Evolvable Physical Self-Replicators
Abstract Building an evolvable physical self-replicating machine
is a grand challenge. The main problem is that the device must
be capable of hereditary variation, das ist, replicating in many
configurations—configurations into which it enters unpredictably
by mutation. Template replication is the solution found by nature.
A scalable device must also be capable of miniaturization, and so
have few or no moving and electronic parts. Here a significant step
toward this goal is presented in the form of a physical template
replicator made from small plastic pieces containing embedded
magnets that float on an air-hockey-type table and undergo stochastic
Bewegung. Our units replicate by a process analogous to the replication
of DNA, except without the involvement of enzymes. Building a
physical rather than a computational model forces us to confront
several problems that have analogues on the nano scale. Insbesondere,
replication must be maintained by preventing side reactions such as
spontaneous ligation, cyclization, product inhibition, and elongation
at staggered ends. The last of these results in ever-lengthening
sequences in a process known as the elongation catastrophe. Der
extreme specificity of structure required by the monomers is
indirect evidence that some kind of natural selection took place
prior to the existence of nucleotide analogues during the origin of life.
Nathaniel Virgo**,†
University of Sussex
Max Planck Institute for
Biogeochemistry
Chrisantha Fernando*,†,‡,§
University of Sussex
Queen Mary, University of London
MRC National Institute for Medical
Forschung
Bill Bigge†
University of Sussex
†
Phil Husbands
University of Sussex
Schlüsselwörter
Physical self-replicator, Vorlage
replication, elongation catastrophe, origin
des Lebens
A version of this paper with color figures is
available online at http://dx.doi.org/10.1162/
artl_a_00056. Subscription required.
1 Einführung
In todayʼs highly evolved biology, a complex array of enzymes is necessary for the replication of DNA
and RNA polymers. These enzymes function as complex molecular machines and are themselves the
results of evolution. Such complex enzymes would not have been available at the origin of life, Und
so an important question is how the replication of molecular sequences can have occurred without
ihnen. In this article we investigate the possibility of non-enzymatic template replication [11, 12]. Das ist ein
mechanism by which information-carrying polymer strands could have replicated without the need
for enzymes. The proposed mechanism is similar to the replication of modern nucleotide sequences in
that it is based on the pairing of complementary strands; the difference is that in non-enzymatic template
replication, free monomers form into a complementary strand spontaneously rather than being guided
into place by enzymes.
* Contact author.
** Now at the Max Planck Institute for Biogeochemistry, 07745 Jena, Deutschland.
† Department of Informatics, University of Sussex, Falmer, Brighton BN1 9RH, Vereinigtes Königreich. Email: ctf20@sussex.ac.uk (C.F.).
‡ School of Electronic Engineering and Computer Science, Queen Mary, University of London, Mile End Road, London E1 4NS, Vereinigtes Königreich.
§ MRC National Institute for Medical Research, London NW7 1AA, Vereinigtes Königreich.
© 2012 Massachusetts Institute of Technology
Artificial Life 18: 129–142 (2012)
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
N. Virgo et al.
Evolvable Physical Self-Replicators
Figur 1. A generic illustration of template replication and two side reactions that must be avoided. (A) Homologous
template directed ligation (self-replication) results in the correct duplicaton of a sequence. (B) Ein neues (incorrect) se-
quence is formed by non-templated spontaneous ligation. (C) Elongation of the original sequence by partially homologous
template ligation at staggered ends. Sehen [4] for a full analysis of the elongation catastrophe.
Non-enzymatic template replication has been demonstrated in chemical systems (z.B., [13]). Wie-
immer, it is not straightforward to achieve, because there are a number of side reactions whose rates must
be made low enough to allow sufficient autocatalytic flux that one cycle of replication produces more
units than the cycle started with. Mit anderen Worten, the rate of decay of the constituents of the auto-
catalytic cycle must not exceed the rate of growth of the autocatalytic cycle. Possible side reactions are
shown diagrammatically in Figure 1. Our work consists of a macroscopic version of non-enzymatic
template replication constructed out of inert components that float around at random on an air-hockey
table. In constructing it, we had to solve many of the same problems that arise in the chemical domain.
The complexity of the resulting “molecules” suggests that non-enzymatic template replication might not
be easy enough to achieve for it to have arisen spontaneously, before the origins of natural selection. Der
probability of assembling such specific molecules by chance seems as low as the probability of the ran-
dom assembly of Paleyʼs watch, and the assembly of two or more of these molecules seems even more
unimaginable. Whereas we can respond to Paley by citing Darwinian evolution, what kind of evolution
could have taken place prior to the existence of nucleotides?
Our work is an improvement on the first chemical self-replicating system to be constructed,
which took the form of an end-blocked hexanucleotide sequence GGCGCC that catalyzes the tem-
plated ligation of CGG and CCG trimers [13]. Such a system does not contain units of evolution,
because there is no hereditary variation, and hence it transmits zero bits of information from gen-
eration to generation. This work also improves on the constructions of Lional Penrose and Roger
Penrose in being 2D rather than 1D [8–10]. It is also an improvement on previously constructed
macroscopic “self-replicating” systems. Zum Beispiel, the 2D macroscopic system designed by Jarle
Breivik succumbed to both the problem of spontaneous ligation and the elongation catastrophe [1].
The first templates formed by spontaneous (non-templated) ligation were replicated much more
slowly than the spontaneous formation of novel sequences from monomers. Außerdem, diese
130
Artificial Life Volume 18, Nummer 2
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
N. Virgo et al.
Evolvable Physical Self-Replicators
templates continued to grow and ultimately fell victim to the elongation catastrophe, in which
sequences become trapped in longer and longer strands, eventually being unable to replicate [4]. Das
is an anterior problem to the problem of Eigenʼs error catastrophe, which requires that mutations
must occur at a low rate for the maintenance of a long sequence [2]. Without reducing side reactions,
the problem of mutation does not even arise, for there is no replicator to be mutated. A minimal
evolvable system must therefore exhibit the replication of dimers with a low rate of assembly of
incorrect or elongated sequences. At least, the average rate of replication of a seed dimer should be
greater than the rate of formation of all other sequences put together. The mechanical 2D stochastic
self-replicator presented here has limited rates of non-catalyzed spontaneous self-assembly (ligation)
of monomers, limited partial homologous templated ligation (elongation at staggered double strand
ends), limited cyclization (formation of cyclic complexes), and limited product inhibition (double
strands getting stuck together).
Nächste, we describe the design of the monomers and show the results of a seeding experiment to
distinguish self-replication from self-assembly. This is the only demonstration so far of an evolvable
2D template replication system that is capable of low rates of spontaneous elongation yet high rates
of self-replication (without the use of monomers containing complex electronically implemented finite
state machines) [5, 14].
2 Methoden
We constructed a macroscopic self-replicating object out of inert and unpowered components that
float at random on an air-hockey table; siehe Abbildung 2. They operate by (approximate) Brownian
Bewegung; the properties of each are determined by the shape and electromagnetic properties of the
“molecules”; and they approximately obey mass-action kinetics. The system is capable of multiplica-
tion, Variation, and heredity and therefore can be considered to contain units of evolution as defined
by John Maynard Smith [7]. The minimal evolvable self-replicating system shown here consists of
dimers (polymers of length two) that can replicate three possible sequences, AB, BA, or AA(BB), als
in Abbildung 1.
Figur 2. The design of the air-hockey-style table containing the monomers. Sails on each monomer are blown by a
perimeter of small fans. Another fan below the table passes air through small holes to suspend monomers over the
table like small hovercraft.
Artificial Life Volume 18, Nummer 2
131
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
N. Virgo et al.
Evolvable Physical Self-Replicators
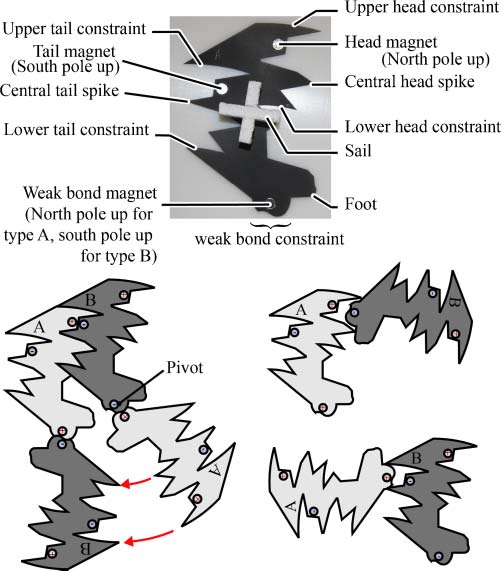
Figur 3. The design of the monomers. The top photograph shows the names used in the text for important parts of the
Design. In the photograph, the two monomers are distinguished by the color of their polystyrene sails (white for A, black for
B), whereas in diagrams, type B is shown in a darker shade of gray. The lower left diagram shows the mechanism by which
templated ligation takes place (but see also Figure 4). The lower right diagram shows how the design prevents the weak bond
magnets from bonding to the strong bond magnets.
A frictionless table, similar to those used for air hockey, was purpose built and consisted of a flat
plastic surface perforated with an array of 1.5-mm-diameter holes, spaced at intervals of 10 mm. Ein
enclosure underneath this surface was pressurized with a powerful fan to produce a steady jet of air
from each hole, allowing suitably shaped objects to float above the table surface. Surrounding the
table was a set of approximately 20 small fans that could be arranged to cause a stochastic motion of
the pucks, albeit with a significant rotational element; see Video A in the supplementary material.1
There was no temperature oscillation as in Breivikʼs experiment, das ist, the fans always rotated at the
same speed. The walls of the table allowed approximately elastic collisions. The puck design is shown
in Abbildung 3.
The pucks are 1.5 mm thick and made of plastic. The bases of the pucks are flat, allowing hovercraft-
type low-friction floating of the puck above the table. The pucks were fabricated using a Versalaser
cutter. Rapid fabrication of new designs was possible for prototyping. Pucks contain molybdenum disk
magnets that can be oriented with the north or south pole facing upwards, allowing specification of
attractive or repulsive interaction pairs.
The final design has the following features. The strong “phosphodiester bonds” (P) must not
form spontaneously. This is achieved by embedding the magnets deep within the puck and produc-
ing a lock-and-key-type join that can only form if the pucks collide at a very specific orientation. Das
orientation tends to occur only when the two monomers are “hydrogen bonded” (H) to a dimer tem-
plate, and not when two pucks collide against each other as untemplated monomers. Once the pucks
make the p-bond, the magnets are very close together, so the bond is strong. Thus the p-bond is
1 http://www.cogs.susx.ac.uk/users/ctf20/dphil_2005/Photos/Magnetic%20Chemistry.m4v
132
Artificial Life Volume 18, Nummer 2
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
N. Virgo et al.
Evolvable Physical Self-Replicators
difficult to form due to steric constraints, but once formed is strong due to close magnets and me-
chanical rigidity. The h-bonds consist of an interaction between magnets that are further apart when
the bond is formed, so that the bond is weaker. Auch, there is a curve on the surface of the bond to allow
pucks to rotate when h-bonded. This rotation brings the two h-bonded monomers into the appropriate
configuration for the p-bond to form.
To reduce product inhibition, the pucks are shaped in such a way that two p-bonded dimers
cannot be joined at both h-bonds, due to the presence of a protruding part that we call the foot (sehen
Figur 3). Daher, as the p-bond forms, it breaks one of the two h-bonds. The remaining h-bond is suffi-
ciently weak that the two dimers can separate and undergo another round of replication.
There are two types of monomer, labeled A and B, which differ only in the orientation of the mag-
nets that form their h-bonds. A-type monomers can only form weak (h-)bonds with B-type monomers,
und umgekehrt. Strong (p-)bonds can be formed between any pair of monomers, giving rise to four types
of strong-bonded dimer: AA, AB, BA, and BB. Template replication produces a new dimer that is both
the complement and the reverse of the original. This results in three separate autocatalytic cycles: {AB},
with the reaction AB + A + B → 2AB; {BA}, with the reaction BA + A + B → 2BA; Und {AA, BB},
with the reactions AA + 2B → AA + BB and BB + 2A → AA + BB (siehe Abbildung 4).
Misalignment with the generation of a staggered end (or dangling end, as they are often called) can
cause AA dimers to be extended via catalysis to AAA trimers, and similarly for the BB type, by partial
homologous ligation (siehe Abbildung 5). Jedoch, in all the experiments conducted we did not observe the
production of tetramers by partial homologous ligation. Wichtig, misalignment did not tend to oc-
cur for AB and BA dimers, which cannot catalyze partial homologous ligation-dependent elongations
unless another species of dimer is also present in the system. The explanation is given in Figure 5.
Zusammenfassend, there are three principles that we used to limit the elongation catastrophe in this
simple system: (1) impossibility of formation of non-complementary h-bonded pairs, (2) a high
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 4. The autocatalytic cycle for replication of an AB dimer. (A) A type-B monomer joins to the dimer. (B) A type-A
monomer joins to the other h-bond and swivels into place via the mechanism shown in Figure 1. Catalysis can also take place
if the monomers join in the opposite order; in this case both monomers must swivel on their weak bonds, which often occurs
when the configuration collides with another object. (C) A p-bond is formed by template-directed ligation, and simultaneously
one of the h-bonds is broken. A collision with another molecule or the table edge is required in order for this step to occur.
(D) Another collision breaks the remaining weak bond, and the two strands separate, completing the cycle.
Artificial Life Volume 18, Nummer 2
133
N. Virgo et al.
Evolvable Physical Self-Replicators
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 5. (A) The formation of a BBB trimer due to partial homologous ligation. The production of AAA and BBB trimers
in this way is fairly common in our system (siehe Abbildung 9). (B) Staggered bonding is not possible between two AB dimers
(or two BA dimers), because it would require the formation of an h-bond between magnets of the same polarity. (C) Es ist
in theory possible for a further partial homologous ligation to extend a BBB trimer into a BBBB tetramer. Jedoch, Wir
did not observe this in any of our trials. We suspect this is because the two polymers have a high moment of inertia
about the weak bondʼs pivot point, destabilizing the bond and making it likely to break. (D) Polymers of length greater
than two cannot replicate in the same way as dimers, because the foot mechanism does not allow the strong-bond
constraints to align with the weak-bond pivot.
moment of inertia at the pivot point of a staggered end, Und (3) improper alignment of p-bond
passive hooks during an attempted templated ligation for N-mers where N > 2. In combination,
these three factors significantly reduced the elongation catastrophe by limiting partial homologous
ligation. The curved passive hooks previously described also helped by reducing the extent of non-
catalyzed ligation.
To understand the system, it is useful to use a comparative method and consider the “phylogeny
of designs” leading to the current designs. It took approximately 30 iterations to produce the final
Design, some of which can be seen in Figure 6. The following problems had to be solved:
ich. The strong (p-)bonds must be unlikely to form spontaneously, das ist, we had to minimize
spontaneous generation.
134
Artificial Life Volume 18, Nummer 2
N. Virgo et al.
Evolvable Physical Self-Replicators
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 6. A selection of unsuccessful iterations of the design, illustrating the ways in which various problems were solved.
The designs are shown in chronological order. See text for details. In the electronic version, magnets are shown in red or
blue according as the north or south pole is oriented upwards. The weak bond magnets, whose orientation depends on
the polymer type, are shown in white.
Artificial Life Volume 18, Nummer 2
135
N. Virgo et al.
Evolvable Physical Self-Replicators
ii. There must not be any reactions that catalyze p-bond formation, other than the intended
template mechanism. Zum Beispiel, if two pairs of monomers joined by h-bonds come together,
they must not line up at the angle needed to form p-bonds.
iii. Once formed, the strong bonds must be strong enough that they rarely break. (In the final
design they were strong enough not to break at all.)
iv. The strong bonds must form easily when catalyzed by the weak bonds.
v. The weak (h-)bonds must form easily.
vi. The weak bonds must also break easily. This facilitates strand separation, as well as freeing
up monomers that have become weak-bonded to other monomers, which would otherwise
not be able to participate in catalysis.
vii. Once a dimer has catalyzed the creation of another, the two strands must be able to separate;
this is the problem of product inhibition.
viii. The magnet in the weak bond must not be able to attach strongly to the magnet in the
head or tail of another puck. Such unwanted bonds inhibit catalysis by occupying the bond
points, and can also give rise to configurations that can catalyze the wrong type of dimer.
ix. The puck must be able to float effectively on the table. Designs with long thin protruding parts,
or uneven weight distributions, can drag on the tableʼs surface.
X. The pucks must not tend to jump off the tableʼs surface and become stacked on top of
one another. This tends to happen if two magnets with the same polarity are forced close
to one another, or if the design features spikes that are too sharp.
Of these, problems i and ii were by far the hardest to solve. In most of our designs, einschließlich der
final one, the strong bond works by requiring the two pucks to collide at a very precise angle. In vielen
of the designs, if the collision occurred at a slightly different angle, a strong bond would often form
Trotzdem. This is because the head and tail magnets would tend to make the pucks slide into place to
form a strong bond, or else the two pucks would sit together in a configuration where they could
easily be nudged into the right position to form a strong bond. This was solved in the final design by
adding long spikes to the strong-bond constraints, in such a way that the magnets tend to pull the
pucks away from, rather than towards, the strong-bond configuration if the pucks are not correctly
lined up. Jedoch, the pucks do still occasionally collide at the angle needed to form a strong bond.
Since we could not substantially reduce the rate at which this occurs, we instead focused on increasing
the rate of catalysis.
We addressed problem iv by designing the weak bond to act as a pivot that guides the strong bonds
into place. Problem v was solved by making the weak bond protrude as much as possible from the
body of the puck. This increases the range of relative angles at which two pucks can be oriented while
still being able to form a weak bond. Problem vi was addressed by making the weak bond into a pivot
that can swing fairly freely. As the joint hinges, the two magnets are pushed further apart, so that the
bond can break if it swings far enough. This could be fine-tuned by making very small changes to the
magnetsʼ positions.
The foot mechanism was introduced to solve problem vii. Problem viii was solved in the final
design by the spikes in the head and tail sections (siehe Abbildung 3). These also help with problem i.
The remaining problems were solved primarily by trial and error.
Figur 6 shows a selection of previous iterations of the design, illustrating some of these prob-
lems and how they were solved. Design (A) was ineffective because weak bonds formed only rarely.
This is because the pucks have to be fairly specifically oriented with respect to one another in order
for the weak bond magnets to come in range of each other. Zusätzlich, the weak bond magnet of
an A-type monomer can bond strongly to the tail magnet of another monomer, blocking catalysis.
These two problems are solved in design (B) by making the weak bond protrude from the body of
136
Artificial Life Volume 18, Nummer 2
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
N. Virgo et al.
Evolvable Physical Self-Replicators
the puck, and by redesigning the strong bond so that the magnets are recessed away from the puckʼs
edge. Jedoch, it is relatively easy for strong bonds to form spontaneously in this design, and they
can also be catalyzed by the edge of the table. The spikes added to the strong bonds in design (C) help
to prevent spontaneous strong-bond formation, but they also interfere with the catalysis mechanism.
This design also features a hump on the opposite side to the weak bond; this is to prevent the edge of
the table from catalyzing bonds. Design (D) is the first to feature a weak bond that is designed to pivot
around a particular point, with a correspondingly curved set of strong-bond constraints. Jedoch,
strong bonds can still form spontaneously quite easily, and weak-bond formation is relatively rare.
Design (e) has a strong bond that is held together using repulsion rather than attraction (somit
the head and tail magnets are of the same polarity). Bedauerlicherweise, this tends to result in the magnets
jumping off the table to stack on top of one another, since that is energetically preferable to their
being near one another in a repulsive configuration. The weak bonds have also been redesigned to
be easier to form. Design (F ) is similar but uses attracting magnets again; its main problems are that
strand separation is very slow, and spontaneous strong-bond formation is still possible. Design (G) Ist
the first to feature a mechanism to break one of the weak bonds when a strong bond is catalyzed (zwei
dimers cannot fit together in such a way that they are joined at both weak bonds). Jedoch, Die
spontaneous formation of strong bonds is still possible, as is the formation of unwanted bonds
between the weak and strong bond magnets. Design (H) uses Velcro rather than magnets for the
strong bonds in an attempt to solve these problems. This idea was discarded because Velcro produces
a loose joint, which means the strong bonds do not align accurately enough for catalysis to take place.
Jedoch, we realized in testing this design that widening the lock-and-key structures on the strong
bonds helps to prevent spontaneous strong-bond formation.
Design (ich) is close to the final design and works fairly effectively. Its two remaining problems are
that unwanted weak-strong bonds can form (although they are quite weak), and monomers can be
attracted together by the strong-bond magnets in such a way that a strong bond can form if they are
nudged in the right way. These problems are solved in the final design by the addition of the central
head and tail spikes (siehe Abbildung 2) and by making the other spikes a lot larger.
We produced a total of 14 monomers, seven of type A and seven of type B. A total of 48 ex-
periments were performed with the final design, each lasting 25 min. Of these, 36 were seeded trials,
meaning that one dimer was added to a system containing the remaining 12 monomers. The system
is allowed to run for a few minutes before adding the dimer, to ensure that the initial conditions do
not affect the outcome. After the dimer was added, we counted the number of each type of polymer
jeden 2.5 min.
Of the 36 seeded trials, 12 were seeded with a type-AB dimer, 12 with type BA, 6 with type AA,
Und 6 with type BB. Since AA and BB are two phases of the same replicator, those two are plotted as
a single set of 12 Versuche.
The control experiment involves initializing the system with seven A-type monomers and seven
B-type ones, and is again run for 25 min. Twelve such experiments were conducted.
3 Ergebnisse
Figur 7 shows a series of photographs illustrating a successful replication cycle. The results from the
repeated trials are summarized below and in Figures 8 Und 9. We count as a side reaction the produc-
tion of any oligomer other than the seed type. In the AA/BB case we count AAA and BBB as copies
of the original rather than as side products, because there is no mechanism to prevent the formation
of these trimers, and because they can still catalyze the production of new BB or AA dimers.
In 19 out of the 36 seeded trials, no side reactions took place during the 25 min of the trial. In these
successful trials, an average of 4.3 duplicates (oder, in the AA/BB case, elongations) of the seed were
created in addition to the seed itself. The maximum possible number of copies is six in the AB or
BA case, or five in the AA/BB case, with a mismatched pair of monomers left over. This best-case
performance was achieved in four of the trials.
Artificial Life Volume 18, Nummer 2
137
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
N. Virgo et al.
Evolvable Physical Self-Replicators
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
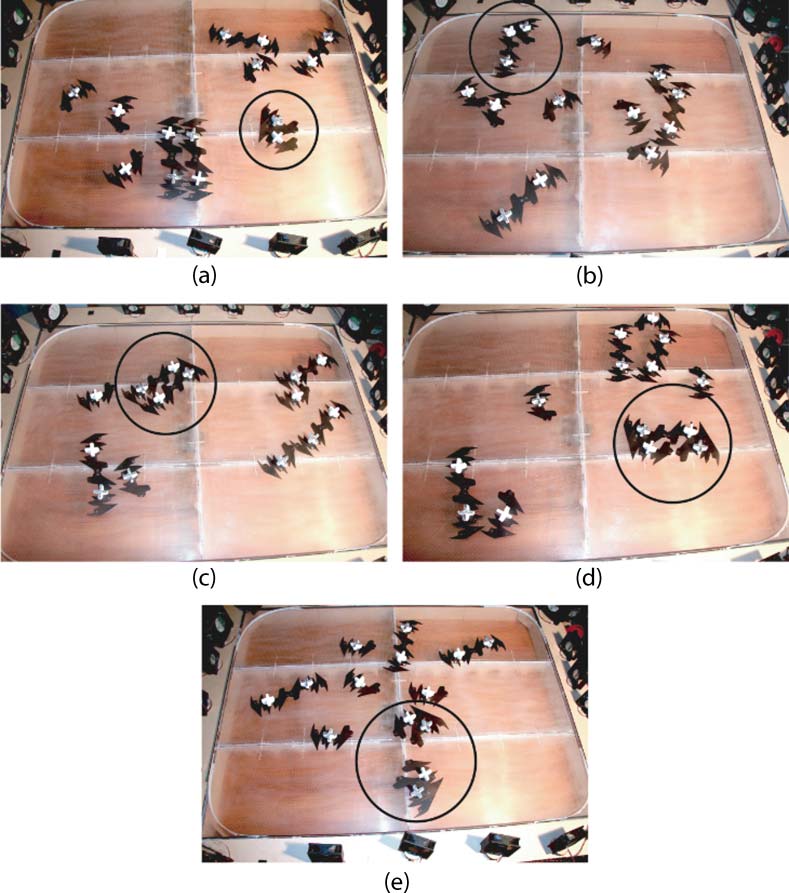
Figur 7. Photographs showing one round of the self-replication cycle. (A) An AB dimer (circled) is placed in a system
containing six A monomers (with white-topped sails) and six B monomers (black-topped sails). (B) A B monomer joins
to the A part of the dimer via a weak bond. (C) An A monomer joins via a weak bond to the B part of the dimer, and its
head constraints interlock slightly with the other monomerʼs tail constraints. (D) A collision with the tableʼs edge or
another molecule pushes the two monomers together, so that they form a strong bond. This breaks one of the two
weak bonds. Note that both dimers are of type AB. (e) Further collisions break the remaining weak bond, and the two
strands separate. This completes the autocatalytic cycle.
In the remaining 17 seeded trials, a side reaction produced an oligomer of a different species from
the seed. In some trials, this did not substantially disrupt the replication of the seed, but in others,
particularly if the side reaction happened early in the trial, the side product produced more replicates
than the seed dimer, effectively outcompeting it by using up the monomer supply. Under some
circumstances, it is also possible for the side product to join to a dimer of the seed type in a staggered
Mode (wie in Abbildung 5), catalyzing its elongation into a different species. For these reasons, der Mittelwert
number of duplicates of the seed after 25 min was only 1.7 im 17 trials where side reactions
138
Artificial Life Volume 18, Nummer 2
N. Virgo et al.
Evolvable Physical Self-Replicators
occurred, oder 3.1 over all 36 seeded trials. In four out of the 12 unseeded trials, there were no side
Reaktionen, meaning that only monomers were present after 25 min. Over all 12 unseeded trials, ein
Durchschnitt von 2.7 oligomers were produced, of various species.
Time series data are shown in Figures 8 Und 9, averaged over each of the four sets of 12 Versuche. In
Figur 8, all the side-reaction products are lumped into a single category. The error bars show that the
domination of duplicates of the seed over all other species is statistically significant to within a 95%
confidence interval at every time step.
4 Diskussion
One of the advantages of building a physical model rather than a computer simulation is that we
were forced to confront any problems that arose by redesigning the monomers, rather than by chang-
ing the parameters of a simulation. Many of the problems that arose have direct analogues on the
molecular scale, in particular spontaneous ligation, elongation at staggered ends, cyclization, und Prod-
uct inhibition.
The analogy with real chemistry is not a precise one. Our physical system is missing some con-
straints that apply to real chemistry (such as the requirement that a molecule be composed of atoms),
and introduces extra constraints of its own, such as the requirement that a puck be able to float
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 8. Time series plots showing the results of letting the system run for 25 min, seeded with one dimer of a particular
type, or with no dimer. In this plot, all polymers apart from those of the seed type are lumped into a single category. In
the case of the trials seeded with AA or BB, we count AA, BB, AAA, and BBB as a single category, since these can all be
produced by the catalysis process from the seed type. Each plot shows the average over 12 Versuche. The error bars show a
95% confidence interval.
Artificial Life Volume 18, Nummer 2
139
N. Virgo et al.
Evolvable Physical Self-Replicators
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figur 9. Time series data from the same trials as in Figure 8, with the reaction products split up by length. Note in
particular the drop in concentration of AA and BB dimers toward the end of the trial as they are converted into AAA
and BBB via elongation at staggered ends. (Error bars are omitted because they would be overlapping.)
effectively on the tableʼs surface. Our magnets all produced forces with the same range, whereas real
chemistry involves forces on several scales. The spatial and temporal scales are very different, and real
chemical systems probably react at a very low rate compared to the frequency of collisions. Despite
these differences, we suspect that a molecule capable of uncatalyzed template replication would re-
quire a similar level of complexity to our macroscopic monomers.
This presents some difficulty for the hypothesis that non-enzymatic template replication could
have been the origin of natural selection. If complex and specific (and hence large) molecules are
required for template replication, then it is hard to see how they could have arisen, except perhaps as
a product of an existing protometabolism that was already capable of evolution by natural selection
via some other mechanism. Several mechanisms have been proposed for achieving heredity without
the need for macromolecules; sehen [3] for a more detailed discussion and a proposal for how pre-
template natural selection could take place. This mechanism is based on networks of reactions be-
tween many simple molecules. These reaction networks do not need to have a precise form (und in
fact are chosen randomly in their model), and thus they avoid all the problems of specificity that arise
from needing to suppress side reactions.
It would be interesting to continue research along these lines, to see to what extent the problems
we have identified can be solved in macroscopic systems. Insbesondere, our system is not able to
reproduce strings of an arbitrary length, and for this a different mechanism would be required.
Another problem to be overcome is how to achieve high-fidelity replication in the face of a noisy
Umfeld. In some cases a varying environment may actually be beneficial, as demonstrated by
the use of temperature oscillations in Brevikʼs experiment.
140
Artificial Life Volume 18, Nummer 2
N. Virgo et al.
5 Conclusions
Evolvable Physical Self-Replicators
Self-replication by non-enzymatic template replication in a macroscopic system, using passive, inert
components, has been demonstrated. Producing such macroscopic template replication required mini-
mizing a number of side reactions and other difficulties that have analogues in the chemical domain.
These include spontaneous ligation, elongation at staggered ends, cyclization, and product inhibition. In
order for these to be minimized simultaneously, we had to design very specific and fairly complex
monomers. This suggests that template replication is hard in chemical systems, and perhaps must have
arisen through natural selection rather than preceding it [3].
The hexanucleotide replicator of von Kiedrowski was not evolvable, because no mutant of the orig-
inal sequence was capable of self-replication. Außerdem, the ends of the molecules were blocked, Also
that elongation was impossible. The replicators of Penrose and Penrose worked in 1D or had blocked
ends to prevent elongation. Breivikʼs model was not evolvable for the opposite reason: There was too
much spontaneous generation of novel sequences and an elongation catastrophe. Note that such spon-
taneous formation of novel sequences is quite different from mutations that preserve correlations be-
tween existing traits and their fitness. The latter permits Darwinian evolution; the former permits only
random search.
There is no doubt that the elongation catastrophe will be faced in all nano-scale self-replicating
systems as well. Natürlich, technology may allow such problems to be solved somewhat trivially if
monomers are allowed to contain switchable electromagnetic bonds [6] and can implement a finite
state machine [5], thus avoiding issues of product inhibition and mismatching by simply allowing
bonds to be made arbitrarily or on the basis of perfect local information. Jedoch, this arbitrary
programmability limits their utility in providing insight into possible molecular mechanisms of non-
enzymatic template replication that depend on carefully evolved steric and force constraints, welches ist
one of our main motivations here.
Danksagungen
The work was funded by the EU Grant E-FLUX FP7. Many thanks to Simon McGregor, Chris Buckley,
Eors Szathmáry, Roderich Gross, and Francesco Mondada. Special thanks to Jarle Breivik and Guenter
von Kiedrowski, whose work was the inspiration for this project.
Verweise
1. Breivik, J. (2001). Self-organization of template-replicating polymers and the spontaneous rise of genetic
Information. Entropy, 3, 273–279.
2. Eigen, M. (1971). Selforganization of matter and the evolution of biological macromolecules.
Naturwissenschaften, 58, 465–523.
3. Fernando, C., & Rowe, J. (2007). Natural selection in chemical evolution. Journal of Theoretical Biology,
247, 152–167.
4. Fernando, C. T., von Kiedrowski, G., & Szathmáry, E. (2007). A stochastic model of nonenzymatic
nucleic acid replication: “Elongators” sequester replicators. Journal of Molecular Evolution, 64,
572–585.
5. Griffith, S., Goldwater, D., & Jacobson, J. M. (2005). Self-replication from random parts. Natur,
437, 636.
6. Groß, R. M., Magnenat, S., Küchler, L., Massaras, V., Bonani, M., & Mondada, F. (2009). Towards an
autonomous evolution of non-biological physical organisms. In Proceedings of the 10th European Conference
on Artificial Life. Berlin: Springer-Verlag.
7. Maynard Smith, J. (1986). The problems of biology. Oxford, Vereinigtes Königreich: Oxford University Press.
8. Penrose, L. S. (1958). Mechanics of self-reproduction. Annals of Human Genetics, 23, 59–72.
9. Penrose, L. S. (1959). Self-reproducing machines. Scientific American, 200, 105–114.
Artificial Life Volume 18, Nummer 2
141
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
N. Virgo et al.
Evolvable Physical Self-Replicators
10. Penrose, L. S., & Penrose, R. (1957). A self-reproducing analogue. Natur, 179, 1183.
11. Szathmáry, E. (2000). The evolution of replicators. Philosophical Transactions of the Royal Society (London) Serie B,
355, 1669–1676.
12. Szathmáry, E. (2006). The origin of replicators and reproducers. Philosophical Transactions of the Royal Society
(London) Serie B, 361, 1761–1776.
13. Von Kiedrowski, G. (1986). A self-replicating hexadeoxy nucleotide. Angewandte Chemie International Edition
in English, 25, 932–935.
14. Zykov, V., Mytilinaios, E., Adams, B., & Lipson, H. (2005). Self-reproducing machines. Natur, 435,
163–164.
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
e
D
u
A
R
T
l
/
/
l
A
R
T
ich
C
e
–
P
D
F
/
/
/
/
1
8
2
1
2
9
1
6
6
2
8
9
2
A
R
T
l
/
_
A
_
0
0
0
5
6
P
D
.
F
B
j
G
u
e
S
T
T
Ö
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
142
Artificial Life Volume 18, Nummer 2