Hippocampal–Cortical Encoding Activity Predicts
the Precision of Episodic Memory
Saana M. Korkki1, Franziska R. Richter2, and Jon S. Simons1
Abstrakt
■ Our recollections of past experiences can vary in both the
number of specific event details accessible from memory and
the precision with which such details are reconstructed. Prior
neuroimaging evidence suggests the success and precision of
episodic recollection to rely on distinct neural substrates during
memory retrieval. Im Gegensatz, the specific encoding mecha-
nisms supporting later memory precision, and whether they dif-
fer from those underlying successful memory formation in
allgemein, are currently unknown. Hier, we combined continuous
measures of memory retrieval with model-based analyses of
behavioral and neuroimaging data to tease apart the encoding
correlates of successful memory formation and mnemonic pre-
Entscheidung. In the MRI scanner, participants encoded object-scene
displays and later reconstructed features of studied objects using
a continuous scale. We observed overlapping encoding activity
in inferior prefrontal and posterior perceptual regions to predict
both which object features were later remembered versus
forgotten and the precision with which they were reconstructed
from memory. Im Gegensatz, hippocampal encoding activity
significantly predicted the precision, but not overall success,
of subsequent memory retrieval. The current results align with
theoretical accounts proposing the hippocampus to be critical
for representation of high-fidelity associative information and
suggest a contribution of shared cortical encoding mechanisms
to the formation of both accessible and precise memory
Darstellungen. ■
EINFÜHRUNG
Our memories are not an exact reproduction of the past
but can range from high-fidelity, precise reconstructions
of previous experiences to less precise, lower-resolution
Darstellungen. Behavioral evidence suggests such varia-
tion in mnemonic precision to be distinguishable from
the general success of memory retrieval (Harlow &
Yonelinas, 2016; Richter, Cooper, Bays, & Simons, 2016;
Brady, Konkle, Gill, Oliva, & Alvarez, 2013; Harlow &
Donaldson, 2013; but see Schurgin, Wixted, & Brady,
2020). Although the likelihood of successful retrieval of
information from memory and the precision of the
retrieved information correlate across individuals, most
variance in each measure is nevertheless unrelated to
the other (Richter et al., 2016). Darüber hinaus, these two
aspects of objective memory performance appear to be
associated with separable subjective characteristics
(Harlow & Yonelinas, 2016) and are differentially sensitive
to various experimental manipulations (z.B., Berens,
Richards, & Horner, 2020; Sun et al., 2017; Xie & Zhang,
2017; Sutterer & Awh, 2016) as well as to memory impair-
ments in distinct populations (Korkki, Richter,
Jeyarathnarajah, & Simons, 2020; Nilakantan, Bridge,
VanHaerents, & Voss, 2018; Cooper et al., 2017), eliciting
1University of Cambridge, 2University of Leiden
© 2021 Massachusetts Institute of Technology
proposals that they may at least to some degree reflect a
dissociable neurocognitive basis.
In der Tat, prior neuroimaging evidence indicates the
success and precision of episodic recollection to recruit
distinct regions of the posterior-medial network during
memory retrieval (Richter et al., 2016). Whereas retrieval
activity in the hippocampus (HC) has been observed to
increase for successful in comparison to unsuccessful
retrieval, trial-wise variation in memory precision appears
to correlate with retrieval-related activity in the lateral pa-
rietal cortex (Richter et al., 2016), although others high-
light a role for medial temporal regions also (Montchal,
Reagh, & Yassa, 2019; Stevenson et al., 2018). Jedoch,
despite increased interest in the neural basis of mnemonic
precision, the focus of prior studies has been on retrieval
mechanisms (z.B., Cooper & Ritchey, 2019; Montchal
et al., 2019; Stevenson et al., 2018; Richter et al., 2016),
whereas the encoding substrates supporting the forma-
tion of precise memory representations, and whether
they differ from those supporting successful encoding
in general, remain unresolved.
Successful episodic memory formation is typically asso-
ciated with activity increases in a network of medial tem-
poral, lateral prefrontal, and posterior perceptual regions
(Kim, 2011; Spaniol et al., 2009). The HC receives input
from content-specific perceptual regions and is thought
to bind disparate event features into a coherent memory
representation (Cooper & Ritchey, 2020; Ranganath,
Zeitschrift für kognitive Neurowissenschaften 33:11, S. 2328–2341
https://doi.org/10.1162/jocn_a_01770
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
J
Ö
C
N
_
A
_
0
1
7
7
0
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
2010; Davachi, 2006; Paller & Wagner, 2002) and allow for
the storage of similar experiences in an orthogonalized,
or nonoverlapping, manner (Norman & O’Reilly, 2003;
O’Reilly & McClelland, 1994). Lateral prefrontal regions,
andererseits, are involved in the strategic and
controlled encoding of information into memory via pro-
cesses such as attentional selection, elaboration, and inte-
gration of information relevant for current task goals
(Blumenfeld & Ranganath, 2007; Simons & Spiers, 2003).
The specific neural substrates supporting successful
memory formation have been found to exhibit process
specificity, varying for instance according to the depth of
stimulus processing engaged in at encoding (Park,
Uncapher, & Rugg, 2008; Otten & Rugg, 2001) and the type
of retrieval process later recruited (Staresina & Davachi,
2006; Ranganath et al., 2004). Darüber hinaus, encoding correlates
appear sensitive to more subtle differences in the quality of
retained representations, including their objective amount
of detail (Cooper & Ritchey, 2020; Qin, van Marle,
Hermans, & Fernandez, 2011), and subjective ratings of
memory vividness or confidence (Kensinger, Addis, &
Atapattu, 2011; Qin et al., 2011). Jedoch, while beginning
to elucidate the encoding mechanisms underlying variation
in more qualitative aspects of later retrieval, prior studies
have typically been limited by the use of categorical measures
of the quantity of details remembered, or participants’ subjec-
tive reports, which may not directly map onto more graded
variations in objective memory precision.
It is possible that, in addition to relying on distinct
brain regions during retrieval (Richter et al., 2016), Die
success and precision of episodic recollection may be
supported by at least partly separable neural mechanisms
during memory encoding. Zum Beispiel, the successful
retrieval of information from memory may depend on
the strength of an association between a retrieval cue
and the target memory, thus drawing in particular on
associative encoding processes supported by the HC
and the prefrontal cortex (Blumenfeld & Ranganath,
2007; Davachi, 2006). Im Gegensatz, the precision with
which specific mnemonic features can be reconstructed
from memory may closely relate to the fidelity of stimulus
encoding in posterior perceptual regions (Emrich,
Riggall, LaRocque, & Postle, 2013) and/or to hippocampal
function supporting the formation of distinct and de-
tailed memory traces that can be later reconstructed with
high precision (Moscovitch, Cabeza, Winocur, & Nadel,
2016). In der Tat, an association between hippocampal en-
coding activity and subsequent mnemonic precision
would align with prior accounts suggesting the HC to
be critical for representation of high-fidelity relational in-
formation across perception and memory (Ekstrom &
Yonelinas, 2020; Kolarik et al., 2016; Aly, Ranganath, &
Yonelinas, 2013; Yonelinas, 2013). Alternativ, it is pos-
sible that, contrary to dissociable neural substrates ob-
served during retrieval (Richter et al., 2016), Die
successful and precise encoding of information into
memory may rely on shared neural mechanisms that
perhaps act to increase the strength of the memory more
generally, rendering it both accessible and precise at
retrieval.
In the current study, we employed continuous mea-
sures of memory retrieval and model-based analyses of
behavioral and neuroimaging data to elucidate the en-
coding substrates of mnemonic precision. In the MRI
scanner, participants encoded visual stimulus displays de-
picting an object overlaid on a scene background. The lo-
cation and color of the objects were drawn from circular
Räume, and at retrieval, participants recreated these attri-
butes of the studied items using a continuous response
dial. This approach allowed us to segregate encoding
activity supporting later successful memory retrieval from
that supporting subsequent mnemonic precision in a
manner not afforded by more typical categorical measures
of retrieval performance (z.B., old/new, remember/know),
thus providing novel insight into the encoding mecha-
nisms supporting the acquisition of precise episodic
memories.
METHODEN
Teilnehmer
Twenty-one young adults (18–29 years old) participated in
the current experiment. All participants were right-
handed, native English speakers, and had normal or
corrected-to-normal vision, no color blindness, and no
current or historical diagnosis of any neurological, psychi-
atric, or developmental disorder, or learning difficulty.
Participants indicated no current use of any psychoactive
medication and no medical or other contradictions to MRI
scanning. One participant was excluded from all analyses
because of excessive movement (>4 mm) in the scanner,
leaving 20 participants to contribute to the present analy-
ses (eight men, 12 Frauen; Durchschnittsalter = 22.15 Jahre, SD =
3.10 Jahre). The participants were recruited via the
University of Cambridge Psychology Department Sona vol-
unteer recruitment system (Sona Systems, Ltd.) and com-
munity advertisements and were reimbursed with £30 for
their participation. All participants gave written informed
consent in a manner approved by the Cambridge
Psychology Research Ethics Committee.
Materials
Stimuli for the memory task comprised 180 images of
outdoor scenes and 180 images of distinct everyday
Objekte. The images were obtained from existing stimuli
sets (scenes: Richter et al., 2016; Objekte: Brady et al.,
2013) and Google image search. Each object image was
randomly paired with a scene image to generate 180 trial-
unique study displays (size = 750 × 750 pixels). Across
the study displays, we varied the appearance of two ob-
ject features: color and location. For each display, Objekt
color and location were randomly selected from circular
Korkki, Richter, and Simons
2329
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
J
Ö
C
N
_
A
_
0
1
7
7
0
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
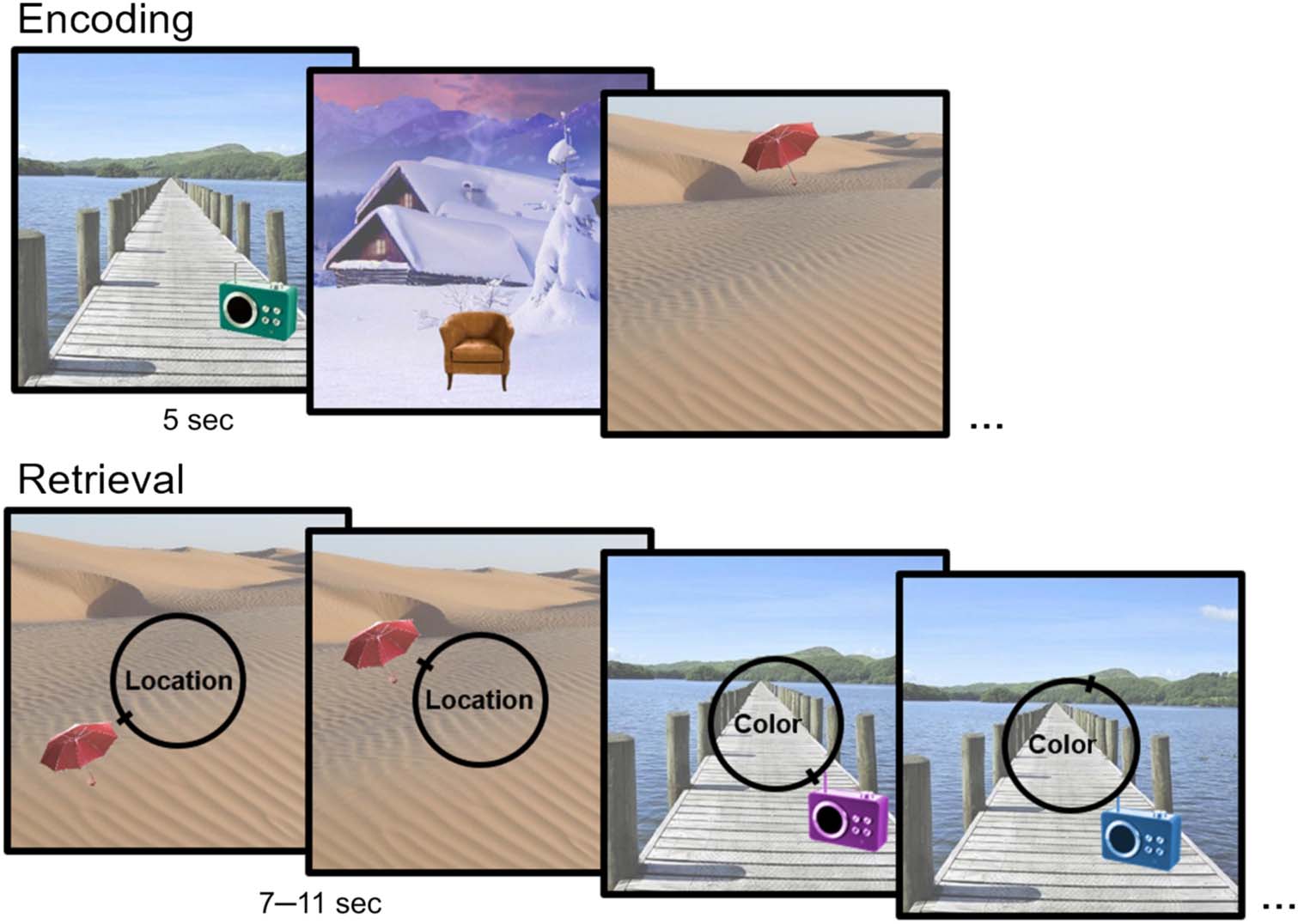
Figur 1. Example study and
test trials of the memory task.
At study, participants viewed
stimuli displays consisting of
one object overlaid on a scene
background (stimulus duration:
5 Sek). The location and color
of the objects at study were
randomly chosen from circular
parameter spaces (0–360°). Bei
test, participants recreated
either the location or color
of each studied object using a
360° continuous response dial,
allowing for a fine-grained
assessment of memory fidelity.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
J
Ö
C
N
_
A
_
0
1
7
7
0
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
parameter spaces (0–360°; vgl. Cooper et al., 2017; Richter
et al., 2016; siehe Abbildung 1). All participants viewed the
same study displays in a randomized order.
Design and Procedure
Before the scan, participants read the instructions and
undertook practice trials of the memory task. The task
was modified from Richter et al. (2016) by reducing the
number of objects presented at study and the number of
features later tested, to result in one feature retrieval trial
per study display. In Summe, participants completed nine
study-test blocks over nine functional runs (one study
and one test phase per run). At study, participants sequen-
tially viewed 20 object-background displays (stimulus
Dauer: 5 Sek) and were instructed to try and memorize
the appearance of each display, including the location and
color of the object. The study phase was followed by a
10-sec delay during which a “Get Ready” message was pre-
sented on a black screen. After this delay, Teilnehmer
were asked to reconstruct either the location or the color
of each object viewed in the preceding study phase (eins
feature question per each encoding trial, insgesamt 20
retrieval trials per block). At retrieval, the test object
reappeared on its associated background with a central
cue word “Location” or “Color” indicating the type of
feature tested on that trial. The initial appearance of the
tested feature was randomly selected from a circular
parameter space (0–360°), whereas the appearance of
the untested feature remained unchanged from study to
test. Mit anderen Worten, for location trials, the test object
reappeared in its original color, but in a randomly selected
location, whereas for color trials, the test object reap-
peared in its original location but in a randomly chosen
color. Participants were asked to recreate the object’s
original features as accurately as they could by moving a
slider around a 360° response dial using their middle
and index fingers on a button box and were able to confirm
their answer by pressing a third key with their thumb. Der
retrieval phase was self-paced with the constraint of a
minimum trial length of 7 sec and a maximum RT of
11 Sek. Participants on average produced RTs that were
well under this limit (M = 5.64 Sek, SD = 0.68 Sek), Und
the percentage of trials where response selection was not
confirmed in time was very low (M = 1.36%, SD = 1.77%).
Note that if a participant failed to confirm their answer
innerhalb 11 Sek, their last position on the response wheel
was recorded as their answer for that trial.
Participants completed 90 location and 90 color trials
in Summe (10 trials of each type per task block). The type
of feature tested for each object was randomized across
displays but constant across participants so that all partic-
ipants answered the same feature question for each study
display. To ensure that memory was tested for feature
values spanning the entire circular space, the randomiza-
tion was conducted with a constraint of roughly equal
number (d.h., 20–25) of target feature values sampled
from each quadrant around the circular space for both
the location and color conditions. The order of study
and test displays was then randomized across participants
with the constraint of no more than four encoding or
retrieval trials in a row for which the same type of feature
was tested. Study and test trials were separated by a fix-
ation cross with jittered duration between 0.4 Und 2.4 Sek
2330
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 11
(mean ISI duration: 1 Sek) drawn from a Poisson distribu-
tion. After the first five of the nine task blocks, partici-
pants were given a 10-min break from the memory task
in the scanner, during which a diffusion-weighted struc-
tural scan was acquired (analysis of diffusion-weighted
data not reported here).
Behavioral Analysis
For each trial, we calculated participants’ retrieval error
as the angular difference between their response value
and the target feature value (0 ± 180°). To distinguish
the likelihood of successful memory retrieval from the
precision of the retrieved information, we fitted a two-
component mixture model (Bays, Catalao, & Husain,
2009; Zhang & Luck, 2008) to each participant’s retrieval
error data using maximum likelihood estimation (Code
available at www.paulbays.com/code/JV10/index.php).
This mixture model has been shown to characterize
long-term memory performance in similar tasks (z.B.,
Korkki et al., 2020; Richter et al., 2016; Brady et al.,
2013) and has been employed to gain insights about
the neural basis of the precision of episodic recollection
(Cooper & Ritchey, 2019; Stevenson et al., 2018; Richter
et al., 2016). The model assumes that two distinct
sources of error contribute to participants’ retrieval per-
formance across trials: variability, das ist, noise, in suc-
cessful retrieval of target features and the presence of
random guess responses where memory retrieval has
failed to bring any diagnostic information about the tar-
get to mind. These two sources of error are modeled by a
von Mises distribution (circular equivalent of a Gaussian
distribution) centered at a mean error of 0° from the
target value, with a concentration, K, and a circular uni-
form distribution with a probability, pU, jeweils.
Precision of memory retrieval can be estimated as the
concentration parameter (K; higher values reflect higher
precision) of the target von Mises distribution and the
likelihood of successful memory retrieval ( pT ) as the
probability of responses stemming from the target von
Mises distribution ( pT = 1 − pU ). Consistent with prior
Studien (Korkki et al., 2020; Richter et al., 2016), Das
two-component model was found to fit the current data
better than an alternative one-component model where
participants’ responses were assumed to stem from a
von Mises distribution around the target feature value
nur (mean Bayesian information criterion for the one-
component model: 386.10; mean Bayesian information
criterion for the two-component model: 317.62; Modelle
fitted to individual participants’ data across the feature
Bedingungen).
MRI Acquisition
MRI scanning took place at the University of Cambridge
Medical Research Council Cognition and Brain Sciences
Unit using a 3-T Siemens Tim Trio scanner with a
32-channel head coil. For each participant, a whole-brain
structural image was acquired using a T1-weighted 3-D
magnetization prepared rapid gradient echo sequence
(repetition time = 2.25 Sek, Echozeit = 3 ms, umdrehen
angle = 9°, Sichtfeld = 256 × 256 × 192 mm, reso-
lution = 1 mm isotropic, GRAPPA acceleration factor 2).
Functional data were acquired over nine runs each com-
prising one task block (one encoding and one retrieval
Phase), using a single-shot EPI sequence (repetition
time = 2 Sek, Echozeit = 30 ms, flip angle = 78°, field
of view = 192 × 192 mm, resolution = 3 mm isotropic).
Each volume consisted of 32 sequential oblique-axial
Scheiben (interslice gap: 0.75 mm) acquired parallel to the
AC–PC transverse plane. Across the participants, der Mittelwert
number of volumes acquired per functional run was 166.09
(SD = 8.08). The scanning protocol further included a
diffusion-weighted structural scan that was acquired after
the first five functional runs (not analyzed here).
fMRI Preprocessing
Data preprocessing and analysis were performed with
SPM 12 (www.fil.ion.ucl.ac.uk/spm/) implemented in
MATLAB R2016a (The MathWorks). The first five volumes
of each functional run were discarded to allow for T1
equilibration. Außerdem, any additional volumes col-
lected after each task block had finished were discarded
for each participant so that the last volume of each run
corresponded to a time point of ∼2 sec after the last fix-
ation cross. The functional images were spatially rea-
ligned to the mean image to correct for head motion
and temporally interpolated to the middle slice to correct
for differences in slice acquisition time. The anatomical
image was coregistered to the mean EPI image, bias-
corrected and segmented into different tissue classes
(gray matter, white matter, cerebrospinal fluid). Diese
segmentations were used to create a study-specific struc-
tural template image using the DARTEL (Diffeomorphic
Anatomical Registration Through Exponentiated Lie
Algebra) toolbox (Aschenbrenner, 2007). The functional data
were normalized to Montreal Neurological Institute space
using DARTEL and spatially smoothed with an isotropic
8-mm FWHM Gaussian kernel.
Main fMRI Analyses
To obtain trial-specific estimates of the success and preci-
sion of memory retrieval for the fMRI analyses, we fitted
the two-component mixture model (von Mises + uniform
distribution) to retrieval error data across all participants
and feature conditions (3600 trials in total). Using the
best-fitting model probability density function, we then
calculated the probability of each error belonging to the
target von Mises distribution over the uniform distribution
and classified errors with at least .05 probability of stem-
ming from the von Mises distribution as “successful” and
errors with less than .05 probability of belonging to the
Korkki, Richter, and Simons
2331
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
J
Ö
C
N
_
A
_
0
1
7
7
0
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
von Mises distribution as “unsuccessful” (vgl. Cooper et al.,
2017; Richter et al., 2016). In terms of degrees, this corre-
sponded to a subsequent retrieval success cutoff of ±51°,
where trials with an absolute error ≤ 51° (range = 0–51°)
were classified as “successful” and trials with an absolute
error > 51° were classified as “unsuccessful” (range =
52–180°). As done in prior studies (Cooper & Ritchey,
2019, 2020; Cooper et al., 2017; Richter et al., 2016), Wir
used the across-participant model-derived cutoff to
ensure that responses of the same error magnitude
were consistently classified as successful or unsuccessful
across individuals as well as to avoid any bias in the error
cutoffs related to differences in individual model fits.
We further note that using feature-specific cutoffs,
rather than the threshold estimated across all retrieval
Versuche, did not change the significance of our main
results. For trials classified as successfully encoded, A
trial-specific measure of memory precision was further
calculated as 180° minus [participant’s absolute retrieval
error on that trial] so that higher values (smaller error)
reflected higher precision (range = 129–180°; vgl. Cooper
et al., 2017; Richter et al., 2016).
For each participant, a first-level general linear model
was constructed containing three regressors correspond-
ing to each event of interest (successful location encod-
ing, successful color encoding, and unsuccessful
encoding) and a fourth regressor modeling the retrieval
Versuche. For the successful encoding trials, the trial-specific
estimates of memory precision were included as para-
metric modulators comprising two additional regressors
im Modell. The precision parametric modulators were
rescaled to range between 0 Und 1 to facilitate the direct
comparison of success and precision-related effects and
were mean centered for each participant. Neural activity
was modeled with a boxcar function convolved with the
canonical hemodynamic response function, with a dura-
tion of 5 sec for the encoding trials and a variable dura-
tion (7–11 sec) for the retrieval trials, capturing the
duration of the study and test displays, jeweils. Six
participant-specific movement parameters estimated dur-
ing realignment (three rigid-body translations, drei Rota-
tionen) were further included as covariates in the first-level
model to capture any residual movement-related arti-
Fakten. Because of the small number of guessing trials in
each functional run, data from all functional runs were
concatenated for each participant, and nine constant
block regressors were included as additional covariates.
Autocorrelation in the data was estimated with an AR(1)
Modell, and a temporal high-pass filter with a 1/128-Hz
cutoff was used to eliminate low-frequency noise. Erste-
level participant-specific parameter estimates were sub-
mitted to second-level random effects analyses.
Contrasts
The contrasts for the fMRI analyses focused on identify-
ing regions where encoding activity positively predicted
the subsequent success and/or precision of episodic
memory retrieval (d.h., increases in BOLD signal for suc-
cessful encoding or higher memory precision). To exam-
ine encoding activity associated with the subsequent
success of memory retrieval, we contrasted encoding tri-
als for which memory retrieval subsequently succeeded
against trials for which memory retrieval subsequently
failed (“subsequent retrieval success effects”). Zu identifizieren
encoding activity predicting the later precision of memory
retrieval, positive associations between BOLD signal
and the precision parametric modulator were examined
(d.h., linear relationship between BOLD signal and preci-
sion parametric modulator; “subsequent precision
effects”). We further assessed the overlap between subse-
quent success and subsequent precision effects using
conjunction analyses. Conjunction analyses were con-
ducted testing against the conjunction null hypothesis
to ensure that regions identified in this analysis displayed
reliable encoding activity associated with each individual
Kontrast, das ist, both subsequent success and subse-
quent precision of memory retrieval (see Nichols, Brett,
Andersson, Wager, & Polina, 2005). Darüber hinaus, Wir
assessed the specificity of the subsequent success and
subsequent precision effects by conducting exclusive
masking of each subsequent memory contrast by the
andere (d.h., subsequent retrieval success masked by
subsequent precision contrast and vice versa). Dafür
Analyse, the mask image was thresholded at p < .050
uncorrected (cf. Uncapher, Otten, & Rugg, 2006; Smith,
Henson, Dolan, & Rugg, 2004).
Because of a relatively low number of guess trials per
feature condition for some individuals, it was not possi-
ble to investigate feature-specific subsequent success
effects. Furthermore, analysis of feature-specific subse-
quent precision effects did not yield any significant differ-
ences across the ROIs ( ps > .208) or the whole brain
( ps > .303). Daher, our analyses focused on examining
BOLD activity predicting the subsequent success and
precision of memory retrieval across the feature condi-
tionen, consistent with the approach taken in previous
studies employing a similar paradigm (Cooper et al.,
2017; Richter et al., 2016).
ROIs
The main analyses focused on a small number of a priori
ROIs implicated by meta-analytic evidence in supporting
the successful formation of episodic memories for visual
Information (Kim, 2011; Spaniol et al., 2009). Speziell,
the ROIs included the HC, the inferior frontal gyrus
(IFG), and the fusiform gyrus (FFG). Given evidence for
greater consistency of subsequent memory effects in the
left hemisphere (Spaniol et al., 2009), left-lateralized ROIs
wurden benutzt, each comprising the left anatomical region as
defined by the automated anatomical labeling atlas
(Tzourio-Mazoyer et al., 2002). Statistical significance within
each anatomical ROI was assessed using small-volume
2332
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 11
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
J
Ö
C
N
_
A
_
0
1
7
7
0
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
correction with a peak-level family-wise error (FWE)-
corrected (based on random field theory) threshold of
P < .05, correcting for the number of voxels in each ROI.
In addition to the ROI analyses, we sought to identify any
additional brain regions displaying a relationship between
encoding activity and the subsequent success and/or preci-
sion of memory retrieval in exploratory whole-brain analy-
ses conducted at a whole-brain FWE-corrected threshold
of p < .05, with a minimum extent of 5 contiguous voxels.
Additional Control Analyses
In addition to the main fMRI analyses described above, we
conducted two additional analyses to assess whether
BOLD signal in any of the ROIs was associated with trial-
wise variation in participants’ memory error for trials clas-
sified as “unsuccessful” based on the model-derived cutoff
(absolute error > 51°), das ist, when variation in memory
error was assumed to be driven by guessing, or when col-
lapsing across all encoding trials without assuming a
model-based separation between successful and unsuc-
cessful retrieval. For the first analysis, the first-level general
linear model was identical to what was described above,
but with the addition of a parametric modulator for unsuc-
cessful trials also that reflected trial-wise variation in partic-
ipants’ subsequent memory error (180° – absolute error;
range: 0–128°). For the second, model-free, Analyse, alle
encoding trials in the location and color condition were
modeled with one regressor each, and a parametric
modulator reflecting trial-wise variation in subsequent
memory error (180° – absolute error; range: 0–180°) War
added for each condition. Parametric modulators were
mean centered, and the contrast of interest investigated
linear increases in BOLD signal with decreasing memory
Fehler.
ERGEBNISSE
Verhaltensergebnisse
For each trial, we calculated participants’ retrieval error as
the angular difference between their response value and
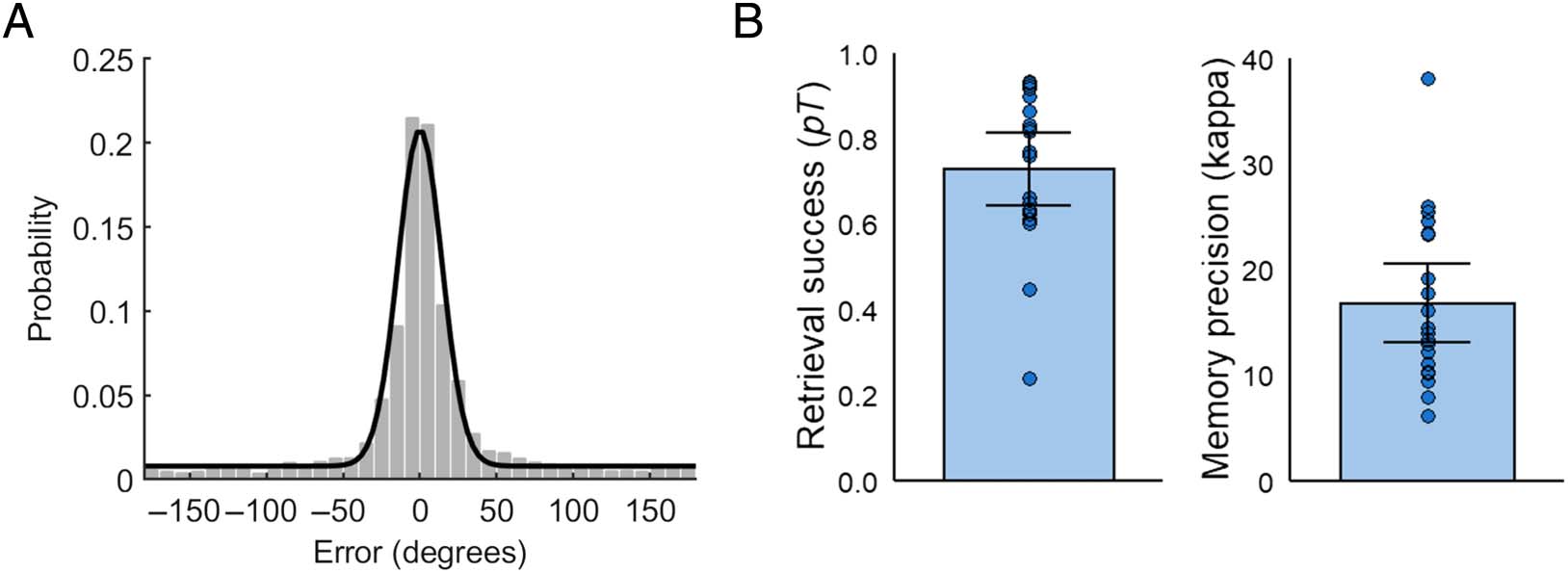
the target feature value (0 ± 180°; see Figure 2A). Across
participants and feature conditions, overall task perfor-
Mance, as measured by the mean absolute retrieval error,
was 30.43° (SD = 15.04°), with a significantly higher mean
absolute error in the color (M = 34.48°, SD = 15.90°) In
comparison to the location condition (M = 26.37°, SD =
15.80°), T(19) = 3.63, p = .002, d = 0.81. To further
decompose the specific sources of error contributing to
participants’ overall performance, we fitted the two-
component mixture model (von Mises + uniform distri-
bution) to each individual participant’s retrieval error data
using maximum likelihood estimation (Bays et al., 2009).
The mean model-estimated probability of successful
memory retrieval, defined as the probability of responses
stemming from a von Mises distribution centered at the
target feature value ( pT ), War .73 (SD = .18) across par-
ticipants and feature conditions (see Figure 2B). Der
mean model-estimated precision of memory retrieval,
estimated as the concentration parameter, Κ, of the target
von Mises distribution, War 16.79 (SD = 7.92) across par-
ticipants and feature conditions (see Figure 2B; note that
this value of Κ is comparable to an SD of approximately
14.20°). Mean memory precision (K ) was significantly
higher in the location (M = 34.65, SD = 27.24) in compar-
ison to the color condition (M = 10.94, SD = 7.15), T(19) =
4.04, p = .001, d = 0.90, whereas mean probability of
successful memory retrieval ( pT ) did not significantly
differ between the two feature conditions (location: M =
0.75, SD = 0.18; color: M = 0.73, SD = 0.20), T(19) = 0.65,
p = .524. Consistent with previous results (Richter et al.,
2016), we also observed a moderate positive correlation
Figur 2. (A) Distribution of retrieval errors (response – target) across all trials and participants. Black line illustrates response probabilities predicted
by the two-component mixture model (von Mises + uniform distribution; model fitted to data across all participants for visualization). (B) Mean
model-estimated probability of successful memory retrieval ( pT ) and memory precision (Κ) across participants. Error bars display 95% confidence
interval of the mean and data points of individual participant parameter estimates.
Korkki, Richter, and Simons
2333
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
J
Ö
C
N
_
A
_
0
1
7
7
0
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
between estimates of the probability of successful mem-
ory retrieval and memory precision across participants,
rs = .54, p = .014.
fMRI Results
Encoding Activity Predicting Subsequent Retrieval
Success and Memory Precision in a Priori ROIs
Our ROI analyses focused on examining whether encoding
activity in three regions typically displaying subsequent
memory effects for visual information, nämlich, the HC,
the IFG, and the FFG, differentially contributes to the later
success and precision of episodic memory retrieval. We first
examined increases in encoding activity for trials that were
subsequently successfully retrieved (absolute retrieval error
≤ 51°) in contrast to trials that were subsequently forgotten
(absolute retrieval error > 51°; note that only one object
feature, d.h., location or color, was reconstructed for each
encoding display). Within our anatomical ROIs, Wir
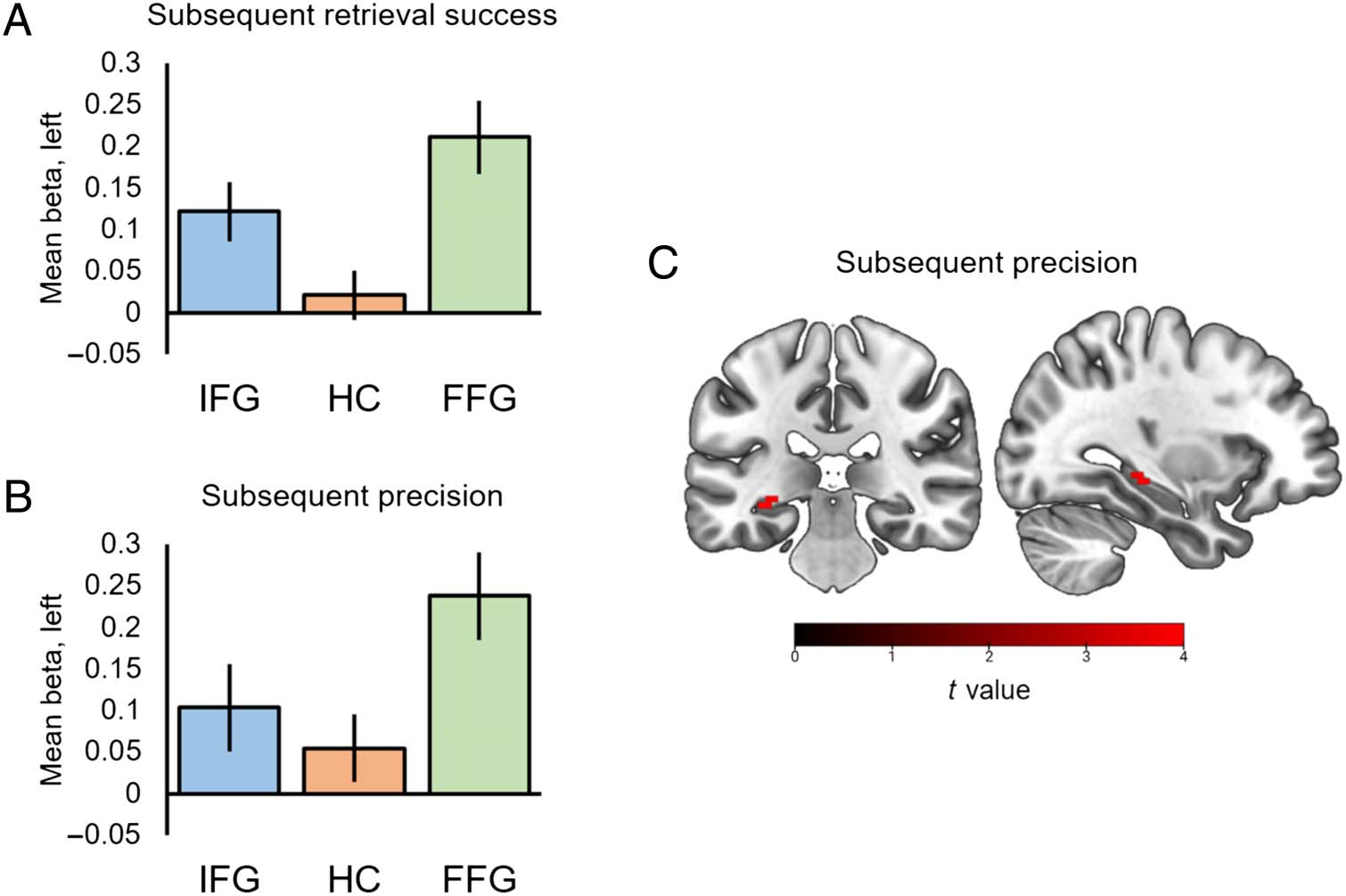
observed increased encoding activity in the IFG, T(19) =
6.75, p = .001, peak: −36, 27, 18, and the FFG, T(19) =
8.88, P < .001, peak: −30, −63, −9, to predict whether
object features were later successfully retrieved from
memory or forgotten (peak-level FWE-corrected within
each ROI; see Figures 3A and 4A). In contrast, no significant
subsequent retrieval success effects were detected in the
HC ( ps > .151).
We next examined whether encoding activity in these
regions predicted the graded precision with which object
features were later successfully retrieved from memory
(linear relationship between BOLD signal and precision
parametric modulator). In addition to predicting which
trials were successfully remembered, encoding activity
in the IFG, T(19) = 5.63, p = .011, peak: −57, 15, 15,
and the FFG, T(19) = 6.27, p = .001, peak: −33, −75,
−18, positively correlated with the precision of later
memory retrieval (see Figures 3B and 4B). Außerdem,
increased encoding activity in the HC, T(19) = 4.20, p =
.029, peak: −33, −30, −9, was associated with greater
mnemonic precision for object features (see Figure 3B
and C). As a control analysis, we further investigated
whether BOLD signal in any of the ROIs predicted trial-
wise variation in memory error across trials classified as
unsuccessful (d.h., when variation in memory error was
assumed to be driven by guessing). No significant associ-
ations between BOLD signal and subsequent memory
error were detected for trials classified as unsuccessful
in any of the ROIs ( ps > .238).
Daher, results from the ROI analyses suggest encoding
activity in the inferior frontal and fusiform cortex to sup-
port both the later success and precision of memory re-
trieval, whereas significant increases in BOLD signal in
the HC were observed for subsequent memory precision
nur. We next sought to assess whether encoding activity
predicting these two aspects of later retrieval perfor-
mance overlapped in any of the ROIs. Conjunction anal-
yses indicated significant overlap between subsequent
success and subsequent precision effects in both the
IFG, T(19) = 4.86, p = .007, peak: −42, 3, 27, und das
FFG, T(19) = 6.12, P < .001, peak: −42, −57, −12,
whereas no significant overlap was detected in the HC
( ps > .778). Außerdem, hippocampal encoding activity
still predicted the subsequent precision of memory
retrieval after exclusive masking with the subsequent
retrieval success contrast (mask thresholded at p > .050
unkorrigiert), T(19) = 4.20, p = .029, peak: −33, −30,
−9. Andererseits, significant subsequent retrieval
Figur 3. Mean parameter
estimates for (A) subsequent
success (successful >
unsuccessful) Und (B)
subsequent precision (positive
association between BOLD
signal and precision parametric
modulator) effects in the left
IFG, HC, and FFG. Error bars
display ±1 SEM. (C) Encoding
activity correlating with the
subsequent precision of
memory retrieval in the
hippocampal ROI (visualized
at an uncorrected threshold
of p < .001).
2334
Journal of Cognitive Neuroscience
Volume 33, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
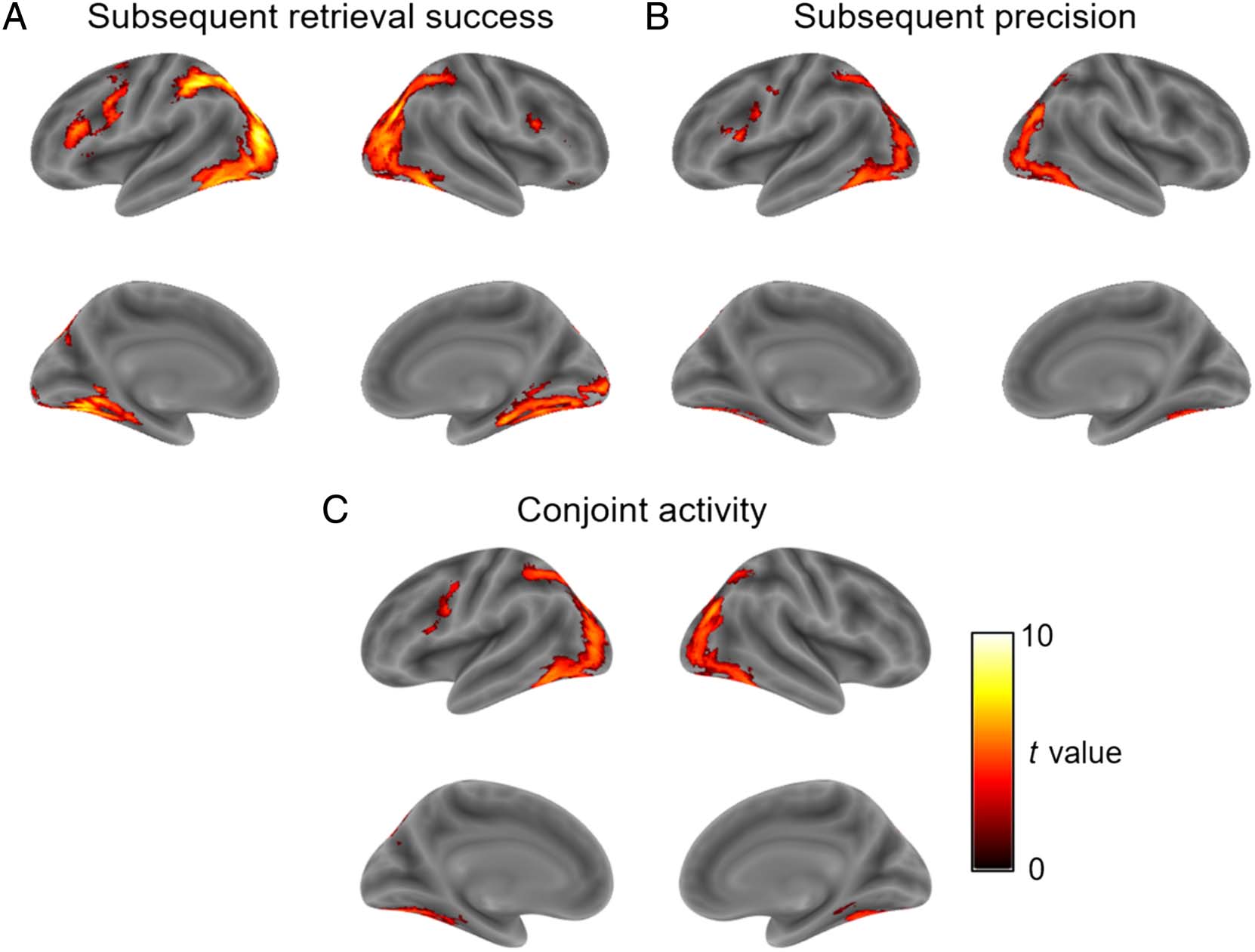
Figure 4. Encoding activity predicting the (A) subsequent success (successful > unsuccessful) Und (B) subsequent precision (positive association
between BOLD activity and precision parametric modulator) of memory retrieval, Und (C) overlap between encoding activity predicting both later
success and precision of memory retrieval. Visualized at an uncorrected threshold of p < .001, with a minimum cluster of 10 voxels.
success effects were detected in the IFG, t(19) = 5.76, p =
.006, peak = −36, 27, 15, and the FFG, t(19) = 4.35, p =
.039, peak = −18, −45, −12, after exclusive masking with
the subsequent precision contrast, consistent with the
observation of more widespread subsequent retrieval
success than subsequent precision effects in these two
regions (see Figure 4). No significant subsequent preci-
sion effects were observed in these two regions ( ps >
.052) after exclusive masking with the subsequent retrieval
success contrast.
Encoding Activity Predicting Variation in Memory Error
across All Encoding Trials in a Priori ROIs
In addition to the model-based analyses described above,
we investigated whether trial-by-trial variation in BOLD
signal in any of the ROIs was associated with trial-by-trial
variation in subsequent memory error across all encoding
Versuche, without assuming a categorical distinction between
successful and unsuccessful retrieval. Consistent with the
pattern of results observed in the model-based analyses,
which indicated encoding activity in the IFG and FFG to
be sensitive to both the subsequent success and subse-
quent precision of memory retrieval, we observed encod-
ing activity in these two regions to also predict the
magnitude of subsequent memory error when collapsing
across all encoding trials [IFG: T(19) = 7.03, p = .001,
peak: −51, 9, 27; FFG: T(19) = 12.18, P < .001, peak:
−42, −57, −12]. In contrast, trial-wise variation in mem-
ory error was not significantly associated with encoding
activity in the HC when examining all encoding trials
( ps > .323).
Encoding Activity Predicting Subsequent Retrieval
Success and Memory Precision across the Whole Brain
To identify any additional brain regions, beyond our a
priori ROIs, where encoding activity predicted the later
success and/or precision of memory retrieval, we further
performed complementary whole-brain analyses. Activity
in several regions of the dorsal and ventral visual streams,
including the middle occipital gyrus, inferior parietal
gyrus, FFG, and inferior temporal gyrus, was found to
predict which object features were later successfully
remembered versus forgotten (siehe Tabelle 1 and Figure 4A).
For subsequent memory precision, no significant voxels
survived a whole-brain peak-level corrected significance
threshold ( P < .050 FWE-corrected, k > 5), although
we note that three clusters spanning the left inferior
temporal gyrus, middle occipital gyrus, and cerebellum,
T(19) = 7.08, P < .001, the right middle occipital gyrus
and FFG, t(19) = 5.84, p < .001, and the left IFG, t(19) =
Korkki, Richter, and Simons
2335
Table 1. Encoding Activity Associated with the Subsequent Success (Successful > Unsuccessful) and Subsequent Precision (Positive
Relationship between BOLD Activity and Precision Parametric Modulator) of Memory Retrieval in the Whole-Brain Analyses, P < .050
FWE-Corrected at Peak Level, k > 5
Region
Voxels
X
Subsequent retrieval success
L middle occipital gyrus
L inferior parietal gyrus
R middle occipital gyrus
R inferior temporal gyrus
R middle occipital gyrus
R FFG
Subsequent precision
No significant voxels
Conjoint activity
L middle occipital gyrus
R middle occipital gyrus
L FFG
L middle occipital gyrus
L = left; R = right.
355
87
63
15
21
8
30
41
30
22
−36
−33
33
45
42
30
−24
33
−42
−39
j
−87
−45
−75
−54
−81
−27
−75
−78
−57
−84
z
12
39
30
−12
9
−21
30
24
−12
6
T
P
10.50
9.41
8.83
8.27
7.95
7.33
6.59
6.39
6.12
6.01
<.001
<.001
.001
.003
.005
.016
.002
.004
.008
.010
5.63, p < .001, survived FWE correction at the cluster
level (cluster-forming threshold p < .001 uncorrected;
see Figure 4B for whole-brain results visualized at an
uncorrected threshold). Whole-brain conjunction analy-
ses further indicated significant overlap between subse-
quent success and subsequent precision effects in the
middle occipital and fusiform gyri (see Table 1 and
Figure 4C). Exclusive masking of each subsequent
memory contrast by the other did not reveal any further
regions where encoding activity significantly predicted
only the subsequent success or the subsequent precision
of memory retrieval.
DISCUSSION
Amid growing interest in the neural substrates underlying
the precision of episodic memory, prior studies have pre-
dominantly focused on retrieval processes (Cooper &
Ritchey, 2019; Montchal et al., 2019; Stevenson et al.,
2018; Richter et al., 2016), leaving the specific encoding
mechanisms supporting the acquisition of high-fidelity
memories largely uncharacterized. Here, we employed
continuous measures of memory retrieval in combination
with model-based analyses of fMRI data to segregate the
encoding activity supporting the later success and preci-
sion of episodic retrieval. We observed encoding activity
in overlapping cortical regions, including the IFG, FFG,
and middle occipital gyrus, to predict both which object
features were later successfully retrieved from memory
versus forgotten and the precision with which they were
reconstructed. In contrast, encoding activity in the HC
significantly predicted the precision of later memory
retrieval only. Together, these findings highlight a
hippocampal–cortical basis for the formation of precise
memories of perceptual information and provide novel
insight into the encoding substrates supporting the
accessibility and precision of episodic memory.
The current finding demonstrating a relationship
between trial-by-trial variation in hippocampal encoding
activity and later memory precision is consistent with
previous accounts emphasizing a critical role for this
region in supporting detailed episodic memories (Robin
& Moscovitch, 2017; Moscovitch et al., 2016). Related to
our current findings, prior neuroimaging studies have
found hippocampal encoding activity to correlate with
measures of later retrieval quality, such as participants’
subjective ratings of memory confidence (Kirwan,
Wixted, & Squire, 2008; Preston et al., 2008), or the objec-
tive amount of detail recalled (Cooper & Ritchey, 2020;
Qin et al., 2011). Moreover, trial-wise variation in hippo-
campal encoding activity has been found to predict the
specificity of subsequent neural reinstatement of mne-
monic content (Danker, Tompary, & Davachi, 2016;
Wing, Ritchey, & Cabeza, 2015), providing support for
the idea that hippocampal function at encoding may in
part determine the fidelity with which information can
2336
Journal of Cognitive Neuroscience
Volume 33, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
be later recalled. Emerging evidence suggests the posterior
HC to be particularly important for supporting the fine-
grained representation of perceptual details (Brunec et al.,
2018; Poppenk, Evensmoen, Moscovitch, & Nadel, 2013),
consistent with our current finding of the peak of the
subsequent precision effect being located in the posterior
part of the HC.
In contrast, we did not observe significant subsequent
retrieval success effects in the HC in our current para-
digm. Furthermore, hippocampal encoding activity still
predicted the precision of later memory retrieval after ex-
clusive masking with the subsequent retrieval success
contrast, suggesting specificity of this effect to memory
precision. The lack of significant retrieval success effects
in the HC may seem surprising given previous evidence
for hippocampal encoding increases for successful versus
unsuccessful encoding of associative information (Staresina
& Davachi, 2008; Davachi, 2006); however, we note that
prior studies have not attempted to distinguish memory
precision-related activity from that related to successful
encoding in general, both of which may be associated with
accurate performance in a categorical memory task. Similar
to the pattern of results observed here, others have further
observed hippocampal encoding activity to predict graded
variation in participants’ subjective ratings of memory con-
fidence only for responses above a certain threshold, while
not categorically distinguishing between remembered and
forgotten items (Shrager, Kirwan, & Squire, 2008).
Theoretical accounts postulate that hippocampal involve-
ment across cognitive domains may be explained by re-
quirement for representation of high-fidelity (i.e., highly
precise) and high-dimensional (i.e., comprising multiple as-
sociations) information (Ekstrom & Yonelinas, 2020;
Yonelinas, 2013). This account aligns with our current find-
ing of greater hippocampal encoding activity with greater
precision of object feature bindings, although we note that,
while requiring binding of multiple event attributes (i.e., ob-
ject identity to color and spatial location), event complexity
was not explicitly manipulated here and participants recon-
structed only one feature of each studied object while the
untested feature remained unchanged from study to test.
Future studies manipulating the number and type of object
attributes encoded, and testing memory for multiple fea-
tures, can more directly evaluate the relationship of hippo-
campal encoding activity to remembered event complexity.
Our findings are further in line with patient evidence dem-
onstrating medial temporal lesions to disproportionately
impair both short-term and long-term memory for high-
fidelity associations (Nilakantan et al., 2018; Koen,
Borders, Petzold, & Yonelinas, 2017) and suggest a poten-
tial role for a deficient hippocampal encoding mechanism
in such impairments.
A prior study employing a similar paradigm to the one
used here found hippocampal retrieval activity to be
associated with the success, but not precision, of episodic
memory retrieval (Richter et al., 2016). Although likely
not directly mapping onto prior distinctions made in the
literature, it is possible that this apparent difference
between encoding and retrieval effects in the HC could
reflect differential demands on hippocampal function
during memory encoding and retrieval. More specifically,
hippocampal pattern separation during memory encoding
may be critical for the storage of differentiated memory
representations that can be later retrieved with high
precision, in particular when feature overlap is high (i.e.,
when multiple objects encoded in similar colors or
spatial locations; Xie, Park, Zaghloul, & Zhang, 2020;
Moscovitch et al., 2016; Yassa & Stark, 2011; Norman &
O’Reilly, 2003; Bakker, Kirwan, Miller, & Stark, 2008). At
retrieval, hippocampal pattern completion is thought to
enable access to stored memory representations when
presented with a noisy or partial cue, resulting in a thresh-
olded memory signal where only items above a certain
criteria elicit successful retrieval (Norman, 2010; Norman
& O’Reilly, 2003). Interestingly, some evidence suggests
that hippocampal response during perception may be
more graded, supporting fine-grained perceptual discrim-
ination (Elfman, Aly, & Yonelinas, 2014; Aly et al., 2013), a
proposal consistent with the pattern of memory-related
activity observed here.
Beyond the HC, we observed activity in overlapping
cortical regions, including the IFG, FFG, and middle
occipital gyrus, to predict both the later success and pre-
cision of episodic memory retrieval. Our finding of left
inferior frontal involvement in subsequent retrieval suc-
cess and precision is consistent with previous evidence
implicating this region in cognitive control of memory
encoding, supporting successful memory formation
across a range of encoding tasks and mnemonic content
(Blumenfeld, Parks, Yonelinas, & Ranganath, 2011;
Park & Rugg, 2008, 2011; Murray & Ranganath, 2007;
Blumenfeld & Ranganath, 2006). Specifically, ventrolat-
eral regions of the prefrontal cortex have been proposed
to support the attentional selection and elaborative en-
coding of goal-relevant information, leading to formation
of strong and distinctive memory traces for specific item
features (Blumenfeld, Lee, & D’Esposito, 2014;
Blumenfeld & Ranganath, 2007; Simons & Spiers,
2003). Such selective encoding processes supported by
this region may act to enhance the representation of
goal-relevant features in posterior perceptual regions
(Sprague, Saproo, & Serences, 2015; Gilbert & Li, 2013;
Xue et al., 2013; Chun & Turk-Browne, 2007) and/or mod-
ulate hippocampal encoding more directly (Aly & Turk-
Browne, 2017; Carr, Engel, & Knowlton, 2013), aiding the
formation of durable and precise memory representations.
The current results further emphasize the role of per-
ceptual regions in supporting the formation of accessible
and precise memory traces. Specifically, we observed
encoding activity in the FFG, a region typically associated
with object perception and memory ( Vaidya, Zhao,
Desmond, & Gabrieli, 2002; Bar et al., 2001; Haxby et al.,
2001), to predict both the later success and precision of
object feature retrieval. This finding is consistent with
Korkki, Richter, and Simons
2337
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
previous studies that have observed memory-related
activity increases in the FFG during episodic encoding
(reviewed in Kim, 2011; Spaniol et al., 2009), potentially
playing an important role in the formation of detailed
object representations (Kensinger, Garoff-Eaton, &
Schacter, 2007; Garoff, Slotnick, & Schacter, 2005), and
with evidence suggesting representational specificity in
the occipitotemporal cortex during encoding to predict
subsequent memory performance (Gordon, Rissman,
Kiani, & Wagner, 2014; Ward, Chun, & Kuhl, 2013; Xue
et al., 2010). Beyond our ROIs, we further observed that
encoding activity in a wider network of ventral and
dorsal visual regions predicted the subsequent success of
memory retrieval. Of these regions, conjoint subsequent
retrieval success and precision effects were observed in
the middle occipital gyrus. The involvement of a broad
set of ventral and dorsal visual regions aligns with demands
of the current task for processing various visual attributes
of the study displays.
We further note that the interpretation of the mixture
model parameters as reflecting two distinct sources of
memory error in the context of long-term retrieval has
recently been challenged (Schurgin et al., 2020).
Specifically, Schurgin et al. (2020) suggest errors in visual
working memory, and at least under specific constraints
also in long-term memory, to be explained by a single-
parameter signal detection model when taking the non-
linear relationship between physical and psychological
stimulus spaces into account (Schurgin et al., 2020).
Although the ability of this model to account for selective
changes in retrieval success or precision observed in
previous studies of long-term memory (e.g., Nilakantan
et al., 2018; Cooper et al., 2017; Nilakantan, Bridge,
Gagnon, VanHaerents, & Voss, 2017; Sutterer & Awh,
2016), as well as to generalize to other stimulus spaces,
such as spatial locations employed here, remains unclear,
we note that our current findings regarding encoding
activity in the inferior frontal and ventral visual cortex
are not inconsistent with a single-parameter conceptual-
ization. It is possible that the common subsequent suc-
cess and subsequent precision effects observed in these
regions could reflect a single dimension of memory
strength or quality. Indeed, encoding activity in these
two regions was also found to predict trial-wise variation
in memory error when collapsing across all encoding
trials, although we note that no such effects were still
observed if examining trials classified as unsuccessful only.
However, we did not observe hippocampal encoding
activity to predict trial-wise variation in memory error
when collapsing across all encoding trials (or for trials
classified as unsuccessful), suggesting a benefit of the
mixture modeling approach for characterizing memory-
related activity in the HC. We further note that, although
our current approach of using model-derived retrieval
success thresholds estimated at the group level ensured
consistent classification of trials to conditions across
participants, this nevertheless means that our threshold
estimate was not sensitive to individual differences in
memory precision.
In summary, the current study aimed to elucidate the
encoding mechanisms supporting the formation of acces-
sible and precise memory traces. We observed encoding
activity in prefrontal and posterior perceptual regions to
support both the later success and precision of episodic
memory retrieval, suggesting a shared role in the forma-
tion of strong and durable memory traces that are readily
accessible from memory and can be reconstructed with a
high degree of precision. In contrast, activity in the HC was
found to significantly predict later memory precision only,
consistent with accounts emphasizing the importance of
this region in supporting high-fidelity representation of
associative information across cognitive domains.
Acknowledgments
We are grateful to Paul Bays for valuable advice and to the staff
of the MRC Cognition and Brain Sciences Unit MRI facility for
scanning assistance. For the purpose of open access, the author
has applied a CC BY public copyright license to any Author
Accepted Manuscript version arising from this submission.
Reprint requests should be sent to Saana M. Korkki or Jon S.
Simons, Department of Psychology, University of Cambridge,
Cambridge CB2 3EB, United Kingdom, or via e-mail: smk62@cam
.ac.uk, jss30@cam.ac.uk.
Author Contributions
Saana M. Korkki: Conceptualization; Formal analysis;
Investigation; Methodology; Writing—Original draft,
Writing—Review & editing. Franziska R. Richter:
Conceptualization; Formal analysis; Investigation;
Methodology; Supervision; Writing—Review & editing.
Jon S. Simons: Conceptualization; Formal analysis;
Funding acquisition; Methodology; Supervision; Writing
—Review & editing.
Funding Information
This study was funded by Biotechnology and Biological
Sciences Research Council (https://dx.doi.org/10.13039
/501100000268), grant number: BB/ L02263X/1, and
James S. McDonnell Foundation Scholar (https://dx.doi
.org/10.13039/100000913), grant number: #220020333,
and was carried out within the University of Cambridge
Behavioural and Clinical Neuroscience Institute, funded
by a joint award from the Medical Research Council
and the Wellcome Trust.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
2338
Journal of Cognitive Neuroscience
Volume 33, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/ W = .108, and W/ W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/ W = .102, and W/ W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explic-
itly when selecting which articles to cite and gives them
the opportunity to report their article’s gender citation
balance. The authors of this article report its proportions
of citations by gender category to be as follows: M/M =
.556, W/M = .286, M/ W = .063, and W/ W = .095.
REFERENCES
Aly, M., Ranganath, C., & Yonelinas, A. P. (2013). Detecting
changes in scenes: The hippocampus is critical for strength-
based perception. Neuron, 78, 1127–1137. https://doi.org/10
.1016/j.neuron.2013.04.018, PubMed: 23791201
Aly, M., & Turk-Browne, N. B. (2017). How hippocampal memory
shapes, and is shaped by, attention. In D.E. Hannula &
M.C. Duff (Eds.), The hippocampus from cells to systems:
Structure, connectivity, and functional contributions
to memory and flexible cognition. (pp. 369–403). Cham:
Springer. https://doi.org/10.1007/978-3-319-50406-3_12
Ashburner, J. (2007). A fast diffeomorphic image registration
algorithm. Neuroimage, 38, 95–113. https://doi.org/10.1016/j
.neuroimage.2007.07.007, PubMed: 17761438
Bakker, A., Kirwan, C. B., Miller, M., & Stark, C. E. L. (2008).
Pattern separation in the human hippocampal CA3 and
dentate gyrus. Science, 319, 1640–1642. https://doi.org/10
.1126/science.1152882, PubMed: 18356518
Bar, M., Tootell, R. B. H., Schacter, D. L., Greve, D. N., Fischl, B.,
Mendola, J. D., et al. (2001). Cortical mechanisms specific
to explicit visual object recognition. Neuron, 29, 529–535.
https://doi.org/10.1016/S0896-6273(01)00224-0, PubMed:
11239441
Bays, P. M., Catalao, R. F. G., & Husain, M. (2009). The precision
of visual working memory is set by allocation of a shared
resource. Journal of Vision, 9, 1–11. https://doi.org/10.1167/9
.10.7, PubMed: 19810788
Berens, S. C., Richards, B. A., & Horner, A. J. (2020).
Dissociating memory accessibility and precision in forgetting.
Nature Human Behaviour, 4, 866–877. https://doi.org/10
.1038/s41562-020-0888-8, PubMed: 32514041
Blumenfeld, R. S., Lee, T. G., & D’Esposito, M. (2014). The
effects of lateral prefrontal transcranial magnetic stimulation
on item memory encoding. Neuropsychologia, 53, 197–202.
https://doi.org/10.1016/j.neuropsychologia.2013.11.021,
PubMed: 24316198
Blumenfeld, R. S., Parks, C. M., Yonelinas, A. P., & Ranganath, C.
(2011). Putting the pieces together: The role of dorsolateral
prefrontal cortex in relational memory encoding. Journal of
Cognitive Neuroscience, 23, 257–265. https://doi.org/10.1162
/jocn.2010.21459, PubMed: 20146616
Blumenfeld, R. S., & Ranganath, C. (2006). Dorsolateral prefrontal
cortex promotes long-term memory formation through its
role in working memory organization. Journal of Neuroscience,
26, 916–925. https://doi.org/10.1523/JNEUROSCI.2353-05.2006,
PubMed: 16421311
Blumenfeld, R. S., & Ranganath, C. (2007). Prefrontal cortex and
long-term memory encoding: An integrative review of findings
from neuropsychology and neuroimaging. Neuroscientist,
13, 280–291. https://doi.org/10.1177/1073858407299290,
PubMed: 17519370
Brady, T. F., Konkle, T., Gill, J., Oliva, A., & Alvarez, G. A. (2013).
Visual long-term memory has the same limit on fidelity as
visual working memory. Psychological Science, 24, 981–990.
https://doi.org/10.1177/0956797612465439, PubMed:
23630219
Brunec, I. K., Bellana, B., Ozubko, J. D., Man, V., Robin, J., Liu,
Z.-X., et al. (2018). Multiple scales of representation along
the hippocampal anteroposterior axis in humans. Current
Biology, 28, 2129.e6–2135.e6. https://doi.org/10.1016/j.cub
.2018.05.016, PubMed: 29937352
Carr, V. A., Engel, S. A., & Knowlton, B. J. (2013). Top–down
modulation of hippocampal encoding activity as measured
by high-resolution functional MRI. Neuropsychologia, 51,
1829–1837. https://doi.org/10.1016/j.neuropsychologia.2013
.06.026, PubMed: 23838003
Chun, M. M., & Turk-Browne, N. B. (2007). Interactions
between attention and memory. Current Opinion in
Neurobiology, 17, 177–184. https://doi.org/10.1016/j.conb
.2007.03.005, PubMed: 17379501
Cooper, R. A., Richter, F. R., Bays, P. M., Plaisted-Grant, K. C.,
Baron-Cohen, S., & Simons, J. S. (2017). Reduced
hippocampal functional connectivity during episodic
memory retrieval in autism. Cerebral Cortex, 27, 888–902.
https://doi.org/10.1093/cercor/bhw417, PubMed: 28057726
Cooper, R. A., & Ritchey, M. (2019). Cortico-hippocampal
network connections support the multidimensional quality of
episodic memory. eLife, 8, e45591. https://doi.org/10.7554
/eLife.45591, PubMed: 30900990
Cooper, R. A., & Ritchey, M. (2020). Progression from feature-
specific brain activity to hippocampal binding during episodic
encoding. Journal of Neuroscience, 40, 1701–1709. https://
doi.org/10.1523/JNEUROSCI.1971-19.2019, PubMed:
31826947
Danker, J. F., Tompary, A., & Davachi, L. (2016). Trial-by-trial
hippocampal encoding activation predicts the fidelity of
cortical reinstatement during subsequent retrieval. Cerebral
Cortex, 27, 3515–3524. https://doi.org/10.1093/cercor/bhw146,
PubMed: 27288317
Davachi, L. (2006). Item, context and relational episodic
encoding in humans. Current Opinion in Neurobiology,
16, 693–700. https://doi.org/10.1016/j.conb.2006.10.012,
PubMed: 17097284
Ekstrom, A. D., & Yonelinas, A. P. (2020). Precision, binding,
and the hippocampus: Precisely what are we talking about?
Neuropsychologia, 138, 107341. https://doi.org/10.1016/j
.neuropsychologia.2020.107341, PubMed: 31945386
Elfman, K. W., Aly, M., & Yonelinas, A. P. (2014).
Neurocomputational account of memory and perception:
Thresholded and graded signals in the hippocampus.
Hippocampus, 24, 1672–1686. https://doi.org/10.1002/hipo
.22345, PubMed: 25112784
Emrich, S. M., Riggall, A. C., LaRocque, J. J., & Postle, B. R.
(2013). Distributed patterns of activity in sensory cortex
reflect the precision of multiple items maintained in visual
short-term memory. Journal of Neuroscience, 33,
6516–6523. https://doi.org/10.1523/JNEUROSCI.5732-12
.2013, PubMed: 23575849
Garoff, R. J., Slotnick, S. D., & Schacter, D. L. (2005). The neural
origins of specific and general memory: The role of the
fusiform cortex. Neuropsychologia, 43, 847–859. https://doi
.org/10.1016/j.neuropsychologia.2004.09.014, PubMed:
15716157
Gilbert, C. D., & Li, W. (2013). Top–down influences on visual
processing. Nature Reviews Neuroscience, 14, 350–363.
https://doi.org/10.1038/nrn3476, PubMed: 23595013
Gordon, A. M., Rissman, J., Kiani, R., & Wagner, A. D. (2014).
Cortical reinstatement mediates the relationship between
content-specific encoding activity and subsequent
Korkki, Richter, and Simons
2339
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
recollection decisions. Cerebral Cortex, 24, 3350–3364.
https://doi.org/10.1093/cercor/bht194, PubMed: 23921785
Harlow, I. M., & Donaldson, D. I. (2013). Source accuracy data
reveal the thresholded nature of human episodic memory.
Psychonomic Bulletin & Review, 20, 318–325. https://doi.org
/10.3758/s13423-012-0340-9, PubMed: 23192370
Harlow, I. M., & Yonelinas, A. P. (2016). Distinguishing between
the success and precision of recollection. Memory, 24,
114–127. https://doi.org/10.1080/09658211.2014.988162,
PubMed: 25494616
Haxby, J. V., Gobbini, M. I., Furey, M. L., Ishai, A., Schouten, J. L., &
Pietrini, P. (2001). Distributed and overlapping representations
of faces and objects in ventral temporal cortex. Science, 293,
2425–2430. https://doi.org/10.1126/science.1063736, PubMed:
11577229
Kensinger, E. A., Addis, D. R., & Atapattu, R. K. (2011).
Amygdala activity at encoding corresponds with memory
vividness and with memory for select episodic details.
Neuropsychologia, 49, 663–673. https://doi.org/10.1016/j
.neuropsychologia.2011.01.017, PubMed: 21262244
Kensinger, E. A., Garoff-Eaton, R. J., & Schacter, D. L. (2007).
How negative emotion enhances the visual specificity of a
memory. Journal of Cognitive Neuroscience, 19, 1872–1887.
https://doi.org/10.1162/jocn.2007.19.11.1872, PubMed:
17958489
Kim, H. (2011). Neural activity that predicts subsequent
memory and forgetting: A meta-analysis of 74 fMRI studies.
Neuroimage, 54, 2446–2461. https://doi.org/10.1016/j
.neuroimage.2010.09.045, PubMed: 20869446
Kirwan, C. B., Wixted, J. T., & Squire, L. R. (2008). Activity in the
medial temporal lobe predicts memory strength, whereas
activity in the prefrontal cortex predicts recollection. Journal
of Neuroscience, 28, 10541–10548. https://doi.org/10.1523
/JNEUROSCI.3456-08.2008
Koen, J. D., Borders, A. A., Petzold, M. T., & Yonelinas, A. P.
(2017). Visual short-term memory for high resolution
associations is impaired in patients with medial temporal
lobe damage. Hippocampus, 27, 184–193. https://doi.org/10
.1002/hipo.22682, PubMed: 27859914
Kolarik, B. S., Shahlaie, K., Hassan, A., Borders, A. A., Kaufman,
K. C., Gurkoff, G., et al. (2016). Impairments in precision,
rather than spatial strategy, characterize performance on the
virtual Morris Water Maze: A case study. Neuropsychologia,
80, 90–101. https://doi.org/10.1016/j.neuropsychologia.2015
.11.013, PubMed: 26593960
Korkki, S. M., Richter, F. R., Jeyarathnarajah, P., & Simons, J. S.
(2020). Healthy ageing reduces the precision of episodic
memory retrieval. Psychology and Aging, 35, 124–142.
https://doi.org/10.1037/pag0000432, PubMed: 31928030
Montchal, M. E., Reagh, Z. M., & Yassa, M. A. (2019). Precise
temporal memories are supported by the lateral
entorhinal cortex in humans. Nature Neuroscience, 22,
284–288. https://doi.org/10.1038/s41593-018-0303-1, PubMed:
30643291
Moscovitch, M., Cabeza, R., Winocur, G., & Nadel, L. (2016).
Episodic memory and beyond: The hippocampus and
neocortex in transformation. Annual Review of Psychology,
67, 105–134. https://doi.org/10.1146/annurev-psych-113011
-143733, PubMed: 26726963
Murray, L. J., & Ranganath, C. (2007). The dorsolateral
prefrontal cortex contributes to successful relational memory
encoding. Journal of Neuroscience, 27, 5515–5522. https://
doi.org/10.1523/JNEUROSCI.0406-07.2007, PubMed:
17507573
Nichols, T., Brett, M., Andersson, J., Wager, T., & Poline, J.-B.
(2005). Valid conjunction inference with the minimum
statistic. Neuroimage, 25, 653–660. https://doi.org/10.1016/j
.neuroimage.2004.12.005, PubMed: 15808966
Nilakantan, A. S., Bridge, D. J., Gagnon, E. P., VanHaerents, S. A.,
& Voss, J. L. (2017). Stimulation of the posterior cortical–
hippocampal network enhances precision of memory
recollection. Current Biology, 27, 465–470. https://doi.org/10
.1016/j.cub.2016.12.042, PubMed: 28111154
Nilakantan, A. S., Bridge, D. J., VanHaerents, S., & Voss, J. L.
(2018). Distinguishing the precision of spatial recollection
from its success: Evidence from healthy aging and unilateral
mesial temporal lobe resection. Neuropsychologia, 119,
101–106. https://doi.org/10.1016/j.neuropsychologia.2018.07
.035, PubMed: 30086364
Norman, K. A. (2010). How hippocampus and cortex contribute
to recognition memory: Revisiting the complementary
learning systems model. Hippocampus, 20, 1217–1227.
https://doi.org/10.1002/hipo.20855, PubMed: 20857486
Norman, K. A., & O’Reilly, R. C. (2003). Modeling hippocampal
and neocortical contributions to recognition memory: A
complementary-learning-systems approach. Psychological
Review, 110, 611–646. https://doi.org/10.1037/0033-295X
.110.4.611, PubMed: 14599236
O’Reilly, R. C., & McClelland, J. L. (1994). Hippocampal
conjunctive encoding, storage, and recall: Avoiding a trade-
off. Hippocampus, 4, 661–682. https://doi.org/10.1002/hipo
.450040605, PubMed: 7704110
Otten, L. J., & Rugg, M. D. (2001). Task-dependency of the
neural correlates of episodic encoding as measured by fMRI.
Cerebral Cortex, 11, 1150–1160. https://doi.org/10.1093
/cercor/11.12.1150
Paller, K. A., & Wagner, A. D. (2002). Observing the transformation
of experience into memory. Trends in Cognitive Sciences,
6, 93–102. https://doi.org/10.1016/S1364-6613(00)01845-3,
PubMed: 15866193
Park, H., & Rugg, M. D. (2008). Neural correlates of successful
encoding of semantically and phonologically mediated inter-
item associations. Neuroimage, 43, 165–172. https://doi.org
/10.1016/j.neuroimage.2008.06.044, PubMed: 18675362
Park, H., & Rugg, M. D. (2011). Neural correlates of encoding
within- and across-domain inter-item associations. Journal of
Cognitive Neuroscience, 23, 2533–2543. https://doi.org/10
.1162/jocn.2011.21611, PubMed: 21254802
Park, H., Uncapher, M. R., & Rugg, M. D. (2008). Effects of study
task on the neural correlates of source encoding. Learning &
Memory, 15, 417–425. https://doi.org/10.1101/lm.878908
Poppenk, J., Evensmoen, H. R., Moscovitch, M., & Nadel, L.
(2013). Long-axis specialization of the human hippocampus.
Trends in Cognitive Sciences, 17, 230–240. https://doi.org/10
.1016/j.tics.2013.03.005, PubMed: 23597720
Preston, A. R., Bornstein, A. M., Hutchinson, J. B., Gaare, M. E.,
Glover, G. H., & Wagner, A. D. (2010). High-resolution fMRI
of content-sensitive subsequent memory responses in human
medial temporal lobe. Journal of Cognitive Neuroscience,
22, 156–173. https://doi.org/10.1162/jocn.2009.21195
Qin, S., van Marle, H. J. F., Hermans, E. J., & Fernandez, G.
(2011). Subjective sense of memory strength and the
objective amount of information accurately remembered are
related to distinct neural correlates at encoding. Journal of
Neuroscience, 31, 8920–8927. https://doi.org/10.1523
/JNEUROSCI.2587-10.2011, PubMed: 21677175
Ranganath, C. (2010). A unified framework for the functional
organization of the medial temporal lobes and the
phenomenology of episodic memory. Hippocampus, 20,
1263–1290. https://doi.org/10.1002/hipo.20852, PubMed:
20928833
Ranganath, C., Yonelinas, A. P., Cohen, M. X., Dy, C. J., Tom, S. M.,
& D’Esposito, M. (2004). Dissociable correlates of
recollection and familiarity within the medial temporal lobes.
Neuropsychologia, 42, 2–13. https://doi.org/10.1016/j
.neuropsychologia.2003.07.006, PubMed: 14615072
2340
Journal of Cognitive Neuroscience
Volume 33, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Richter, F. R., Cooper, R. A., Bays, P. M., & Simons, J. S. (2016).
Distinct neural mechanisms underlie the success, precision,
and vividness of episodic memory. eLife, 5, e18260. https://
doi.org/10.7554/eLife.18260, PubMed: 27776631
Robin, J., & Moscovitch, M. (2017). Details, gist and schema:
Hippocampal–neocortical interactions underlying recent and
remote episodic and spatial memory. Current Opinion in
Behavioral Sciences, 17, 114–123. https://doi.org/10.1016/j
.cobeha.2017.07.016
Schurgin, M. W., Wixted, J. T., & Brady, T. F. (2020).
Psychophysical scaling reveals a unified theory of visual
memory strength. Nature Human Behaviour, 4, 1156–1172.
https://doi.org/10.1038/s41562-020-00938-0, PubMed:
32895546
Shrager, Y., Kirwan, C. B., & Squire, L. R. (2008). Activity in both
hippocampus and perirhinal cortex predicts the memory
strength of subsequently remembered information. Neuron,
59, 547–553. https://doi.org/10.1016/j.neuron.2008.07.022,
PubMed: 18760691
Simons, J. S., & Spiers, H. J. (2003). Prefrontal and medial
temporal lobe interactions in long-term memory. Nature
Reviews Neuroscience, 4, 637–648. https://doi.org/10.1038
/nrn1178, PubMed: 12894239
Smith, A. P. R., Henson, R. N. A., Dolan, R. J., & Rugg, M. D.
(2004). fMRI correlates of the episodic retrieval of emotional
contexts. Neuroimage, 22, 868–878. https://doi.org/10.1016/j
.neuroimage.2004.01.049, PubMed: 15193617
Spaniol, J., Davidson, P. S. R., Kim, A. S. N., Han, H.,
Moscovitch, M., & Grady, C. L. (2009). Event-related fMRI
studies of episodic encoding and retrieval: Meta-analyses
using activation likelihood estimation. Neuropsychologia, 47,
1765–1779. https://doi.org/10.1016/j.neuropsychologia.2009
.02.028, PubMed: 19428409
Sprague, T. C., Saproo, S., & Serences, J. T. (2015). Visual
attention mitigates information loss in small- and large-scale
neural codes. Trends in Cognitive Sciences, 19, 215–226.
https://doi.org/10.1016/j.tics.2015.02.005, PubMed: 25769502
Staresina, B. P., & Davachi, L. (2006). Differential encoding
mechanisms for subsequent associative recognition and free
recall. Journal of Neuroscience, 26, 9162–9172. https://doi
.org/10.1523/JNEUROSCI.2877-06.2006, PubMed: 16957073
Staresina, B. P., & Davachi, L. (2008). Selective and shared
contributions of the hippocampus and perirhinal cortex to
episodic item and associative encoding. Journal of Cognitive
Neuroscience, 20, 1478–1489. https://doi.org/10.1162/jocn
.2008.20104, PubMed: 18303974
Stevenson, R. F., Zheng, J., Mnatsakanyan, L., Vadera, S., Knight,
R. T., Lin, J. J., et al. (2018). Hippocampal CA1 gamma power
predicts the precision of spatial memory judgments.
Proceedings of the National Academy of Sciences, U.S.A.,
115, 10148–10153. https://doi.org/10.1073/pnas.1805724115,
PubMed: 30224452
Sun, S. Z., Fidalgo, C., Barense, M. D., Lee, A. C. H., Cant, J. S., &
Ferber, S. (2017). Erasing and blurring memories: The
differential impact of interference on separate aspects of
forgetting. Journal of Experimental Psychology: General,
146, 1606–1630. https://doi.org/10.1037/xge0000359,
PubMed: 28933892
Sutterer, D. W., & Awh, E. (2016). Retrieval practice enhances
the accessibility but not the quality of memory. Psychonomic
Bulletin & Review, 23, 831–841. https://doi.org/10.3758
/s13423-015-0937-x, PubMed: 26404635
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neuroimage, 15, 273–289. https://doi.org/10.1006/nimg
.2001.0978
Uncapher, M. R., Otten, L. J., & Rugg, M. D. (2006). Episodic
encoding is more than the sum of its parts: An fMRI
investigation of multifeatural contextual encoding. Neuron,
52, 547–556. https://doi.org/10.1016/j.neuron.2006.08.011,
PubMed: 17088219
Vaidya, C. J., Zhao, M., Desmond, J. E., & Gabrieli, J. D. E.
(2002). Evidence for cortical encoding specificity in episodic
memory: Memory-induced re-activation of picture processing
areas. Neuropsychologia, 40, 2136–2143. https://doi.org/10
.1016/S0028-3932(02)00053-2, PubMed: 12208009
Ward, E. J., Chun, M. M., & Kuhl, B. A. (2013). Repetition
suppression and multi-voxel pattern similarity differentially
track implicit and explicit visual memory. Journal of
Neuroscience, 33, 14749–14757. https://doi.org/10.1523
/JNEUROSCI.4889-12.2013, PubMed: 24027275
Wing, E. A., Ritchey, M., & Cabeza, R. (2015). Reinstatement of
individual past events revealed by the similarity of distributed
activation patterns during encoding and retrieval. Journal of
Cognitive Neuroscience, 27, 679–691. https://doi.org/10.1162
/jocn_a_00740, PubMed: 25313659
Xie, W., Park, H.-B., Zaghloul, K. A., & Zhang, W. (2020).
Correlated individual differences in the estimated precision
of working memory and long-term memory: Commentary
on the study by Biderman, Luria, Teodorescu, Hajaj, and
Goshen-Gottstein (2019). Psychological Science, 31, 345–348.
https://doi.org/10.1177/0956797620903718, PubMed: 32049590
Xie, W., & Zhang, W. (2017). Negative emotion enhances
mnemonic precision and subjective feelings of remembering
in visual long-term memory. Cognition, 166, 73–83. https://
doi.org/10.1016/j.cognition.2017.05.025, PubMed: 28554087
Xue, G., Dong, Q., Chen, C., Lu, Z., Mumford, J. A., & Poldrack,
R. A. (2010). Greater neural pattern similarity across
repetitions is associated with better memory. Science, 330,
97–101. https://doi.org/10.1126/science.1193125, PubMed:
20829453
Xue, G., Dong, Q., Chen, C., Lu, Z.-L., Mumford, J. A., &
Poldrack, R. A. (2013). Complementary role of frontoparietal
activity and cortical pattern similarity in successful episodic
memory encoding. Cerebral Cortex, 23, 1562–1571. https://
doi.org/10.1093/cercor/bhs143, PubMed: 22645250
Yassa, M. A., & Stark, C. E. L. (2011). Pattern separation in the
hippocampus. Trends in Neurosciences, 34, 515–525. https://
doi.org/10.1016/j.tins.2011.06.006, PubMed: 21788086
Yonelinas, A. P. (2013). The hippocampus supports high-
resolution binding in the service of perception, working
memory and long-term memory. Behavioural Brain
Research, 254, 34–44. https://doi.org/10.1016/j.bbr.2013.05
.030, PubMed: 23721964
Zhang, W., & Luck, S. J. (2008). Discrete fixed-resolution
representations in visual working memory. Nature, 453,
233–235. https://doi.org/10.1038/nature06860, PubMed:
18385672
Korkki, Richter, and Simons
2341
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
1
2
3
2
8
1
9
6
5
7
0
5
/
/
j
o
c
n
_
a
_
0
1
7
7
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3