Greater Visual Working Memory Capacity for
Visually Matched Stimuli When They Are
Perceived as Meaningful
Isabel E. Asp1
, Viola S. Störmer1,2, and Timothy F. Brady1
Abstrakt
■ Almost all models of visual working memory—the cognitive sys-
tem that holds visual information in an active state—assume it has a
fixed capacity: Some models propose a limit of three to four objects,
where others propose there is a fixed pool of resources for each
basic visual feature. Recent findings, Jedoch, suggest that memory
performance is improved for real-world objects. What supports
these increases in capacity? Hier, we test whether the meaningful-
ness of a stimulus alone influences working memory capacity while
controlling for visual complexity and directly assessing the active
component of working memory using EEG. Participants remem-
bered ambiguous stimuli that could either be perceived as a face
or as meaningless shapes. Participants had higher performance
and increased neural delay activity when the memory display con-
sisted of more meaningful stimuli. Critically, by asking participants
whether they perceived the stimuli as a face or not, we also show
that these increases in visual working memory capacity and recruit-
ment of additional neural resources are because of the subjective
perception of the stimulus and thus cannot be driven by physical
properties of the stimulus. Broadly, this suggests that the capacity
for active storage in visual working memory is not fixed but that
more meaningful stimuli recruit additional working memory
resources, allowing them to be better remembered. ■
EINFÜHRUNG
Visual working memory is a cognitive system used to hold
visual information in an active state, protecting it from in-
terference from new sensory inputs. The capacity of visual
working memory is clearly highly limited. Jedoch, nearly
all theories of visual working memory go beyond arguing
capacity is limited and argue that, for a given set of stimuli,
there is a “fixed” capacity, Zum Beispiel, a certain fixed
number of objects that can be remembered regardless of
complexity (z.B., of three to four objects; Awh, Barton, &
Vogel, 2007), or a fixed resource pool that can be distrib-
uted among items (Bays, Catalao, & Husain, 2009). Das
fixed capacity is often claimed to arise from limits in atten-
tional capacity like a certain number of attentional spot-
lights for maintaining items in the focus of attention
(z.B., Cowan, 2005) or from more neurally inspired re-
source limits like a probabilistically fixed number of action
potentials because of divisive normalization (z.B., Bays,
2015; Ma, Husain, & Bays, 2014). Daher, although adding
new visual features, like colors in addition to orientations,
may allow for parallel or object-based storage of these
features in distinct neural populations and thus “more
capacity” (z.B., Fougnie, Cormiea, & Alvarez, 2013), for a
given set of the same stimuli—with the same objects,
colors, and orientations present—capacity is argued to
be fixed by many working memory models. Ähnlich, für
1Universität von Kalifornien, 2Dartmouth College
© 2021 Massachusetts Institute of Technology
sufficiently complex stimuli that already contain all of the
low-level features that can be processed in parallel (orien-
Station, color, usw.; Wolfe & Horowitz, 2017), many fixed-
capacity models make the prediction that there is no
longer an additional benefit from parallel storage.
Stattdessen, when each item to be remembered is more com-
plex, each item simply requires more memory resources
(z.B., Luria, Sessa, Gotler, Jolicœur, & Dell’Acqua, 2010),
resulting in worse performance at the same set size. Für
Beispiel, some theories say that when items become
complex enough, each item requires more than one “slot”
in memory (Luck, 2008).
How do these working memory limits affect memory for
more realistic stimuli, like real objects, that are not just
conjunctions of simple features? Einerseits, if visual
working memory is best described as a set of parallel
mechanisms for storing simple, low-level features and sus-
taining them, then realistic objects, just like very complex
polygons, should simply take more of the same fixed pool
of working memory resources compared to simple stimuli
(as in Luria et al., 2010), because real objects tend to be
more visually complex than simple features alone. An
die andere Hand, visual working memory may not be
thought of solely as reflecting the maintenance of a set
of low-level visual features: Stattdessen, perception of stimuli
as meaningful may allow the recruitment of additional
features and neural populations, which can be used to
support working memory storage. Zum Beispiel, perceiving
a stimulus as a face may allow the same stimulus to be
Zeitschrift für kognitive Neurowissenschaften 33:5, S. 902–918
https://doi.org/10.1162/jocn_a_01693
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
represented in terms of not just low-level but also face-
specific visual features (pose, Geschlecht, usw.), und von
higher-level face regions in addition to low-level visual re-
gions, effectively creating additional working memory ca-
pacity. This would suggest that meaningful perception of
stimuli changes the nature of visual working memory ca-
pacity limits, allowing the recruitment of additional
resources.
Consistent with the idea that meaningful stimuli may be
better represented in visual working memory, some behav-
ioral work has shown that familiarity and knowledge
improve performance in short-term memory tasks (z.B.,
Zimmer & Fischer, 2020; Ngiam, Khaw, Holcombe, &
Goodbourn, 2018; Brady, Konkle, & Alvarez, 2009; Curby,
Glazek, & Gauthier, 2009; Jackson & Raymond, 2008;
Alvarez & Cavanagh, 2004). Zum Beispiel, familiar faces
appear to be easier to remember than unfamiliar faces
( Jackson & Raymond, 2008), and familiar letters, eher
than letters from unfamiliar alphabets, are more easily
remembered (Ngiam et al., 2018). Such benefits can even
result from moderate amounts of training (z.B., on Chinese
Figuren: Zimmer & Fischer, 2020; Zimmer, Popp, Reith,
& Krick, 2012). Such results—related to idea of “chunk-
ing”—are broadly consistent with the idea that working
memory capacity is impacted by knowledge. Jedoch, Es
is unclear what is driving these effects on improved perfor-
Mance. Zum Beispiel, in addition to the core storage capac-
ity of items actively in the focus of attention, it has been
repeatedly found that activated forms of long-term memory
that can be used for short-term storage of information
(z.B., Cowan, 2005) take advantage of knowledge (z.B.,
long-term working memory; Ericsson & Kintsch, 1995).
Daher, it is plausible that these effects of knowledge do
not result from any change in “core” working memory ca-
pacity, but the recruitment of other, more passive forms of
storage. Zum Beispiel, “chunking” has been argued to re-
flect some form of passive storage rather than a change
in the active capacity of working memory per se (Huang
& Awh, 2018), and there is some evidence for this in certain
situations, where significant training with a small set of
stimuli seems to allow them to be encoded in more ab-
stract and more efficient ways (z.B., Zimmer & Fischer,
2020). Zusätzlich, some authors have claimed that differ-
ences in capacity for different stimuli result from confusa-
bility of the item pairs in the test (z.B., a decision effect),
rather than a true effect on how much information can be
actively maintained in working memory (Awh et al., 2007).
Some recent work has found that when people are
asked to remember real-world objects (z.B., an umbrella,
ein Stuhl) instead of simple isolated features (z.B., colored
squares), not only is behavioral working memory capacity
erhöht (Brady & Störmer, 2020A, 2020B; Brady,
Störmer, & Alvarez, 2016), but so is a neural measure of
working memory engagement (Brady et al., 2016: the con-
tralateral delay activity [CDA]; Vogel & Machizawa, 2004).
Das ist, people perform better and show more neural de-
lay activity with more realistic objects, consistent with the
recruitment of additional memory resources for the more
realistic stimuli, rather than these items taking either more
or less of the same fixed capacity or relying on alternative
memory systems. Jedoch, real-world objects differ in
many ways from simple isolated features: Zum Beispiel,
they are both visually more complex and also uniquely
connect to existing knowledge (d.h., are meaningful).
Daher, it remains unknown what the critical factor is that
allows the recruitment of additional memory resources.
Außerdem, the measured neural activity during the
working memory delay period (the CDA) may be sensitive
to the visual complexity of stimuli independently of
memory capacity per se, making it difficult to interpret
changes in neural activity as being solely because of
changes in working memory engagement.
Hier, we test whether this neurally active component of
working memory storage, measured using the CDA, Ist
affected by the meaningfulness of a stimulus while control-
ling for visual complexity, das ist, whether people genu-
inely recruit additional working memory resources for
novel stimuli that can be processed in a meaningful way.
We asked participants to remember two-tone images that
were either perceived as meaningful or not for short pe-
riods, and measured memory performance (Experimente
1 Und 2) as well as sustained neural activity during the
maintenance period using electroencephalography as a di-
rect measure of the active component of working memory
(EEG; Experiment 3). Stimuli consisted of ambiguous face
Bilder (“Mooney images”; Mooney, 1957) that were care-
fully matched so that different images would be perceived
as a face by different people (Brady, Alvarez, & Störmer,
2019; Schwiedrzik, Melloni, & Schurger, 2018); das ist,
the same Mooney face would be perceived as meaningful
by one person but as not meaningful by someone else.
Daher, among these face stimuli, there were, on average,
no physical differences between images that one participant
would perceive as a face or perceive as a set of black-and-
white arbitrary shapes (not meaningful). In addition to
these ambiguous face stimuli, we also showed participants
arbitrary shapes that matched the overall low-level features
of the faces (Experimente 1 Und 3) or inverted versions of
the faces (Experiment 2), which provided an objective non-
face condition. The set of experiments allowed us to exam-
ine both the cognitive and neural consequences of visual
information being perceived as meaningful or not and
thus to examine whether active storage in working memory
per se was changed by this perception, independent of the
physical attributes of the stimulus.
EXPERIMENT 1: BEHAVIORAL CAPACITY
FOR MOONEY FACES VERSUS
UNRECOGNIZABLE SHAPES
Experiment 1 examined behavioral working memory
capacity for Mooney faces perceived as faces versus not
perceived as faces and for scrambled Mooney faces.
Asp, Störmer, and Brady
903
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Methoden
Teilnehmer
All participants gave written informed consent before
beginning the experiment. Experiment 1 was preregis-
tered (aspredicted.org/ blind.php?x=k8rm4k), Und, In
accordance with the preregistration, 24 undergraduates
from the University of California, San Diego, participated
for course credit (16 Frauen, ages 18–28 years). All exper-
iments were approved by the institutional review board
of the University of California, San Diego.
Stimuli
The stimuli used consisted of 300 Bilder (150 ambiguous
Mooney faces and 150 shuffled Mooney faces that appeared
as unrecognizable shapes). The ambiguous face images
were previously ranked as moderately difficult to recognize
a face, gaining the status “ambiguous” instead of “easy” (für
Einzelheiten, see Brady et al., 2019; Schwiedrzik et al., 2018,
which used these stimuli in a long-term memory context).
This difficulty was chosen in an attempt to create a more
evenly distributed subjective report of faces seen. The un-
recognizable shape stimuli were created by segmenting the
Mooney face images into continuous regions of white or
black, labeling these as “parts,” inverting each region, Und
then creating new images from this library of parts, Thema
to the constraint that no two parts of a newly generated
image overlapped (Figure 1B). The target/foil images for
the two-alternative forced-choice (2-AFC) test were ran-
domly selected per trial; Jedoch, the foil image always
matched the condition of the target (d.h., target: unrecog-
nizable shapes; foil: unrecognizable shapes). To ensure that
participants saw each stimulus an equal number of times
and so that we could analyze item effects, jeder Reiz
appeared 1 time as a target, 1 time as a foil, and twice as a
distractor, giving a total of 300 trials per participant.
Verfahren
During the experiment, participants viewed three images
centered around fixation for 1000 ms (Figure 1A). Wir
used long encoding times because of previous evidence
that the role of meaningfulness may require long encoding
times to become apparent (Brady & Störmer, 2020A; Brady
et al., 2016). The memory display of each trial would con-
sist of either zero, eins, zwei, or three ambiguous Mooney
faces with unrecognizable shape stimuli making up any re-
mainder so that the memory set would always consist of
three images. Als solche, we varied how many of these im-
ages were ambiguous face stimuli versus unrecognizable
shape stimuli throughout the experiment. While the im-
ages were on the screen, participants were to maintain
their eyes on the center of the screen. After the images dis-
appeared, there was an 800-msec delay period with only
image location markers on the screen. Following the de-
lay, one of the location markers enlarged for 500 ms, In-
dicating the location of the to-be-tested image. The target
image and a foil image of the same condition (d.h., face or
unrecognizable shapes) were then presented one above
the other, with the space between the two images cen-
tered at the prior location marker. Participants were asked
to indicate which of the two images was part of the recent
set of studied images by pressing a key on the keyboard
(top image: upper arrow key; bottom image: lower arrow
key). The two images remained on the screen until the
participant made a selection. Once the participant pressed
a key, they were prompted to recall and respond with how
many images they had perceived as faces during encoding
(0, 1, 2, oder 3) using the number pad. The experiment con-
sisted of five blocks with 60 trials in each. The trial condi-
tions were randomized throughout the experiment, Und
each trial condition was presented evenly within each
block. Prior to the experiment, all participants completed
two mini-blocks of four trials each as practice for the full
five blocks of the experiment that followed. None of the
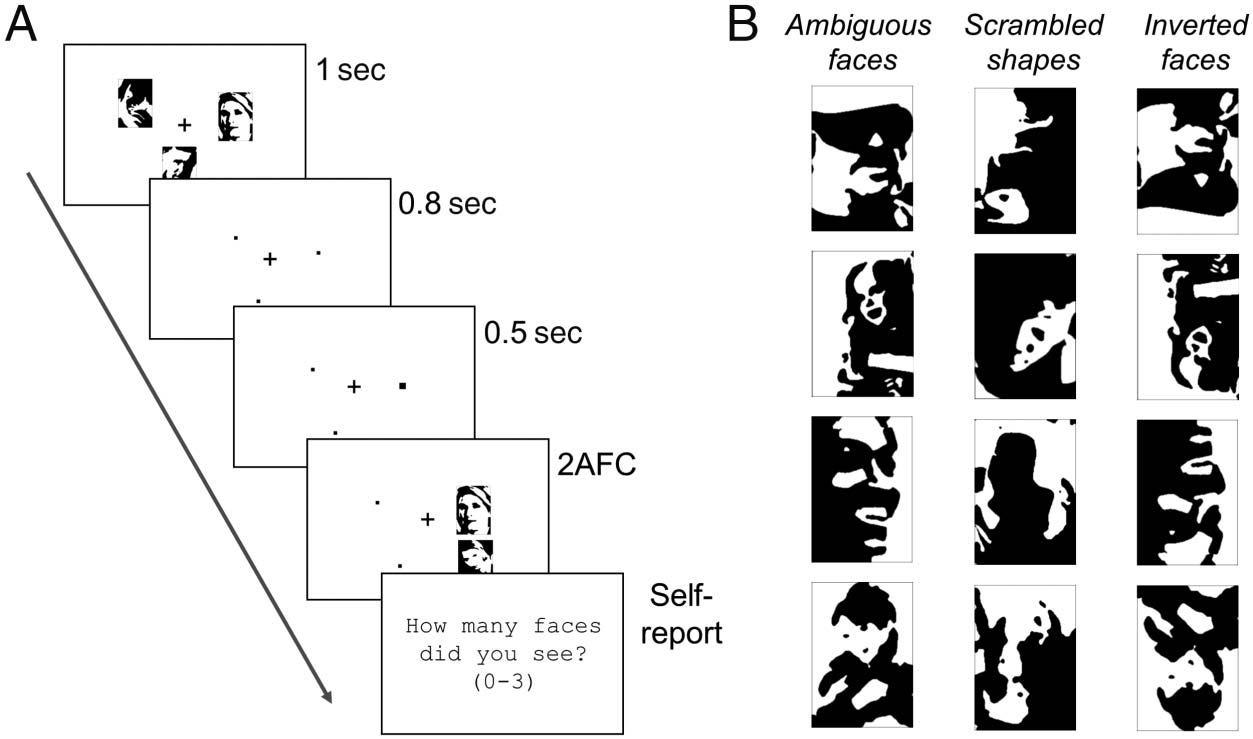
Figur 1. (A) Methods for
Experimente 1 Und 2.
Participants saw a memory
display with three images that
could either be ambiguous
Mooney faces or nonfaces. Der
nonfaces in Experiment 1 war
scrambled versions of the
Mooney faces (“unrecognizable
shapes”) and in Experiment 2
were inverted Mooney faces.
After a short delay, Teilnehmer
were shown two images and
had to indicate which one
was part of the memory display
(top vs. bottom). Subsequently,
they were asked how many
faces they perceived during
encoding. (B) Example stimuli:
ambiguous Mooney faces
(used in all Experiments), unrecognizable shape stimuli made from scrambling Mooney images (used in Experiments 1 Und 3),
inverted ambiguous Mooney images (used in Experiment 2).
904
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 5
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
images used in the practice trials were used in the full
Experiment.
Statistical Analyses
Experiment 1 has a within-subject, one-factor design with
four levels (the number of stimuli that are upright faces,
0-3), as well as another factor, the subjectively reported
number of faces seen (also 0–3). The dependent measure
is accuracy in the 2-AFC memory test. As described in the
preregistration, our main analysis uses a generalized linear
mixed-effect model, taking all of the trials accuracy data
(0/1, binomial) and fitting a model that treats both partic-
ipants and items as random effects. We ask—separately
and together—about the effect of the objective number
of faces shown (0, 1, 2, 3) and subjective number of
reported faces (0, 1, 2, 3). Insbesondere, to look at the effect
of objective number of faces alone, we examine the effect
of the objective number of faces shown after taking into
account random effects of participants and items:
Correct ∼1 þ ObjectiveFaceNum þ 1jItem
D
Þ
þ 1jSubject
D
Þ
In a separate model, the effect of both objective and sub-
jective number of faces and their interaction is estimated,
again treating both subjects and items as random effects:
Correct ∼1 þ ObjectiveFaceNum * SubjectiveFaceNum
þ 1jItem
D
Þ þ 1jSubject
D
Þ
We fit these models using the fitglme function in MATLAB
(The MathWorks, Inc.), with a binomial distribution. We ob-
tained significance ( p values) using MATLAB’s defaults for
the ANOVA function (a Type III F test). These models allow
us to examine how accuracy is impacted by both the objec-
tive and subjective number of faces present, while taking
into account item effects.
We also assess performance across the objectively
shown number of faces with a repeated-measures
ANOVA, which does not take into account the effects of
individual items but may be more interpretable for
readers unfamiliar with the mixed-effect approach.
Ergebnisse
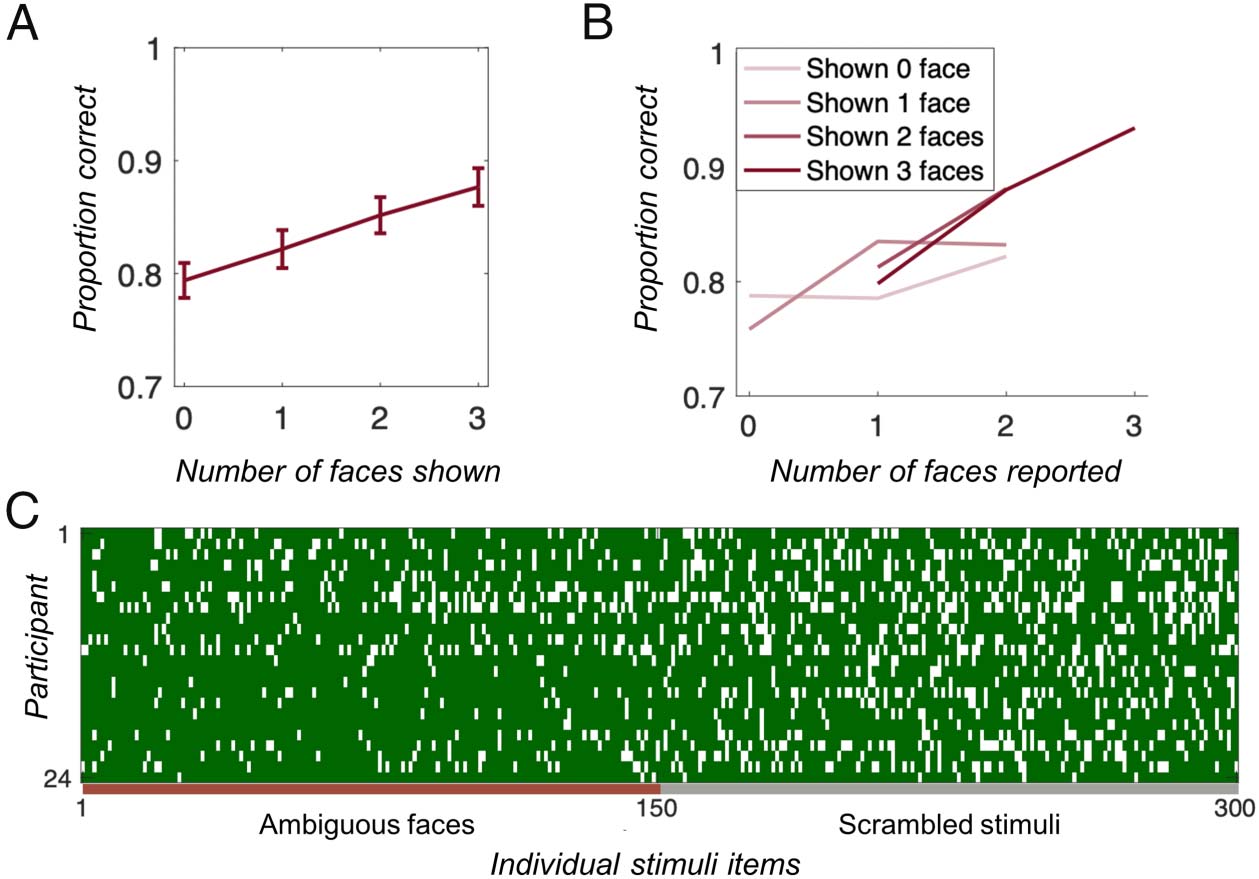
Participants remembered images more accurately when
the memory display objectively contained more faces:
Recognition accuracy increased when more faces, eher
than unrecognizable shape images, were presented dur-
ing the study phase, F(3, 69) = 18.7, P < .0001, despite
the unrecognizable shapes being made from the same un-
derlying set of shapes and correspondingly having the
same low- and mid-level features present as the faces. A
binomial general linear mixed-effect model showed this
finding was robust not only across individuals but also
after taking into account the effect of items (individual
faces/unrecognizable shapes): β = 0.18, F(1, 7198) =
34.2, p < .0001.
Rather than focusing solely on the objective number of
face stimuli present, we also analyzed the data as a func-
tion of both the objective number of faces present and
the subjective number of faces participants reported per-
ceiving. We hypothesized that faces that were present
objectively would only be beneficial for memory perfor-
mance when those faces were perceived by an individual,
that is, an interaction between these two factors. Thus, we
performed a general linear mixed-effect analysis, taking
into account the random effect of individuals and items
and examined the effect of both the objectively shown
and subjectively reported number of faces. We found a
significant effect of Number of Subjective Faces (β =
0.15, F(1, 7196) = 4.82, p = .028), no significant effect
of the Objective Number of Faces (β = −0.03, F(1,
7196) = 0.36, p = .55), and an interaction between these
two factors: (β = 0.09, F(1, 7196) = 6.82, p = .009), such
that when more objective faces were shown, the subjec-
tive perception of these faces was critical to performance
rather than their mere presence (Figure 2A–2B). In other
words, the effect of Subjective Number of Faces seen on
accuracy was larger when objectively more faces were
shown. Furthermore, because items were included as ran-
dom effects in the general linear mixed-effect model, we
can conclude that the memory improvement was robust
across the images. To further investigate whether there
were reliable item effects, such that certain images were
more likely to be remembered, we plotted individual per-
formance (correct/incorrect) for each stimulus tested and
did not find any reliable item effects, which further indi-
cates there was no consistency in which images were
more likely to be remembered (Figure 2C).
EXPERIMENT 2: BEHAVIORAL CAPACITY FOR
MOONEY FACES VERSUS INVERTED FACES
In Experiment 1, the ambiguous faces and scrambled faces
are tightly matched for visual features, as the scrambled
unrecognizable shapes were made by segmenting and
scrambling the set of Mooney images used in the study.
However, the stimuli do differ, as the scrambling process
did not perfectly preserve low- and mid-level features.
Thus, in Experiment 2, to ensure that the same physical
stimuli led to the same effect of recognition on memory
performance, we used a different control: inverted versions
of the ambiguous Mooney images. As in Experiment 1, the
design and analysis plan for Experiment 2 was preregistered
(aspredicted.org/blind.php?x=tt4pq2). The use of inverted
images more tightly controls for image features between
the recognizable and unrecognizable images since the same
low- and mid-level features are present, although it also
allows for the possibility that some inverted faces are recog-
nized and/or that participants might mentally rotate the im-
ages some proportion of the time. Convergence between
Experiment 1 and Experiment 2 would suggest that the
Asp, Störmer, and Brady
905
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
j
o
c
n
_
a
_
0
1
6
9
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Results of Experiment
1. (A–B) Accuracy (proportion
correct) for the 2-AFC memory
match judgment depended on
(A) how many faces were shown
in the memory display (error
bars are based on within-subject
SEM ) (B) and how many faces
participants reported seeing,
such that more faces perceived
tended to result in better
performance when more faces
were shown. Given the
uncertainty in both the x and y
variables, the within-subject
nature of the relevant
comparison, and the differing
number of trials at each point,
no error bars capture the
relevant variation in these
points. However, to get a sense
of the uncertainty per point,
Table 1 shows the proportion
of trials in each point. (C)
Individual trials, where green indicates a correct response and white indicates an incorrect response. The lack of vertical columns indicates that
individual items did not drive the effect, as although participants remembered faces better than scrambled images, participants were not consistent in
which faces or scrambled images tended to be accurately reported.
effects we report are robust to how we choose to control
for low- and mid-level features.
Methods
Participants
Experiment 2 was preregistered (aspredicted.org/blind.
php?x=tt4pq2), and, in accordance with the preregistra-
tion, 24 undergraduates from the University of California,
San Diego, participated, also for course credit (19 women,
ages 18–32 years). One participant had performance below
our preregistered exclusion criterion (average memory
performance across conditions below chance) and was
replaced.
Stimuli and Procedure
Experiment 2 was identical to Experiment 1 except that,
rather than using scrambled Mooney faces images, it used
inverted Mooney faces (Figure 1B). Thus, each trial con-
sisted of either zero, one, two, or three upright ambiguous
faces with inverted faces making up any remainder so that
the memory set would always consist of three images.
This more tightly controls for image features between
the recognizable and unrecognizable images, although it
also allows for the possibility that some inverted faces are
perceived as faces and/or that participants might mentally
rotate the images some proportion of the time.
Statistical Analyses
Analysis was identical to Experiment 1.
Results
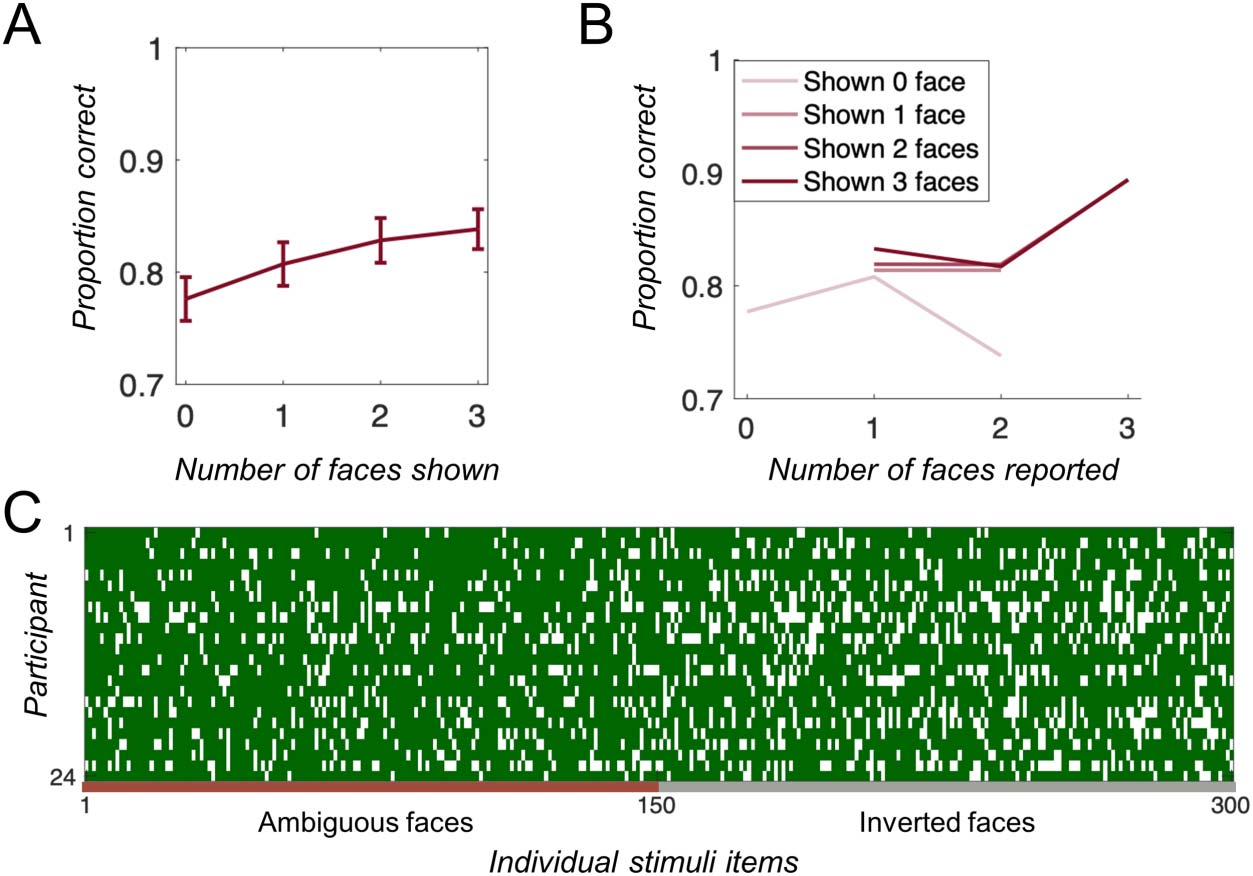
Recognition accuracy increased when more faces, rather
than inverted faces, were presented during the study
phase, F(3, 69) = 6.83, p = .0004. A binomial general linear
mixed-effect model showed this finding was robust not
only across individuals but also after taking into account
the effect of Items: β = 0.11, F(1, 7198) = 14.3, p = .0002.
To examine the effect of Subjective Perception of Faces,
we again performed a general linear mixed-effect analysis
taking into account the random effect of participants and
items and examining the effect of both the objectively
shown and subjectively reported number of faces. We
found no main effect of Number of Subjective Faces (β =
0.03, F(1, 7196) = 0.23, p = .63), no main effect of the
Objective Number of Faces (β = −0.09, F(1, 7196) =
2.98, p = .08), but an interaction between these two factors:
(β = 0.11, F(1, 7196) = 13.1, p = .0003), such that, when
more upright faces were shown, the subjective percep-
tion of these faces was critical to performance rather
than their mere presence (Figure 3A–3B). That is, just like
in Experiment 1, there was an effect of the Subjective
Number of Faces reported that was larger when more
faces were objectively present. In addition, looking at in-
dividual performance for each stimulus tested revealed no
evidence of any reliable Item effects, which again suggests
there was no consistency in which images were more
likely to be remembered (Figure 3C).
Whereas Experiment 1 found a main effect of these two
factors in addition to an interaction, here, we see only an
interaction with no significant main effects. This arises
because of a difference in how frequently people report
different numbers of subjective faces, despite the same
effects of both subjective and objective faces on
906
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
j
o
c
n
_
a
_
0
1
6
9
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Results of Experiment
2. (A–B) Accuracy (proportion
correct) for the 2-AFC memory
match judgment depended on
(A) how many faces were shown
in the memory display (B) and
how many faces participants
reported seeing, such that more
faces perceived tended to result
in better performance when
more faces were present. Given
the uncertainty in both the
x and y variables, the within-
subject nature of the relevant
comparison, and the differing
number of trials at each point,
no error bars capture the
relevant variation in these
points. However, to get a sense
of the uncertainty per point,
Table 1 shows the proportion of
trials in each point. (C)
Individual trials, where green
indicates a correct response and
white indicates an incorrect response. The lack of vertical columns indicates that individual items did not drive the effect, as although participants
remembered faces better than inverted-face images, participants were not consistent in which faces or inverted-face images tended to be accurately
reported.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
j
o
c
n
_
a
_
0
1
6
9
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
performance in both experiments. In particular, it arises
because, in Experiment 2, people saw faces more fre-
quently in stimuli that did not have faces than they did
in Experiment 1 (Table 1). Most likely, this is because in-
version is just less effective than the scrambling in reduc-
ing face perception (e.g., Experiment 1, when zero face
was shown, 50% of the time 0 were reported as shown;
in Experiment 2, when zero face was shown, 25% of the
Table 1. Frequency of Total Trials Reporting Each Subjective
Number of Faces for Each Objective Number of Faces for Each
of the Three Experiments
Experiment 1 Reported 0 Reported 1 Reported 2 Reported 3
Shown 0
Shown 1
Shown 2
Shown 3
51%
22%
11%
6%
32%
52%
31%
19%
16%
24%
48%
37%
2%
3%
10%
39%
Experiment 2 Reported 0 Reported 1 Reported 2 Reported 3
Shown 0
Shown 1
Shown 2
Shown 3
24%
14%
9%
6%
30%
31%
25%
15%
25%
41%
45%
40%
12%
14%
22%
39%
Experiment 3 Reported 0 Reported 1 Reported 2 Reported 3
Shown 1
Shown 3
21%
5%
38%
25%
31%
42%
10%
28%
time 0 were reported as shown). Thus, a larger proportion
of trials happen to be low objective/low subjective in
Experiment 1 than Experiment 2. This difference between
experiments is consistent with the general advice to not
interpret main effects when interactions are present:
Although the effect of both objective and subjective face
perception on performance appears the same in the
two experiments, the significance of the main effects is
different because of the different proportion of trials in
each bin.
EXPERIMENT 3: ACTIVE STORAGE ASSESSED
VIA THE CDA
Experiments 1 and 2 show a strong behavioral advantage
not only to presenting participants with faces rather than
meaningless shapes or inverted faces to remember, but
with participants’ own perception of those stimuli as faces.
This was true even after taking into account the effects of
individual items, consistent with our previous work in
which we found almost no reliability in which items were
perceived as faces from the same stimulus set (Brady et al.,
2019). These data provide initial evidence that visual work-
ing memory performance may benefit from the perceived
meaningfulness of the stimulus: When remembering an
image that connects to a concept (e.g., a face), that image
is remembered better than when the same visual informa-
tion is not perceived as meaningful but only seen as some-
thing arbitrary (e.g., meaningless black and white shapes).
This contrasts with a view where working memory capacity
is fixed by the total number of items, independent of the
content of those items (e.g., three to four objects regard-
less of complexity; Awh et al., 2007) and/or is based solely
Asp, Störmer, and Brady
907
on the persistence of sensory information in low-level
visual cortex, with images being held in a perceptual-like
unprocessed state (e.g., Serences, 2016) and thus only
the number of distinct low-level features being relevant
to “how much” capacity is available. Instead, consistent
with previous work (e.g., Ngiam et al., 2018; Brady et al.,
2016; Jackson & Raymond, 2008; Alvarez & Cavanagh,
2004) but with even tighter control on stimulus features
than this previous work, we find that recognized and
familiar stimuli result in better behavioral performance
than unrecognized or unfamiliar stimuli.
Similar effects of meaningfulness and improved mem-
ory performance have previously been found in episodic
long-term memory (Brady et al., 2019; Wiseman & Neisser;
1974; Bartlett, 1932), and it has been repeatedly found that
activated forms of long-term memory that can be used for
short-term storage of information (e.g., Cowan, 2005) take
advantage of knowledge (e.g., Ericsson & Kintsch, 1995).
In addition, some authors have claimed that effects like we
find here derive solely from confusability of the item pairs
in the test (e.g., a decision effect), rather than a true effect
of working memory maintenance (Awh et al., 2007). Thus,
to test whether the increase in memory performance ob-
served here was because of increases in active storage of
information in working memory, or reflect a lack of pro-
cess purity because of the use of other memory systems,
such as episodic long-term memory, or a decision effect,
we recorded the brain’s electrophysiological responses
that provide a direct measure of working memory engage-
ment in Experiment 3.
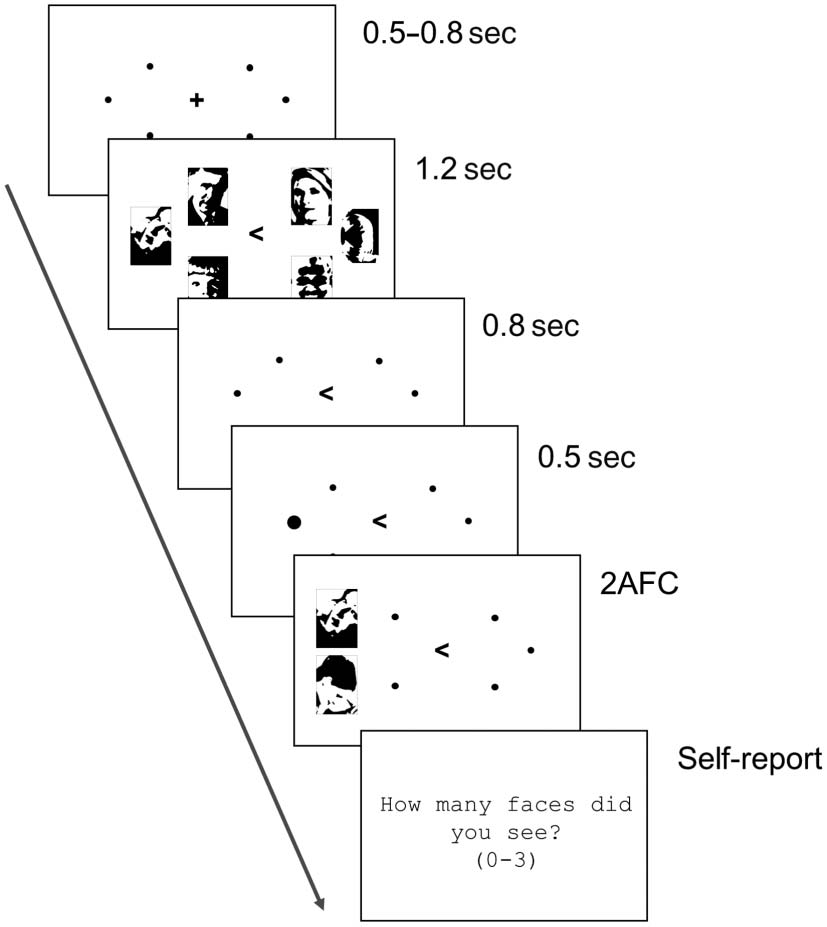
In particular, we adapted the experiment to an EEG ver-
sion of the task and showed the stimuli on a lateralized dis-
play with three images in each visual half-field, and cued
participants with a central arrow to only memorize one
side of the display (either left or right; Figure 4). This al-
lowed us to measure the CDA, a neural marker of active
maintenance of visual information ( Vogel & Machizawa,
2004). Just like in Experiment 1, we presented ambiguous
face stimuli and scrambled versions of them, but this ex-
periment only included two conditions: one face versus
three faces, to obtain a sufficient number of trials.
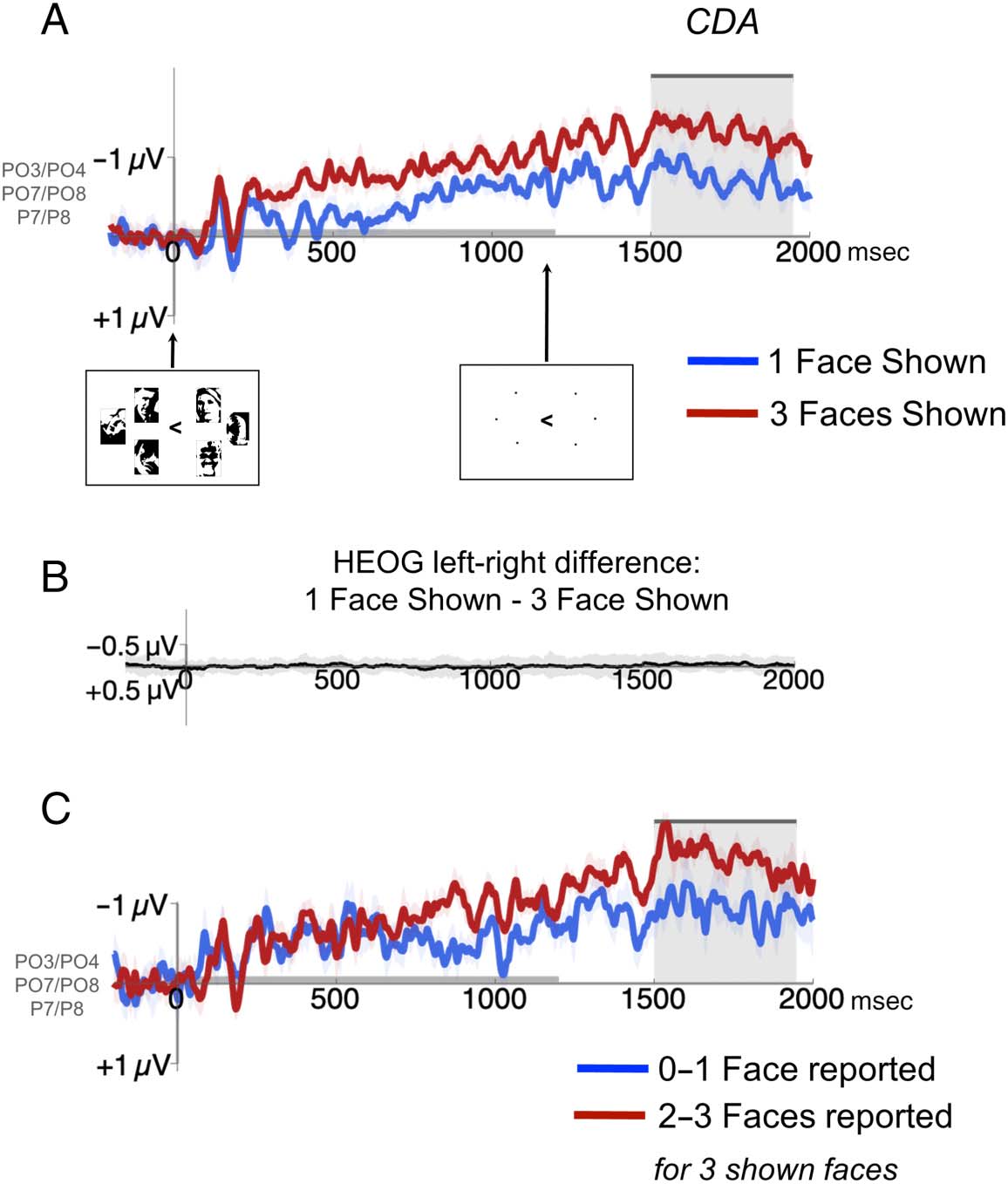
The CDA is a lateralized slow wave that occurs during the
retention interval and tracks how much information is ac-
tively maintained (Luria, Balaban, Awh, & Vogel, 2016;
Vogel, McCollough, & Machizawa, 2005), and thus can be
used to infer that the information is being held actively in
mind rather than stored in a passive way using a long-term
memory system (Brady et al., 2016), and can distinguish be-
tween a change in how much information is maintained
versus decision-level effects like confusion at test (Luria
et al., 2010; Awh et al., 2007). It is often taken as an index
of the focus of attention in visual working memory—for ex-
ample, the active storage component of the memory sys-
tem, as it has been shown to increase with the number of
items participants hold in working memory ( Vogel &
Machizawa, 2004) and decrease when items are dropped
from working memory ( Williams & Woodman, 2012;
Figure 4. Methods of Experiment 3 (EEG experiment). Participants were
cued to remember the items on one side of the display only (left or right).
They then had to remember the three images shown on that side of
the display and, then, after a delay, report which image was seen in a
particular location. They then self-reported how many faces they had
perceived on the to-be-remembered side of the initial display (0–3).
Vogel et al., 2005); furthermore, the CDA correlates with
the individual’s working memory capacity (Störmer, Li,
Heekeren, & Lindenberger, 2013; Vogel & Machizawa,
2004) and disappears when items have been consolidated
into episodic long-term memory (Schurgin, Cunningham,
Egeth, & Brady, 2018; Carlisle, Arita, Pardo, & Woodman,
2011). Finally, and of particular interest to this study, the
CDA is not sensitive to the perceptual effort of encoding
stimuli into memory or differentiating them at test, instead
reflecting memory, not effort (Ikkai, McCollough, & Vogel,
2010). Thus, Experiment 3 allows us to test whether the in-
crease in memory performance observed in Experiments 1
and 2 was because of increases in active storage of informa-
tion in working memory—if it is reflected in the CDA—or
whether it reflects a lack of process purity because of the
use of other memory systems, such as episodic long-term
memory, or reflects a decision effect rather than any mem-
ory representation change.
Methods
Participants
Twenty-three participants completed Experiment 3 and
received course credit or $10 per hour for participation.
Data from three participants in Experiment 3 had to be ex-
cluded because of artifacts in the EEG (> 50% of trials ex-
eingeschlossen), and data from one participant had to be excluded
because part of the session was not recorded. Of the
908
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 5
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
remaining 19 Teilnehmer (13 Frauen, ages 18–34 years)
all had normal or corrected-to-normal vision.
Stimuli and Procedure
Materials and stimuli were almost identical to those used
in Experiment 1. Jedoch, in Experiment 3, EEG activity
was measured throughout the experiment to examine the
CDA. The CDA is a lateralized slow wave that occurs during
the retention interval and tracks how much information is
actively maintained (Luria et al., 2016; Vogel et al., 2005).
Daher, in Experiment 3, we asked whether the greater
working memory capacity observed for recognizable ver-
sus unrecognizable images resulted in greater CDA as well
as greater performance. If the behavioral performance in-
crease is mirrored by a CDA increase, this would suggest
that the capacity increase is mediated by greater ability to
actively hold meaningful information in visual working
memory—the recruitment of additional neural resources
for more meaningful stimuli—rather than the use of other
memory systems (like passive storage in visual long-term
Erinnerung).
In order to record the lateralized CDA component,
stimuli were presented on a lateralized display with three
images on the left and three images on the right visual
half-field, and participants were cued with a central arrow
on which side to remember (Figur 4). Außerdem, Zu
obtain a sufficient number of trials, we only included
conditions in which either three ambiguous faces or one
ambiguous face (and two unrecognizable shape images)
was shown on each side. Because there were now six stim-
uli being displayed in a single study screen, the number of
times each image was repeated increased. Face images
were repeated ∼8–9 times, and unrecognizable shape im-
ages were repeated about 4 times throughout the experi-
ment. This was because of the trial conditions that were
chosen (one-face and three-face trials). Wichtig, Wenn
the participant successfully attended to one side only,
the high repetition would not be overtly noticeable, Sei-
cause only half of the number of repetitions was on the
task-relevant side of the display.
Each trial started with the presentation of six dots that
served as placeholders; three were positioned on the left
half of the screen in a semicircle, and the other three were
presented on the right half of the screen. These dots ap-
peared for 500–800 msec ( jittered) and served as location
markers for the later images. Then six images appeared on
the screen as well as a central arrow cue; three images
were positioned on the left half of the screen, und das
other three were presented on the right half of the screen.
The central arrow cue, indicating which side to memorize,
appeared at the same time as the stimuli. The images were
presented for 1200 msec to allow ample time to recognize
the arrow cue, shift attention to the cued side covertly, Und
encode the images. Participants were required to maintain
fixation while the images were on the screen as well as
during the delay period. daher, participants could only
use their peripheral vision to study the images on the cued
Seite, and compliance with this was ensured by rejecting
trials with horizontal eye motion from the analysis (sehen
Electrophysiological Recordings and Analysis section).
After the images disappeared, there was an 800-msec delay
period with only location markers on the screen.
Following the delay, one of the location markers on the
cued side was enlarged and presented for 500 ms, indi-
cating the location of the to-be-tested image. Once the test
images appeared, participants were allowed to blink and
move their eyes. The experiment consisted of 10 blocks
mit 32 trials each. The trial conditions (one vs. three faces
in each study array) were randomized throughout the ex-
periment, and each trial condition was presented evenly
within each block. Prior to the experiment, all participants
completed two mini-blocks of four trials each as practice
for the full 10 experimental blocks that followed. None of
the images used in the practice trials were used in the full
Experiment.
Electrophysiological Recordings and Analysis
EEG activity was recorded continuously from 32 Ag/AgCI
electrodes arranged according to the 10–20 system,
mounted in an elastic cap and amplified by an ActiCHamp
amplifier (Brain Products GmbH). The horizontal EOG
(HEOG) was acquired using a bipolar pair of electrodes po-
sitioned at the external ocular canthi, and the vertical EOG
was measured at electrode FP1, located above the left eye.
All scalp electrodes were referenced to an electrode on the
right mastoid online. The EEG was filtered online with a cas-
caded integrator-comb antialiasing filter with a half-power
cutoff at 130 Hz and then digitized with a 500-Hz sampling
rate. Signal processing was performed with MATLAB using
the EEGLAB (Delorme & Makeig, 2004) and ERPLAB tool-
boxes (Lopez-Calderon & Luck, 2014) and custom-written
scripts. Continuous EEG data were filtered off-line with a
bandpass of 0.01–112 Hz (half amplitude cutoff, with a
slope of −12 dB/octave). Data were epoched into trials,
and artifact rejection was performed for the interval −200
Zu +2000 msec relative to the memory display onset, Und
trials with horizontal eye movements, blinks, or excessive
muscle movements were excluded from the analysis.
Artifacts were detected using a semi-automated procedure,
in which differences between minimum and maximum volt-
ages were compared with a threshold value. Threshold
values were determined by visual inspection for each partic-
ipant individually. After using the artifact detection func-
tions implemented in ERPLAB (Lopez-Calderon & Luck,
2014; peak-to-peak for blinks at channel FP1 and a step
function to detect horizontal eye movements at the
HEOG channel), we rechecked the artifact detection visu-
ally to ensure that all eye movements and blinks were de-
tected correctly. If additional artifacts were spotted, diese
trials were excluded as well. For the subjects retained in
the final data set (n = 19), this resulted in an average re-
jection rate of 18.3%.
Asp, Störmer, and Brady
909
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Artifact-free data were rereferenced to the average of the
left and right mastoids, digitally low-pass filtered (half-
amplitude cutoff at 30 Hz; slope of 12 dB/octave), and base-
lined to the 200-msec prestimulus interval. ERPs elicited by
the memory display were averaged separately for each con-
dition (one face shown, three faces shown) and were then
collapsed across to-be-remembered hemifield (links, Rechts)
and hemisphere of recording (links, Rechts) to obtain wave-
forms recorded contralaterally and ipsilaterally to the
to-be-remembered side. Endlich, because the CDA is quan-
tified as the difference in activity across hemispheres, Wir
calculated contralateral-minus-ipsilateral difference wave-
forms for each condition. For each participant, der Mittelwert
CDA amplitude was measured with respect to the 200-msec
prestimulus baseline at six posterior electrode sites (PO3/
PO4/PO7/PO8/P7/P8) beginning 300 msec after the offset
of the memory display until the test probe appeared (in diesem
Experiment, 1500- to 1950-msec post memory display on-
set), consistent with previous work (Brady et al., 2016;
McCollough, Machizawa, & Vogel, 2007). The resulting
mean amplitudes were our measure of the CDA magnitude.
The exact electrode sites were chosen a priori based on a
pilot participant that was not used in the main analysis. Wir
did not use the same sites as our previous work (Brady et al.,
2016) because the EEG caps used were slightly different (Die
previous work was done at a different university with a differ-
ent system). Jedoch, to ensure that the results were not
because of the specific electrodes we used, we repeated all
statistical analysis using electrode sites PO3/PO4/PO7/PO8
that match our previous work (Brady et al., 2016); the rele-
vant effects remain significant (difference in CDA amplitude
between objectively shown one face versus three faces: p =
.028; difference in CDA amplitude between subjectively re-
portiert 0/1 vs. 2/3 faces: p = .042).
To ensure that any lateralized effects indexed by the
CDA component were not because of differences in spuri-
ous eye movements between the conditions, we com-
pared the HEOG waveforms for remember-left versus
remember-right conditions during the CDA time window,
separately for the two set sizes (one face vs. three faces),
and found no reliable difference for each of the left versus
right comparisons, T(18) = 1.64, p = .12, for the one-face
condition, and t(18) = 1.38, p = .19, for the 3-face condi-
tion. Most importantly, the left-minus-right difference
waves did not differ between the conditions, T(18) =
0.16, p = .87 (see Figure 5B), demonstrating that the
CDA differences we observed between conditions are
not because of differences in eye movements.
Figur 5. Results of Experiment
3. (A) Contralateral-minus-
ipsilateral difference waveform
over parietal-occipital electrode
sites for when one face was
shown (blue line) versus three
faces were shown (red line).
The CDA is measured 300 ms
after offset of the memory
display until the test probe
appears (shaded gray area).
Shaded areas represent
within-subject standard errors
of the mean at each time point.
(B) We found no eye drifts that
could affect the CDA magnitude
differentially: As reported in the
Methods section, we found
no significant HEOG difference
for remember-left versus
remember-right trials in either
condition separately. In
addition, plotted here is the
difference-of-differences: Wenn
small differences in left–right
eye drift caused the difference
in CDA between one face
shown versus three faces
shown, this would be reflected
in a deviation from 0 in diesem
plot, which is not present. (C)
Contralateral-minus-ipsilateral
difference waveform over
parietal-occipital electrode sites
for when three faces were
shown, broken down by when zero to one face were reported (blue line) versus two to three faces were reported (red line). The CDA is measured
300 msec after offset of the memory display until the test probe appears (shaded gray area). Shaded areas represented within-subject standard errors
of the mean at each time point.
910
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 5
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Statistical Analyses
Experiment 3 has a within-subject, one-factor design with
two levels (the number of stimuli that are faces, 1 oder 3) Und
another factor of the subjectively reported number of
faces (0-3).
The behavioral dependent measure is again accuracy
in the 2-AFC memory test. In addition to fitting the same
generalized linear mixed-effect model from Experiments 1
Und 2, the effect of the objective number of faces can now
also be compared via a paired t test because there are only
two levels.
The neural measure in Experiment 3 is the CDA. Für
comparing CDA based on the objective and subjective
number of faces, we also rely on a linear mixed-effect
model to take into account not only random effects of
subjects but also items, taking advantage of the benefits
of a mixed- effects analysis for electrophysiology (Bagiella,
Sloan, & Heitjan, 2000). Insbesondere, we calculate CDA on
a per trial basis by: (1) first subtracting the baseline sepa-
rately for each relevant electrode for each trial, (2) then sub-
tracting contralateral electrodes from ipsilateral electrodes,
and then (3) averaging over the CDA time window. Dann,
we fit a model that takes into account either only the objec-
tive number of faces:
CDA ∼ 1 þ ObjectiveFaceNum þ 1jItem
D
Þ þ 1jSubject
D
Þ
or both the objective and subjectively reported number of
faces:
CDA ∼ 1 þ ObjectiveFaceNum * SubjectiveFaceNum
Þ
Þ þ 1jSubject
þ 1jItem
D
D
We fit these models using the fitlme function in MATLAB,
using a rank-based inverse normal transformation on CDA
Werte (Bliss, 1967). Nearly identical results were obtained
when using untransformed CDA values, but the residuals
were not normally distributed, so the transformation was
necessary to meet the assumptions of the statistical test.
We obtained significance ( p values) using MATLAB’s de-
faults for the anova function (a Type III F test). Zusätzlich,
to test the effect solely from the objective number of faces
without controlling for item effects, we report a paired
t test of CDA magnitude.
The main measure of the effect of objectively shown
number of faces and subjectively reported number of faces
is the mixed-effect model, which makes use of all partici-
pants and all trials and takes into account item effects.
Jedoch, for a follow-up subjective-report analysis, Wir
also report the effect of looking only at performance with
three objective faces present, which uses only a subset
of the data used in the main mixed-effect analysis but
may be more interpretable for readers unfamiliar with
the mixed-effect approach. To do so, we averaged ERPs ac-
cording to individual’s reports of how many faces they re-
ported seeing on each trial but only included those trials
on which we showed three ambiguous faces. To have a suf-
ficient number of trials in each condition, we collapsed
across reports where participants said they saw zero or
one face, and two or three faces. Außerdem, for this par-
ticular analysis, we a priori restricted our EEG analysis to
those participants who had a minimum of 20 trials in each
bin, ergebend 14 usable participants for this analysis. An
average, the remaining participants had well over 20 Versuche
per bin, with the average number of trials per bin being 67
in these participants.
Ergebnisse
In line with Experiments 1 Und 2, we found that recogni-
tion accuracy increased when more faces, rather than un-
recognizable shapes, were presented during the study
Phase, T(18) = 2.63, p = .017, and this was also held after
taking into account the effect of items (β = 0.076, F(1,
4964) = 4.1, p = .04).
Wichtig, this performance benefit was accompa-
nied by increased CDA for the three- versus one-face con-
dition, T(18) = 2.92, p = .009, which was also robust when
taking into account the random effect of items (β = .035,
F(1, 7198) = 6.12, p = .01). Daher, there is a change not
only in behavioral performance, but this change is also
reflected in increased CDA when more faces are shown,
suggesting more information is actively held in visual
working memory for faces than nonfaces.
To test to what extent the increase in working memory
capacity was because of subjectively perceiving the stimuli
as faces, we again looked at behavioral performance as a
function of subjectively reported number of faces. Wir
found both a main effect of Number of Subjective Faces
(β = 0.21, F(1, 4962) = 5.46, p = .019), a main effect of
the Objective Number of Faces (β = 0.16, F(1, 4962) =
4.35, p = .037), Und, in diesem Fall, no significant interaction
(β = −0.07, F(1, 4962) = 2.7, p = .098). This provides
evidence that not only the objective number of faces
shown, but also the subjective number of faces reported,
drives behavioral performance.
Wichtig, the effect of subjectively reported number
of perceived faces was also found in the CDA data, Wo
we observed a main effect of Number of Subjective Faces
(β = −0.09, F(1, 4962) = 0.23, p = .012), no main effect of
the Objective Number of Faces (β = −0.04, F(1, 4962) =
2.98, p = .23), and an interaction between these two
factors (β = 0.04, F(1, 4962) = 13.1, p = .006), such that
when more objective faces were shown, the subjective
perception of these faces was critical to performance
rather than their mere presence. This suggests that not
only behavioral performance but also neural activity in-
dicative of working memory storage is modulated by
the subjective number of perceived faces.
Whereas this mixed-effect analysis makes use of all of
the ∼5000 trials of data to estimate the effect of objective
and subjective number of faces, we also performed a
follow-up analysis that split the data into trials where par-
ticipants reported seeing zero or one face and trials where
participants reported seeing two or three faces (only using
Asp, Störmer, and Brady
911
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
trials in which we showed three ambiguous faces). This ad-
dresses the role of subjective perception when all three stim-
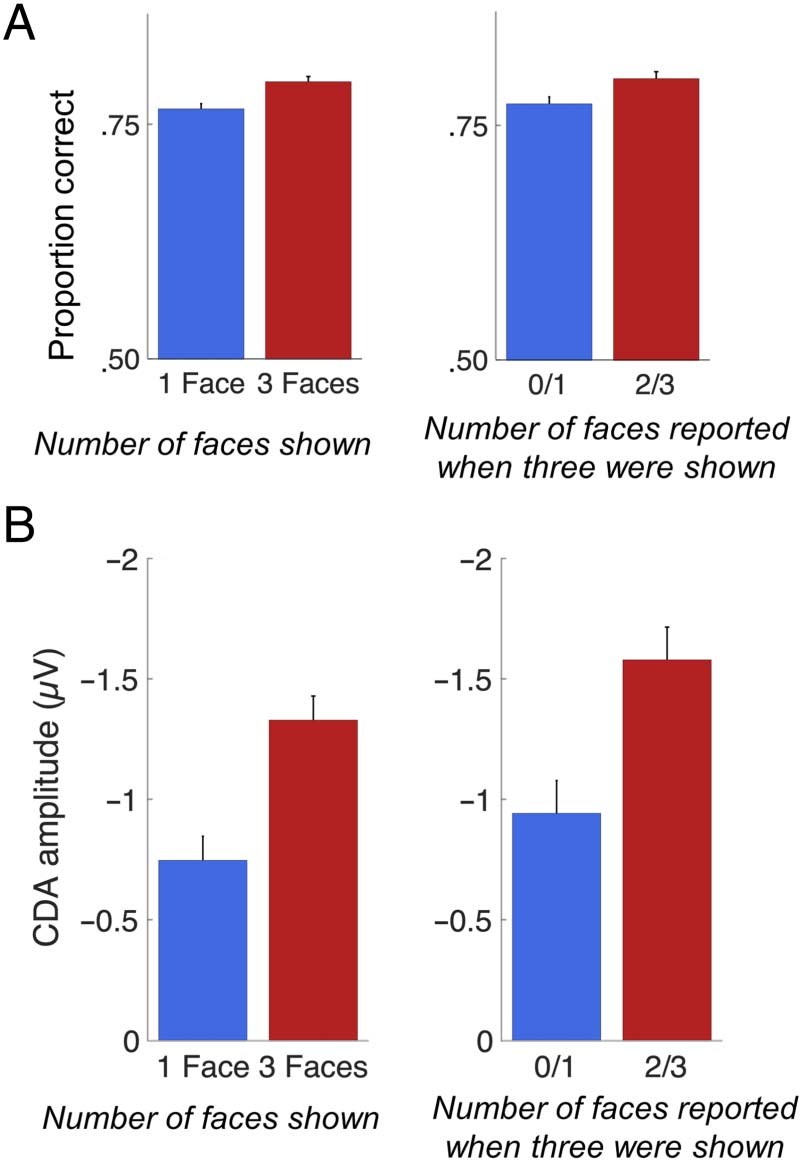
uli are faces. We found that performance was higher on trials
where participants reported seeing more faces versus fewer
faces, T(18) = 2.83, p = .011 (see Figure 6A). Zusätzlich,
looking at individuals who had enough trials to analyze
the magnitude of the CDA for each bin, the CDA ampli-
tude was larger for trials where more faces were perceived
during encoding compared to trials where fewer faces
were seen, T(13) = 2.35, p = .035 (Figuren 5 and 6B).
Tisch 1 reports the frequency with which trials were
present in these bins, for both this experiment and
Experimente 1 Und 2. Wie erwartet, and consistent with this
Analyse, participants do not report seeing the exact num-
ber of faces that were being presented to them, allowing
us to examine both the subjective perception of the stim-
uli as well as the objective number of faces presented.
Zusammen, these data show that active storage in visual
working memory is modulated by meaningfully perceived
Reize, with such stimuli recruiting additional storage ca-
pacity. This provides evidence against accounts where be-
havioral benefits for familiarity (z.B., Ngiam et al., 2018;
Jackson & Raymond, 2008; Alvarez & Cavanagh, 2004)
arise solely from decision-level effects like reduced
Figur 6. Results of Experiment 3. (A) Accuracy (proportion correct)
for the 2-AFC memory match judgment was higher when the memory
display consisted of three faces versus one face (links), and when
participants reported seeing two or three faces versus zero or one face
on three-face trials (Rechts). (B) Mean CDA amplitude was higher when
the memory display consisted of three faces versus one face (links), Und
when participants reported seeing two or three faces versus zero or one
Gesicht (Rechts).
confusability of stimuli at test (Awh et al., 2007), or arise
solely from the recruitment of other memory systems, wie
visual long-term memory.
GENERAL DISCUSSION
Zusammen, the behavioral and EEG results suggest that vi-
sual working memory performance is heavily influenced
by the type of information being stored. Speziell, Wir
found that an observer’s ability to extract meaningful
information from a stimulus enables them to hold more
information actively in visual working memory, even for
physically identical stimuli, as the same stimulus is remem-
bered better when it is perceived as a face than when it is
nicht. Daher, our data suggest that visual working memory
capacity is not fixed to a certain number of objects regard-
less of physical complexity or a globally fixed pool of
resources, even for physically identical stimuli with the
same orientations, spatial frequencies, and other visual
information present. Stattdessen, it critically depends on the
type of information that is remembered—not how visually
complex that information is, but rather whether that infor-
mation can be perceived as a meaningful feature or not.
Our data are broadly consistent with models where more
features being present allow participants to store more
information in memory (z.B., Brady et al., 2016; Wyble,
Swan, & Callahan-Flintoft, 2016; Luck & Vogel, 2013), mit
the added idea that these additional features are not
simply low-level features. Stattdessen, additional meaningful
face-level features are available for stimuli perceived as
faces. Zusätzlich, our data argue against theories of visual
working memory that propose that when having to re-
member the visual details of an object, as in the current
Aufgabe, persistence of sensory-like activity in low-level visual
Regionen (z.B., V1) alone is responsible for memory perfor-
Mance, instead arguing for a major role for high-level inter-
pretations of these visual details, possibly via persistence
in higher-level visual areas that are selectively recruited for
meaningful stimuli, such as the fusiform face area (z.B.,
Druzgal & D'Esposito, 2001) or via the modulation of
low-level regions by high-level interpretation.
Active Storage Measured via the CDA
Active storage of information, and thus the use of working
Erinnerung, is reflected in persistent neural activity in frontal
and parietal brain regions (Buschman, Siegel, Roy, &
Müller, 2011), and can be measured in the electroenceph-
alogram with the CDA. The CDA is often taken as an index
of the focus of attention in visual working memory—for
Beispiel, the active storage component of the memory
System. Insbesondere, evidence for its selectivity to active
storage in visual working memory includes the following:
(1) Its magnitude increases with the number of items par-
ticipants hold in working memory ( Vogel & Machizawa,
2004) and decreases when items are dropped from
Arbeitsgedächtnis ( Williams & Woodman, 2012; Vogel
912
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 5
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
et al., 2005); (2) the CDA correlates with individual’s
working memory capacity (Störmer et al., 2013; Vogel &
Machizawa, 2004); (3) und am wichtigsten, the CDA
disappears when items have been consolidated into epi-
sodic long-term memory (Schurgin et al., 2018; Carlisle
et al., 2011). Daher, the fact that we found an increased
CDA for recognizable versus unrecognizable images is con-
sistent with the idea that the capacity increase observed
behaviorally is mediated by greater ability to actively hold
meaningful information in visual working memory rather
than the use of other memory systems (like passive storage
in visual long-term memory) or arising solely from decision-
level effects like reduced confusability of stimuli at test
(as proposed by Awh et al., 2007).
Our data are not consistent with an account where peo-
ple have a fixed capacity in terms of the number of objects
regardless of complexity (z.B., Awh et al., 2007) or other
memory resources and that unrecognizable shape stimuli
simply take up more of these “slots” or resources since
they cannot be organized (as has sometimes been claimed
about polygons and other complex stimuli; see Luria et al.,
2010). This is because such an account predicts either
equivalent CDA engagement in the two conditions,
Weil, in both cases, working memory is “full” or, wenn überhaupt-
thing, greater CDA in the condition with the unrecogniz-
able shapes, as they each require multiple slots or greater
resource usage in memory. Stattdessen, we find that the face
stimuli not only result in better performance but also
greater sustained neural activity as measured by the
CDA, suggesting the recruitment of additional memory
resources for the meaningful stimuli. This is in direct con-
trast to the predictions of fixed capacity accounts of visual
Arbeitsgedächtnis, which argue that only additional low-
level features can be used to recruit additional capacity.
Daher, our data provide evidence that meaningfulness—
rather than the visual information on our retinas—underlies
visual working memory capacity.
In the current work, we test only set size 3. Es ist möglich
that testing different set sizes would reveal a difference
between “how many” items are remembered versus “how
accurately” each is perceived/remembered, with meaning-
fulness having a differential impact on such factors. We be-
lieve the current evidence from both fitting continuous
report data (z.B., Schurgin, Wixted, & Brady, 2020; Bays,
2015; Van den Berg, Schienbein, Chou, George, & Ma, 2012) Und
from receiver operating characteristic analysis (Robinson,
Benjamin, & Irwin, 2020) strongly favors a view where items
simply vary continuously in how much noise they are en-
coded with or how much noise is accumulated during the
delay, rather than a view where a large fraction of items that
were seen 1 sec ago are completely lost as though they had
never been seen. Because the most up-to-date frameworks
do not feature all-or-none forgetting of items from memory,
and thus reject this distinction between “how many” and
“how precisely” items are remembered, we do not focus
on it here: Stattdessen, we emphasize only that additional
memory capacity can be recruited for meaningful stimuli.
Our finding of increased active storage for more mean-
ingful stimuli differs from other studies that found in-
creased behavioral performance for familiar relative to
unfamiliar stimuli, similar to our behavioral effects, Aber
did not observe an increase in neural delay activity for
the familiar stimuli. Zum Beispiel, when Zimmer et al.
(2012) and Zimmer and Fischer (2020) trained partici-
pants on a subset of Chinese characters, they showed
higher behavioral performance in a working memory task
for these learned stimuli, but less neural delay activity for
the familiar characters relative to unfamiliar characters, bei
least at low set sizes. Zimmer et al. suggested that memory
for familiar characters was supported by long-term mem-
ories acquired during training (z.B., word form acquisition;
Zimmer & Fischer, 2020; Zimmer et al., 2012)—effectively,
people had learned abstract labels for the characters,
which could be used to support performance in the work-
ing memory task with less stimulus-specific information
needing to be encoded. This is broadly consistent with
our suggestion about the role of active storage in working
Erinnerung: In our study, the stimuli were generally novel,
and perceiving a face in the stimulus allowed participants
to encode additional stimulus-specific information (z.B.,
face-specific features) that could help support memory,
resulting in increased delay activity. Im Gegensatz, im
Zimmer et al. arbeiten, because the stimuli were well learned,
fewer features needed to be encoded to support memory
for the learned stimuli—a simple label (verbal or concep-
tual) is sufficient to remember them—and so participants
did not store as many specific visual features to succeed at
the task, resulting in reduced delay activity.
Subjective and Objective Effects
In the current work, we show evidence for two effects: Wir
show that subjective perception modulates how accu-
rately Mooney faces are remembered, and we show that
the objective status of the image as a face or not (entweder
scrambled or inverted) also modulates how accurately
these images are remembered. This provides converging
evidence that it is truly the perception of the stimuli as
meaningful that is relevant both for behavioral perfor-
mance and for increased working memory-related neural
Aktivität. Insbesondere, neither analysis, taken alone, con-
trols for all possible alternative accounts: Zum Beispiel,
the analysis conditioning on the subjectively reported
number of faces is inherently correlational. Daher, im-
ory, prestimulus states could modulate both how many
faces participants report and how well they perform in
the memory task. Zusätzlich, small differences in low-level
features or mid-level features could, in theory, account for
the objective face effect. Jedoch, taken together, the two
results support our main conclusion that seeing something
as a face results in higher memory capacity, given that we
find the same effect when we show participants objectively
more faces, a causal experimental manipulation, und in
participants’ subjective reporting of their experience.
Asp, Störmer, and Brady
913
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Differential Attention to Faces
Might the benefit of face perception to memory in these
studies arise in part because people are more interested
in faces, and so differentially attend to faces within a trial?
This cannot straightforwardly account for the benefit to
overall performance of more faces being present, das ist
because our conditions do not refer to trials where a “face”
was probed versus not probed (which could be affected by
differential attention), but instead to average performance
for a random item from the entire display, regardless of
which item was probed—so if nonfaces were impaired
but faces helped by the allocation of resources, aber die
same overall pool of resources was used, this would aver-
age out and result in no overall benefit or cost.
Our data can also be used to test this account directly: A
differential attention account predicts that, as more faces
are present and/or perceived, performance on trials where
nonfaces are probed will drop by a large amount, as people
preferentially encode the faces. We do not see any evi-
dence for this in our data. Sorting by objective number
of faces, in Experiment 1, we find performance on nonface
trials is 79% with zero face present, 77% with one face pres-
ent, Und 78% with two faces present. In Experiment 2, pro-
formance on nonface trials is 78% with zero face present,
77% with one present, Und 76% with two faces present.
Fitting a mixed-effect model that accounts for subject
and item effects to only data from trials where a nonface
is probed also reveals no effect of the number of objective
or subjective faces on performance and no interaction (alle
p > .10 in both experiments). Daher, we see no evidence
that our effects are driven by differential attention to
faces at the cost of nonfaces, within the same pool of
resources—instead, our data show that participants sim-
ply remember face stimuli better than nonface stimuli re-
gardless of how many of each are present, consistent with
our interpretation that perceiving a stimulus as a face
allows the use of face-specific features and, entsprechen-
ingly, the recruitment of additional working capacity.
Knowledge and Visual Working Memory
The observed benefits for working memory performance
from perceived faces and for remembering real-world
Objekte (Brady & Störmer, 2020A, 2020B; Brady et al.,
2016) suggest that meaningful processing is the critical
attribute that gives rise to enhanced active storage, ähnlich
to the role of meaning in long-term memory (Konkle,
Brady, Alvarez, & Oliva, 2010; McWeeny, Jung, Hay, &
Ellis, 1987; Bower, Karlin, & Dueck, 1975). What deter-
mines the meaningfulness of an object? The present
findings show that when a visual stimulus is associated
with a high-level object category (d.h., a face), this can be
sufficient to increase working memory performance. Das
suggests that high-level visual templates such as knowing
what a face looks like can provide a link to connect an
incoming stimulus to existing knowledge and familiar
categories. In this case, meaningfulness may mostly
depend on high-level visually derived categories, aber in
other cases, Zum Beispiel, real-world objects, it seems plau-
sible that other, even richer conceptual associations are
being activated to induce meaningfulness. Such percep-
tion may allow the engagement of more high-level brain
Regionen, allowing people to maintain neural activity over
the delay not only in low-level sensory areas but also in
high-level regions relevant to the processing of the stimu-
lus, which may be higher-level visual regions, or other
brain regions activated by semantic knowledge. Für
Beispiel, fMRI studies have shown that when participants
remember faces, the fusiform face area is activated
(Salmela, Ölander, Muukkonen, & Bays, 2019; Druzgal &
D'Esposito, 2001), and when participants remember
images of hands, somatosensory regions are recruited
(Galvez-Pol, Calvo-Merino, Capilla, & Forster, 2018).
Another recent study showed that meaningful rather than
perceptually matched but nonmeaningful stimuli results
in greater activity in the ventral stream during visual
working memory tasks (Stojanoski, Emrich, & Cusack,
2019), again consistent with the idea that stimuli that con-
nect to knowledge are represented in other, zusätzlich
higher-level visual areas during working memory tasks.
Daher, it is possible that, in some sense, the improved
capacity we observe arises even during perception (oder
encoding) of the stimuli: Recruitment of higher-level
regions while viewing the stimuli provides the foundation
for these regions to play a role in working memory.
Consistent with this, we recently showed that how stim-
uli are processed during encoding is critical in determining
whether benefits for meaningful and real-world objects
arise (Brady & Störmer, 2020A). In a series of experiments,
we demonstrate that sequential encoding of real-world
objects—where each stimulus is shown one at a time, daher
allowing focused and item-based encoding—results in
larger object benefits relative to simultaneous encoding
of all stimuli at once, as often done in working memory
Studien (and as done here). We believe that sequential
encoding is one particularly effective way to ensure that
participants recognize an individual item’s identity, can
connect it to existing knowledge, and therefore recruit
additional neural resources to hold that item in memory.
Another way such deep encoding of stimuli can be
achieved is by using long encoding times, wie wir es getan haben
here and in other studies (Brady & Störmer, 2020B; Brady
et al., 2016); Jedoch, it seems that long encoding times
are not always sufficient in eliciting a benefit for meaning-
ful objects (Li, Xiong, Theeuwes, & Wang, 2020; Quirk,
Adam, & Vogel, 2020; but see Brady & Störmer, 2020B,
for an alternative explanation for these differences),
suggesting that variation in how participants encode a par-
ticular memory display is an important consideration
when studying memory for meaningful stimuli. In diesem
Studie, Zum Beispiel, participants were encouraged to try
to connect the memory stimuli to knowledge—that is,
try to see them as faces, as we asked them after every trial
914
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 5
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
how many faces they perceived. Daher, it seems critical that
items are processed in depth during encoding in order to
achieve the benefits promoted by meaningfulness.
Whether—and to what extent—the increases in capac-
ity we observe happen during encoding and persist across
the delay, or occur only from reduced noise accumulation
during storage, is an interesting question for future re-
suchen. We believe that either interpretation is very much
in line with our main conclusion—namely, that working
memory capacity is not fixed but adaptable to the type
of stimuli being remembered. In der Tat, it is plausible that
these two possibilities are fundamentally the same, Wenn, als
many propose, visual working memory is really “sustained
perception,” with attention-like mechanisms continuing
to keep the perceptual activity going over a delay (z.B.,
Chun, 2011). Under such an account, the improved per-
sistence of perceived faces over the delay is exactly the same
thing as improved encoding of them into working memory,
and improved encoding is likely necessary to allow the
greater maintenance activity we observe. Consistent with
Das, an exploratory analysis of Experiment 3 suggests that
the CDA benefits here arise even while the stimuli are on
the screen: Excluding the initial perceptual processing
window (300 ms), we find a reliable amplitude differ-
ence between one versus three objective faces during
the time the stimuli are on the screen, T(18) = 2.96, p =
.0084. This is consistent with other work finding CDA-
like e_k; activity is present even while stimuli are on the
screen, if they must be held actively in mind during this
Zeit (Tsubomi, Fukuda, Watanabe, & Vogel, 2013; Drew,
Horowitz, Wolfe, & Vogel, 2012; Emrich, Al-Aidroos, Pratt,
& Ferber, 2009).
Many studies have shown that existing knowledge or
familiarity with a stimulus improves the ability to maintain
information not only in long-term memory but also over
short delays, often termed long-term working memory
(Ericsson & Kintsch, 1995). Jedoch, it has been unclear
the extent to which such benefits arise from changes in
active storage in working memory per se, or from the
usage of “long-term” memory systems or other forms of
more passive storage that can be utilized in the short-term
maintenance of information. Both neurally and cognitively,
the active component of working memory (sometimes re-
ferred to as the “focus of attention”) is particularly impor-
tant as it seems to be the element of working memory
most responsible for capacity limits and correlations with
broader cognitive abilities (z.B., Cowan, 2005). Durch con-
trast, other forms of memory sometimes called working
memory but which are nonactive—like activated long-
term memories or long-term working memory—are
widely acknowledged to be influenced by knowledge
and meaningfulness (Ericsson & Kintsch, 1995), but are
often considered not to be core elements of working
memory capacity (Feldmann-Wüstefeld, Vogel, & Awh,
2018). We here show that this active component of
Arbeitsgedächtnis (as indexed by the CDA) is strongly
affected by the meaningfulness of the stimulus being
remembered. If a stimulus connects to a concept and is
thus perceived as meaningful, the active storage compo-
nent of memory capacity is greater.
Whereas a large body of work has shown benefits for ob-
jects of expertise in short-term storage (z.B., Curby et al.,
2009), one line of recent work has claimed that expertise
effects in visual working memory may sometimes arise
from enhanced consolidation of items into memory rather
than increased capacity per se (Xie & Zhang, 2017B, 2018),
although capacity increases have also been found (Xie &
Zhang, 2017A). Wichtig, in the current work with our
within-subject, perceptual manipulation of meaningful
recognition and concurrent electrophysiological recordings,
we show that active storage in working memory itself is
impacted by meaning, with no change over the time course
of the CDA component as suggested by a consolidation-
based account. We also use an encoding time that is long
enough to avoid encoding limits (1000 ms), even accord-
ing to the work arguing for consolidation limits (z.B., Xie &
Zhang, 2018). The kind of meaningfulness Xie and Zhang
manipulate is quite different than in the current work, als
unlike the perception of a stimulus as a face-enhanced
memory in their experiments likely requires retrieval of
specific information from long-term memory.
How might conceptual knowledge or other crystallized
long-term memories enhance active storage in working
Erinnerung? One possibility is that once a stimulus is per-
ceived as meaningful (z.B., as a face), more useful informa-
tion is being extracted and stored relative to when a
stimulus is perceived as meaningless. In that case, the abil-
ity to maintain this information in working memory flexi-
bly adjusts depending on the stimulus input—possibly
allowing a broader set of features to be remembered and
actively maintained for holistic objects relative to arbitrary
shapes. This would also limit interference between the
neural populations that must be held active (z.B., Cohen,
Konkle, Rhee, Nakayama, & Alvarez, 2014). Weil
having more crystallized knowledge about an object or
object category results in more relevant neural popula-
tions being activated, attentional mechanisms can main-
tain more active information successfully when dealing
with meaningful objects ( Wyble et al., 2016). Daher, knowl-
edge may serve as a “hook” that helps recruit the critical
neural populations that are both more relevant and more
distinct, thereby facilitating active maintenance of mean-
ingful information.
Reprint requests should be sent to Isabel Asp or Timothy Brady,
Abteilung für Psychologie, Universität von Kalifornien, San Diego,
9500 Gilman Dr. #0109, La Jolla, CA 92093, or via email: iasp
@ucsd.edu; timbrady@ucsd.edu.
Autorenbeiträge
Isabel E. Asp: Konzeptualisierung; Datenkuration; Formal
Analyse; Untersuchung; Methodik; Schreiben – Original
Entwurf. Viola S. Störmer: Konzeptualisierung; Formale Analyse;
Asp, Störmer, and Brady
915
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Akquise von Fördermitteln; Untersuchung; Methodik; Aufsicht;
Validierung; Schreiben – Rezension & Bearbeitung. Timothy F. Brady:
Konzeptualisierung; Datenkuration; Formale Analyse; Funding
Erwerb; Untersuchung; Methodik; Project administra-
tion; Software; Aufsicht; Validierung; Visualisierung;
Schreiben – Originalentwurf; Schreiben – Rezension & Bearbeitung.
Informationen zur Finanzierung
Timothy F. Brady and Viola S. Störmer, Division of
Behavioral and Cognitive Sciences (dx.doi.org/10.13039
/100000169), grant number: BCS-1829434.
Vielfalt in der Zitierpraxis
A retrospective analysis of the citations in every article pub-
in dieser Zeitschrift aufgeführt von 2010 Zu 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) Veröffentlichung
in the Journal of Cognitive Neuroscience ( JoCN) während
this period were M(ein)/M = .408, W(Oman)/M = .335,
M/W = .108, and W/W = .149, the comparable propor-
tions for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, S. 3–7). Folglich, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
VERWEISE
Alvarez, G. A., & Cavanagh, P. (2004). The capacity of visual
short-term memory is set both by visual information load and
by number of objects. Psychological Science, 15, 106–111.
DOI: https://doi.org/10.1111/j.0963-7214.2004.01502006.X,
PMID: 14738517
Awh, E., Barton, B., & Vogel, E. K. (2007). Visual working
memory represents a fixed number of items regardless of
complexity. Psychological Science, 18, 622–628. DOI:
https://doi.org/10.1111/j.1467-9280.2007.01949.x, PMID:
17614871
Bagiella, E., Sloan, R. P., & Heitjan, D. F. (2000). Mixed-effects
models in psychophysiology. Psychophysiology, 37, 13–20.
DOI: https://doi.org/10.1111/1469-8986.3710013, PMID:
10705763
Bartlett, F. C. (1932). Remembering: An experimental and
social study. Cambridge: Cambridge University Press.
Bays, P. M. (2015). Spikes not slots: Noise in neural populations
limits working memory. Trends in den Kognitionswissenschaften, 19,
431–438. DOI: https://doi.org/10.1016/j.tics.2015.06.004,
PMID: 26160026
Bays, P. M., Catalao, R. F., & Husain, M. (2009). The precision of
visual working memory is set by allocation of a shared
resource. Journal of Vision, 9, 7. DOI: https://doi.org/10
.1167/9.10.7, PMID: 19810788, PMCID: PMC3118422
Bliss, C. ICH. (1967). Statistics in biology: Statistical methods for
research in the natural sciences. New York: McGraw-Hill.
DOI: https://doi.org/10.1002/bimj.19690110327
Bower, G. H., Karlin, M. B., & Dueck, A. (1975).
Comprehension and memory for pictures. Memory &
Cognition, 3, 216–220. DOI: https://doi.org/10.3758
/BF03212900, PMID: 21287062
Brady, T. F., Alvarez, G. A., & Störmer, V. S. (2019). Die Rolle
of meaning in visual memory: Face-selective brain activity
predicts memory for ambiguous face stimuli. Zeitschrift für
Neurowissenschaften, 39, 1100–1108. DOI: https://doi.org/10.1523
/JNEUROSCI.1693-18.2018, PMID: 30541914, PMCID:
PMC6363929
Brady, T. F., Konkle, T., & Alvarez, G. A. (2009). Compression
in visual working memory: Using statistical regularities to
form more efficient memory representations. Zeitschrift für
Experimental Psychology: Allgemein, 138, 487–502. DOI:
https://doi.org/10.1037/a0016797, PMID: 19883132
Brady, T. F., & Störmer, V. S (2020A). The role of meaning in
visual working memory: Real-world objects, but not simple
Merkmale, benefit from deeper processing. PsyArxiv. DOI:
https://doi.org/10.31234/osf.io/kzvdg
Brady, T. F., & Störmer, V. S (2020B). Comparing memory
capacity across stimuli requires maximally dissimilar foils:
Using deep convolutional neural networks to understand
visual working memory capacity for real-world objects.
PsyArxiv. DOI: https://doi.org/10.31234/osf.io/25t76
Brady, T. F., Störmer, V. S., & Alvarez, G. A. (2016). Arbeiten
memory is not fixed-capacity: More active storage capacity for
real-world objects than for simple stimuli. Verfahren der
Nationale Akademie der Wissenschaften, USA., 113, 7459–7464. DOI:
https://doi.org/10.1073/pnas.1520027113, PMID: 27325767,
PMCID: PMC4941470
Buschman, T. J., Siegel, M., Roy, J. E., & Müller, E. K. (2011).
Neural substrates of cognitive capacity limitations. Verfahren
der Nationalen Akademie der Wissenschaften, USA., 108, 11252–11255.
DOI: https://doi.org/10.1073/pnas.1104666108, PMID:
21690375, PMCID: PMC3131328
Carlisle, N. B., Arita, J. T., Pardo, D., & Woodman, G. F. (2011).
Attentional templates in visual working memory. Zeitschrift für
Neurowissenschaften, 31, 9315–9322. DOI: https://doi.org/10.1523
/JNEUROSCI.1097-11.2011, PMID: 21697381, PMCID:
PMC3147306
Chun, M. M. (2011). Visual working memory as visual attention
sustained internally over time. Neuropsychologie, 49,
1407–1409. DOI: https://doi.org/10.1016/j.neuropsychologia
.2011.01.029, PMID: 21295047
Cohen, M. A., Konkle, T., Rhee, J. Y., Nakayama, K., & Alvarez,
G. A. (2014). Processing multiple visual objects is limited by
overlap in neural channels. Verfahren des Nationalen
Akademie der Wissenschaften, USA., 111, 8955–8960. DOI: https://
doi.org/10.1073/pnas.1317860111, PMID: 24889618, PMCID:
PMC4066506
Cowan, N. (2005). Working memory capacity. New York:
Psychology Press. DOI: https://doi.org/10.4324/9780203342398
Curby, K. M., Glazek, K., & Gauthier, ICH. (2009). A visual short-
term memory advantage for objects of expertise. Zeitschrift
of Experimental Psychology: Human Perception and
Performance, 35, 94–107. DOI: https://doi.org/10.1037/0096
-1523.35.1.94, PMID: 19170473, PMCID: PMC4159943
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Zeitschrift für Neurowissenschaften
Methoden, 134, 9–21. DOI: https://doi.org/10.1016/j.jneumeth
.2003.10.009, PMID: 15102499
Drew, T., Horowitz, T. S., Wolfe, J. M., & Vogel, E. K. (2012).
Neural measures of dynamic changes in attentive tracking
load. Zeitschrift für kognitive Neurowissenschaften, 24, 440–450. DOI:
https://doi.org/10.1162/jocn_a_00107, PMID: 21812558
Druzgal, T. J., & D'Esposito, M. (2001). Activity in fusiform face
area modulated as a function of working memory load.
Kognitive Gehirnforschung, 10, 355–364. DOI: https://doi.org
/10.1016/S0926-6410(00)00056-2, PMID: 11167061
916
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 5
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Emrich, S. M., Al-Aidroos, N., Pratt, J., & Ferber, S. (2009). Visual
search elicits the electrophysiological marker of visual
Arbeitsgedächtnis. PLoS One, 4, e8042. DOI: https://doi
.org/10.1371/journal.pone.0008042, PMID: 19956663,
PMCID: PMC2777337
Ericsson, K. A., & Kintsch, W. (1995). Long-term working
Erinnerung. Psychological Review, 102, 211–245. DOI: https://
doi.org/10.1037/0033-295X.102.2.211, PMID: 7740089
Feldmann-Wüstefeld, T., Vogel, E. K., & Awh, E. (2018).
Contralateral delay activity reflects working memory storage,
not the current focus of spatial attention within visual
Arbeitsgedächtnis. Zeitschrift für kognitive Neurowissenschaften, 30,
1185–1196. DOI: https://doi.org/10.1162/jocn_a_01271,
PMID: 29694260, PMCID: PMC6283407
Fougnie, D., Cormiea, S. M., Alvarez, G. A. (2013). Object
benefits without object-based representations. Zeitschrift für
Experimental Psychology: Allgemein, 142, 621–626. DOI:
https://doi.org/10.1037/a0030300, PMID: 23067063
Galvez-Pol, A., Calvo-Merino, B., Capilla, A., & Forster, B.
(2018). Persistent recruitment of somatosensory cortex
during active maintenance of hand images in working
Erinnerung. Neurobild, 174, 153–163, DOI: https://doi.org
/10.1016/j.neuroimage.2018.03.024, PMID: 29548846.
Huang, L., & Awh, E. (2018). Chunking in working memory via
content-free labels. Wissenschaftliche Berichte, 8, 23. DOI: https://doi
.org/10.1038/s41598-017-18157-5, PMID: 29311568, PMCID:
PMC5758528
Ikkai, A., McCollough, A. W., & Vogel, E. K. (2010). Contralateral
delay activity provides a neural measure of the number of
representations in visual working memory. Zeitschrift für
Neurophysiologie, 103, 1963–1968. DOI: https://doi.org/10
.1152/jn.00978.2009, PMID: 20147415, PMCID: PMC2853266
Jackson, M. C., & Raymond, J. E. (2008). Familiarity enhances
visual working memory for faces. Journal of Experimental
Psychologie: Human Perception and Performance, 34,
556–568. DOI: https://doi.org/10.1037/0096-1523.34.3.556,
PMID: 18505323, PMCID: PMC4262787
Konkle, T., Brady, T. F., Alvarez, G. A., & Oliva, A. (2010).
Conceptual distinctiveness supports detailed visual long-term
memory for real-world objects. Journal of Experimental
Psychologie: Allgemein, 139, 558–578. DOI: https://doi.org
/10.1037/a0019165, PMID: 20677899, PMCID: PMC3398125
Li, X., Xiong, Z., Theeuwes, J., & Wang, B. (2020). Visual
memory benefits from prolonged encoding time regardless
of stimulus type. Journal of Experimental psychology.
Learning, Memory, and Cognition, 46, 1998–2005. DOI:
https://doi.org/10.1037/xlm0000847, PMID: 32437186
Lopez-Calderon, J., & Luck, S. J. (2014). ERPLAB: An open-
source toolbox for the analysis of event-related potentials.
Grenzen der menschlichen Neurowissenschaften, 8, 213. DOI: https://
doi.org/10.3389/fnhum.2014.00213, PMID: 24782741,
PMCID: PMC3995046
Luck, S. J. (2008). Visual short-term memory. In Visual memory
(S. 43–85). Oxford University Press. DOI: https://doi.org
/10.1093/acprof:oso/9780195305487.003.0003
Luck, S. J., & Vogel, E. K. (2013). Visual working memory
Kapazität: From psychophysics and neurobiology to individual
differences. Trends in den Kognitionswissenschaften, 17, 391–400. DOI:
https://doi.org/10.1016/j.tics.2013.06.006, PMID: 23850263,
PMCID: PMC3729738
Luria, R., Balaban, H., Awh, E., & Vogel, E. K. (2016). Der
contralateral delay activity as a neural measure of visual
Arbeitsgedächtnis. Neuroscience and Biobehavioral Reviews,
62, 100–108. DOI: https://doi.org/10.1016/j.neubiorev.2016
.01.003, PMID: 26802451, PMCID: PMC4869985
Luria, R., Sessa, P., Gotler, A., Jolicœur, P., & Dell’Acqua, R.
(2010). Visual short-term memory capacity for simple and
complex objects. Zeitschrift für kognitive Neurowissenschaften, 22,
496–512. DOI: https://doi.org/10.1162/jocn.2009.21214,
PMID: 19301998
Ma, W. J., Husain, M., & Bays, P. M. (2014). Changing concepts
of working memory. Naturneurowissenschaften, 17, 347–356.
DOI: https://doi.org/10.1038/nn.3655, PMID: 24569831,
PMCID: PMC4159388
McCollough, A. W., Machizawa, M. G., & Vogel, E. K. (2007).
Electrophysiological measures of maintaining representations
in visual working memory. Kortex, 43, 77–94. DOI: https://
doi.org/10.1016/S0010-9452(08)70447-7
McWeeny, K. H., Jung, A. W., Hay, D. C., & Ellis, A. W. (1987).
Putting names to faces. British Journal of Psychology, 78, 143–149.
DOI: https://doi.org/10.1111/j.2044-8295.1987.tb02235.x
Mooney, C. M. (1957). Age in the development of closure ability
in children. Canadian Journal of Psychology/Revue
Canadienne de Psychologie, 11, 219–226. DOI: https://doi
.org/10.1037/h0083717, PMID: 13489559
Ngiam, W. X., Khaw, K. L., Holcombe, A. O., & Goodbourn, P. T.
(2018). Visual working memory for letters varies with
familiarity but not complexity. Journal of Experimental
Psychologie: Learning, Memory, and Cognition, 45,
1761–1775. DOI: https://doi.org/10.31234/osf.io/cxkw5
Quirk, C., Adam, K. C. S., & Vogel, E. K. (2020). No evidence for
an object working memory capacity benefit with extended
viewing time. eNeuro, 7, ENEURO.0150-20.2020. DOI:
https://doi.org/10.1523/ENEURO.0150-20.2020, PMID:
32859722, PMCID: PMC7519167
Robinson, M. M., Benjamin, A. S., & Irwin, D. E. (2020). Is there
a K in capacity? Assessing the structure of visual short-term
Erinnerung. Cognitive Psychology, 121, 101305, DOI: https://
doi.org/10.1016/j.cogpsych.2020.101305, PMID: 32531272
Salmela, V. R., Ölander, K., Muukkonen, ICH., & Bays, P. M. (2019).
Recall of facial expressions and simple orientations reveals
competition for resources at multiple levels of the visual
hierarchy. Journal of Vision, 19, 8. DOI: https://doi.org
/10.1167/19.3.8, PMID: 30897626, PMCID: PMC6432740
Schurgin, M. W., Cunningham, C. A., Egeth, H. E., & Brady, T. F.
(2018). Visual long-term memory can replace active storage in
visual working memory. bioRxiv preprint. DOI: https://doi
.org/10.1101/381848
Schurgin, M. W., Wixted, J. T., & Brady, T. F. (2020). Psychophysical
scaling reveals a unified theory of visual memory strength.
Natur menschliches Verhalten, 4, 1156–1172. DOI: https://doi
.org/10.1038/s41562-020-00938-0, PMID: 32895546
Schwiedrzik, C. M., Melloni, L., & Schurger, A. (2018). Mooney
face stimuli for visual perception research. PLoS One, 13,
e0200106. DOI: https://doi.org/10.1371/journal.pone.0200106,
PMID: 29979727, PMCID: PMC6034866
Serences, J. T. (2016). Neural mechanisms of information
storage in visual short-term memory. Sehforschung, 128,
53–67. DOI: https://doi.org/10.1016/j.visres.2016.09.010,
PMID: 27668990, PMCID: PMC5079778
Stojanoski, B., Emrich, S. M., & Cusack, R. (2019). Representation
of semantic information in ventral areas during encoding is
associated with improved visual short-term memory. bioRxiv.
DOI: https://doi.org/10.1101/2019.12.13.875542
Störmer, V. S., Li, SC., Heekeren, H. R., & Lindenberger, U.
(2013). Normative shifts of cortical mechanisms of encoding
contribute to adult age differences in visual-spatial working
Erinnerung. Neurobild, 73, 167–175. DOI: https://doi.org
/10.1016/j.neuroimage.2013.02.004, PMID: 23415947
Tsubomi, H., Fukuda, K., Watanabe, K., & Vogel, E. K. (2013).
Neural limits to representing objects still within view. Zeitschrift
der Neurowissenschaften, 33, 8257–8263. DOI: https://doi.org/10
.1523/JNEUROSCI.5348-12.2013, PMID: 23658165, PMCID:
PMC4049283
Van den Berg, R., Schienbein, H., Chou, W. C., George, R., & Ma, W. J.
(2012). Variability in encoding precision accounts for visual
Asp, Störmer, and Brady
917
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
short-term memory limitations. Verfahren des Nationalen
Akademie der Wissenschaften, USA., 109, 8780–8785. DOI: https://
doi.org/10.1073/pnas.1117465109, PMID: 22582168, PMCID:
PMC3365149
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity predicts
individual differences in visual working memory capacity.
Natur, 428, 748–751. DOI: https://doi.org/10.1038
/nature02447, PMID: 15085132
Vogel, E. K., McCollough, A. W., & Machizawa, M. G. (2005).
Neural measures reveal individual differences in controlling
access to working memory. Natur, 438, 500–503. DOI:
https://doi.org/10.1038/nature04171, PMID: 16306992
Williams, M., & Woodman, G. F. (2012). Directed forgetting and
directed remembering in visual working memory. Zeitschrift für
Experimental Psychology: Learning, Memory, and Cognition,
38, 1206–1220. DOI: https://doi.org/10.1037/a0027389, PMID:
22409182, PMCID: PMC3817833
Wiseman, S., & Neisser, U. (1974). Perceptual Organization as
a determinant of visual recognition memory. amerikanisch
Journal of Psychology, 87, 675–681. DOI: https://doi.org
/10.2307/1421974, PMID: 4463729
Wolfe, J. M., & Horowitz, T. S. (2017). Five factors that guide
attention in visual search. Natur menschliches Verhalten, 1,
0058. DOI: https://doi.org/10.1038/s41562-017-0058
Wyble, B., Swan, G., & Callahan-Flintoft, C. (2016). Measuring
visual memory in its native format. Trends im kognitiven Bereich
Wissenschaften, 20, 790–791. DOI: https://doi.org/10.1016/j.tics
.2016.08.012, PMID: 27623428
Xie, W., & Zhang, W. (2018). Familiarity speeds up visual short-
term memory consolidation: Electrophysiological evidence
from contralateral delay activities. Zeitschrift für Kognition
Neurowissenschaften, 30, 1–13. DOI: https://doi.org/10.1162/jocn
_a_01188, PMID: 28891784
Xie, W., & Zhang, W. (2017A). Familiarity increases the number
of retained Pokémon in visual short-term memory. Memory
& Cognition. 45, 677–689. DOI: https://doi.org/10.3758
/s13421-016-0679-7, PMID: 27933560
Xie, W., & Zhang, W. (2017B). Familiarity speeds up visual
short-term memory consolidation. Journal of Experimental
Psychologie: Human Perception and Performance, 43,
1207–1221. DOI: https://doi.org/10.1037/xhp0000355,
PMID: 28287761
Zimmer, H. D., & Fischer, B. (2020). Visual working memory
of Chinese characters and expertise: the expert’s memory
advantage is based on long-term knowledge of visual word
Formen. Grenzen in der Psychologie, 11, 516. DOI: https://doi
.org/10.3389/fpsyg.2020.00516, PMID: 32362852, PMCID:
PMC7180225
Zimmer, H. D., Popp, C., Reith, W., & Krick, C. (2012). Gains of
item-specific training in visual working memory and their
neural correlates. Gehirnforschung, 1466, 44–55. DOI: https://
doi.org/10.1016/j.brainres.2012.05.019, PMID: 22659025
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
/
J
/
Ö
C
N
A
R
T
ich
C
e
–
P
D
l
F
/
/
/
/
3
3
5
9
0
2
1
9
0
0
8
6
4
/
J
Ö
C
N
_
A
_
0
1
6
9
3
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
918
Zeitschrift für kognitive Neurowissenschaften
Volumen 33, Nummer 5