Effects of Pulvinar Inactivation on Spatial Decision-making
between Equal and Asymmetric Reward Options
Melanie Wilke1,2,3, Igor Kagan1,3, and Richard A. Andersen1
Abstrakt
■ The ability to selectively process visual inputs and to decide
between multiple movement options in an adaptive manner is
critical for survival. Such decisions are known to be influenced
by factors such as reward expectation and visual saliency. Der
dorsal pulvinar connects to a multitude of cortical areas that are
involved in visuospatial memory and integrate information
about upcoming eye movements with expected reward values.
Jedoch, it is unclear whether the dorsal pulvinar is critically
involved in spatial memory and reward-based oculomotor deci-
sion behavior. To examine this, we reversibly inactivated the
dorsal portion of the pulvinar while monkeys performed a de-
layed memory saccade task that included choices between
equally or unequally rewarded options. Pulvinar inactivation
resulted in a delay of saccade initiation toward memorized con-
tralesional targets but did not affect spatial memory. Weiter-
mehr, pulvinar inactivation caused a pronounced choice bias
toward the ipsilesional hemifield when the reward value in
the two hemifields was equal. Jedoch, this choice bias could
be alleviated by placing a high reward target into the contra-
lesional hemifield. The bias was less affected by the manipulation
of relative visual saliency between the two competing targets.
These results suggest that the dorsal pulvinar is involved in deter-
mining the behavioral desirability of movement goals while being
less critical for spatial memory and reward processing. ■
EINFÜHRUNG
Survival in an environment that contains a large amount
of information requires organisms to focus on a subset of
incoming sensory information and to strategically decide
between multiple response options. Spatial working
memory and the ability to direct eye movements to explore
spatial locations of potential interest constitute important
cognitive components of this selection process. Auswahl
of eye movement targets is guided by both the visual
saliency of the stimuli as well as motivational values (Opris
& Bruce, 2005). Zum Beispiel, primates typically saccade to
stimuli that predict high reward and saccade parameters
such as time of initiation, accuracy, and frequency of
choices are influenced by the predicted reward outcome
(Milstein & Dorris, 2007; McCoy & Platt, 2005; Lauwereyns,
Watanabe, Coe, & Hikosaka, 2002; Takikawa, Kawagoe,
Itoh, verstopft, & Hikosaka, 2002).
Although much of the research on visuospatial pro-
cessing and reward-based saccadic decision-making
has focused on cortical areas, there is ample evidence
that the dorsal pulvinar may play a critical role in these
functions (Grieve, Acuna, & Cudeiro, 2000). (1) Anatomical
evidence: Unlike the ventral pulvinar, which is primarily
interconnected with visual areas and receives input from
the superficial layers of the superior colliculus (SC), Die
dorsal pulvinar has reciprocal connections with cortical
1California Institute of Technology, 2University of Goettingen,
3German Primate Center, Leibniz Institute for Primate Research,
Goettingen, Deutschland
© 2013 Massachusetts Institute of Technology
areas that maintain spatial information for upcoming eye
movements and are modulated by expected reward values
(Das hier & Lyon, 2007; Stepniewska, 2004). Speziell, Die
medial portion of the dorsal pulvinar, which we focus
on in the current study, is reciprocally interconnected
with the posterior parietal cortex, the upper bank of the
STS (Cappe, Morel, & Rouiller, 2007; Gutierrez, Cola, Seltzer,
& Cusick, 2000; Yeterian & Pandya, 1989; Asanuma, Andersen,
& Cowan, 1985), the pFC (Gutierrez et al., 2000; Selemon &
Goldman-Rakic, 1988), and limbic structures such as the
posterior parahippocampal gyrus and the posterior cingu-
late cortex (Baleydier & Mauguiere, 1985). This connec-
tivity with cortical areas known to be involved in the
processing of behavioral relevance has led to the hypoth-
esis that the pulvinar enhances the processing of biologi-
cally relevant stimuli by interaction with those regions
(Padmala, Lim, & Pessoa, 2010). (2) Physiological evidence:
Firing rates and response variability of pulvinar neurons in
monkeys are modulated by visual attention and perception
(Wilke, Mueller, & Leopold, 2009; Bender & Youakim,
2001; Benevento & Port, 1995; Robinson & Petersen, 1992;
Petersen, Robinson, & Keys, 1985), and neuroimaging
studies in humans demonstrated attention-related BOLD
signals in the pulvinar (Hulme, Whiteley, & Shipp, 2010;
Padmala et al., 2010; Kastner et al., 2004). Pulvinar neurons
exhibit presaccadic and postsaccadic response changes
(Berman & Wurtz, 2011; Benevento & Port, 1995; Robinson,
McClurkin, Kertzman, & Petersen, 1991; Robinson, McClurkin,
& Kertzman, 1990), and visual responses are enhanced
when a stimulus represents the goal of a saccade (Petersen
Zeitschrift für kognitive Neurowissenschaften 25:8, S. 1270–1283
doi:10.1162/jocn_a_00399
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
Ö
9
C
5
N
1
_
3
A
/
_
J
0
Ö
0
C
3
N
9
9
_
A
P
_
D
0
0
B
3
j
9
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
et al., 1985). Zusätzlich, fMRI studies in humans and
monkeys demonstrated BOLD modulation in the pulvinar
during saccade execution (Krebs et al., 2010; Bäcker, Patel,
Corbetta, & Snyder, 2006). (3) Lesion evidence: Perhaps,
the most convincing evidence for a critical role of the
pulvinar in spatial orienting and eye movement decisions
can be derived from lesion studies. Pulvinar lesions in
humans (Arend, Rafal, & Station, 2008; Karnath, Himmelbach,
& Rorden, 2002; Danziger, Station, Owen, & Rafal, 2001; Rafal
& Posner, 1987; Zihl & von Cramon, 1979) and monkeys
(Wilke, Turchi, Schmied, & Leopold, 2010; Petersen, Robinson,
& Morris, 1987) lead to spatial neglect symptoms, einschließlich
difficulties to direct attention toward the contralesional
field. Although the causal role of the pulvinar in the selec-
tion of eye movement targets remains less explored, recent
studies reported lesion-induced deficits in oculomotor
integration and saccadic decision-making (Van der Stigchel,
Arend, van Koningsbruggen, & Rafal, 2010; Wilke, Turchi,
et al., 2010; Arend, Machado, et al., 2008). Speziell, A
previous study in monkeys that employed a visually guided
saccade task showed that reversible inactivation of the
dorsal pulvinar is associated with a choice bias toward the
ipsilesional hemifield although performance toward single
targets remained largely intact (Wilke, Turchi, et al., 2010).
The primary goal of the current study was to investi-
gate how inactivation of the dorsal pulvinar influences
spatial memory and reward-based oculomotor decision
behavior. We were particularly interested to see whether
the spatial choice bias following pulvinar inactivation that
has been observed in the previous study (Wilke, Turchi,
et al., 2010) occurs at the level of perceptual extinction or
at later stages of the decision-making process. Zu diesem Zweck,
we employed a delayed memory saccade task with inter-
leaved instructed (single target) and choice (bilateral targets)
trials and varied the target-associated reward magnitude. In einem
subset of experiments, we also varied the visual saliency by
using targets with different luminance levels. We hypothe-
sized that, if the choice bias was primarily because of
perceptual extinction, it would persist even when the
stimulus in the contralesional field indicated a higher
reward in comparison with the ipsilesional field. In con-
trast, if the biased spatial choice behavior was because of
a motivational/intentional movement bias, wir erwartet
the high reward stimulus to increase the willingness to
perform an eye movement toward the contralesional space.
METHODEN
All surgical and animal care procedures were done in
accordance with National Institutes of Health guidelines
and were approved by the California Institute of Technol-
Ogy, Animal Care and Use Committee.
(PEEK) headpost embedded in Palacos bone cement
(BioMet) attached to the cranium with short ceramic
screws (Thomas Recording), under general anesthesia.
Monkeys were chronically implanted with a 22-gauge
guide PEEK cannula (Plastics One, Roanoke, VA) pene-
trating the dura and targeting the dorsal pulvinar in the
left hemisphere. The top portion of this guide cannula was
set in a corrugated ceramic cylinder, which was affixed to
the skull with ceramic screws and bone cement and served
as a guide for (acute) insertion of the 28-gauge internal
PEEK cannula during the experiments. Before each sur-
gery, we acquired high-resolution anatomical MRI (sehen
Structural MRI Acquisition). Anatomical MRI scans were
transformed into an AC–PC plane, and cannula positions
were planned by comparing the coronal MRI slices with
the combined MRI and histology atlas of the macaque brain
(Saleem & Logothetis, 2006).
Pulvinar Inactivation
Microinfusions of the GABA-A agonist 4,5,6,7-tetrahydro-
isoxazolo[5,4-C]-pyridin-3-ol (THIP; Tocris Bioscience,
Ellisville, MO) were made in each inactivation session
via a sterile 28-gauge internal cannula. To confirm the injec-
tion locations, the MR contrast agent gadolinium (Magnevist,
Berlex Imaging, Montville, NJ) was added to the solution of
weak phosphate buffered saline, and anatomical MR images
were acquired. The THIP was dissolved in PBS (along with
the gadolinium), and the solution (pH 7.0–7.5) was sterile
filtered (Corning, Inc., Corning, New York) before injection. Speci-
ficity of drug effects was further validated in an additional
control session in which only the vehicle and gadolinium
were injected. Total injection volumes ranged from 2.0 Zu
4.5 μl of 6.6 mg/ml of THIP, which were delivered at a rate
of 0.7–1.0 μl/min using a 100-μl gas-tight Hamilton syringe
driven by a digital infusion pump (Harvard Apparatus,
Holliston, MA). The infusions were carried out while the
animals were awake and sitting in a primate chair, mit
their heads restrained via implanted head posts, Vor
the MRI scanning. All injections in both monkeys were
made in the left pulvinar. We conducted 13 inactivation
sessions into the dorsal pulvinar in monkey F (two injec-
tion locations in the left pulvinar, seven and six sessions)
and five sessions in monkey R (one injection location in
the left pulvinar; Figure 1A). Because the results from the
two injection sites in monkey F were similar, we pooled the
data for the sake of clarity. Kontrolle (no inactivation) Daten
collection was interleaved with drug injection sessions.
Behavioral effects following THIP injections into the pulvi-
nar typically lasted several hours. The minimum interval
between injections was 2 Tage.
Preparation
Two male rhesus macaques (Macaca mulatta) weighing
8–10 kg were implanted with an MR-compatible plastic
Structural MRI Acquisition
Anatomical images were acquired in a Bruker Biospec
4.7-T/60-cm vertical bore scanner equipped with a BGA38S2
gradient coil. A linear birdcage volume RF coil (Bruker;
Wilke, Kagan, and Andersen
1271
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
Ö
9
C
5
N
1
_
3
A
/
_
J
0
Ö
0
C
3
N
9
9
_
A
P
_
D
0
0
B
3
j
9
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
/
.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
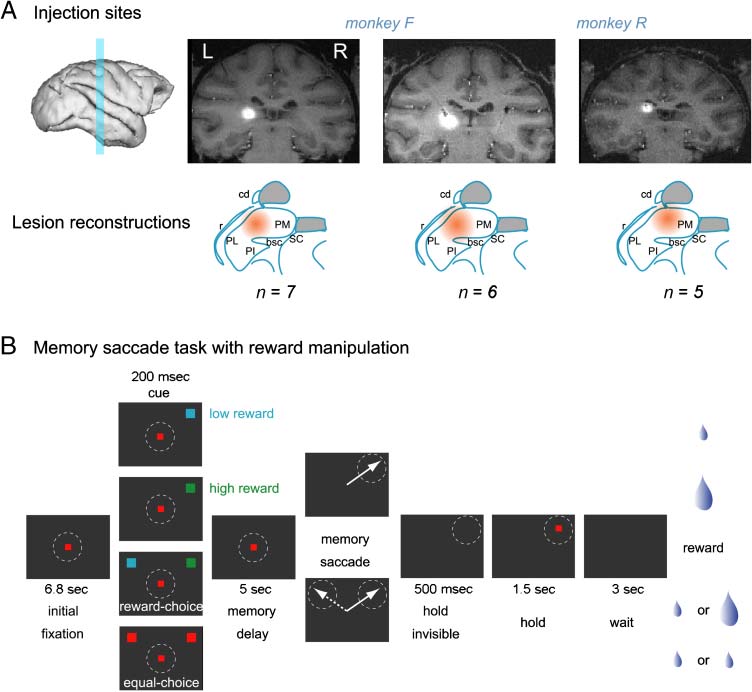
Figur 1. Inactivation sites
and experimental design.
(A) Inactivation sites in the
two monkeys visualized with
coinjections of gadolinium
MR contrast agent. The top
shows coronal MR images
(0.8-mm thickness) around the
tip of the cannula between 20
Und 40 min after THIP injection.
The coinjected gadolinium
appears white. In all cases,
the center of the injection
was between +3 Und +5 mm
in Horseley–Clark coordinates.
The bottom shows the
estimated injection locations
(orange). Estimation was
performed by overlaying the
pulvinar landmarks derived
from the anatomical monkey
atlas on the aligned and
scaled MRI slices (Saleem &
Logothetis, 2006). Injections
shown represent the typical
injection volume used in
the current study (3–4 μl).
Abkürzungen: bc = brachium
of SC; cd = caudate; PM =
medial pulvinar (target
Struktur); PL = lateral pulvinar;
PI = inferior pulvinar; r =
reticular nucleus. (B) Memory-
guided saccade task. A trial
started with the presentation of a
fixation spot followed by the
presentation of a saccade cue in either the left or right or in both hemifields, selected randomly from 18 possible positions. In instructed trials, only a
single target was presented. In choice trials, two targets appeared simultaneously in the left and right hemifields equidistantly from the fixation point
at the same vertical position. The color of the cue signaled the amount of upcoming reward (Blau: low reward, Grün: high reward, Rot: niedrig [equal]
reward). Monkeys were trained on this cue–reward association before the start of the inactivation experiments. Monkeys were required to keep fixating
following cue offset and to make a saccade following a 5-sec memory delay. The offset of the fixation point served as signal to execute the saccade.
monkeys F and R) or a quadrature surface RF coil (H. Merkle;
monkey F), allowed whole-head coverage. Monkeys were
previously adapted to the scanner environment and sat
awake in a custom-made MR-compatible chair (Kagan,
Iyer, Lindner, & Andersen, 2010). Coronal anatomical
T1-weighted images were acquired using a MDEFT
sequence with an in-plane resolution of 0.5 mm and a
slice thickness of 0.8 mm.
Behavioral Tasks
Behavioral data were acquired between 50 Und 180 min
after the start of the injection.
Memory-guided Saccade Task with
Variable Reward Magnitude
Monkeys were tested with a delayed memory saccade
task consisting of two major trial types: (1) instructed
Versuche, in which the presentation of a single cue defined
the saccade target unequivocally; Und (2) choice trials,
in which two targets were presented in opposite hemi-
fields equidistant from the central fixation, and monkeys
could perform a saccade toward either one. The trial se-
quence is depicted in Figure 1B. Monkeys were required
to fixate a red square in the middle of the screen. Nach
6.8 Sek, either one (instructed) or two (Auswahl) cues of
0.37° square size were presented for 200 ms, indicating
the target position(S) for the saccade and the associated
reward magnitude. Following cue offset, monkeys were
required to maintain the target position in memory and
keep central fixation for another 5 Sek (memory period).
The end of this memory delay period was signaled by the
offset of the fixation spot (“go signal”), and the monkey
had to perform a correct saccade within 500 ms. Der
saccade was judged to be correct if the eye position fell
within a radius of 5° and was held for additional 500 ms,
after which the confirmation target at the previously cued
position reappeared. The visual cues were randomly
chosen from 18 (10°–16° eccentricity) Standorte, nine in
the left and nine in the right visual hemifield. Incorrect
trials with fixation breaks or incorrect saccades were
1272
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 8
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
Ö
9
C
5
N
1
_
3
A
/
_
J
0
Ö
0
C
3
N
9
9
_
A
P
_
D
0
0
B
3
j
9
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
/
.
F
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
aborted; successful trials were rewarded with water.
Before the start of the experiments, monkeys were trained
to associate the color of the cue with a certain reward
magnitude that was delivered after a successful memory
saccade (Figure 1B). Speziell, a blue cue predicted
low reward, whereas a green cue predicted high reward
(Verhältnis: 1/3, the liquid volumes ranged from 0.22–0.33 ml
for low reward to 0.66–0.99 ml for high reward). Experi-
ments started when monkeys selected the high reward
cue in >90% of the choice trials during training sessions.
In instructed trials, the monkey had to perform a correct
saccade even for low reward targets; ansonsten, the trial
with low reward magnitude was repeated until a correct
response was made. Except for this low reward persistence,
all trials types (target locations, instructed/choice, reward
condition) were randomly interleaved.
Memory-guided Saccade Task with
Variable Contrast Levels
To test for effects of visual saliency on saccade performance
and choices, we varied in a subset of sessions the luminance
contrast of the saccade cues (10% vs. 100% of the maximal
red luminance presented on the black background) while
keeping the reward amount constant. Although we could
not measure the absolute luminance of the stimuli because
we used the LCD goggles with the incorporated lens optics
for stimulus presentation, the low contrast cue was faint
but suprathreshold for goggle-adapted humans in the dark
Zimmer (similar to the scanner bore). All other aspects of
the task were the same as described for the reward ex-
periments above. In the six control and inactivation ses-
sions where we tested the effect of luminance contrast,
those were interleaved with the reward magnitude ex-
periment in a counterbalanced block-wise manner, mit
each block having a duration of 20 min.
Stimulus Presentation, Online Behavioral Control,
and Data Acquisition
Visual stimuli were presented on 800 × 600, 60-Hz LCD
goggles (Resonance Technology, Northridge, CA) sub-
tending 30° × 24° of visual angle using custom OpenGL
Software. Eye position was monitored at 60 Hz and 0.5°–1°
accuracy with a mini-IR camera (Resonance Technology or
Arrington Research, Scottsdale, AZ) and recorded together
with stimulus and timing information. Online behavioral
control and feedback were implemented in a LabVIEW
RT (National Instruments, Austin, TX).
Eye Movement Recording and Analysis
The position of the eye was monitored in real time and
stored off-line. Trials in which the monkey did not main-
tain fixation or exceeded the saccade reaction time criteria
(500 msec from the offset of the fixation point) war
aborted. The central fixation window radius was 3°–4°,
and the peripheral saccadic target window radius was 5°.
At 60-Hz sampling rate of the video-based IR eye tracker,
saccades of >0.33° amplitude can in principle be de-
tected by a 20°/sec velocity threshold. Jedoch, Weil
of instrumental noise in the eye tracker, ergebend
∼0.5° accuracy, we limited our analysis to saccades of
>1° that could be reliably identified using a custom
combined velocity and acceleration algorithm.
Preliminary data analysis did not reveal a systematic
effect of pulvinar inactivation on particular target posi-
tions within a given hemifield. Daher, results for the dif-
ferent locations within a given hemifield were pooled
together. Unless otherwise specified, data were submitted
to a three-way ANOVA with the factors Drug (control vs.
inactivation), Reward (low vs. hoch), and Hemifield (links
vs. Rechts). This analysis was followed by a two-tailed t test
for statistical comparisons separately for each hemifield.
The significance level was set to p < .05.
RESULTS
We tested two monkeys following injection of the GABA-
A agonist THIP into the dorsal pulvinar (13 sessions in
monkey F, 5 sessions in monkey R, and the same num-
ber of interleaved control sessions). Injection locations
were initially planned by means of presurgical anatomical
MRI and verified in each session by imaging the spread of
the coinjected contrast agent gadolinium associated with
the injection (Experimental Methods). Imaging data indi-
cate that inactivation was primarily in the medio-dorsal
pulvinar (Figure 1A). We use the terms “ipsilesional”
and “contralesional” with respect to the inactivated hemi-
sphere (e.g., after an injection into the left hemisphere,
the left visual hemifield is “ipsilesional”).
Inactivation Effects on Memory Saccades with
Different Rewards: Instructed Trials
To examine the effect of pulvinar inactivation on saccades
toward memorized targets, we employed a delayed memory
saccade task with single targets (Figure 1B, “instructed”).
Targets were associated with different reward magnitudes,
which were signaled by the color of the cue. Monkeys were
trained on this association before the beginning of the ex-
periments. We evaluated two different types of saccade
errors: (1) fixation break saccades (premature saccade
before the go signal but after the cue was presented) and
(2) incorrect saccades (saccade after the go signal but to
the wrong position). After inactivation, there was no sub-
stantial impairment in the monkeyʼs ability to correctly
remember the cued target position and saccade toward it
(Figure 2A). In control sessions, there was a tendency for
fixation aborts toward cues signaling higher reward, which
reached significance for the right hemifield in monkey F
and for both hemifields in monkey R (two-tailed t test,
p < .05; Figure 2B, Table 1). In addition, the proportion
of correctly executed saccades after the delay period
Wilke, Kagan, and Andersen
1273
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
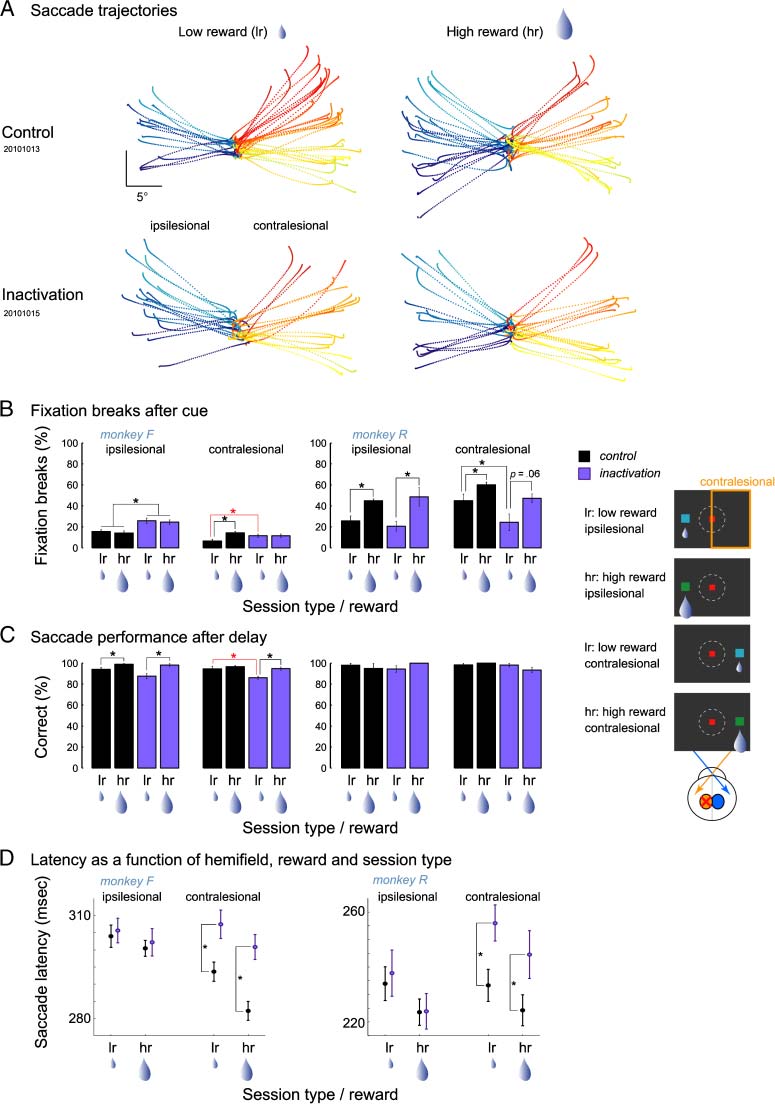
Figure 2. Instructed saccades. (A) Trajectories of instructed memory saccades toward single targets in a typical control and inactivation session
in monkey F. Saccades were made to targets in 18 possible directions at an eccentricity between 10° and 16°. Saccadic eye movements (dotted
curves) in control (top) and inactivation sessions (bottom) for low (left) and high (right) reward. Trajectory colors denote the locations of the
saccade targets (red–yellow: right, cyan–blue: left; scale bar, 5°). Here, and in all other figures, the left hemifield is ipsilesional, and right hemifield
is contralesional after the inactivation. (B) Effect of inactivation on fixation aborts. The percent of fixation aborts following cue presentation for
each hemifield, reward condition, and monkey. Data were averaged over sessions and pooled over the nine target positions in each hemifield. Here
and in other figures, purple bars represent data obtained after inactivation; black bars, data from control sessions. Error bars indicate SE across
sessions, and stars denote p < .05 for the specific comparison as indicated by connecting lines (red asterisk denotes contralesional deficits). Inset
on the right illustrates the four instructed cue conditions. (C) Effect of inactivation on saccade performance. The percent of correct saccades that
monkeys made after they successfully maintained fixation during the delay period for each hemifield, reward condition, and monkey. Same notations
as in (B). Red star indicates the contralesional decrease of correct low reward saccades in monkey F, the only contralesional deficit besides the
prolongation of saccade latencies observed for the instructed trials. (D) Mean saccade latencies as a function of reward and hemifield. Saccade latencies
toward the contralesional space became longer in both monkeys after inactivation.
1274
Journal of Cognitive Neuroscience
Volume 25, Number 8
Table 1. Task Performance in Instructed Trials for Control and Inactivation Sessions
Fixation Breaks (%)
Saccade Errors (%)
Overall
Performance (%)
Latency (msec)
Low
Reward
High
Reward
Low
Reward
High
Reward
Low
Reward
High
Reward
Low
Reward
High
Reward
15.6*
25.8*
14.1*
24.5*
5.9**
12.4**
6.5*,**
14.2**
5.4*
11.6*
11.5
13.9*,**
25.5**
44.9**
20.1
48.6
45.0*,**
60.0**
2.0
1.7
1.7
24.4*
47.3
1.82
1.0**
1.8**
3.3
5.2**
5.0
0
0
6.5
79.5*,**
85.1*,**
65.5*
88.5*
76.2*,**
72.8**
74.5
74.1*
82.9
83.9**
52.6**
51.5
53.8*,**
39.9**
74.8*
49.3
304
306
294*,**
307*
234
238
233*
256*
300
302
282*
301*
224
224
224*
245*
HF
LH
LH
RH
RH
LH
LH
RH
RH
Monkey
Session
F
R
Control
Inactivation
Control
Inactivation
Control
Inactivation
Control
Inactivation
LH = left (ipsilesional) hemifield; RH = right (contralesional) hemifield. Significant inactivation effects are in bold.
*p < .05 for comparison between control versus inactivation (separate two-tailed t tests within each reward condition and hemifield).
**p < .05 for comparison between low reward versus high reward (separate two-tailed t tests within session and hemifield).
was significantly increased in monkey F for high rewarded
positions, and saccade latencies tended to be shorter in
those trials in both monkeys (Figure 2C, Table 1).
In both monkeys, inactivation affected the pattern of
fixation aborts, as indicated by an interaction between
the factors Drug × Hemifield (ANOVA, p < .05 in monkey F,
p = .06 in monkey R). Specifically, monkey F showed
a significant increase of fixation aborts toward cues in the
ipsilesional hemifield, whereas fixation aborts toward
the contralesional hemifield decreased following inactiva-
tion in monkey R (Figure 2B, Table 1). The only signifi-
cant contralesional impairments in instructed trials were
the increased fixation breaks and slightly decreased sac-
cade performance for low-reward contralesional targets in
monkey F (Figure 2B, red asterisk).
The most pronounced and consistent effect of inactiva-
tion on instructed trials was observed for saccade latencies.
In both monkeys, inactivation resulted in a significant in-
crease of saccade latencies for contralesional positions
(ANOVA, Drug × Hemifield, p < .05; Figure 2D). Separate
analysis for the ipsilesional and contralesional hemifields
revealed a significant effect of inactivation for both low-
and high-reward contralesional targets (two-tailed t test,
p < .05) but not for ipsilesional targets ( p > .7; Tisch 1).
No significant interaction between Drug × Reward or
Drug × Reward × Hemifield was found in instructed trials
for any performance measure or saccade latencies, weiter
indicating that pulvinar inactivation did not interfere with
reward processing per se (Figure 2A–D).
Effects on Saccade Choices between Equal and
Unequal Options
To investigate how pulvinar inactivation altered selection
between two response options, we next examined the
inactivation effects on choices between two simul-
taneously presented cues in both hemifields. The two
cues predicted either equal or differing reward magni-
tudes, as indicated by their color (Figure 1B, “choice”).
Although individual spatial preferences were apparent in
control sessions (monkey F preferred right hemifield, Und
monkey R preferred left hemifield under conditions of equal
reward), monkeys still chose both sides of space when the
two cues did not differ in respect to expected reward mag-
nitude (Figure 3A and B). Following inactivation, Affen
showed a strong decrease of contralesional choices (Feige-
ure 3A and B) when selecting between equal rewards.
Speziell, monkey F chose right (contralesional to be)
targets in 78% of the trials in control sessions; equal reward
choices toward the contralesional field decreased to 42%
following pulvinar inactivation (one-way ANOVA, main effect
of drug, P < .05). A similar effect was found in monkey R
(control: 31% vs. inactivation: 5%, p < .05; Figure 3B).
We next evaluated the effect of reward expectation on
choices between targets with unequal reward magnitude.
As predicted from previous studies (McCoy & Platt, 2005;
Platt & Glimcher, 1999), in control sessions, monkeys
typically selected the target that predicted the higher reward
(>95%). For the inactivation sessions, we hypothesized
that perceptual extinction during bilateral cue presen-
tation would be expressed as a tendency to choose ipsi-
lesional targets even if they yielded the low reward. An
die andere Hand, if pulvinar inactivation does not act
primarily on the perceptual processing but, zum Beispiel,
leads to an increased effort to perform a contralesional
Bewegung (thus decreasing “desirability” of contralesional
Raum), we expected that a higher reward magnitude for
contralesional positions will increase the proportion of
choices toward this side. The pattern of results supports
the latter hypothesis. After inactivation, monkeys still
Wilke, Kagan, and Andersen
1275
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
Ö
9
C
5
N
1
_
3
A
/
_
J
0
Ö
0
C
3
N
9
9
_
A
P
_
D
0
0
B
3
j
9
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
/
F
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
D
Ö
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
Ö
9
C
5
N
1
_
3
A
/
_
J
0
Ö
0
C
3
N
9
9
_
A
P
_
D
0
0
B
3
j
9
G
9
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
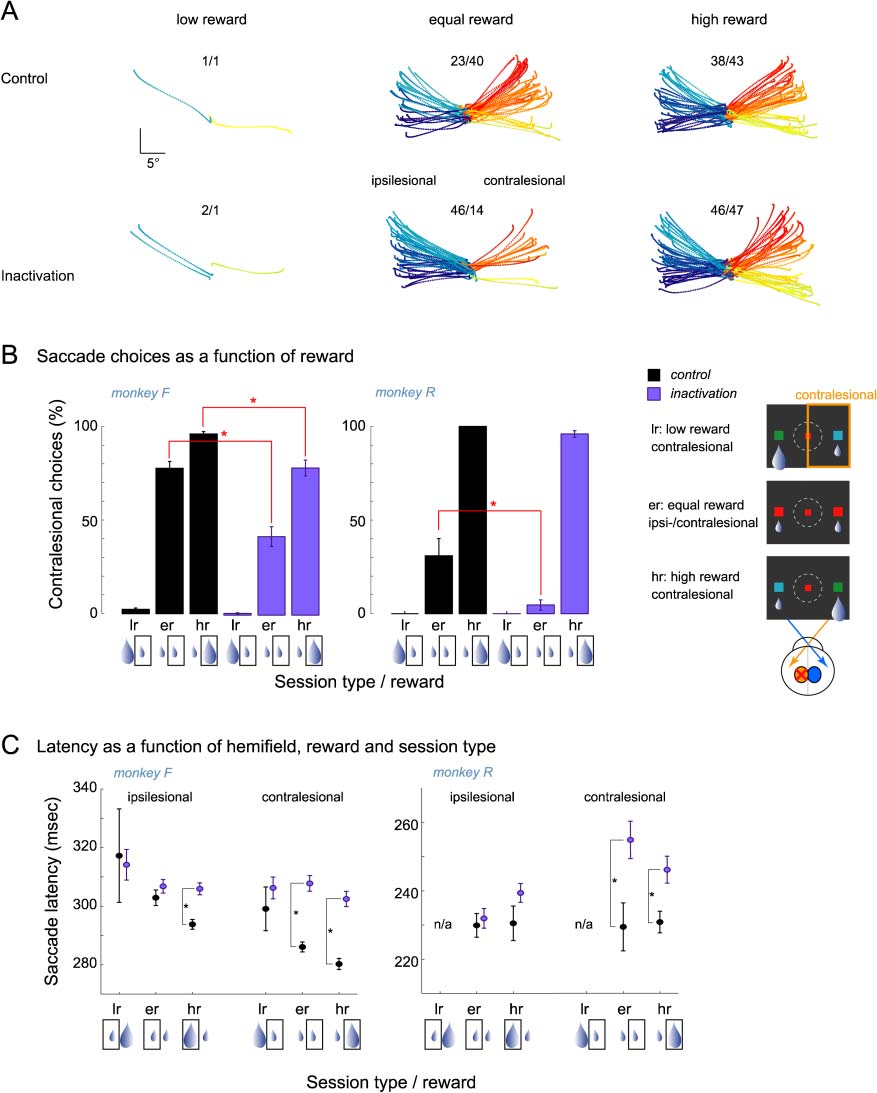
Figur 3. Choice behavior. (A) Trajectories of selected memory saccades in choice trials in a typical control and inactivation session in monkey F
(same sessions as in Figure 2A for instructed saccades). Note the inactivation-induced decrease of choices toward the contralesional space in the
equal reward condition as opposed to the unaffected contralesional choice behavior for targets associated with high reward. The numbers above the
trajectories indicate the number of saccades made to the left and right hemifields. Other notations as in Figure 2A. (B) Percentage of saccades toward
the contralesional (Rechts) hemifield in choice trials as a function of reward and session type. In both monkeys, the percentage of contralesional
choices decreased significantly during selection between equal rewards but remained high when a large reward was placed in the contralesional
hemifield. Inset on the right illustrates the three-choice cue conditions. The rectangular frame around the small or large reward icon in each left/right
reward pair under the horizontal axis denotes the corresponding selection. (C) Saccade latencies for ipsilesional and contralesional choices as a
function of expected reward. Latencies for low reward choices were not plotted in monkey R because of the lack of those trials. Note that saccade
latencies toward the contralesional space became longer in both monkeys after inactivation, similar to instructed saccades.
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
1276
Zeitschrift für kognitive Neurowissenschaften
Volumen 25, Nummer 8
strongly preferred contralesional over ipsilesional target
positions when the contralesional cue indicated the higher
reward (monkey F: control = 96% vs. inactivation = 79%;
monkey R: control = 100% vs. inactivation = 96%). Daher,
although inactivation somewhat reduced the proportion
of contralesional high-reward choices (ANOVA for high-
reward choices, main effect of Drug: monkey F, P <
.05; monkey R, p = .09), reward expectation largely
overcame the inactivation-induced spatial choice bias in
both monkeys.
However, despite almost complete alleviation of the
choice bias by the high reward, we still observed in-
creased latencies to contralesional low- and high-reward
targets, as in instructed saccades (Figure 3C, Table 2).
Accordingly, a separate analysis for each hemifield re-
vealed a significant inactivation effect for contralesional
saccades in both monkeys (two-tailed t test, monkeys F
and R, p < .05). Although the latency increase after in-
activation was more pronounced for contralesional targets,
monkey F also showed a latency increase for ipsilesional
high-reward saccades (monkey F, p < .05; monkey R,
p = .37). Taken together, this pattern of results indicates
at least a partial dissociation between response selection
and saccade initiation deficits.
Effect of Visual Saliency on Saccade Choices
The alleviation of the spatial choice bias by high re-
ward association suggested that the bias following pul-
vinar inactivation in the equal reward condition was
not caused by a perceptual deficit but rather by an atten-
tional/intentional impairment. Nonetheless, we wondered
whether monkeys overcame their choice bias because the
cue associated with the high reward value had acquired a
higher perceptual saliency. To assess the influence of visual
saliency on choice behavior more directly, we conducted
six control and six inactivation sessions in monkey F, where
we directly varied the luminance of the cues, without
reward manipulation. The temporal structure of the task
was as in the reward experiments described above (Fig-
ure 1B). In instructed trials, a single target of either low
(10%) or high luminance (100% of the luminance used in
the regular equal reward condition) was shown, and in
choice trials, the two targets, presented simultaneously
in both hemifields, were either of equal or differing lumi-
nance (Figure 4A, right inset). As illustrated in Figure 4A,
pulvinar inactivation significantly increased the number
of fixation aborts (ANOVA, main effect of Drug, p < .05;
Table 3). Fixation aborts after inactivation were more
pronounced toward the ipsilesional hemifield, although
this tendency was also present for contralesional positions
(ANOVA, Hemifield × Drug, p = .49). Saccade errors were
not significantly affected in either hemifield (main effect of
Drug, p = .09; Hemifield × Drug, p = .07; Figure 4B).
No significant interaction between Contrast × Drug or
Contrast × Drug × Hemifield was found for either error
type ( p > .4), indicating that pulvinar lesions did not
specifically impair processing of low luminance saccade
Hinweise (Figure 4A and B). Inactivation increased saccade
latencies for both ipsilesional and contralesional stimuli,
although this comparison did not reach statistical sig-
nificance ( p > .07; Figure 4C).
Most relevant for our question is the analysis of choices
between targets with equal or different luminance con-
Bünde (Figure 4D, links). As in the reward experiment de-
scribed above, inactivation significantly decreased the
proportion of contralesional choices when the luminance
in both hemifields was equal (Kontrolle: 72% vs. inactivation:
22%; one-way ANOVA, main effect of Drug, P < .05).
In control sessions, when given the choice between two
Table 2. Saccade Latencies in Choice Trials for Control and Inactivation Sessions
Monkey
F
R
Session
Control
Inactivation
Control
Inactivation
Control
Inactivation
Control
Inactivation
HF
LH
LH
RH
RH
LH
LH
RH
RH
Latency (msec)
Low Reward
Equal Reward
High Reward
317*
314
299
306
n.a.
n.a.
n.a.
n.a.
303
307
287**
308**
231
233
231
256
294*,**
306**
280**
302**
232
240
232**
247**
n.a. = not applicable because of lack of trials.
*Next to the equal (low) reward value denotes p < .05 for comparison between equal (low) reward versus high reward (separate two-tailed t tests for
within session and hemifield).
**Next to the control value denotes p < .05 for comparison between control versus inactivation (separate two-tailed t tests for within each reward
condition and session type).
Wilke, Kagan, and Andersen
1277
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
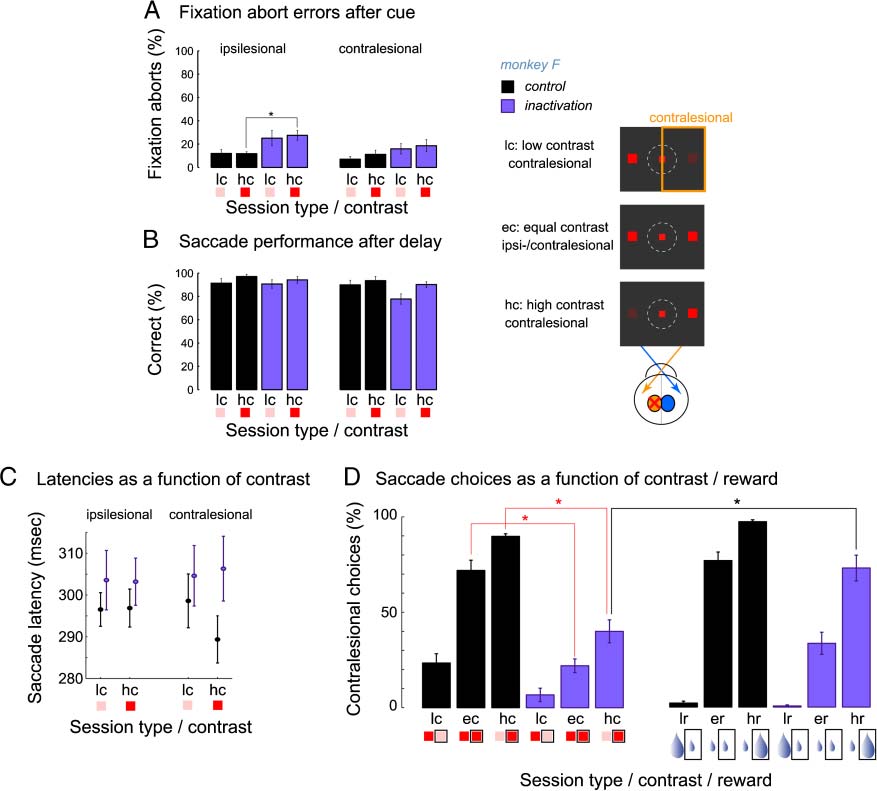
Figure 4. Effect of luminance contrast on saccade behavior and choices. Memory-guided saccade task with luminance contrast manipulation
(monkey F, six control and six inactivation sessions). The general task structure was as described in Figure 1B. The luminance contrast of the
targets was 10% (low contrast) or 100% (high contrast). Positions of low and high contrast cues were randomly varied between 18 possible
target positions. Reward magnitude was held constant for all contrast levels. Inset on the right illustrates the three-choice cue conditions
(single-target instructed conditions are not shown). (A) The percent of fixation aborts in instructed trials following cue presentation for each
hemifield and contrast condition. (B) Proportion of correct saccades in instructed trials for each hemifield and contrast condition. (C) Mean saccade
latency in instructed trials as a function of contrast and hemifield. (D, left) Percentage of saccades toward the contralesional (right) hemifield in
choice trials as a function of contrast and session type, in contrast manipulation blocks. Note that, after inactivation, even when a high contrast
target was presented in the contralesional field together with a low contrast target in the ipsilesional field, the monkey still more often selected
the ipsilesional target (60%). The rectangular frame around the small (faint) or large (saturated) contrast icon in each left/right contrast pair
under the horizontal axis denotes the corresponding selection. (D, right) Percentage of saccades toward the contralesional (right) hemifield in
choice trials as a function of reward and session type, in reward manipulation blocks, tested in the same sessions with contrast manipulation
blocks. These data are a subset of 13 control and 13 inactivation reward manipulation sessions shown in Figure 3B for monkey F.
targets with different luminance, the monkey preferred
the high over the low luminance targets (90% choices of
high luminance in the right, contralesional hemifield to be;
and 77% in the left hemifield). Following inactivation,
a high luminance target in the contralesional hemifield
coupled with a low luminance target in the ipsilesional
hemifield somewhat increased the proportion of contra-
lesional choices as compared with the equal luminance
condition (40% vs. 22%, respectively). Nonetheless, as de-
scribed in the previous section, a high reward stimulus
alleviated the ipsilesional selection bias more effectively
than manipulation of visual saliency (monkey F, high-
reward contralesional selection of 79% overall). In the
subset of six inactivation sessions in monkey F where we
interleaved both the reward and luminance contrast manip-
ulations, contralesional choice was 70% for high reward
vs. 40% for high contrast, which was significantly different
(Figure 4D, right; Table 4).
The higher effectiveness of reward as opposed to the
luminance manipulation is also expressed by a significant
1278
Journal of Cognitive Neuroscience
Volume 25, Number 8
Table 3. Task Performance in Instructed Trials in Control and Inactivation Sessions, for Low and High Contrast Trials
Monkey
Session
F
Control
Inactivation
Control
Inactivation
HF
LH
LH
RH
RH
Fixation Breaks (%)
Saccade Errors (%)
Overall
Performance (%)
Latency (msec)
Low
Contrast
High
Contrast
Low
Contrast
High
Contrast
Low
Contrast
High
Contrast
Low
Contrast
High
Contrast
11.9
25.1
7.0
15.9
11.8*
27.5*
11.2
18.6
8.6
9.4
10.1
22.2
2.9
5.8
6.5
9.9
80.2
67.4
83.5*
64.8*
85.6*
68.2*
83.6
73.82
297
303
299
304
297
303
289
306
Significant inactivation effects are printed in bold. None of the comparisons between low contrast versus high contrast reached significance (separate
two-tailed t tests for within session and hemifield).
*p < .05 for comparison between control versus inactivation (separate two-tailed t tests within each contrast condition and session type).
interaction in the ANOVA between Drug × “Salience
manipulation” (i.e., reward vs. luminance; p < .05).
DISCUSSION
Lesions in the dorsal pulvinar lead to spatial deficits such
as an ipsilesional oculomotor choice bias. In this study,
we tested whether pulvinar inactivation affects saccades
toward memorized spatial positions and reward-based
target selection. We were particularly interested to see
whether the inactivation-induced spatial choice bias
could be overcome by manipulation of cue-associated
reward values. Although pulvinar inactivation increased
memory saccade latencies for contralesional targets, it
did not impair the ability to perform correct memory sac-
cades. Most importantly, we found that the strong ipsi-
lesional choice bias that was present under equal
reward conditions was strongly reduced when the cue in
the contralesional field predicted the high reward.
Pulvinar and Oculomotor Behavior
Inactivation resulted in an increase of memory saccade
latencies toward contralesional targets. Such an increase
Table 4. Percent of Right Hemifield (Contralesional) Choices
for Six Control and Six Inactivation Sessions as a Function of
Saliency Manipulation
Choices Towards RH (%)
Control
Inactivation
Low Equal High
Low Equal High
Salience
Manipulation
Reward
Luminance
2
23
77
72
97
90
1
7
33
22
70
40
Reward and luminance manipulations were conducted in control and
inactivation sessions in interleaved counterbalanced blocks within each
session. Significant effects of manipulation in inactivation sessions are
printed in bold.
for contralesional positions was not found in a previous
study that employed a speeded visually guided saccade
task (Wilke, Turchi, et al., 2010). It is conceivable that
pulvinar inactivation results in an intentional movement
deficit, which is more pronounced when no visual stim-
ulus is present. The dorsal pulvinar is reciprocally inter-
connected with several areas in the parietofrontal cortex
such as the lateral intraparietal (LIP) area, FEF, and the
dorsolateral pFC (dlPFC), where electrophysiological stud-
ies in monkeys have reported firing rate changes during
cue and delay-period intervals in the context of memory
saccade tasks (Takeda & Funahashi, 2002; Chafee &
Goldman-Rakic, 1998; Gnadt & Andersen, 1988). Corre-
spondingly, several studies demonstrated that lesions in
these areas impair performance in memory saccade tasks
(Wilke, Kagan, & Andersen, 2012; Liu, Yttri, & Snyder,
2010; Dias & Segraves, 1999; Li, Mazzoni, & Andersen,
1999; Funahashi, Bruce, & Goldman-Rakic, 1993). These
studies revealed that LIP and FEF inactivation were asso-
ciated with longer latencies and hypometric saccades
toward contralesional positions (Dias & Segraves, 1999; Li
et al., 1999), whereas dlPFC lesions did not result in longer
latencies but were characterized by erroneous saccade
directions (Funahashi et al., 1993). Thus, our findings are
consistent with the view that dorsal pulvinar lesions lead to
increased memory saccade latencies by affecting activity
within and/or between LIP and FEF areas. At the same time,
one of the monkeys showed an increase of fixation aborts
toward single ipsilesional cues after pulvinar inactivation.
This impairment of saccade inhibition is reminiscent of
the higher number of ipsilesional express saccades follow-
ing pulvinar inactivation that has been reported in a pre-
vious study that used a direct saccade paradigm ( Wilke,
Turchi, et al., 2010). Nonetheless, the effects of pulvinar
inactivation on saccade performance toward single tar-
gets were modest overall, and we thus conclude that the
medio-dorsal pulvinar is less critical for perceptual and
mnemonic processes but is more important for the for-
mation of movement intentions such as saccade planning
and initiation.
Wilke, Kagan, and Andersen
1279
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Pulvinar and Equal Reward Choices
We found that pulvinar inactivation led to a strong reduc-
tion of contralesional memory-guided saccade choices if
a saccade toward the ipsilesional space yielded the same
amount of reward. This choice bias is consistent with a
previous inactivation study that employed a visually guided
saccade task in monkeys ( Wilke, Turchi, et al., 2010)
and also corresponds to studies in humans with pulvinar
lesions (Arend, Machado, et al., 2008). Although we cannot
rule out the possibility of stochastic perceptual extinction,
monkeys typically did choose the contralesional target
when the cue color signaled the higher reward (see
below), and we thus interpret this bias not as primary
detection deficit but as movement intention deficit.
One possibility is that pulvinar inactivation increased
the thresholds to trigger a saccade toward the contra-
lesional space, whereas the thresholds for positions in
the ipsilesional space either decreased or remained the
same. This interpretation is consistent with the finding that
pulvinar inactivation resulted in longer memory saccade
latencies for contralesional targets even in the absence of
ipsilesional competition. Because the dorsal pulvinar
exchanges reciprocal connections with fronto-parietal
cortical areas and area STP in the dorsal bank of the STS,
which support visuospatial attention and eye movement
behavior, those deficits could be interpreted as dysfunction
in those thalamo-cortical networks (Saalmann & Kastner,
2011; Kaas & Lyon, 2007; Sherman, 2005; Gutierrez et al.,
2000; Scalaidhe, Albright, Rodman, & Gross, 1995;
Blatt, Andersen, & Stoner, 1990). Thus, by applying the
interhemispheric imbalance model (Kinsbourne, 1977),
one could assume that the latency increase and the
reduced choices of contralesional targets are because of
hypoactivity and/or disruption of cortico-cortical communi-
cation in those areas within the lesioned hemisphere. This
interpretation is particularly plausible with regard to pre-
vious findings of similar choice bias following lesions in
LIP (Wilke et al., 2012; Wardak, Olivier, & Duhamel,
2002) and FEF (Schiller & Chou, 1998). In addition, hyper-
activity in response to ipsilesional targets in corresponding
areas within the intact hemisphere could also contribute to
the observed ipsilesional choice bias.
Pulvinar and Reward-based Choices
Consistent with previous work in intact animals, when
reward magnitude differed between the two possible
choices, monkeys strongly preferred the position that
predicted the higher reward (McCoy & Platt, 2005; Opris
& Bruce, 2005). Importantly, this choice pattern remained
almost intact following inactivation, showing that the spa-
tial bias could be overcome by manipulating the reward
associated with the contralesional target. One interpreta-
tion can be that the dorsal pulvinar inactivation does not
impair the perceptual visibility of contralesional targets
but instead affects the intentional, motivational encoding
of contralesional movements. The high reward may have
increased the willingness of the monkeys to exert addi-
tional effort. This is in line with human and animal studies
showing that action decisions are weighted by reward and
the required effort (Kurniawan, Guitart-Masip, & Dolan,
2011). Also consistent with increased effort to overcome
the behavioral extinction are recent findings in a study in
which a decision bias was produced by unilateral revers-
ible inactivation of LIP in monkeys (Wilke et al., 2012). In
trials in which the animals still chose targets in the con-
tralesional hemifield, visual-oculomotor cortical areas in
both hemispheres showed increased BOLD signals, sug-
gesting bihemispheric compensation.
Studies in patients with cortical lesions have shown that
emotional and motivational factors have an impact on
neglect and extinction behavior (Dominguez-Borras, Saj,
Armony, & Vuilleumier, 2012). For example, preliminary
reports indicated that reward-predicting stimuli such
as money symbols can improve the detection perfor-
mance of neglect patients for contralesional targets (Lucas,
Diserens, Leroy, Krattinger, & Vuilleumier, 2005; Mesulam,
1999), and it has been demonstrated that extinction symp-
toms can be improved by adding emotional content to the
stimuli such as presenting angry or happy faces (Fox, 2002;
Vuilleumier et al., 2002; Vuilleumier & Schwartz, 2001),
emotional scenes (Grabowska et al., 2011), or pleasant music
(Soto et al., 2009). Likewise, studies in healthy participants
indicate that food and money incentives improve spatial
attention (Padmala & Pessoa, 2010; Engelmann & Pessoa,
2007) and perceptual processing of reward-associated stim-
ulus features (Hickey, Chelazzi, & Theeuwes, 2010; Pleger,
Blankenburg, Ruff, Driver, & Dolan, 2008). The effects of
stimulus–reward association in the current study are in
agreement with this literature.
Was the selection of high-reward targets driven by their
potentially higher perceptual visibility? The exact nature of
the interaction between reward and attentional/perceptual
processes remains unresolved (Pessoa & Engelmann,
2010), and a recent fMRI study demonstrated consider-
able overlap between neural networks involved in cue-
associated reward processing and attention (Krebs, Boehler,
Roberts, Song, & Woldorff, 2012). Thus, although one may
attempt to dissociate bottom–up visual saliency and action
selection signals, they may not represent entirely separable
processes but aspects of the same competition processes
implemented throughout the visuomotor system (Cisek &
Kalaska, 2010). Nevertheless, we wanted to see how the
direct manipulation of bottom–up visual saliency would
affect the choice behavior. Although the (behaviorally irrel-
evant) visual contrast strongly affected the choice behavior
in control conditions, after inactivation, the alleviation of
the choice bias by high visual saliency of the contralesional
stimulus was considerably weaker than by high reward.
This finding is consistent with studies in neglect patients
showing that manipulating the visual saliency in the con-
tralesional field by prolonging stimulus duration, increas-
ing size or luminance, improves extinction symptoms, but
1280
Journal of Cognitive Neuroscience
Volume 25, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
rather modestly (Bays, Singh-Curry, Gorgoraptis, Driver,
& Husain, 2010; Smania, Martini, Prior, & Marzi, 1996;
Di Pellegrino & De Renzi, 1995; Zihl & von Cramon, 1979).
These results suggest that the perceptual visibility of
contralesional cues was not the main factor guiding the
choice behavior after pulvinar inactivation—monkeys still
saw them and were willing to select them more fre-
quently if they led to high reward. The much greater effect
of reward on behavior after pulvinar inactivation suggests
that there may be separate sources of modulation for
reward and bottom–up visual saliency and that reward
has a greater effect on the decision process. Future studies
that parametrically equalize the effects of reward and visual
contrast would be required to address this possibility.
However, because stimulus detection and saccade re-
sponse were not dissociated in the current study, we
cannot exclude the possibility that perceptual factors also
contributed to the observed deficits. Moreover, in one of
the monkeys, the reward modulation did not completely
overcome the inactivation-induced spatial bias, suggest-
ing that perceptual extinction of contralesional targets
may have occurred in some of the trials.
What could be the neural mechanism for the restora-
tion of contralesional choices when the high reward cue
was placed into the contralesional hemifield? There are
two main possibilities, which are not mutually exclusive:
(1) Modulation of activity in cortical areas that receive
direct input from the pulvinar are involved in oculomotor
planning and are modulated by reward expectation.
Several cortical areas that exchange projections with
the dorso-medial pulvinar (e.g., FEF, dlPFC, LIP, and
the posterior cingulate) link visual cues with oculomotor
behavior and increase their activity with the expected
value of potential eye movements (Ding & Hikosaka,
2006; McCoy, Crowley, Haghighian, Dean, & Platt, 2003;
Leon & Shadlen, 1999; Platt & Glimcher, 1999). It is thus
possible that high reward expectation elevated activity in
those areas within the lesioned hemisphere to a level
that triggered a saccade choice into the contralesional
field. Assuming that activity in those cortical areas at
the side of the lesion was decreased following pulvinar
inactivation (Wilke, Kagan, & Andersen, 2010), the higher
reward in the contralesional field, in comparison with the
ipsilesional field (represented by the intact hemisphere),
may have increased activity in the lesioned hemisphere
over the decision threshold. (2) Alternatively, reward
could have restored contralesional choices by modula-
tion of reward-sensitive subcortical structures, which do
not receive direct projections from the pulvinar. For
example, the increase of contralesional choices with
higher reward could have been mediated by structures
of the oculomotor decision networks that contain spatial
maps such as the caudate (Mort et al., 2003), the SC
(Lauwereyns, Takikawa, et al., 2002), and the substantia
nigra (Sato & Hikosaka, 2002).
In conclusion, we propose that inactivation of the
dorsal pulvinar changes the balance between behavioral
desirability of competing spatial locations while leaving
the visibility, reward processing, spatial memory, and
saccade execution functions largely intact.
Acknowledgments
We thank K. Pejsa and N. Sammons for animal care and
Dr. V. Shcherbatyuk for computer support. This work was
supported by Moore Foundation, National Eye Institute,
National Science Foundation, Defense Advanced Research
Projects Agency, and Boswell Foundation (to R. A.) and the
Herman and Lilly Schilling Foundation (to M. W.).
Reprint requests should be sent to Melanie Wilke, University of
Goettingen, Robert-Koch-Str. 40, Goettingen, 37075, Germany,
or via e-mail: melanie.wilke@med.uni-goettingen.de.
REFERENCES
Arend, I., Machado, L., Ward, R., McGrath, M., Ro, T., & Rafal, R. D.
(2008). The role of the human pulvinar in visual attention and
action: Evidence from temporal-order judgment, saccade
decision, and antisaccade tasks. Progress in Brain Research,
171, 475–483.
Arend, I., Rafal, R., & Ward, R. (2008). Spatial and temporal
deficits are regionally dissociable in patients with pulvinar
lesions. Brain, 131, 2140–2152.
Asanuma, C., Andersen, R. A., & Cowan, W. M. (1985).
The thalamic relations of the caudal inferior parietal
lobule and the lateral prefrontal cortex in monkeys:
Divergent cortical projections from cell clusters in
the medial pulvinar nucleus. Journal of Comparative
Neurology, 241, 357–381.
Baker, J. T., Patel, G. H., Corbetta, M., & Snyder, L. H. (2006).
Distribution of activity across the monkey cerebral cortical
surface, thalamus and midbrain during rapid, visually guided
saccades. Cerebral Cortex, 16, 447–459.
Baleydier, C., & Mauguiere, F. (1985). Anatomical evidence for
medial pulvinar connections with the posterior cingulate cortex,
the retrosplenial area, and the posterior parahippocampal
gyrus in monkeys. Journal of Comparative Neurology, 232,
219–228.
Bays, P. M., Singh-Curry, V., Gorgoraptis, N., Driver, J., &
Husain, M. (2010). Integration of goal- and stimulus-related
visual signals revealed by damage to human parietal cortex.
Journal of Neuroscience, 30, 5968–5978.
Bender, D. B., & Youakim, M. (2001). Effect of attentive fixation
in macaque thalamus and cortex. Journal of Neurophysiology,
85, 219–234.
Benevento, L. A., & Port, J. D. (1995). Single neurons with both
form/color differential responses and saccade-related
responses in the nonretinotopic pulvinar of the behaving
macaque monkey. Visual Neuroscience, 12, 523–544.
Berman, R. A., & Wurtz, R. H. (2011). Signals conveyed in the
pulvinar pathway from superior colliculus to cortical area MT.
Journal of Neuroscience, 31, 373–384.
Blatt, G. J., Andersen, R. A., & Stoner, G. R. (1990). Visual
receptive field organization and cortico-cortical connections
of the lateral intraparietal area (area LIP) in the macaque.
Journal of Comparative Neurology, 299, 421–445.
Cappe, C., Morel, A., & Rouiller, E. M. (2007). Thalamocortical
and the dual pattern of corticothalamic projections of the
posterior parietal cortex in macaque monkeys. Neuroscience,
146, 1371–1387.
Chafee, M. V., & Goldman-Rakic, P. S. (1998). Matching patterns
of activity in primate prefrontal area 8a and parietal area 7ip
Wilke, Kagan, and Andersen
1281
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
neurons during a spatial working memory task. Journal of
Neurophysiology, 79, 2919–2940.
Cisek, P., & Kalaska, J. F. (2010). Neural mechanisms for
interacting with a world full of action choices. Annual Review
of Neuroscience, 33, 269–298.
Danziger, S., Ward, R., Owen, V., & Rafal, R. (2001). The effects
of unilateral pulvinar damage in humans on reflexive orienting
and filtering of irrelevant information. Behavioural Neurology,
13, 95–104.
Di Pellegrino, G., & De Renzi, E. (1995). An experimental
investigation on the nature of extinction. Neuropsychologia,
33, 153–170.
Krebs, R. M., Boehler, C. N., Roberts, K. C., Song, A. W., &
Woldorff, M. G. (2012). The involvement of the dopaminergic
midbrain and cortico-striatal-thalamic circuits in the integration
of reward prospect and attentional task demands. Cerebral
Cortex, 22, 607–615.
Krebs, R. M., Woldorff, M. G., Tempelmann, C., Bodammer, N.,
Noesselt, T., Boehler, C. N., et al. (2010). High-field fMRI
reveals brain activation patterns underlying saccade execution
in the human superior colliculus. PloS One, 5, e8691.
Kurniawan, I. T., Guitart-Masip, M., & Dolan, R. J. (2011).
Dopamine and effort-based decision making. Frontiers in
Neuroscience, 5, 81.
Dias, E. C., & Segraves, M. A. (1999). Muscimol-induced
Lauwereyns, J., Takikawa, Y., Kawagoe, R., Kobayashi, S.,
inactivation of monkey frontal eye field: Effects on visually
and memory-guided saccades. Journal of Neurophysiology,
81, 2191–2214.
Ding, L., & Hikosaka, O. (2006). Comparison of reward
modulation in the frontal eye field and caudate of the
macaque. Journal of Neuroscience, 26, 6695–6703.
Dominguez-Borras, J., Saj, A., Armony, J. L., & Vuilleumier, P.
(2012). Emotional processing and its impact on unilateral
neglect and extinction. Neuropsychologia, 50, 1054–1071.
Engelmann, J. B., & Pessoa, L. (2007). Motivation sharpens
exogenous spatial attention. Emotion, 7, 668–674.
Fox, E. (2002). Processing emotional facial expressions: The
role of anxiety and awareness. Cognitive, Affective &
Behavioral Neuroscience, 2, 52–63.
Funahashi, S., Bruce, C. J., & Goldman-Rakic, P. S. (1993).
Dorsolateral prefrontal lesions and oculomotor delayed-
response performance: Evidence for mnemonic “scotomas.”
Journal of Neuroscience, 13, 1479–1497.
Gnadt, J. W., & Andersen, R. A. (1988). Memory related motor
planning activity in posterior parietal cortex of macaque.
Experimental Brain Research, 70, 216–220.
Grabowska, A., Marchewka, A., Seniow, J., Polanowska, K.,
Jednorog, K., Krolicki, L., et al. (2011). Emotionally
negative stimuli can overcome attentional deficits in
patients with visuo-spatial hemineglect. Neuropsychologia,
49, 3327–3337.
Grieve, K. L., Acuna, C., & Cudeiro, J. (2000). The primate
pulvinar nuclei: Vision and action. Trends in Neurosciences,
23, 35–39.
Gutierrez, C., Cola, M. G., Seltzer, B., & Cusick, C. (2000).
Neurochemical and connectional organization of the dorsal
pulvinar complex in monkeys. Journal of Comparative
Neurology, 419, 61–86.
Hickey, C., Chelazzi, L., & Theeuwes, J. (2010). Reward changes
salience in human vision via the anterior cingulate. Journal of
Neuroscience, 30, 11096–11103.
Hulme, O. J., Whiteley, L., & Shipp, S. (2010). Spatially
distributed encoding of covert attentional shifts in human
thalamus. Journal of Neurophysiology, 104, 3644–3656.
Kaas, J. H., & Lyon, D. C. (2007). Pulvinar contributions to the
dorsal and ventral streams of visual processing in primates.
Brain Research Reviews, 55, 285–296.
Kagan, I., Iyer, A., Lindner, A., & Andersen, R. A. (2010). Space
representation for eye movements is more contralateral in
monkeys than in humans. Proceedings of the National
Academy of Sciences, U.S.A., 107, 7933–7938.
Karnath, H. O., Himmelbach, M., & Rorden, C. (2002). The
subcortical anatomy of human spatial neglect: Putamen,
caudate nucleus and pulvinar. Brain, 125, 350–360.
Kastner, S., OʼConnor, D. H., Fukui, M. M., Fehd, H. M.,
Herwig, U., & Pinsk, M. A. (2004). Functional imaging of the
human lateral geniculate nucleus and pulvinar. Journal of
Neurophysiology, 91, 438–448.
Kinsbourne, M. (1977). Hemi-neglect and hemisphere rivalry.
Advances in Neurology, 18, 41–49.
Koizumi, M., Coe, B., et al. (2002). Feature-based anticipation
of cues that predict reward in monkey caudate nucleus.
Neuron, 33, 463–473.
Lauwereyns, J., Watanabe, K., Coe, B., & Hikosaka, O. (2002). A
neural correlate of response bias in monkey caudate nucleus.
Nature, 418, 413–417.
Leon, M. I., & Shadlen, M. N. (1999). Effect of expected reward
magnitude on the response of neurons in the dorsolateral
prefrontal cortex of the macaque. Neuron, 24, 415–425.
Li, C. S., Mazzoni, P., & Andersen, R. A. (1999). Effect of
reversible inactivation of macaque lateral intraparietal area on
visual and memory saccades. Journal of Neurophysiology,
81, 1827–1838.
Liu, Y., Yttri, E. A., & Snyder, L. H. (2010). Intention and
attention: Different functional roles for LIPd and LIPv.
Nature Neuroscience, 13, 495–500.
Lucas, N., Diserens, K., Leroy, R., Krattinger, S., & Vuilleumier,
P. (2005). Gambling against neglect: Modulation of spatial
attention by implicit reward learning. Journal of Cognitive
Neuroscience, Suppl. S, 151.
McCoy, A. N., Crowley, J. C., Haghighian, G., Dean, H. L., &
Platt, M. L. (2003). Saccade reward signals in posterior
cingulate cortex. Neuron, 40, 1031–1040.
McCoy, A. N., & Platt, M. L. (2005). Expectations and outcomes:
Decision-making in the primate brain. Journal of Comparative
Physiology A: Neuroethology, Sensory, Neural, and Behavioral
Physiology, 191, 201–211.
Mesulam, M. M. (1999). Spatial attention and neglect: Parietal,
frontal and cingulate contributions to the mental representation
and attentional targeting of salient extrapersonal events.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 354, 1325–1346.
Milstein, D. M., & Dorris, M. C. (2007). The influence of expected
value on saccadic preparation. Journal of Neuroscience, 27,
4810–4818.
Mort, D. J., Malhotra, P., Mannan, S. K., Rorden, C., Pambakian, A.,
Kennard, C., et al. (2003). The anatomy of visual neglect. Brain,
126, 1986–1997.
Opris, I., & Bruce, C. J. (2005). Neural circuitry of judgment and
decision mechanisms. Brain Research, Brain Research
Reviews, 48, 509–526.
Padmala, S., Lim, S. L., & Pessoa, L. (2010). Pulvinar and
affective significance: Responses track moment-to-moment
stimulus visibility. Frontiers in Human Neuroscience,
4, 64.
Padmala, S., & Pessoa, L. (2010). Interactions between
cognition and motivation during response inhibition.
Neuropsychologia, 48, 558–565.
Pessoa, L., & Engelmann, J. B. (2010). Embedding reward signals
into perception and cognition. Frontiers in Neuroscience,
4, 17.
Petersen, S. E., Robinson, D. L., & Keys, W. (1985). Pulvinar
nuclei of the behaving rhesus monkey: Visual responses
and their modulation. Journal of Neurophysiology, 54,
867–886.
1282
Journal of Cognitive Neuroscience
Volume 25, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Petersen, S. E., Robinson, D. L., & Morris, J. D. (1987).
Contributions of the pulvinar to visual spatial attention.
Neuropsychologia, 25, 97–105.
Platt, M. L., & Glimcher, P. W. (1999). Neural correlates of
decision variables in parietal cortex. Nature, 400, 233–238.
Pleger, B., Blankenburg, F., Ruff, C. C., Driver, J., & Dolan, R. J.
(2008). Reward facilitates tactile judgments and modulates
hemodynamic responses in human primary somatosensory
cortex. Journal of Neuroscience, 28, 8161–8168.
Rafal, R. D., & Posner, M. I. (1987). Deficits in human visual
spatial attention following thalamic lesions. Proceedings of
the National Academy of Sciences, U.S.A., 84, 7349–7353.
Robinson, D. L., McClurkin, J. W., & Kertzman, C. (1990).
Orbital position and eye movement influences on visual
responses in the pulvinar nuclei of the behaving macaque.
Experimental Brain Research, 82, 235–246.
Robinson, D. L., McClurkin, J. W., Kertzman, C., & Petersen, S. E.
(1991). Visual responses of pulvinar and collicular neurons
during eye movements of awake, trained macaques. Journal
of Neurophysiology, 66, 485–496.
Robinson, D. L., & Petersen, S. E. (1992). The pulvinar and
visual salience. Trends in Neurosciences, 15, 127–132.
Saalmann, Y. B., & Kastner, S. (2011). Cognitive and perceptual
functions of the visual thalamus. Neuron, 71, 209–223.
Saleem, K., & Logothetis, N. K. (2006). A combined MRI and
histology atlas of the rhesus monkey brain. New York:
Academic Press.
Sato, M., & Hikosaka, O. (2002). Role of primate substantia
nigra pars reticulata in reward-oriented saccadic eye
movement. Journal of Neuroscience, 22, 2363–2373.
Scalaidhe, S. P., Albright, T. D., Rodman, H. R., & Gross, C. G.
(1995). Effects of superior temporal polysensory area lesions
on eye movements in the macaque monkey. Journal of
Neurophysiology, 73, 1–19.
Schiller, P. H., & Chou, I. H. (1998). The effects of frontal eye
field and dorsomedial frontal cortex lesions on visually
guided eye movements. Nature Neuroscience, 1, 248–253.
Selemon, L. D., & Goldman-Rakic, P. S. (1988). Common
cortical and subcortical targets of the dorsolateral prefrontal
and posterior parietal cortices in the rhesus monkey: Evidence
for a distributed neural network subserving spatially guided
behavior. Journal of Neuroscience, 8, 4049–4068.
Sherman, S. M. (2005). Thalamic relays and cortical functioning.
Progress in Brain Research, 149, 107–126.
Smania, N., Martini, M. C., Prior, M., & Marzi, C. A. (1996). Input
and response determinants of visual extinction: A case study.
Cortex, 32, 567–591.
Soto, D., Funes, M. J., Guzman-Garcia, A., Warbrick, T.,
Rotshtein, P., & Humphreys, G. W. (2009). Pleasant music
overcomes the loss of awareness in patients with visual
neglect. Proceedings of the National Academy of Sciences,
U.S.A., 106, 6011–6016.
Stepniewska, I. (2004). The pulvinar complex. In J. H. Kaas &
C. E. Collins (Eds.), The primate visual system (pp. 53–80).
London: CRC Press.
Takeda, K., & Funahashi, S. (2002). Prefrontal task-related
activity representing visual cue location or saccade direction
in spatial working memory tasks. Journal of Neurophysiology,
87, 567–588.
Takikawa, Y., Kawagoe, R., Itoh, H., Nakahara, H., & Hikosaka,
O. (2002). Modulation of saccadic eye movements by
predicted reward outcome. Experimental Brain Research,
142, 284–291.
Van der Stigchel, S., Arend, I., van Koningsbruggen, M. G., &
Rafal, R. D. (2010). Oculomotor integration in patients
with a pulvinar lesion. Neuropsychologia, 48, 3497–3504.
Vuilleumier, P., Armony, J. L., Clarke, K., Husain, M., Driver, J.,
& Dolan, R. J. (2002). Neural response to emotional faces
with and without awareness: Event-related fMRI in a parietal
patient with visual extinction and spatial neglect.
Neuropsychologia, 40, 2156–2166.
Vuilleumier, P., & Schwartz, S. (2001). Emotional facial
expressions capture attention. Neurology, 56, 153–158.
Wardak, C., Olivier, E., & Duhamel, J. R. (2002). Saccadic target
selection deficits after lateral intraparietal area inactivation in
monkeys. Journal of Neuroscience, 22, 9877–9884.
Wilke, M., Kagan, I., & Andersen, R. A. (2010). Pulvinar
inactivation alters cortical responses during spatial
decision making (Conference abstract). Society for
Neuroscience Meeting, San Diego.
Wilke, M., Kagan, I., & Andersen, R. A. (2012). Functional
imaging reveals rapid reorganization of cortical activity after
parietal inactivation in monkeys. Proceedings of the National
Academy of Sciences, U.S.A., 109, 8274–8279.
Wilke, M., Mueller, K. M., & Leopold, D. A. (2009). Neural
activity in the visual thalamus reflects perceptual suppression.
Proceedings of the National Academy of Sciences, U.S.A.,
106, 9465–9470.
Wilke, M., Turchi, J., Smith, K., & Leopold, D. A. (2010). Pulvinar
inactivation disrupts selection of movement plans. Journal of
Neuroscience, 30, 8650–8659.
Yeterian, E. H., & Pandya, D. N. (1989). Thalamic connections
of the cortex of the superior temporal sulcus in the
rhesus monkey. Journal of Comparative Neurology,
282, 80–97.
Zihl, J., & von Cramon, D. (1979). The contribution of the
“second” visual system to directed visual attention in man.
Brain, 102, 835–856.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
8
2
5
1
/
2
8
7
/
0
1
1
2
9
7
4
0
5
/
6
1
9
7
3
7
o
9
c
5
n
1
_
3
a
/
_
j
0
o
0
c
3
n
9
9
_
a
p
_
d
0
0
b
3
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Wilke, Kagan, and Andersen
1283