemisphere, in particular the
right prefrontal and occipital cortices and much of the
right intraparietal sulcus. Intersections of the correlates
between numerical magnitude and physical size distance
effects were observed in bilateral regions of the anterior
intraparietal sulcus. The authors concluded that during
comparative judgements, these continuous quantities
are engaged in common parietal representations.
It is important to note that the studies by Pinel and
colleagues (Pinel et al., 2004; Pinel et al., 2001) rest on the
basic assumption that for any given task, the first stage
is to translate the input into the appropriate representa-
tion, and in the case of comparison, the stimuli are con-
verted to analogue representations of quantity (Dehaene,
1992). Although Pinel et al.’s (2004) findings support
2050
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
F
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
common parietal representations in processing numer-
ical magnitude and physical size, it was also observed
that comparative judgements of numerical magnitudes
yielded larger activation in several parietal regions (bilat-
eral inferior parietal foci and the left intraparietal sulcus)
than those of physical sizes. No explanation was provided
by the authors for such a difference.
Problems with Previous Studies
of Numerical Stroop
In previous number Stroop studies, the attended and
unattended dimensions are not properly balanced. Für
Beispiel, nine numbers have been used to create two
levels of numerical distance, distant and close pairs (In
addition to distance 0 for neutral pairs), whereas three
physical sizes were often employed to create large,
small, and neutral stimuli; somit, only one level of
physical distance (in addition to distance 0 for neutral
pairs) was used. This meant that the two competing
dimensions were not appropriately matched,
limiting
the inferences about the amount of information avail-
able in the task-relevant and -irrelevant channels (z.B.,
Girelli et al., 2000; Henik & Tzelgov, 1982). Rubinsten
et al. (2002) used three levels of numerical distance
and two levels of physical distance, Und, more recently,
Pinel et al. (2004) varied both numerical distance and
physical distance, but the stimuli were grouped in the
Analysen, so that the factor distance consisted of only
two levels (distant and close pairs). With such a design,
Pinel et al. failed to observe any distance effect in either
numerical magnitude or physical size when they were
task irrelevant.
In the present experiment, we parametrically varied
both numerical distance and physical distance and test-
ed for distance effects under both task-relevant and
-irrelevant conditions. Hier, we viewed distance effects
as indicators for refined information processing; Das
contrasts with the coarse large–small dichotomous clas-
sification implicated by the Stroop effect.
Neural Basis of Conflict Resolution
Specific brain regions have been implicated in process-
ing and resolution of informational conflict from task-
irrelevant features. Insbesondere, enhanced activation in
the anterior cingulate cortex (ACC) has been reported in
color-word Stroop tasks where subjects have to name
the color of a conflicting color word, Zum Beispiel, Die
word GREEN printed in red, compared to a noncon-
flicting stimulus (z.B., Derbyshire et al., 1998; Fuhrmann
et al., 1995; George et al., 1994; Bench et al., 1993;
Pardo et al., 1990). Ähnlich, greater ACC activation has
also been observed during incompatible compared to
compatible trials, in flanker tasks (z.B., Durston et al.,
2003; Bunge, Dudukovic, Thomason, Vaidya, & Gabriela,
2002; van Veen, Cohen, Botvinick, Stenger, & Fuhrmann;
2001; Casey et al., 2000; Hazeltine, Poldrack, & Gabriela,
2000; Botvinick, Nystrom, Fissell, Fuhrmann, & Cohen,
1999). These findings have been interpreted as suggest-
ing a role in conflict resolution.
Jedoch, more recent research has indicated that the
precise function of the ACC rests in the detection of
conflict, rather than in the resolution of conflict per se.
The model proposed by Botvinick, Mutiger, Barch, Fuhrmann,
and Cohen (2001) assumes that conflict monitoring
(z.B., detection) influences cognitive control. In partic-
ular, during a period of high conflict, more attention is
directed to the relevant task, and if another conflict
follows close in time, less interference is expected due to
the already heightened state of the system. In other
Wörter, the model predicts that not only behavior, Aber
also ACC activation would be affected by (1) Variationen
in trial type frequency and (2) the preceding trial’s
trial type.
Evidence to support Botvinick et al.’s (2001) Modell
has come from various types of interference tasks. In einem
color-word Stroop task, Carter et al. (2000) reported
that ACC activation during incongruent trials was higher
when such trials were infrequent than when they were
frequent, paralleling the behavioral finding that incon-
gruent trials induced more conflict when such trials
were rare. Ähnlich,
in a flanker task, greater ACC
activation was observed in incompatible trials that fol-
lowed compatible trials than those that followed incom-
patible trials (Botvinick et al., 1999), a finding replicated
and extended by Durston et al. (2003). Außerdem, in einem
go/no-go task, the increase in ACC activation to a no-go
trial is a function of increasing number of preceding go
Versuche (Durston et al., 2002). All these findings are
consistent with the involvement of the ACC in conflict
detection and support Botvinick et al.’s (2001) proposal
that high ACC activation triggers an increased attention
on the relevant task, leading to less interference in cases
where conflicts are frequent or consecutive.
Another well-established finding is that ACC activity has
been associated with error commission (z.B., Critchley,
Tang, Glaser, Butterworth, & Dolan, 2005; Mutiger, Barch,
Gray, Molfese, & Snyder, 2001; Menon, Adleman, White,
Glover, & Reiss, 2001; Falkenstein, Hoormann, Christ,
& Hohnsbein, 2000; Kiehl, Kiddle, & Hopfinger, 2000;
Carter et al., 1998; Gehring, Goss, Coles, Meyer, &
Donchin, 1993; für eine Rezension, see Botvinick, Cohen, &
Fuhrmann, 2004). Behavioral and electromyographic obser-
vations indicate that errors in speeded response tasks
are frequently associated with response conflict (Yeung,
Botvinick, & Cohen, 2004). The reason that errors are
committed even when there is a conflict detection system
is that even as an error response is being executed,
ongoing processing of the stimulus often leads to a
belated activation of the correct response, giving rise to
a transient period during with both correct and incorrect
responses are activated. Daher, ACC activation during er-
rors reflects the detection of a postresponse conflict.
Tang et al.
2051
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
Conflict can occur at numerous levels of information
Verarbeitung, ranging from perceptual representation to
response selection. Several studies have reported that
the ACC is engaged most strongly during conflicts at
the level of response selection (Milham, Banich, &
Barad, 2003; Nelson, Reuter-Lorenz, Sylvester, Jonides,
& Schmied, 2003; Weissman, Giesbrecht, Song, Mangun, &
Woldorff, 2003; Bunge et al., 2002; Milham et al., 2001;
van Veen et al., 2001). Andererseits, da war ein
lack of substantial ACC activation when comparing be-
tween conflict and nonconflict trials in a color-word
matching Stroop task (Zysset, Mu¨ller, Lohmann, & von
Cramon, 2001). Stattdessen, Zysset et al. (2001) observed
enhanced activation in regions along the left inferior
Stirnfurche (IFS) during conflict trials compared to
nonconflict trials (neutral and congruent trials) and con-
cluded that ‘‘regions along the IFS appear to be involved
in solving interference effect and task management.’’
Research employing other paradigms supports this.
Go/no-go and stop-signal tasks require subjects to per-
form speeded responses on ‘‘go’’ trials and to inhibit
their response on ‘‘no-go’’ or ‘‘stop’’ trials. Such re-
sponse inhibition has been reported to activate regions
along the right inferior frontal gyrus in neuroimaging
Studien (z.B., Bunge et al., 2002; Durston et al., 2002;
Menon et al., 2001; Garavan, Ross, & Stein, 1999; Rubia
et al., 1999; Konishi, Nakajima, Uchida, Sekihara, &
Miyashita, 1998, 1999). In some studies, enhanced acti-
vation was observed bilaterally (z.B., Menon et al., 2001).
The idea that ACC activation is associated with conflict
detection suggests an effect on attentional allocation (für
a review, see Botvinick et al., 2004). Andererseits,
there is evidence to suggest that regions in the inferior
frontal cortex are responsible for inhibiting prepotent
responses (z.B., Bunge et al., 2002; Durston et al., 2002;
Menon et al., 2001; Garavan et al., 1999; Rubia et al., 1999;
Konishi et al., 1998, 1999) and are perhaps involved in
conflict resolution (Zysset et al., 2001). More recently,
van Veen and Carter (2005) reported distinct neural sub-
strates for cognitive (semantic) interference and response
conflict in a modified color-word Stroop task. Bilateral
middle frontal gyri, right superior frontal gyrus, und das
ACC were activated in response to cognitive interference.
Response conflict activated the bilateral middle/inferior
frontal gyri and a more anterior region in the ACC. Con-
junction analysis revealed no overlap in activation be-
tween the two contrasts.
The present study investigates the neural fate of task-
irrelevant information using a modified version of the
number Stroop paradigm with two comparable dimen-
sionen, nämlich, numerical magnitude and physical size.
The parametric design allows us to vary and measure
systematically the amount of interference exerted by
the task-irrelevant channels during conflict trials in
each of the comparison tasks, and hence characterize
the information-processing fate of task. A reversed nu-
merical distance effect in physical comparisons would
indicate that exact numerical values had been com-
puted despite task irrelevance (Girelli et al., 2000; Henik
& Tzelgov, 1982). Außerdem, the current design al-
lows us to test, for the first time, for a reversed physical
distance effect in numerical comparisons (d.h., Aufgabe-
irrelevant condition). The reversed effects are indicators
for autonomous information processing.
In terms of brain activation, parietal activation levels
were predicted to be inversely related to numerical
distance in line with previous findings (Pinel et al.,
2004; Pinel et al., 2001). The key issue in the experiment
was whether task relevance modulates parietal activity. Wenn
the fate of task-irrelevant information in this task was
autonomous processing, then an absence of a modula-
tion would be suggestive. The second key issue was to
identify the neural basis of the resolution of the conflict
between task-relevant and irrelevant information. Wieder,
balanced dimensions in a parametric design could reveal
activations specific to conflict and to errors arising from
conflict. We expected that conflict and errors would
modulate activity in frontal rather than parietal regions
where numbers and size are primarily represented.
METHODEN
Tasks
The two tasks were numerical magnitude and physical size
comparisons. Subjects had to select the larger number
numerically or physically according to the task require-
ment. Subjects responded by pressing the left or right
button to indicate the side of the larger relevant attribute.
Reaction times and responses were recorded. A program
written in Cogent (which runs on a MATLAB Version 6.1
platform; The MathWorks, Natick, MA) was used.
Stimuli
The stimuli, presented on a screen situated outside
the scanner, were reflected onto a mirror (of size 20 (cid:1)
9 cm2) placed inside the scanner. In jedem Versuch, two di-
gits appeared simultaneously in white on a black back-
Boden. Each presentation lasted 1000 msec with an
interstimulus interval of 3000 ms.
The stimuli were Arabic digits (1 Zu 9) in Arial font.
Each digit might appear in one of the nine different sizes
(subtending from approximately 3.88 Zu 9.78 with a mean
ratio of 1.1 between adjacent sizes). Four numerical
distances (ND = 1, 2, 3, Und 4) and four physical
distances (PD = 1, 2, 3, Und 4 Einheiten) wurden benutzt. Der
pairs used were as follows: 2 3, 3 4, 6 7, 7 8 (for ND of 1);
1 3, 2 4, 6 8, 7 9 (for ND of 2); 1 4, 2 5, 5 8, 6 9 (for ND of
3); 1 5, 2 6, 4 8, 5 9 (for ND of 4). The equivalent physical
sizes were used to systematically vary PD.
There were three experimental conditions: congru-
ent, when the numerically larger digit was physically
larger (z.B., 2 6); incongruent, when the numerically
2052
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
larger digit was physically smaller (z.B., 2 6) or vice
versa; and neutral, when the two digits were of the
same physical size in the numerical comparison task
(z.B., 2 6) or when the same digit appeared in different
sizes in the physical comparison task (z.B., 2 2).
The total number of trials in each task was 217 (d.h., 64
congruent + 64 neutral + 64 incongruent + 25 blank =
217 Versuche). Stimuli were presented in a pseudorandom
order to avoid carryover effects.
Subjects
Es gab 18 right-handed subjects (11 men and 7
Frauen), gealtert 21 Zu 38 Jahre (M = 25.0 Jahre, SD =
4.01 Jahre). They performed two tasks (numerical and
physical comparison tasks); half of them participated in
the numerical task first, and the other half in the phys-
ical task first. All subjects had normal or corrected-to-
normal eyesight.
Scanning Procedures and Imaging
Data Processing
Whole-brain functional magnetic resonance imaging
(fMRT) data was acquired on a 1.5-T Magnetom VISION
System (Siemens Sonata, Erlangen, Deutschland). Function-
al
images were obtained with a gradient-echo-planar
sequence using blood oxygenation level-dependent
(BOLD) Kontrast, each comprising a full-brain volume
von 28 contiguous axial slices, 3.5-mm thickness). Bd-
umes were acquired continuously with a repetition time
(TR) von 2.52 Sek. A total of 275 scans were acquired for
each participant in two sessions (etwa 10 min
jede), with the first six volumes subsequently discarded
to allow for T1 equilibration effects. During fMRI scan-
ning, pupil diameter was recorded online by an in-
frared eye tracker. The data were analyzed using SPM2
(Wellcome Department of Imaging Neuroscience; www.
fil.ion.ucl.ac.uk/spm) implemented in MATLAB 6.1.0.450
Release 12.1. Individual scans were realigned, slice time
corrected, normalized to the MNI template with voxels
von 2 (cid:1) 2 (cid:1) 2 mm3 and spatially smoothed by an 8-mm
full width half-maximum (FWHM) Gaussian kernel using
standard SPM methods.
Event-related activity for each voxel, for each condi-
tion and each subject was modeled using a canonical
hemodynamic response function plus temporal and
dispersion derivatives. Statistical parametric maps of
the t statistic (SPM{T}) were generated for each subject
and the contrast images were further smoothed by an
8-mm FWHM Gaussian kernel.
At the second-level random-effects analysis, A 2 (cid:1) 4
analysis of variance (ANOVA) model was applied; Die
factors were task (numerical comparison task and phys-
ical comparison task) and trial type (congruent, neutral,
incongruent, and error trials). Congruent and incon-
gruent trials were modeled parametrically with respect
to task-relevant and task-irrelevant distance, and neu-
tral trials were modeled parametrically with respect
to task-relevant distance. This allowed us to construct
t contrasts to test specifically for distance-modulated ac-
tivities. Our model was optimized to detect linear in-
crease/decrease in activities related to numerical and/
or physical distance. In der vorliegenden Studie, congruent
and neutral trials were classified as nonconflict trials,
whereas incongruent trials were conflict trials. To com-
pare between conflict and nonconflict trials, and error
and correct trials in each task, t contrasts were con-
structed. Threshold significance was set at .001 uncor-
rected for multiple comparisons.
ERGEBNISSE
Behavioral Data
Errors were incorrect responses made in the compari-
son tasks, das ist, trials where subjects pressed the
wrong key. Reaction time outliers were identified by
the standard SPSS procedure (values that lay more than
1.5 times the interquartile range above the third quartile
oder 1.5 times the interquartile range below the first
quartile) and removed. ANOVAs were used to analyze
mean error rates and mean reaction times, and when-
ever Mauchly’s test of sphericity assumption was violat-
Hrsg, the Greenhouse–Geisser epsilon was used to correct
the degrees of freedom. A 2 (cid:1) 3 repeated measures
ANOVA was conducted on mean error rates. The factors
were task (numerical comparison task and physical
comparison task) and congruent (congruent, neutral,
and incongruent). The ANOVA revealed a significant
main effect of congruity, F(1,22) = 38.10, p = .001; A
nonsignificant main effect of task, F(1,17) = 3.34, ns;
and a nonsignificant Task (cid:1) Congruity interaction,
F(1,21) < 1, ns. Tests of within-subjects contrasts re-
vealed a significant difference in error rates between
incongruent and neutral trials, F(1,17) = 52.00, p < .001,
and a nonsignificant difference between neutral and
congruent trials, F(1,17) < 1, ns. The mean error rates
were 11.55%, 4.43%, and 3.95%, respectively.
A 2 (cid:1) 3 repeated measures ANOVA was conducted on
mean reaction times. The factors were task (numerical
and physical comparison tasks) and congruent (congru-
ent, neutral, and incongruent). The ANOVA revealed
a significant main effect of task, F(1,17) = 16.55, p =
.001, a significant main effect of congruity, F(2,34) =
156.62, p < .001, and a significant Task (cid:1) Congruity
interaction, F(2,34) = 8.07, p = .001. The mean reaction
times for numerical and physical comparison tasks were
617 and 570 msec, respectively.
Considering only the numerical comparison task, the
main effect of congruity was significant, F(2,34) =
112.52, p < .001. Tests of within-subjects contrasts
revealed a significant difference between congruent
Tang et al.
2053
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
2
p
2
d
0
f
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
o
9
c
/
n
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
j
2
o
2
c
0
n
4
.
9
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
2
t
.
o
2
n
0
0
4
8
9
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
and neutral trials, F(1,17) = 47.84, p < .001, and
between the latter and incongruent trials, F(1,17) =
81.60, p < .001. The mean reaction times were 585,
614, and 652 msec, respectively. Similarly for the phys-
ical comparison task, the main effect of congruity was
significant, F(2,34) = 41.62, p < .001. Tests of within-
subjects contrasts revealed a significant difference be-
tween congruent and neutral trials, F(1,17) = 5.16,
p < .050, and between the latter and incongruent trials,
F(1,17) = 56.16, p < .001. The mean reaction times were
553, 564, and 593 msec, respectively.
Further analyses were conducted on mean reaction
times to test for distance effects in both the task-relevant
and -irrelevant dimensions for each task focusing on
incongruent trials, because only here was there infor-
mational conflict. For the numerical comparison task, a
2 (cid:1) 4 repeated measures ANOVA was conducted on
mean reaction times of the incongruent trials. The fac-
tors were task relevance (task-relevant and -irrelevant)
and distance (1, 2, 3, and 4). The ANOVA revealed a
significant Attention (cid:1) Task Relevance interaction,
F(3,51) = 13.02, p < .001. There was no significant
main effect (all nonsignificant). Further analyses were
conducted at each level of task relevance.
At the task-relevant level, the factor distance (in this
case, the numerical distance) showed a significant main
effect, F(3,51) = 8.61, p < .001. Tests of within-subjects
contrasts revealed a significant negative linear trend for
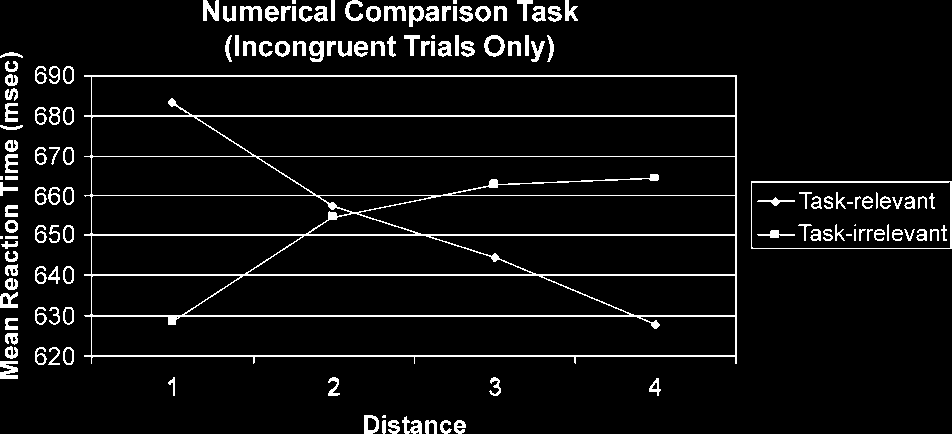
this factor, F(1,17) = 19.20, p < .001 (see Figure 1).
No other trend was significant. At the task-irrelevant
level, the factor distance (in this case, the physical dis-
tance) showed a significant main effect, F(3,51) = 4.78,
p = .005. Tests of within-subjects contrasts revealed a
significant positive linear trend for this factor, F(1,17) =
7.51, p < .050 (see Figure 1). No other trend was
significant.
When considering the physical comparison task, we
conducted a 2 (cid:1) 4 repeated measures ANOVA on mean
reaction times of the incongruent trials. The factors were
task relevance (task-relevant and -irrelevant) and dis-
tance (1, 2, 3, and 4). The ANOVA revealed a significant
main effect of task, F(1,17) = 19.11, p < .001, a
Figure 1. Distance effects in task-relevant and -irrelevant
dimensions of the numerical magnitude comparison task.
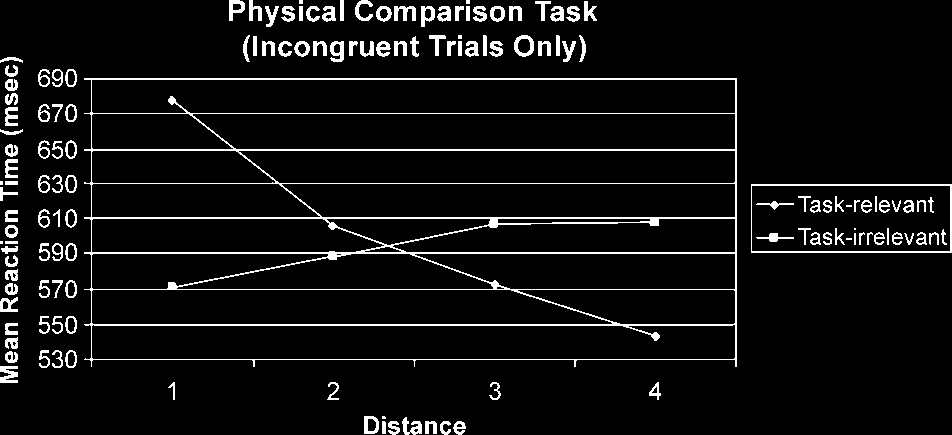
Figure 2. Distance effects in task-relevant and -irrelevant
dimensions of the physical size task.
significant main effect of distance, F(3,51) = 16.26,
p < .001, and a significant Attention (cid:1) Task Relevance
interaction, F(3,51) = 55.58, p < .001. Further analyses
were conducted at each level of task relevance.
At the task-relevant level, the factor distance (in this
case, the physical distance) showed a significant main
effect, F(2,33) = 53.49, p < .001. Tests of within-subjects
contrasts revealed a significant negative linear trend,
F(1,17) = 96.44, p < .001, and a significant quadratic
trend, F(1,17) = 8.98, p < .010, for this factor (see
Figure 2). At the task-irrelevant level, the factor distance
(in this case, the numerical distance) showed a signifi-
cant main effect, F(3,51) = 7.59, p < .001. Tests of
within-subjects contrasts revealed a significant positive
linear trend, F(1,17) = 30.95, p < .001 (see Figure 2). No
other trend was significant.
In summary, during conflict trials, the task-relevant
dimension showed a classic distance effect (indicated by
a negative linear trend), whereas the task-irrelevant
dimension showed a reversed distance effect (indicated
by a positive trend), regardless of task.
Functional Imaging Data
Functional imaging data analysis at the first level allowed
for neural responses associated with congruent, neutral,
incongruent, and error trials to be modeled indepen-
dently. Second-level t contrasts were constructed to test
for brain regions associated with the parametric modu-
lation of numerical distance and physical distance during
neutral conditions. An F contrast was then constructed
to compare processing of the two dimensions. Analysis
on the conflict trials allowed us to test for a Task (cid:1)
Distance interaction. To identify regions involved in
conflict and error trials, t contrasts were used. Conjunc-
tion analyses by inclusive masking were also performed
to identify common regions for conflict and error pro-
cessing across numerical and physical comparison tasks.
Brain activations are summarized in Tables 1–3. Voxels
reported are in Talairach coordinate space.
In neutral trials, distance only varied parametrically in
the task-relevant dimension, that is, only numerical
2054
Journal of Cognitive Neuroscience
Volume 18, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
2
p
2
d
0
f
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
o
9
c
/
n
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
j
2
o
2
c
0
n
4
.
9
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
2
t
.
o
2
n
0
0
4
8
9
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
distance was manipulated in the numerical task, and
only physical distance in the physical task. No parietal
region parametrically modulated by numerical distance
or physical distance during these trials were revealed by
t contrasts. However, the F contrast (see Table 1)
comparing the processing of numerical distance and
physical distance during neutral trials revealed several
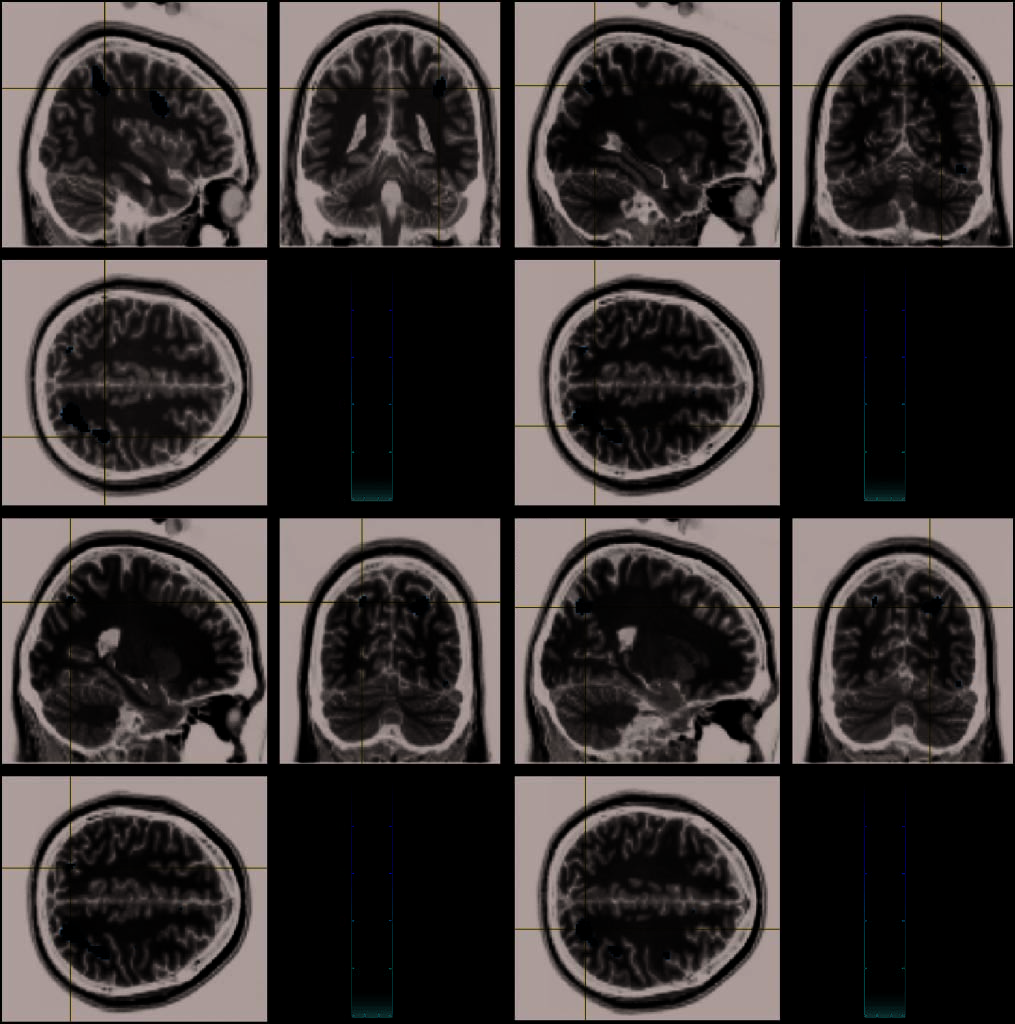
parietal regions that showed enhanced activation in
processing numerical relative to physical distance in-
cluding the right inferior parietal lobule [40 (cid:2)39 42],
right precuneus [22 (cid:2)64 42], right inferior parietal
lobule [32 (cid:2)56 45], and left superior parietal
lobule
[(cid:2)22 (cid:2)66 46], as well as the bilateral inferior frontal gyri
and right temporal and occipital regions (see Table 1
and Figure 3). Small volume correction searches (5-mm
radius) were performed with reference to Pinel et al.
(2004) and revealed that the right inferior parietal lobule
[38 (cid:2)41 43] also showed enhanced activation processing
numerical distance compared to physical distance. No
enhanced activation was observed processing physical
distance relative to numerical distance. Conjunction by
inclusive masking across the two tasks revealed no
commonly activated voxel.
However, the parietal regions that showed enhanced
activation processing numerical relative to physical dis-
tance were not affected by task requirement during
conflict trials. The F contrast constructed to test for a
Task (numerical task and physical task) (cid:1) Dimension
(numerical distance and physical distance) revealed no
significant difference in parietal activation.
To test for differences in brain activity between con-
flict and nonconflict trials, t contrasts were constructed
(see Table 2). In the numerical task, enhanced activation
in conflict trials compared to nonconflict trials was
observed in right inferior frontal [44 9 31] and middle
frontal [36 52 (cid:2)14] gyri, left fusiform gyrus [(cid:2)44 (cid:2)49
(cid:2)13], right occipital lobe [34 (cid:2)76 (cid:2)1], and various other
regions. In the physical task, enhanced activation in
conflict trials was observed only in the left inferior
frontal gyrus [(cid:2)40 30 8]. Conjunction by inclusive
masking across the two tasks revealed no commonly
activated voxel. To test for differences in brain activity
between error and correct trials, t contrasts were con-
structed (see Table 3). Conjunction by inclusive masking
(corrected for familywise error at 0.05) was performed
across task and revealed enhanced activation in error
trials compared to correct trials in bilateral
inferior
frontal gyri: left [(cid:2)32 17 (cid:2)11] and right [42 27 (cid:2)6],
and several regions along the bilateral superior temporal
gyri (see Table 3).
DISCUSSION
In the present experiment we examined processing of
numerical magnitude and physical size using bidimen-
sional stimuli. By varying the two dimensions, we were
able to create conflicting situations as well as systemat-
ically vary the amount of interference for each of task-
relevant dimensions. We made efforts to match the two
Table 1. An F Contrast Revealed Regions that Showed Enhanced Activation when Processing Numerical Distance Compared
with Physical Distance
Talairach Coordinates
Voxels
z Score
x
y
z
Brain Area
F contrast (numerical distance > physical distance)
758
169
55
501
95
24
23 (SVC)
4.72
3.86
3.69
3.68
3.53
3.45
3.45
3.39
3.22
3.67
46
(cid:2)36
46
40
22
32
50
46
(cid:2)22
38
7
17
(cid:2)77
(cid:2)39
(cid:2)64
(cid:2)56
(cid:2)53
(cid:2)61
(cid:2)66
(cid:2)41
24
(cid:2)3
17
42
42
45
(cid:2)9
(cid:2)9
46
41
Right frontal lobe; inferior frontal gyrus; white matter
Left frontal lobe; inferior frontal gyrus; gray matter; BA 47
Right occipital lobe; middle occipital gyrus; white matter
Right parietal lobe; inferior parietal lobule; gray matter; BA 40
Right parietal lobe; precuneus; white matter
Right parietal lobe; inferior parietal lobule; white matter
Right temporal lobe; subgyral; white matter
Right occipital lobe; subgyral; white matter
Left parietal lobe; superior parietal lobule; gray matter; BA 7
Right inferior parietal lobule
No significant voxel was found in the opposition direction. Regions that showed enhanced activation under task-relevant conditions compared
with task-irrelevant conditions with respect to increasing distance in numerical and physical distance were revealed by t contrasts. BA =
Brodmann’s area; SVC = small volume correction.
Tang et al.
2055
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
.
F
.
Ö
N
1
8
M
A
j
2
0
2
1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
F
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Figur 3. Parietal regions showing enhanced activation when processing numerical distance relative to physical size (clockwise from top
links: regions in the right inferior parietal lobule ([40 (cid:2)39 42] Und [32 (cid:2)56 45]), right precuneus [22 (cid:2)64 42], and left superior parietal
lobule [(cid:2)22 (cid:2)66 46]).
dimensions by difficulty. In terms of error commission,
the two tasks did not differ in either rate or pattern.
Jedoch, with respect to reaction times, the physical
comparison task was significantly faster than the numer-
ical comparison task. Such a difference could be ex-
plained by the different nature of these tasks—physical
size comparisons require processing at the perceptual
Ebene, whereas numerical magnitude comparisons re-
quire higher cognitive processing. On this basis, es ist
not surprising that processing time was shorter for
the former. It is important to note that the two tasks
showed the same reaction time patterns with respect
to congruity—significant interference and facilitation in
both numerical and physical tasks; mit anderen Worten, Die
classical Stroop effect. Although not perfectly matched
in terms of reaction time, the Stroop effect was observed
in both tasks, indicating that two directions influenced
one another and, somit, the present paradigm is a more
2056
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 12
Tisch 2. Regions Revealed by t Contrasts Showed Enhanced Activation during Conflict Trials Compared with Nonconflict
Trials in Numerical and Physical Comparison Tasks
Voxels
z Score
X
j
z
Brain Area
Numerical task conflict trials > numerical task nonconflict trials—t contrast
Talairach Coordinates
224
112
613
42
34
16
56
14
28
4.32
4.08
4.08
3.93
3.60
3.73
3.66
3.43
3.42
3.28
3.25
32
(cid:2)10
14
18
8
16
(cid:2)22
36
34
(cid:2)44
44
(cid:2)48
12
(cid:2)5
6
6
16
(cid:2)17
52
(cid:2)76
(cid:2)49
9
48
51
11
9
0
49
5
(cid:2)14
(cid:2)1
(cid:2)13
31
Right superior parietal lobule; BA 7
Left superior frontal gyrus; BA 6
Right thalamus; ventral anterior nucleus
Right sublobar; lentiform nucleus; putamen
Right sublobar; Schwanz; caudate head
Right superior frontal gyrus
Left sublobar; extranuclear
Right middle frontal gyrus
Right occipital lobe; subgyral
Left fusiform gyrus
Right inferior frontal gyrus
Physical task conflict trials > physical task nonconflict trials—t contrast
12
3.20
(cid:2)40
30
8
Left inferior frontal gyrus
suitable choice for studying informational conflict than
the traditional color-word task in which direction of
influence is unidirectional—from word to color.
The Stroop effect we observed is consistent with the
number Stroop literature (Girelli et al., 2000; Tzelgov
et al., 1992; Foltz et al., 1984; Henik & Tzelgov, 1982;
Besner & Coltheart, 1979). Daher, subjects were unable
to ignore the irrelevant information regardless of task.
Facilitation was reflected by reaction times but not error
Tarife, consistent with the general finding in the Stroop
literature that facilitation is virtually always substantially
smaller than interference (see review, MacLeod, 1991).
Distance effects (indicated by negative linear trends)
were observed in task-relevant channels, das ist, numer-
ical distance effect in the numerical task and physical
distance effect in the physical task. Reversed distance
effects were observed in both tasks, das ist, a reversed
numerical distance effect in the physical task (konsistent
with Girelli et al., 2000; Henik & Tzelgov, 1982) und das
novel finding of a reversed physical distance effect in the
numerical task. Because distance effect was used as an
indicator of refined information processing, the ob-
served reversed distance effects under task-irrelevant
conditions suggest that numerical distance and physical
distance are processed in an autonomous fashion, sim-
ilar to task-relevant processing.
As proposed in the Introduction, the reversed dis-
tance effect can be explained in terms of amount of
interference. Information that would normally require
little effort to process under task-relevant conditions
(z.B., the salient difference between two numbers with
a large physical distance) is harder to ignore under
task-irrelevant conditions (z.B.,
in numerical compari-
Sohn). Im Gegensatz, information that requires more effort
to process under task-relevant conditions (z.B., numer-
ical comparison of two numerically close numbers)
would exert little interference when such information
is to be ignored under task-irrelevant conditions (In
physical comparison).
Our brain imaging data revealed that when the task-
irrelevant dimensions were kept constant (d.h., in neutral
Versuche), the parietal lobes (regions in the right inferior
parietal lobule, right precuneus, and left superior pari-
etal lobule) showed enhanced activation when process-
ing numerical distance compared to physical distance.
This extends the findings of Pinel et al. (2004) WHO
observed enhanced activation in parietal lobes (Regionen
in bilateral
inferior parietal foci and left intraparietal
Sulkus) during numerical comparisons compared to
physical comparisons, implying a processing difference
between numerical and physical dimensions.
Although there was no evidence to suggest that
processing numerical magnitude and physical size acti-
vated different parietal regions, the enhanced activa-
tion when processing numerical distance relative to
physical distance indicates a quantitative difference in
the processing of these two dimensions. This is consist-
ent with (although does not demonstrate) the distinction
Tang et al.
2057
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
F
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
Tisch 3. Regions Revealed by t Contrasts Showed Enhanced Activation during Error Trials Compared with Correct Trials
in Numerical and Physical Comparison Tasks
Voxels
z Score
X
j
Numerical task error trials > numerical task correct trials—t contrast
Talairach Coordinates
2107
1200
619
3058
341
127
392
72
166
22
53
27
21
6.88
5.52
5.32
3.99
4.93
4.66
4.56
4.56
4.47
4.08
4.13
4.03
3.70
3.65
3.46
3.34
3.25
3.17
3
51
(cid:2)32
(cid:2)32
(cid:2)51
42
55
34
6
(cid:2)4
(cid:2)18
0
4
(cid:2)50
36
(cid:2)46
(cid:2)26
(cid:2)59
(cid:2)42
(cid:2)26
17
10
(cid:2)46
27
20
16
(cid:2)15
(cid:2)15
(cid:2)77
26
(cid:2)22
(cid:2)2
(cid:2)5
13
32
(cid:2)31
Physical task error trials > physical task correct trials—t contrast
1807
1837
909
335
149
5.83
5.08
5.00
4.43
4.89
4.15
4.04
53
50
53
32
(cid:2)34
(cid:2)16
4
(cid:2)42
(cid:2)27
23
19
17
(cid:2)73
(cid:2)35
z
11
(cid:2)7
(cid:2)11
(cid:2)29
13
(cid:2)6
12
(cid:2)29
3
3
(cid:2)25
15
(cid:2)19
31
(cid:2)18
20
24
31
13
(cid:2)5
1
(cid:2)9
(cid:2)11
(cid:2)25
(cid:2)35
Brain Area
Right superior temporal gyrus
Right temporal subgyral
Left inferior frontal gyrus; BA 47
Left superior frontal gyrus
Left superior frontal gyrus
Right inferior frontal gyrus
Right inferior frontal gyrus; BA 45
Right superior temporal gyrus
Right thalamus; medial dorsal nucleus
Left thalamus; medial dorsal nucleus
Left postertior lobe; uvula
Interhemispheric
Right brainstem; pons
Left precentral gyrus; BA 6
Right temporal subgyral
Left frontal subgyral
Left frontal subgyral
Left inferior parietal lobule
Right superior temporal gyrus
Right middle temporal gyrus
Right inferior frontal gyrus
Right inferior frontal gyrus; BA 47
Left inferior frontal gyrus; BA 47
Left posterior lobe; uvula
Right brainstem; medulla
Conjunction by masking across tasks (error trials > correct trials), corrected for family wise error at .050
715
72
181
37
32
6.88
5.52
5.32
4.93
4.66
53
51
(cid:2)32
(cid:2)51
42
(cid:2)42
(cid:2)26
17
(cid:2)46
27
11
(cid:2)7
(cid:2)11
13
(cid:2)6
Right superior temporal gyrus
Right superior temporal subgyral
Left inferior frontal gyrus; BA 47
Left superior temporal gyrus
Right inferior frontal gyrus
A conjunction (by inclusive masking) across the two tasks revealed regions that were commonly activated during error trials.
2058
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
made by Zorzi and Butterworth (1999) between numer-
ical magnitudes that are conceptualized as ‘‘discrete
numerosities’’ and physical sizes that are represented
in an analogue (or continuous) Mode, and with the
suggestion that comparative judgements on discrete
numerosity representations evoked by numbers call for
higher processing requirements compared with those
on analogue representations evoked by physical sizes.
We postulate that the processing streams of these two
types of representation converge at a cognitive level
(where conflict occurs when the streams of information
are incompatible).
In line with our a priori prediction, the parietal
Regionen (in right inferior parietal lobule, right precuneus,
and left superior parietal lobule) that showed enhanced
activation processing numerical relative to physical dis-
tance were not affected by task requirement in conflict
situations. Mit anderen Worten, these parietal regions were
equally active whether or not required by the task to
process numerical magnitudes. The lack of difference in
parietal activation level across numerical and physical
comparison tasks provides a strong evidence for auton-
omous processing of numerical magnitude.
Interessant, we found no evidence to suggest that
parietal activation was parametrically modulated by ei-
ther numerical or physical distance. This appears in-
consistent with numerical-distance-modulated parietal
regions identified by Pinel and colleagues (Pinel et al.,
2004; Pinel et al., 2001). Jedoch, a closer look their
findings suggests that the numerical-distance-modulated
parietal activation might not be as robust as the authors
behauptet. In the Pinel et al. (2001) Studie, numerical
distances were not modeled parametrically. The re-
ported distance-modulated parietal activation was in fact
a main effect of distance (across three levels) rather than
a significant linear decrease in activation with increasing
numerical distance. When masking the main effect of
distance for close > medium and medium > far, Die
only surviving significant region was in the precuneus.
Darüber hinaus, the reported numerical distance-modulated
parietal regions in the bilateral intraparietal sulci and the
right precuneus were from a single-subject analysis. Von
the four subjects tested, only two showed these effects;
one failed to show any strong correlation between brain
activation and numerical distance. The inconsistent find-
ings cast doubts on whether parietal activation is truly
modulated by numerical distance.
The current paradigm is arguably more complex than
that used by Pinel et al. (2001), and this could mean that
any numerical-distance-modulated parietal activation was
less likely to emerge. Pinel et al. (2001) used a compar-
ison task to a fixed reference, in which subjects had to
judge whether a visually presented number was smaller
or larger than the reference (65). Such a task is probably
easier than the current paradigm that involves comparing
bidimensional stimuli. Because Pinel et al.’s stimuli only
varied in one dimension (numerical magnitude), Dort
was neither irrelevant information nor conflict to influ-
ence subjects’ judgements, and the subjects were likely to
perform the task without much difficulty.
When confronted with a conflict, subjects had to
inhibit the task-irrelevant information in order to per-
form the task correctly. The enhanced activation in the
right inferior frontal gyrus in conflict trials compared to
nonconflict trials during numerical comparisons is con-
sistent with findings in go/no-go and stop-signal tasks
(z.B., Bunge et al., 2002; Menon et al., 2001; Garavan
et al., 1999; Rubia et al., 1999; Konishi et al., 1998, 1999).
In Durston et al.’s (2002) go/no-go task, activity associ-
ated with successful response inhibition extended to the
right middle frontal gyrus. In the current experiment,
enhanced activation in this area was also observed in
conflict trials during numerical comparisons. The cur-
rent findings provide support for the suggestion that
regions in the inferior frontal cortex are responsible for
inhibiting prepotent responses and perhaps even con-
flict resolution.
The left fusiform gyrus showed enhanced activation in
conflict trials relative to nonconflict trials during numer-
ical comparisons. Such activation does not only reflect
the recognition of the visual shapes of the numbers, Aber
also suggests that the computation over the perceptual
properties of the numbers interacted with the semantic
processing of the numbers during numerical compari-
sons. This suggestion is further supported by the en-
hanced activation in the right occipital lobe—an area
involved in visual processing—in conflict trials during
numerical comparisons.
During physical comparisons, enhanced activation
in the left inferior frontal gyrus was observed in conflict
trials relative to nonconflict trials, but there was no evi-
dence to suggest that differential activity in the visual
Kortex, supporting the idea that the source of inter-
ference comes from higher cognitive processing of the
Zahlen.
There was also evidence to suggest that conflict may
also occur at the response level. In van Veen and
Carter’s (2005) Studie, bilateral middle frontal gyri and
right superior frontal gyrus were activated in response to
cognitive interference, whereas response conflict acti-
vated the bilateral middle/inferior frontal gyri. The acti-
vations observed in this study—bilateral inferior frontal
gyri, right middle and right superior frontal gyri—imply
that there were two sources to the conflict, nämlich, bei
cognitive and response levels.
No evidence was found to support an involvement of
the ACC in conflict resolution. Small volume correction
searches (5-mm radius) were performed with reference
to ACC coordinates identified by Menon et al. (2001),
van Veen et al. (2001), Bush et al. (1999), and Barch et al.
(1997), but no significant voxel was found. This appears
inconsistent with van Veen and Carter’s (2005) finding
that different parts of the ACC were involved in cognitive
interference and response conflict. Jedoch, in a related
Tang et al.
2059
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
article using the present paradigm (Critchley et al.,
2005), we constructed a model optimized for the detec-
tion of ACC activation and reported such an activation
during conflict trials. Critchley et al. (2005) reported a
region in the ACC [6 22 38] very close to that reported
by van Veen and Carter during semantic conflict [7 18
42]. This provides strong evidence for a semantic level
conflict. dennoch, we have stated earlier that there
was another possible source of conflict, nämlich, Bei der
response level, which has been reflected by the activa-
tion in bilateral inferior frontal gyri.
Darüber hinaus, we found no evidence to support the in-
volvement of the ACC in error commission. We per-
formed small volume correction searches (5-mm radius)
with reference to ACC coordinates identified by Menon
et al. (2001), who reported its associated activity with
error processing and found no significant effects. Wir
have proposed an alternative explanation that the
ACC acts as an interface between cognitive and bio-
behavioral systems based on evidence that its activation
strongly predicted trial-by-trial variability in autonomic
response magnitude (reflected by changes in pupil size)
that peaked during error trials (Critchley et al., 2005).
Zusammenfassend, we set out to investigate whether nu-
merical magnitude and physical size are processed in an
autonomous fashion. By parametrically varying the
amount of interference exerted by the task-irrelevant
dimension on the task-relevant one in either task, A
reversed distance effect—the indicator of refined infor-
mation processing—was observed in each of the dimen-
sions when these were task irrelevant. Such a novel
finding strongly supports autonomous, refined infor-
mation processing in both dimensions. The reversed
distance effect appears to be a robust indicator of
information processing and may prove to be a more
sensitive measure than the Stroop effect itself in Stroop
variants where an interference effect is not easily elic-
ited. Parietal regions have been described as compris-
ing a notation-independent representation of their
semantic content as quantities (Dehaene & Cohen,
1995). We found no evidence to suggest that parietal
regions were modulated by numerical distance. Wie-
immer, we provided strong evidence for the autono-
mous processing of numerical magnitude. Endlich, Wir
conclude that prefrontal regions, especially bilateral
inferior frontal and right middle frontal gyri, are asso-
ciated with conflict resolution when these sources of
conflict could come from both cognitive and response
levels.
Danksagungen
This research was supported by the Wellcome Trust via a
program grant to H. D. C. and R. J. D.
Reprint requests should be sent to Joey Tang, Institute of
Cognitive Neuroscience, Alexandra House, 17 Queen Square,
London WC1N 3AR, Vereinigtes Königreich, oder per E-Mail: joey.tang@ucl.ac.uk.
VERWEISE
Barch, D. M., Mutiger, T. S., Nystrom, L. E., Forman, S. D.,
Noll, D. C., & Cohen, J. D. (1997). Dissociating working
memory from task difficulty in human prefrontal cortex.
Neuropsychologie, 35, 1373–1380.
Bench, C., Frith, C., Grasby, P., Friston, K., Paulesu, E.,
Frackowiak, R., et al. (1993). Investigations of the
functional anatomy of attention using the Stroop test.
Neuropsychologie, 31, 907–922.
Besner, D., & Coltheart, M. (1979). Ideographic and
alphabetic processing in skilled reading of English.
Neuropsychologie, 17, 467–472.
Botvinick, M., Nystrom, L., Fissell, K., Fuhrmann, C., &
Cohen, J. (1999). Conflict monitoring vs. Auswahl-
for-action in anterior cingulate cortex. Natur, 402,
179–181.
Botvinick, M. M., Mutiger, T. S., Barch, D. M., Fuhrmann, C. S.,
& Cohen, J. D. (2001). Conflict monitoring and
cognitive control. Psychological Review, 108, 624–652.
Botvinick, M. M., Cohen, J. D., & Fuhrmann, C. S. (2004).
Conflict monitoring and anterior cingulate cortex: Ein
update. Trends in den Kognitionswissenschaften, 12, 539–546.
Mutiger, T. S., Barch, D. M., Gray, J. R., Molfese, D. L., &
Snyder, A. (2001). Anterior cingulate cortex and response
conflict: Effects of frequency, inhibition and errors.
Hirnrinde, 11, 825–836.
Buckley, P. B., & Gillman, C. B. (1974). Comparisons of
digits and dot patterns. Journal of Experimental
Psychologie, 103, 1131–1136.
Bunge, S. A., Dudukovic, N. M., Thomason, M. E., Vaidya,
C. J., & Gabriela, J. D. E. (2002). Development of frontal
lobe contributions to cognitive control in children:
Evidence from fMRI. Neuron, 33, 301–311.
Busch, G., Frazier, J. A., Rauch, S. L., Seidman, L. J., Whalen,
P. J., Jenike, M. A., et al. (1999). Anterior cingulate cortex
dysfunction in attention deficit/hyperactivity disorder
revealed by fMRI and the counting Stroop. Biologisch
Psychiatrie, 45, 1542–1552.
Fuhrmann, C., Mintun, M., & Cohen, J. (1995). Interference and
15Ö
facilitation effects during selective attention: An H2
PET study of Stroop task performance. Neurobild, 2,
264–272.
Fuhrmann, C. S., Beaver, T. S., Barch, D. M., Botvinick, M. M.,
Noll, D., & Cohen, J. D. (1998). Anterior cingulate cortex,
error detection, and the online monitoring of performance.
Wissenschaft, 280, 747–749.
Fuhrmann, C. S., McDonald, A. M., Botvinick, M., Ross, L. L.,
Stenger, V. A., Noll, D., et al. (2000). Parsing executive
processes: Strategic vs. evaluative functions of the anterior
cingulate cortex. Proceedings of the National Academy
of Sciences, USA., 97, 1944–1948.
Casey, B. J., Thomas, K. M., Welsh, T. F., Badgaiyan, R.,
Eccard, C. H., Jennings, J. R., et al. (2000). Dissociation
of response conflict, attentional control, and expectancy
with functional magnetic resonance imaging (fMRT).
Verfahren der Nationalen Akademie der Wissenschaften, USA.,
97, 8728–8733.
Critchley, H. D., Tang, J., Glaser, D., Butterworth, B., &
Dolan, R. (2005). Anterior cingulate activity during error
and autonomic response. Neurobild, 27, 885–895.
Dehaene, S. (1992). Varieties of numerical abilities.
Cognition, 44, 1–42.
Dehaene, S., & Cohen, L. (1995). Towards an anatomical
and functional model of number processing. Mathematical
Cognition, 1, 83–120.
Derbyshire, S., Vogt, B., & Jones, A. (1998). Pain and Stroop
interference tasks activate separate processing modules in
2060
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
anterior cingulate cortex. Experimentelle Hirnforschung,
118, 52–60.
Duncan, E. M., & MacFarland, C. E. (1980). Isolating the
effects of symbolic distance and semantic congruity in
comparative judgments: An additive factors analysis.
Memory & Cognition, 8, 612–622.
Durston, S., Davidson, M. C., Thomas, K. M., Worden, M.,
Tottenham, N., Martinez, A., et al. (2003). Parametric
manipulation of conflict and response competition using
rapid mixed-trial event related fMRI. Neurobild, 20,
2135–2141.
Durston, S., Thomas, K. M., Yang, Y., Ulug˘, A. M., Zimmerman,
R. D., & Casey, B. J. (2002). A neural basis for the
development of inhibitory control. Developmental
Wissenschaft, 5, F9–F16.
Falkenstein, M., Hoormann, J., Christ, S., & Hohnsbein, J.
(2000). ERP components on reaction errors and their
functional significance: A tutorial. Biological Psychology,
51, 87–107.
Fias, W., Lammertyn, J., Reynvoet, B., Dupont, P., & Orban,
G. A. (2003). Parietal representation of symbolic and
nonsymbolic magnitude. Zeitschrift für Kognition
Neurowissenschaften, 15, 47–56.
Foltz, G. S., Poltrock, S. E., & Potts, G. R. (1984). Mental
comparisons of size and magnitude. Zeitschrift für
Experimental Psychology: Learning, Memory, Und
Cognition, 10, 442–453.
Garavan, H., Ross, T. J., & Stein, E. A. (1999). Right hemispheric
dominance of inhibitory control: An event-related fMRI
Studie. Verfahren der Nationalen Akademie der Wissenschaften,
USA., 96, 8301–8306.
Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., &
Donchin, E. (1993). A neural system for error detection
and compensation. Psychological Science, 4, 385–390.
George, M., Ketter, T., Parekh, P., Rosinsky, N., Ring, H.,
Casey, B., et al. (1994). Regional brain activity when
selecting a response despite interference: An H2
study of the Stroop and an emotional Stroop. Human
Brain Mapping, 1, 194–209.
15O PET
Girelli, L., Lucangeli, D., & Butterworth, B. (2000). Der
development of automaticity in accessing number magnitude.
Journal of Experimental Child Psychology, 76, 104–122.
Glaser, M. O., & Glaser, W. R. (1982). Time course analysis
of the Stroop phenomenon. Journal of Experimental
Psychologie: Human Perception and Performance, 8,
875–894.
Hazeltine, E., Poldrack, R., & Gabriela, J. D. (2000). Neuronal
activation during response competition. Zeitschrift für
Cognitive Neuroscience, 12, 118–129.
Henik, A., & Tzelgov, J. (1982). Is three greater than five:
The relation between physical and semantic size in
comparison tasks. Memory and Cognition, 10, 389–395.
Hinrichs, J. V., Yurko, D. S., & Hu, J. H. (1981). Two-digit
number comparison: Use of place information. Zeitschrift
of Experimental Psychology: Human Perception and
Performance, 7, 890–901.
Kiehl, K. A., Kiddle, P. F., & Hopfinger, J. B. (2000).
Error processing and the rostral anterior cingulate: Ein
ereignisbezogene fMRT-Studie. Psychophysiology, 37, 216–223.
Konishi, S., Nakajima, K., Uchida, ICH., Kameyama, M., &
Miyashita, Y. (1999). Common inhibitory mechanism in
human inferior prefrontal cortex revealed by event-related
fMRT. Gehirn, 122, 981–991.
Konishi, S., Nakajima, K., Uchida, ICH., Sekihara, K., &
Miyashita, Y. (1998). No-go dominant brain activity in
human inferior prefrontal cortex revealed by functional
magnetic resonance imaging. European Journal of
Neurowissenschaften, 10, 1209–1213.
Logan, G. D. (1980). Attention and automaticity in Stroop
and priming tasks: Theory and data. Cognitive Psychology,
12, 523–553.
MacLeod, C. M. (1991). Half a century of research on the
Stroop effect: An integrative review. Psychological Bulletin,
109, 163–203.
Menon, V., Adleman, N. E., White, C. D., Glover, G. H., &
Reiss, A. L. (2001). Kartierung des menschlichen Gehirns, 12, 131–143.
Milham, M. P., Banich, M. T., & Barad, V. (2003). Competition
for priority in processing increases prefrontal cortex’s
involvement in top-down control: An event-related fMRI
study of the Stroop Task. Kognitive Gehirnforschung, 17,
212–222.
Milham, M. P., Banich, M. T., Webb, A., Barad, V., Cohen,
N. J, Wszalek, T., et al. (2001). The relative involvement
of anterior cingulate and prefrontal cortex in attentional
control depends on nature of conflict. Cognitive Brain
Forschung, 12, 467–473.
Moyer, R. S., & Landauer, T. K. (1967). The time required
for numerical inequality. Natur, 215, 1519–1520.
Nelson, J. K., Reuter-Lorenz, P. A., Sylvester, C.-Y. C.,
Jonides, J., & Schmied, E. E. (2003). Dissociable neural
mechanisms underlying response-based and familiarity-
based conflict in working memory. Verfahren der
Nationale Akademie der Wissenschaften, USA., 100, 11171–11175.
Pardo, J., Pardo, P., Janer, K., & Rachel, M. (1990).
The anterior cingulate cortex mediates processing
selection in the Stroop attentional conflict paradigm.
Verfahren der Nationalen Akademie der Wissenschaften,
USA., 87, 256–259.
Pinel, P., Dehaene, S., Rivie`re, D., & Le Bihan, D. (2001).
Modulation of parietal activation by semantic distance in
a number comparison task. Neurobild, 14, 1013–1026.
Pinel, P., Le Clec’H, G., van de Moortele, P. F., Naccache, L.,
Le Bihan, D., & Dehaene, S. (1999). Event-related fMRI
analysis of the cerebral circuit for number comparison.
NeuroReport, 10, 1473–1479.
Pinel, P., Piazza, M., Le Bihan, D., & Dehaene, S. (2004).
Distributed and overlapping cerebral representations
of number, Größe, and luminance during comparative
judgments. Neuron, 41, 983–993.
Posner, M. ICH., & Snyder, C. R. R. (1975). Attention and
cognitive control. In R. L. Solso (Ed.), Information
processing cognition: The Loyola symposium (S. 55–85).
Hillsdale, NJ: Erlbaum.
Rubia, K., Overmeyer, S. O., Taylor, E., Brammer, M.,
Williams, S., Simmons, A., et al. (1999). Hypofrontality
in attention deficit hyperactivity disorder during higher
order motor control: A study with functional MRI.
American Journal of Psychiatry, 156, 891–896.
Rubinsten, O., Henik, A., Berger, A., & Shahar-Shalev, S.
(2002). The development of internal representations of
magnitude and their association with Arabic numerals.
Journal of Experimental Child Psychology, 81, 74–92.
Shallice, T. (1988). From neuropsychology to mental
Struktur. Cambridge: Cambridge University Press.
Shiffrin, R. M., & Shneider, W. (1977). Controlled and
automatic human information processing: II. Perceptual
learning, automatic attending, and general theory.
Psychological Review, 84, 127–190.
Stroop, J. R. (1935). Studies of interference in serial
verbal reactions. Journal of Experimental Psychology,
18, 643–662.
Tzelgov, J., Meyer, J., & Henik, A. (1992). Automatic and
intentional processing of numerical information. Zeitschrift
of Experimental Psychology: Learning, Memory, Und
Cognition, 18, 166–179.
van Veen, V., & Fuhrmann, S. C. (2005). Separating semantic
Tang et al.
2061
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
.
.
.
.
.
F
Ö
N
1
8
M
A
j
2
0
2
1
conflict and response conflict in the Stroop task: A
functional MRI study. Neurobild, 27, 497–504.
van Veen, V., Cohen, J. D., Botvinick, M. M., Stenger, V. A.,
& Fuhrmann, C. S. (2001). Anterior cingulate cortex, conflict
Überwachung, and levels of processing. Neurobild, 14,
1302–1308.
Weissman, D. H., Giesbrecht, B., Song, A. W., Mangun, G. R.,
& Woldorff, M. G. (2003). Conflict monitoring in the
human anterior cingulate cortex during selective attention
to global and local object features. Neurobild, 19,
1361–1368.
Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004). Der
neural basis of error-detection: Conflict monitoring
and the error-related negativity. Psychological Review,
111, 931–959.
Zbrodoff, N. J., & Logan, G. D. (1986). On the autonomy
of mental process: A case study of arithmetic. Zeitschrift
of Experimental Psychology: Allgemein, 115, 118–130.
Zorzi, M., & Butterworth, B. (1999). A computational
model of number comparison. In M. Hahn & S. C.
Stoness (Hrsg.), Proceedings of the Twenty-First Annual
Conference of the Cognitive Science Society (S. 772–777).
Hillsdale, NJ: Erlbaum.
Zysset, S., Mu¨ller, K., Lohmann, G., & von Cramon, D. Y.
(2001). Color-word matching Stroop task: Separating
interference and response conflict. Neurobild, 13, 29–36.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
D
H
T
Ö
T
w
P
N
:
Ö
/
A
/
D
M
e
ich
D
T
F
P
R
Ö
R
M
C
.
H
S
ich
P
l
v
D
e
ich
R
R
e
C
C
H
T
.
A
M
ich
R
e
.
D
C
u
Ö
M
Ö
/
C
J
N
Ö
A
C
R
N
T
ich
C
/
A
e
R
–
P
T
D
ich
C
1
l
8
e
1
–
2
P
2
D
0
F
4
/
9
1
8
1
/
9
1
3
5
2
6
/
8
2
7
0
4
Ö
9
C
/
N
1
2
7
0
5
0
5
6
9
1
9
8
9
/
1
J
2
Ö
2
C
0
N
4
.
9
2
P
0
D
0
6
B
.
j
1
G
8
u
.
e
1
S
2
T
.
Ö
2
N
0
0
4
8
9
S
.
e
P
P
D
F
e
M
B
B
j
e
R
G
2
u
0
e
2
S
3
T
/
J
T
F
.
.
.
.
.
Ö
N
1
8
M
A
j
2
0
2
1
2062
Zeitschrift für kognitive Neurowissenschaften
Volumen 18, Nummer 12